Embed Size (px)
Citation preview

R
A
ICKa
Kb
c
h
•
•
•
•
•
a
A
R
R
A
A
K
B
A
C
M
S
B
P
E
1
n
h
0
Behavioural Brain Research 313 (2016) 1–7
Contents lists available at ScienceDirect
Behavioural Brain Research
journa l homepage: www.e lsev ier .com/ locate /bbr
esearch report
tryptic hydrolysate from bovine milk as1-casein enhancespentobarbital-induced sleep in mice via the GABAA receptor
rene Joy I. dela Pena a, Hee Jin Kim a, June Bryan de la Pena a, Mikyung Kim a,hrislean Jun Botanas a, Kyung Yi You a, Taeseon Woo a, Yong Soo Leeb, Jae-Chul Jung c,yung-Mi Kim c, Jae Hoon Cheong a,∗
Uimyung Research Institute for Neuroscience, Department of Pharmacy, Sahmyook University, 815 Hwarang-ro, Nowon-gu, Seoul 139-742, Republic of
orea
Department of Pharmacy, Duksung Women’s University, 33 Samyang-ro 144-gil, Dobong-gu, Seoul, 01369, Republic of Korea
Life Science Research Institute, NOVAREX Co., Ltd., Ochang, Cheongwon, Chungbuk 28126, Republic of Korea
i g h l i g h t s
Bovine aS1-casein tryptic hydrolysate did not alter the locomotor activity and motor function of mice.
Bovine aS1-casein tryptic hydrolysate potentiated the sleep induced by pentobarbital sodium in mice.
Bovine aS1-casein tryptic hydrolysate increased the slow (delta) EEG wave in rats.
Bovine aS1-casein tryptic hydrolysate increased Cl− influx, in vitro, which was blocked by bicuculline.
Bovine aS1-casein tryptic hydrolysate has sleep-promoting properties with no or minimal sedative effects.
r t i c l e i n f o
rticle history:
eceived 6 May 2016
eceived in revised form 6 July 2016
ccepted 8 July 2016
vailable online 9 July 2016
eywords:
ovine aS1-casein tryptic hydrolysate (CH)
lpha-casozepine
asein
ilk
leep
enzodiazepine
entobarbital
a b s t r a c t
Studies have shown that enzymatic hydrolysis of casein, the primary protein component of cow’s
milk, produces peptides with various biological activities, and some of these peptides may have sleep-
promoting effects. In the present study, we evaluated the sedative and sleep-promoting effects of bovine
aS1-casein tryptic hydrolysate (CH), containing a decapeptide aS1-casein known as alpha-casozepine.
CH was orally administered to ICR mice at various concentrations (75, 150, 300, or 500 mg/kg). An hour
after administration, assessment of its sedative (open-field and rota-rod tests) and sleep-potentiating
effects (pentobarbital-induced sleeping test and EEG monitoring) were conducted. Although a trend can
be observed, CH treatment did not significantly alter the spontaneous locomotor activity and motor
function of mice in the open-field and rota-rod tests. On the other hand, CH (150 mg/kg, respectively)
enhanced the sleep induced by pentobarbital sodium in mice. It also promoted slow-wave (delta) EEG
activity in rats; a pattern indicative of sleep or relaxation. These behavioral results indicate that CH
has sleep-promoting effects, but no or has minimal sedative effects. To elucidate the probable mecha-
EGnism behind the effects of CH, we examined its action on intracellular chloride ion influx in cultured
human neuroblastoma cells. CH dose-dependently increased chloride ion influx, which was blocked by
co-administration of bicuculline, a competitive GABAA receptor antagonist. Taken together, the results of
the present study suggest that CH has sleep-promoting properties which are probably mediated through
ride i
the GABAA receptor–chlo. Introduction
Sleep plays a vital role in our health and well-being. A “good
ight’s sleep” can soothe and restore a person’s physical and psy-
∗ Corresponding author.
E-mail address: [email protected] (J.H. Cheong).
o
a
s
ttp://dx.doi.org/10.1016/j.bbr.2016.07.013
166-4328/© 2016 Elsevier B.V. All rights reserved.
on channel complex.
© 2016 Elsevier B.V. All rights reserved.
chological state after a long and tiring day. Adequate and quality
sleep can also protect an individual from some diseases and acci-
dents, and can generally promote a better quality of life [1,2].
