Embed Size (px)
Citation preview

BIOLOGY OF REPRODUCTION 83, 988–996 (2010)Published online before print 11 August 2010.DOI 10.1095/biolreprod.110.085654
Aberrant Growth and Pattern Formation in Peromyscus HybridPlacental Development1
Amanda R. Duselis and Paul B. Vrana2
Department of Biological Chemistry, School of Medicine, University of California Irvine, Irvine, California
ABSTRACT
Crosses between the North American deer mouse speciesPeromyscus maniculatus (BW) and P. polionotus (PO) producedramatic asymmetric developmental effects. BW females matedto PO males (female bw 3 male po) produce viable growth-retarded offspring. In contrast, PO females mated to BW males(female PO 3 male BW) produce overgrown but dysmorphicconceptuses. Most female PO 3 male BW offspring are dead bymidgestation; those surviving to later time points displaynumerous defects reminiscent of several diseases. The hybrideffects are particularly pronounced in the placenta. Here weexamine placental morphological defects via histology and insitu hybridization as well as the relationship between growthand mortality in the female PO 3 male BW cross. These assaysindicate altered hybrid fetal:placental ratios by the equivalent ofmouse (Mus) Embryonic Day (E) 13 and disorganization andlabyrinth defects in female PO3male BW placentas and confirmearlier suggestions of a severely reduced junctional zone in thefemale bw 3 male po hybrids. Further, we show that bothcellular proliferation and death are abnormal in the hybridsthrough BrdU incorporation and TUNEL assays, respectively.Together the data indicate that the origin of the effects is prior tothe equivalent of Mus E10. Finally, as the majority of these assayshad not previously been performed on Peromyscus, these studiesprovide comparative data on wild-type placentation.
apoptosis, developmental biology, placenta, trophoblast
INTRODUCTION
The placenta and associated extraembryonic tissues performmultiple functions during mammalian prenatal development [1,2]. In addition to mediating maternal-fetal nutrient/wastetransfer and immune relationships, these tissues are majorendocrine organs and contribute to early pattern formation andhematopoiesis [3]. Thus, perturbations of placental functionmay result in fetal and/or maternal health consequences [4–6].However, there are few mammalian models for studying thepotential interactions of natural genetic variants to producedevelopmental dysplasias. For example, the genomes ofcommonly used rodent strains do not represent naturallyoccurring allelic combinations and have been affected byinbreeding [7, 8]. Similarly, other domesticated mammals are
known to have been altered by human selection (e.g., dogs,cows) [9].
Deer mice (Peromyscus) offer such an opportunity. Thesenative North American cricetid rodents are .25 million yearsdiverged from both laboratory mice (Mus) and Rats (Rattus)and not capable of interbreeding with them. Crosses betweentwo species, P. maniculatus (stock ¼ BW) and P. polionotus(stock ¼ PO), produce asymmetric effects on growth anddevelopment. Both parental stock strains were derived fromsingle wild populations [10].
While PO and BW animals have identical term fetal andplacental weights, BW females crossed with PO males producegrowth-retarded offspring [11, 12]. These animals remainsmaller than either parental species throughout life (denoted asfemale bw 3 male po to indicate the effect). Litter sizes of thefemale bw 3 male po hybrid cross are slightly smaller thanthose of either parental strain, indicating some prenatal lethality[13]. However, the growth-retarded survivors do not exhibit anoticeably shortened life span (PO, BW live to 4þ yr), arefertile, and the sexes are equally represented.
In contrast, PO females mated to BW males (female PO 3male BW) produce overgrown and severely dysmorphicconceptuses. Roughly half of all female PO 3 male BWbreedings end in complete death of the litter by the timegestation is two-thirds complete [14, 15]. Female PO 3 maleBW hybrids that survive to late gestation display numerousdevelopmental defects, many reminiscent of human syndromes[14]. Rare female PO 3 male BW litters that reach parturitiontypically result in maternal death because of an inability to passthe hybrid offspring through the birth canal [13].
The placenta is particularly affected in both crosses [13,16, 17]. The female bw 3 male po placentas average half theweight of those from the parental strains, while those of thefemale PO 3 male BW cross weigh about three times that ofthe parental strains. An additional portion (;10%) of femalePO 3 male BW conceptuses lack any obvious embryonicstructures [14]. These conceptuses typically resemble pla-cental tissue and thus may be analogous to hydatidiformmoles.
The hybrid placentas (particularly female PO 3 male BW)have been shown to misexpress extracellular matrix (ECM)-related loci, loci-encoding cell-cycle regulators, and locisubject to genomic imprinting [18–20]. Numerous imprintedgene products have been documented as playing significantroles in mammalian placental growth and development [21].Similar misexpression or loss of imprinting (LOI) of imprintedgenes has been linked to the molar phenotype [22, 23].However, neither the relationship of the altered geneexpression to the hybrid phenotypes nor the nature of theplacental defects have yet been elucidated.
