Embed Size (px)
Citation preview

of June 3, 2018.This information is current as
Responses to Cardiac Allograftsand Increased Allospecific Antibody Absence of Recipient CCR5 Promotes Early
Andrew C. Novick, Hiroshi Toma and Robert L. FairchildHiroyuki Amano, Alice Bickerstaff, Charles G. Orosz,
http://www.jimmunol.org/content/174/10/6499doi: 10.4049/jimmunol.174.10.6499
2005; 174:6499-6508; ;J Immunol
Referenceshttp://www.jimmunol.org/content/174/10/6499.full#ref-list-1
, 19 of which you can access for free at: cites 36 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2005 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 3, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 3, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Absence of Recipient CCR5 Promotes Early and IncreasedAllospecific Antibody Responses to Cardiac Allografts1
Hiroyuki Amano,*§ Alice Bickerstaff,¶ Charles G. Orosz,¶ Andrew C. Novick,* Hiroshi Toma,§
and Robert L. Fairchild2*†‡
Acute rejection is mediated by T cell infiltration of allografts, but mechanisms mediating the delayed rejection of allografts inchemokine receptor-deficient recipients remain unclear. The rejection of vascularized, MHC-mismatched cardiac allografts byCCR5�/� recipients was investigated. Heart grafts from A/J (H-2a) donors were rejected by wild-type C57BL/6 (H-2b) recipientson day 8–10 posttransplant vs day 8–11 by CCR5�/� recipients. When compared with grafts from wild-type recipients, however,significant decreases in CD4� and CD8� T cells and macrophages were observed in rejecting allografts from CCR5-deficientrecipients. These decreases were accompanied by significantly lower numbers of alloreactive T cells developing to IFN-�-, but notIL-4-producing cells in the CCR5�/� recipients, suggesting suboptimal priming of T cells in the knockout recipients. CCR5 wasmore prominently expressed on activated CD4� than CD8� T cells in the spleens of allograft wild-type recipients and on CD4�
T cells infiltrating the cardiac allografts. Rejecting cardiac allografts from wild-type recipients had low level deposition of C3d thatwas restricted to the graft vessels. Rejecting allografts from CCR5�/� recipients had intense C3d deposition in the vessels as wellas on capillaries throughout the graft parenchyma similar to that observed during rejection in donor-sensitized recipients. Titersof donor-reactive Abs in the serum of CCR5�/� recipients were almost 20-fold higher than those induced in wild-type recipients,and the high titers appeared as early as day 6 posttransplant. These results suggest dysregulation of alloreactive Ab responses andAb-mediated cardiac allograft rejection in the absence of recipient CCR5. The Journal of Immunology, 2005, 174: 6499–6508.
A key event during the allograft rejection process is therecruitment of alloantigen-activated T cells into thegraft, where they are activated to express the effector
functions that mediate graft tissue destruction. This recruitment isdirected by both adhesion molecules and chemokines and theirrespective receptors. T cell attractant chemokines includingCXCL9/monokine induced by IFN-� (Mig),3 CXCL10/IFN-�-in-ducible protein-10, and CCL5/RANTES are expressed during re-jection, and several clinical studies have indicated a correlationbetween the expression of these chemokines in graft biopsies withongoing rejection episodes (1–4). The importance of chemokinesin rejection is further indicated by studies in animal models dem-onstrating prolonged allograft survival when donors or recipientswith targeted deletions in genes encoding specific chemokines orreceptors are used or when recipients are treated with Abs to thesemolecules (5–10). However, in complete MHC-mismatched donorto recipient strain combinations, targeting a single chemokine orreceptor in the absence of adjunctive therapy has only resulted indelayed rejection of allografts.
The rejection of allografts when a specific chemokine or che-mokine receptor is antagonized has been interpreted to indicate theredundancy in the chemokine system and the ability of other che-mokine/receptor interactions to replace the target. Previous studiesfrom this laboratory indicated the prolonged survival of cardiacallografts when recipients were treated with Abs to the T cell che-moattractant Mig (9). At the time of rejection, there were manyCCR5-expressing cells in the cardiac allografts, suggesting thatmononuclear cell expression of CCR5 may substitute for Mig-CXCR3 interactions in directing T cells into the grafts. A recentstudy reported the prolonged survival of cardiac allografts inCCR5�/� recipients (11). In clinical renal transplantation, recipi-ents expressing a homozygous mutation encoding a nonfunctionalCCR5 have better graft outcomes than recipients expressing func-tional receptors (12). These studies suggest that expression ofCCR5 may play an important role in directing T cells into allo-grafts to mediate acute rejection. In addition to directing T cellrecruitment, in vitro studies have indicated that CCR5 ligands,including MIP-1�, MIP-1�, and RANTES, amplify T cell activa-tion during culture with anti-CD3 mAb or APCs (13–15). In lightof these in vitro studies, we have investigated the activation ofalloantigen-specific T cells as well as mechanisms of rejection inCCR5-deficient recipients of MHC-mismatched cardiac allografts.The results indicate decreased T cell infiltration into rejecting al-lografts in CCR5�/� recipients that is due to reduced effector Tcell priming to alloantigens, and suggest that alternative mecha-nisms mediate cardiac allograft rejection in CCR5�/� recipients.
Materials and MethodsAnimals
A/J (H-2a), BALB/c (H-2d), P/J (H-2p), and C57BL/6 (H-2b) mice wereobtained through C. Reeder at the National Cancer Institute (Frederick,MD). CCR5�/� mice were obtained from The Jackson Laboratory. Adultmales of 8–12 wk of age were used throughout this study.
*Glickman Urological Institute and †Department of Immunology, Cleveland ClinicFoundation, Cleveland, OH 44195; ‡Department of Pathology, Case Western ReserveUniversity School of Medicine, Cleveland, OH 44106; §Department of Urology, To-kyo Women’s Medical School, Tokyo, Japan; and ¶Department of Surgery, Trans-plantation Division, Ohio State University College of Medicine, Columbus, OH43210
Received for publication March 30, 2004. Accepted for publication March 9, 2005.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by National Institutes of Health Grants AI40459 andAI51620.2 Address correspondence and reprint requests to Dr. Robert L. Fairchild, NB3-79,Lerner Research Institute, Cleveland Clinic Foundation, 9500 Euclid Avenue, Cleve-land, OH 44195-0001. E-mail address: [email protected] Abbreviations used in this paper: Mig, monokine induced by IFN-�; DC, dendriticcell.
