Embed Size (px)
Citation preview

International Journal for Parasitology 39 (2009) 1611–1616
Contents lists available at ScienceDirect
International Journal for Parasitology
journal homepage: www.elsevier .com/locate / i jpara
Acanthamoeba affects the integrity of human brain microvascular endothelialcells and degrades the tight junction proteins
Naveed Ahmed Khan *, Ruqaiyyah SiddiquiSchool of Veterinary Medicine and Science, University of Nottingham, Sutton Bonington, UK
a r t i c l e i n f o a b s t r a c t
Article history:Received 21 April 2009Received in revised form 11 June 2009Accepted 12 June 2009
Keywords:AcanthamoebaBlood–brain barrierHuman brain microvascular endothelialcellsTight junctionsTransendothelial electrical resistanceRhoAOccludinZonula occludens
0020-7519/$36.00 � 2009 Australian Society for Paradoi:10.1016/j.ijpara.2009.06.004
* Corresponding author. Address: School of VeterUniversity of Nottingham, College Road, Sutton Bonin(0)115 951 6409; fax: +44 (0)115 951 6440.
E-mail address: [email protected] (N
Haematogenous spread is a key step in the development of Acanthamoeba granulomatous encephalitis,however it is not clear how circulating amoebae cross the blood–brain barrier to enter the CNS to producedisease. Using the primary human brain microvascular endothelial cells (HBMEC), which constitute theblood–brain barrier, here it is shown that Acanthamoeba abolishes the HBMEC transendothelial electricalresistance. Using traversal assays, it was observed that Acanthamoeba crosses the HBMEC monolayers.The primary interactions of Acanthamoeba with the HBMEC resulted in increased protein tyrosine phos-phorylations and the activation of RhoA, suggesting host–parasite cross-talk. Furthermore, Western blotassays revealed that Acanthamoeba degraded occludin and zonula occludens-1 proteins in a Rho kinase-dependent manner. Overall, these findings suggest that Acanthamoeba affects the integrity of the mono-layer and traverses the HBMEC by targeting the tight junction proteins.
� 2009 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
1. Introduction
Acanthamoeba granulomatous encephalitis is a rare infectionthat almost always proves fatal. The majority of Acanthamoebaencephalitis cases occur in immunocompromised individuals. Thisinfection is of concern in view of (i) an increasing immunocompro-mised population, and (ii) individuals undergoing immunosup-pressive therapy and excessive use of steroids. Perhaps the mostdistressing aspect is the limited availability of effective treatmentagainst Acanthamoeba granulomatous encephalitis, which at pres-ent has a case fatality rate of more than 90%. The gross pathologyof the autopsied brain often shows severe oedema and haemor-rhagic necrosis and encephalitis (reviewed in Marciano-Cabraland Cabral 2003; Khan, 2006, 2008; Visvesvara et al., 2007).
It is widely accepted that the route of entry for Acanthamoebaincludes the respiratory tract, leading to amoebae invasion of thealveolar blood vessels, followed by haematogenous spread. Skin le-sions may provide direct entry into the bloodstream, bypassing thelower respiratory tract. Acanthamoeba entry into the CNS mostlikely occurs through the blood–brain barrier, which is highly
sitology Inc. Published by Elsevier
inary Medicine and Science,gton LE12 5RD, UK. Tel.: +44
.A. Khan).
selective in regulating the entry of microbes and/or molecules.The olfactory neuroepithelium route (i.e. invasion of the olfactorypart of the nasal epithelium and migration along nerve fibres, fol-lowed by invasion of the olfactory bulb) provides another route ofentry into the CNS and has been studied in experimental models(Culbertson et al., 1959; Cerva, 1967; Martinez, 1985, 1991; Marti-nez and Visvesvara, 1997). It is worth noting that the olfactory partof the nasal mucosa in the rat (rodents) is 50% of the total surfacearea of the nose, whilst in humans, the olfactory part of the nasalmucosa is only 3–8% (Illum, 2000; Graff and Pollack, 2005). Thesmaller fraction of nasal mucosa associated with the olfactory epi-thelium in humans can be associated with a lower likelihood ofthis route leading to human disease. Furthermore, the widespreaddistribution of lesions in the brain, as observed in Acanthamoebagranulomatous encephalitis patients, would fit better with hae-matogenous spread than olfactory access. Acanthamoeba entry intothe CNS most likely occurs through the endothelial lining of thecerebral capillaries (Martinez, 1985, 1991; Martinez and Visvesv-ara, 1997). Hematoxylin–eosin-stained sections of the brain tissueof Acanthamoeba granulomatous encephalitis patients exhibit largenumbers of amoebae in the perivascular space, indicating theinvolvement of the cerebral capillaries as the sites of entry intothe CNS. Affected tissues other than the CNS may include s.c. tis-sue, skin, liver, lungs, kidneys, adrenals, pancreas, prostate, lymphnodes and bone marrow, which further suggest haematogenous
Ltd. All rights reserved.

