Embed Size (px)
Citation preview

INTRODUCTION
The mental representation of numbers andquantities is a fundamental ability of the mind. It isinvolved in a variety of human activities rangingfrom our everyday handling of objects and usageof money, measures of distance and time, throughto astronomy and computer sciences. However,some aspects of numerical cognition appear to beshared with many non-human species, suggestingthat the complex representation of numbers inhumans may have biological foundations that arealso available to other animals (for reviews, seeDehaene, 1997; Butterworth, 1999). Monkeys(Brannon and Terrace, 1998), as well as rats (Meckand Church, 1983) and birds (Fetterman, 1993) candiscriminate between different numbers of objectsor events. Similarly, human infants demonstrate anearly ability to distinguish the numerosity ofvarious entities, including visual stimuli, sounds, oractions (Wynn, 1998; Brannon, 2002). Therefore,in contrast to traditional theories assuming thatnumerical competence emerges from thedevelopment of linguistic and reasoning skills (e.g.Piaget, 1941), more recent hypotheses have beenput forward according to which elementary numberrepresentation may rely in part on specializedsystems in the brain that are intimately associatedwith object individuation (Simon, 1997) or visuo-spatial functions (Dehaene, 1992, 1997).
In support of the role of a spatial code innumber representation, psychophysical studies inhealthy humans have suggested that numberprocessing may operate on an analogical magnitude
format that is spatially organized by numericalproximity (Dehaene et al., 1998). Such spatialrepresentation of numbers has been described as a“mental number line” along which numbers arepositioned from the left to right side according totheir progressively increasing magnitude(Restle,1970; Seron et al., 1992; Dehaene et al., 1993).Small numbers would be mentally representedfurther toward the left along this line, and largenumbers more toward the right. Consistent withthis proposal, people show spatial compatibilityeffects when responding to numbers, with fasterreaction times to small numbers using the left handand faster reaction times to large numbers usingthe right hand (Dehaene et al., 1993; Bächtold etal., 1998). This can be found even when numbermagnitude is irrelevant to the task, suggesting thatnumber processing may involve an automaticaccess to spatial codes, a phenomenon dubbed asthe Spatial Numerical Association of ResponseCodes (SNARC) effect (Dehaene et al., 1993).
Moreover, neuropsychology studies in patientswith brain damage (Benson and Weir, 1972;Grafman et al., 1982; Dehaene and Cohen, 1991;Delazer and Benke, 1997; Butterworth, 1999;Cohen et al., 2000) and recent findings fromfunctional neuroimaging (Rueckert et al., 1996;Chochon et al., 1999; Dehaene et al., 1999; Pinel etal., 1999; Gruber et al., 2001; Pinel et al., 2001)suggest that number processing is criticallyassociated with neural circuits in bilateral parietallobes. According to these studies, left parietalregions might be more important for exactarithmetic abilities based on conceptual knowledge
Cortex, (2004) 40, 000-000
ABSTRACTTHE NUMBER SPACE AND NEGLECT
Patrik Vuilleumier1, Stéphanie Ortigue2 and Peter Brugger3
(1Laboratory of Behavioural Neurology and Imaging of Cognition, Geneva; 2Functional Brain Mapping Laboratory,University Hospital Geneva; 3Department of Neurology, Neuropsychology Unit, University Hospital Zürich)
ABSTRACT
Recent cognitive models of numerical abilities have postulated that number processing may in part rely on arepresentation of quantities where magnitude is organized by spatial proximity, along a “mental number line” extendingfrom left to right. We describe four experiments that examined whether such a spatial representation of number would beaffected by the presence of unilateral neglect after right hemisphere damage. When asked to judge whether a single numbershown at fixation was smaller or larger than “5”, patients with neglect were selectively slower to respond to “4”, but whenasked to compare numbers to “7” they were selectively slower to respond to “6”. This is consistent with a representationaldeficit for numbers located to the left of a reference point along the mental number line and was not found in other rightbrain-damage patients without neglect. No effect of represented number position was found in a non-numerical taskrequiring judgements of the physical size of single digit characters. Finally, when asked to classify numbers as indicatinghours earlier or later than six o’clock, neglect patients showed a reverse pattern with slower responses to numbers largerthan “6’, consistent a representational deficit for hour numbers located on the left side of an imagined clock-face. Ourfindings demonstrate that unilateral spatial neglect may produce specific representational deficits in number processing thatimplicate different spatial representation according to the task demands.
Key words: ??????

and fact retrieval, whereas right parietal regionsmight be more important for approximatecalculations based on an analogical quantityrepresentation (Chochon et al., 1999; Cohen et al.,2000). Remarkably, many parietal areas activatedduring number processing are also known to beimplicated in visuo-spatial functions, includingspatial attention (Gitelman et al., 1999; Corbetta etal., 2000), oculo-motor planning (Corbetta et al.,1998), mental rotation (Kosslyn et al., 1998), andother spatial coordinate-transformation tasks (Zackset al., 1999). On the other hand, recent findingssuggest that estimation of numerosity for small setsof discrete items (i.e., ? 4) may recruit specific brainareas in extrastriate cortex, outside parietal regionsaffected by spatial neglect (Sathian et al., 1999;Vuilleumier and Rafal, 1999; Fink et al., 2001).
Our study examined whether any spatialrepresentation in number processing would beaffected by the presence of unilateral neglect afterright hemisphere damage. Spatial neglect ischaracterized by a profound disturbance in therepresentation of contralesional space, with animpaired ability to direct attention and respond tostimuli located on the affected side (Bisiach andVallar, 1988; Vallar, 1998; Driver and Vuilleumier,2001). Neglect typically follows focal brain lesionsin parietal cortex (Vallar, 1993) or other regionsinterconnected with parietal cortex (Mesulam,1999; Karnath, 2001). Since the seminalobservations by Bisiach (Bisiach and Luzzatti,1978; Bisiach et al., 1979; Bisiach, 1993), therehas been abundant evidence that unilateral spatialneglect can also concern mental representations invisual imagery (Guariglia et al., 1991; Bartolomeoet al., 1994; Coslett, 1997; Ortigue et al., 2001) ormemory (Meador et al., 1987; Beschin et al.,2000). Moreover, a recent report suggested that the“mental number line” might be distorted in patientswith right brain damage and left neglect (Zorzi etal., 2002), who showed a systematic misplacementof the midpoint towards higher values whenjudging a numerical interval that was presentedaurally (e.g., stating that five is halfway betweentwo and six). This result might be consistent withrepresentational neglect for numbers along themental number line, resembling the rightwarddeviation observed during the bisection of physicalline in these patients (Bisiach and Vallar, 1988;Vallar, 1998). Alternatively, it might also reflect anon-spatial bias for the larger of two numbers witha systematic response strategy to remove anapproximate amount from the latter (rather than aspatial deviation towards “right-sided” numbers onthe mental line), possibly related to previoussuggestions that the right hemisphere quantitysystems might be preferentially bias towards therepresentation of small numbers (Cipolotti et al.,1991; Stanescu-Cosson et al., 2000).
