Embed Size (px)
Citation preview

A2 WJEC BIOLOGY
UNIT 4
Sexual reproduction
in plants
Biology Department - Gower College Swansea
The generalised structure of flowers to be able to compare wind and insect pollinated flowers
Learners should be able to recognise and label a halfflower of a typical regular, diocotyledonous, insect
pollinated flower to include: receptacle, calyx, sepal, corolla, petal, stamen, filament, anther, carpel, ovary,
ovule, style and stigma.
They should be able to identify differences between an insect and a wind-pollinated flower in terms of function
of flower parts and adaptations to different methods of pollination.
The development of pollen and ovules, including examination of prepared slides of anther and ovary
The role of mitosis and meiosis in the development of pollen grains in an anther to include:
• mitosis to produce large numbers of pollen mother cells followed by meiosis to produce a tetrad of four hap-
loid cells;
• the role of the tapetum in pollen grain development;
• development and structure of a mature pollen grain, including subsequent mitotic divisions of the nucleus;
dehiscence and pollen dispersal.
The role of mitosis and meiosis in the development of an ovule in the ovary to include:
• meiosis of a megaspore mother cell in the nucellus to produce four haploid megaspores;
• the growth and subsequent development of one of the megaspores including three mitotic divisions to pro-
duce eight haploid nuclei within the embryo sac.
Learners should be able to recognise structures in a mature ovule to include: funicle, integuments, micropyle,
embryo sac, female gamete, two synergids, two polar nuclei, three antipodal cells.

Cross and self-pollination
Pollination is the transfer of pollen from an anther to a stigma. Learners should understand the
genetic
consequences of self-pollination and cross-pollination and appreciate how meiosis and random
fertilisation result in increased genetic variation through cross-pollination. There are different
adaptations of flowers that promote cross-pollination. These include irregular flower structure and
chemical self-incompatibility.
Learners should understand the events following pollination to include:
• mitosis of the pollen grain nucleus to produce two male gametes and a pollen tube nucleus
• germination of a pollen grain on a compatible stigma;
• growth of a pollen tube (under the control of the pollen tube nucleus) through the digestion of the
style through the secretion of hydrolase enzymes
• entry of the pollen tube into the embryo sac through the micropyle.
Both male gametes are involved in separate fertilisation events:
• one male gamete enters the embryo sac and fuses with the female gamete to produce a diploid
zygote
the second male gamete fuses with the two polar nuclei to form a triploid primary endosperm
nucleus.
The formation and structure of seed and fruit as shown by broad bean and maize
The events that take place following double fertilisation to produce seeds and fruits include:
• the ovule developing into a seed;
• the diploid zygote divides by mitosis to form the diploid embryo, consisting of plumule, radicle and
one or two cotyledons;
• the triploid endosperm nucleus divides by mitosis to form endosperm tissue, an important food
storage tissue in cereal grains, e.g. wheat;
• the integuments develop into the testa;
• the micropyle remains as a pore in the testa
• the ovary wall develops into a fruit wall enclosing the seeds.
Learners should be able to identify and label diagrams of broad bean and maize seeds to include:
hilum (scar of the funicle), micropyle, testa, position of radicle, plumule, cotyledons.
Seeds have evolved as a survival strategy for a terrestrial mode of life. Plants have developed
different mechanisms to enable the dispersal of seeds. This reduces competition following germina-
tion and increases the chance of growth into mature plants.

The process of germination of Vicia faba (broad bean)
Seeds can remain dormant until suitable conditions are present, i.e. availability of water, oxygen and
a suitable temperature. Germination involves the rapid onset of biochemical activity and growth of a
seedling until the plant can carry out photosynthesis and become independent of the food stores
contained in the cotyledons.
The stages of germination in a non-endospermic seed, e.g. broad bean include:
• water being imbibed through the micropyle; the cotyledons swelling and the testa being split to al-
low
entry of more oxygen for aerobic respiration;
• food reserves from the cotyledons, starch and proteins, are mobilised through hydrolysis (and also
lipids in some seeds);
• providing sources of energy for respiration and growth of the plumule and radical. In endospermic
seeds, e.g. maize, gibberellin, a plant hormone, is involved in the process of germination:
• following imbibition of water gibberellin is released by the embryo and diffuses to the aleurone layer
which contains proteins;
• gibberellins induces the production of hydrolytic enzymes, e.g. amylase which break down stored
nutrients
• glucose and other nutrients diffuse to the embryo where they are used in aerobic respiration and

