Embed Size (px)
Citation preview

Ž .Brain Research 788 1998 232–236
Research report
Abnormal localization of iron regulatory protein in Alzheimer’s disease
Mark A. Smith a,) , Kristina Wehr b, Peggy L.R. Harris a, Sandra L. Siedlak a, James R. Connor c,George Perry a
a Institute of Pathology, Case Western ReserÕe UniÕersity, CleÕeland, OH 44106, USAb ( )European Molecular Biology Laboratory EMBL , Heidelberg 69117, Germany
c Department of Neuroscience and Anatomy, PennsylÕania State UniÕersity College of Medicine, M.S. Hershey Medical Center, Hershey, PA, USA
Accepted 23 December 1997
Abstract
A role for altered iron metabolism in the pathogenesis of Alzheimer’s disease has been suggested by several reports associating thecardinal neuropathologic lesions with markers of free radical-induced damage and redox-active iron. We hypothesized that the abnormal
Ž .distribution of iron in Alzheimer brain might result from alterations in iron regulatory proteins IRP such as IRP-1 and IRP-2, the maincontrol elements of cellular iron homeostasis. Here, we report that while IRP-1 is present at similar levels in both Alzheimer and controlbrain tissue, IRP-2 shows striking differences and is associated with intraneuronal lesions, including neurofibrillary tangles, senile plaqueneurites and neuropil threads. Since IRP-2 colocalizes with redox-active iron, our results suggest that alterations in IRP-2 might bedirectly linked to impaired iron homeostasis in Alzheimer’s disease. q 1998 Elsevier Science B.V.
Keywords: Alzheimer’s disease; Iron; Metabolism; Oxidative stress
1. Introduction
ŽIron regulatory proteins IRP; previously known as.IRE-BP or IRF are cytoplasmic mRNA binding proteins
that respond to cellular iron concentrations by regulatingthe translation of proteins involved in iron uptake, storage,
Ž w x.and utilization reviewed in Ref. 14 . For example, thetranslation of ferritin, transferrin receptor and erythroid5-aminolevulinic acid synthase are regulated by the inter-action of IRP with conserved RNA stem-loop structures
Ž . Žcalled iron-responsive elements IREs reviewed in Refs.w x.37,10 . With iron chelation or low cytoplasmic ironconcentration, IRP, in its apo-form, binds with high affin-ity to the iron-responsive element and is able to stabilizetransferrin receptor mRNA. Conversely, a high concentra-tion of free iron prevents binding to IREs through assem-bly of an iron–sulfur cluster and conversion of IRP to itsinactive holoform. Thus, IRPs regulate iron homeostasisby enabling the tight coordination between the cytoplasmicconcentration of free iron, cellular iron uptake and the
) Corresponding author. Institute of Pathology, Case Western ReserveUniversity, 2085 Adelbert Road, Cleveland, OH 44106, USA. Fax:q1-216-368-8964; E-mail: [email protected]
synthesis of ferritin and heme. Moreover, IRPs may regu-late energy metabolism since the mRNA encoding mito-chondrial aconitase, the rate limiting enzyme of the TCAcycle, also contains IREs.
In Alzheimer’s disease, there is significant evidence forw xdisrupted iron homeostasis 2,3,7,34,38 . In particular, re-
dox-active iron, associated with neurofibrillary tangles andw xsenile plaques 34 , is a likely contributor to the pervasive
Žoxidative damage found in the disease reviewed in Ref.w x.30,31 . Furthermore, aberrant iron homeostasis may par-ticipate in pathophysiologic energy metabolism in
Ž w x.Alzheimer’s disease reviewed in 6 . Given the impor-tance of IRPs in iron homeostasis, oxidative stress, andenergy metabolism, we investigated IRP-1 and IRP-2 ex-pression in Alzheimer’s disease.
