Embed Size (px)
Citation preview

www.elsevier.com/locate/ijpara
International Journal for Parasitology 37 (2007) 131–138
Review Article
Acanthamoeba invasion of the central nervous system
Naveed Ahmed Khan *
School of Biological and Chemical Sciences, Birkbeck College, University of London, London WC1E 7HX, UK
Received 18 September 2006; received in revised form 9 November 2006; accepted 15 November 2006
Abstract
Pathogenic Acanthamoeba are known to infect the CNS, resulting in fatal granulomatous encephalitis. The mechanisms associatedwith the pathogenesis remain unclear; however pathophysiological complications involving the CNS most likely include induction ofpro-inflammatory responses, invasion of the blood–brain barrier and the connective tissue and neuronal damage leading to brain dys-function. The routes of entry include the olfactory neuroepithelium pathway and/or lower respiratory tract, followed by haematogenousspread. Skin lesions may provide direct entry into the bloodstream, bypassing the lower respiratory tract. For the haematogenous route,entry of amoebae into the CNS most likely occurs at the sites of the blood–brain barrier. Recent studies have identified several molecularmechanisms associated with Acanthamoeba traversal of the blood–brain barrier and targeting those may help develop therapeutic inter-ventions and/or design preventative strategies.� 2006 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
Keywords: Acanthamoeba; Granulomatous encephalitis; Central nervous system; Blood–brain barrier
1. Introduction
Acanthamoeba is an opportunistic protozoan pathogenthat is widely distributed in the environment. PathogenicAcanthamoeba have been shown to cause serious humandiseases, including painful keratitis resulting in blindnessand fatal granulomatous encephalitis (reviewed in Marci-ano-Cabral and Cabral, 2003; Shuster and Visvesvara,2004; Clarke and Niederkorn, 2006; Khan, 2006). Howev-er, the pathogenesis and pathophysiology associated withAcanthamoeba infections are still far from clear. Althoughthe current methods of treatment against Acanthamoeba
keratitis are shown to be somewhat effective, treatment ofAcanthamoeba granulomatous encephalitis (AGE) is moreproblematic. There is no single drug that is shown to beeffective in AGE infections. Current methods of treatmentinclude a combinatorial approach, where a mixture ofdrugs is administered, and even then the prognosis remainsextremely poor. There is an urgent need for improved anti-
0020-7519/$30.00 � 2006 Australian Society for Parasitology Inc. Published b
doi:10.1016/j.ijpara.2006.11.010
* Tel.: +44 0207 079 0797; fax: +44 0207 631 6246.E-mail address: [email protected]
microbial chemotherapy and/or alternative strategies todevelop therapeutic interventions. The purpose of thisreview is to discuss our current understanding of Acantha-
moeba invasion of the CNS, a pre-requisite in the develop-ment of AGE.
2. Acanthamoeba granulomatous encephalitis
Central nervous system infections due to pathogenicAcanthamoeba mostly occur in chronically ill or immuno-compromised patients due to human immunodeficiencyvirus infection, diabetes, immunosuppressive therapy,malignancies, malnutrition or alcoholism. The clinicalsymptoms may resemble viral, bacterial or tuberculosismeningitis involving headache, fever, behavioural changes,hemiparesis, lethargy, stiff neck, aphasia, ataxia, vomiting,nausea, cranial nerve palsies, increased intracranial pressureand seizures, finally leading to death. Post-mortem exami-nation often shows severe oedema and hemorrhagic necro-sis (Martinez, 1985, 1991; Martinez and Visvesvara, 1997).The pathophysiological complications involving the CNSmost likely include induction of pro-inflammatory responses,
y Elsevier Ltd. All rights reserved.

132 N.A. Khan / International Journal for Parasitology 37 (2007) 131–138
invasion of the blood–brain barrier and the connective tis-sue and neuronal damage leading to the brain dysfunction(Shin et al., 2001; Benedetto and Auriault, 2002a,b; Mat-tana et al., 2002; Marciano-Cabral and Cabral, 2003;Marciano-Cabral et al., 2004; Shuster and Visvesvara,2004). Cutaneous infections due to Acanthamoeba can lastfor months but the involvement of the CNS can result infatal consequences within days or weeks. A granulomatousresponse may be absent or minimal in patients with aseverely impaired immune system that is interpreted asimpairment of cellular immune response (Martinez, 1985,1991).