However, millions of people in the world do not get enough amount
of sleep or have difficulty falling or staying asleep (i.e. sleep dis-
rders). These conditions can lead to impairments in well-being
nd the development of various health problems such as obe-
ity, diabetes, and hypertension [1,3–5]. To manage sleep disorders

ral Br
a
d
b
s
s
S
b
b
(
b
p
s
p
m
T
o
t
a
m
a
d
e
(
t
2
2
w
S
c
r
w
m
a
A
t
t
r
U
2
L
m
I
d
0
L
c
p
i
w
i
g
2
5
d
6
m
a
a
t
N
d
2
U
fi
t
3
d
a
t
f
2
s
d
i
e
i
(
r
t
w
2
w
r
w
fi
I
A
r
p
2 I.J.I. dela Pena et al. / Behaviou
nd avoid or mitigate sleep-related problems, sedative-hypnotics
rugs (e.g. benzodiazepines, diazepam) are commonly prescribed
y physicians. However, long-term use of these synthetic sub-
tances may lead to tolerance, dependence, and other unwanted
ide-effects [6,7]. Thus, there is a growing interest in the develop-
ment of new sleep-aids that don’t have or have lesser side effects
than sedative-hypnotic drugs, and most preferably from natural
sources.
Milk has long been known and used as natural sleep-aid.
Recently, we reported that milk collected during night time (Night
milk) produces sedative and sleep-promoting effects in mice, an
effect comparable to that of the benzodiazepine drug, diazepam
[8]. This effect of Night milk is attributed to its rich stores of
sleep-promoting components such as tryptophan and melatonin.
Another milk component that is thought to contribute to its sleep-
promoting properties is casein, the primary protein constituent
of cow’s milk. Studies have shown that enzymatic hydrolysis of
casein produces peptides with various biological activities [9,10].
Of these, bovine aS1-casein tryptic hydrolysate (CH), containing a
decapeptide aS1-casein known as alpha-casozepine, was shown to
have anxiolytic, anticonvulsant, and anti-stress effects [1,11–14].
tudies have also shown that CH can alleviate sleep disturbances in
oth animals and humans [1,12]. These effects of CH are thought to
e mediated through its affinity to the gamma amino butyric acid
GABA)A receptors. This receptor is also the main binding site of
enzodiazepine-type drugs. Despite the putative sleep-promoting
roperties of CH, no study has directly assessed the sedative and
leep-promoting effects of CH.
Thus, in the present study we evaluated the sedative and sleep-
romoting effects of CH in mice. CH was orally administered to
ice in dosages of 75, 150, 300, or 500 mg/kg of body weight.
hen, behavioral changes consequential to CH treatment (acute
r repeated) were observed in various experimental procedures
hat would characterize the sedative (open-field and rota-rod test)
nd sleep-promoting (pentobarbital-induced sleeping test and EEG
onitoring) activities of a substance. The most commonly used
nd prescribed benzodiazepine, diazepam, was used as a reference
rug in these experiments. Moreover, to further characterize the
ffects of CH and its possible mechanism, changes in chloride ion
Cl−) influx (GABAergic neurotransmission) was assessed in cul-
ured human neuroblastoma cells (SH- SY5Y), in vitro.
. Materials and methods
.1. Animals
Male ICR mice (25–30 g) and Sprague-Dawley rats (250–300 g)
ere obtained from Hanlim Laboratory Animals Co. (Hwasung,
outh Korea). They were housed in groups in a temperature-
ontrolled (22 ± 2 ◦C) and humidity-controlled (55 ± 5%) animal
oom on a 12/12 h light/dark (07:00–19:00 h light) schedule. They
ere acclimatized to the facility for seven days before the com-
encement of any experiments. Food and water were freely
vailable, except the night before and during the experiments.
ll experiments were performed between 10:00 to 16:00. Animal
reatment and maintenance were carried out in accordance with
he Principles of Laboratory Animal Care (NIH publication No. 85-23
evised 1985) and the Animal Care and Use Guidelines of Sahmyook
niversity, Korea (SYUIACUC 2015-009).
.2. Drugs and materials
Bovine aS1-casein tryptic hydrolysate (CH, also known as
actium®) was obtained from Ingredia (Arras, France). N-(6-
ethoxyquinolyl) acetoethylester (MQAE) was purchased from
t
d
d
r
ain Research 313 (2016) 1–7
nvitrogen Co. (Carlsbad, CA, USA). Bicuculline was dissolved in
imethyl sulfoxide (DMSO), with a maximum concentration of
.1%. Pentobarbital sodium was obtained from Hanlim Pharm. Co.,
td (Seoul, South Korea). Diazepam and other materials were pur-
hased from Sigma-Aldrich Co. (St. Louis, Mo, USA). CH was freshly
repared in sterile distilled water and was orally (p.o.) given to mice
n dosages 75, 150, 300, or 500 mg/kg (CH-treated group). Diazepam
as diluted in physiological saline and was intraperitoneally (i.p.)
njected at 1 mg/kg (positive control group). Animals in the control
roup received the vehicle (distilled water, p.o.).
.3. Psychopharmacological evaluation
All experiments were conducted 60 min after the 1st (acute) and
th (repeated) day of treatment. Experiments were performed as
escribed in our previous studies [8,15], but with minor modifica-
tions.