To understand the basis of these defects, we thereforeassessed hybrid placental patterning, growth dynamics, andtheir potential association with female PO 3 male BW hybridmortality. We also assessed parental strain and hybrid placentalgrowth, morphology, and gene expression over the time
1Supported by NSF #MCB-0517754 and NIH grant #P40 RR014279.2Correspondence and current address: Paul B. Vrana, PeromyscusGenetic Stock Center, Department of Biological Sciences, University ofSouth Carolina, Columbia, SC 29208. FAX: 803 576 5780;e-mail: [email protected] or [email protected]
Received: 29 April 2010.First decision: 29 May 2010.Accepted: 28 July 2010.� 2010 by the Society for the Study of Reproduction, Inc.eISSN: 1529-7268 http://www.biolreprod.orgISSN: 0006-3363
988
Dow
nloaded from w
ww
.biolreprod.org.

corresponding to Mus Embryonic Day (E) 10–16. Peromyscusdevelopment differs from that of Mus in that it is approximately4 days longer prior to implantation. That is, a Peromyscusembryo E14 is the equivalent of a Mus E10 embryo. However,we refer to Mus equivalent developmental time points to avoidconfusion.
Peromyscus placentas are discoidal and similar in morphol-ogy to those of other muroid rodents [24]. As in other muroids,Peromyscus placentation results in three distinct layers: 1) thelabyrinth, where maternal and fetal blood supplies meet; 2) thejunctional zone (or spongiotrophoblast), which gives rise toinvasive and endocrine cells; and 3) the maternal decidua—thatpart of the endometrium that has been remodeled by invasivezygotic cells and serves as the attachment point. Themorphology of the junctional zone/labyrinth boundary andmaternal interface exhibit some differences from those of housemice (Mus) and other genera. Studies to date indicate that thefemale bw 3 male po placental phenotype is due to a reductionof the junctional zone [16, 17]. The structural changesoccurring in female PO 3 male BW placentation are lessclear, however, as are the dynamics of the abnormalproliferation. Here we investigate these changes throughhistology and gene expression of loci known to exhibit hybridmisexpression and/or trophoblast lineage markers. In addition,we assess BrdU incorporation and TUNEL assays to pursuethese questions.
MATERIALS AND METHODS
Animals
We purchased PO and BW animals from the Peromyscus Genetic StockCenter (http://stkctr.biol.sc.edu). Animals were housed and bred underapproved institutional animal care protocols of the University of California,Irvine, or the University of South Carolina and in accordance with the Guidefor the Care and Use of Laboratory Animals. Animals were housed with foodand water ad libitum on a 16:8-h light:dark cycle. We bred PO females withBW males and BW females with PO males to obtain reciprocal hybrids. Malesand females were placed in the same cage with a separation cage top to allowthe estrous cycle of the female to be induced by the presence of a male withoutcontact. After 3 days, a normal cage top was used so that females and malescould mate at will. Vaginal smears were performed two times a day (morningand evening) and visually assessed for sperm. We designated Day 0 ofdevelopment when sperm was found in vagina of cycling females. Placentas(and associated fetuses) were dissected at each time point listed and for eachgrouping (PO and BW, female bw 3 male po, and female PO 3 male BW).Placentas were then weighed (wet), and the average for each group and timepoint was determined.
Histological Analysis
Placentas were fixed in 4% paraformaldehyde. Fixed samples weredehydrated through ascending concentrations of ethanol and then sent toHistoserv Inc. (www.histoservinc.com) for sectioning and hematoxylin andeosin staining. The laminin (catalog #ab7463) antibody was purchased fromAbcam (www.abcam.com) and used according to the manufacturers’ protocol.
In Situ Hybridization
Detection of messenger RNA by in situ hybridization in tissue sections wasperformed by standard methods [25]. Peromyscus sequences (;500 base-pair
[bp] cDNA fragment from exons 1–2 of the H19 gene, ;400-bp fragmentspanning exons 2–3 of Cdkn1c, and a full-length Esx1 cDNA) were cloned intothe Topo TA dual promoter cloning vector (Invitrogen). Sp6- and T7-derivedRNA probes were generated from linearized plasmids in the presence ofdigoxygenin to generate both sense and anti-sense hybridization probes. Tpbpain situ hybridization was performed with a full-length Mus cDNA probe. TheTpbpa and Esx1 procedures were performed by Histoserv, Inc. For bothhistology and in situ hybridizations, at least three sections from threeindividuals per time point and subgrouping were examined. Sense-strandprobes for each gene were also hybridized; none exhibited staining.
Gene Expression
Quantitative real-time PCR for Esx1 was performed as previouslydescribed. The Peromyscus Rpl32 gene was used as a control. Reactions weredone in triplicate in 96-well plates and run on an ABI 7900 real-time PCRinstrument. Quantitative analysis was conducted according to Critical Factorsfor Successful Real-Time PCR from Qiagen located at www.Qiagen.com. Aminimum of three samples were used for each subgrouping (PO, BW, femalePO 3 male BW and female bw 3 male po) in the analysis.