2. Materials and methods
2.1. Tissue section preparation
Central nervous system tissue was obtained at post-mortem, from patients with clinically- and histopathologi-cally-confirmed Alzheimer’s disease using the
w x ŽCERADrNIA criteria 16 ns5; ages 74, 79, 81, 81 and
0006-8993r98r$19.00 q 1998 Elsevier Science B.V. All rights reserved.Ž .PII S0006-8993 98 00002-X
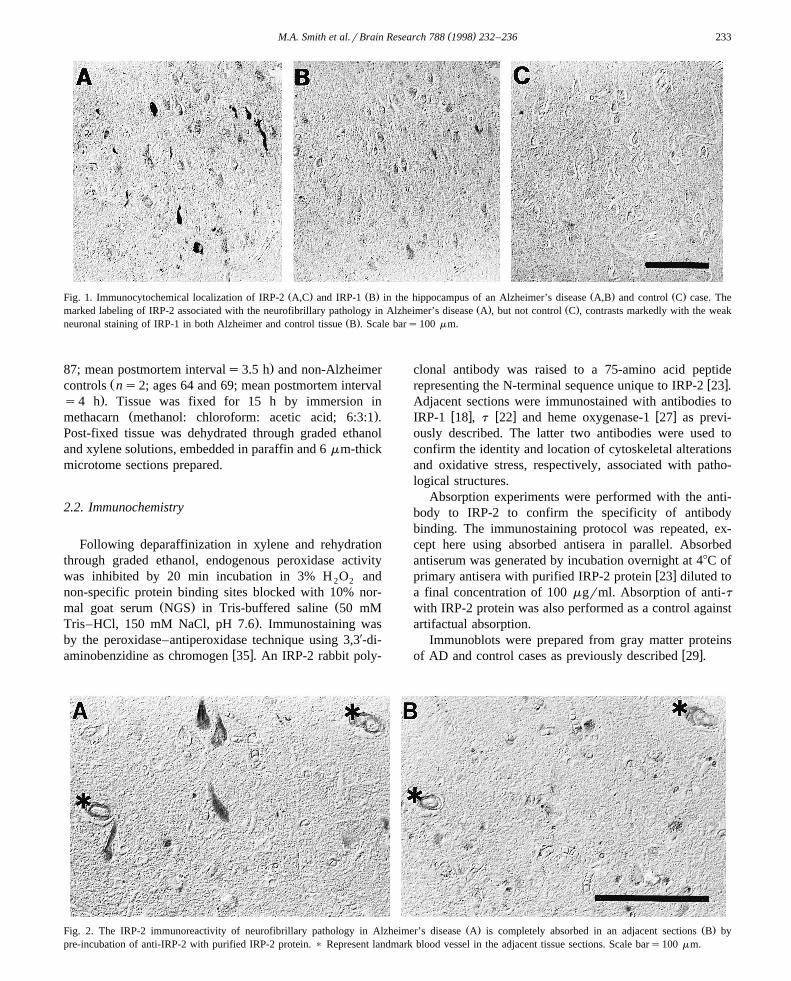
( )M.A. Smith et al.rBrain Research 788 1998 232–236 233
Ž . Ž . Ž . Ž .Fig. 1. Immunocytochemical localization of IRP-2 A,C and IRP-1 B in the hippocampus of an Alzheimer’s disease A,B and control C case. TheŽ . Ž .marked labeling of IRP-2 associated with the neurofibrillary pathology in Alzheimer’s disease A , but not control C , contrasts markedly with the weak
Ž .neuronal staining of IRP-1 in both Alzheimer and control tissue B . Scale bars100 mm.
.87; mean postmortem intervals3.5 h and non-AlzheimerŽcontrols ns2; ages 64 and 69; mean postmortem interval
.s4 h . Tissue was fixed for 15 h by immersion inŽ .methacarn methanol: chloroform: acetic acid; 6:3:1 .
Post-fixed tissue was dehydrated through graded ethanoland xylene solutions, embedded in paraffin and 6 mm-thickmicrotome sections prepared.