3. Strategies of Acanthamoeba invasion of the central
nervous system: at the tissue level
The mechanisms by which foreign molecules/com-pounds enter the CNS have gained particular attentionfrom the scientific community. The understanding of path-ways by which CNS protective barriers are breached is notonly important for pathogen/toxin entry into the CNSleading to pathophysiological events but equally crucialfor our ability to devise mechanisms for the rationaledesign of therapeutic interventions. The routes of Acantha-
moeba entry into the CNS were described in landmarkstudies by Culbertson et al. (1959) and Cerva (1967). Itwas shown that intranasal, intrapulmonary and intracardi-al inoculations of amobae into experimental animals leadto their invasion of the CNS, suggesting that amoebaecan enter the CNS by different routes. This is consistentwith clinical findings and it is now widely accepted thatentry of amoebae into the brain most likely occurs eitherthrough the olfactory neuroepithelium route and/or viathe bloodstream. In the former, amoebae invade the nasalmucosa and migrate along nerve fibers, followed by inva-sion of the olfactory bulb. However, this review is focussedon the latter, where Acanthamoeba enter the lungs via thenasal route. Next, amoebae traverse the lungs into thebloodstream, followed by haematogenous spread. Finally,Acanthamoeba cross the blood–brain barrier, enter theCNS and produce disease. This was shown by findings thatamoebae that were instilled intranasally into mice developpulmonary disease, followed by neurological signs (Marti-nez et al., 1975). Pathological examination of these miceshowed broncho-pneumonia associated with encephalitiswith amoebic trophozoites and cysts in the lung and brain,suggesting the involvement of haematogenous spread(Martinez et al., 1975). Skin lesions may provide directentry of amoebae into the bloodstream, thus bypassingthe lower respiratory tract. Affected tissues other than theCNS may include subcutaneous tissue, skin, liver, lungs,kidneys, adrenals, pancreas, prostate, lymph nodes andbone marrow, which further suggest haematogenousspread pre-mortem. Blood vessels serving the brain are col-lectively called the blood–brain barrier. Thus amoebaeentry into the CNS may occur at the sites of the blood–brain barrier (Martinez et al., 1975; Martinez, 1985,
1991). The blood–brain barrier is a diffusion barrier essen-tial for the normal function of the CNS (Tuomanen, 1996).It can be further divided into two levels: a barrier betweenblood and brain extracellular fluid located at the cerebralcapillary endothelium (Fig. 1A) and a barrier betweenblood and CSF located at the choroid plexus epitheliumand the arachnoid membrane (Fig. 1B). The former is com-posed of basement membrane (containing laminin, colla-gen IV, proteoglycans (especially heparan sulphate),fibronectins, nidogen and entactin), astrocytes (a type ofglial cells) that extend foot-like projections around the out-side of the capillaries, pericytes that are embedded withinthe basement membrane and endothelial cells (Fig. 2).Endothelial cells are characterised by the presence of tightjunctions (consisting of three integral membrane proteins,claudin, occludin, junction adhesion molecule and cyto-plasmic proteins, zonula occludens-1 (ZO-1, ZO-2 andZO-3) and adherens junctions (composed of membraneprotein, cadherin that bind to catenin that are linked tothe actin cytoskeleton and form adhesive contacts betweenthe cells) (Fig. 3). Zonula occludens are accessory proteinsthat provide structural support and bind to all three inte-gral membrane proteins of the tight junctions, further link-ing membrane proteins to the actin cytoskeleton(Tuomanen, 1996; Huber et al., 2001; Kim et al., 2006).In contrast, the blood–CSF barrier (at the choroids plexus)is found within each of the four cerebral ventricles and isresponsible for the production of CSF. Here, endothelialcells are fenestrated to allow penetration of the blood con-tents but are surrounded by epithelial cells of the choroidplexus (Fig. 1). Both tight junctions and adherens junctionsare present in the epithelial cells of the choroid plexus(Fig. 3). Of interest, claudins-1, -2, -11, occludin and ZO-1 are present in the choroid plexus epithelial cells (blood–CSF barrier), whereas claudins-1, -5, -11, occludin andZO-1 are present in the blood–brain barrier (Wolburget al., 2001).
4. Acanthamoeba traversal of the blood–brain barrier: at thecellular level
As indicated above, Acanthamoeba invasion and pene-tration of the CNS may occur at the blood–brain barrierthat is located at two sites: (i) the cerebral capillary endothe-lium and (ii) the choroid plexus. To this end, several lines ofevidence suggest that Acanthamoeba entry into the CNSmost likely occurs at the cerebral capillary endothelium.This has been demonstrated with observations that lesionsare most frequent in the brain parenchyma of AGE-infectedpatients who develop encephalitis (Martinez et al., 1975;Martinez, 1985, 1991; Martinez and Visvesvara, 1997).H&E-stained sections of brain tissue exhibit large numbersof amoebae in the perivascular space. In contrast, patho-gens that enter via the choroid plexus route will end up inthe CSF. Thus, the pathology in such cases will first appearin the ventricles, and only at later stages with extensiveinflammation will it involve the brain parenchyma, a finding

Blood-side
Brain-side
Endothelialcells
Blood-side
CSF
Endothelialcells
Epithelial cells
Cerebral capillaryendothelium
Choroid plexus
Astrocytes
Tight junctions
Fenestratedendothelium
Tight junctions
A
B
Fig. 1. The blood–brain barrier can be further divided into two levels, a barrier between blood and brain extracellular fluid located at the cerebral capillaryendothelium (A) and a barrier between blood and CSF located at the choroid plexus epithelium (B). Of note, amoebae entry into the CNS most likelyoccurs at the cerebral capillary endothelium.
Pericyte
Astrocyte Basement membrane
Tight junction
Astrocyte
Endothelium
Astrocyte
Blood
Fig. 2. Schematic illustration of the blood–brain barrier at the cerebralcapillary endothelium. The endothelial cells are surrounded by basementmembrane composed of collagen type IV, laminin, fibronectins andheparan sulfate proteoglycans which is ensheathed by astrocytes. Theendothelial cells from brain capillaries are distinct from the endothelialcells from peripheral capillaries in the presence of tight junctions,increased mitochondria and association with astrocytes.
