Embed Size (px)
Citation preview

Abundance and geochemical significance of C2n dialkylalkanesand highly branched C3n alkanes in diverse Meso- and
Neoproterozoic sediments.
Paul F. Greenwooda,*, Khaled R. Arourib,Graham A. Logana,c, Roger E. Summonsc,d
aGeoscience Australia, GPO Box 378, Canberra, ACT, 2601. AustraliabGeology and Geophysics, School of Earth and Environmental Sciences, University of Adelaide, SA 5005, Australia
cAustralian Centre for Astrobiology, Macquarie University, NSW 2109, AustraliadDeparment of Earth Atmospheric and Planetary Sciences, Massachusetts Institute of Technology, Cambridge MA 02139-4307, USA
Received 20 March 2003; accepted 24 October 2003
(returned to author for revision 27 June 2003)
Abstract
Several series of branched alkanes with quaternary carbon centers have been detected by GC–MS in the saturatedhydrocarbon fractions of Neoproterozoic sedimentary rocks associated with fossilised microbial mats (Officer Basin,South Australia) and of shallow water carbonates (Spitsbergen, East Greenland and Baffin Island, Canada). A series of5,5-diethylalkanes was predominant in most of these sediments and was typically accompanied by related series in
lower abundance which, based on mass spectral characteristics, are tentatively assigned as 6,6-diethylalkanes, 5-butyl,5-ethylalkanes and 6-butyl, 6-ethylalkanes. Each series displays strong odd or even carbon number preferences. Severalof the sediments contained additional series of structurally undefined C3n alkanes, notable for very negative retention
time factors which become more negative with increasing molecular weight—indicating that branching increases witheach C3 addition. The strongly sequential carbon preference of the respective C2n and C3n branched alkane series, and apredominantly light 13C signature of the 5,5-diethylalkanes (cf. co-occurring n-alkanes), are consistent with an origin
from organisms with a distinct physiology and/or carbon source. The relatively wide occurrence and high abundance ofthese branched alkanes in Proterozoic sediments containing benthic microbial mats suggests a relationship with specificpaleoenvironmental conditions, for example strong water column redox gradients, that may have been more prevalentthan in the Phanerozoic.
# 2003 Elsevier Ltd. All rights reserved.
1. Introduction
The organic matter content of Neoproterozoicsediments has revealed useful information regarding
microbial diversity (Summons and Walter, 1990) and
paleoenvironments (Logan et al., 1997, 1999, 2001) in
the period of rapid and dramatic global change leadingup to the Phanerozoic. Many of the hydrocarbonsencountered in the Neoproterozoic are the same as
those found in Phanerozoic rocks and oils and, for themost part, their precursor biolipids can be found inextant organisms, establishing a direct ‘biomarker’ link.However, some classes of compounds, for example the
C20+ mid-chain methyl alkanes that are so abundant inProterozoic oils from Oman (e.g. Hold et al., 1999) andSiberia (Fowler and Douglas, 1987), have no generally
0146-6380/$ - see front matter # 2003 Elsevier Ltd. All rights reserved.
doi:10.1016/j.orggeochem.2003.10.013
Organic Geochemistry 35 (2004) 331–346
www.elsevier.com/locate/orggeochem
* Corresponding author at present address: School of
Applied Chemistry, Curtin University of Technology, PO Box
U 1987, Perth, WA, 6845, Australia.
E-mail address: [email protected]
(P.F. Greenwood).

recognized counterparts in modern biota. Despite thesegaps in knowledge, studies of these ancient sedimentsand their molecular fossils are important because theymay provide insights into unusual sedimentary environ-
ments and microbiota in the period leading up to thefirst appearance of complex multicellular organisms.High concentrations of unusual branched alkanes
were reported to occur in Neoproterozoic sedimentsfrom the Tanana Formation of the Officer Basin, Aus-tralia (Arouri et al., 2000a,b). Whilst the exact form and
identity of these biomarkers was not established, theyappeared to be directly associated with fossilised micro-bial mat communities that occur in the same sediments.
Microbial mat communities, particularly those wherecyanobacteria are the predominant organisms, are wellknown for having distinctive patterns of short-chain(C15–C20) methyl alkanes. These same hydrocarbons
have been identified in cyanobacterial cultures (Kosteret al., 1999; Dembitsky, et al., 2001), as well as modernand ancient sediments associated with cyanobacterial
mat assemblages (Shiea et al., 1990, 1991; Robinson andEglinton, 1990; Summons and Walter, 1990; Kenig etal., 1995). Neoproterozoic sediments throughout the
Australian Centralian Superbasin are well-preservedand typically contain evidence of fossilised cyano-bacteria and protists (Schopf, 1968; Grey, 1998). Asso-
ciated bitumens are also often rich in low molecularweight organic remains of ancient algae and bacteria(Summons and Walter, 1990; Logan et al., 1997, 1999).A series of monomethyl alkanes and a series of C19–
C33 branched alkanes with a strong odd carbon num-ber preference that were originally mistakenly assignedas 3,7-dimethylalkanes on the basis of similar GC and
MS data to published literature (Mycke et al., 1988),have been reported from ancient microbial mats in the1640 Ma Barney Creek Formation in Australia, (Logan
et al., 2001). A separate series of mid-chain branchedalkanes, consisting predominantly of even carbonnumbers ranging from C22 to C36 were also found inBarney Creek sediments in association with an assem-
blage of large filamentous microfossils (Logan et al.,2001). The much broader range of carbon numbers ofthese unusually prominent methyl alkanes and their
carbon isotopic signatures suggest that they may derivefrom a non-cyanobacterial source.The present study was initiated in an attempt to
identify the unusual branched hydrocarbon productsdetected in the Tanana Formation sediments by Arouriet al. (2000a,b) through close examination of mass
spectral data and gas chromatographic elution patterns.Their distribution in co-eval sediments from the North-ern Hemisphere associated with well-documentedmicrobial organic matter types was investigated by
analysis of sediments from 700 to 800MaNeoproterozoiccarbonate successions of Svalbard and East Greenlandand in the Victor Bay Formation of Baffin Island,
Canada. Where chromatographic resolution allowed,the d13C composition of several hydrocarbons was alsomeasured.
2. Experimental
2.1. Sample descriptions
2.1.1. Tanana formation
Drill core intersections of the Neoproterozoic TananaFormation in the Officer Basin of South Australia wereoriginally sampled for organic geochemical analyses of
large and well-preserved leiospheroid and acantho-morph acritarchs (Arouri et al., 1999,2000b). In addi-tion to the acritarchs, sediments from these locationsoccasionally contained large filamentous microfossils
and microbial mats in various states of preservation.Palynological examination and scanning electron
microscopy showed that the mats in these sediments
were biologically diverse (Arouri et al., 2000b). Abun-dant filamentous and coccoid microfossils, together withamorphous and finely disseminated organic matter
associated with microbially-formed fine carbonate crys-tals characterize the mat horizons. These features, toge-ther with varied biomarker patterns were used to
categorise several distinct organic facies (Arouri et al.,2000a). The prominent series of branched alkanes weredetected in Microbial Mat Type 1 and Type 2 facies.Petrographic analysis indicated the Tanana Formation
sediments had thermal alteration index 42.8 consistentwith an early oil window maturity (Arouri et al., 2000b).
2.1.2. Neoproterozoic carbonate successions ofSpitsbergen (Svalbard) and East GreenlandA suite of samples representing different formations,
lithologies and biofacies were provided by ProfessorAndrew Knoll, Harvard University and have been thesubject of detailed sedimentologic, micropaleontologicand chemostratigraphic analysis (e.g. Knoll et al.,
1986a,b,1990,1991; Green et al., 1987,1988; Swett andKnoll, 1989). Geological and palynological data of thesesamples are shown in Table 1 along with Knoll’s sample
keys and description.The Spitsbergen and East Greenland sediments from
the 700 to 800 Ma Neoproterozoic carbonate succes-
sions are renowned for their distinctive benthic micro-fossils (Green et al., 1987, 1988; Knoll et al., 1986a). Inparticular, the upper Eleonore Bay Group, which com-
prises about 4000 m of sandstones, shales and carbo-nates, contains an unusual fossil assemblage in a 1200 munit known as the Limestone-Dolomite Series, or LDS(Green et al., 1988). Within the LDS, Bed 18 consists of
oolite and pisolite beds 10–50 cm thick intercalated withother sediments and with a range of sedimentologicalfeatures suggesting a shallow, sub-tidal to intertidal
332 P.F. Greenwood et al. / Organic Geochemistry 35 (2004) 331–346

coastal regime. There are mat clasts, stromatolites andflake conglomerates as well as fossiliferous ooids and
pisoids. Cyanobacterial fossils represented by fila-mentous and coccoid forms dominate although thereare also protists such as Melanocyrillium present. Of
particular note are numerous epilithic, interstitial andendolithic taxa that colonised the sediments, some ofwhich (e.g. Polybessurus bipartitus) resemble forms thatcan be found today in the Bahamas (Green et al., 1987).