2.2. Immunochemistry
Following deparaffinization in xylene and rehydrationthrough graded ethanol, endogenous peroxidase activitywas inhibited by 20 min incubation in 3% H O and2 2
non-specific protein binding sites blocked with 10% nor-Ž . Žmal goat serum NGS in Tris-buffered saline 50 mM
.Tris–HCl, 150 mM NaCl, pH 7.6 . Immunostaining wasby the peroxidase–antiperoxidase technique using 3,3X-di-
w xaminobenzidine as chromogen 35 . An IRP-2 rabbit poly-
clonal antibody was raised to a 75-amino acid peptidew xrepresenting the N-terminal sequence unique to IRP-2 23 .
Adjacent sections were immunostained with antibodies tow x w x w xIRP-1 18 , t 22 and heme oxygenase-1 27 as previ-
ously described. The latter two antibodies were used toconfirm the identity and location of cytoskeletal alterationsand oxidative stress, respectively, associated with patho-logical structures.
Absorption experiments were performed with the anti-body to IRP-2 to confirm the specificity of antibodybinding. The immunostaining protocol was repeated, ex-cept here using absorbed antisera in parallel. Absorbedantiserum was generated by incubation overnight at 48C of
w xprimary antisera with purified IRP-2 protein 23 diluted toa final concentration of 100 mgrml. Absorption of anti-twith IRP-2 protein was also performed as a control againstartifactual absorption.
Immunoblots were prepared from gray matter proteinsw xof AD and control cases as previously described 29 .
Ž . Ž .Fig. 2. The IRP-2 immunoreactivity of neurofibrillary pathology in Alzheimer’s disease A is completely absorbed in an adjacent sections B bypre-incubation of anti-IRP-2 with purified IRP-2 protein. ) Represent landmark blood vessel in the adjacent tissue sections. Scale bars100 mm.

( )M.A. Smith et al.rBrain Research 788 1998 232–236234
3. Results
While antisera to IRP-1 weakly stained neurons in bothŽ .disease and control tissue Fig. 1B , anti-IRP-2 revealed
striking differences between Alzheimer’s disease and con-Ž .trol brain Fig. 1A,C . Indeed, anti-IRP-2 labeling was not
Ž .significant in control brain Fig. 1C , but strong anti-IRP-2immunoreactivity was present in the intracellular neurofib-rillary pathology of Alzheimer’s disease, including neu-
Žrofibrillary tangles and neuropil threads Fig. 1A and Fig..2A . Staining adjacent tissue sections with antibodies to t
or heme oxygenase-1 showed near total overlap of theIRP-2 immunostaining profiles with those recognized by t
or heme oxygenase-1 with IRP-2 only labeling a subsetŽ .approximately 30–50% of the neurofibrillary lesions rec-ognized by either of the other probes. Extracellular neu-rofibrillary tangles were not labeled by IRP-2 antisera. Toconfirm the specificity of IRP-2 labeling, several controlexperiments were performed in parallel. Absorption withpurified IRP-2 protein completely abolished immuno-
Ž .staining of the IRP-2 antisera Fig. 2 , whereas no effectswere observed by absorption of the antiserum to t withIRP-2 protein. No specific staining was found with pre-im-mune rabbit antisera or with rabbit antibodies againstirrelevant epitopes, and as an additional control for thespecificity of the IRP antibodies, immunoblot analysesrevealed proteins corresponding to the molecular weight
w xspecies previously reported for human brain 11 and withno differences between AD and control cases.