N.A. Khan / International Journal for Parasitology 37 (2007) 131–138 133
that is normally inconsistent with AGE pathological fea-tures and further suggests that traversal of the blood–brainbarrier by amoebae most likely occurs at the cerebral capil-lary endothelium. However, CSF is continuous with theextracellular fluid of the brain parenchyma, and flows uni-directionally from ventricles down to the lumbar region,thus spinal taps of patients may aid in the early diagnosisof AGE, which is a pre-requisite in the successful treatmentof AGE infection (Petry et al., 2006).
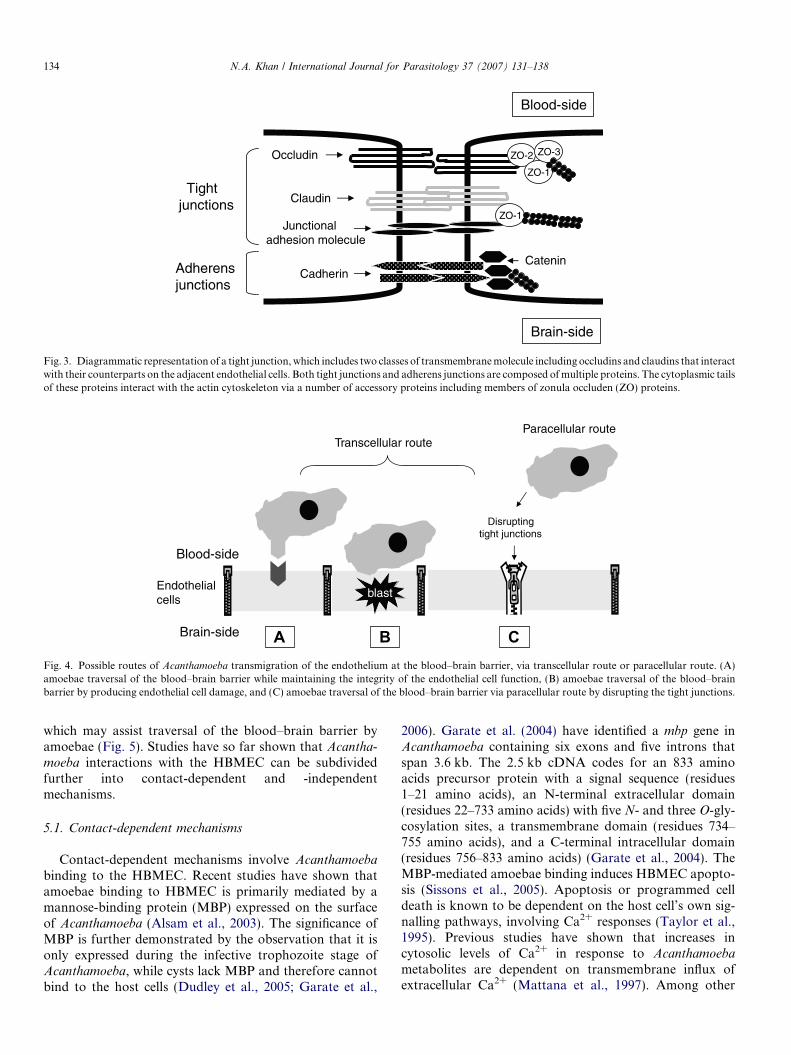
At the cellular-level, pathogens can penetrate the blood–brain barrier either through the cell (transcellular route) orbetween the cells (paracellular route) (Fig. 4). Pathogensmay use the paracellular route either by injuring the cellor traversing the cell while maintaining the integrity ofthe cell monolayers (Fig. 4). In addition, leukocyte-facili-tated entry into the CNS using Trojan horse mechanismsis employed by many intracellular pathogens (Drevetsand Leenen, 2000). Given the size of Acanthamoeba, i.e.,12–35 lm, it is likely that Acanthamoeba transmigrationof the endothelium at the sites of the blood–brain barrieroccurs via the transcellular route by producing endothelialcell damage, and/or via the paracellular route by targetingthe tight junctions (Fig. 4).
5. Acanthamoeba invasion of the blood–brain barrier: at the
molecular level
The study of Acanthamoeba–host cell interactions at themolecular-level is crucial for the rationale development oftherapeutic interventions. Recent availability of in vitro
cultures of primary human brain microvascular endothelialcells (HBMEC), which constitute the blood–brain barrier,have been of tremendous value in the identification andcharacterisation of molecular determinants that arerequired in host–parasite interactions (Stins et al., 1997;Alsam et al., 2003). Our recent studies have been focussedon Acanthamoeba interactions with the primary HBMECand showed that amoebae produce HBMEC dysfunction,
ZO-2
ZO-1
ZO-3
ZO-1
Catenin Cadherin
Junctionaladhesion molecule
Claudin
Occludin
Blood-side
Brain-side
Tightjunctions
Adherensjunctions
Fig. 3. Diagrammatic representation of a tight junction, which includes two classes of transmembrane molecule including occludins and claudins that interactwith their counterparts on the adjacent endothelial cells. Both tight junctions and adherens junctions are composed of multiple proteins. The cytoplasmic tailsof these proteins interact with the actin cytoskeleton via a number of accessory proteins including members of zonula occluden (ZO) proteins.