1612 N.A. Khan, R. Siddiqui / International Journal for Parasitology 39 (2009) 1611–1616
spread. Overall, these findings suggest that unlike Naegleria fowleriwhich uses the olfactory neuroepithelium route, Acanthamoeba in-vades the CNS via the blood–brain barrier. The mechanisms associ-ated with Acanthamoeba traversal of the blood–brain barrier arenot clear. The present study determined the effects of Acantha-moeba on the human brain microvascular endothelial cells thatconstitute the blood–brain barrier. The findings revealed that Acan-thamoeba induced blood–brain barrier perturbations by targetingthe tight junction proteins.
2. Materials and methods
2.1. Culture of Acanthamoeba
All chemicals were purchased from Sigma Laboratories (Poole,Dorset, England), unless otherwise stated. For the present study,Acanthamoeba castellanii belonging to the T4 genotype was ob-tained from the American Type Culture Collection (ATCC 50492).As, based on the 18S rRNA gene sequencing, most of the clinicalisolates of Acanthamoeba (from keratitis, encephalitis and cutane-ous infections) as well as environmental isolates have been typedas the T4 genotype, the aforementioned isolate was used as a rep-resentative of the T4 genotype. Acanthamoeba cultures were grownwithout shaking in 15-ml of PYG medium (proteose peptone 0.75%(w/v), yeast extract 0.75% (w/v) and glucose 1.5% (w/v)) in T-75 tis-sue culture flasks at 30 �C as previously described (Sissons et al.,2005) and media refreshed 17–20 h prior to experiments. Acantha-moeba adhering to flasks represented trophozoite forms and wereused for all assays.
2.2. Human brain microvascular endothelial cell cultures
The primary human brain microvascular endothelial cells(HBMEC) were used as previously described (Stins et al., 1997; Al-sam et al., 2003). Briefly, small fragments of cerebral cortex, de-rived from individuals who had undergone surgical resections forseizure disorder at Johns Hopkins University School of Medicine(USA) were used for HBMEC isolation following approval fromthe University Ethics Committee. The primary endothelial cellswere purified by fluorescent activated cell sorting, and their puritytested using endothelial markers such as expression of F-VIII, car-bonic anhydrase IV and uptake of acetylated low density lipopro-tein (DiI-AcLDL) as previously described (Stins et al., 1997; Alsamet al., 2003) and resulted in >99% pure endothelial cultures. TheHBMEC were routinely grown on rat-tail collagen-coated dishesin complete medium (RPMI-1640 containing 10% heat-inactivatedFBS, 10% Nu-Serum, 2 mM glutamine, 1 mM pyruvate, penicillin(100 U/ml), streptomycin (100 lg/ml), non-essential amino acidsand vitamins) (Invitrogen, Paisley, England).
2.3. Traversal assays
The HBMEC were seeded into the upper chamber of collagen-coated Transwells, in a 24-well Transwell plate (Corning Costar,Corning Ltd., Hemel Hempstead, England), and grown to conflu-ency for at least 2 days in HBMEC culture media as described pre-viously (Stins et al., 2001; Alsam et al., 2005). Following thisincubation, the media were replaced with fresh culture mediacontaining 500 nM hydrocortisone and cultured for an additional3–5 days. The integrity of monolayers was monitored by measur-ing transendothelial electrical resistance (TEER) using a tissue-resistant measurement chamber and a voltohmmeter (EVOM,World Precision Instruments). The HBMEC monolayers exhibitingTEER of more than 200 X/cm2 were subsequently used for tra-versal assays. To determine the effects of Acanthamoeba on TEER,
amoebae were added to the upper chamber of the Transwells(5 � 105 amoebae; total volume 200 ll; counted using a haemocy-tometer) and incubated for up to 8 h. Following this incubation, theintegrity of the HBMEC monolayer was determined by measuringTEER as described above. In addition, the effects of Acanthamoebaconditioned medium (CM) on HBMEC monolayer integrity weredetermined. The CM was produced by incubating Acanthamoeba(5 � 106 amoebae/ml) in RPMI-1640 for 17–20 h. Following thisincubation, cell-free CM were collected by centrifugation and usedfor traversal assays.
To evaluate the ability of Acanthamoeba to cross the HBMECmonolayer, amoebae were added to the upper chamber andallowed to cross the HBMEC monolayer for up to 8 h in the pres-ence or absence of a rho kinase inhibitor, Y27632 (Merck Chemi-cals, UK). Subsequently, 100 ll samples were taken from thebottom chamber and the number of amoebae determined byhaemocytometer counting.