2.3.1. Open-field test (OFT)
The open-field apparatus was a square arena measuring
42 × 42 × 42 cm. CH, diazepam, or the vehicle was given to mice
0 min before the start of each session. To start an experiment,
ice were gently placed in the center of the open-field and were
llowed to spontaneously explore the whole arena. Mice were
llowed to habituate to the open-field arena for 2 min to eliminate
he bias of novelty. Then, an automated system (Ethovision, Noldus,
etherlands) recorded the distance moved (cm) and movement
uration (seconds) of each subject for 10 min.
.3.2. Rota-rod test
The rotating rod or rota-rod apparatus used was purchased from
go Basile Corporation (Model 7650; Varese, Italy). It was set at a
xed speed of 36 rotations per minute (rpm). A day before the actual
est, mice were habituated and trained to run on the rotating rod for
min. Mice with poor performance were excluded. On the actual
ay of the experiment, mice were given CH, diazepam, or vehicle
nd placed on the rotating rod. Each session lasts for 10 min. Latency
ime before the first fall (seconds) and the number of falls (falling
requency) were recorded.
.3.3. Pentobarbital-induced sleeping test
Experiments were performed in a standard mice cage, mea-
uring 50 × 30 × 50 cm, with aspen beddings. Mice were given CH,
iazepam, or vehicle before the start of the test. To start a test and to
nduce sleep, pentobarbital sodium (42 mg/kg, i.p.) was injected to
ach mouse. Immediately after pentobarbital injection, mice were
ndividually placed in cages for observation. The disappearance
onset of sleep) and reappearance (duration of sleep) of the righting
eflex were monitored and recorded, for a maximum of 2 h. Animals
hat did not sleep within 15 min after pentobarbital administration
ere excluded from the experiment.
.3.4. Electrophysiological (EEG) recording
To assess the changes in brain electrical wave activities, rats
ere surgically implanted with wireless telemetry and were
ecorded by LabChart 7.3 software (AD Instruments, USA). Animals
ere anesthetized with pentobarbital sodium (50 mg/kg, i.p.) and
xed in a stereotaxic apparatus (51600 Stoelting Co., Wood Dale,
llinois). A two-channel digital transmitter (Telemetry Research,
uckland, NZ) was intraperitoneally (abdomen) implanted to each
at then electrodes were shunted towards the skull. Electrodes were
ositioned into the left frontal and right occipital cortical regions
hen fixed with a screw and stabilized with dental cement. Seven-
ay post-surgery (recovery period), rats were given vehicle, CH, or
iazepam, and then 60 min after, electroencephalographic (EEG)
ecording were started. Two frequency bands, delta (0.5–4 Hz; slow

ral Br
w
F
2
s
C
4
c
o
s
a
p
r
m
s
c
a
2
u
n
h
A
s
s
3
3
t
g
m
a
t
d
F
I
s
3
w
O
p
r
4
s
(
d
a
e
t
o
b
t
I.J.I. dela Pena et al. / Behaviou
ave) and alpha (8–13 Hz; fast wave), were quantified using Fast
ourier-transformed (FFT).
.3.5. Intracellular Cl− measurement assay
The effects of the CH, or CH + bicuculline, on intracellular Cl−
concentration ([Cl−]i) in SH-SY5Y human neuroblastoma cells
were evaluated. Relative changes were measured using the Cl−-
ensitive indicator, N-(6-methoxyquinolyl) acetoetylester (MQAE).
ells were washed twice and re-suspended at a concentration of
× 105 cells/ml in Hank’s solution. For loading MQAE into the cells,
ells were incubated overnight with the dye at a final concentration
f 5 mM at room temperature. Fluorescence (excitation wavelength
et at 365 nm, emission wavelength at 450 nm) was monitored in
well-stirred cuvette. Experiments were executed at room tem-
erature to minimize fluorescent dye loss. Data are presented as
elative fluorescence F/F0, where F0 is the fluorescence without Cl−
ions and F is the fluorescence as a function of time. The F/F0values
are directly proportional to [Cl−]i. Fluorescence values were modi-
fied for background fluorescence which was separately determined
using an HEPES-buffered KSCN solution containing five mM valino-
ycin to maximally quench the MQAE ion-selective signal. In a
eparate experiment the F0 value was determined by bathing the
ells with Cl−-free (KNO3) solution containing 10 mM tributyltin
nd 10 mM nigericin.
.4. Statistical analysis
All data were expressed as mean ± S.E.M. Data were analyzed
sing one-way analysis of variance (ANOVA). When statistically sig-
ificant differences were found, Dunnett’s test was used as a post
oc test to determine the statistical differences between groups.
ll statistical analyses were conducted using GraphPad Prism Ver-
ion 5.02 software (California, USA). Differences were considered
tatistically significant when p < 0.05.