4. Discussion
In this study, we immunocytochemically demonstratethe selective association of IRP-2, but not iron IRP-1, withthe pathologic hallmarks of Alzheimer’s disease. Pro-nounced IRP-2 immunoreactivity in Alzheimer’s disease islocalized to neurofibrillary tangles and neuropil threadsŽ .i.e., the neurofibrillary lesions . That only a subset ofneurofibrillary tangles are labeled by IRP-2 suggests thatalterations in IRP-2 occur at specific chronological stagesor, alternatively, that specific epitopes are masked duringthe disease course. In adjacent sections, IRP-2-labelledstructures show overlap with t-positive and HO-1-positivestructures suggesting an association of IRP-2 with cyto-skeletal and oxidative stress abnormalities, respectively.Control brains show little IRP-2 immunoreactivity, con-
w xfirming previous studies of normal brain 11 . While im-munoblot experiments revealed no distinct differences ineither IRP-1 or IRP-2 protein amount in brain ho-mogenates from control and Alzheimer’s disease cases, theincrease of IRP-2 in the cell bodies of susceptible neurons,together with the apparent lack of IRP-1, indicates thatthere is a differential induction or re-localization of IRPproteins in Alzheimer’s disease, perhaps as a consequenceof accumulation of transported protein. In this regard, it is
interesting to note that IRP-2, and not IRP-1, is normallydegraded in the presence of increased iron, in a processrequiring ubiquitination and degradation in proteosomesw x12 . Since IRP-2 lacks the ability to adopt an inactive butstable form, unlike IRP-1, the increased levels of IRP-2 inneurons in Alzheimer’s disease might result from prote-olytic abnormalities previously suggested to play an impor-
w xtant pathogenic role in Alzheimer’s disease 26 . Indeed,decreased ubiquitination or inhibition of proteosome activ-ity would allow for an increase in IRP-2-levels. Accord-ingly, IRP-2-positive neurons show extensive ubiquitinconjugate accumulation, but apparently not degradation, of
w xseveral cytoskeletal proteins 17,21 .As stated earlier, when iron is low, IRP-1 and IRP-2
w x Ž . w xboth bind 13 to an iron responsive element IRE 8 onthe 5X cap structure of the ferritin HmRNA blockingbinding and subsequent translation. When cellular ironincreases, the IRP is released by removal of the 4Fe–4Scluster and translation of ferritin can proceed. Oxidativestress and nitric oxide also influence the interactions be-tween IRPs and IRE. Both of these factors can induce
w xbinding of IRP to IRE 10 , but not identically since IRP-2activity is unaffected by H O while for IRP-1, the re-2 2
sponse to oxidative stress is faster than with either ironw xdepletion or nitric oxide 19 . In the present study, we
report an increase in IRP-2 in the brains of cases ofAlzheimer’s disease and demonstrate that IRP-2 overlaps
Ž .with HO-1 positive oxidatively stressed structures.Therefore, if the increased IRP-2 in Alzheimer’s disease isactively bound to the IRE, it could explain the observationthat iron concentrations are increased in cases ofAlzheimer’s disease brain with a concurrent decrease in
w xferritin 4,15 . Such an increase in iron without an appro-priate increase in ferritin, to detoxify the iron, would leavethe neuron vulnerable to reactive oxygen species. In sup-port of such a scenario, there is a similar distribution of
w xIRP-2 and oxidative changes proteins 20,24,27–29,32,33suggesting a possible common mechanism for both loss ofintracellular iron homeostasis and oxidative stress inAlzheimer’s disease. Additional support of this is the
w xrecent histochemical localization of redox-active iron 34that shows a strikingly similar distribution to the aberrantlocalization of IRP-2. Furthermore, the altered IRP-2 pre-sented here in Alzheimer’s disease complements previouswork showing evidence for perturbations in ironmetabolism including alterations in iron, transferrin and
w xferritin 2,3,7,34,38 .Although IRP-1 and IRP-2 bind IREs with equal affin-
w xity, each also binds unique alternative ligands 1 . There-fore, the increase of IRP-2 in Alzheimer’s disease couldhave downstream effects, other than on iron homeostasis,such as blockage of mitochondrial aconitase synthesis and
w xconsequent metabolic alterations 9 . Furthermore, it isnoted that exon 13 of the amyloid b-protein precursorŽ .b PP , a protein that is linked to Alzheimer’s diseaseŽ w x. w xreviewed in Ref. 25 , contains an IRE-like domain 36 .