The Journal of Immunology
Copyright © 2005 by The American Association of Immunologists, Inc. 0022-1767/05/$02.00
by guest on June 3, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Antibodies
The following Abs were used for immunohistological and flow cytometryanalyses: FITC-conjugated goat anti-mouse IgG Ab (Pierce); FITC-conju-gated rat anti-mouse IgG1, IgG2a, IgG2b, IgG3, IgA, and IgM Abs, PerCP-conjugated GK1.5 (rat anti-mouse CD4 mAb) and 53-6.7 (rat anti-mouseCD8 mAb), FITC-conjugated PgP-1, Ly-24 (anti-mouse CD44 mAb), bi-otinylated and PE-conjugated rat anti-mouse CCR5 mAb, and FITC-con-jugated goat anti-mouse IgM and IgG Ab (BD Pharmingen); rat anti-mousemacrophage mAb, F4/80 (Serotec); goat anti-mouse CXCR3 (C20) andgoat anti-mouse CCR5 antiserum (Santa Cruz Biotechnology); PE-conju-gated donkey anti-goat IgG Ab (Jackson ImmunoResearch Laboratories);and FITC-conjugated rabbit anti-C3d Ab (DakoCytomation).
Preparation of bone marrow-derived dendritic cells (DC)
Bone marrow cells were flushed from the femurs and tibia of A/J mice andcultured for 7 days in medium containing 10 ng/ml GM-CSF and 10 ng/mlIL-4 to generate mature DC. After the culture, the DC were isolated bypositive selection using CD11c� MACS cell separation magnetic beads(Miltenyi Biotec), and 1.6 � 106 DC were injected i.v. into wild-typeC57BL/6 and B6.CCR5�/� mice.
Heterotopic cardiac transplant
Cardiac transplants were performed using the method of Corry et al. (16).Briefly, donor and recipient mice were anesthetized with phenobarbital.Donor hearts were harvested and placed in chilled lactated Ringer’s solu-tion while the recipient mice were prepared. The donor heart was anasta-mosed to the recipient abdominal aorta and vena cava using microsurgicaltechniques. Upon completion of the anastamoses and organ perfusion, thetransplanted hearts resumed spontaneous contraction. The strength andquality of cardiac impulses were graded by palpation each day, as previ-ously described (9). Rejection of cardiac grafts was considered complete bythe cessation of impulse and was confirmed visually for each graft bylaparotomy. In C57BL/6 recipients, complete rejection of A/J cardiacgrafts occurs between 8 and 10 days after transplantation. Cardiac isograftsin C57BL/6 recipients functioned for �100 days. The significance in al-lograft survival between recipient groups was analyzed by log rank test,and p � 0.01 was considered a significant difference between groups.
Immunohistology
Heart grafts were retrieved from recipients at the time of rejection, em-bedded in OCT compound (Sakura Finetek), and frozen at �80°C. Sec-tions were cut at 8 �m and mounted onto slides. For immunohistochem-istry, sections were fixed in acetone for 10 min and air dried. Slides wereimmersed in PBS for 10 min and then in 0.03% H2O2 for 10 min to elim-inate endogenous peroxidase activity. The slides were then stained for 1 hwith 5 �g/ml anti-CD4 mAb (GK1.5), anti-CD8 mAb (53-6.7), or anti-macrophage F4/80 mAb in 0.05 M Tris-HCl with 1% BSA. Control slideswere incubated with rat IgG as the primary Ab. After three washes in PBSfor 5 min each, slides were incubated for 20 min with biotinylated goatanti-rat IgG diluted 1/300 in PBS. After three washes in PBS, slides wereincubated with streptavidin-HRP (DakoCytomation) for 20 min andwashed another three times. To prepare the substrate-chromagen solution,a 10 mg tablet of 3,3�-diaminobenzidine (Sigma-Aldrich) was dissolved in15 ml of PBS plus 12 �l of 30% H2O2. The solution was applied to theslides, which were incubated for 3–7 min and then rinsed in dH2O to stopthe reaction. The slides were counterstained with hematoxylin for 3 minand rinsed with tap water. The slides were dehydrated and viewed underlight microscopy, and the images were captured using ImagePro Plus (Me-dia Cybernetics). Numbers of cells staining positive were counted in eightrandom fields from three different tissue sections from three differentgrafts, and the significance between mean numbers of positive cells perfield in different treatment groups was tested using Mann-Whitney U test.To investigate colocalization of CCR5 with graft-infiltrating CD4 and CD8T cells and macrophages, adjacent sections were cut and placed on separateslides as mirror images (17). One slide was stained with goat anti-mouseCCR5 antiserum and the other with Ab to detect infiltrating CD4 or CD8T cells or macrophages, as above.
For immunofluorescent staining of C3d, 8-�m frozen sections werefixed in acetone for 10 min and air dried. Slides were immersed in PBS for10 min, and FITC-conjugated rabbit anti-C3d Ab diluted 1/100 in 1%BSA/PBS was applied for 30 min in a humid chamber. After three washeswith PBS, the slides were mounted with VECTASHIELD with 4�,6�-dia-
midino-2-phenylindole (Vector Laboratories) and viewed under a fluores-cent microscope.
ELISPOT assay
Priming of alloantigen-specific T cells from heart allograft recipients wasinvestigated by enumerating IFN-�-producing and IL-4 T cells using ELIS-POT assays, as previously described (18, 19). Briefly, ELISA spot plates(Unifilter 350; Polyfiltronics) were coated with 2 �g/ml IFN-�- or IL-4-specific mAb and incubated overnight at 4°C. The plates were blocked with1% BSA/PBS and then washed four times with PBS. Spleen cell suspen-sions from graft recipients were prepared on day 7 posttransplant and usedas responder cells. Spleen cells from A/J, C57BL/6, and DBA mice wereprepared and treated with mitomycin C for use as stimulator cells in theassay, as described above. Responder and stimulator cells (1:2) were cul-tured in serum-free HL-1 medium (BioWhittaker) supplemented with 1mM L-glutamine. After 24 h of cell culture at 37°C in 5% CO2, cells wereremoved from the plate by extensive washing with PBS. Biotinylated anti-IFN-� mAb (2 �g/ml) or anti-IL-4 (4 �g/ml) was added, and the plate wasincubated for 6 h at room temperature. The plate was washed three timeswith PBS/0.05% Tween 20, and streptavidin-conjugated alkaline phospha-tase was added to each well. After 2 h at room temperature, the plates werewashed with PBS, and NBT-5-bromo-4-cloro-3-indolyl substrate (Kirke-gaard & Perry Laboratories) was added for the detection of IFN-�- orIL-4-producing cells. The resulting spots were counted with an Immuno-Spot Series I analyzer (Cellular Technology) that was designed to detectELISA spots with predetermined criteria for spot size, shape, and colori-metric density.
Flow cytometry
The expression of CXCR3 and CCR5 on CD4� and CD8� T cells in thespleens of naive mice and heart allograft recipients and on graft-infiltratingT cells was tested by flow cytometry. Spleen cell suspensions were pre-pared, and 106 cell aliquots were washed three times with staining buffer(Dulbecco’s PBS with 2% FCS/0.2% NaN3) and stained with PerCP ratanti-mouse CD4 or anti-mouse CD8� mAb, FITC anti-mouse CD44 mAb,and goat anti-mouse CXCR3 or biotinylated anti-mouse CCR5 Ab for 25min on ice. The cells were washed twice, and the cells were stained withPE-streptavidin to detect CCR5� cells or PE anti-goat IgG to detectCXCR3� cells. After washing five times, the cells were analyzed by flowcytometry using a FACScan (BD Biosciences). The CD4� or CD8� T cellswere gated and analyzed for expression of CD44 and CXCR3 or CCR5. Toisolate allograft-infiltrating cells, the allograft was retrieved on day 7 post-transplant, cut into small pieces, and incubated at 37°C in 20 ml of amixture containing 1 mM sodium pyruvate, 10 mM HEPES, 1000 U/mlcollagenase, 100 �g/ml DNase, and 1000 U/ml hyaluronidase in HBSS.After 1 h, the tissue was pressed through a cell strainer, the cells werecollected and washed three times with HBSS, and then 106 cell aliquotswere stained with anti-CD4, anti-CD8, and anti-CCR5 Abs.