Proliferation and Apoptosis Assays
Females with litters at the equivalent of Mus E10, E13, and E16 wereinjected with BrdU at 1.5 ml/100 g and euthanized 1 h later. Sections werestained for apoptosis and amount of proliferation with the In situ Cell DeathDetection Kit, AP (catalog #11684809910) and 5-bromo-20-deoxy-uridineLabeling Detection Kit II (catalog #1299964) assay kits from Roche AppliedScience. The number of BrdU-incorporated and TUNEL-positive cells werecounted under 303 magnification. The numbers presented were derived byaveraging at least three sections from four individuals per time group from eachsubgrouping (the two parental strains—denoted as PO/BW, female bw 3 malepo, and female PO 3 male BW).
RESULTS
Placental Growth Versus Hybrid Survival
The placental weights of the parental strains and reciprocalhybrids at three time points (equivalent to Mus ages E10, 13,and 16) are summarized in Table 1. The reduction in female bw3 male po placental weight is apparent at the earliest stageexamined and throughout the remainder of gestation (i.e.,including times later than E16).
Surprisingly, placentas of the female PO 3 male BWconceptuses we examined at E10 do not vary significantly inweight from those of the parental stock animals. However, wedid observe developmental defects in the E10 female PO 3male BW conceptuses. By E13, however, the average female
TABLE 1. Peromyscus placental weights (g) at selected ages.a
Group/ageb E10 E13 �E16
PO/BW 0.075 6 0.013 0.101 6 0.014 0.180 6 0.020bw 3 po 0.046 6 0.007 0.041 6 0.004 0.091 6 0.007PO 3 BW 0.083 6 0.006 0.278 6 0.140 0.458 6 1.00
a Mean weights are shown, 6 1 SD.b All ages are Mus equivalent.
FIG. 1. Average placental:fetal weight ratios from E10 to E16. Genotypeis indicated in key. With the exception of the parental strains (PO/BW) andfemale bw 3 male po at E14, all three classes differed significantly fromeach other after E12 as judged by the Student t-test; (P , 0.001) asimplemented in Microsoft Excel. All ages are Mus equivalent.
HYBRID PLACENTAL GROWTH AND PATTERN FORMATION 989
Dow
nloaded from w
ww
.biolreprod.org.

PO 3 male BW placental weight is greater than that of maturePO/BW embryos (;E15 or later).
Our studies indicate that the majority of female PO 3 maleBW conceptuses are dead by E16. While many of these deathsoccur prior to E10, overall female PO 3 male BW mortalitycontinues to increase from E10 to E16. We had previouslyobserved that greater placental weight was correlated with thedegree of LOI in backcross (PO females 3 F1 males) thatmimics the female PO 3 male BW cross [26]. Similarly, thedegree of LOI in mixed-background animals is associated withprenatal mortality. We therefore considered whether increasedplacental growth might be generally associated with the femalePO 3 male BW mortality.
To test this hypothesis, we plotted placental weights oftimed pregnancies vs. the percentage of female PO 3 male BWconceptuses alive in litters at that age (Supplemental Fig. S1available at www.biolreprod.org). These data provided littleevidence of a specific association at any particular age. Apossible exception is seen from E12 to E13; the percentage oflive conceptuses observed at E12 was only ;50%, while only;30% of those observed at E13 were alive. However,examination of timed pregnancies indicates that the cumulativetotal of live conceptuses by this point is well below 50%(meaning that there may be a selection bias such thatconceptuses that survive to E13 are atypical). This observation,combined with the phenotypic heterogeneity of the female PO3 male BW conceptuses, makes determining such associationsdifficult.
We also assessed fetal:placental weight ratios as a measureof placental efficiency [27]. As expected, the average value of
this ratio increases steadily over the second half of gestationin parental strain (PO or BW) animals (Fig. 1); it reaches 10the day prior to parturition. While the trajectory of the femalebw 3 male po ratio is more erratic, it reaches slightly greatervalues than that of the parental strains. This increased ratioreflects the observation that female bw 3 male po placentalweight is more affected than fetal weight. In contrast, themaximum average female PO 3 male BW ratio is approxi-mately six. The lowered ratio in female PO 3 male BWconceptuses may reflect both placental defects and celllineage shifts toward extraembryonic fates (as suggested bythe molar conceptuses).