Here we report a series of four experiments thatfurther investigated the spatial nature of impaired
number processing in neglect. Patients and controlswere tested on number comparison tasks previouslyused in normal subjects (Dehaene, 1989; Bächtoldet al., 1998; Pinel et al., 1999). We systematicallymanipulated the nature and spatial parameters ofthe task in order to determine whether neglect mayspecifically affect the representation of positionalproximity along the mental number line(Experiments 1 and 2) and specifically occurduring numerical judgments, as opposed tophysical size or other non-numerical judgements(Experiments 3 and 4).
EXPERIMENT 1: NUMBER SIZE COMPARISON
This experiment used a simple numbercomparison task in which a number stimulus has tobe judged as smaller or larger than a fixed referencenumber. This task is thought to involve a criticalmagnitude evaluation stage whereby numbersaccess an analogical representation based on spatialproximity (Restle, 1970; Dehaene, 1992, 1997).Our experiment therefore tested whether spatialneglect would selectively impair comparisons madefor numbers located leftward from the referencepoint along the “mental number line”.
Methods
Participants included seven patients with righthemisphere damage (RHD) who had left neglect (6males, mean age 61.2 ± sd 12); seven patients withRHD who had no clinical sign of neglect (5 males,mean age 64.7 ± sd 9); and seven healthy controlsubjects (4 males, mean age 41.0 ± sd 8). Allpatients with RHD had suffered a recent focal strokein the middle or posterior cerebral artery territory(12 ischemic, 2 hemorrhagic, all ≤ 3 months post-onset). Lesions were confirmed by either CT or MRIscans in all cases. All patients were tested onstandard clinical tests (see Mayer et al., 1999)including line bisection, cancellation, reading,drawing, and figural fluency tasks. Neglect wasdetermined by the presence of left-sided omissionsin at least two or more of these tests, whereasabsence of neglect was determined by a lack ofomissions in all tests. Two patients in the neglectgroup (NEG) had left hemianopia and two othershad left visual extinction, whereas three patients inthe group without neglect (NONEG) had lefthemianopia and none visual extinction. No patientshowed neglect in visual mental imagery as testedby the description of familiar places (see Ortigue etal., 2001), and none of them had dyscalculia instandard clinical tests including basic arithmeticoperations with 1 and 2-digit numbers (see Mayer etal., 1999). Normal controls (CONT) had no historyof past neurological or psychiatric disease.
All participants were tested on the number sizecomparison task in one or two sessions. Stimuli
2 Patrik Vuilleumier and Others

were single digits (~ 2° of visual angle) presentedeach in turn at the centre of a computer screen (~50 cm in front of the subject). The task was tojudge as quickly as possible whether the digit wassmaller or larger than FIVE. All digits from 1 to 9,except 5, were shown in random order, precededby a fixation cross (700 msec), and remained onthe screen until a response was made. This wasfollowed by an interval of 1 sec before the nexttrial. Each digit was presented 12 times, resultingin a total of 96 trials per blocks.
Responses were made by key-presses, with onekey assigned to “smaller” and the other to “larger”judgments. Since several patients had left-sideparesis due to their stroke, all subjects wererequired to use their right hand. To minimize therole of a simple SNARC effect due to using theright hand (i.e., generally slower responses to “left-sided” numbers below 5) (see Dehaene et al., 1993;Bächtold et al., 1998), most subjects were tested ontwo separate blocks using reversed key assignments(e.g., first with a left-sided key for “smaller”responses and then with a left-sided key for“larger” responses). Only two patients with neglectand two other without neglect had to be tested onlyon the most natural response mapping (“smaller”responses on left key) because of a failure toswitch to the alternative mapping (i.e. with manyerrors by persevering on the previously learnedresponse key assignment). However, including thisresponse-key factor or excluding these patientsfrom analysis did not affect the basic resultsdescribed below. All participants received 10practice trials before each block. Nothing in thetask instructions emphasized any spatial aspect inperformance or in imagined number position.
Results
The mean number of errors in each block and themean response times (RTs) for correct answers toeach digit (1-4 and 6-9) were calculated for eachsubject. Errors occurred on less than 3.7% of trialsin all subjects and did not differ between patientswith neglect (mean 1.3 ± sd 1.6), patients withoutneglect (2.2 ± 2.3) and normal controls (2.1 ± sd 1.3).An ANOVA on these error rates indicated nosignificant difference between the three groups [F (2, 32) = 1.58, p = .24], but a main effect ofresponse-key mapping [F (1, 32) = 11.0, p = .002]due less errors occurring with the more compatible(“smaller” on left side, mean 1.14) than incompatibleassignment (“smaller” on right side, mean 2.7) in allgroups. There was a no interaction between groupand response mapping [F (2, 32) = 0.73].
RTs on correct trials are shown in Figure 1 as afunction of the number stimulus, collapsed acrossall response-key mappings. A striking difference inthe pattern of RTs was observed between the threegroups. Not only were NEG patients generallyslower than NONEG patients and NONEG patientsin turn slower than CONT subjects, but morecritically NEG patients showed a uniquely steepincrease in RTs for numbers close to the centralreference “5”. Most strikingly, NEG patients hadparticularly slower RTs for the number “4” whichwould be immediately positioned to the left of “5”on the mental number line. This seems consistentwith a lateralized impairment in the mentalrepresentation of numbers on the left side of areference point along the number line, despite thefact that all stimuli were presented at the centre ofthe screen in the absence of any other competing
Neglect in number space 3
Fig. 1 – Experiment 1 (A) and 2 (B). Mean response times (RTs) in the number size judgment tasks where each number had to beclassified as larger or smaller than 5 (A), or as larger or smaller than 7 (B). RTs are collapsed across the different response-key mappingconditions. RHD = right hemisphere-damaged patients, LHD = left hemisphere-damaged patients.
stimulus and without any specific instruction to usespatial imagery. This effect was not observed inNONEG patients and CONT subjects who onlyexhibited a mild and symmetrical RT increasearound the central “5”, consistent with a normaldistance effect in such number comparison tasks(Dehaene et al., 1998). This pattern wascorroborated by a mixed ANOVA showing maineffects of group [F (2, 32) = 39.9, p < .001] andnumber stimulus [from 1-4 and 6-9, F (7, 224) =15.4, p < .001], together with a highly significantinteraction between group and number [F (14, 224)= 7.1, p < .001]. Overall, the RT difference to “4”versus “6” was > 150 msec in 5/7 NEG, but noneof NONEG and CONT subjects.