. Results
.1. Open-field test
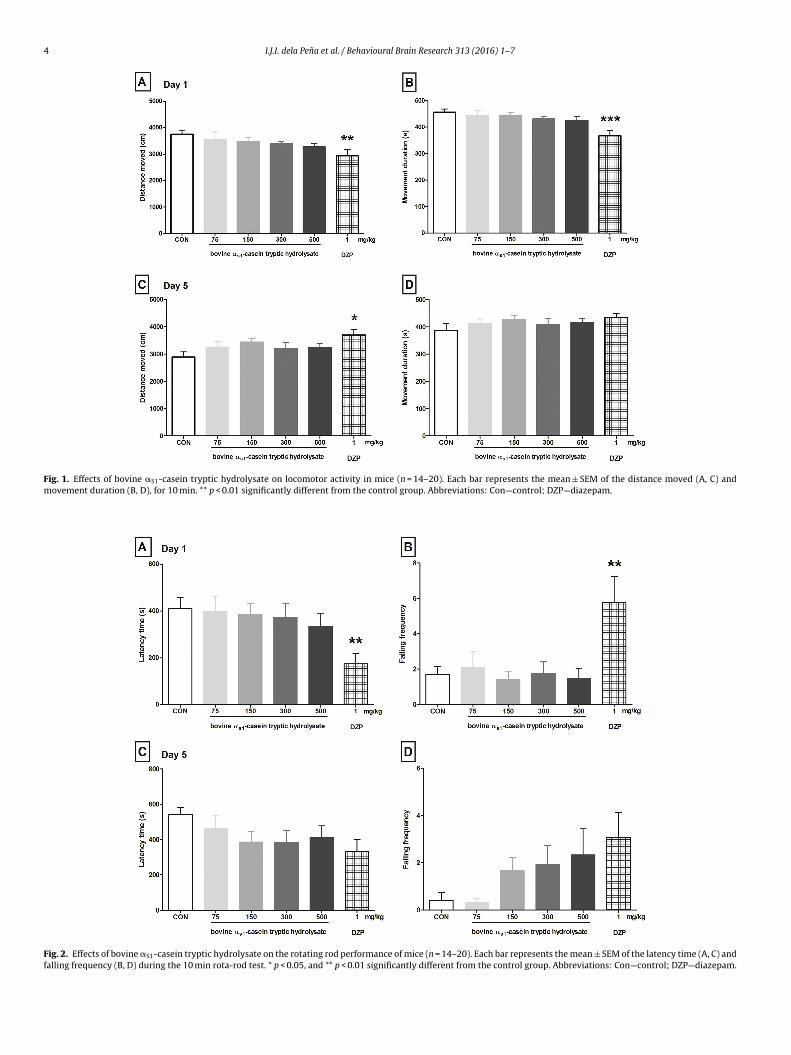
Fig. 1 shows the distance moved (A, C) and the movement dura-
ion (B, D) of animals treated with CH, diazepam, or vehicle (control
roup). One-way ANOVA showed significant differences in distance
oved (D1 [F (5, 92) = 2.37, p < 0.05], D5 [F (5, 74) = 2.15, p < 0.05)
nd movement duration (D1 [F (5, 92) = 4.49, p < 0.01]) between
he experimental groups. Acute (single) treatment of diazepam
emonstrated a significant decrease in distance moved ([q = 3.204];
ig. 1A) and movement duration ([q = 4.197]; Fig. 1B) compared to
the vehicle-treated (control) group, exhibiting diazepam’s sedative
effect. However, repeated treatment showed a significant increase
in distance moved ([q = 3.114]; Fig. 1C). CH-treated mice showed
a decreasing trend in distance moved and movement duration;
however, it did not show statistical significance compared with the
control group.
3.2. Rotarod test
Fig. 2 shows the latency to first fall (A, C) and falling frequency
(B, D) of mice treated with CH, diazepam, or vehicle (control group).
One-way ANOVA showed significant differences in latency to first
fall (D1 [F (5, 92) = 2.83, p < 0.05]) and falling frequency (D1 [F (5,
92) = 4.24, p < 0.01]) between the experimental groups. Diazepam-
treated mice significantly showed decrease in endurance/latency
time (D1 [q = 3.194]; Fig. 2A) and increased falling frequency (D1
[q = 3.477]; Fig. 2B) on the rotating rod, as compared to the control.
i
p
ain Research 313 (2016) 1–7 3
n support to the open-field test, CH-treated mice did not show a
ignificant difference in latency time and falling frequency.