Female PO 3 Male BW Hybrid Placental Hemorrhaging
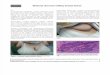
One cause of the female PO 3 male BW placentalinefficiency is suggested by the frequent occurrence ofmacroscopic blood pools (i.e., indicating intraplacental hem-orrhaging). We used hematoxylin and eosin staining to assessthe nature and ontogeny of this major defect in placentalpattern formation (Fig. 2). The female PO 3 male BW bloodpools we have observed are more often located at the placentaledges. However, as shown in Figure 2, large pools occupying acentral sinus occur at least as early as E10 (note that the degreeof hemorrhaging is heterogeneous). This suggests that apossible underlying cause of embryonic death may be failureof maternal-fetal nutrient transfer due to blood vessel instabilityin the labyrinth. Moreover, our previous research suggested adeficiency of some ECM-related proteins in female PO 3 maleBW placentas [20].
FIG. 2. Hematoxylin and eosin-stained histological sections illustrating the hemorrhaging and disorganization observed in the female PO 3 male BWplacentas. Genotype is indicated at top and age (Mus equivalent) at left. Original magnification 31; bars¼ 1 mm.
990 DUSELIS AND VRANA
Dow
nloaded from w
ww
.biolreprod.org.

To further investigate this possibility, we stained placentalsections with a laminin antibody. These proteins are keycomponents of the basal lamina and therefore useful forenhancing the clarity of placental blood vessels [28].Examination of the stained sections suggests a reduction infemale PO 3 male BW embryonic blood vessels (i.e., relativeto PO/BW; Fig. 3). Moreover, the walls of the maternal bloodsinuses within the female PO 3 male BW labyrinth typicallyappear thinner and often compromised. These defects areconsistent with the previously documented reduction in femalePO 3 male BW placental collagen expression as well as similarhemorrhaging in the associated embryos [11, 20].
Labyrinth and Junctional Zone Markers
The product of the X-chromosome-linked Esx1 gene is ahomeodomain transcription factor critical for proper morpho-genesis of vascularization within the labyrinth layer [29, 30].Ablation of mouse Esx1 expression results in enlargedplacentas with little demarcation between the labyrinth andjunctional zone. The Esx1 mutant placentas also exhibitabnormal capillary dilation and branching. In addition to thesesimilarities between the knockout and female PO 3 male BWplacentas, Esx1 is tightly linked to a Peromyscus locus mappedon the basis of association with the placental hypertrophy [26,31]. Thus, variation in this gene may contribute to aspects ofthe female PO 3 male BW dysgenesis.
Our previous study indicated Esx1 overexpression in femalebw 3 male po placentas—possibly reflecting the relatively highproportion of labyrinthine cells in this hybrid cross. Wereassessed Esx1 expression using the same quantitative PCRassay at E10, E13, and E16 (Fig. 4A). The temporallyexpanded data reveals that the female bw 3 male po Esx1overexpression is transitory; at E13, female bw 3 male poexpression drops significantly below that of PO/BW. While theaverage female PO 3 male BW expression is lower than that ofthe parental strains at E10 and E16, the values substantiallyoverlap. Therefore, while it is unlikely that reduced Esx1
expression levels contribute to the female PO 3 male BWdefects, it is possible that spatial misexpression plays a role.
To investigate this possibility, we performed in situhybridization with a Peromyscus Esx1 probe on placentalsections. As expected, the highest proportion of expressingcells is seen at E10, with few positive cells at E16 in theparental strains or either hybrid type. As in Mus, E10expression appeared highest in the developing labyrinth. AtE13 and E16, however, expressing cells are more dispersedthroughout the placenta (Fig. 4B). Esx1 staining in the E10female bw 3 male po hybrid placentas reflects the apparentreduction of junctional zone trophoblast. Similarly, female PO3 male BW E10 Esx1 expression is more dispersed,presumably reflecting that spatial disorganization has alreadyoccurred.
We also performed in situ hybridization experiments withprobes from two additional loci whose primary placentalexpression is within the labyrinthine layer. Cdkn1c and H19 arelinked imprinted loci that we have previously shown to exhibitfemale PO 3 male BW hybrid misexpression [18–20]. Whileboth are normally expressed from the maternal allele, H19exhibits increased (biallelic) female PO 3 male BW expression,while placental Cdkn1c expression is reduced. The potentialeffects of increased H19 expression are unclear [32]. However,the Cdkn1c product is a negative regulator of growth, and itsdownregulation is associated with hyperproliferation in normaldevelopment and diseases, including hydatidiform moles [33].
Figure 5 illustrates that Cdkn1c stains both the Peromyscusjunctional zone and the labyrinth at E10. Thereafter, expressionis confined largely to the labyrinth, trophoblast giant cells(TGCs), and probable immature TGCs. In contrast, H19 isfound in the labyrinth and mature TGCs at all ages. The H19TGC expression illustrates a similar migration of these cells inPeromyscus as in other rodent species: at E10, the majority ofTGCs are seen between the labyrinth and spongiotrophoblastlayer but by E16 are within the decidua and junctional zone.However, the staining patterns of these two genes suggest a
FIG. 3. Representative examples of lami-nin immunohistochemistry to assess laby-rinth vasculature. Genotype is indicated attop and age at left. Columns representgroups of placentas by genotype: PO/BW,female PO 3 male BW and female bw 3male po, respectively. A–C ¼ E10 placen-tas, D–F ¼ E13 placentas and G–I ¼ E16placentas (all ages are Mus equivalent).Double arrowheads represent embryonicblood vessels; single arrowheads indicatecompromised maternal blood sinuses. Suchtears were never observed in PO/BW orfemale bw 3 male po sections. All pictureswere taken of the labyrinth at 340 magni-fication.