( )M.A. Smith et al.rBrain Research 788 1998 232–236 235
Therefore, since oxidative stress increases the expressionw xof b PP and amyloid-b 39 , the findings presented here
suggest a fundamental link between iron metabolism, ox-idative stress, b PPramyloid-b metabolism, proteolyticabnormalities and Alzheimer’s disease.
In conclusion, the association of IRP-2 with the lesionsof Alzheimer’s disease provides further evidence that is-sues of iron management, protease imbalance, oxidativestress and metabolic abnormalities are involved in diseasepathogenesis and, together with previous studies, supportthe therapeutic use of metal chelators in the treatment of
w xAlzheimer’s disease 5 .
Acknowledgements
This work was supported through grants from the Na-Ž .tional Institutes of Health NIH , the American Health
Ž .Assistance Foundation AHAF , and the AmericanŽ .Alzheimer Association IIRG-94-122 . The authors express
thanks to Avi Friedlich for helpful discussions.
References
w x1 J. Butt, H.Y. Kim, J.P. Basilion, S. Cohen, K. Iwai, C.C. Philpott, S.Altschul, R.D. Klausner, T.A. Rouault, Differences in the RNAbinding sites of iron regulatory proteins and potential target diver-
Ž .sity, Proc. Natl. Acad. Sci. U.S.A. 93 1996 4345–4349.w x2 J.R. Connor, S.L. Menzies, S.M. St. Martin, E.J. Mufson, A histo-
chemical study of iron, transferrin, and ferritin in Alzheimer’sŽ .diseased brains, J. Neurosci. Res. 31 1992 75–83.
w x3 J.R. Connor, B.S. Snyder, J.L. Beard, R.E. Fine, E.J. Mufson,Regional distribution of iron and iron-regulatory proteins in the
Ž .brain in aging and Alzheimer’s disease, J. Neurosci. Res. 31 1992327–335.
w x4 J.R. Connor, B.S. Snyder, P. Arosio, D.A. Loeffler, P. LeWitt, Aquantitative analysis of isoferritins in select regions of aged, Parkin-
Ž .sonian and Alzheimer’s diseased brains, J. Neurochem. 65 1995717–724.
w x5 D.R. Crapper McLachlan, A.J. Dalton, T.P. Kruck, M.Y. Bell, W.L.Smith, W. Kalow, D.F. Andrews, Intramuscular desferrioxamine in
Ž .patients with Alzheimer’s disease, Lancet 337 1991 1304–1308.w x6 C.E. Finch, D.M. Cohen, Aging, metabolism, and Alzheimer dis-
Ž .ease: review and hypotheses, Exp. Neurol. 143 1997 82–102.w x7 P.F. Good, D.P. Perl, L.M. Bierer, J. Schmeidler, Selective accumu-
lation of aluminum and iron in the neurofibrillary tangles ofŽ .Alzheimer’s disease: a laser microprobe LAMMA study, Ann.
Ž .Neurol. 31 1992 286–292.w x8 N.K. Gray, M.W. Hentze, Iron regulatory protein prevents binding
of the 43S translation pre-initiation complex to ferritin and eALASŽ .mRNAs, EMBO J. 13 1994 3882–3891.
w x9 B.R. Henderson, E. Menotti, L.C. Kuhn, Iron regulatory proteins 1and 2 bind distinct sets of RNA target sequences, J. Biol. Chem. 271Ž .1996 4900–4908.
w x10 M.W. Hentze, L.C. Kuhn, Molecular control of vertebrate ironmetabolism: mRNA-based regulatory circuits operated by iron, nitric
Ž .oxide, and oxidative stress, Proc. Natl. Acad. Sci. U.S.A. 93 19968175–8182.