FIGURE 1. Rejection of MHC-mismatched A/J cardiac allografts bywild-type C57BL/6 vs CCR5�/� recipients. Groups of C57BL/6 wild-type(n � 8) and CCR5�/� (n � 7) mice received cardiac allografts from A/Jdonors, and the grafts were monitored daily by palpation for rejection.Allograft rejection was confirmed visually in all recipients by laparatomy.Isografts were maintained �100 days.
6500 Ab-MEDIATED GRAFT REJECTION IN CCR5-DEFICIENT RECIPIENTS
by guest on June 3, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Flow cytometry to detect and measure donor-specific Abs in cardiacallograft recipient serum was performed using a modification of the methoddescribed by Wasowska et al. (20). Thymocyte suspensions from naiveA/J, P/J, and C57/BL6 mice were prepared in HBSS, and 50-�l aliquotscontaining 1.5 � 105 thymocytes were incubated with 50 �l of 4-folddilutions of sera from naive wild-type C57BL/6 and wild-type and CCR5-deficient heart allograft recipients for 1 h on ice. The cells were washedthree times and suspended in 50 �l of staining buffer (Dulbecco’s PBS with2% FCS/0.2% NaN3) containing a mixture of FITC-conjugated goat Absspecific for mice IgG and IgM (BD Pharmingen) for 30 min on ice. Thecells were washed twice, fixed, and analyzed by flow cytometry. The meanchannel fluorescence of each dilution of each serum sample was deter-mined, and the dilution that returned the mean channel fluorescence to thelevel observed when A/J thymocytes were stained with a 1/4 dilution ofnormal wild-type serum was divided by two and reported as the titer. Se-rum from groups of five cardiac allograft recipients was tested, and dif-ferences in mean titer between wild-type and CCR5�/� recipients weredetermined using Mann-Whitney U test.
To test for the presence of donor-reactive IgM and IgG isotypes in naiveand cardiac allograft recipient serum, 106 A/J thymocytes were incubatedwith a 1/20 dilution of serum. The cells were washed three times andstained with FITC-conjugated Abs to mouse IgM and IgG isotypes for 30min on ice. The cells were washed three times and analyzed by flow cy-tometry. Results shown are the percentage of A/J thymocytes that bounddetectable naive and allograft recipient Ab.
ResultsDelayed rejection, but decreased T cell infiltration into heartallografts in CCR5�/� recipients
To begin to investigate the potential role of CCR5 in cardiac al-lograft rejection, the rejection of complete MHC-mismatched A/Jheart grafts by wild-type C57BL/6 and CCR5�/� recipients wascompared. Wild-type recipients rejected the allografts at day 8–10posttransplant (Fig. 1). A modest prolongation to day 8–11 wasobserved in CCR5-deficient recipients, but this prolongation wasnot statistically significant. Tissue sections prepared from rejectingallografts retrieved from wild-type recipients indicated the intensemononuclear cell infiltration typical of acute rejection (Fig. 2). Incontrast, rejecting allografts retrieved from CCR5�/� recipientshad a clear decrease in mononuclear cell infiltration. When frozensections were stained to assess infiltration by CD4� and CD8� Tcells and macrophages, there was a striking decrease in infiltrationby all three leukocyte populations into the rejecting allografts re-trieved from CCR5-deficient recipients (Fig. 3). When randomsections of slides were counted, a 4-fold decrease in infil-trating CD4� and CD8� T cells ( p � 0.01) was observed in the
FIGURE 2. Histological analysis of rejecting allo-grafts from wild-type C57BL/6 vs CCR5�/� recipients.At the time of rejection, A/J cardiac allografts fromwild-type C57BL/6 (a) and CCR5�/� (b) recipientswere retrieved, and formalin-fixed sections were pre-pared and stained with H&E and viewed under lightmicroscopy. Magnification, �200.
FIGURE 3. Immunohistologicalanalysis of rejecting allografts fromwild-type C57BL/6 and CCR5�/� re-cipients. At the time of rejection, A/Jcardiac allografts were retrieved fromwild-type C57BL/6 (d–f) andCCR5�/� (a–c) recipients, and frozensections were prepared and stainedwith Abs to CD4 (a and d), CD8 (b ande), and F4/80 to detect macrophages (cand f). Magnification, �200.
6501The Journal of Immunology
by guest on June 3, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

CCR5-deficient recipients when compared with infiltration bythese T cells in rejecting grafts from wild-type recipients at thetime of rejection (Fig. 4). The number of infiltrating macrophageswas �2-fold less in the CCR5�/� recipient ( p � 0.05).
Expression of CCR5 by heart allograft-infiltrating cells
A potential mechanism underlying the decreased infiltration ofmononuclear cells into heart allografts in CCR5�/� recipients was
FIGURE 4. Mononuclear cell infiltration into cardiac allografts from wild-type vs CCR5�/� recipients. Rejecting A/J allografts from wild-type andCCR5�/� recipients were retrieved, and frozen sections were prepared and stained with specific Abs to detect CD4� (a) and CD8� T cells (b) andmacrophages (c). The slides were viewed at �200, and the number of positively staining cells was counted in eight random fields of three different frozensections from five different grafts. The number of graft-infiltrating CD4� and CD8� T cells (p � 0.01, Mann-Whitney U test) and macrophages (p � 0.05)was significantly less for each cell population in grafts from CCR5�/� recipients (f) when compared with cell infiltration into wild-type recipients (p).
FIGURE 5. Colocalization of CCR5 with allograft-infiltrating CD4� T cells. On day 8 posttransplant, re-jecting A/J allografts were retrieved from C57BL/6 re-cipients. Mirror-image frozen sections were preparedand stained to colocalize CCR5 expression (b, d, and f)with infiltrating CD4 (a) or CD8 (c) T cells and mac-rophages (e) by immunohistochemistry.