Paracellular route
Blood-side
Brain-side
Transcellular route
blast
Disrupting tight junctions
Endothelialcells
A B CFig. 4. Possible routes of Acanthamoeba transmigration of the endothelium at the blood–brain barrier, via transcellular route or paracellular route. (A)amoebae traversal of the blood–brain barrier while maintaining the integrity of the endothelial cell function, (B) amoebae traversal of the blood–brainbarrier by producing endothelial cell damage, and (C) amoebae traversal of the blood–brain barrier via paracellular route by disrupting the tight junctions.
134 N.A. Khan / International Journal for Parasitology 37 (2007) 131–138
which may assist traversal of the blood–brain barrier byamoebae (Fig. 5). Studies have so far shown that Acantha-
moeba interactions with the HBMEC can be subdividedfurther into contact-dependent and -independentmechanisms.
5.1. Contact-dependent mechanisms
Contact-dependent mechanisms involve Acanthamoeba
binding to the HBMEC. Recent studies have shown thatamoebae binding to HBMEC is primarily mediated by amannose-binding protein (MBP) expressed on the surfaceof Acanthamoeba (Alsam et al., 2003). The significance ofMBP is further demonstrated by the observation that it isonly expressed during the infective trophozoite stage ofAcanthamoeba, while cysts lack MBP and therefore cannotbind to the host cells (Dudley et al., 2005; Garate et al.,
2006). Garate et al. (2004) have identified a mbp gene inAcanthamoeba containing six exons and five introns thatspan 3.6 kb. The 2.5 kb cDNA codes for an 833 aminoacids precursor protein with a signal sequence (residues1–21 amino acids), an N-terminal extracellular domain(residues 22–733 amino acids) with five N- and three O-gly-cosylation sites, a transmembrane domain (residues 734–755 amino acids), and a C-terminal intracellular domain(residues 756–833 amino acids) (Garate et al., 2004). TheMBP-mediated amoebae binding induces HBMEC apopto-sis (Sissons et al., 2005). Apoptosis or programmed celldeath is known to be dependent on the host cell’s own sig-nalling pathways, involving Ca2+ responses (Taylor et al.,1995). Previous studies have shown that increases incytosolic levels of Ca2+ in response to Acanthamoeba
metabolites are dependent on transmembrane influx ofextracellular Ca2+ (Mattana et al., 1997). Among other
pRB
Cell death
HBMEC
E2FpRB
PP P
S phase(DNA synthesis) Cell cycle
arrest
PIP2 PIP3
PH
AktE2F
X
Proteases MBP
?
PI3K
Occludin ZO-1
Permeability increase
Targeting tight junctions & ECM
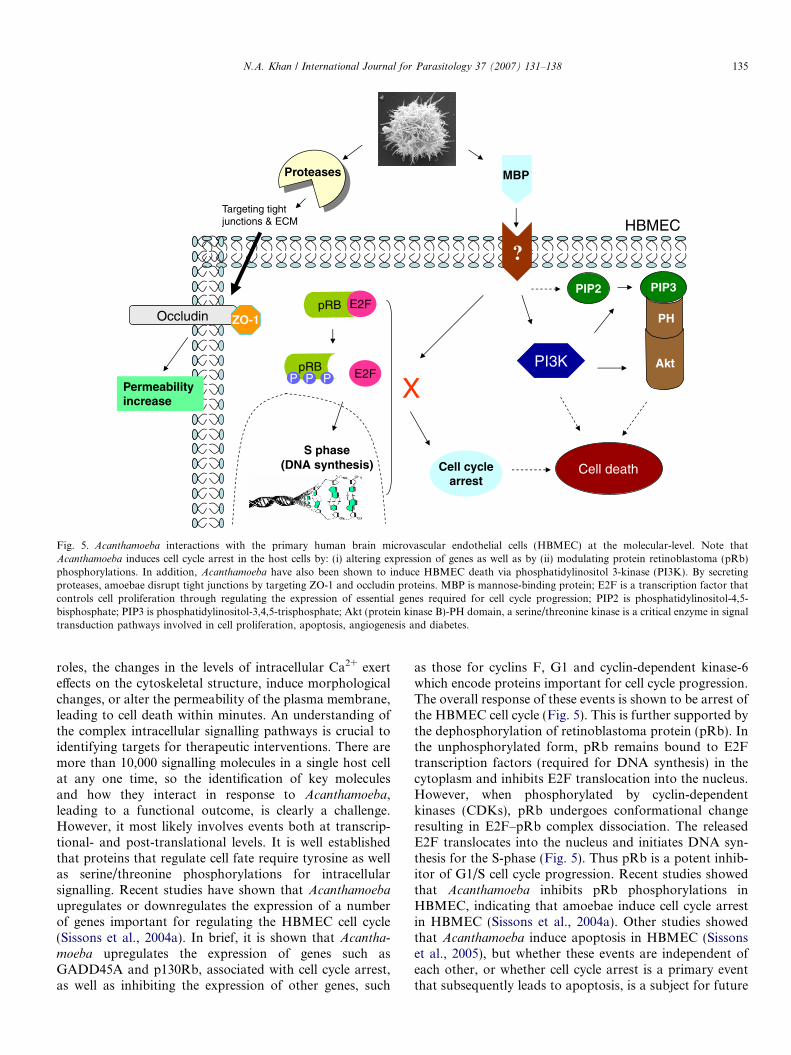
Fig. 5. Acanthamoeba interactions with the primary human brain microvascular endothelial cells (HBMEC) at the molecular-level. Note thatAcanthamoeba induces cell cycle arrest in the host cells by: (i) altering expression of genes as well as by (ii) modulating protein retinoblastoma (pRb)phosphorylations. In addition, Acanthamoeba have also been shown to induce HBMEC death via phosphatidylinositol 3-kinase (PI3K). By secretingproteases, amoebae disrupt tight junctions by targeting ZO-1 and occludin proteins. MBP is mannose-binding protein; E2F is a transcription factor thatcontrols cell proliferation through regulating the expression of essential genes required for cell cycle progression; PIP2 is phosphatidylinositol-4,5-bisphosphate; PIP3 is phosphatidylinositol-3,4,5-trisphosphate; Akt (protein kinase B)-PH domain, a serine/threonine kinase is a critical enzyme in signaltransduction pathways involved in cell proliferation, apoptosis, angiogenesis and diabetes.