HYBRID PLACENTAL GROWTH AND PATTERN FORMATION 991
Dow
nloaded from w
ww
.biolreprod.org.

lack of demarcation between the two zones in female PO 3male BW and are consistent with an early reduction in thefemale bw 3 male po junctional zone/spongiotrophoblast.
We used the well-established marker Tpbpa (i.e., 4311) tofurther assess possible aberrant junctional zone patterning inthe hybrids (Fig. 6). In Mus, this gene is expressed in alljunctional zone cell types except TGCs [34]. Surprisingly, E10Tpbpa staining was most intense in female bw 3 male poplacental sections. However, by E13 the female bw 3 male postaining region was considerably narrower than that of theparental strains and nearly absent at E16. This result suggestsan initial surfeit of female bw 3 male po spongiotrophoblastprogenitors that fail to multiply and/or survive at a normal rate.
In contrast, female PO 3 male BW Tpbpa staining was lesscohesive than that of the parental strains at all three ages: E10expression is scattered and found on the edges of the areasstained by H19 and Cdkn1c. By E13, parental strain Tpbpastaining has expanded considerably, while female PO 3 maleBW is even more scattered. This suggests that very earlyfemale PO 3 male BW placental patterning is at least slightlyperturbed, and the disorganization is magnified as developmentproceeds.
Proliferation and Cell Death
The observed aberrant growth and pattern formation in thehybrid placentas suggests possible imbalances between prolif-eration and apoptosis. As we previously found genes encodingcell-cycle regulators to be affected in the hybrid placentas [20],we investigated whether gross differences in cellular prolifer-ation and death were apparent in the hybrid placentas. Weutilized BrdU incorporation to measure proliferation andTUNEL staining, which identifies cells with DNA fragmenta-tion typically resulting from apoptosis.
In Mus, placental growth shows increasing proliferation andorganization of the placenta until approximately E14.5, when itis considered mature [35]. Reciprocally, regionalized apoptosisin the maternal decidua and fetal glycogen cells start at mid- tolate gestation. Figure 7 indicates that these patterns are verysimilar in wild-type Peromyscus: the number of BrdU-positivecells declines past E10, becoming virtually absent by E16.Correspondingly, the number of PO/BW TUNEL-positiveplacental cells increased over this period.
Both the female PO 3 male BW and the female bw 3 malepo placentas exhibited shifted proliferation/apoptosis curves.Surprisingly, both hybrid types had higher numbers of BrdU-
FIG. 4. Quantitative PCR (A) and in situhybridization assays (B) of placental Esx1expression. In A, genotype is indicated atbottom. Bars indicate standard error. Femalebw 3 male po expression was significantlydifferent (P , 0.001) than the other twoclasses at both E10 and E13 (Mus equiva-lent). In B, genotype is indicated at top andage at left. Note the expanded area of Esx1expression at E10 in both hybrid typesrelative to the parental strains (PO/BW).However, the expressing cells are moredispersed in the female PO 3 male BWhybrids. Arrows indicate areas of relativelyintense staining. Original magnification310; bars ¼ 100 lm.
992 DUSELIS AND VRANA
Dow
nloaded from w
ww
.biolreprod.org.

FIG. 5. Representative results of placental in situ hybridization experiments with Cdkn1c and H19 probes. Columns represent groups of placentas (PO/BW, female PO 3 male BW and female bw 3 male po, respectively). A–H¼ E10 placentas, I–P¼ E13 placentas, Q–X¼ E16 placentas (all ages are Musequivalent). Rows alternate between the two genes as indicated in the leftmost panels. Single arrow¼mature TGCs, double arrow¼ probable immatureTGCs. Brackets in B and D outline spongiotrophoblast. Column 1 pictures were taken at 31 magnification; column 2–4 pictures were taken at 315magnification. Note that the PO/BW 310 magnification picture at E10 (B) appears rotated, as it is a close-up of the rightmost section of A.
HYBRID PLACENTAL GROWTH AND PATTERN FORMATION 993
Dow
nloaded from w
ww
.biolreprod.org.

positive cells, albeit at different points in development: at E10,female bw 3 male po placentas displayed both greatlyincreased numbers of proliferating and dying cells relative tothe parental strains. By E13, however, proliferation had beenreduced to wild-type levels, and cell death was virtually absent.TUNEL-positive cells increased at E16 but remained belowwild-type levels.