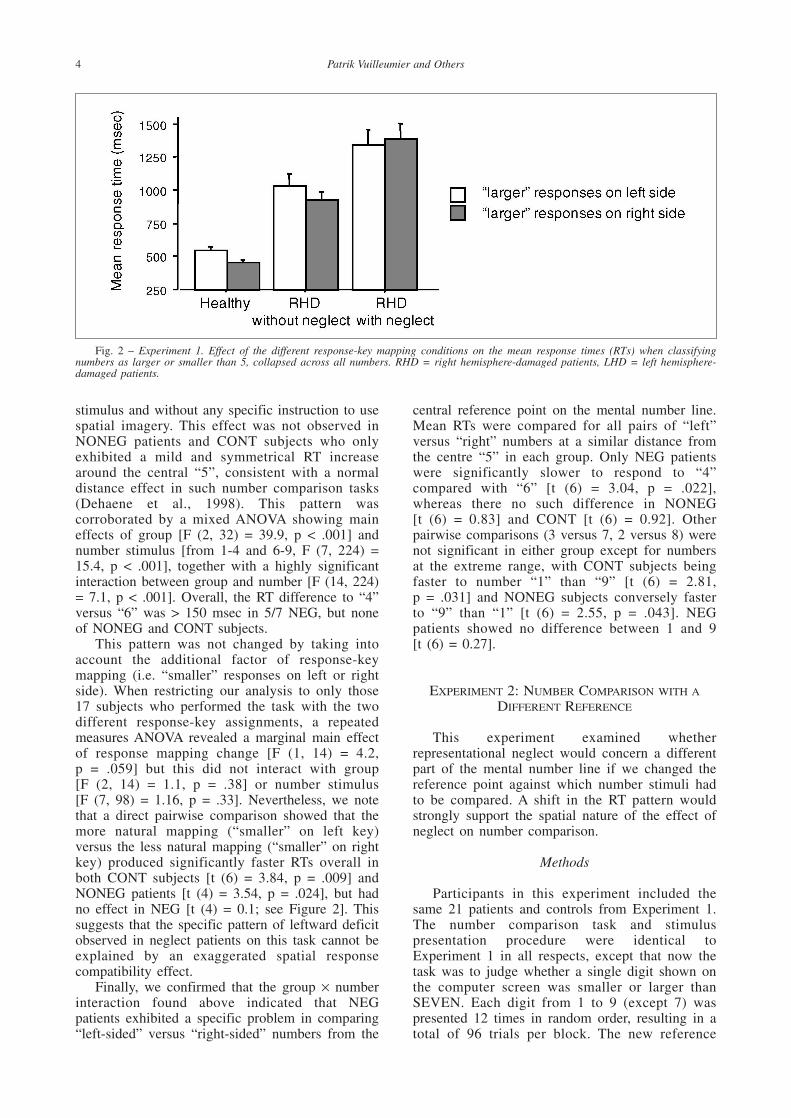
This pattern was not changed by taking intoaccount the additional factor of response-keymapping (i.e. “smaller” responses on left or rightside). When restricting our analysis to only those17 subjects who performed the task with the twodifferent response-key assignments, a repeatedmeasures ANOVA revealed a marginal main effectof response mapping change [F (1, 14) = 4.2, p = .059] but this did not interact with group [F (2, 14) = 1.1, p = .38] or number stimulus [F (7, 98) = 1.16, p = .33]. Nevertheless, we notethat a direct pairwise comparison showed that themore natural mapping (“smaller” on left key)versus the less natural mapping (“smaller” on rightkey) produced significantly faster RTs overall inboth CONT subjects [t (6) = 3.84, p = .009] andNONEG patients [t (4) = 3.54, p = .024], but hadno effect in NEG [t (4) = 0.1; see Figure 2]. Thissuggests that the specific pattern of leftward deficitobserved in neglect patients on this task cannot beexplained by an exaggerated spatial responsecompatibility effect.
Finally, we confirmed that the group × numberinteraction found above indicated that NEGpatients exhibited a specific problem in comparing“left-sided” versus “right-sided” numbers from the
central reference point on the mental number line.Mean RTs were compared for all pairs of “left”versus “right” numbers at a similar distance fromthe centre “5” in each group. Only NEG patientswere significantly slower to respond to “4”compared with “6” [t (6) = 3.04, p = .022],whereas there no such difference in NONEG [t (6) = 0.83] and CONT [t (6) = 0.92]. Otherpairwise comparisons (3 versus 7, 2 versus 8) werenot significant in either group except for numbersat the extreme range, with CONT subjects beingfaster to number “1” than “9” [t (6) = 2.81, p = .031] and NONEG subjects conversely fasterto “9” than “1” [t (6) = 2.55, p = .043]. NEGpatients showed no difference between 1 and 9 [t (6) = 0.27].
EXPERIMENT 2: NUMBER COMPARISON WITH A
DIFFERENT REFERENCE
This experiment examined whetherrepresentational neglect would concern a differentpart of the mental number line if we changed thereference point against which number stimuli hadto be compared. A shift in the RT pattern wouldstrongly support the spatial nature of the effect ofneglect on number comparison.
Methods
Participants in this experiment included thesame 21 patients and controls from Experiment 1.The number comparison task and stimuluspresentation procedure were identical toExperiment 1 in all respects, except that now thetask was to judge whether a single digit shown onthe computer screen was smaller or larger thanSEVEN. Each digit from 1 to 9 (except 7) waspresented 12 times in random order, resulting in atotal of 96 trials per block. The new reference
4 Patrik Vuilleumier and Others
Fig. 2 – Experiment 1. Effect of the different response-key mapping conditions on the mean response times (RTs) when classifyingnumbers as larger or smaller than 5, collapsed across all numbers. RHD = right hemisphere-damaged patients, LHD = left hemisphere-damaged patients.

number “7” was chosen so that the number 6would fall in a relative leftward position in thisexperiment but in a relative rightward position inthe previous experiment, and hence be directlycompared across the two tasks.
This experiment was given on the same day asExperiment 1 in most subjects, or on a differentsession a few days apart in a few patients. Again,two separate blocks were given in which response-key mapping (“smaller” responses on left versusright side) was systematically varied, except in twoNEG and two CONT patients who failed to switchto a new mapping after performing the first block.
Results
Mean number of errors in each block and meanresponse times (RTs) for correct answers to eachdigit (1-6 and 8-9) were calculated as in theprevious experiment. Errors occurred on less than4.5% of trials in all subjects and did not differbetween NEG patients (mean 1.8 ± sd 2.8),NONEG patients (1.0 ± 0.8) and normal CONTsubjects (2.0 ± sd 1.8). An ANOVA indicated nosignificant effect or interaction due to group [F (2, 32) = 1.16] or response-key mapping [F (1, 14) = 0.42].
The pattern of correct RTs again revealed astriking difference between the patients withneglect and the other two groups (Figure 1). As inthe previous experiment, NEG patients were notonly slower than NONEG and CONT for allnumbers, but also exhibited a disproportionateincrease in their RTs to numbers “5” and “6”,particularly compared with “8” and “9”. This isagain suggestive of a specific difficulty inresponding to numbers situated immediately to theleft of the reference “7” along the mental numberline. A mixed ANOVA confirmed significant maineffects of group [F (2, 32) = 40.6, p < .001] andnumber stimulus [1-6 and 8-9, F (7, 224) = 9.4, p < .001], together with a significant interactionbetween group and number [F (14, 224) = 4.5, p < .001]. Similarly as in experiment 1, the RTdifference to “6” versus “7” was > 150 msec in 5/7NEG, but none of NONEG and CONT subjects.