6502 Ab-MEDIATED GRAFT REJECTION IN CCR5-DEFICIENT RECIPIENTS
by guest on June 3, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

the need for CCR5 expression to direct this infiltration. To inves-tigate this, the colocalization of CCR5 on infiltrating CD4� andCD8� T cells and macrophages in rejecting heart allografts re-trieved from wild-type recipients was analyzed. Adjacent, mirror-
image sections were prepared from the heart allografts at the timeof rejection, and one was stained to detect CCR5 expression andthe other stained with Abs to detect the phenotype of the infiltrat-ing cell populations. Infiltrating CD4� T cells clearly expressed
FIGURE 6. Expression of CCR5 on allograft-infil-trating CD4� and CD8� T cells. On day 7 posttrans-plant, A/J allografts were retrieved and digested, and theisolated cells were stained with Abs to detect CCR5 ex-pression on gated graft-infiltrating CD4 (a) and CD8 (b)T cells. The number in each panel indicates the percent-age of graft-infiltrating CD4 or CD8 T cells stainingpositively for CCR5.
FIGURE 7. Expression of CXCR3 and CCR5 on splenic T cells from C57BL/6 recipients of A/J cardiac allografts. C57BL/6 mice received cardiac grafts fromA/J donors, and at the indicated times posttransplant aliquots of spleen cell suspensions from allograft recipients and naive mice were stained with Abs to CD4(a and c) or CD8 (b and d), CXCR3 or CCR5, and CD44. The cells were analyzed by three-color flow cytometry, and the CD4� and CD8� cell populations weregated to analyze expression of CXCR3 or CCR5 vs CD44. Data shown are from a single experiment and are representative of two individual experiments.
6503The Journal of Immunology
by guest on June 3, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

CCR5 (Fig. 5a). In contrast, graft-infiltrating CD8� T cells andmacrophages did not colocalize with CCR5 staining, whereas vas-cular smooth muscle cells known to constitutively express CCR5(21) stained positively (Fig. 5, b and c).
The expression of CCR5 on allograft-infiltrating CD4� andCD8� T cell populations was further investigated using anotherapproach. The allografts were retrieved on day 7 posttransplant,the allograft tissue was digested, and aliquots of the cells werestained with Abs and analyzed by flow cytometry. Although allo-graft infiltration by CD4� T cells was much less intense than in-filtration by CD8� T cells (Figs. 4 and 5), 50% of the infiltratingCD4� T cells expressed CCR5, whereas only 31% of the infiltrat-ing CD8� T cells expressed CCR5 (Fig. 6).
Temporal expression of CXCR3 and CCR5 on T cells fromrecipients of heart allografts
The low expression of CCR5 on allograft-infiltrating CD8� T cellsand macrophages suggested that suboptimal trafficking into theallograft was unlikely to account for the reduced cellular infiltra-tion observed in rejecting heart allografts in CCR5�/� recipients.The expression of chemokine receptors on recipient T cells duringpriming for rejection of heart allografts has remained largely un-defined. To investigate this aspect of the T cell response, the ex-pression of the chemokine receptors CXCR3 and CCR5 was com-pared on CD4� and CD8� T cells in the spleen of allograftrecipients. In addition, the expression of CD44 as an indicator ofcellular activation was used to test chemokine receptor expressionon activated T cell populations in the allograft recipients. Lownumbers of CXCR3-expressing CD4� T cells were observed innaive mice, and this expression was restricted to the CD44high
population (Fig. 7a). In heart allograft recipients, these numbersincreased slightly on day 4 posttransplant and then increased muchmore on days 6 and 8, with the expression again restricted to theCD44high cell population. Although more CXCR3� CD8� thanCD4� T cells were observed in the spleens of naive mice, similarresults were observed with CD8� T cell expression of CXCR3during progression to allograft rejection (Fig. 7b). CD4� T cellexpression of CCR5 followed a similar pattern as CXCR3 expres-
sion, with slight increases at day 4 posttransplant and further in-creases afterward (Fig. 7c). In contrast, expression of CCR5 wasnot as prominent on CD8� T cells in allograft recipients and wasexpressed at lower levels than observed on CD4� T cells (Fig. 7d).At day 4 posttransplant, CCR5 expression was observed only onCD44high cells for each T cell population, but was observed onpopulations of CD44low and CD44high CD4� and CD8� T cells atdays 6 and 8 posttransplant.
Decreased alloreactive T cell priming in CCR5�/� allograftrecipients
The role of CCR5 in alloreactive T cell priming was investigatedby comparing the development of alloreactive T cells from wild-type and CCR5�/� heart allograft recipients with IFN-�- and IL-4-producing cells using ELISPOT assays. The number of IFN-�-producing cells in the CCR5�/� recipient spleen was less than halfthat observed in recipients of wild-type grafts at day 7 posttrans-plant (Fig. 8). The number of alloantigen-specific T cells produc-ing IL-4 was similar in each set of recipients, suggesting that im-mune deviation did not account for the decreased priming ofalloreactive T cells to IFN-�-producing cells observed inCCR5�/� recipients.
The possibilities that the decreased priming of alloreactive Tcells in CCR5-deficient recipients of heart allografts was due to adirect requirement for CCR5 for T cell priming vs an inhibitoryeffect of passenger leukocytes in the allograft on the priming of theCCR5-deficient T cells were addressed. First, purified A/J DCwere administered to wild-type and CCR5�/� recipients, andELISPOT assays were performed 4 days later (Fig. 9a). The num-ber of alloreactive T cells producing IFN-� in CCR5-deficient re-cipients of the DC was 50% that induced in wild-type recipients.Second, heart allograft donor A/J mice were subjected to 1100 radwhole body irradiation to deplete the donors of radiation-sensitivepassenger leukocytes, and 2 days later the hearts were transplantedto wild-type and CCR5�/� recipients. On day 7 posttransplant,graft recipient spleen cells were tested for levels of alloreactive Tcell priming using IFN-� ELISPOT assays (Fig. 9b). Again, the
FIGURE 8. Frequency of donor-specific IFN-�- andIL-4-producing cells in the spleen of wild-type andCCR5�/� recipients of A/J cardiac allografts. On day 7posttransplant, spleen cells from each recipient and fromnaive wild-type mice were analyzed by ELISPOT toenumerate the number of alloantigen-specific T cellsproducing IFN-� (a) and IL-4 (b). Spleens from each setof recipients had similar numbers of T cells as analyzedby flow cytometry and total cell counts. The results arerepresentative of two individual experiments.
FIGURE 9. Frequency of donor-specific IFN-�-pro-ducing cells in response to irradiated allografts and to do-nor DC. a, Wild-type C57BL/6 and CCR5�/� mice re-ceived 1.6 � 106 bone marrow-derived DC i.v., and 4 dayslater spleen cells from each recipient and from naive wild-type mice were analyzed by ELISPOT to enumerate thenumber of alloantigen-specific T cells producing IFN-�. b,A/J mice were treated with 1100 rad whole body irradia-tion, and 2 days later the hearts were transplanted to wild-type C57BL/6 and CCR5�/� mice. On day 7 posttrans-plant, spleen cells from each recipient and from naive wild-type mice were analyzed by ELISPOT to enumerate thenumber of alloantigen-specific T cells producing IFN-�.
6504 Ab-MEDIATED GRAFT REJECTION IN CCR5-DEFICIENT RECIPIENTS
by guest on June 3, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
number of alloreactive T cells producing IFN-� in the CCR5-de-ficient recipients was almost 50% that observed in wild-type re-cipients of the heart allografts from the irradiated donors. Collec-tively, the results in Figs. 8 and 9 indicate the reduced priming ofT cells to alloantigen in the absence of CCR5.