2.4. Western blot assays
To determine the effects of Acanthamoeba on the HBMEC mono-layer tight junction proteins, Western blot assays were performed.Briefly, HBMEC were grown to confluency in six-well plates andmonolayers were incubated with live amoebae (5 � 106 amoe-bae/ml) or their CM for 90 min in the presence or absence of arho kinase inhibitor, Y27632. The CM was produced by incubatingAcanthamoeba (5 � 106 amoebae/ml) in RPMI-1640 for 17–20 h asindicated above. Following this incubation, CM were collected bycentrifugation and incubated with the HBMEC monolayer. For con-trols, HBMEC alone or HBMEC incubated with methanol-fixedamoebae were used. Following incubation, cells were lysed withRIPA lysis buffer (50 mM Tris–HCl (pH 7.4), 0.1% SDS, 0.5%Na-deoxycholate, 10 mM Na-pyrophosphate, 25 mM b-glycero-phosphate, 150 mM NaCl, 2 mM EDTA, 2 mM EGTA, 1% TritonX-100, 1 mM Na3VO4, 50 mM NaF, 1 mM PMSF, 1 lg/ml aprotinin,1 lg/ml leupeptin, 1 lg/ml pepstatin). Samples were centrifuged(10,000g, 4 �C, 6 min), and supernatant collected for proteinquantification (Bio-Rad protein detection kit). Equal amounts ofproteins were electrophoresed using 10% SDS–PAGE under dena-turing conditions. Proteins were transferred onto nitrocellulosemembranes and blocked in 4% blocking buffer (blotting gradenon-fat milk solution; Bio-Rad, Hemel Hempstead, England). Themembranes were blotted with anti-zonula occludens (ZO)-1, andanti-occludin antibodies (Invitrogen), in the blocking buffer over-night at 4 �C, with gentle shaking. The next day, the membraneswere washed and subsequently incubated (60 min, 4 �C) withappropriate horseradish peroxidase-linked secondary antibodies(Cell Signalling Tech, Beverly, MA, USA). Protein bands were visual-ised using an enhanced chemiluminescent detection kit (Amer-sham Biosciences, England).
2.5. GST-rhotekin assays
To determine effects of Acanthamoeba on the activation ofHBMEC RhoA, monolayers were incubated with Acanthamoeba(5 � 106 amoebae) for 90 min, and lysates prepared using RIPAbuffer and protein quantified as described previously (Khan et al.,2002). An equal amount of protein (500 lg to 1 mg) was incubatedwith GST-rhotekin (Upstate Biotech, Lake Placid, NY, USA) for45 min at 4 �C to collect active forms of RhoA, i.e. GTP-RhoA. Theprotein complex was washed with RIPA and resolved by 10%SDS–PAGE and transferred to nitrocellulose membrane. The blotswere blocked with TBST (25 mM Tris, pH 7.4, 150 mM NaCl and0.1% Tween 20) containing 4% skimmed milk for 60 min at 22 �C.After blocking, the membranes were incubated overnight (4 �C)with mouse monoclonal antibody against RhoA (Santa Cruz Bio-

N.A. Khan, R. Siddiqui / International Journal for Parasitology 39 (2009) 1611–1616 1613
tech, Santa Cruz, CA, USA), and subsequently incubated (60 min,22 �C) with horseradish peroxidase-linked secondary antibodyagainst mouse. The antibody bound RhoA was visualised using anenhanced chemiluminescence kit. A single band of RhoA in the21 kDa range was detected. Before performing GST-rhotekin incu-bation, an aliquot of each sample was analysed by Western blot-ting using RhoA-specific antibody showing equal amounts ofproteins in all samples.
3. Results
3.1. Acanthamoeba crosses the HBMEC monolayers
Using an in vitro model of the human blood–brain barrier, Acan-thamoeba crossing of the endothelial barrier was assessed. Amoe-bae were added to the apical side of the HBMEC monolayergrown on Transwell filters and their appearance in the bottomchamber was determined. The live Acanthamoeba traversed theHBMEC monolayers. Approximately 2% of the inoculum(9,375 ± 255 amoebae) were found in the lower chamber within8 h of incubation. In contrast, methanol-fixed amoebae did not tra-verse the endothelial barrier, following addition in the upperchamber.