.3. Pentobarbital-induced sleeping test
Fig. 3 shows the onset/loss of righting reflex (A, C) and total
duration (B, D) of sleep induced by pentobarbital sodium in
mice pretreated with CH, diazepam, or vehicle (control). One-
way ANOVA showed significant differences in sleep onset time
(D1 [F (5, 92) = 11.57, p < 0.001], D5 [F (5, 73) = 2.85, p < 0.01])
and sleeping duration (D1 [F (5, 92) = 34.47, p < 0.001], D5 [F (5,
73) = 5.00, p < 0.001]) between the experimental groups. Diazepam
treatment potentiated pentobarbital-induced sleeping behaviors
as shown by a decreased sleep onset time (D1 [q = 7.148], D5
[q = 3.382]; Fig. 3A) and increased total sleep duration time (D1
[q = 7.148], D5 [q = 4.141]; Fig. 3B & D). CH treatment also poten-
tiated pentobarbital-induced sleep duration at 150 mg/kg (D1
[q = 3.725], D5 [q = 2.616]; Fig. 3B & D).
3.4. Electroencephalogram (EEG) recording
Fig. 4 shows the total power or delta (A, C) and alpha (B, D)
aves in the EEG of rats treated with CH, diazepam, or vehicle.
ne-way ANOVA showed significant differences in slow (delta total
ower) [D1 F (3, 44) = 5.812, p < 0.01] and fast (alpha total power)
[F (3, 44) = 2.992, p < 0.05] waves. Diazepam-treated rats showed a
significant increase in delta waves (D1 [q = 3.939]; Fig. 4A) and a sig-
nificant decrease in alpha waves (D1 [q = 2.942]; Fig. 4B). CH-treated
ats also showed a significant increase (D1 [q = 2.673]; Fig. 4A) in
delta total power and a downward trend in alpha waves; however,
it did not reach statistical significance compared with the vehicle-
treated (control) rats.
3.5. Intracellular Cl− measurement assay
Fig. 5 shows the effects of CH on Cl− influx in human neurob-
lastoma cells in vitro. Treatment of CH increased Cl− influx in a
dose-dependent manner (Fig. 5A). Contrariwise, bicuculline, a com-
petitive GABAA receptor antagonist, inhibited the increased in Cl
influx induced by CH (Fig. 5B).
. Discussion
In the present study, our goal was to evaluate the sedative and
leep-promoting effects of bovine aS1-casein tryptic hydrolysate
CH). We have found that CH has sleep-promoting properties as evi-
enced by an augmented pentobarbital-induced sleep in mice and
n increased slow (delta) EEG wave in rats. This sleep-promoting
ffect is probably mediated through the GABAergic neurotransmit-
er system.
The probable sedative effect of CH was evaluated through the
pen-field and rota-rod test. Consistent with previous studies, the
enzodiazepine drug, diazepam, decreased (after single treatment)
he locomotor activity of mice in the open-field test (Fig. 1). A
decreased locomotor activity in this test is considered to reflect
the sedative effects of a substance [16]. However, mice repeatedly
treated with diazepam showed a significant increase in the open-
field test (Fig. 1). Similar results were observed by Pinna et al. [17],
suggesting that locomotor increase induced by diazepam in repeat-
edly treated subjects might be due to the reduction of neophobia or
drug tolerance [18,19]. Although a trend can be observed, CH did
not significantly alter the locomotor activity of mice. This result
indicates that this substance has no strong sedative effects. Similar
results were observed in the rota-rod test (Fig. 2). The rota- rod test
s another test used to evaluate the sedative and/or muscle-relaxant
roperties of a substance [20]. We have found that CH treatment did
4 I.J.I. dela Pena et al. / Behavioural Brain Research 313 (2016) 1–7
Fig. 1. Effects of bovine aS1-casein tryptic hydrolysate on locomotor activity in mice (n = 14–20). Each bar represents the mean ± SEM of the distance moved (A, C) and
movement duration (B, D), for 10 min. ** p < 0.01 significantly different from the control group. Abbreviations: Con—control; DZP—diazepam.
Fig. 2. Effects of bovine aS1-casein tryptic hydrolysate on the rotating rod performance of mice (n = 14–20). Each bar represents the mean ± SEM of the latency time (A, C) and
falling frequency (B, D) during the 10 min rota-rod test. * p < 0.05, and ** p < 0.01 significantly different from the control group. Abbreviations: Con—control; DZP—diazepam.