Increased alloantigen-specific Ab production in CCR5�/�
allograft recipients
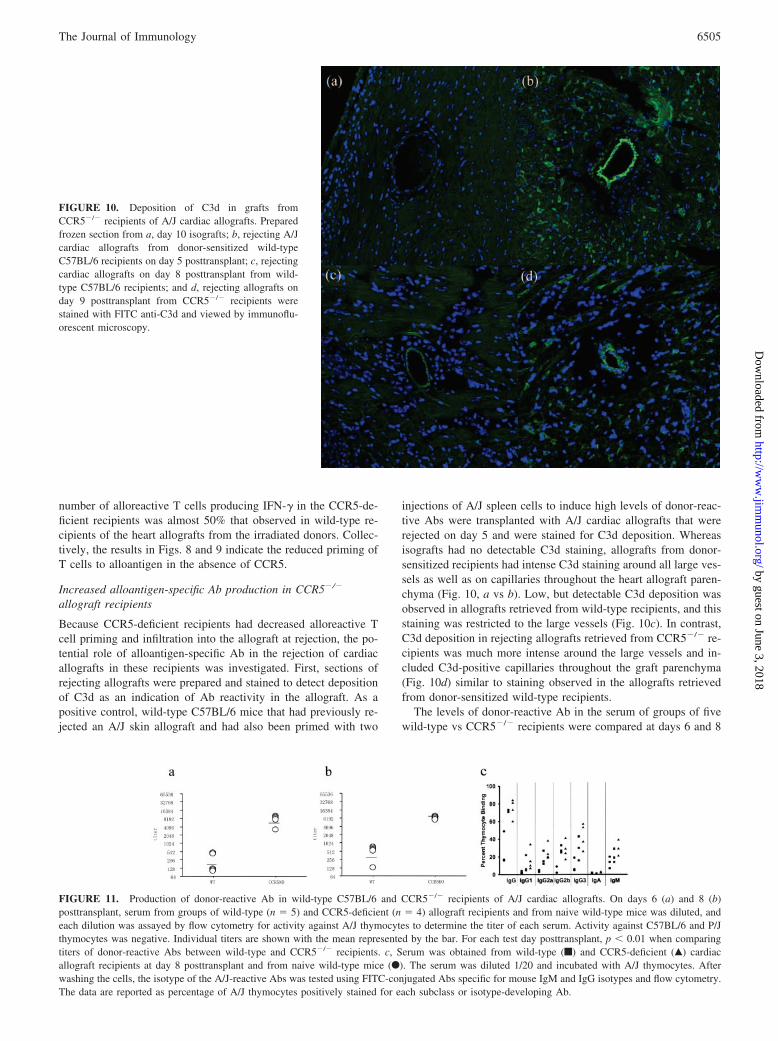
Because CCR5-deficient recipients had decreased alloreactive Tcell priming and infiltration into the allograft at rejection, the po-tential role of alloantigen-specific Ab in the rejection of cardiacallografts in these recipients was investigated. First, sections ofrejecting allografts were prepared and stained to detect depositionof C3d as an indication of Ab reactivity in the allograft. As apositive control, wild-type C57BL/6 mice that had previously re-jected an A/J skin allograft and had also been primed with two
injections of A/J spleen cells to induce high levels of donor-reac-tive Abs were transplanted with A/J cardiac allografts that wererejected on day 5 and were stained for C3d deposition. Whereasisografts had no detectable C3d staining, allografts from donor-sensitized recipients had intense C3d staining around all large ves-sels as well as on capillaries throughout the heart allograft paren-chyma (Fig. 10, a vs b). Low, but detectable C3d deposition wasobserved in allografts retrieved from wild-type recipients, and thisstaining was restricted to the large vessels (Fig. 10c). In contrast,C3d deposition in rejecting allografts retrieved from CCR5�/� re-cipients was much more intense around the large vessels and in-cluded C3d-positive capillaries throughout the graft parenchyma(Fig. 10d) similar to staining observed in the allografts retrievedfrom donor-sensitized wild-type recipients.
The levels of donor-reactive Ab in the serum of groups of fivewild-type vs CCR5�/� recipients were compared at days 6 and 8
FIGURE 10. Deposition of C3d in grafts fromCCR5�/� recipients of A/J cardiac allografts. Preparedfrozen section from a, day 10 isografts; b, rejecting A/Jcardiac allografts from donor-sensitized wild-typeC57BL/6 recipients on day 5 posttransplant; c, rejectingcardiac allografts on day 8 posttransplant from wild-type C57BL/6 recipients; and d, rejecting allografts onday 9 posttransplant from CCR5�/� recipients werestained with FITC anti-C3d and viewed by immunoflu-orescent microscopy.
FIGURE 11. Production of donor-reactive Ab in wild-type C57BL/6 and CCR5�/� recipients of A/J cardiac allografts. On days 6 (a) and 8 (b)posttransplant, serum from groups of wild-type (n � 5) and CCR5-deficient (n � 4) allograft recipients and from naive wild-type mice was diluted, andeach dilution was assayed by flow cytometry for activity against A/J thymocytes to determine the titer of each serum. Activity against C57BL/6 and P/Jthymocytes was negative. Individual titers are shown with the mean represented by the bar. For each test day posttransplant, p � 0.01 when comparingtiters of donor-reactive Abs between wild-type and CCR5�/� recipients. c, Serum was obtained from wild-type (f) and CCR5-deficient (Œ) cardiacallograft recipients at day 8 posttransplant and from naive wild-type mice (F). The serum was diluted 1/20 and incubated with A/J thymocytes. Afterwashing the cells, the isotype of the A/J-reactive Abs was tested using FITC-conjugated Abs specific for mouse IgM and IgG isotypes and flow cytometry.The data are reported as percentage of A/J thymocytes positively stained for each subclass or isotype-developing Ab.
6505The Journal of Immunology
by guest on June 3, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

posttransplant. In wild-type heart allograft recipients, donor-reac-tive Ab titers were at low levels at day 6 (281.6 210.3) post-transplant and were slightly increased by day 8 (435.2 171.7),the day of allograft rejection (Fig. 11, a and b). In contrast, hightiters of donor-reactive Abs were observed in CCR5�/� recipientsas early as day 6 posttransplant (7372.8 1831.7), and at day 8posttransplant (8192 0.0) were 18.8 times the levels observed inwild-type recipients. No reactivity against third-party P/J alloge-neic targets was observed in serum from either allograft recipientgroup (data not shown). Although increases in donor-reactive IgGAb of all isotypes were observed in the serum of CCR5-deficientallograft recipients, no differences in donor-specific IgG isotypeproduction were observed at day 8 posttransplant in serum fromCCR5-deficient allograft recipients when compared with wild-typerecipients (Fig. 11c).