2.1.3. Victor Bay Formation, Baffin Island (Canada)The Victor Bay Formation occurs within the thick
(�6000 m), and relatively undeformed, sedimentarypackage of the Bylot Supergroup on Northern BaffinIsland and nearby Bylot Island (Kah et al., 1999). The
ages of these rock units are not well constrained but,based on regional correlations and recent Pb isotopegeochronology dating of sedimentary carbonates, a
depositional age of approximately 1200 Ma has beensuggested (Kah et al., 2001). The Victor Bay Formationtogether with the underlying Society Cliffs and overlyingAthole Point formations, comprise �1500 m of pre-
dominantly platform, shelf and slope carbonates. TheVictor Bay Formation itself contains a variety of car-bonate and siliclastic lithologies representing peritidal
shelf to subtidal shelf and slope environments. Unlikethe younger sediments from Svalbard and East Green-
land, the Baffin Island units are fossil-poor and havecarbonates with d13C values in a relatively narrow rangebetween +3.5 and �1.0, lacking the extreme excursions
that characterize the Neoproterozoic. Two samples,AGSO #4193 and AGSO #4194, from the Victor BayFormation were analysed (Table 1).
2.2. GC–MS analysis
Saturated hydrocarbon fractions were obtained using
standard Soxhlet extraction (i.e. 93:7 dichloro-methane:methanol, 72 hrs) and column chromato-graphy procedures (i.e. petroleum ether eluant on silica/
alumina stationary phase) and analysed by GC–MSusing full scan and selected ion recording methods. Sum-mons et al. (1995) and references therein give further
details of procedure for the isolation of the aliphatic frac-tion. The GC–MS analyses were performed using either(1) a Hewlett-Packard 5890II gas chromatograph equip-ped with a split/splitless injector and a 25 m� 0.32 mm i.d
� 0.52 mm film thickness DB5 column interfaced to anUltimaQ Autospec double focusing mass spectrometer, or(2) an Agilent 6890 GC equipped with an on-column
Table 1
Sample details for sediments from Neoproterozoic carbonate successions of Spitsbergen, East Greenland and Baffin Island samples
Sample nos.
Well/formation L ithology and biofaciesAGSO #3912
86-P-13 Draken T idal flat carbonates, muds, microbial mats with occasional thin shales. Mats depositedduring relatively quiet periods have been ripped up and washed by storms into adjacent
lagoons and then silicified (Knoll and Swett, 1990; Knoll et al., 1991). Diverse and
well-preserved microflora
AGSO #3913
86-P-152 Elbobreen B lack shale near lower boundary of formation. Chuaria compressions and vase-shapedmicrofossils. This is a relatively deep basinal facies associated with maximum flooding.
AGSO #3914
86-P-105Backlundtoppen
P
d
artially silicified laminated dolomicrite, within predominantly oolitic unit from a storm
ominated shelf environment (Knoll and Swett, 1990)
AGSO #3915
86-P-9 Draken B lack chert nodules in dolomitic micrite. part of peritidal facies packageAGSO #3916
86-M-43 Elbobreen B lack shale, part of shallowing upward series above basal flooding event (86-P-152, above);still below fairweather wave base
AGSO #3917
86-F-5 Kingsbreen B lack shale in siliciclastic, storm-dominated shelf successionAGSO #3919
86-G-68 Svanbergfjellet T he sample is a silcified tidal flat microbialite from the lower part of the formationAGSO #3920
86-P-42 Backlundtoppen L ower half of the formation in oolite dominated, upper half mostly stromatolites, withflooding by siliciclastics at top
AGSO #3921
83-E-19-5 Bed 18, B ed 18 of Limestone–Dolomite Series (Green et al., 1987). Silicified dolosiltite—interbedded with fossiliferous oolites
AGSO # 3922
86-P-66 Grusdievbreen B lack calcisiltite/micrite in carbonate dominated inner shelf successionAGSO #3923
86-G-13 Svanbergfjellet S ilicified black microbially laminated limestone—peritidal to inner shelf in the lowerSvanbergfjellet Fm
AGSO #3924
86-ST-5 Tokammane B lack shale, near maximum flooding above phosphatic condensed interval—mid-shelfenvironment, with an abundance of acritarchs (Knoll and Swett, 1987)
AGSO #4185
P-6450 Elbobreen C alcareous black shale just below oolitic dolostone, near top of shallowing upwardElbobreen sequence, noted above
AGSO #4193
87-SS-172 Victor Bay B oth samples are thinly intercalated black shales and flat, mechanically laminated micritesand calcisiltites deposited in deepening upward subtidal shelf environment (Kah et al., 1999)
AGSO #4194 87-SS-225 Victor BayP.F. Greenwood et al. / Organic Geochemistry 35 (2004) 331–346 333

injector and a 60 m � 0.32 mm i.d. � 0.25 mm filmthickness DB1 column interfaced to an Agilent 5973mass selective detector (MSD). A typical gas chromato-graph oven temperature program was an initial 40 �C
(held for 2 min) raised at 2–6 �C/min to 300 �C (held for25 min). Mass spectral parameters included a full scanmass range of 50–550 Da or selected ion data for 14n+1
Da, where n=6–14 (i.e., 85, 99. . . 197 Da), an electronenergy of 70 eV and scan speeds 41 s/decade.
3. Results and discussion
At least two different types of methyl alkane serieswere detected in the Neoproterozoic-aged sedimentsfrom each of the represented formations. The totalion chromatogram (TIC) and several reconstructed ion
chromatograms of the saturate fraction of a TananaFormation sample (1234 m deep) from the Munta-1 wellare shown in Fig. 1. Microscopic and molecular
analyses distinguished three distinct organic facies inTanana sediments (Arouri et al., 2000a), and the 1234m, Munta-1 sample is representative of Microbial Mat
Facies 2 (MMF2). The TIC comprises a large unre-solved complex mixture (UCM) from which resolvedC16–C20 n-alkanes are only partially distinct from the
baseline (Fig. 1a) due to their relatively low abundance.The major resolved compounds in the TIC form twodistinctive series which progress by addition of C3 units,from C18 to at least C30. These unusual C3n hydro-
carbons (series E and F) elute at much earlier retentiontimes than n-alkanes of equivalent carbon number. Sev-eral methyl alkane series with strong even or odd carbon
number predominance (i.e., C2n series A–D) were alsodetected and resolved by subtraction of related ion pairs(Fig. 1b–e).
Hydrocarbons of series A–E, but not F, were detectedwith varied relative abundances in the Spitsbergen,E. Greenland and Baffin Island sediments.