3.2. Acanthamoeba affects the HBMEC monolayer integrity
The live amoebae have the ability to cross the HBMEC monolay-ers. Next, the integrity of the endothelial monolayer was moni-tored by measuring TEER. Both live Acanthamoeba and CMincubated with HBMEC monolayer abolished the endothelial TEERwithin 8 h (Fig. 1). This is consistent with our previous findingswhich showed that HBMEC incubated with amoebae exhibited in-creased BSA crossing (Alsam et al., 2005). The HBMEC monolayerincubated alone exhibited TEER of approximately 250 X/cm2
(Fig. 1, dotted line), which did not change significantly when incu-bated with methanol-fixed Acanthamoeba (Fig. 1). Notably, amoe-bae incubation with the HBMEC monolayer for 1 h was notassociated with significant changes in the endothelial TEER, whilstAcanthamoeba CM significantly affected the integrity of the HBMEC
Live amoebae Conditionedmedium
Fixed amoebae0
50
100
150
200
250
300
TEER
(Ohm
/cm
2 )
Fig. 1. Acanthamoeba affected the integrity of the human brain microvascularendothelial cell (HBMEC) monolayer. Live amoebae (5 � 105 amoebae; total volume200 ll) and their conditioned medium (CM; produced by incubating amoebaeovernight in RPMI-1640 as described in Section 2) as well as methanol-fixedamoebae (5 � 105 amoebae; total volume 200 ll) were added to the upperchambers of Transwell tissue culture inserts grown with HBMEC monolayers. Theintegrity of the HBMEC monolayer was determined by measuring transendothelialelectrical resistance (TEER) as described in Section 2. Note that HBMEC incubatedalone exhibited TEER of approximately 250 X/cm2 as indicated by dotted lines.Following 8 h incubation, both live Acanthamoeba and CM, but not methanol-fixedamoebae, abolished HBMEC monolayer TEER (P < 0.01, using paired t-test, one-taildistribution). Results represent the average of three independent experimentsperformed in duplicate. Error bars indicate SD.
monolayer within as little as 30 min, as determined by decreasedTEER (data not shown).
3.3. Acanthamoeba and its conditioned medium induced activation ofRhoA
Rho GTPases affect actin cytoskeletal rearrangements by regu-lating actin polymerisation (Caprioli et al., 1983; Flatau et al.,1997; Schmidt et al., 1997). To assess whether Acanthamoeba in-duced RhoA activation, HBMEC were incubated with amoebaeand their CM, and the total lysates were compared for the activatedGTP form of RhoA. Both live Acanthamoeba and the CM were effec-tive in the activation of RhoA GTPase compared with the HBMECincubated alone (Fig. 2). Of note, rho kinase inhibitor, Y27632, at50 lM concentrations, partially inhibited amoebae-mediated TEERchanges and parasite crossing of the HBMEC monolayer (Fig. 2)(P < 0.05 using paired t-test, one-tail distribution).
3.4. Acanthamoeba and the conditioned medium degraded tightjunction proteins
The high transendothelial electrical resistance in the blood–brain barrier is due to the presence of tight junctions. Our previousstudies have shown that amoebae induced degradation of ZO-1and occludin proteins (Khan, 2007). Given that RhoA become acti-vated in response to Acanthamoeba and the CM, HBMEC were incu-bated with live amoebae and their CM in the presence or absenceof rho kinase inhibitor, Y27632. As expected, Acanthamoeba andthe CM target zonula occludens-1 and occludin (Fig. 3), suggestingthat Acanthamoeba disrupts tight junctions to induce HBMEC per-meability and/or HBMEC monolayer disruptions leading to theblood–brain barrier perturbations. Notably the rho kinase inhibi-tor, Y27632, partially inhibited amoebae-mediated tight junctionproteins disruptions (Fig. 3). Under the experimental settings inthe present study, Y27632 had no effect on the viability of amoe-bae or of endothelial cells as determined by Trypan blue exclusionassay (data not shown).
4. Discussion
Several lines of evidence suggest that Acanthamoeba entry intothe brain occurs at the sites of the blood–brain barrier (Martinez,1985, 1991; Martinez and Visvesvara, 1997; Marciano-Cabral andCabral 2003; Khan, 2006, 2008). This is consistent with severalother microbial pathogens that are known to enter the CNS atthe sites of the blood–brain barrier including, Escherichia coli(Kim, 2001, 2003), group B Streptococcus (Nizet et al., 1997), Strep-tococcus pneumoniae (Ring et al., 1998), Listeria monocytogenes(Greiffenberg et al., 1998), Neisseria meningitidis (Unkmeir et al.,2002), Mycobacterium tuberculosis and L. monocytogenes (Drevetset al., 2004; Join-Lambert et al., 2005), fungal pathogens such asCandida albicans (Jong et al., 2001) and Cryptococcus neoformans(Chang et al., 2004); and protozoa including Trypanosoma spp.(Grab et al., 2004), and Balamuthia mandrillaris (Visvesvara et al.,2007; Matin et al., 2008).