The structure of insect and wind pollinated plants
Carpel
Diagram of the key features of an insect pollinated angiosperm flower
Summary points on the structure of the flower
The anther is the male reproductive organ and is where the male gamete is formed.
The carpel is the female reproductive organ.
The petals are brightly coloured to attract insects for pollination. The flower may also release a
scent to attract pollinators.
They have receptacles containing nectar as a reward for insects.
The ovary is where the embryo will develop after fertilisation in a structure called the embryo sac
which is found in the ovule.
Angiosperm plants and insects are a good example of co-evolution in regards to pollination. This means
that both the plant and the insects have adaptations to encourage and facilitated the transfer of pollen.
Adaptations include:
Brightly colored petals to attract insects.
Sweat nectar for the insect to feed on.
'landing platform' and nectar guides that show the insect where to land and where to search for nectar

Diagram of the key features of a wind pollinated grass flower
Insect pollinated plants Wind pollinated plants
A comparison of the structures of wind and insect pollinated plants.
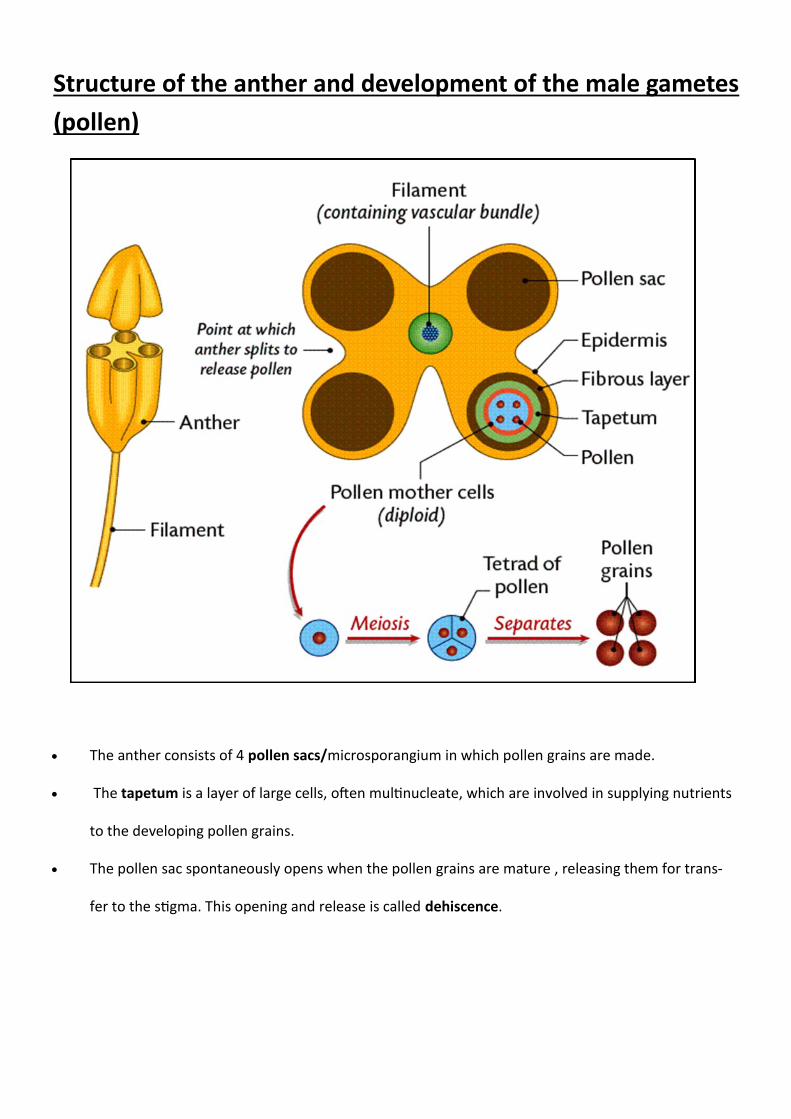
Structure of the anther and development of the male gametes
(pollen)
The anther consists of 4 pollen sacs/microsporangium in which pollen grains are made.
The tapetum is a layer of large cells, often multinucleate, which are involved in supplying nutrients
to the developing pollen grains.
The pollen sac spontaneously opens when the pollen grains are mature , releasing them for trans-
fer to the stigma. This opening and release is called dehiscence.