3.1. GC–MS features of series A–D branched alkanes
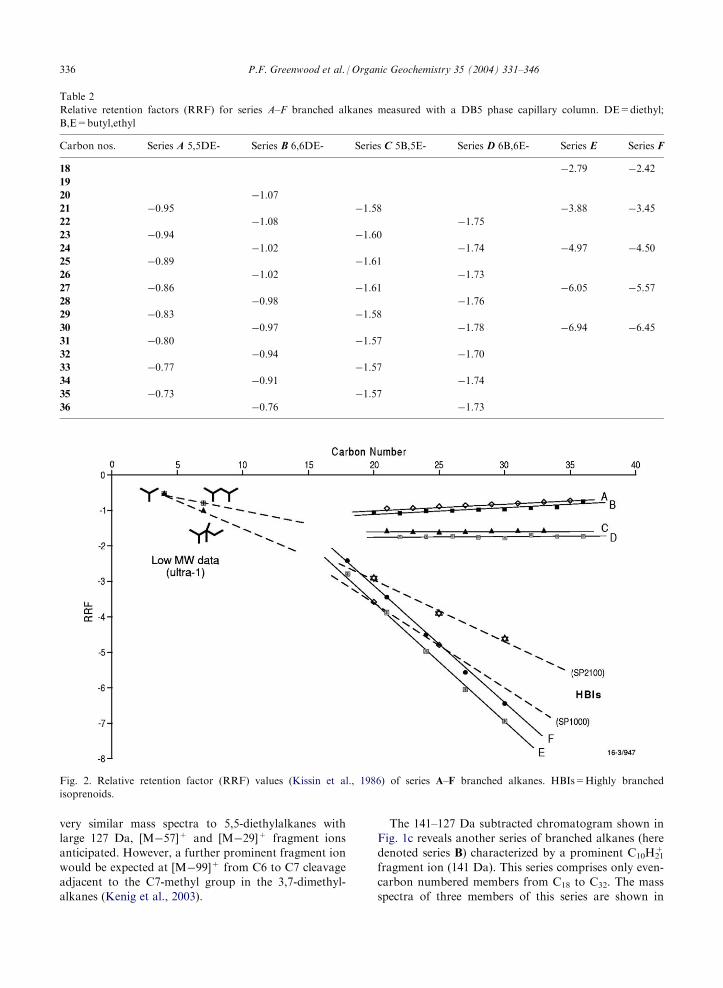
3.1.1. Relative retention factors
Relative retention factors (RRF), described by Kissinet al. (1986), provide a numerical representation of theGC elution characteristics of branched alkanes com-
pared to n-alkanes of the same carbon number. They arecalculated by the following equation:
RRFðXnÞ ¼KFXn�nð100Þ
100
Where,
KFðXnÞ ¼ 100ðn� 1Þ þ100ðRTXn � RTCn�1Þ
RTCn � RTCn�1
RT=retention time, X=branched alkane; Cn=normalalkane with n carbons.The RRF values of series A–D as measured on a DB5
column are listed in Table 2 and plotted in Fig. 2. The
negative values reflect their early elution compared withn-alkanes of the same carbon number and this can beattributed to alkyl-branching which reduces the struc-
tural volume of the hydrocarbon and the interaction ithas with the GC column phase. The hydrocarbonswithin each A–D series show a linear trend of RRF
values indicative of homologous or pseudo-homologousseries. Series C (��1.59) and D (��1.75) have notice-ably more negative RRF values than A (��0.85) and B
(��0.95) indicating a relatively higher degree ofbranching in the former (Fig. 2).
3.1.2. Mass spectral characteristics
The mass spectra of several members of series A
are shown in Fig. 3. The carbon number of these(and series B–F) hydrocarbons was assigned on the
basis of a corresponding odd electron/even mass par-ent typically detected in very low abundance. Theseries A hydrocarbons are characterized by a major
peak at 127 Da (C9H19+). This ion is consistently
higher in abundance than the 113 Da ion, an unusualfeature for acyclic aliphatic hydrocarbons which for-
tuitously provides for selective recognition of the ser-ies via the 127–113 Da subtracted ion chromatogram(Fig. 1b).The chromatographic and mass spectral data repor-
ted here are almost identical to those published for aseries of odd carbon numbered alkanes detected insediments of various ages and paleoenvironments (see
Table 3). Largely based on mass spectral interpretation,these compounds were consistently and incorrectlyassigned as 3, 7-dimethylalkanes (Structure I, Appendix)
or 3, (o-8)-dimethylalkanes (e.g., Dill et al., 1988; Fla-viano et al., 1994). Closer consideration of their massspectra and the synthesis and analytical correlation ofone member of the series by Kenig et al. (2002, 2003)
demonstrated that the previous characterizations wereerroneous and that the series are actually 5,5-diethyl-alkanes (II).
The quaternary carbon containing fragment (C9H19+)
from the 5, 5-diethylalkanes (II) gives rise to an abun-dant 127 Da ion, and the presence of two ethyl groups
accounts exactly for the [M�29]+ ion (where M=par-ent ion MW) being twice as abundant as the [M�57]+
fragment ion (i.e. only one butyl group). Furthermore,
whilst hydrogen rearrangement can accompany frag-mentation adjacent to a quaternary carbon, it does nottypically occur to such an extent that the even mass/oddelectron fragment ion is of similar or higher abundance
to its corresponding odd mass/even electron fragmention at plus one Da (Kenig et al., 2002). The previouslyproposed 3,7-dimethylalkanes would probably yield
334 P.F. Greenwood et al. / Organic Geochemistry 35 (2004) 331–346
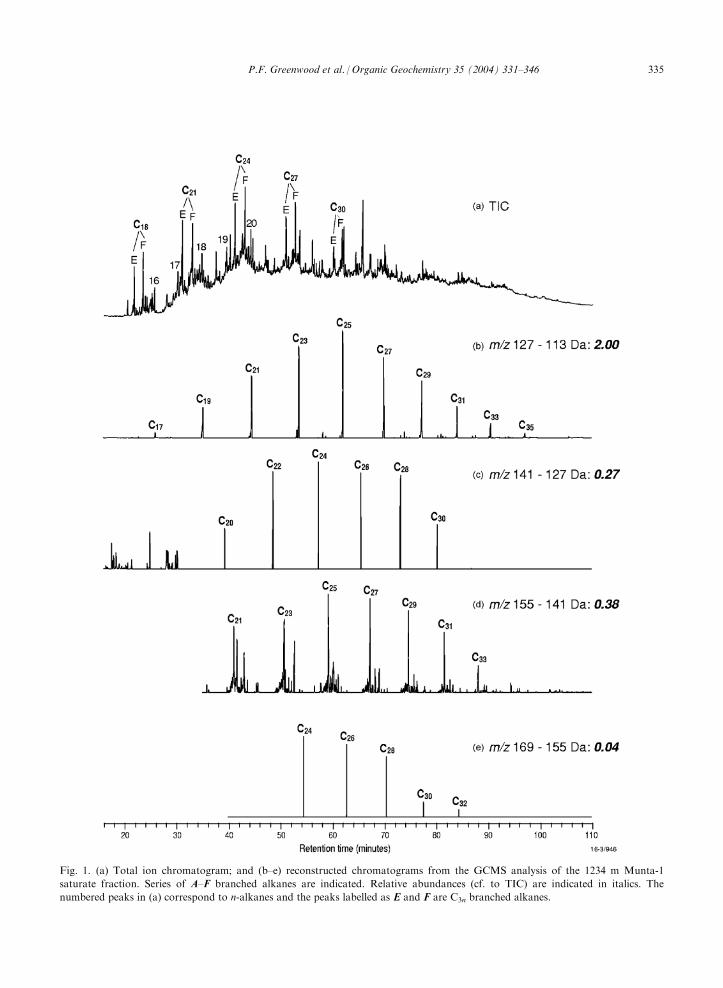
Fig. 1. (a) Total ion chromatogram; and (b–e) reconstructed chromatograms from the GCMS analysis of the 1234 m Munta-1
saturate fraction. Series of A–F branched alkanes are indicated. Relative abundances (cf. to TIC) are indicated in italics. The
numbered peaks in (a) correspond to n-alkanes and the peaks labelled as E and F are C3n branched alkanes.
P.F. Greenwood et al. / Organic Geochemistry 35 (2004) 331–346 335
very similar mass spectra to 5,5-diethylalkanes withlarge 127 Da, [M�57]+ and [M�29]+ fragment ionsanticipated. However, a further prominent fragment ion
would be expected at [M�99]+ from C6 to C7 cleavageadjacent to the C7-methyl group in the 3,7-dimethyl-alkanes (Kenig et al., 2003).
The 141–127 Da subtracted chromatogram shown inFig. 1c reveals another series of branched alkanes (heredenoted series B) characterized by a prominent C10H21
+
fragment ion (141 Da). This series comprises only even-carbon numbered members from C18 to C32. The massspectra of three members of this series are shown in
Table 2
Relative retention factors (RRF) for series A–F branched alkanes measured with a DB5 phase capillary column. DE=diethyl;
B,E=butyl,ethyl
Carbon nos.
Series A 5,5DE- Series B 6,6DE- Series C 5B,5E- Series D 6B,6E- Series E Series F18
�2.79 �2.4219
20
�1.0721
�0.95 �1.58 �3.88 �3.4522
�1.08 �1.7523
�0.94 �1.6024
�1.02 �1.74 �4.97 �4.5025
�0.89 �1.6126
�1.02 �1.7327
�0.86 �1.61 �6.05 �5.5728
�0.98 �1.7629
�0.83 �1.5830
�0.97 �1.78 �6.94 �6.4531
�0.80 �1.5732
�0.94 �1.7033
�0.77 �1.5734
�0.91 �1.7435
�0.73 �1.5736
�0.76 �1.73Fig. 2. Relative retention factor (RRF) values (Kissin et al., 1986) of series A–F branched alkanes. HBIs=Highly branched
isoprenoids.