However, the mode of blood–brain barrier crossing, i.e. intracel-lular or paracellular, varies between different pathogens. Using anin vitro model of the HBMEC that constitute the blood–brain bar-rier, here it is shown that Acanthamoeba transmigrates the HBMECmonolayer. For some microbial pathogens such as E. coli K1, it isshown that the blood–brain barrier is breached without affectingits integrity (Kim et al., 1992; Stins et al., 2001). A key characteris-tic to measure the integrity of the blood–brain barrier, is that itexhibits high TEER (approximately 2,000 X/cm2) (Pardridge,1999), whilst endothelial cells from the human placenta exhibit
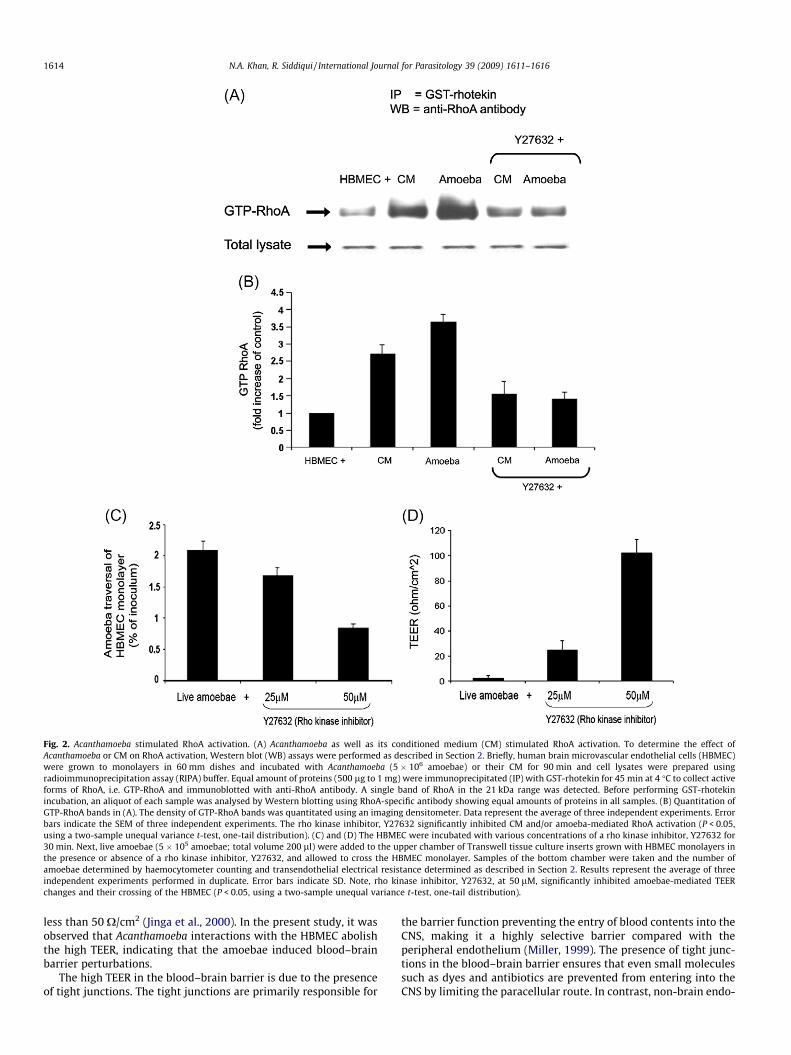
Fig. 2. Acanthamoeba stimulated RhoA activation. (A) Acanthamoeba as well as its conditioned medium (CM) stimulated RhoA activation. To determine the effect ofAcanthamoeba or CM on RhoA activation, Western blot (WB) assays were performed as described in Section 2. Briefly, human brain microvascular endothelial cells (HBMEC)were grown to monolayers in 60 mm dishes and incubated with Acanthamoeba (5 � 106 amoebae) or their CM for 90 min and cell lysates were prepared usingradioimmunoprecipitation assay (RIPA) buffer. Equal amount of proteins (500 lg to 1 mg) were immunoprecipitated (IP) with GST-rhotekin for 45 min at 4 �C to collect activeforms of RhoA, i.e. GTP-RhoA and immunoblotted with anti-RhoA antibody. A single band of RhoA in the 21 kDa range was detected. Before performing GST-rhotekinincubation, an aliquot of each sample was analysed by Western blotting using RhoA-specific antibody showing equal amounts of proteins in all samples. (B) Quantitation ofGTP-RhoA bands in (A). The density of GTP-RhoA bands was quantitated using an imaging densitometer. Data represent the average of three independent experiments. Errorbars indicate the SEM of three independent experiments. The rho kinase inhibitor, Y27632 significantly inhibited CM and/or amoeba-mediated RhoA activation (P < 0.05,using a two-sample unequal variance t-test, one-tail distribution). (C) and (D) The HBMEC were incubated with various concentrations of a rho kinase inhibitor, Y27632 for30 min. Next, live amoebae (5 � 105 amoebae; total volume 200 ll) were added to the upper chamber of Transwell tissue culture inserts grown with HBMEC monolayers inthe presence or absence of a rho kinase inhibitor, Y27632, and allowed to cross the HBMEC monolayer. Samples of the bottom chamber were taken and the number ofamoebae determined by haemocytometer counting and transendothelial electrical resistance determined as described in Section 2. Results represent the average of threeindependent experiments performed in duplicate. Error bars indicate SD. Note, rho kinase inhibitor, Y27632, at 50 lM, significantly inhibited amoebae-mediated TEERchanges and their crossing of the HBMEC (P < 0.05, using a two-sample unequal variance t-test, one-tail distribution).