In contrast, female PO 3 male BW cell death remained wellbelow that of the parental strains at all three time pointsexamined. While BrdU incorporation did rise to levels well inexcess of those exhibited by the parental strains, this increasewas not apparent until E13. The number of female PO 3 maleBW proliferative cells decreased somewhat from E13 to E16,reaching roughly equivalent numbers as that of apoptotic cellsat the latter time point.
The localization of cell death and proliferation in PO/BWand female PO 3 male BW placentas were similar to thoseobserved in Mus (e.g., late gestation apoptosis in the deciduaand glycogen cells) [36, 37]. The abnormally high levels ofproliferation in the female bw 3 male po placentation werecentered within the spongiotrophoblast layer and maternaldecidua, with apoptotic cell located throughout the placenta(data not shown).
DISCUSSION
The placenta is a transient organ whose proper function isnevertheless critical for both maternal and fetal health [6].Placental hypoplasia is associated with intrauterine and othergrowth retardation [2, 4] syndromes, while hyperplasia isassociated with fetal overgrowth syndromes such as Beckwith-Wiedemann [38] as well as gestational diabetes [39] andvarious forms of gestational trophoblast disease [40]. Peromy-scus offer the opportunity for assessing the effects of naturalgenetic variation on placental growth, development, andphysiology as well as an additional comparative model forthese processes as well as disease. We have previously shownaltered expression and epigenetic state of a number ofimprinted loci implicated in human diseases that correlate withthe Peromyscus hybrid phenotypes (e.g., reductions in Cdkn1cand Phlda2 expression) [41].
We have focused primarily on the dramatic placental defectsin female PO 3 male BW dysgenesis and how they mayunderlie fetal mortality. Female PO 3 male BW placentalefficiency (as defined by fetal:placental weight ratio) is reduced
after E12. This period of placental weight increase isimmediately preceded by a rise in fetal mortality. Among thepotential causes of mortality in this time frame are the observedlabyrinth defects; these include unstable maternal blood spaces(i.e., presumably due to the thinner walls) and a decrease in thenumber of fetal capillaries. These defects would be expected toinhibit maternal-fetal nutrient and waste exchange.
FIG. 6. Representative results of Tpbpa insitu placental hybridization. Genotypes andages indicated at the top and left of figure,respectively (all ages are Mus equivalent).The first column pictures were taken at 31magnification, while all others were takenat 310 magnification. Note that the 310pictures are rotated 90 degrees clockwiserelative to the 31 pictures.
FIG. 7. Quantitation of BrdU (A) and TUNEL (B) assays. Average numberof positive cells per section is shown for each genotype (indicated in key);bars indicate standard error. Significant differences (P , 0.001) in A arefemale bw 3 male po (from the other two classes) at E10 and both hybridtypes (from PO/BW) at E13 and E16. All classes significantly differ fromeach other in B except female bw 3 male po from PO/BW at E13 and thetwo hybrid types from each other at E16 (all ages are Mus equivalent).
994 DUSELIS AND VRANA
Dow
nloaded from w
ww
.biolreprod.org.

The in situ hybridization experiments suggest that bothlabyrinth and junctional zone patterning are affected. Becauseof our previous demonstration of a reduction in Phlda2expression [14], we predicted an increase in the number offemale PO 3 male BW Tpbpa staining cells by comparisonwith the Mus knockout of this gene [42]. However, this was notthe case; we hypothesize that female PO 3 male BWoverproliferation (and simultaneous reduction in apoptosis)results in an elevated proportion of improperly differentiatedcells. For example, we frequently observed cells that may beimmature/improperly differentiated TGCs (Fig. 5). Concurrentexperiments assessing TGC and stem-cell markers support thishypothesis. We suggest that the maternal effect that results infemale PO 3 male BW genomic imprinting perturbations [43]also results in epigenetic misregulation of lineage allocationand/or differentiation.
While the maternal effect locus appears to be the majordeterminant of the female PO 3 male BW phenotypes, Esx1 istightly linked to a locus influencing placental overgrowth inthat cross. Similarities to the Mus targeted mutation strengthenthe case of Esx1 as a candidate for this locus. While there is noapparent reduction in female PO 3 male BW Esx1 expression,its spatial patterning is clearly perturbed. As the analysesclearly indicate, the female PO 3 male BW defects genesisoccurs prior to E10, necessitating earlier analyses.
Preliminary analyses also suggest a role for the Xchromosome in the genetics of the female bw 3 male pogrowth retardation. While it is not clear that this potentiallinkage will include Esx1, its transient overexpression andsubsequent sharp decline are intriguing. While the female bw 3male po E16 expression is wild type, the earlier misexpressionand differences in spatial expression may be sufficient tocontribute to the aberrant placental patterning.
The junctional zone marker Tpbpa also appears overex-pressed in E10 female bw 3 male po placentas as judged by insitu hybridization. However, this region is reduced relative tothe parental strains by E13, corroborating our observation thatthe higher rate of cell death particularly targets this region.Concurrent analyses also indicate a premature reduction inTGC numbers.