336 P.F. Greenwood et al. / Organic Geochemistry 35 (2004) 331–346
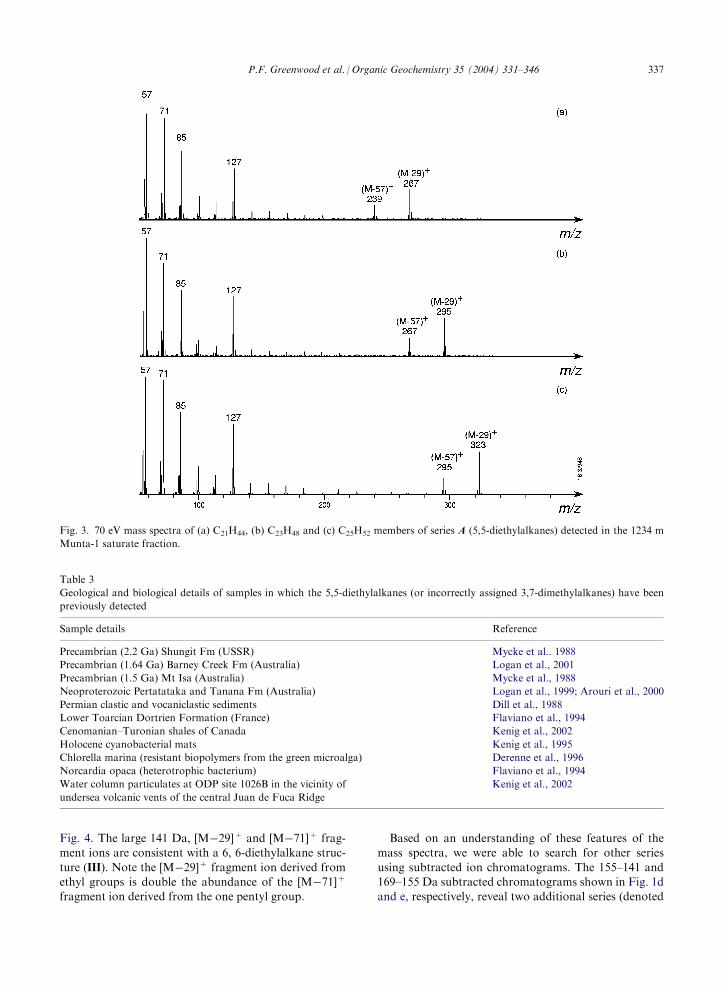
Fig. 4. The large 141 Da, [M�29]+ and [M�71]+ frag-ment ions are consistent with a 6, 6-diethylalkane struc-
ture (III). Note the [M�29]+ fragment ion derived fromethyl groups is double the abundance of the [M�71]+
fragment ion derived from the one pentyl group.
Based on an understanding of these features of themass spectra, we were able to search for other series
using subtracted ion chromatograms. The 155–141 and169–155 Da subtracted chromatograms shown in Fig. 1dand e, respectively, reveal two additional series (denoted
Fig. 3. 70 eV mass spectra of (a) C21H44, (b) C23H48 and (c) C25H52 members of series A (5,5-diethylalkanes) detected in the 1234 m
Munta-1 saturate fraction.
Table 3
Geological and biological details of samples in which the 5,5-diethylalkanes (or incorrectly assigned 3,7-dimethylalkanes) have been
previously detected
Sample details
ReferencePrecambrian (2.2 Ga) Shungit Fm (USSR)
Mycke et al.. 1988Precambrian (1.64 Ga) Barney Creek Fm (Australia)
Logan et al., 2001Precambrian (1.5 Ga) Mt Isa (Australia)
Mycke et al., 1988Neoproterozoic Pertatataka and Tanana Fm (Australia)
Logan et al., 1999; Arouri et al., 2000Permian clastic and vocaniclastic sediments
Dill et al., 1988Lower Toarcian Dortrien Formation (France)
Flaviano et al., 1994Cenomanian–Turonian shales of Canada
Kenig et al., 2002Holocene cyanobacterial mats
Kenig et al., 1995Chlorella marina (resistant biopolymers from the green microalga)
Derenne et al., 1996Norcardia opaca (heterotrophic bacterium)
Flaviano et al., 1994Water column particulates at ODP site 1026B in the vicinity of
undersea volcanic vents of the central Juan de Fuca Ridge
Kenig et al., 2002
P.F. Greenwood et al. / Organic Geochemistry 35 (2004) 331–346 337
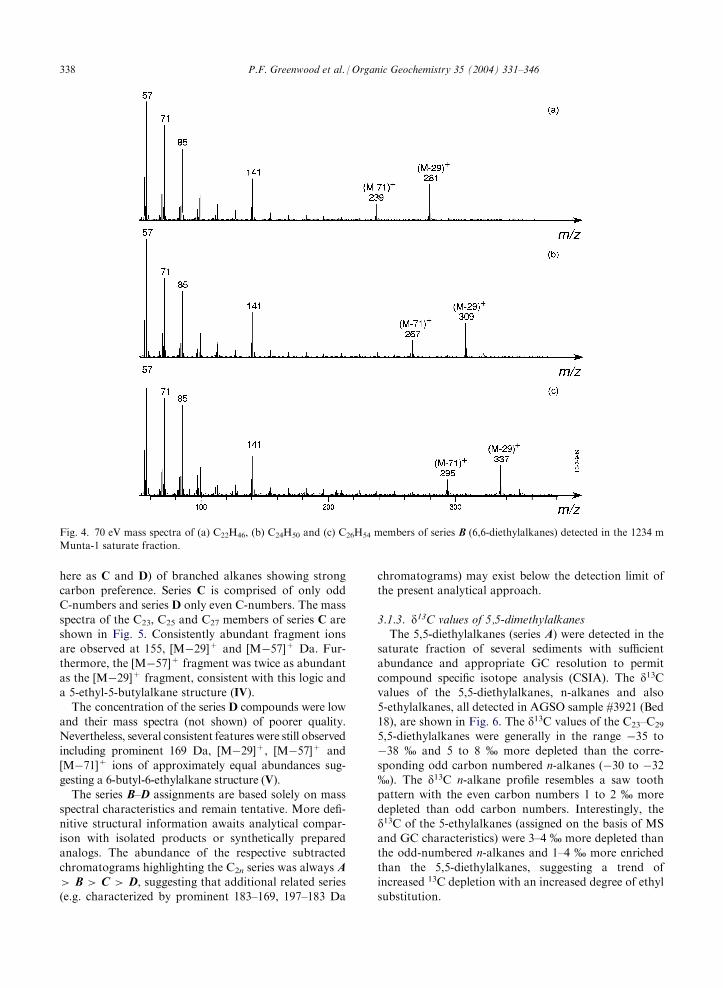
here as C and D) of branched alkanes showing strongcarbon preference. Series C is comprised of only oddC-numbers and series D only even C-numbers. The mass
spectra of the C23, C25 and C27 members of series C areshown in Fig. 5. Consistently abundant fragment ionsare observed at 155, [M�29]+ and [M�57]+ Da. Fur-thermore, the [M�57]+ fragment was twice as abundant
as the [M�29]+ fragment, consistent with this logic anda 5-ethyl-5-butylalkane structure (IV).The concentration of the series D compounds were low
and their mass spectra (not shown) of poorer quality.Nevertheless, several consistent features were still observedincluding prominent 169 Da, [M�29]+, [M�57]+ and
[M�71]+ ions of approximately equal abundances sug-gesting a 6-butyl-6-ethylalkane structure (V).The series B–D assignments are based solely on mass
spectral characteristics and remain tentative. More defi-nitive structural information awaits analytical compar-ison with isolated products or synthetically preparedanalogs. The abundance of the respective subtracted
chromatograms highlighting the C2n series was always A> B > C > D, suggesting that additional related series(e.g. characterized by prominent 183–169, 197–183 Da
chromatograms) may exist below the detection limit ofthe present analytical approach.