Again, the additional factor of response-keymapping did not affect this pattern of results whenconsidering subjects tested under the two differentkey assignment conditions, although it produced asignificant main effect [F (1, 14) = 11.0, p = .005]due to slower RTs with the less compatible(“smaller” response on right key) compared tocompatible mapping (“smaller” response on leftkey) in all three groups. There was no significanttwo-way interaction of response mapping withnumber, but a three-way interaction of responsemapping × number × group [F (14, 224) = 4.14, p < .001]. Post-hoc tests showed that this was dueto the incompatible versus compatible mappingslowing responses for all numbers in NONEG
patients [main effect, F (1, 4) = 12.2, p = .025],but not in CONT subjects whose responses wereslowed specifically for number “6” [main effect ofmapping: F (1, 4) = 3.6, p = .105; mapping ×number interaction: F (7, 42) = 3.11, p = 009]. Bycontrast, NEG patients showed no significant effector interaction of response mapping (p ≥ .22). Thisis similar to Experiment 1 and suggests that thedeficit observed in neglect for numbers to the leftof “7” was not due to an exaggerated responsecompatibility effect due to using the right hand.
We also compared mean RTs for pairs of “left”versus “right” numbers at a similar distance fromthe center “7” in each group. NEG patients wereslower to respond to “6” compared with “8” [t (6) = 3.56, p = .012] and to “5” compared with“9” [t (6) = 3.67, p = .002]. By contrast, NONEGpatients were slower only for “5” compared with“9” [t (6) = 2.96, p = .025], and CONT showed noasymmetry for either pairs [t (6) < 1.17, p < .29].
Finally, we also directly examined RTs for thenumber “6” in this and the previous experiment, asthis same number fell to the right of the reference“5” in the first task but to the left of reference “7”in the second task. Responses to number “6” weresignificantly slower in the second versus firstexperiment in NEG patients [mean 1547 vs. 1349ms, t (6) = 5.38, p = .001], but not in NONEG[1216 vs. 1093 ms, t (6) = 1.49, p = .18] andCONT subjects [503 vs. 556 ms, t (6) = 1.34, p = .22]. Such RT changes in NEG occurred purelydue to changes in task requirements as presentationof the number (at the centre of the screen) and itsrelative probability (1:8) were exactly the same inthe two experiments.
EXPERIMENT 3: STROOP SIZE EFFECTS
The preceding results show that patients withleft neglect are impaired at making number sizecomparisons when the stimulus to be judged issmaller than a reference point, and hencepresumably located on a relative leftward positionalong the mental number line. This would beconsistent with a form of representational neglectfor numerical entities coded in a spatial manner.Patients might fail to orient attention to the left ofa currently relevant reference frame (Restle, 1970),as happens for other perceptual and imagerydeficits in neglect (Bisiach and Luzzatti, 1978;Bisiach, 1993). Alternatively, one could supposethat representational neglect for numbers in the sizecomparison tasks might result from a distortion ofquantities represented on the left side of the mentalline, and thus concern to some degree all left-sidednumbers along the line, irrespective of task andreference point. Experiment 3 tested this idea usinga Stroop-like interference task in which numericalquantity (small or large number) could becongruent or incongruent with physical features of
Neglect in number space 5

the stimulus (small or large character). We askedwhether an impaired representation of numberquantity would cause a selective loss of Stroopeffects, affecting numbers at a relative leftwardposition (e.g. below 5) more than numbers in arelative rightward position (e.g. above 5).
Methods
Participants in this experiment included thesame patients and controls as Experiment 1.Stimuli included all digits (1 to 9) and couldappear either as a small (~ 1.5° of visual angle) orlarge character (~ 4° of visual angle). Digits werepresented each in turn at the centre of a computerscreen, preceded by a central fixation cross (700ms). Each digit from 1 to 9 appeared 12 times.Half had a small size and half a large size,resulting in a total of 108 trials per block, given inrandom order. The task was to judge whether thesize of the character was physically small or large,by pressing one of two response keys as quickly aspossible. Numerical size had to be ignored. Stimuliremained on the screen until a response was made.Each trial was preceded by a blank interval of 1sec. All participants received 10 practice trialsbefore test blocks.
Results
Errors were slightly more frequent in bothpatient groups than normal controls (NEG mean:6.4 ± sd 2.6; NONEG: 4.0 ± sd 2.4; CONT: 2.8 ±sd 1.4). However, an ANOVA of error ratesshowed that the factor of group did not produce areliable effect [F (2, 18) = 1.127, p = .31], whereas
the factor of physical/numerical congruencyshowed a significant main effect [F (1, 18) = 6.58, p = .019] without any interaction with the factor ofnumber range [below or above 5, F (1, 18) = 2.98, p = .11) or with group F (2, 18) = .39]. Thus,errors were more frequent when the physical sizeof the stimulus was incongruent versus congruentwith numerical quantity (mean 3.1 ± sd 2.1 versus6.5 ± sd 2.4 across groups).
RTs on correct responses showed a similarStroop-like effect (Figure 3). All participants werefaster when judging the physical size of stimuliwhose numerical quantity was congruent with size(e.g. a small “1” or a big “9”), as opposed toincongruent (e.g. a big “1” or a small “9”). AnANOVA on RTs using the factors of group,physical/numerical congruency, and stimulus type(number 1 to 9) confirmed significant main effectsof congruency [F (2, 18) = 4.87, p < .001] andgroup [F (1, 18) = 11.1, p < .001], without anyother effects and interaction. The same results wereobtained when considering only the dichotomousfactor of number range (below versus above five,[F (1, 18) = 0.8]. This intact Stroop size effects inNEG patients indicate that representational neglectfor numbers does not affect the access to numericalquantities, but more probably a spatialrepresentation of imagined number position.
EXPERIMENT 4: SNARC AROUND THE CLOCK
This experiment further examined whetherrepresentational neglect for numbers could concerna different part of the mental number line, and affectlarger numbers (i.e., above 6) more than smaller
6 Patrik Vuilleumier and Others
Fig. 3 – Experiment 3. Mean response times (RTs) in the Stroop task, where each number had to be classified according to itsphysical size on the screen (large or small character), showing an interference as a function of congruency with the numerical size(larger or smaller than 5). RHD = right hemisphere-damaged patients, LHD = left hemisphere-damaged patients.