N.A. Khan / International Journal for Parasitology 37 (2007) 131–138 135
roles, the changes in the levels of intracellular Ca2+ exerteffects on the cytoskeletal structure, induce morphologicalchanges, or alter the permeability of the plasma membrane,leading to cell death within minutes. An understanding ofthe complex intracellular signalling pathways is crucial toidentifying targets for therapeutic interventions. There aremore than 10,000 signalling molecules in a single host cellat any one time, so the identification of key moleculesand how they interact in response to Acanthamoeba,leading to a functional outcome, is clearly a challenge.However, it most likely involves events both at transcrip-tional- and post-translational levels. It is well establishedthat proteins that regulate cell fate require tyrosine as wellas serine/threonine phosphorylations for intracellularsignalling. Recent studies have shown that Acanthamoeba
upregulates or downregulates the expression of a numberof genes important for regulating the HBMEC cell cycle(Sissons et al., 2004a). In brief, it is shown that Acantha-
moeba upregulates the expression of genes such asGADD45A and p130Rb, associated with cell cycle arrest,as well as inhibiting the expression of other genes, such
as those for cyclins F, G1 and cyclin-dependent kinase-6which encode proteins important for cell cycle progression.The overall response of these events is shown to be arrest ofthe HBMEC cell cycle (Fig. 5). This is further supported bythe dephosphorylation of retinoblastoma protein (pRb). Inthe unphosphorylated form, pRb remains bound to E2Ftranscription factors (required for DNA synthesis) in thecytoplasm and inhibits E2F translocation into the nucleus.However, when phosphorylated by cyclin-dependentkinases (CDKs), pRb undergoes conformational changeresulting in E2F–pRb complex dissociation. The releasedE2F translocates into the nucleus and initiates DNA syn-thesis for the S-phase (Fig. 5). Thus pRb is a potent inhib-itor of G1/S cell cycle progression. Recent studies showedthat Acanthamoeba inhibits pRb phosphorylations inHBMEC, indicating that amoebae induce cell cycle arrestin HBMEC (Sissons et al., 2004a). Other studies showedthat Acanthamoeba induce apoptosis in HBMEC (Sissonset al., 2005), but whether these events are independent ofeach other, or whether cell cycle arrest is a primary eventthat subsequently leads to apoptosis, is a subject for future

Fig. 6. Acanthamoeba-induced perturbation of tight junctions barrier.Acanthamoeba were incubated with RPMI-1640 for 24 h at 30 �C. Nextday, cell-free medium (Acanthamoeba conditioned medium, ACM) wascollected by centrifugation. The ACM was incubated with the confluenthuman brain microvascular endothelial cell (HBMEC) monolayers forvarious intervals of time and cells were lysed by RIPA lysis buffer. Equalamounts of cell lysates were used for Western blotting assays using anti-ZO-1 (A) and anti-occludin (B) antibodies. Note that ACM-induced lossof ZO-1 and occludin within 30 min. Results are representative of threeindependent experiments.
136 N.A. Khan / International Journal for Parasitology 37 (2007) 131–138
studies. Recent studies have shown that Acanthamoeba-mediated HBMEC is dependent on the activation of phos-phatidylinositol 3-kinase (PI3K) (Fig. 5). This was con-firmed using LY294002, a specific PI3K inhibitor as wellas using HBMEC expressing mutant p85, i.e., a regulatorysubunit of PI3K (dominant-negative PI3K). The path-way(s) downstream of PI3K leading to Acanthamoeba-mediated HBMEC death remain unidentified. Studies inmyeloma cells showed that activation of proapoptotic mol-ecules, Bak and Bax, loss of mitochondrial membranepotential and release of cytochrome c are likely effectormolecules downstream of PI3K-mediated host cell death(Thyrell et al., 2004). Overall, these studies showed thatAcanthamoeba binds to HBMEC in a MBP-mediated man-ner, inducing intracellular signalling pathways leading toapoptosis. In addition, the adhesion of Acanthamoeba tothe HBMEC leads to secondary events such as phagocyto-sis. The ability of Acanthamoeba to form food cups oramoebastomes during incubations with the host cells sug-gests that phagocytosis plays an important role in Acantha-
moeba pathogenesis (Pettit et al., 1996; Khan, 2001; Alsamet al., 2005a). In support of this, it is shown that cytocha-lasin D (a toxin that blocks actin polymerization) inhibitsAcanthamoeba-mediated HBMEC death, confirming thatactin-mediated cytoskeletal rearrangements of Acantha-
moeba are important in their ability to phagocytoseHBMEC (unpublished data). Overall, these findings sug-gest that MBP is crucial in Acanthamoeba binding to thetarget cells, as well as its phagocytotic processes.