It is as yet unclear whether the female bw 3 male po fetaland postnatal growth retardation is dependent on the placentaldefects. These and many other questions make this uniquesystem ripe for additional study. More detailed analyses arenow feasible because of recent increases in Peromyscusresources, including BW genome sequencing in progress(www.ncbi.nlm.nih.gov/Traces/home), more than 90 000 ESTsin public databases, a nascent genetic map [44], and improvedreproductive technology (unpublished data).
ACKNOWLEDGMENTS
The Tpbpa probe was courtesy of Dr. Steven Lipkin of Weill CornellMedical College. We thank Dr. Grant MacGregor (University ofCalifornia, Irvine) and Dr. Steven Kistler (University of South Carolina)for assistance in microscopy/digital photography.
REFERENCES
1. Moffett A, Loke YW, McLaren A (eds.). Biology and Pathology ofTrophoblast. Cambridge, UK: Cambridge University Press; 2006.
2. Fowden AL, Sferruzzi-Perri AN, Coan PM, Constancia M, Burton GJ.Placental efficiency and adaptation: endocrine regulation. J Physiol 2009;587:3459–3472.
3. Barcena A, Kapidzic M, Muench MO, Gormley M, Scott MA, Weier JF,Ferlatte C, Fisher SJ. The human placenta is a hematopoietic organ duringthe embryonic and fetal periods of development. Dev Biol 2009; 327:24–33.
4. Cross JC. Placental function in development and disease. Reprod FertilDev 2006; 18:71–76.
5. Myatt L. Placental adaptive responses and fetal programming. J Physiol2006; 572:25–30.
6. Fisher SJ. The placenta dilemma. Semin Reprod Med 2000; 18:1–6.7. Beck JA, Lloyd S, Hafezparast M, Lennon-Pierce M, Eppig JT, Festing
MF, Fisher EM. Genealogies of mouse inbred strains. Nat Genet 2000; 24:23–25.
8. Smale L, Heideman PD, French JA. Behavioral neuroendocrinology innontraditional species of mammals: things the ‘‘knockout’’ mouse can’ttell us. Horm Behav 2005; 48:474–483.
9. Dobney K, Larson G. Genetics and animal domestication: new windowson an elusive process. J Zool 2006; 269:261–271.
10. Dewey MJ, Dawson WD. Deer mice: ‘‘the Drosophila of North Americanmammalogy.’’ Genesis 2001; 29:105–109.
11. Dawson WD. Fertility and size inheritance in a Peromyscus species cross.Evolution 1965; 19:44–55.
12. Dawson WD. Postnatal development in Peromyscus maniculatus-polio-notus hybrids I, developmental landmarks and litter mortality. Ohio J Sci1966; 66:518–522.
13. Maddock MB, Chang MC. Reproductive failure and maternal-fetalrelationship in a Peromyscus species cross. J Exp Zool 1979; 209:417–426.
14. Duselis AR, Vrana PB. Assessment and disease comparisons of hybriddevelopmental defects. Hum Mol Genet 2007; 16:808–819.
15. Liu TT. Prenatal mortality in Peromyscus with special reference to itsbearing on reduced fertility in some interspecific and intersubspecificcrosses. Contrib Lab Vert Biol 1953; 60:1–32.
16. Rogers JF, Dawson WD. Foetal and placental size in a Peromyscus speciescross. J Reprod Fertil 1970; 21:255–262.
17. Vrana PB, Matteson PG, Schmidt JV, Ingram RS, Joyce A, Prince KL,Dewey MJ, Tilghman SM. Genomic imprinting of a placental lactogen inPeromyscus. Dev Genes Evol 2001; 211:523–532.
18. Vrana PB, Guan XJ, Ingram RS, Tilghman SM. Genomic imprinting isdisrupted in interspecific Peromyscus hybrids. Nat Genet 1998; 20:362–365.
19. Wiley CD, Matundan HH, Duselis AR, Isaacs AT, Vrana PB. Patterns ofhybrid loss of imprinting reveal tissue- and cluster-specific regulation.PLoS ONE 2008; 3:e3572.
20. Duselis AR, Obergfell C, Mack JA, O’Neill MJ, Nguyen QK, O’Neill RJ,Vrana PB. Cell-cycle and extra-cellular matrix gene expression changesduring deer mouse (Peromyscus) hybrid placental development. ReprodFertil Dev 2007; 19:695–708.
21. Maccani MA, Marsit CJ. Epigenetics in the placenta. Am J ReprodImmunol 2009; 62:78–89.
22. Judson H, Hayward BE, Sheridan E, Bonthron DT. A global disorder ofimprinting in the human female germ line. Nature 2002; 416:539–542.
23. El-Maarri O, Seoud M, Coullin P, Herbiniaux U, Oldenburg J, Rouleau G,Slim R. Maternal alleles acquiring paternal methylation patterns inbiparental complete hydatidiform moles. Hum Mol Genet 2003; 12:1405–1413.