numbers (i.e. below 6) if the task implied to codenumber position in a reverse left-right orientation.To this end, we used the Clock Task (Bächtold etal., 1998) which has previously been designed totest SR-compatibility in representational space inhealthy people and requires subjects to classifynumbers as indicating hours earlier or later than sixo’clock. The original version of the Clock Task(Bächtold et al., 1998) revealed a reaction timeadvantage of the left hand for responding to largenumbers (representing hours on the left side of theclock face) and of the right hand for small numbers(on the right half of the clock face).
Methods
We tested a new group of 26 right-handedpatients with a single unilateral cortical lesioninvolving the parietal cortex of either hemisphere(18 RHD, 8 LHD, none of them with any signs ofaphasia or dyscalulia). All lesions were shown byCT or MRI to be largely confined to posteriorparietal regions. Of the RHD patients, 10 (4women; mean age 53.8 ± sd 12.5) showed signs ofleft-sided neglect, whereas 8 (1 woman; mean age56.2 ± sd 12.7) did not. Of the 8 LHD patients (2women; mean age 55.6 ± sd 10.3), none showedsigns of right-sided neglect on any of the clinicaltasks. These tasks were similar to those used toassess neglect in the patients participating inExperiments 1 to 3 and assessed line bisection,reading, drawing, and figural fluency performance.A new group of 40 right-handed elderly volunteerswithout past neurological or psychiatric history wasalso tested, including 20 women (mean age = 55.4years ± sd 8.3 years) and 20 men (mean age = 56.7years ± sd 6.5 years).
The Clock Task required unimanual responsesto centrally displayed arabic numerals ranging from1 to 11 (without number 6). Participants wereasked to decide, as fast and as accurately aspossible, whether the presented numbercorresponded to an hour earlier or later than sixo’clock. Two runs were administered, one with a“COMPATIBLE” spatial mapping of responseeffector with side of the clock face (i.e. left-sidekey for earlier than 6, right-side key for later than6), and one with an “INCOMPATIBLE” responsemapping (right key for earlier than 6, left key forlater than 6). In all subjects, response effectorswere the first (index) and third (ring) fingers ofone hand (always the ipsilesional for the patients).Thus, all subjects used their dominant right hand,except LHD patients who all used their left hand toprevent potentially confounding effects of righthand weakness. All participants were tested on thetwo runs within the same day. Stimuli were shownfor 1 sec at the centre of a computer screen(Helvetica, ~ 2° visual angle). Within each run(“COMPATIBLE” and “INCOMPATIBLE”), eachdigit was presented 10 times in pseudorandom
order, for a total of 100 trials per condition. Theorder of runs was counterbalanced within eachsubject group. All participants received 20 practicetrials before each run, but were not trained using adisplay explicitly representing a clockface image(unlike in Bächtold et al., 1998).
Results
The number of correct decisions was computedfor each response compatibility condition in eachparticipant. Accuracy was high in the healthycontrol subjects (mean: 99% correct ± sd 3 forcompatible run, versus 98% ± sd 2 forincompatible run). All three patient groups wereless accurate but still clearly above chance in boththe compatible run (92% ± sd 9%, 89% ± sd 11%,and 95% ± sd 6% correct decisions for LHD-patients, RHD-patients with and RHD-patientswithout neglect, respectively) and the incompatiblerun (84% ± sd 17%, 79% ± sd 20%, and 85% ± sd14% correct, respectively).
The critical results concerned RTs of correctdecisions, computed for each responsecompatibility condition and each side of the clockin each participant. In a first step, we assessedwhether hemispheric lesion side differentiallyaffected RTs to numbers on the left (RT(7-11)) andright side of a clock face (RT(1-5)). RTs were thussubject to a repeated measures ANOVA withGROUP (RH-patients, LH-patients and normalcontrols) as the between subject factor, and SR-COMPATIBILITY (compatible run vs.incompatible run) and SIDE OF IMAGINALCLOCK FACE (left vs. right) as the repeatedmeasures. The results (Figure 4A) revealed a maineffect of GROUP [F (2, 63) = 42.0, p <. 001],mean RTs were 1044 ms (sd = 337 ms) for RH-patients, 723 ms (sd = 204 ms) for LH-patients and600 ms (sd = 107 ms) for normal controls. Bothpatient groups were slower than normal controls(RH-patients: t = 8.8, p < .001, LH-patients: t = 2.3, p < .03), and RH-patients slower than LH-patients (t = 3.0, p < .01). The main effect of SR-compatibility was also significant [F (1, 63) = 15.4,p < .001, due to RTs being overall longer in theincompatible run], as was the main effect of SIDE [F (1, 63) = 21.4, p < .001; due to RT(7-11) beinggenerally longer than RT(1-5)].
Importantly, the interactions between GROUPand SR-compatibility [F (2, 63) = 9.3, p < .001]and GROUP and SIDE OF CLOCK FACE [F (2, 63) = 11.5, p < .001] were significant aswell. This was mainly due to RH-patients showinggreater RT increases in the incompatible run andfor numbers on the left side of the clock (see TableI for single group comparisons). Finally, theinteraction between SR-compatibility and SIDEwas not significant [F (1, 63) = .23, p > .6], whilethe triple interaction between GROUP, SR-compatibility, and SIDE approached significance
Neglect in number space 7

8 Patrik Vuilleumier and Others
Fig. 4 – Experiment 4. (A) Mean reaction times (RTs) when numbers had to be classified as earlier or later than 6 o’clock, as afunction of the response compatibility mapping (compatible runs = right and left hand for later and earlier, respectively; incompatibleruns = left and right hand for later and earlier, respectively). (B) Mean RTs in patients with right brain lesions as a function of number,showing that presence of neglect was associated with slower responses to hours on the left-side of the clock (later than 6) compared withhours on the right side (earlier than 6). RHD = right hemisphere-damaged patients, LHD = left hemisphere-damaged patients.
TABLE II
Means and Standard Deviations of Response Side and Clockface Side Effects for the Four Participant Groups in Experiment 4
Magnitude of RT effect Healthy controls LHD patients RHD patients
Without neglect With neglect
Response compatibility (RTincompatible – RTcompatible) 16.4 68.0 283.5** 201.0*(SD = 78) (SD = 279) (SD = 204) (SD = 328)
Clockface side (RT7-11 – RT1-5) 12.7** 22.6 41.3 144.8**(SD = 31) (SD = 67) (SD = 88) (SD = 99)
All values are in msec. Statistical significance is marked by asteriks: *p < .1; **p < .05; ***p < .01; paired t-tests observed versus expected (i.e., zero) values,all p-values two-tailed.

[F (2, 63) = 2.8, p < .07; due to largercompatibility effects for the right side of clock facein LHD patients but not other groups].