Among other determinants, ecto-ATPases have beenshown to play a role in Acanthamoeba-mediated HBMECdamage. Ecto-ATPases are glycoproteins present in plasmamembranes that have their active sites facing the externalmedium rather than the cytoplasm and are known tohydrolyse extracellular adenosine 5 0-triphosphate (ATP)and other nucleoside triphosphates (Sissons et al., 2004b).The resultant adenosine diphosphate (ADP) can have toxiceffects on host cells by binding to P2y2 purinergic receptors.Recent studies have shown that a P2 receptor antagonist,suramin, inhibits Acanthamoeba-mediated HBMEC death(Mattana et al., 2002; Sissons et al., 2004b), indicating thatecto-ATPases may play a role in Acanthamoeba pathogen-esis. Again, exogenous mannose enhances ecto-ATPaseactivities, suggesting that the engagement of Acanthamoeba
adhesin, i.e. MBP, is associated with ecto-ATPase activities(Sissons et al., 2004b).
5.2. Contact-independent mechanisms
Among contact-independent mechanisms, extracellularproteases are seen to be the most potent in amoebae-medi-ated HBMEC monolayer disruptions (Alsam et al., 2005b).Proteases hydrolyse peptide bonds and thus exhibit the abil-ity to degrade various substrates. Acanthamoeba secretelarge amounts of proteases, in particular serine proteases(Khan et al., 2000). Acanthamoeba serine proteases havebeen shown to degrade types I, III and IV collagen, elastin
and fibronectin, which are main components of the extracel-lular matrix, as well as fibrinogen, IgG, IgA, albumin, plas-minogen (involved in proteolytic degradation of ECM), andhaemoglobin (Kong et al., 2000; Sissons et al., 2006). Thishighlights a role for extracellular proteases in facilitatingthe migration of Acanthamoeba from the systemic circula-tion into the deeper lying tissues of the CNS. In additionto serine proteases, there is evidence for metalloproteaseactivity in Acanthamoeba (Mitro et al., 1994; Alfieri et al.,2000; Sissons et al., 2006). The metalloprotease exhibitedproperties of ECM degradation as evidenced by its activityagainst collagens I and III, elastin (elastic fibrils of ECM),and plasminogen (Sissons et al., 2006). Although Acantha-
moeba proteases have been shown to disrupt HBMEC mon-olayers, they do not appear to induce HBMEC cytotoxicityas determined by lactate dehydrogenase release (a markerfor cell cytotoxicity) (Alsam et al., 2005b; Sissons et al.,2006). Acanthamoeba proteases-mediated HBMEC mono-layer disruptions can be blocked by phenylmethylsulfonylfluoride (PMSF), a serine protease inhibitor, suggestingthe role of serine proteases in blood–brain barrier perturba-tions (Alsam et al., 2005b; Sissons et al., 2006). Further-more, recent studies have shown that Acanthamoeba
proteases target ZO-1 and occludin (Figs. 5 and 6) (S.Alsam, unpublished data). This was demonstrated usinganti-ZO-1 and anti-occludin antibodies in Western blottingassays as previously described (Khan et al., 2002). As indi-cated above, ZO-1 and occludin are involved in the forma-tion of tight junctions, suggesting that Acanthamoeba
disrupt tight junctions to induce HBMEC permeabilityand/or HBMEC monolayer disruptions, leading to blood–brain barrier perturbations.

N.A. Khan / International Journal for Parasitology 37 (2007) 131–138 137
6. Conclusion
Acanthamoeba traversal of the blood–brain barrier is akey step in AGE pathogenesis that most likely occurs atthe sites of the cerebral capillary endothelium. The abilityof Acanthamoeba to bind, phagocytose, induce cell cyclearrest and produce HBMEC death as well as disruptingtight junctions by secreting proteases, suggests that eitherthe transcellular and/or the paracellular route may act aspotential modes of amoebae entry into the CNS. Our under-standing of the molecular mechanisms associated with AGEpathogenesis is beginning to emerge but is still in its infancy.Future studies are required to identify molecular interac-tions between pathogens and hosts. These would help toidentify targets for the rationale development of therapeuticinterventions and/or the design of preventative measures.
Acknowledgements
The author is grateful to Selwa Alsam, James Sissons,Ricky Dudley, Suk-Yul Jung, Ruqaiyyah Siddiqui and Ab-dul Matin, School of Biological and Chemical Sciences,Birkbeck, University of London, England, UK; KwangSik Kim and Monique Stins, Division of Infectious Diseas-es, Johns Hopkins University School of Medicine, Balti-more, MD, USA, for assistance. This work wassupported by grants from the Faculty Research Fund,Central Research Fund, University of London, NuffieldFoundation and The Royal Society.