3.1.3. d13C values of 5,5-dimethylalkanesThe 5,5-diethylalkanes (series A) were detected in the
saturate fraction of several sediments with sufficientabundance and appropriate GC resolution to permit
compound specific isotope analysis (CSIA). The d13Cvalues of the 5,5-diethylalkanes, n-alkanes and also5-ethylalkanes, all detected in AGSO sample #3921 (Bed
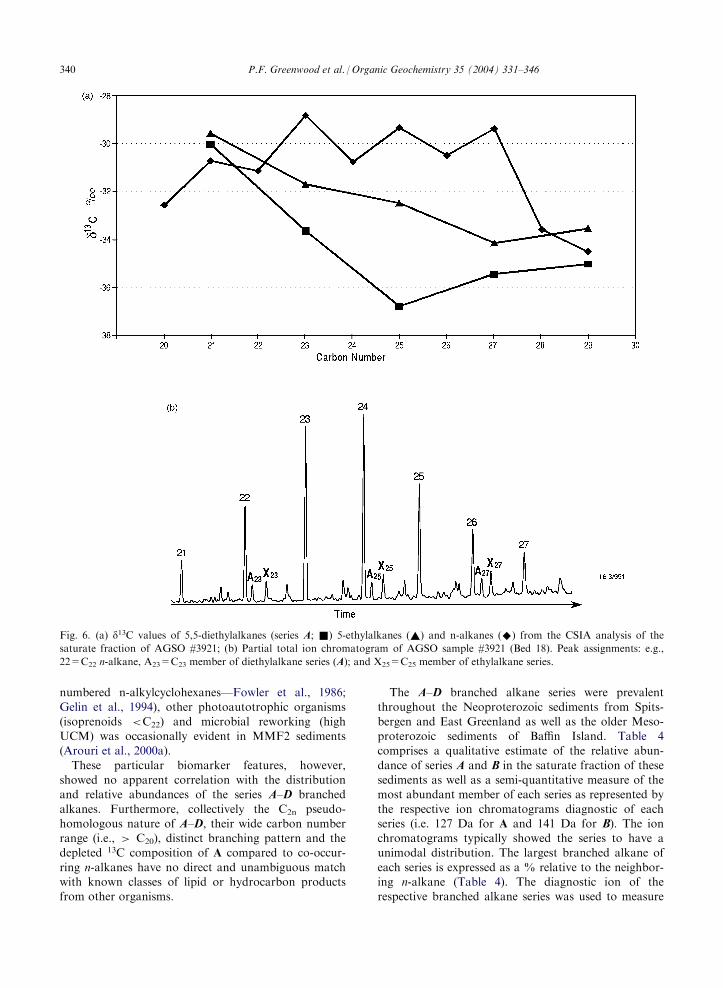
18), are shown in Fig. 6. The d13C values of the C23–C295,5-diethylalkanes were generally in the range �35 to�38 % and 5 to 8 % more depleted than the corre-
sponding odd carbon numbered n-alkanes (�30 to �32%). The d13C n-alkane profile resembles a saw toothpattern with the even carbon numbers 1 to 2 % more
depleted than odd carbon numbers. Interestingly, thed13C of the 5-ethylalkanes (assigned on the basis of MSand GC characteristics) were 3–4 % more depleted thanthe odd-numbered n-alkanes and 1–4 % more enriched
than the 5,5-diethylalkanes, suggesting a trend ofincreased 13C depletion with an increased degree of ethylsubstitution.
Fig. 4. 70 eV mass spectra of (a) C22H46, (b) C24H50 and (c) C26H54 members of series B (6,6-diethylalkanes) detected in the 1234 m
Munta-1 saturate fraction.
338 P.F. Greenwood et al. / Organic Geochemistry 35 (2004) 331–346

3.2. Geochemical significance of series A–D diethyl- andbutyl-, ethyl-alkanes
The C2n features of odd or even carbon number pref-erence and butyl and/or ethyl substitution observed forthe A–D series of branched hydrocarbons is consistentwith the biosynthetic elongation of a specific precursor
by acetate addition as is the case with other poly-methylenic lipid classes. They may derive from thedefunctionalisation of specific precursors, for example
biolipids containing carboxyl or hydroxyl groups.Methyl substitutions at specific chain locations arecommon among bacterial lipids, fatty acids for example
having preferential methylation sites at C-2, C-3 andC-10 (e.g. Dowling et al., 1986; Pond et al., 1985; Kosteret al., 1999). A biological origin is more likely than for-
mation via geosynthetic processes (e.g., thermal crack-ing of a longer precursor molecule), chemosynthesis orcatalyzed rearrangement of n-alkenes during catagenesis(Logan et al., 1999), all of which would contribute to
random carbon number and isomer distributions.Similar distributions of 5,5-diethylalkanes and the
previously reported, but incorrectly assigned, 3,7-
dimethylalkanes have been reported in a wide variety ofsamples ranging from extant biopolymers to Paleopro-terozoic sediments known to contain microbial mats
(Table 3). The series B–D branched alkanes have not beenso widely reported although Simons et al. (2002) recentlyreported B and C in Cenomanian–Turonian sediments.In the Tanana Formation sediments (Arouri et al.,
2000a), the seriesA–D branched alkanes were consistentlydetected in moderate to high abundance inMicrobial MatFacies 1 (MMF1) and occasionally in minor to trace
amounts in MMF2. Other hydrocarbon markers andfeatures diagnostic of specific bacterial types were occa-sionally detected in MMF1 sediments. For example,
prominent C16–C20 monomethylalkanes and 2a- and3b-methylhopanes (Summons and Jahnke, 1992; Sum-mons et al., 1999) are indicative of input from cyano-
bacteria and/or methanotrophs. Progression beyond C20of the monomethylalkane series, an even carbon numberpreference at the high MW end and the odd carbon siteof branching are features consistent with distributions
of mid-chain branched alkanes attributed to sulfide oxi-dizing bacteria (Logan et al., 1999). Additionalmolecular evidence for algae (e.g., even carbon-
Fig. 5. 70 eV mass spectra of (a) C23H48, (b) C25H52 and (c) C27H56 members of series series C (5-butyl, 5-ethylalkanes) detected in the
1234 m Munta-1 saturate fraction.
P.F. Greenwood et al. / Organic Geochemistry 35 (2004) 331–346 339
numbered n-alkylcyclohexanes—Fowler et al., 1986;Gelin et al., 1994), other photoautotrophic organisms
(isoprenoids <C22) and microbial reworking (highUCM) was occasionally evident in MMF2 sediments(Arouri et al., 2000a).
These particular biomarker features, however,showed no apparent correlation with the distributionand relative abundances of the series A–D branched
alkanes. Furthermore, collectively the C2n pseudo-homologous nature of A–D, their wide carbon numberrange (i.e., > C20), distinct branching pattern and thedepleted 13C composition of A compared to co-occur-
ring n-alkanes have no direct and unambiguous matchwith known classes of lipid or hydrocarbon productsfrom other organisms.
The A–D branched alkane series were prevalentthroughout the Neoproterozoic sediments from Spits-
bergen and East Greenland as well as the older Meso-proterozoic sediments of Baffin Island. Table 4comprises a qualitative estimate of the relative abun-
dance of series A and B in the saturate fraction of thesesediments as well as a semi-quantitative measure of themost abundant member of each series as represented by
the respective ion chromatograms diagnostic of eachseries (i.e. 127 Da for A and 141 Da for B). The ionchromatograms typically showed the series to have aunimodal distribution. The largest branched alkane of
each series is expressed as a % relative to the neighbor-ing n-alkane (Table 4). The diagnostic ion of therespective branched alkane series was used to measure
Fig. 6. (a) d13C values of 5,5-diethylalkanes (series A; &) 5-ethylalkanes (~) and n-alkanes (^) from the CSIA analysis of the
saturate fraction of AGSO #3921; (b) Partial total ion chromatogram of AGSO sample #3921 (Bed 18). Peak assignments: e.g.,
22=C22 n-alkane, A23=C23 member of diethylalkane series (A); and X25=C25 member of ethylalkane series.
340 P.F. Greenwood et al. / Organic Geochemistry 35 (2004) 331–346

the abundance of both the branched alkane and neigh-
boring n-alkane, so are biased towards the detection ofthe branched alkane. AGSO sample #3912, for example,contains both series A and B in very high relative
abundances with the 5,5-diethylpentacosane (200% cf.C30 n-alkane based on m/z 127 data) and 6, 6-diethylte-tracosane (80% cf. C29 n-alkane based on m/z 141 data)
being the most abundant members of series A and B,respectively.The relative abundances of the A–D series hydro-
carbons detected in the Spitsbergen and Baffin Island
sediments showed no overt geological or lithologicalassociation, although there was a persistent presence oflow contents of organic matter of apparent benthic
microbial origin. Examination of the broader secularand sedimentary distribution patterns of series A
(Table 3) presents some clues. Their presence in water
expelled from a contemporary sub-sea hydrothermalvent (Axial volcano) and modern to Holocene micro-bial mats (Abu Dhabi) indicates that the source
organism(s) are prokaryotic and presently extant(Kenig et al., 2002). Based on their high relative abun-dances in some samples, the source organisms appar-ently can represent significant concentrations and
quantities of biomass in some kinds of habitat. Thepresence of series A in a deep sea hydrothermal envir-onment discounts photoautotrophs as a likely source.