In a second step, we assessed whether, withinthe group of patients with RH lesions, presence orabsence of neglect differentially affected RT(7-11)and RT(1-5), respectively. RTs of correct decisionswere subject to a repeated measures ANOVA withNEGLECT (present vs. absent) as the betweensubject factor, and SR-compatibility (compatiblerun vs. incompatible run) and SIDE OF CLOCKFACE (left vs. right) as the repeated measures.This revealed a main effect of SR-compatibility [F (1, 16) = 13.3, p < .005, with longer RTs in theincompatible run] and a main effect of SIDE [F (1, 16) = 17.3, p < .001, with RT(7-11) longerthan RT(1-5)]. Critically, the main effect ofNEGLECT was not significant [F (1, 16) = 1.1, p > .3] but the interaction between NEGLECT andSIDE was [F (1, 16) = 5.4, p < .05]. Over bothruns, patients without neglect had comparable RTsto small and large numbers (t = 1.3, p > .2),whereas patients with neglect were much slower tolarge than to small numbers (t = 4.6, p < .005;Figure 4B), that is, to numbers positioned on theleft-side of the clock. No other interactionapproached significance.
DISCUSSION
The present findings demonstrate that unilateralspatial neglect may impair simple numberprocessing tasks in a way that suggests a spatiallylateralized disturbance in the mental representationof numbers. This occurred even though stimuliwere presented at fixation and the tasks did notexplicitly require the use of a spatial strategy.When asked to judge whether numbers weresmaller or larger than “5”, patients with neglectwere much slower to make a response to “4”compared with “6” and other higher numbers, butwhen asked to judge numbers as smaller or largerthan “7” they were now much slower to respond to“6” compared with higher numbers. Such effectsseem consistent with a difficulty to orient attentiontoward an internal representation of numberslocated on the left side of a relevant referencepoint along the “mental number line”. IncreasedRTs for “left-sided” numbers predominantlyaffected those immediately preceding the referencepoint (“4” in the 5-task; “6” in the 7-task), whichis consistent with a classical distance effect incomparison tasks whereby the discriminationbetween two numbers becomes harder as theirnumerical distance decreases (Moyer and Landauer,1967; Dehaene, 1989; Dehaene et al., 1998). Bycontrast, in the Stroop task requiring judgments ofphysical size instead of numerical magnitude, ourneglect patients showed no significant deficit as afunction of number range that would indicate an
abnormal representation of quantity for smallnumbers (“left-sided” on the mental line); and inthe Clock task requiring to judge numbers asreferring to hours rather than to cardinality, neglectpatients showed an inverse pattern with slower RTsfor larger versus smaller numbers, respectivelylocated on the left versus right side of an imaginedclock face. Taken together, these results provideevidence that spatial neglect may cause arepresentational deficit that specifically affects themental manipulation of number based on positionalproximity, rather than the abstract knowledge ofnumerical quantities.
Our findings converge with other recent resultssuggesting that neglect in right-brain damagepatients may disrupt the mental bisection ofnumerical intervals (Zorzi et al., 2002) and thatnumber processing in healthy subjects can inducespatial biases in the bisection of physical lines(Fischer, 2001). In addition, our data indicate thatinformation about relative position (i.e., “4”precedes “5” and “6” precedes “7” along themental number line) must still be normallyrepresented despite right hemisphere damage andleft spatial neglect. This is suggested by theselective slowing of responses to numbersimmediately preceding the reference points in the5-task and 7-task. These findings not only lendsupport to recent proposals that spatial codes areautomatically activated when humans perform tasksrequiring the processing of numerical quantity(Dehaene, 1992, 1997), but also bolster the earlieridea that certain mental operations on numbers mayinvolve literal shifts of attention acting on aspatially-organized internal image (Restle, 1970). Asimilar role of lateralized mental images is likely toaccount for spatial compatibilty effects in normalsubjects’ performance in the Clock task (Bächtoldet al., 1998) and representational deficits in neglectpatients (Bisiach and Luzzatti, 1978; Bisiach,1993).
Our results in neglect patients also reveal thatthe activation of spatial codes during numberprocessing is not entirely obligatory, but dependson the nature of the task and the currently relevantframe of reference. First, spatial neglect affectedperformance only when the task requiredjudgements of numerical magnitude or relativetime, but not with judgements of physical size.Second, spatial neglect affected different numbersduring comparison in the 5-task or 7-task. This isconsistent with other observations that spatialneglect can be manifested for different parts of thesame mental representation depending on taskinstructions or imagined viewpoint (Bisiach andLuzzatti, 1978; Meador et al., 1987; Bisiach, 1993;Ortigue et al., 2001). Thus, in the first report ofrepresentational neglect, Bisiach and Luzzattialready described that asking patients to describe ascene from memory from a given vantage pointcould reveal neglect for the contralesional half of
Neglect in number space 9

the scene, although the same information could beprovided if the patient was then asked to describethe scene from an opposite viewpoint. Our findingsof a relative representational neglect for number“4” in the 5-task, but for “6” in the 7-task,illustrate a similar phenomenon in a seeminglymore abstract numerical space. Both the simplenumber comparison and clock tasks mightconstitute particularly useful tests forrepresentational neglect, as a spatial representationof number magnitude (Dehaene, 1997) and analogtimes (Boroditsky, 2000) might both be universal.Theses tests may also be more sensitive than othertraditional tests of representational neglect, sincenone of the neglect patients in our study showedany significant signs of imaginal neglect whentested on scene or map description (Bartolomeo etal., 1994; Rode et al., 1995; Ortigue et al., 2001).Moreover, in both experiments 1 and 2, a RTdifference to “left” versus “right” sided numbersexceeded 150 msec in 70% of patients withneglect, but none of other subjects.
A spatial impairment in number processing inneglect patients is consistent with a closefunctional relationship between the representationof numerical quantity and visuospatial processesmediated by right parietal cortex (Dehaene et al.,1998; Chochon et al., 1999; Göbel et al., 2001).Functional imaging studies have consistentlyshown activation of right intraparietal cortex duringnumber comparison tasks similar to ours (Chochonet al., 1999; Pinel et al., 1999; Pinel et al., 2001).Similar regions are also activated by mentalimagery tasks involving the representation of aclockface (Formisano et al., 2002), as well as byshifts of spatial attention (Gitelman et al., 1999;Corbetta et al., 2000). However, our study does notaddress the issue of whether distinct versusoverlapping regions in parietal cortex mediatenumber processing and other visuospatial functions(Simon et al., 2002). Brain lesions in our patientswere usually large and likely to encompass morethan a single parietal sub-region. We incidentallynote that among our neglect patients, two who hadpredominant subcortical lesions showed the samepattern as two others with more focal parietaldamage, but remote parietal diaschisis cannot beexcluded in such cases (Perani et al., 1987).
We finally note an apparently paradoxicaldifference between the number comparison tasks(exp. 1 and 2) and the clock task (exp. 4) forspatial response compatibility effects. In thenumber tasks, neglect patients showed reducedcompatibility effects when key-press responseswere made according to a more natural mapping(“smaller” versus “larger” responses on left versusright keys, respectively). In the clock task,however, neglect patients showed preserved oreven exaggerated compatibility effects. Thisdifference could be related either to the differentnature of the tasks or to different patients tested in
each conditions. It might be a consequence of amore ambiguous SR mapping in the clock task,with a conflict between the traditional SNARCeffect (“larger” responses on the right side) and itsreversal in this task (see Bächtold et al., 1998).Alternatively, one might speculate that damage toparietal networks that are implicated in the mentalmanipulation of numerical quantities might alsoweaken response compatibility effects based onleft-to-right orientation for numbers but not forother meaningful objects such as a clock face,which can presumably be represented in othervisual areas. In any case, it must be emphasizedthat in none of the tasks employed in the presentstudy did such response compatibility interact withthe effects of stimulus number and representationalneglect.