The pollen grain contains the male gamete – this is called the generative nucleus. This divides later
by mitosis to form 2 sperm nuclei.
the pollen grain also contains the tube nucleus. The tube nucleus controls the development of the
pollen tube
The pollen grain has a highly textured surface called the exine.
The pollen grain protects the generative nucleus from desiccation.
The textured surface of the pollen grain aids in it’s attachment to the stigma.
The pollen grain is the vehicle to transport the male gamete (generative nucleus).
Scanning electron micrograph of the surface
of pollen grains.

The structure of the carpel (the female reproductive organ)
and the female gamete
Structure of the female carpel.
Detailed structure of the ovary.
The ovule is made up of the
integuments, nucellus and the
megasporocyte
The megasporocyte divides by meiosis
and mitosis (see next page) to produce
the embryo sac
The funicle/funiculus connects the
ovule to the ovary wall. The area at the
point of connect ion is called the
placenta

Within the ovule, the megaspore mother cell divides by meiosis and then by mitosis three times to
form 8 haploid nuclei. Initially these are present in the embryo sac without separation but are then
walled off to produce separate cells.
one of these will be the female gamete (the egg cell or ovum)
two other nuclei form the polar nuclei which remain together in the embryo sac without being
walled off until after fertilisation
Two cells called the synergid cells form either side of the egg cell and are involved in releasing
chemical signals to guide the pollen tube to the egg cell. They disintegrate after fertilisation.
The antipodal cells form at the other end of the embryo sac but have no function and
disintegrate.

Pollination
Before fertilisation can occur in plants there has to be the transfer of pollen to the stigma – this is called
pollination. Pollination occurs by means of insects transferring pollen from the anther to the stigma.
Pollination can also occur via the wind – this is called wind pollination.
There are two types of pollination
1. Self pollination.
This occurs when pollen is transferred from the anther to the stigma of the same plant. This pollination
reduces variability.
2. Cross pollination.
This occurs when pollen is transferred from the anther to the stigma of a different plant. This pollina-
tion can promote the generation of variation in the offspring.
The relative merits of self- and cross-pollination We have seen that insect and wind pollination are both ways of achieving cross-pollination and that self-
pollination is possible in some flowers. There are advantages associated with both self- and cross-
pollination. In fact, many plant species show both types.
Self-pollination has the advantage that it is very reliable, particularly if the plants are widely scattered. It
is also advantageous in harsh environments, such as high on mountains where insects and other
pollinators are scarce. The major disadvantage of self-pollination is that it results in less genetic variation.
Self fertilisation occurs, with gametes from the same parent fusing. This is an extreme form of
inbreeding (inbreeding is sexual reproduction between genetically similar individuals). Inbreeding makes
it more likely that both parents will possess the same harmful recessive alleles, making it more likely that
the alleles will come together and be expressed in the offspring. Also, the reduced genetic variation re-
stricts the opportunities for natural selection to occur, and therefore for adaptation to changes in the
environment. Evolution of the species is therefore restricted.
Cross-pollination is a form of outbreeding, that is sexual reproduction between genetically different indi-
viduals - the more genetically different, the, greater the outbreeding. The advantage of cross- pollination
is that it results in more genetic variation than self-pollination. This improves the chances of the species
surviving environmental change and adapting well to its environment because it provides more variants
for natural selection.
Cross-pollination results in greater genetic variation for the simple reason that the gametes are produced
by genetically different individuals, and therefore show more genetic variation.