w x11 J. Hu, J.R. Connor, Demonstration and characterization of the ironŽ .regulatory protein in human brain, J. Neurochem. 67 1996 838–844.
w x12 K. Iwai, R.D. Klausner, T.A. Rouault, Requirements for iron-regu-lated degradation of the RNA binding protein, iron regulatory pro-
Ž .tein 2, EMBO J. 14 1995 5350–5357.w x13 H.Y. Kim, R.D. Klausner, T.A. Rouault, Translational repressor
activity is equivalent and is quantitatively predicted by in vitro RNAbinding for two iron-responsive element-binding proteins, IRP1 and
Ž .IRP2, J. Biol. Chem. 270 1994 4983–4986.w x14 R.D. Klausner, T.A. Rouault, J.B. Harford, Regulating the fate of
Ž .mRNA: the control of cellular iron metabolism, Cell 72 199319–28.
w x15 D.A. Loeffler, J.R. Connor, P.L. Juneau, B.S. Snyder, L. Kanaley,A.J. DeMaggio, H. Nguyen, C.M. Brickman, P.A. LeWitt, Transfer-rin and iron in normal, Alzheimer’s disease, and Parkinson’s disease
Ž .brain regions, J. Neurochem. 65 1995 710–724.w x16 S.S. Mirra, A. Heyman, D. McKeel, S.M. Sumi, B.J. Crain, L.M.
Brownlee, F.S. Vogel, J.P. Hughes, G. van Belle, L. Berg, TheŽ .consortium to establish a registry for Alzheimer’s disease CERAD :
II. Standardization of the neuropathologic assessment of Alzheimer’sŽ .disease, Neurology 41 1991 479–486.
w x17 H. Mori, J. Kondo, Y. Ihara, Ubiquitin is a component of pairedŽ .helical filaments in Alzheimer’s disease, Science 235 1987 1641–
1644.w x18 K. Pantopoulos, N.K. Gray, M.W. Hentze, Differential regulation of
Ž .two related RNA-binding proteins, iron regulatory protein IRP andŽ .IRPB, RNA 1 1995 155–163.
w x19 K. Pantopoulos, G. Weiss, M.W. Hentze, Nitric oxide and oxidativeŽ .stress H O control mammalian iron metabolism by different2 2
Ž .pathways, Mol. Cell. Biol. 16 1996 3781–3788.w x20 M.A. Pappolla, R.A. Omar, K.S. Kim, N.K. Robakis, Immunohisto-
chemical evidence of oxidative stress in Alzheimer’s disease, Am. J.Ž .Pathol. 140 1992 621–628.
w x21 G. Perry, R. Friedman, G. Shaw, V. Chau, Ubiquitin is detected inneurofibrillary tangles and senile plaque neurites of Alzheimer dis-
Ž .ease brains, Proc. Natl. Acad. Sci. U.S.A. 84 1987 3033–3036.w x22 G. Perry, M. Kawai, M. Tabaton, M. Onorato, P. Mulvihill, P.
Richey, A. Morandi, J.A. Connolly, P. Gambetti, Neuropil threadsof Alzheimer’s disease show a marked alteration of the normal
Ž .cytoskeleton, J. Neurosci. 11 1991 1748–1755.w x23 T.A. Rouault, C.K. Tang, S. Kaptain, W.H. Burgess, D.J. Haile, F.