Reports of their occurrence in Phanerozoic sediments
are rare whereas they do appear to be more abundantin the Proterozoic. Mesozoic occurrences are in blackshales which, in the case of the Cenomanian–Turonian
at least, are attributed to oceanic anoxia (Simons et al.,2002).It has been hypothesized that Earth’s oceans were
largely anoxic at the beginning of the Paleoproterozoic,anoxic and sulfidic by the Mesoproterozoic and largelyventilated by the end of the Neoproterozoic (Canfield,1998; Anbar and Knoll, 2002). During this period there
was a major shift in the redox chemistry of marine sul-fur and nitrogen species and there would have existedsteep redox gradients, concomitant with shifts in spe-
ciation of both S and N in the water column as there arein sediments today. Organisms having some role in thesulfur cycle, or possibly the nitrogen cycle, could have
exploited these redox gradients and been moreprevalent. This would explain an enhanced expressionof their lipids in Proterozoic sediments compared to
Phanerozoic ones where the marine water column wasonly rarely affected by anoxia. A detailed search ofmodern environments where there are steep redoxgradients supporting primary production by microbes is
a plausible approach to finding further examples ofthese lipids and, through the associated biogeochemicalprocesses, the responsible microbes.
Table 4
Relative abundance of C2n (series A and B) and C3n (series E) branched alkanes detected in saturate fractions of Spitsbergen, East
Greenland and Baffin Island sediments
Samples
Series A—5,5-diethylalkanes (127 Da)
Series B—6,6-diethyl
alkanes (141 Da)
Series E—C3nalkanes (155 Da)
Relativeabundancea
Largest(cf. n-alk)b
Relativeabundancea
Largest(cf. n-alk)b
Relativeabundancea
Largest(cf. n-alk)b
AGSO #3912 (86-P-13)
++++ A29 (200) +++ B28 (80) +++ E24 (110) AGSO #3913 (86-P-152) ++ A29 (40) + B30 (20) + E21 (40) AGSO #3914 (Biod.c) (86-P-105) +++ A29 (210) + B30 (100) ++ E21 (740)cAGSO #3915 (86-P-9)
++++ A29 (210) ++ B28 (70) ++ E24 (200) AGSO #3916 (86-M-43) + A23 (20) + B28 (30) ++ E27 (140) AGSO #3917 (86-F-5) +++ A29 (320) ++ B28 (100) +++ E27 (60)AGSO #3918 (Biod.c) (86-F-6)
+ A29 (20) ND � ++ E30 (30) AGSO #3919 (86-G-68) + A23 (10) ND � +++ E24 (90)AGSO #3920 (Biod.c) (86-P-42)
+ A23 (3) ND � ++ E24 (690)cAGSO #3921 (83-E-19-5)
++++ A27 (140) ++ B28 (80) + E27 (70) AGSO #3922 (86-P-66) ++++ A27 (220) ++ B28 (80) + E27 (30)AGSO #3923 (86-G-13)
++++ A29 (320) ++ B28 (100) + E27 (60) AGSO #3924 (86-ST-5) + A25 (40) ND � + E30 (30) AGSO #4185 (P-6450) ++ A27 (50) + B28 (20) ++ E24 (60)AGSO #4193 (87-SS-172)
++++ A25 (250) ++ B28 (160) ++++ E27 (790) AGSO #4194 (87-SS-225) ++++ A25 (220) ++ B28 (180) ++++ E27 (430)ND=not detected.a +, ++, +++, ++++ reflects relative abundance.b % Compared with the n-alkane eluting just prior to the most abundant branched alkane. An=Series A branched alkane with n
carbon numbers.c Saturate hydrocarbon distribution shows evidence of biodegradation.
P.F. Greenwood et al. / Organic Geochemistry 35 (2004) 331–346 341
3.3. GC–MS of C3n series E and F branched alkanes
3.3.1. Relative retention factorsThe series E and F compounds detected in Tanana
sediments elute at very early retention times comparedto n-alkanes of corresponding carbon number indicatinga very high degree of branching. The RRF values of E
and F are extremely negative and decrease with increas-ing MW (Table 2, Fig. 2). These values are of a similarorder to those reported previously for highly branched
isoprenoids (HBIs), notwithstanding that the latter weremeasured on columns with different stationary phases(Barrick et al., 1980; Prahl et al., 1980). HBIs (e.g.,VI and
VII of Appendix) are ubiquitous in Cenozoic to modernmarine sediments and are some of the most highly methylsubstituted compounds known (Robson and Rowland,1986; Rowland and Robson, 1990). The RRF profiles of
E and F are more negative than the HBI series suggestingthey exhibit an even higher degree of branching. Thedecrease in the RRF values with increasingMW indicates
increased branching with each C-3 addition.
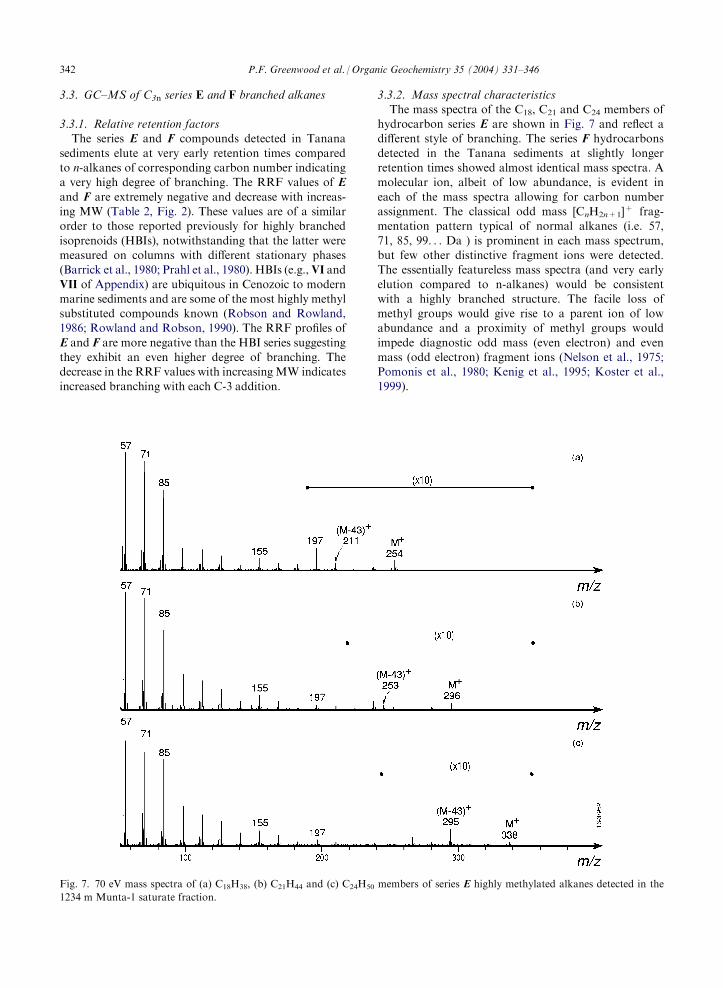
3.3.2. Mass spectral characteristicsThe mass spectra of the C18, C21 and C24 members of
hydrocarbon series E are shown in Fig. 7 and reflect adifferent style of branching. The series F hydrocarbons
detected in the Tanana sediments at slightly longerretention times showed almost identical mass spectra. Amolecular ion, albeit of low abundance, is evident in
each of the mass spectra allowing for carbon numberassignment. The classical odd mass [CnH2n+1]
+ frag-mentation pattern typical of normal alkanes (i.e. 57,
71, 85, 99. . . Da ) is prominent in each mass spectrum,but few other distinctive fragment ions were detected.The essentially featureless mass spectra (and very early
elution compared to n-alkanes) would be consistentwith a highly branched structure. The facile loss ofmethyl groups would give rise to a parent ion of lowabundance and a proximity of methyl groups would
impede diagnostic odd mass (even electron) and evenmass (odd electron) fragment ions (Nelson et al., 1975;Pomonis et al., 1980; Kenig et al., 1995; Koster et al.,
1999).
Fig. 7. 70 eV mass spectra of (a) C18H38, (b) C21H44 and (c) C24H50 members of series E highly methylated alkanes detected in the
1234 m Munta-1 saturate fraction.