Mechanisms favoring cross-pollination
We have seen that cross-pollination brings genetic advantages and that elaborate mechanisms exist to
increase the likelihood and efficiency of crosspollination. The main ones are listed here but other, often
unique, mechanisms can be found.
Dioecious plants. When a species, such as willow, produces separate male and female plants it is
described as dioecious. Self pollination is impossible in such species, but the number of dioecious
plant species is very few. They are often trees: holly, yew and poplar are other examples.
Monoecious plants. Monoecious species, such as oak and birch, are those which produce separate
male and female flowers on the same plant. This encourages cross-pollination between adjacent
plants, while still allowing self-pollination among the flowers of the same plant.
Protandry and protogyny. Anthers and stigmas sometimes mature at different times, thus
encouraging cross-pollination. If the anthers mature first, as in the white deadnettle, it is known as
protandry. The term used when the stigmas mature first, as in the bluebell, is protogyny. Usually
there is an overlapping period when both anthers and stigmas are ripe, allowing for self-
pollination as well.
Self-incompatibility. Even if self-pollination occurs, self fertilisation is often made less likely or
impossible by slow or zero growth of the pollen tubes. This is termed incompatibility and is
genetically determined. An extreme example is clover, which is totally self-incompatible.
Structural Incompatibility. The diagram below shows this mechanism in the primrose (Primula vulgaris) which has 2 flower types (a) thrum and (b) pin. These 2 types were originally described by Darwin. He suggested that when an insect visits a pin flower it inserts its proboscis (feeding tube) into the flower tube to feed on the nectar, and pollen from the anthers would stick onto the proboscis about halfway down. If the insect then visits a thrum flower the pollen already on its proboscis is at the right level to meet and stick to the stigma about halfway down the flower tube, leading to cross fertilisation. At the same time pollen from the anthers at the top of the thrum flower tube is likely to stick to the top of the insect proboscis, the perfect position for cross-fertilising the next pin flower the insect visits.
(a) (b)
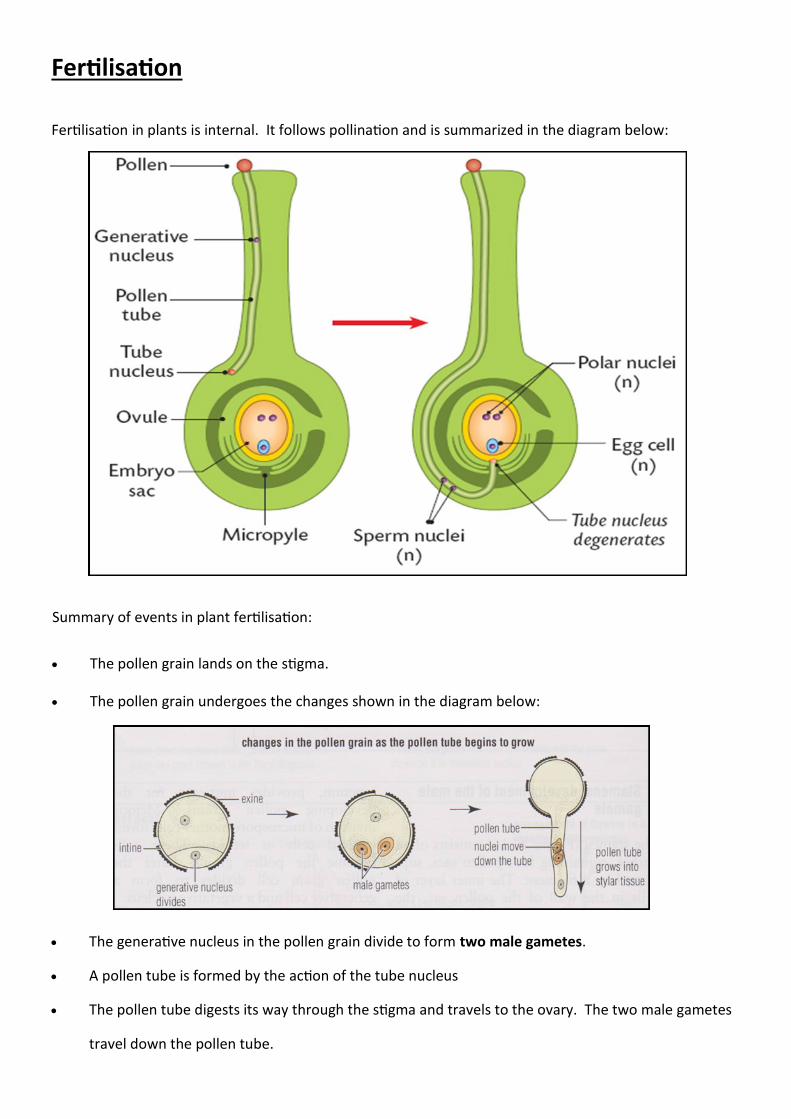
Fertilisation
Fertilisation in plants is internal. It follows pollination and is summarized in the diagram below:
Summary of events in plant fertilisation:
The pollen grain lands on the stigma.
The pollen grain undergoes the changes shown in the diagram below:
The generative nucleus in the pollen grain divide to form two male gametes.
A pollen tube is formed by the action of the tube nucleus
The pollen tube digests its way through the stigma and travels to the ovary. The two male gametes
travel down the pollen tube.