1614 N.A. Khan, R. Siddiqui / International Journal for Parasitology 39 (2009) 1611–1616
less than 50 X/cm2 (Jinga et al., 2000). In the present study, it wasobserved that Acanthamoeba interactions with the HBMEC abolishthe high TEER, indicating that the amoebae induced blood–brainbarrier perturbations.
The high TEER in the blood–brain barrier is due to the presenceof tight junctions. The tight junctions are primarily responsible for
the barrier function preventing the entry of blood contents into theCNS, making it a highly selective barrier compared with theperipheral endothelium (Miller, 1999). The presence of tight junc-tions in the blood–brain barrier ensures that even small moleculessuch as dyes and antibiotics are prevented from entering into theCNS by limiting the paracellular route. In contrast, non-brain endo-

Fig. 3. Acanthamoeba degraded tight junction proteins. To determine the effect ofAcanthamoeba or conditioned medium (CM) tight junction proteins, Western blot(WB) assays were performed as described in Section 2. Briefly, human brainmicrovascular endothelial cells (HBMEC) were grown to monolayers in 60 mmdishes, incubated with various concentrations of a rho kinase inhibitor, Y27632, for30 min prior to incubation with Acanthamoeba (5 � 106 amoebae) or their CM for90 min, in the presence or absence of a rho kinase inhibitor, Y27632, and cells werelysed by radioimmunoprecipitation assay (RIPA) lysis buffer. Equal amounts ofproteins were used for Western blot assays using anti-zonula occludens (ZO)-1, andanti-occludin antibodies. Note that both live amoebae and their CM induced loss ofZO-1 and occludin within 90 min, which was partially blocked with rho kinaseinhibitor, Y27632. Results are representative of three independent experiments.
Acanthamoeba
MBP-mediatedadhesion
Serine proteases
Rho activation
Rho kinase
P-MLCMLC
phosphatase
Redistribution/alterationof ZO-1 & occludin
Tight junctionpermeability increase
Fig. 4. Diagrammatic representation of Acanthamoeba-mediated Rho activation viaserine proteases and/or mannose-binding protein (MBP), leading to blood–brainbarrier permeability changes. RhoA activation induces myosin light-chain (MLC)phosphorylation (P-MLC) via Rho kinase causing redistribution/alteration of tightjunction proteins, zonula occludens-1 (ZO-1) and occludin, resulting in theelevation of barrier permeability.
N.A. Khan, R. Siddiqui / International Journal for Parasitology 39 (2009) 1611–1616 1615
thelium is more permissive. The tight junctions consist of integralmembrane proteins (i.e. occludin, claudin, junction adhesion mol-ecule, endothelial cell-selective adhesion molecules), and cytoplas-mic proteins ZO-1, ZO-2 and ZO-3. Occludin is a phosphoproteinand its cytoplasmic domain is directly associated with zonula pro-teins. Claudins bind to claudins on the adjacent endothelial cells toform the tight junctions. The carboxy termini of claudins binds tothe cytoplasmic ZO proteins. Claudin-5 and claudin-3 are localisedin the tight junctions of the brain endothelial cells. Junctionaladhesion molecules and the endothelial cell-selective adhesionmolecules are also localised in the tight junctions of the blood–brain barrier. The accessory proteins of the ZOs provide structuralsupport and bind to integral membrane proteins of the tight junc-tions, further linking membrane proteins to the actin cytoskeleton(Tuomanen, 1996; Huber et al., 2001; Kim et al., 2006). It is inter-esting that Acanthamoeba CM degraded occludin and ZO proteinssuggesting that amoebae target the paracellular route to disruptthe blood–brain barrier. In addition, the present studies showedthat Acanthamoeba CM induces activation of Rho-associated intra-cellular signalling cascades in HBMEC. Rho-associated pathwayscould disturb the function of tight junctions, thus leading to the in-creased blood–brain barrier permeability. For example, recentstudies in intestinal epithelial cells have shown that RhoA regu-lates phosphorylation of myosin light-chain phosphorylations,causing structural changes and redistribution of ZO-1 and occludinleading to increased blood–brain barrier permeability (Shen et al.,2006). Based on other eukaryotic cells, a speculative diagram onthe activation of RhoA leading to the blood–brain barrier perturba-tions is depicted in Fig. 4.
Although the precise molecular mechanisms by which Acantha-moeba traverses the blood–brain barrier remain incompletelyunderstood, it is thought that they are likely to involve both para-site (adhesins, proteases, phospholipases) as well as host determi-nants (IL-beta, IL-alpha, TNF-alpha, IFN-gamma, host cellapoptosis). The overall outcome is increased permeability and/or
apoptosis of the brain endothelial cells, which promote blood–brain barrier disruptions leading to the parasite invasion of theCNS, a key step in the development of granulomatous encephalitis.The understanding of the cross-talk in parasite–host interactions,as well as between the blood–brain barrier and the CNS in diseasewill provide insights to explain the neuropathogenesis and mayhelp develop novel therapeutic interventions.