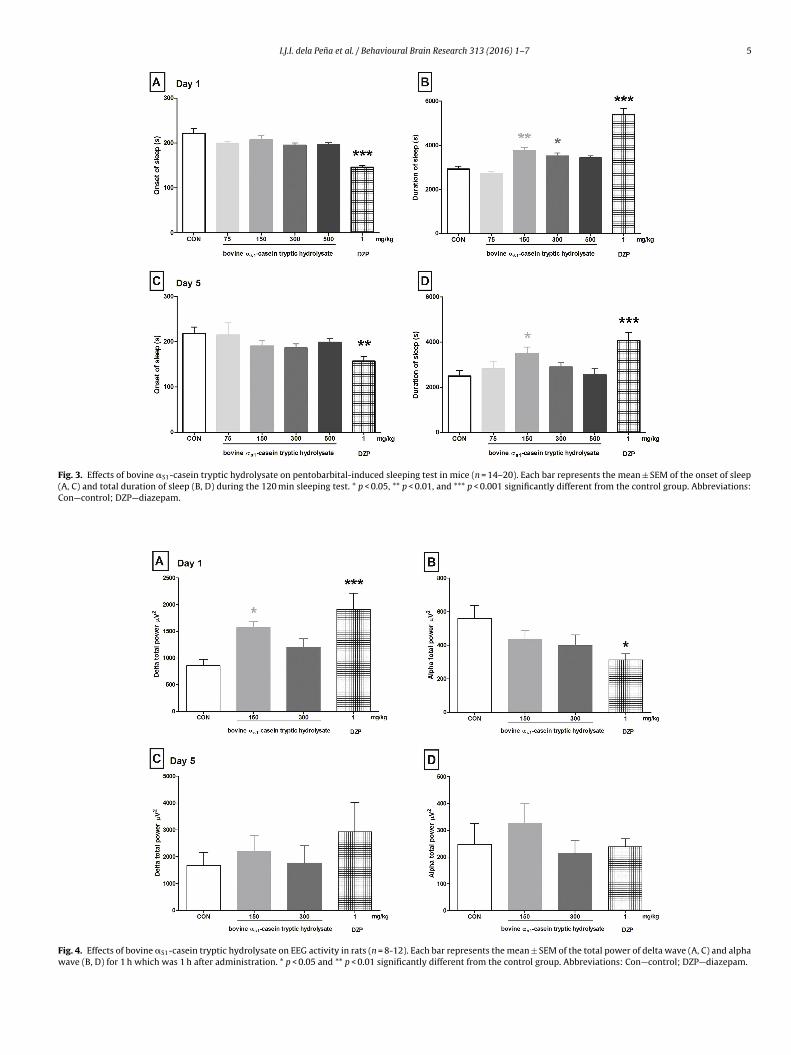
I.J.I. dela Pena et al. / Behavioural Brain Research 313 (2016) 1–7 5
Fig. 3. Effects of bovine aS1-casein tryptic hydrolysate on pentobarbital-induced sleeping test in mice (n = 14–20). Each bar represents the mean ± SEM of the onset of sleep
(A, C) and total duration of sleep (B, D) during the 120 min sleeping test. * p < 0.05, ** p < 0.01, and *** p < 0.001 significantly different from the control group. Abbreviations:
Con—control; DZP—diazepam.
Fig. 4. Effects of bovine aS1-casein tryptic hydrolysate on EEG activity in rats (n = 8-12). Each bar represents the mean ± SEM of the total power of delta wave (A, C) and alpha
wave (B, D) for 1 h which was 1 h after administration. * p < 0.05 and ** p < 0.01 significantly different from the control group. Abbreviations: Con—control; DZP—diazepam.

6 I.J.I. dela Pena et al. / Behavioural Brain Research 313 (2016) 1–7
a cell
a thoxy
p
n
d
l
O
m
t
p
b
c
i
s
e
[
[
[
[
[
Fig. 5. Effects of a bovine aS1-casein tryptic hydrolysate on [Cl−]i in neuroblastom
nd the emission wavelength at 450 nm using the Cl−-sensitive indicator, N-(6me
eak (F/F0). Abbreviations: BIC—bicuculline.
ot cause any significant motor alteration in this test. Diazepam still
emonstrated sedative effects as evidenced by decreased falling
atency and increased falling frequency. Together, the results of the
FT and rota-rod tests suggest that CH does not have or have very
inimal sedative effects.
In the pentobarbital-induced sleeping test, diazepam reduced
he sleep onset and increased the sleep duration of mice. CH also
otentiated the duration of sleep but not the onset of sleep induced
y pentobarbital sodium (Fig. 3). This result suggests that CH pos-
sesses sleep-enhancing effect, but it might lack sleep-inducing
effect. Substances that potentiate the pentobarbital-induced sleep
possess centrally acting sleep-promoting properties [21,22]. The
result of the pentobarbital-induced sleeping test is further sup-
ported by the results of the EEG monitoring in rats. EEG is
considered to be a vital and sensitive parameter during sleep
examination and is used to evaluate and diagnose sleep-related
problems/sleep disorders [23]. Treatment with CH (150 mg/kg) sig-
nificantly increased slow (delta) wave activity (Fig. 4), similar to
that of diazepam. This result is considered to be an indication of
sleep. Generally, high frequency (alpha) EEG waves are decreased
during sleep and, as the stages of sleep progress, delta waves gov-
ern the EEG tracing [23–25]. Together these results indicate that
CH has sleep-enhancing effects. This finding is in concordance with
previous studies showing that CH improves sleep in stressed sub-
jects [1,11,12] and consistent with the notion that this substance
ontributes to the sleep-promoting effects of milk.