The pollen tube enters the embryo sac via the micropyle.
The tube nucleus degenerates.
One of the male gametes fertilizes the egg cell (ovum) which lies directly above the micropyle. This
forms a zygote. The second male gamete fuses with the polar nuclei to form a triploid nucleus
(which has three sets of chromosomes compared to the normal two). This is called double
fertilisation and only occurs in angiosperms.
The fertilised egg cell will develop into the embryo
The fertilised polar nuclei will develop into the endosperm which is a food supply for the growing
embryo.
the embryo sac will form into the seed and the ovary will form into the fruit.
The integuments will form the testa which is the hard outer coat of the seed.
Diagram showing the changes in the embryo sac after fertilisation.

Development of the embryo and the seed
Diagram above shows the further development of the embryo sac in a dicotyledon plant. Note the for-
mation of the zygote which is made up of cotyledons, a plumule and radical. Note in the bottom dia-
gram the testa which is the coat of the seed. The testa is a protective layer.
The cotyledons are a food source.
The plumule will from the shoot of the new plant.
The radical will from the roots of the new plant.
Diagram of the Broad Bean (dicotyledon) seed –
external features. Diagram of the Broad Bean seed – internal features.

Germination: the necessary conditions
The essential environmental factors for germination are an adequate supply of water, a suitable
temperature and an appropriate partial pressure of oxygen. The essential internal conditions required for
germination are the maturity of the embryo and the overcoming of dormancy.
1) Water uptake
Germination can commence after the uptake of water by the seed. Water is absorbed through the
micropyle and the testa. Water absorption is imbibition, the process of uptake of water by the dry colloi-
dal substances of the seed, which include lipids and the dry cell wall substances of the dehydrated seed
tissue. The resultant swelling of the seed may rupture the testa. Subsequent movement of water in the
hydrated seed tissue is by osmosis.
Water is essential to the vacuolation of the growing cells and for the activation of the enzymes that
catalyse the biochemical reactions of germination. Turgor of plant cells is a force for cell expansion when
the walls are in a condition permitting stretching. Water is a reagent in the hydrolysis of stored food
substances, and is required for the translocation of hydrolysed food reserves - the sugars, amino acids
and fatty acids - to the sites of growth in the embryo.
2) Temperature
The optimum temperature for germination is the optimum for the enzymes involved in mobilisation of
food reserves, provided that other factors are nonlimiting. This temperature varies from species to
species; wheat seeds germinate in the temperature range 1-35°C, and maize in the range 5-45°C.
3) Oxygen
Respiration makes available the energy for metabolism and growth. Germinating seeds respire very
rapidly, and require oxygen for aerobic respiration. Seeds will not germinate in the total absence of
oxygen. Diffusion of oxygen through the testa may be slow, however, and in the early stages of
germination seeds may rely on some anaerobic respiration, at least until the testa has ruptured.
4)Mobilisation of stored food
The stored foods of seeds consist of carbohydrates, lipids and proteins. Starch forms the major food
reserve of most seeds, but in the sunflower and some other seeds oil makes up about half of the food
stored. In pea and bean seeds protein is an additional important reserve food.
Food is stored in seeds in an insoluble form, and must be hydrolysed to soluble substances early in
germination, following the hydration of the seed. Stored food is hydrolysed to produce the substrates for
respiration (sugars) and the building blocks for synthesis (substances such as sugars, amino acids and fatty
acids). Some hydrolytic enzymes already exist in the seed and await only water uptake and an
appropriate temperature to become activated. Other enzymes are produced in response to hormones
released from the developing embryo. Release of hydrolytic enzymes is triggered by gibberellic acid (GA/
gibberellin), which is produced in the embryo and diffuses to the food store. GA works directly on the nu-
clei of the food storage cells, activating the genes that code for hydrolytic enzymes. Since GA is formed
by the embryo only after sufficient water has been absorbed, mobilisation of food reserves is linked to
germination.






