In summary, the current results demonstrate thatrepresentational neglect can extend to abstractnumber tasks and thus support the idea that spatialcoding might provide an important foundation forthe development of numerical cognitive abilities.As foreseen by the work of Bisiach and colleagues(Bisiach et al., 1985; Bisiach et al., 1990; Bisiachand Berti, 1995), these findings underscore theimportance of representing space for acting notonly on the external world but also on moreabstract mental contents.
Acknowledgements. We thank Daniel Bächtold for hishelp in testing the patients in Experiment 4; WernerSurbeck and Veronika Lister for collecting normative data;Marianne Regard for her blind scoring of patients’ neglect;and Brian Butterworth, Stanislas Dehaene, Jon Driver, andManuella Piazza for discussion and comments. This workwas in part supported by grants from the Swiss NationalScience Foundation to PV (632-65935.01) and PB (3100-051057.97/1).
REFERENCES
BÄCHTOLD D, BAUMÜLLER M and BRUGGER P. Stimulus-responsecompatibility in representational space. Neuropsychologia, 36:731-735, 1998.
BARTOLOMEO P, D’ERME P and GAINOTTI G. The relationshipbetween visuospatial and representational neglect. Neurology,44: 1710-14, 1994.
BENSON DF and WEIR WF. Acalculia: Acquired anarithmetia.Cortex, 8: 465-72., 1972.
BESCHIN N, BASSO A and DELLA SALA S. Perceiving left andimagining right: Dissociation in neglect. Cortex, 36: 401-14,2000.
BISIACH E. Mental representation in unilateral neglect and relateddisorders: The twentieth Bartlett Memorial Lecture. QuarterlyJournal of Experimental Psychology, 46A: 435-462, 1993.
BISIACH E and BERTI A. Consciousness in dyschiria. In GAZZANIGAM, The cognitive neurosciences, 1331-40, Cambridge, MA:MIT Press, 1995, pp.
BISIACH E, BERTI A and VALLAR G. Analogical and logicaldisorders underlying unilateral neglect of space. In POSNER MIand MARIN OS, Attention & Performance, vol. 9, 239-246,Hillsdale, NJ: Erlbaum, 1985, pp.
BISIACH E and LUZZATTI C. Unilateral neglect of representationalspace. Cortex, 14: 129-133, 1978.
BISIACH E, LUZZATTI C and PERANI D. Unilateral neglect,representational schema and consciousness. Brain, 102: 609-618, 1979.
BISIACH E, MEREGALLI S and BERTI A. Mechanisms of productioncontrol and belief fixation in human visuospatial processing:Clinical evidence from unilateral neglect and
10 Patrik Vuilleumier and Others

misrepresentation. In Computational and clinical approachesto pattern recognition and concept formation: Quantitativeanalyses of behavior, vol. 9, edited by ML COMMONS, RGHERRNSTEIN, SM KOSSLYN and DB MUMFORD, pp. 3-21,Erlbaum, Hillsdale, NJ, 1990.
BISIACH E and VALLAR G. Hemineglect in humans. In F BOLLERand J GRAFMAN, Handbook of Neuropsychology, vol. 1, 195-222, Amsterdam: Elsevier, 1988, pp.
BORODITSKY L. Metaphoric structuring: Understanding timethrough spatial metaphors. Cognition, 75: 1-28., 2000.
BRANNON EM. The development of ordinal numerical knowledgein infancy. Cognition, 83: 223-40., 2002.
BRANNON EM and TERRACE HS. Ordering of the numerosities 1 to9 by monkeys. Science, 282: 746-9, 1998.
BUTTERWORTH B. The Mathematical Brain, London: Macmillan,1999.
CHOCHON F, COHEN L, VAN DE MOORTELE PF and DEHAENE S.Differential contributions of the left and right inferior parietallobules to number processing. J Cogn Neurosci, 11: 617-30,1999.
CIPOLOTTI L, BUTTERWORTH B and DENES G. A specific deficit fornumbers in a case of dense acalculia. Brain, 114: 2619-37,1991.
COHEN L, DEHAENE S, CHOCHON F, LEHERICY S and NACCACHE L.Language and calculation within the parietal lobe: Acombined cognitive, anatomical and fMRI study.Neuropsychologia, 38: 1426-40, 2000.
CORBETTA M, AKBUDAK E, CONTURO TE, SNYDER AZ, OLLINGERJM, DRURY HA, LINENWEBER MR, PETERSEN SE, RAICHLEME, VANESSEN DC and SHULMAN GL. A common network offunctional areas for attention and eye movements. Neuron, 21:761-773, 1998.
CORBETTA M, KINCADE JM, OLLINGER JM, MCAVOY MP andSHULMAN GL. Voluntary orienting is dissociated from targetdetection in human posterior parietal cortex. NatureNeuroscience, 3: 292-297, 2000.
COSLETT HB. Neglect in vision and visual imagery: A doubledissociation. Brain, 120: 1163-1171, 1997.
DEHAENE S. The psychophysics of numerical comparison: Areexamination of apparently incompatible data. PerceptPsychophys, 45: 557-66, 1989.
DEHAENE S. Varieties of numerical abilities. Cognition, 44: 1-42,1992.
DEHAENE S. The number sense, Oxford: Oxford University Press,1997.
DEHAENE S, BOSSINI S and GIRAUX P. The mental representation ofparity and number magnitude. J Exp Psychol Gen, 122: 371-396, 1993.
DEHAENE S and COHEN L. Two mental calculation systems: A casestudy of severe acalculia with preserved approximation.Neuropsychologia, 29: 1045-54, 1991.
DEHAENE S, DEHAENE-LAMBERTZ G and COHEN L. Abstractrepresentations of numbers in the animal and human brain.Trends Neurosci, 21: 355-61, 1998.
DEHAENE S, SPELKE E, PINEL P, STANESCU R and TSIVKIN S.Sources of mathematical thinking: Behavioral and brain-imaging evidence. Science, 284: 970-4, 1999.
DELAZER M and BENKE T. Arithmetic facts without meaning.Cortex, 33: 697-710, 1997.
DRIVER J and VUILLEUMIER P. Perceptual awareness and its loss in unilateral neglect and extinction. Cognition, 79: 39-88,2001.
FETTERMAN JG. Numerosity discrimination: both time and number matter. J Exp Psychol Anim Behav Process, 19: 149-64, 1993.