It may seem confounding that in our experiments CH did not
nduce sedative effects but has sleep-promoting effects. However,
everal studies have suggested that sedative and sleep-promoting
ffects are related but dissociable processes [26,27]. Studies have
suggested that different brain regions mediate these effects. For
instance, a recent study has shown that slow-wave sleep, similar
to the one observed in the present study, is mediated by GABAergic
neurons in the medullary parafacial zone [28], while the seda-
tive effects induced by GABAergic agents are probably mediated
through the tuberomammillary nucleus [29]. Taking these things
into consideration, it is quite rational to conclude that CH pos-
sesses sleep-enhancing effects but is devoid of or has minimal
sedative effects. Further studies are needed to validate this claim
and to dissect the mechanisms by which these effects are medi-
ated. Regardless, our behavioral experiments have demonstrated
that CH has sleep-promoting effects in rodents.
We also investigated the probable mechanism behind the sleep-
enhancing effect of CH. It was reported that CH have a similar
structure with GABA and have an affinity for benzodiazepine
receptors, specifically GABAA receptor subtype [13]. Therefore,
we studied its action on Cl− influx neuroblastoma cells. Indeed,
CH significantly increased Cl− influx in a dose-dependent man-
ner (Fig. 5). Moreover, this CH-induced Cl− influx was blocked
by co-administration of bicuculline, a competitive GABAA recep-
tor antagonist. Altogether, these results strongly suggest that the[
s (SH-SY5Y). Fluorescence was monitored in the excitation wavelength at 365 nm
quinoyl) acetoetylester (MQAE). Contents of influxed Cl− ion were expressed as a
sleep-promoting effect of CH is mediated through GABAergic neu-
rotransmission, notably through the GABAA receptor-Cl− channel
complex.
In conclusion, the present study suggests that CH is capable of
facilitating and promoting sleep, with no or minimal sedative prop-
erties. The sleep-enhancing effects of CH are probably mediated
through the GABAA receptor-Cl− channel complex. CH might be
used as a natural sleep aid for managing sleep-related problems.
Acknowledgement
This work was supported by the Chungbuk Bio International
R&D Project 2014-1-01. We also would like to thank Ingredia SA
(Arras, France) for their supporting knowledge on Lactium®.
References
[1] Z. de Saint-Hilaire, M. Messaoudi, D. Desor, T. Kobayashi, Effects of a bovinealpha S1-casein tryptic hydrolysate (CTH) on sleep disorder in Japanesegeneral population, Open Sleep J. 2 (2009) 26–32.
[2] M.E. Thase, Correlates and consequences of chronic insomnia, Gen. Hosp.Psychiatry 27 (2) (2005) 100–112.
[3] Y. Hayashino, S. Fukuhara, Y. Suzukamo, T. Okamura, T. Tanaka, H. Ueshima,Relation between sleep quality and quantity, quality of life, and risk ofdeveloping diabetes in healthy workers in Japan: the High-risk andPopulation Strategy for Occupational Health Promotion (HIPOP-OHP) Study,BMC Public Health 7 (1) (2007) 1.
[4] K. Spiegel, R. Leproult, E.V. Cauter, Impact of sleep debt on metabolic andendocrine function, Lancet 354 (9188) (1999) 1435–1439.
[5] M. Kuppermann, D.P. Lubeck, P.D. Mazonson, D.L. Patrick, A.L. Stewart, D.P.Buesching, S.K. Filer, Sleep problems and their correlates in a workingpopulation, J. Gen. Intern. Med. 10 (1) (1995) 25–32.
[6] S. Uzun, O. Kozumplik, M. Jakovljevic, B. Sedic, Side effects of treatment withbenzodiazepines, Psychiatr. Danub. 22 (1) (2010) 90–93.
[7] J.H. Woods, G. Winger, Current benzodiazepine issues, Psychopharmacology118 (2) (1995) 107–115.
[8] I.J.I. dela Pena, E. Hong, J.B. de la Pena, H.J. Kim, C.J. Botanas, Y.S. Hong, Y.S.Hwang, B.S. Moon, J.H. Cheong, Milk collected at night induces sedative andanxiolytic-like effects and augments pentobarbital-induced sleeping behaviorin mice, J. Med. Food 18 (11) (2015) 1255–1261.
[9] N.P. Shah, Effects of milk-derived bioactives: an overview, Br. J. Nutr. 84 (S1)(2000) 3–10.
10] H. Teschemacher, Casein-derived opioid peptides: physiological significance,Adv. Biosci. 65 (1987) 41.
11] J.H. Kim, D. Desor, Y.T. Kim, W.J. Yoon, K.S. Kim, J.S. Jun, K.H. Pyun, I. Shim,Efficacy of as1-casein hydrolysate on stress-related symptoms in women,Eur. J. Clin. Nutr. 61 (4) (2007) 536–541.