Acknowledgements
The authors are grateful to Selwa Alsam, University of Bedford-shire, England, UK; and Kwang Sik Kim and Monique Stins, Divisionof Infectious Diseases, Johns Hopkins University School of Medi-cine, Baltimore, MD, USA for support and helpful discussions. Thiswork is partly funded by the University of Nottingham, UK.
References
Alsam, S., Kim, K.S., Stins, M., Rivas, A.O., Sissons, J., Khan, N.A., 2003. Acanthamoebainteractions with human brain microvascular endothelial cells. Microb.Pathogen. 35, 235–241.
Alsam, S., Sissons, J., Jayasekera, S., Khan, N.A., 2005. Extracellular proteases ofAcanthamoeba castellanii (encephalitis isolate belonging to T1 genotype)contribute to increased permeability in an in vitro model of the humanblood–brain barrier. J. Infect. 51, 150–156.
Caprioli, A., Falbo, V., Roda, L.G., Ruggeri, F.M., Zona, C., 1983. Partial purification andcharacterization of an Escherichia coli toxic factor that induces morphologicalcell alterations. Infect. Immun. 39, 1300–1306.
Cerva, L., 1967. Intranasal, intrapulmonary and intracardial inoculation ofexperimental animals with Hartmanella castellanii. Folia Parasitol. 14, 207–215.
Chang, Y.C., Stins, M.F., McCaffery, M.J., Miller, G.F., Pare, D.R., Dam, T., Paul, M., Kim,K.S., Kwon-Chung, K.J., 2004. Cryptococcal yeast cells invade the central nervoussystem via transcellular penetration of the blood–brain barrier. Infect. Immun.72, 4985–4995.
Culbertson, C.G., Smith, J.W., Cohen, H.K., Minner, J.R., 1959. Experimental infectionof mice and monkeys by Acanthamoeba. Am. J. Pathol. 35, 185–197.
Drevets, D.A., Leenen, P.J., Greenfield, R.A., 2004. Invasion of the central nervoussystem by intracellular bacteria. Clin. Microbiol. Rev. 17, 323–347.
Flatau, G., Lemichez, E., Gauthier, M., Chardin, P., Paris, S., Fiorentini, C., Boquet, P.,1997. Toxin-induced activation of the G protein p21 Rho by deamidation ofglutamine. Nature 387, 729–733.
Grab, D.J., Nikolskaia, O., Kim, Y.V., Lonsdale-Eccles, J.D., Ito, S., Hara, T., Fukuma, T.,Nyarko, E., Kim, K.J., Stins, M., Delannoy, M., Rodgers, J., Kim, K.S., 2004. African

1616 N.A. Khan, R. Siddiqui / International Journal for Parasitology 39 (2009) 1611–1616
trypanosome interactions with an in vitro model of the human blood–brainbarrier. J. Parasitol. 90, 970–979.
Graff, C.L., Pollack, G.M., 2005. Nasal drug administration: potential for targetedcentral nervous system delivery. J. Pharm. Sci. 94, 1187–1195.
Greiffenberg, L., Goebel, W., Kim, K.S., Kuhn, M., 1998. Interaction of Listeriamonocytogenes with human brain microvascular endothelial cells: InlB-dependent invasion, long-term intracellular growth and spread frommacrophages to endothelial cells. Infect. Immun. 66, 5260–5267.
Huber, J.D., Egleton, R.D., Davis, T.P., 2001. Molecular physiology andpathophysiology of tight junctions in the blood–brain barrier. TrendsNeurosci. 24, 719–725.
Illum, L., 2000. Transport of drugs from the nasal cavity to the central nervoussystem. Eur. J. Pharm. Sci. 11, 1–18.
Jinga, V.V., Gafencu, A., Antohe, F., Constantinescu, E., Heltianu, C., Raicu, M.,Manolescu, I., Hunziker, W., Simionescu, M., 2000. Establishment of a purevascular endothelial cell line from human placenta. Placenta 21, 325–336.
Jong, A.Y., Stins, M.F., Huang, S.H., Kim, K.S., 2001. Traversal of C. albicans acrosshuman blood–brain barrier in vitro. Infect. Immun. 69, 4536–4544.
Join-Lambert, O.F., Ezine, S., LeMonnier, A., Jaubert, F., Okabe, M., Berche, P., Kayal,S., 2005. Listeria monocytogenes-infected bone marrow myeloid cells promotebacterial invasion of the central nervous system. Cell. Microbiol. 7, 167–180.
Khan, N.A., Wang, Y., Kim, K.J., Chung, J.W., Wass, C.A., Kim, K.S., 2002. Cytotoxicnecrotizing factor-1 contributes to Escherichia coli K1 invasion of the centralnervous system. J. Biol. Chem. 277, 15607–15612.