342 P.F. Greenwood et al. / Organic Geochemistry 35 (2004) 331–346

3.4. Geochemical significance of series E and F highlybranched hydrocarbons
Neither of the E and F hydrocarbons are known in
extant organisms or modern organic sediments and nobiological reason has been identified for their occurrencein ancient sediments. However, such a sequence of
compounds that are systematically elongated by C-3units is almost certainly a consequence of biosynthesis.The C3n branched alkanes were detected in only a
small number of samples associated with microbial matfacies 2 (MMF2) of the Tanana Formation. They werenot detected in completely autochthonous MMF2 sedi-
ments nor the other organic facies (Arouri et al., 2000a).This mat facies was originally deposited in shallow-water extending from the peritidal shelf to upper slope-apron zones. The rocks containing these mats can be
distinguished into light and dark colored sediments, thedarker color due to autochthonous mats together withother kerogen fragments pervading the abundant silt
laminae, but E and F were occasionally detected insediments of both colors. Some MMF2 sediments showbiomarker evidence for cyanobacteria (e.g., C16, C18 and
C20 mid-chain monomethylalkanes) and/or thermo-acidophilic bacteria (C17 and C19 alkylcyclohexanes; deRosa et al., 1972; Oshima and Ariga, 1975). Pyrite
indicative of sulfate reducing bacteria was also detectedin some darker colored sediments. Again, however,none of the potentially diagnostic molecular featuresshowed any firm correlation with the distribution of E
and F throughout the MMF2 sediments.Series E, but not F, branched alkanes were detected in
all of the Spitsbergen and Baffin Island samples (Table 4).
The relative abundances of E varied from very high(AGSO# 3912, 3917, 1919) to low (AGSO# 3913, 3921,3922, 3923, 3924) in the Spitsbergen sediments and was
very high in both Baffin Island sediments (AGSO #4193and #4194). The saturate fractions of several Spitsbergensamples (AGSO# 3914, 3918, 3920) showed evidence ofbiodegradation which enhances the abundances of low
MW series E hydrocarbons relative to the n-alkanes oflower carbon numbers eluting just prior to them (Table 4).
4. Conclusions
Pseudo-homologous series of odd carbon numberedC17–C35 5,5-diethylalkanes (series A) and 5-butyl, 5-ethyl-alkanes (C); and even carbon numbered C16–C34 6,6-
diethylalkanes (B) and 6-butyl, 6-ethylalkanes (D) areprevalent in Neoproterozoic sediments associated withbenthic microbial mats. Two additional series denoted E
and F, with a different and presently unknown style of
branching, have a more restricted distribution. The identi-fication of A as 5,5-diethylalkanes is based on the Keniget al. (2002) synthesis of the C19 homologue while the
other assignments are tentatively based on interpretationof mass spectral data and relative gas chromatographicretention times. Further work, such as comparisons withadditional authentic standards prepared by synthesis, is
needed to confirm these assignments while the structureelucidation of the E and F compounds will likely requireisolation and further spectroscopic investigation.
The presence of these hydrocarbons in very complexorganic matrices, such as with microbial mat-derivedorganic matter, makes them difficult to detect unless
specific ion profiles or subtracted ion chromatogramsare examined closely. The paucity of reports of theirexistence is probably a consequence of this and their
generally low abundance in Phanerozoic sediments. Useof the diagnostic subtracted ion chromatograms employedhere to detect the C2n series A–D hydrocarbons maylead to further examples of their natural occurrence and,
consequently, more clues about possible source organisms.Identification of the series E and F hydrocarbons is
even more problematic due to a lack of diagnostic mass
spectral features and this has almost certainly con-tributed to the infrequency of their detection.The series A–F branched hydrocarbons appear to
reflect inputs of organic matter from prokaryoticmicrobes although their exact sources remain enigmatic.The C2n distributions of A–D show no direct correlation
with the C3n distributions of E and F in any of theTanana, Spitsbergen and Baffin Island formations and,as a consequence, the various series may be proxies fordifferent kinds of physiologies.
Possible source organisms are specific bacteria thatexploit the redox gradients that prevail in hydrothermalsystems and anoxic and/or euxinic environments and
include organisms involved in either the sulfur or nitrogencycles.
Acknowledgements
The authors gratefully acknowledge the provision of
samples and contextual information by ProfessorAndrew Knoll, Harvard University. Analytical assis-tance for the extraction, purification and isotopic ana-
lysis of hydrocarbon fractions was provided by JanetHope, Geoscience Australia. Reviews by Steve Rowlandand an anonymous reviewer greatly improved an early
draft of the manuscript. PFG and GAL publish with thepermission of the CEO of Geoscience Australia.
Associate Editor — Steve Rowland
Appendix
Representative molecular structures: (I) 3, 7-dimethyl-nonadecane; (II) 5, 5-diethylheptadecane (=a member
P.F. Greenwood et al. / Organic Geochemistry 35 (2004) 331–346 343

of series A) (III) 6, 6-diethylheptadecane; (IV) 5-butyl-,
5-ethylheptadecane; (V) 6-butyl-, 6-ethylheptadecane;(VI) 2,6,10-trimethyl-7-(3-methylpentyl)-dodecane (C20
HBI); (VII) 2,6,10,14,18-pentamethyl-7-(3-methylpen-tyl)-nonadecane (C30 HBI).
References
Anbar, A.D., Knoll, A.K., 2002. Proterozoic ocean chemistry
and evolution: a bioinorganic bridge? Science 297, 1137–1142.
Arouri, K., Conaghan, P.J., Walter, M.R., Bischoff, G.C.O.,
Grey, K., 2000a. Reconnaissance sedimentology and hydro-
carbon biomarkers of Ediacarian microbial mats and acri-
tarchs, lower Ungoolya Group, Officer Basin. Precambrian
Research 100, 235–280.
Arouri, K., Greenwood, P.F., Walter, M., 1999. A possible
chlorophycean affinity of some Neoproterozoic acritarchs.
Organic Geochemistry 30, 1323–1337.
Arouri, K., Greenwood, P.F., Walter, M., 2000b. Biological
affinities of Neoproterozoic acritarchs from Australia:
microscopic and chemical characterisation. Organic Geo-
chemistry 31, 75–89.
Barrick, R.C., Hedges, J.I., Peterson, M.L., 1980. Hydrocarbon
geochemistry of the Puget sound region—I. Sedimentary
acyclic hydrocarbons. Geochimica et Cosmochimica Acta 44,
1349–1362.
Canfield, D.E., 1998. A new model for Proterozoic ocean
chemistry. Nature 396, 450–453.
Dembitsky, V.M., Dor, I., Shkrob, I., Aki, M., 2001. Branched
alkanes and other apolar compounds produced by the cya-
nobacterium Microcoleus vaginatus from the Negev Desert.
Russian Journal of Bioorganic Chemistry 27, 110–119.
Derenne, S., Largeau, C., Berkaloff, C., 1996. First example of
an algeanan yielding aromatic-rich pyrolysate: possible geo-
chemical implications on marine kerogen formation. Organic
Geochemistry 24, 617–627.
de Rosa, M., Gambacorta, A., Minale, L., Bullock, J.D., 1972.
The formation of o-cyclohexyl-fatty acids from shikimate in
an acidophilic thermophilic bacillus. Biochemistry Journal
128, 751–754.
Dill, H., Teschner, M., Wehner, H., 1988. Petrography, inor-
ganic and organic geochemistry of Lower Permian carbo-
naceous fan sequences (‘‘Brandschiefer Series’’), Federal
Republic of Germany; constraints to their paleogeography
and assessment of their source rock potential. Chemical
Geology 67, 307–325.
Dowling, N.J.E., Widdel, F., White, D.C., 1986. Phospholipid
ester-linked fatty acid biomarkers of acetate-oxidizing sul-
phate-reducers and other sulfide-forming bacteria. Journal of
General Microbiology 132, 1815–1825.
Flaviano, C., Le Berre, F., Derenne, S., Largeau, C., Connan,
J., 1994. First indications of the formation of kerogen
amorphous fractions by selective preservation; role of non-
hydrolysable macromolecular constituents of eubacterial cell
walls. Organic Geochemistry 22, 759–771.
Fowler, M.G., Douglas, A.G., 1987. Saturated hydrocarbon
biomarkers in oils of Late Precambrian age from Eastern
Siberia. Organic Geochemistry 11, 201–213.
Fowler, M.G., Abolins, P., Douglas, A.G., 1986. Monocyclic
alkanes in Ordovician organic matter. Organic Geochemistry
10, 815–823.
Gelin, F., De Leeuw, J.W., Sinninghe Damste, J.S., Derenne,
S., Metzger, P., 1994. The similarity of chemical structures of
soluble aliphatic polyaldehyde and insoluble algaenan in the
green microalga Botryococcus braunii race A as revealed by
analytical pyrolysis. Organic Geochemistry 21, 423–435.
344 P.F. Greenwood et al. / Organic Geochemistry 35 (2004) 331–346

Green, J.W., Knoll, A.H., Golubic, S., Swett, K., 1987. Paleo-
biology of distinctive benthic microfossils from the Upper
Proterozoic Limestone-Dolomite ‘‘Series’’, Central East
Greenland. American Journal of Botany 74, 928–940.