Finally, potential differences in alloantibody responses to im-munization with A/J spleen cells in wild-type vs CCR5�/� micewere tested. Groups of wild-type and CCR5�/� mice were immu-nized with two injections of A/J spleen cells, and 14 days after thesecond injection the titers of A/J-reactive Abs in the serum weretested (Fig. 12). Similar to the induction of donor-reactive Abs inheart allograft recipients, CCR5-deficient mice produced muchhigher titers of donor-reactive Abs than wild-type mice immunizedwith the A/J spleen cells (1536.0 868.8 vs 213.3 73.9).
DiscussionChemokines are important mediators directing leukocyte traffick-ing to tissue sites of inflammation and cellular positioning duringthe development of lymphoid tissue architecture (22, 23). In ad-dition to these functions, chemokines such as IL-8 activate integrinfunction and promote leukocyte adhesion to the vascular endothe-lium (24, 25). In vitro studies have indicated that RANTES aug-ments the activation of T cells by anti-CD3 Ab to produce IL-2 andproliferate (13, 14). The role of chemokines in the activation of Tcells during the initiation of in vivo responses remains poorlyunderstood.
The key characteristic of acute allograft rejection is alloantigen-primed T cell and macrophage infiltration into the allograft. Thisinfiltration is largely mediated by the coordinated functions of ad-hesion molecules and chemokines. A great deal of interest hascentered on the ability of strategies neutralizing the function ofthese sets of molecules to inhibit cellular infiltration into allograftsand promote long-term graft survival. Studies from several labo-ratories have indicated that antagonism of specific chemokines or
their receptors delays or inhibits rejection of skin, cardiac, islet,and small bowel allografts in rodent models (6, 26–28). Much ofthe early work was focused on CXCR3 and its ligands IFN-�-inducible protein-10 and Mig, potent chemoattractants for Ag-primed T cells. Studies from Hancock et al. (7) indicated pro-longed survival of MHC-mismatched cardiac allografts from 7 to8 days in wild-type recipients to beyond 60 days posttransplant inCXCR3�/� recipients. Previous studies from this laboratory re-ported the absence of class II MHC disparate skin allograft rejec-tion in recipients treated with Abs to Mig (26). Treatment withMig-specific Abs also significantly prolongs the survival of MHC-mismatched cardiac allografts to day 18–25 posttransplant (9). Atthe time of cardiac allograft rejection in Mig-Ab-treated recipients,high expression of CCR5 was observed in the grafts, suggesting apotential compensation for Mig antagonism by CCR5-binding che-mokines in directing T cell graft infiltration.
The role of CCR5 and its primary ligands, MIP-1�, MIP-1�,and RANTES, in allograft rejection has generated a considerableamount of recent interest. In human renal transplant patients, ex-pression of a nonfunctional CCR5, CCR532, is associated withdecreases in acute rejection and better graft outcomes (12). Im-proved lung allograft survival in rat recipients is observed by treat-ment with anti-RANTES Abs (5). In a mouse model, allogeneicislets in wild-type recipients are rejected by day 10 posttransplant,whereas 80% of islet allografts are rejected by day 30 in CCR5�/�
recipients and the remainder survive beyond day 100 (27).A recent study indicated prolongation of BALB/c cardiac allo-
graft survival from day 7–8 posttransplant in C57BL/6 wild-typerecipients to day 20 in CCR5�/� recipients (11). Comparison ofallograft histology at day 7 posttransplant indicated significant re-ductions in T cell and macrophage infiltration into BALB/c allo-grafts in CCR5-deficient recipients when compared with rejectingallografts in wild-type recipients. The data in the current studyindicate a more modest prolongation of A/J cardiac allograft sur-vival in CCR5�/� recipients. This difference in survival is notapparently due to the use of A/J mice as the allograft donor, as wehave also observed rejection of BALB/c cardiac allografts at day10/11 posttransplant in CCR5�/� recipients. Although survival ofA/J cardiac allografts is only extended 2–3 days in CCR5-deficientrecipients, there are striking decreases in T cell and macrophageinfiltration at the time of rejection when compared with rejectingallografts in wild-type recipients that raised questions regardingmechanisms underlying this rejection in CCR5�/� recipients.
Rejecting cardiac allografts in CCR5�/� recipients stained in-tensely with Abs to C3d, an indication of potential Ab depositionin the grafts. The histology of the allografts appeared identical withrejecting allografts from donor-sensitized wild-type recipients withheavy deposition of C3d around the large vessels and capillaries ofthe allograft. Consistent with this staining, CCR5�/� allograft re-cipients had high serum levels of donor-specific Ab, with titers�20-fold that observed in wild-type recipients. The presence ofthis Ab response appeared within 6 days after transplantation at atime when CCR5 expression was detectable on activated CD4� Tcells in wild-type recipients of the cardiac allografts. These resultsare strongly suggestive of an Ab-mediated rejection mechanism inthe absence of recipient CCR5.
Several recent studies are supportive of a potential role of CCR5in the regulation of Ab responses. The addition of RANTES andMIP-1� to human B cell cultures has been shown to up-regulateIgG4 and IgE production (29), although the requirement for CCR1or CCR5 for this effect remains unclear. Immunization ofCCR5�/� mice with T-dependent Ags also enhances Ig responses(30), but the consequence of this increase on an ongoing immuneresponse such as during an infection or to an allograft has not been
FIGURE 12. Induction of higher alloreactive Ab responses in CCR5-deficient than wild-type mice to immunization with A/J splenocytes.Groups of four wild-type C57BL/6 and CCR5�/� mice were immunizedwith 4 � 107 A/J spleen cells i.p. on days 0 and �7, and 14 days laterserum from immunized and naive mice was diluted and tested for reactivityto A/J and C57BL/6 thymocytes to determine titers of A/J-reactive Ab.Values of p � 0.05 when titers of wild-type and CCR5�/� mice arecompared.
6506 Ab-MEDIATED GRAFT REJECTION IN CCR5-DEFICIENT RECIPIENTS
by guest on June 3, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

previously addressed. In conjunction with these studies, the currentresults suggest dysregulation of Ab responses in the absence ofCCR5. The mechanism underlying this dysregulation remains un-clear. In contrast to activated CD4� T cells, we could not detectexpression of CCR5 on B cells from C57BL/6 recipients of cardiacallografts, suggesting that CCR5-mediated regulation of Ig re-sponses is mediated through T cells. The absence of CCR5 on Bcells has also been observed in BALB/c mice, whereas B cellsfrom NOD mice experiencing active diabetes expressed CCR5(31). The alloreactive CD4 T cell response in CCR5-deficient re-cipients did not deviate to an IL-4-producing phenotype, suggest-ing that another CCR5-expressing T cell population may regulatethis Ab response to the cardiac allograft. An intriguing possibilityarises from studies indicating that a population of �� T cells re-stricts the magnitude of Ab responses in lpr mice and CCR5 isexpressed by many populations of these T cells (32, 33). In pre-liminary studies, we have observed the presence of CCR5� �� Tcells in the spleens of wild-type recipients of cardiac allografts.Experiments to test the potential role of these T cells in regulatingalloantibody responses to cardiac allografts as well as the absenceof such cells in CCR5-deficient recipients are in progress.