24. King BF, Hastings RA. The comparative fine structure of the interhemalmembrane of chorioallantoic placentas from six genera of myomorphrodents. Am J Anat 1977; 149:165–179.
25. Wilkinson D, Nieto MA. Detection of messenger RNA by in situhybridization to tissue sections and whole mounts. Methods Enzymol1993; 225:361–373.
26. Vrana PB, Fossella JA, Matteson P, del Rio T, O’Neill MJ, Tilghman SM.Genetic and epigenetic incompatibilities underlie hybrid dysgenesis inPeromyscus. Nat Genet 2000; 25:120–124.
27. van Rens BT, de Koning G, Bergsma R, van der Lende T. Preweaningpiglet mortality in relation to placental efficiency. J Anim Sci 2005; 83:144–151.
28. Natale D, Starovic M, Cross JC. Phenotypic analysis of the mouseplacenta. Methods Mol Med 2006; 121:275–293.
29. Li Y, Lemaire P, Behringer RR. Esx1, a novel X chromosome-linkedhomeobox gene expressed in mouse extraembryonic tissues and malegerm cells. Dev Biol 1997; 188:85–95.
30. Li Y, Behringer RR. Esx1 is an X-chromosome-imprinted regulator ofplacental development and fetal growth. Nat Genet 1998; 20:309–311.
31. Loschiavo M, Nguyen QK, Duselis AR, Vrana PB. Mapping andidentification of candidate loci responsible for peromyscus hybridovergrowth. Mamm Genome 2007; 18:75–85.
32. Thorvaldsen JL, Duran KL, Bartolomei MS. Deletion of the H19differentially methylated domain results in loss of imprinted expressionof H19 and Igf2. Genes Dev 1998; 12:3693–3702.
33. Fisher RA, Hodges MD, Rees HC, Sebire NJ, Seckl MJ, Newlands ES,
HYBRID PLACENTAL GROWTH AND PATTERN FORMATION 995
Dow
nloaded from w
ww
.biolreprod.org.

Genest DR, Castrillon DH. The maternally transcribed gene p57KIP2(CDNK1C) is abnormally expressed in both androgenetic and biparental
complete hydatidiform moles. Hum Mol Genet 2002; 11:3267–3272.
34. Lescisin KR, Varmuza S, Rossant J. Isolation and characterization of a
novel trophoblast-specific cDNA in the mouse. Genes Dev 1988; 2:1639–
1646.
35. Watson E, Cross JC. Development of structures and transport functions in
the mouse placenta. Physiology (Bethesda) 2005; 20:180–193.
36. Jurisicova A, Detmar J, Caniggia I. Molecular mechanisms of trophoblast
survival: from implantation to birth. Birth Defects Res C Embryo Today
2005; 75:262–280.
37. Huppertz B, Herrler A. Regulation of proliferation and apoptosis during
development of the preimplantation embryo and the placenta. Birth
Defects Res C Embryo Today 2005; 75:249–261.
38. McCowan LME, Becroft DMO. Beckwith-Wiedemann syndrome, pla-
cental abnormalities, and gestational proteinuric hypertension. Obstet
Gynecol 1994; 83:813–817.
39. Yu Y, Singh U, Shi W, Konno T, Soares MJ, Geyer R, Fundele R.
Influence of murine maternal diabetes on placental morphology, geneexpression, and function. Arch Physiol Biochem 2008; 114:99–110.
40. Cheung AN. Pathology of gestational trophoblastic diseases. Best PractRes Clin Obstet Gynaecol 2003; 17:849–868.
41. Apostolidou S, Abu-Amero S, O’Donoghue K, Frost J, Olafsdottir O,Chavele KM, Whittaker JC, Loughna P, Stanier P, Moore GE. Elevatedplacental expression of the imprinted PHLDA2 gene is associated withlow birth weight. J Mol Med 2006; 85:379–387.
42. Frank D, Fortino W, Clark L, Musalo R, Wang W, Saxena A, Li CM, ReikW, Ludwig T, Tycko B. Placental overgrowth in mice lacking theimprinted gene Ipl. Proc Natl Acad Sci U S A 2002; 99:7490–7495.
43. Duselis AR, Wiley CD, O’Neill MJ, Vrana PB. Genetic evidence for amaternal effect locus controlling genomic imprinting and growth. Genesis2005; 43:155–165.
44. Ramsdell CM, Lewandowski AA, Weston Glenn JL, Vrana PB, O’NeillRJ, Dewey MJ. Comparative genome mapping of the deer mouse(Peromyscus maniculatus) reveals greater similarity to rat (Rattusnorvegicus) than to the lab mouse (Mus musculus). BMC Evol Biol2008; 8:65–78.
996 DUSELIS AND VRANA
Dow
nloaded from w
ww
.biolreprod.org.