References
Alfieri, S.C., Correia, C.E., Motegi, S.A., Pral, E.M., 2000. Proteinaseactivities in total extracts and in medium conditioned by Acanthamoe-
ba polyphaga trophozoites. J. Parasitol. 86, 220–227.Alsam, S., Kim, K.S., Stins, M., Rivas, A.O., Sissons, J., Khan, N.A.,
2003. Acanthamoeba interactions with human brain microvascularendothelial cells. Microb. Pathogen. 35, 235–241.
Alsam, S., Sissons, J., Dudley, R., Khan, N.A., 2005a. Mechanismsassociated with Acanthamoeba castellanii (T4) phagocytosis. Parasitol.Res. 96, 402–409.
Alsam, S., Sissons, J., Jayasekera, S., Khan, N.A., 2005b. Extracellularproteases of Acanthamoeba castellanii (encephalitis isolate belonging toT1 genotype) contribute to increased permeability in an in vitro modelof the human blood–brain barrier. J. Infect. 51, 150–156.
Benedetto, N., Auriault, C., 2002a. Prolactin-cytokine network in thedefence against Acanthamoeba castellanii in murine microglia [correct-ed]. Eur. Cytokine Netw. 13, 447–455.
Benedetto, N., Auriault, C., 2002b. Complex network of cytokinesactivating murine microglial cell activity against Acanthamoeba
castellanii. Eur. Cytokine Netw. 13, 351–357.Cerva, L., 1967. Intranasal, intrapulmonary and intracardial inoculation
of experimental animals with Hartmanella castellanii. Folia Parasitol.(Prague) 14, 207–215.
Clarke, D.W., Niederkorn, J.Y., 2006. The immunobiology of Acantha-
moeba keratitis. Microb. Infect. 8, 1400–1405.Culbertson, C.G., Smith, J.W., Cohen, H.K., Minner, J.R., 1959.
Experimental infection of mice and monkeys by Acanthamoeba. Am.J. Pathol. 35, 185–197.
Drevets, D.A., Leenen, P.J.M., 2000. Leukocyte-facilitated entry ofintracellular pathogens into the central nervous system. Microb.Infect. 2, 1609–1618.
Dudley, R., Matin, A., Alsam, S., Sissons, J., Mahsood, A.H., Khan,N.A., 2005. Acanthamoeba isolates belonging to T1, T2, T3, T4 but notT7 encyst in response to increased osmolarity and cysts do not bind tohuman corneal epithelial cells. Acta Trop. 95, 100–108.
Garate, M., Cao, Z., Bateman, E., Panjwani, N., 2004. Cloning andcharacterization of a novel mannose-binding protein of Acanthamoeba.J. Biol. Chem. 279, 29849–29856.
Garate, M., Marchant, J., Cubillos, I., Cao, Z., Khan, N.A., Panjwani, N.,2006. Pathogenicity of Acanthamoeba is associated with the expressionof the mannose-binding protein. Invest. Ophthalmol. Vis. Sci. 47,1056–1062.
Huber, J.D., Egleton, R.D., Davis, T.P., 2001. Molecular physiology andpathophysiology of tight junctions in the blood–brain barrier. Trend.Neurosci. 24, 719–725.
Khan, N.A., Jarroll, E.L., Panjwani, N., Cao, Z., Paget, T.A., 2000.Proteases as markers of differentiation of pathogenic and non-pathogenic Acanthamoeba. J. Clin. Microbiol. 38, 2858–2861.
Khan, N.A., 2001. Pathogenicity, morphology and differentiation ofAcanthamoeba. Curr. Microbiol. 43, 391–395.
Khan, N.A., Wang, Y., Kim, K.J., Woong, J., Wass, C.A., Kim, K.S.,2002. Role of cytotoxic necrotizing factor-1 in Escherichia coli K1invasion into the central nervous system. J. Biol. Chem. 277 (18),15607–15612.
Khan, N.A., 2006. Acanthamoeba: biology and increasing importance inhuman health. FEMS Microbiol. Rev. 30, 564–595.
Kim, J.H., Kim, J.H., Park, J.A., Lee, S., Kim, W.J., Yu, Y.S., Kim, K.,2006. Blood-neural barrier: intercellular communication at glio-vascular interface. J. Biochem. Mol. Biol. 39, 339–345.
Kong, H.H., Kim, T.H., Chung, D.I., 2000. Purification and character-ization of a secretory proteinase of Acanthamoeba healyi isolates fromGAE. J. Parasitol. 86, 12–17.
Marciano-Cabral, F., Cabral, G., 2003. Acanthamoeba spp. as agents ofdisease in humans. Clin. Microbiol. Rev. 16, 273–307.
Marciano-Cabral, F., Ludwick, C., Puffenbarger, R.A., Cabral, G.A.,2004. Differential stimulation of microglial pro-inflammatory cyto-kines by Acanthamoeba culbertsoni versus Acanthamoeba castellanii. J.Eukaryot. Microbiol. 51, 472–479.