Khan, N.A., 2006. Acanthamoeba: biology and increasing importance in humanhealth. FEMS Microbiol. Rev. 30, 564–595.
Khan, N.A., 2007. Acanthamoeba invasion of the central nervous system. Int. J.Parasitol. 37, 131–138.
Khan, N.A., 2008. Acanthamoeba and the blood–brain barrier: the breakthrough. J.Med. Microbiol. 57, 1051–1057.
Kim, J.H., Kim, J.H., Park, J.A., Lee, S., Kim, W.J., Yu, Y.S., Kim, K., 2006. Blood–neuralbarrier: intercellular communication at glio-vascular interface. J. Biochem. Mol.Biol. 39, 339–345.
Kim, K.S., Itabashi, H., Gemski, P., Sadoff, J., Warren, R.L., Cross, A.S., 1992. The K1capsule is the critical determinant in the development of Escherichia colimeningitis in the rat. J. Clin. Invest. 90, 897–905.
Kim, K.S., 2001. E. coli translocation at the blood–brain barrier. Infect. Immun. 69,5217–5222.
Kim, K.S., 2003. Neurological diseases: pathogenesis of bacterial meningitis: frombacteremia to neuronal injury. Nat. Rev. Neurosci. 4, 376–385.
Marciano-Cabral, F., Cabral, G., 2003. Acanthamoeba spp. as agents of disease inhumans. Clin. Microbiol. Rev. 16, 273–307.
Martinez, A.J., 1985. Free-living Amebas: Natural History, Prevention, Diagnosis,Pathology and Treatment of Disease. CRC Press, Boca Raton, FL.
Martinez, A.J., 1991. Infections of the central nervous system due to Acanthamoeba.Rev. Infect. Dis. 13, S399–S402.
Martinez, A.J., Visvesvara, G.S., 1997. Free-living, amphizoic and opportunisticamebas. Brain Pathol. 7, 583–598.
Matin, A., Siddiqui, R., Jayasekera, S., Khan, N.A., 2008. Increasing importance ofBalamuthia mandrillaris. Clin. Microbiol. Rev. 21, 435–448.
Miller, D.W., 1999. Immunobiology of the blood–brain barrier. J. Neurovirol. 5, 570–578.
Nizet, V., Kim, K.S., Stins, M., Jonas, M., Nguyen, D., Rubens, C.E., 1997. Invasion ofbrain microvascular endothelial cells by group B Streptococci. Infect. Immun. 65,5074–5081.
Pardridge, W.M., 1999. Blood–brain barrier: biology and methodology. J. Neurovirol.5, 556–569.
Ring, A., Weiser, J.N., Tuomanen, E.I., 1998. Pneumococcal trafficking across theblood–brain barrier. Molecular analysis of a novel bi-directional pathway. J.Clin. Invest. 102, 347–360.
Schmidt, G., Sehr, P., Wilm, M., Selzer, J., Mann, M., Aktories, K., 1997. Gln 63 of Rhois deamidated by Escherichia coli cytotoxic necrotizing factor-1. Nature 387,725–729.
Shen, L., Black, E.D., Witkowski, E.D., Lencer, W.I., Guerriero, V., Schneeberger, E.E.,Turner, J.R., 2006. Myosin light chain phosphorylation regulates barrier functionby remodeling tight junction structure. J. Cell. Sci. 119, 2095–2106.
Sissons, J., Kim, K.S., Stins, M., Jayasekera, S., Alsam, S., Khan, N.A., 2005.Acanthamoeba castellanii induces host cell death via a phosphatidylinositol 3-kinase-dependent mechanism. Infect. Immun. 73, 2704–2708.
Stins, M.F., Gilles, F., Kim, K.S., 1997. Selective expression of adhesion molecules onhuman brain microvascular endothelial cells. J. Neuroimmunol. 76, 81–90.
Stins, M.F., Badger, J., Kim, K.S., 2001. Bacterial invasion and transcytosis intransfected human brain microvascular endothelial cells. Microb. Pathog. 30,19–28.
Tuomanen, E., 1996. Entry of pathogens into the central nervous system. FEMSMicrobiol. Rev. 18, 289–299.
Unkmeir, A., Latsch, K., Dietrich, G., Wintermeyer, E., Schinke, B., Schwender, S., Kim,K.S., Eigenthaler, M., Frosch, M., 2002. Fibronectin mediates Opc-dependentinternalization of Neisseria meningitidis in human brain microvascularendothelial cells. Mol. Microbiol. 46, 933–946.
Visvesvara, G.S., Moura, H., Schuster, F.L., 2007. Pathogenic and opportunistic free-living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri,and Sappinia diploidea. FEMS Immunol. Med. Microbiol. 50, 1–26.





