Two potential explanations were considered to account for thedecreased T cell and macrophage infiltration into cardiac allograftsin CCR5�/� recipients. First, there could be defective recruitmentof alloantigen-primed T cells into the allograft that would supporta role for CCR5 in directing this graft infiltration. Based on thedemonstrated role of CCR5-binding chemokines on T cell activa-tion in vitro (13, 14), an alternative mechanism could be an alter-ation in T cell priming to graft alloantigens. Previous studies usingmixed lymphocyte reactivity as an assessment of alloreactive Tcell priming in cardiac allograft recipients indicated no differencebetween wild-type and CCR5�/� recipients (11). However, in al-logeneic islet recipients, a significant decrease in the number ofalloreactive T cells producing IFN-� was observed in CCR5-de-ficient recipients (27). This decrease was apparently not due todefective alloreactive T cell priming, as the number of alloreactiveT cells producing IL-4 was increased in CCR5�/� recipients, sug-gesting a switch to a type 2 cytokine-producing phenotype in theabsence of CCR5. In the current study, CCR5�/� recipients ofheart allografts also had decreases in the numbers of alloantigen-reactive T cells producing IFN-�, but this was not compensated byan increase in the number of IL-4-producing cells. It is also worthnoting studies by several laboratories indicating infiltration andrejection of cardiac allografts by alloreactive T cells producingtype 2 cytokines (34–36). These results suggest that alloreactive Tcell deviation to type 2 cytokine production is unlikely to accountfor the decreased cellular infiltration into the cardiac allograftsobserved in CCR5-deficient recipients.
The results of the current study indicate a striking enhancementof donor-specific Ab responses in CCR5-deficient recipients ofcardiac allografts and Ab deposition in the graft. In light of the lowlevels of mononuclear cell infiltration observed in these allograftsat the time of rejection, the high levels of alloreactive Ab inducedin the absence of CCR5 are likely to be a factor in mediatingrejection of the graft. There is a considerable interest in the po-tential use of chemokine receptor antagonists to inhibit allograftrejection in clinical transplantation. Differences in alloreactive re-sponses and allograft rejection are to be expected in wild-typerecipients treated with chemokine receptor antagonists vs chemo-kine receptor-deficient recipients. Nevertheless, the results of thecurrent study raise caution regarding the use of CCR5 antagonismas a therapeutic strategy in which administration of such an an-tagonist may promote Ab responses and allograft rejection.
AcknowledgmentsWe thank Tarek El-Sawy, Peter Heeger, Anna Valujskikh, Wink Baldwin,and Angus Thomson for helpful suggestions concerning these studies;Danielle Kish for help with generating bone marrow-derived DC; and thestaff of the Cleveland Clinic Foundation Biological Resources Unit forexcellent care of the animals used in this study.
DisclosuresThe authors have no financial conflict of interest.
References1. Melter, M., A. Evxeni, M. E. Reinders, J. C. Fang, G. McMahon, P. Ganz,
W. W. Hancock, and D. M. Briscoe. 2001. Expression of the chemokine receptorCXCR3 and its ligand IP-10 during human cardiac allograft rejection. Circulation104: 2558–2564.
2. Sederer, S., Y. Cui, F. Eitner, T. Goodpaster, K. L. Hudkins, M. Mack,J. P. Cartron, D. Schlondorff, and C. E. Alpers. 2001. Expression of chemokinesand chemokine receptors during human renal transplant rejection.Am. J. Kidney Dis. 37: 518–531.
3. Zhao, D. X. M., Y. Y. Hu, G. G. Miller, A. D. Luster, R. N. Mitchell, andP. Libby. 2002. Differential expression of the IFN-�-inducible CXCR3-bindingchemokines, and IFN-inducible protein 10, monokine induced by IFN, and IFN-inducible T cell � chemoattractant in human cardiac allografts: association withcardiac allograft vasculopathy and acute rejection. J. Immunol. 169: 1556–1560.
4. Fahmy, N. M., M. H. Yamani, R. C. Starling, N. B. Ratliff, J. B. Young,P. M. McCarthy, J. Feng, A. C. Novick, and R. L. Fairchild. 2003. Chemokineand chemokine receptor gene expression indicates acute rejection of human car-diac transplants. Transplantation 75: 72–78.
5. Belperio, J. A., M. A. Burdick, M. P. Keane, Y. Y. Xue, J. P. Lynch,B. L. Daugherty, S. L. Kunkel, and R. M. Strieter. 2000. The role of the CCchemokine, RANTES, in acute lung allograft rejection. J. Immunol. 165: 461–472.
6. Gao, W., P. S. Topham, J. A. King, S. T. Smiley, V. Csizmadia, B. Lu,C. J. Gerard, and W. W. Hancock. 2000. Targeting of the chemokine receptorCCR1 suppresses development of acute and chronic cardiac allograft rejection.J. Clin. Invest. 105: 35–44.
7. Hancock, W. W., B. Lu, W. Goa, V. Csizmadia, K. Faia, J. A. King, S. T. Smiley,M. Ling, N. P. Gerard, and C. Gerard. 2000. Requirement of the chemokinereceptor CXCR3 for acute allograft rejection. J. Exp. Med. 192: 1515–1520.
8. Hancock, W. W., W. Gao, V. Csizmadia, K. L. Faia, N. Shemmeri, andA. D. Luster. 2001. Donor-derived IP-10 initiates development of acute allograftrejection. J. Exp. Med. 193: 975–980.
9. Miura, M., K. Morita, H. Kobayashi, T. A. Hamilton, M. A. Burdick,R. M. Strieter, and R. L. Fairchild. 2001. Monokine induced by IFN-� is a dom-inant factor directing T cells into murine cardiac allografts during acute rejection.J. Immunol. 167: 3494–3504.
10. Morita, K., M. Miura, D. R. Paolone, T. M. Engeman, A. Kapoor, D. G. Remick,and R. L. Fairchild. 2001. Early chemokine cascades in murine cardiac graftsregulate T cell recruitment and progression of acute allograft rejection. J. Immu-nol. 167: 2979–2984.
11. Gao, W., K. L. Faia, V. Csizmadia, S. T. Smiley, D. Soler, J. A. King,T. M. Danoff, and W. W. Hancock. 2001. Beneficial effects of targeting CCR5 inallograft recipients. Transplantation 72: 1199–1205.
12. Fischereder, M., B. Luckow, B. Hocher, R. P. Wuthrich, U. Rothenpieler,H. Schneeberger, U. Panzer, R. A. Stahl, I. A. Hauser, K. Budde, et al. 2001. CCchemokine receptor 5 and renal-transplant survival. Lancet 357: 1758–1761.
13. Bacon, K. B., B. A. Premack, P. Gardner, and T. J. Schall. 1995. Activation ofdual T cell signaling pathways by the chemokine RANTES. Science 269:1727–1730.