Green, J.W., Knoll, A.H., Swett, K., 1988. Microfossils from
oolites and pisolites of the Upper Proterozoic Eleonore Bay
Group, Central East Greenland. Journal of Paleontology 62,
835–852.
Grey, K., 1998. Ediacarian Palynology of Australia. PhD
Thesis, Macquarie University, (Unpublished).
Hold, I.M., Schouten, S., Jellma, J., Sinninghe Damste, J.S.,
1999. Origin of free and bound mid-chain methyl alkanes in
oils, bitumens and kerogens of the marine, Infracambrian
Huqf Formation (Oman). Organic Geochemistry 30, 1411–
1428.
Kah, L.C., Lyons, T.W., Chesley, J.T., 2001. Geochemistry of
a 1.2 Ga carbonate-evaporite succession, northern Baffin and
Bylot Islands: Implications for Mesoproterozoic marine
evolution. Precambrian Research 111, 203–234.
Kah, L.C., Sherman, A.G., Narbonne, G.M., Knoll, A.H.,
Kaufman, A.J., 1999. d13C stratigraphy of the Proterozoic
Bylot Supergroup, Baffin Island, Canada: implications for
regional lithostratigraphic correlations. Canadian Journal of
Earth Sciences 36, 311–332.
Kenig, F., Simons, D.-J.H., Crich, D., Cowen, J.P., Ventura,
G.T., Brown, T.C., Rehbein, T., 2002. Alkanes with a qua-
ternary carbon centre: a 2,200 Myr record of sulfide oxidiz-
ing bacteria. Geochimica et Cosmochimica Acta 66, A393.
(abstr.).
Kenig, F., Simons, D.-J.H., Crich, D., Cowen, J.P., Ventura,
G.T., Rehbein-Khalily, T., Brown, T.C., Anderson, K.B.,
2003. Branched aliphatic alkanes with quaternary substituted
carbon atoms in modern and ancient geologic samples. Pro-
ceedings of the National Academy of Sciences USA 10/22,
12554–12558.
Kenig, F., Sinninghe Damste, J.S., Kock-van Dalen, A.C.,
Rijpstra, W.I.C., Huc, A.Y., de Leeuw, J.W., 1995. Occur-
rence and origin of mono-, di-, and trimethylalkanes in
modern and Holocene cyanobacterial mats from Abu Dhabi,
United Arab Emirates. Geochimica et Cosmochimica Acta
59, 2999–3015.
Kissin, Y.V., Feulmer, G.P., Payne, W.B., 1986. Gas chroma-
tographic analysis of polymethyl-substituted alkanes. Jour-
nal of Chromatographic Science 24, 164–169.
Knoll, A.H., Golubic, S., Green, J., Swett, K., 1986a. Organi-
cally preserved microbial endoliths from the Late Proter-
ozoic of East Greenland. Nature 321, 856–857.
Knoll, A.H., Hayes, J.M., Kaufman, A.J., Swett, K., Lambert,
I.B., 1986b. Secular variations in carbon isotope ratios from
Upper Proterozoic successions of Svalbard and East Green-
land. Nature 321, 832–838.
Knoll, A.H., Swett, K., 1990. Carbonate deposition during the
later Proterozoic Era: An example from Spitsbergen. Amer-
ican Journal of Science 290A, 104–132.
Knoll, A.H., Swett, K., Mark, J., 1991. Paleobiology of a
Neoproterozoic tidal flat/lagoonal complex; the Draken
conglomerate formation, Spitsbergen. Journal of Paleontol-
ogy 65, 531–570.
Koster, J., Volkman, J.K., Rullkotter, J., Scholz-Bottcher,
B.M., Reithmeier, J., Fischer, U., 1999. Mono-, di- and tri-
methyl-branched alkanes in cultures of the filamentous cya-
nobacterium Calothrix scopulorum. Organic Geochemistry
30, 1367–1379.
Logan, G.A., Calver, C.R., Gorjan, P., Summons, R.E., Hayes,
J.M., Walter, M.R., 1999. Terminal Proterozoic mid shelf
benthic microbial mats in the Centralian Superbasin and
their environmental significance. Geochimica et Cosmochi-
mica Acta 63, 1345–1358.
Logan, G.A., Hinman, M.R., Walter, M.R., Summons, R.E.,
2001. Biogeochemistry of the 1640 Ma McArthur River
(HYC) lead zinc ore and host sediments, Northern Territory,
Australia. Geochimica et Cosmochimica Acta 65, 2317–2336.
Logan, G.A., Summons, R.E., Hayes, J.M., 1997. An isotopic
biogeochemical study of Neoproterozoic and Early Cam-
brian sediments from the Centralian Superbasin, Australia.
Geochimica et Cosmochimica Acta 61, 5391–5409.
Mycke, B., Michaelis, W., Degens, E.T., 1988. Biomarkers in
sedimentary sulfides of Precambrian age. Organic Geochem-
istry 13, 619–625.
Nelson, D.R., Sukkestad, D.R., 1975. Normal and branched
alkanes from cast skins of the grasshopper Schistocerca vaga
(Scudder). Journal of Lipid Research 16, 13–18.
Oshima, M., Ariga, T., 1975. o-Cyclohexyl fatty acids in acid-ophilic bacteria. Journal Biological Chemistry 256, 6963–
6968.
Pomonis, J.G., Nelson, D.R., Fatland, C., 1980. Insect hydro-
carbons 2. Mass spectra of dimethylalkanes and the effect of
the number of methylene units between methyl groups on
fragmentation. Journal of Chemical Ecology 6, 965–973.
Pond, J.L., Langworthy, T.A., Holzer, G., 1985. Long-chain
diols: A new class of membrane lipids from a thermophilic
bacterium. Science 213, 1134–1136.
Prahl, F.G., Bennet, J.T., Carpenter, R., 1980. The early diag-
enesis of aliphatic hydrocarbons and organic matter in sedi-
mentary particles from dabob Bay, Washington. Geochimica
et Cosmochimica Acta 44, 1967–1976.
Robinson, N., Eglinton, G., 1990. Lipid chemistry of Icelandic
hot spring microbial mats. Organic Geochemistry 15, 291–
298.
Robson, J.N., Rowland, S.J., 1986. Identification of novel
widely distributed sedimentary acyclic sesterpenoids. Nature
324, 561–563.
Rowland, S.J., Robson, J.N., 1990. The widespread occurrence
of highly branched acyclic C20, C25 and C30 hydrocarbons in
recent sediments and biota- a review. Marine Environment
Research 30, 191–216.
Schopf, J.W., 1968. Microflora of the Bitter Springs, late Pre-
cambrian, central Australia. Journal of Paleontology 42,
651–688.
Shiea, J., Brassell, S.C., Ward, D.M., 1990. Mid-chain bran-
ched mono- and dimethyl alkanes in hot spring cyano-
bacterial mats; a direct biogenic source for branched
alkanes in ancient sediments? Organic Geochemistry 15,
223–231.
Shiea, J., Brassell, S.C., Ward, D.M., 1991. Comparative ana-
lysis of extractable lipids in hot spring microbial mats and
their component photosynthetic bacteria. Organic Geo-
chemistry 17, 309–319.
Simons, D.-J.H., Kenig, F., Crich, D., Schroder-Adams, C.J.,
2002. Significance of novel branched alkanes with quaternary
carbon centers in black shales. Geochimica et Cosmochimica
Acta 66, A718. (abstr.).
P.F. Greenwood et al. / Organic Geochemistry 35 (2004) 331–346 345

Summons, R.E., Jahnke, L.L., 1992. Identification of the
methylhopanes in sediments and petroleum. Geochimica et
Cosmochimica Acta 54, 247–251.
Summons, R.E., Jahnke, L.L., Logan, G.A., Hope, J.M., 1999.
2-Methylhopanoids as biomarkers for cyanobacterial oxy-
genic photosynthesis. Nature 398, 554–557.
Summons, R.E., Murray, A.P., Boreham, C.J., Gorter, J., 1995.
Chemostratigraphy and the composition of oils from the
Perth Basin, Western Australia. The APPEA Journal 35, 613–
632.
Summons, R.E., Walter, M.R., 1990. Molecular fossils and
microfossils of prokaryotes and protists from Proterozoic
sediments. American Journal of Science 290A, 212–244.
Swett, K., Knoll, A.H., 1989. Marine pisolites from the Upper
Proterozoic carbonates of East Greenland and Spitsbergen.
Sedimentology 36, 75–93.
346 P.F. Greenwood et al. / Organic Geochemistry 35 (2004) 331–346





