Samaniego, O.W. McBride, J.B. Harford, R.D. Klausner, Cloning ofthe cDNA encoding an RNA regulatory protein—the human iron-re-sponsive element-binding protein, Proc. Natl. Acad. Sci. U.S.A. 87Ž .1990 7958–7962.
w x24 L.M. Sayre, D.A. Zelasko, P.L.R. Harris, G. Perry, R.G. Salomon,M.A. Smith, 4-Hydroxynonenal-derived advanced lipid peroxidationend products are increased in Alzheimer’s disease, J. Neurochem. 68Ž .1997 2092–2097.
w x25 D.J. Selkoe, Amyloid beta-protein and the genetics of Alzheimer’sŽ .disease, J. Biol. Chem. 271 1996 18295–18298.
w x26 M.A. Smith, G. Perry, Alzheimer disease: an imbalance of prote-Ž .olytic regulation?, Med. Hypotheses 42 1994 277–279.
w x27 M.A. Smith, R.K. Kutty, P.L. Richey, S.-D. Yan, D. Stern, G.J.Chader, B. Wiggert, R.B. Petersen, G. Perry, Heme oxygenase-1 isassociated with the neurofibrillary pathology of Alzheimer’s disease,
Ž .Am. J. Pathol. 145 1994 42–47.w x28 M.A. Smith, S. Taneda, P.L. Richey, S. Miyata, S.-D. Yan, D. Stern,
L.M. Sayre, V.M. Monnier, G. Perry, Advanced Maillard reactionend products are associated with Alzheimer disease pathology, Proc.
Ž .Natl. Acad. Sci. U.S.A. 91 1994 5710–5714.w x29 M.A. Smith, M. Rudnicka-Nawrot, P.L. Richey, D. Praprotnik, P.
Mulvihill, C.A. Miller, L.M. Sayre, G. Perry, Carbonyl-relatedposttranslational modification of neurofilament protein in the neu-rofibrillary pathology of Alzheimer’s disease, J. Neurochem. 64Ž .1995 2660–2666.
w x30 M.A. Smith, L.M. Sayre, V.M. Monnier, G. Perry, Radical AGEingŽ .in Alzheimer’s disease, Trends Neurosci. 18 1995 172–176.

( )M.A. Smith et al.rBrain Research 788 1998 232–236236
w x31 M.A. Smith, L. Sayre, G. Perry, Is Alzheimer’s a disease ofŽ .oxidative stress?, Alzheimer Dis. Rev. 1 1996 63–67.
w x32 M.A. Smith, G. Perry, P.L. Richey, L.M. Sayre, V.E. Anderson,M.F. Beal, N. Kowall, Oxidative damage in Alzheimer’s, Nature
Ž .382 1996 120–121.w x33 M.A. Smith, P.L.R. Harris, L.M. Sayre, J.S. Beckman, G. Perry,
Widespread peroxynitrite-mediated damage in Alzheimer’s disease,Ž .J. Neurosci. 17 1997 2653–2657.
w x34 M.A. Smith, P.L.R. Harris, L.M. Sayre, G. Perry, Iron accumulationin Alzheimer disease is a source of redox-generated free radicals,
Ž .Proc. Natl. Acad. Sci. U.S.A. 94 1997 9866–9868.w x Ž .35 L.A. Sternberger Ed. , Immunocytochemistry, Wiley, New York,
1986.
w x Ž .36 R.E. Tanzi, B.T. Hyman, Alzheimer’s mutation, Nature 350 1991564.
w x Ž .37 E.C. Theil, Iron regulatory elements IREs : a family of mRNAŽ .non-coding sequences, Biochem. J. 304 1994 1–11.
w x38 C.M. Thompson, W.R. Markesbery, W.D. Ehmann, Y.-X. Mao,D.E. Vance, Regional brain trace-element studies in Alzheimer’s
Ž .disease, Neurotoxicology 9 1988 1–7.w x39 S.D. Yan, S.F. Yan, X. Chen, J. Fu, M. Chen, P. Kuppusamy, M.A.
Smith, G. Perry, G.C. Godman, P. Nawroth, J.L. Zweier, D. Stern,Non-enzymatically glycated tau in Alzheimer’s disease induces neu-ronal oxidant stress resulting in cytokine gene expression and release
Ž .of amyloid b-peptide, Nat. Med. 1 1995 693–699.


