14. Taub, D. D., S. M. Turcovski-Corrales, M. L. Key, D. L. Longo, andW. J. Murphy. 1996. Chemokines and T lymphocyte activation. I. � chemokinescostimulate human T lymphocyte activation in vitro. J. Immunol. 156:2095–2103.
15. Karpus, W. J., N. W. Lukacs, K. J. Kennedy, W. S. Smith, S. D. Hurst, andT. A. Barrett. 1997. Differential CC chemokine-induced enhancement of T helpercell cytokine production. J. Immunol. 158: 4129–4136.
16. Corry, R. J., H. J. Winn, and P. S. Russell. 1973. Primarily vascularized allograftsof hearts in mice. Transplantation 16: 343–350.
17. Osamura, R. Y., K. Watanabe, I. Tanaka, Y. Nakai, and H. Imura. 1981. Com-parative immunohistochemical studies of �-melanocyte stimulating hormone (�-MSH) and adrenocorticotrophic hormone (ACTH) in the bovine and human pi-tuitaries. Acta Endocrinol. 96: 458–463.
18. Benichou, G., A. Valujskikh, and P. S. Heeger. 1999. Contributions of direct andindirect alloreactivity during allograft rejection in mice. J. Immunol. 162:352–358.
19. Zhang, Q.-W., D. D. Kish, and R. L. Fairchild. 2003. Absence of allograftICAM-1 attenuates alloantigen-specific T cell priming, but not primed T celltrafficking into the graft, to mediate acute rejection. J. Immunol. 170: 5530–5537.
20. Wasowska, B. A., Z. Qian, D. L. Cangellow, E. Behrens, K. Van Tran, J. Layton,F. Sanfilippo, and W. M. Baldwin. 2001. Passive transfer of alloantibodies re-stores acute cardiac rejection in IgKO mice. Transplantation 71: 727–736.
21. Mueller, A., and P. G. Strange. 2004. The chemokine receptor CCR5. Int. J. Bio-chem. Cell Biol. 36: 35–38.
6507The Journal of Immunology
by guest on June 3, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

22. Rossi, D., and A. Zlotnik. 2000. The biology of chemokines and their receptors.Annu. Rev. Immunol. 18: 217–242.
23. Campbell, J. J., and E. C. Butcher. 2000. Chemokines in tissue-specific andmicroenvironment-specific lymphocyte homing. Curr. Opin. Immunol. 12:336–341.
24. Constantin, G., M. Majeed, C. Giagulli, L. Piccio, J. Y. Kim, E. C. Butcher, andC. Laudanna. 2000. Chemokines trigger immediate �2 integrin affinity and mo-bility changes: differential regulation and roles in lymphocyte arrest under flow.Immunity 13: 759–769.
25. Laudanna, C., J. Y. Kim, G. Constantin, and E. C. Butcher. 2002. Rapid leuko-cyte integrin activation by chemokines. Immunol. Rev. 186: 37–46.
26. Koga, S., M. B. Auerbach, T. M. Engeman, A. C. Novick, H. Toma, andR. L. Fairchild. 1999. T cell infiltration into class II MHC-disparate allografts andacute rejection is dependent on the IFN-� induced chemokine Mig. J. Immunol.163: 4878–4885.
27. Abdi, R., R. N. Smith, L. Makhlouf, N. Najafian, A. D. Luster, H. Auchincloss,and M. H. Sayegh. 2002. The role of CC chemokine receptor 5 (CCR5) in isletallograft rejection. Diabetes 51: 2489–2495.
28. Zhang, Z., L. Kaptanoglu, Y. Tang, D. Ivancic, S. M. Rao, A. D. Luster,T. A. Barrett, and J. Fryer. 2004. IP-10-induced recruitment of CXCR3 host Tcells is required for small bowel allograft rejection. Gastroenterology 126: 809–818.
29. Kimata, H., A. Yoshida, C. Ishioka, M. Fujimoto, I. Lindley, and K. Furusho.1996. RANTES and macrophage inflammatory protein 1� selectively enhanceimmunoglobulin (IgE) and IgG4 production by human B cells. J. Exp. Med. 183:2397–2402.
30. Zhou, Y., T. Kurihara, R.-P. Ryseck, Y. Yang, C. Ryan, J. Loy, G. Warr, andR. Bravo. 1998. Impaired macrophage function and enhanced T cell-dependentimmune response in mice lacking CCR5, the mouse homologue of the majorHIV-1 coreceptor. J. Immunol. 160: 4018–4025.
31. Carvalho-Pinto, C., M. I. Garcia, L. Gomez, A. Ballesteros, A. Zaballos,J. M. Pflores, M. Mellado, J. M. Rodriguez-Frade, D. Balomenos, andC. Martinez-A. 2004. Leukocyte attraction through the CCR5 receptor controlsprogress form insulitis to diabetes in non-obese diabetic mice. Eur. J. Immunol.34: 548–557.
32. Seagal, J., and D. Melamed. 2004. Contribution of �� and �� T cells to thegeneration of primary immunoglobulin G-driven autoimmune response in immu-noglobulin-�-deficient/lpr mice. Immunology 112: 265–273.
33. Brandes, M., K. Willmann, A. B. Lang, K.-H. Nam, C. Jin, M. B. Brenner,C. T. Morita, and B. Moser. 2003. Flexible migration program regulates �� T-cellinvolvement in humoral immunity. Blood 15: 3693–3701.
34. Piccotti, J. R., S. Y. Chan, R. E. Goodman, J. Magram, E. J. Eichwald, andD. K. Bishop. 1996. IL-12 antagonism induces T helper 2 responses, yet exac-erbates cardiac allograft rejection: evidence against a dominant protective role forT helper 2 cytokines in alloimmunity. J. Immunol. 157: 1951–1957.
35. VanBuskirk, A. M., M. E. Wakely, and C. G. Orosz. 1996. Transfusion of po-larized TH2-like cell populations into SCID mouse cardiac allograft recipientsresults in acute allograft rejection. Transplantation 62: 229–238.
36. Matesic, D., A. Valujskikh, E. Pearlman, A. W. Higgins, A. C. Gilliam, andP. S. Heeger. 1998. Type 2 immune deviation has differential effects on alloreac-tive CD4� and CD8� T cells. J. Immunol. 161: 5236–5244.
6508 Ab-MEDIATED GRAFT REJECTION IN CCR5-DEFICIENT RECIPIENTS
by guest on June 3, 2018http://w
ww
.jimm
unol.org/D
ownloaded from









![Journal of Falkenhagen et al, J Antivir Antiretrovir 213 ... · CCR5 gene via Zinc finger nucleases [4], cleavage of CCR5 mRNA by multimeric ribozymes [5], inhibition of CCR5 mRNA](https://img.pdfslide.us/doc/110x75/5fd3f8f670db7b30b42beea9/journal-of-falkenhagen-et-al-j-antivir-antiretrovir-213-ccr5-gene-via-zinc.jpg)



