FINK GR, MARSHALL JC, GURD J, WEISS PH, ZAFIRIS O, SHAH NJand ZILLES K. Deriving numerosity and shape from identicalvisual displays. Neuroimage, 13: 46-55, 2001.
FISCHER MH. Number processing induces spatial performancebiases. Neurology, 57: 822-826, 2001.
FORMISANO E, LINDEN DE, DI SALLE F, TROJANO L, ESPOSITO F,SACK AT, GROSSI D, ZANELLA FE and GOEBEL R. Tracking themind’s image in the brain I: Time-resolved fMRI duringvisuospatial mental imagery. Neuron, 35: 185-94, 2002.
GITELMAN DR, NOBRE AC, PARRISH TB, LABAR KS, KIM YH,MEYER JR and MESULAM MM. A large-scale distributednetwork for covert spatial attention: Further anatomicaldelineation based on stringent behavioural and cognitivecontrols. Brain, 122: 1093-1106, 1999.
GÖBEL S, WALSH V and RUSHWORTH MFS. The mental numberline and the human angular gyrus. Neuroimage, 14: 1278-89,2001.
GRAFMAN J, PASSAFIUME D, FAGLIONI P and BOLLER F. Calculationdisturbances in adults with focal hemispheric damage. Cortex,18: 37-49, 1982.
GRUBER O, INDEFREY P, STEINMETZ H and KLEINSCHMIDT A.Dissociating neural correlates of cognitive components inmental calculation. Cereb Cortex, 11: 350-9, 2001.
GUARIGLIA C, PADOVANI A, PANTANO P and PIZZAMIGLIO L.Unilateral neglect restricted to visual imagery. Nature, 364:235-237, 1991.
KARNATH HO. New insights into the functions of the superiortemporal cortex. Nat Rev Neurosci, 2: 568-76, 2001.
KOSSLYN SM, DIGIROLAMO GJ, THOMPSON WL and M AN. Mentalrotation of objects verus hands: Neural mechanisms revealedby positron emission tomography. Psychophysiology, 35: 151-161, 1998.
MAYER E, MARTORY MD, PEGNA AJ, LANDIS T, DELAVELLE J andANNONI JM. A pure case of Gerstmann syndrome with asubangular lesion. Brain, 122: 1107-20, 1999.
MEADOR KJ, LORING DW, BOWERS D and HEILMAN KM. Remotememory and neglect syndrome. Neurology, 37: 522-526,1987.
MECK WH and CHURCH RM. A mode control model of countingand timing processes. J Exp Psychol Anim Behav Process, 9:320-34, 1983.
MESULAM MM. Spatial attention and neglect: Parietal, frontal andcingulate contributions to the mental representation andattentional targeting of salient extrapersonal events. PhilosTrans R Soc Lond B Biol Sci, 354: 1325-46, 1999.
MOYER RS and LANDAUER TK. Time required for judgments ofnumerical inequality. Nature, 215: 1519-20, 1967.
ORTIGUE S, VIAUD-DELMON I, ANNONI JM, LANDIS T, MICHEL C,BLANKE O, VUILLEUMIER P and MAYER E. Purerepresentational neglect after right thalamic lesion. AnnalsNeurology, 50: 401-4, 2001.
PERANI D, VALLAR G, CAPPA S, MESSA C and FAZIO F. Aphasia andneglect after subcortical stroke. A clinical/cerebral perfusioncorrelation study. Brain, 110: 1211-29, 1987.
PIAGET J. La genèse du nombre chez l’enfant, Neuchâtel:Delachaux & Niestlé S.A, 1941.
PINEL P, DEHAENE S, RIVIERE D and LEBIHAN D. Modulation ofparietal activation by semantic distance in a numbercomparison task. Neuroimage, 14: 1013-26, 2001.
PINEL P, LE CLEC HG, VAN DE MOORTELE PF, NACCACHE L, LEBIHAN D and DEHAENE S. Event-related fMRI analysis of thecerebral circuit for number comparison. Neuroreport, 10:1473-9, 1999.
RESTLE F. Speed of adding and comparing numbers. J Exp PsycholGen, 91: 191-205, 1970.
RODE G, PERENIN MT and BOISSON D. Négligence de l’espacereprésenté: Mise en évidence par l’évocation mentale de lacarte de France. Rev Neurol, 151: 161-164, 1995.
RUECKERT L, LANGE N, PARTIOT A, APPOLLONIO I, LITVAR I, LEBIHAN D and GRAFMAN J. Vizualizing cortical activationduring mental calculation with fMRI. Neuroimage, 3: 97-103,1996.
SATHIAN K, SIMON TJ, PETERSON S, PATEL GA, HOFFMAN JMand GRAFTON ST. Neural evidence linking visual object enumeration and attention. J Cogn Neurosci, 11: 36-51,1999.
SERON X, PESENTI M, NOEL MP, DELOCHE G and CORNET JA.Images of numbers, or “When 98 is upper left and 6 skyblue”. Cognition, 44: 159-96, 1992.
SIMON O, MANGIN JF, COHEN L, LE BIHAN D and DEHAENE S.Topographical layout of hand, eye, calculation and language-realted areas in the human parietal lobe. Neuron, 33: 475-487,2002.
SIMON TJ. Reconceptualizing the origins of number knowledge: A“non-numerical” account. Cognit Dev, 12: 349-372, 1997.
STANESCU-COSSON R, PINEL P, VAN DE MOORTELE PF, LE BIHAN D,COHEN L and DEHAENE S. Understanding dissociations indyscalculia: A brain imaging study of the impact of numbersize on the cerebral networks for exact and approximatecalculation. Brain, 123: 2240-55, 2000.
VALLAR G. The anatomical basis of spatial neglect in humans. InROBERTSON IH and MARSHALL JC, Unilateral Neglect:Clinical and Experimental Studies, 27-62, Hillsdale, N.J.:Lawrence Erlbaum Associates, Publish, 1993, pp.
VALLAR G. Spatial hemineglect in humans. Trends in CognitiveScience, 2: 87-97, 1998.
VUILLEUMIER P and RAFAL R. “Both” means more than “two”:Localizing and counting in patients with visuospatial neglect.Nat Neurosci, 2: 783-4, 1999.
Neglect in number space 11

WYNN K. Psychological foundations of number: Numericalcompetence in human infants. Trends Cogn. Sci, 2: 296-303,1998.
ZACKS J, RYPMA B, GABRIELI JD, TVERSKY B and GLOVER GH.Imagined transformations of bodies: An fMRI investigation.Neuropsychologia, 37: 1029-40, 1999.
ZORZI M, PRIFTIS K and UMILTA C. Brain damage: Neglect disruptsthe mental number line. Nature, 417: 138-9, 2002.
P. Vuilleumier, Laboratory of Behavioural Neurology and Imaging of Cognition,University Hospital Geneva, Rue Micheli-du-Crest 24, 1211 Geneva 14, Switzerland.E-mail: [email protected]
12 Patrik Vuilleumier and Others