Martinez, A.J., Markowitz, S.M., Duma, R.J., 1975. Experimentalpneumonitis and encephalitis caused by Acanthamoeba in mice:pathogenesis and ultrastructural features. J. Infect. Dis. 131, 692–699.
Martinez, A.J., 1985. Free-living Amebas: Natural History, Prevention,Diagnosis, Pathology and treatment of disease. CRC Press, BocaRaton, FL.
Martinez, A.J., 1991. Infections of the central nervous system due toAcanthamoeba. Rev. Infect. Dis. 13, S399–S402.
Martinez, A.J., Visvesvara, G.S., 1997. Free-living, amphizoic andopportunistic amebas. Brain Pathol. 7, 583–598.
Mattana, A., Bennardini, F., Usai, S., Fiori, P.L., Franconi, F.,Cappuccinelli, P., 1997. Acanthamoeba castellanii metabolites increasethe intracellular calcium level and cause cytotoxicity in wish cells.Microb. Pathog. 23, 85–93.
Mattana, A., Cappai, V., Alberti, L., Serra, C., Fiori, P.L., Cappuccinelli,P., 2002. ADP and other metabolites released from Acanthamoeba
castellanii lead to human monocytic cell death through apoptosis andstimulate the secretion of proinflammatory cytokines. Infect. Immun.70, 4424–4432.
Mitro, K., Bhagavathiammai, A., Zhou, O.M., Bobbett, G., McKerrow,J.H., Chokshi, R., Chokshi, B., James, E.R., 1994. Partial character-ization of the proteolytic secretions of Acanthamoeba polyphaga. Exp.Parasitol. 78, 377–385.
Petry, F., Torzewski, M., Bohl, J., Wilhelm-Schwenkmezger, T., Scheid,P., Walochnik, J., Michel, R., Zoller, L., Werhahn, K.J., Bhakdi, S.,Lackner, K.J., 2006. Early diagnosis of Acanthamoeba infection duringroutine cytological examination of cerebrospinal fluid. J. Clin. Micro-biol. 44, 1903–1904.
Pettit, D.A., Williamson, J., Cabral, G.A., Marciano-Cabral, F., 1996. In
vitro destruction of nerve cell cultures by Acanthamoeba spp.: a

138 N.A. Khan / International Journal for Parasitology 37 (2007) 131–138
transmission and scanning electron microscopy study. J. Parasitol. 82,769–777.
Shin, H.J., Cho, M.S., Jung, S.Y., Kim, H.I., Park, S., Seo, J.H., Yoo,J.C., Im, K.I., 2001. Cytopathic changes in rat microglial cells inducedby pathogenic Acanthamoeba culbertsoni: morphology and cytokinerelease. Clin. Diagn. Lab. Immunol. 8, 837–840.
Shuster, F.L., Visvesvara, G.S., 2004. Free-living amoebae as opportu-nistic and non-opportunistic pathogens of humans and animals. Int. J.Parasitol. 34, 1–27.
Sissons, J., Alsam, S., Jayasekera, S., Kim, K.S., Stins, M., Khan, N.A.,2004a. Acanthamoeba induces cell-cycle arrest in the host cells. J. Med.Microbiol. 53, 711–717.
Sissons, J., Alsam, S., Jayasekera, S., Khan, N.A., 2004b. Ecto-ATPasesof clinical and non-clinical isolates of Acanthamoeba. Microb. Pathog.37, 231–239.
Sissons, J., Kim, K.S., Stins, M., Jayasekera, S., Alsam, S., Khan, N.A.,2005. Acanthamoeba castellanii induces host cell death via a phospha-tidylinositol 3-kinase-dependent mechanism. Infect. Immun. 73,2704–2708.
Sissons, J., Alsam, S., Goldsworthy, G., Lightfoot, M., Jarroll, E.L.,Khan, N.A., 2006. Identification and properties of proteases from anAcanthamoeba isolate capable of producing granulomatous encepha-litis. BMC Microbiol. 6, 42.
Stins, M.F., Gilles, F., Kim, K.S., 1997. Selective expression of adhesionmolecules on human brain microvascular endothelial cells. J. Neuro-immunol. 76, 81–90.
Taylor, W.M., Pidherney, M.S., Alizadeh, H., Niederkorn, J.Y., 1995.In vitro characterization of Acanthamoeba castellanii cytopathic effect.J. Parasitol. 81, 603–609.
Thyrell, L., Hjortsberg, L., Arulampalam, V., Panaretakis, T., Uhles, S.,Zhivotovsky, B., Leibiger, I., Grander, D., Pokrovskaja, K., 2004.Interferon-alpha induced apoptosis in tumor cells is mediated throughPI3K/mTOR signalling pathway. J. Biol. Chem. 279, 24152–24162.
Tuomanen, E., 1996. Entry of pathogens into the central nervous system.FEMS Microbiol. Rev. 18, 289–299.
Wolburg, H., Wolburg-Buchholz, K., Liebner, S., Engelhardt, B., 2001.Claudin-1, claudin-2 and claudin-11 are present in tight junction ofchoroids plexus epithelium of the mouse. Neurosci. Lett. 307, 77–80.





