12] B. Guesdon, M. Messaoudi, C. Lefranc-Millot, G. Fromentin, D. Tomé, P.C. Even,A tryptic hydrolysate from bovine milk as1-casein improves sleep in ratssubjected to chronic mild stress, Peptides 27 (6) (2006) 1476–1482.
13] L. Miclo, E. Perrin, A. Driou, V. Papadopoulos, N. Boujrad, R. Vanderesse, J.Boudier, D. Desor, G. Linden, J. Gaillard, Characterization of a-casozepine, atryptic peptide from bovine as1-casein with benzodiazepine-like activity,FASEB J. 15 (10) (2001) 1780–1782.
14] M. Messaoudi, C. Lefranc-Millot, D. Desor, B. Demagny, L. Bourdon, Effects of atryptic hydrolysate from bovine milk aS1-casein on hemodynamic responses
in healthy human volunteers facing successive mental and physical stresssituations, Eur. J. Clin. Nutr. 44 (2) (2005) 128–132.15] I.J.I. dela Pena, H.L. Lee, S.Y. Yoon, J.B. de la Pena, K.H. Kim, E.Y. Hong, J.H.Cheong, The ethanol extract of Cirsium japonicum increased chloride ioninflux through stimulating GABAA receptor in human neuroblastoma cells

ral Br
[
[
[
[
[
[
[
[
[
[
[
[
[GABAergic parafacial zone is a medullary slow wave sleep-promoting center,Nat. Neurosci. 17 (9) (2014) 1217–1224.
[29] L.E. Nelson, T.Z. Guo, J. Lu, C.B. Saper, N.P. Franks, M. Maze, The sedativecomponent of anesthesia is mediated by GABAA receptors in an endogenoussleep pathway, Nat. Neurosci. 5 (10) (2002) 979–984.
I.J.I. dela Pena et al. / Behaviou
and exhibited anxiolytic-like effects in mice, Drug Discov. Ther. 7 (1) (2013)18–23.
16] X. Zhou, X. Zhao, L. Tang, Y. Zhang, H. Ruan, H. Pi, J. Qiu, J. Wu,Immunomodulatory activity of the rhizomes of Impatiens pritzellii var.hupehensis on collagen-induced arthritis mice, J. Ethnopharmacol. 109 (3)(2007) 505–509.
17] G. Pinna, R.C. Agis-Balboa, A. Zhubi, K. Matsumoto, D.R. Grayson, E. Costa, A.Guidotti, Imidazenil and diazepam increase locomotor activity in miceexposed to protracted social isolation, Proc. Natl. Acad. Sci. U.S.A. 103 (11)(2006) 4275–4280.
18] F. Crestani, J.R. Martin, H. Möhler, U. Rudolph, Mechanism of action of thehypnotic zolpidem in vivo, Br. J. Pharmacol. 131 (7) (2000) 1251–1254.
19] R.M. McKernan, T.W. Rosahl, D.S. Reynolds, C. Sur, K.A. Wafford, J.R. Atack, S.Farrar, et al., Sedative but not anxiolytic properties of benzodiazepines aremediated by the GABAA receptor a1 subtype, Nat. Neurosci. 3 (6) (2000)587–592.
20] S. Farkas, P. Berzsenyi, E. Kárpáti, P. Kocsis, I. Tarnawa, Simplepharmacological test battery to assess efficacy and side effect profile ofcentrally acting muscle relaxant drugs, J. Pharmacol. Toxicol. Methods 52 (2)(2005) 264–273.
21] H. Ma, C. Kim, Y. Ma, S. Nam, D. Kim, S. Woo, J. Hong, K. Oh, Magnolol
enhances pentobarbital-induced sleeping behaviors: possible involvement ofGABAergic systems, Phytother. Res. 23 (9) (2009) 1340–1344.22] F.C.F. De Sousa, B.A. Pereira, V.T.M. Lima, C.D.G. Lacerda, C.T.V. Melo, J.M.Barbosa-Filho, S.M.M. Vasconcelos, G.S.B. Viana, Central nervous systemactivity of yangambin from Ocotea duckei Vattimo (Lauraceae) in mice,
ain Research 313 (2016) 1–7 7
Phytother. Res. 19 (4) (2005) 282–286.23] V.M. Kumar, Sleep and sleep disorders, Indian J. Chest Dis. Allied Sci. 50 (1)
(2008) 129.24] S.M. Stahl, Disorders of sleep and wakefulness and their treatment, in: N.
Muntner (Ed.), Stahl’s Essential Psychopharmacology: Neuroscientific Basisand Practical Applications, Cambridge University Press, New York, 2008, pp.815–862.
25] H. van Lier, W.H. Drinkenburg, Y.J. van Eeten, A.M. Coenen, Effects ofdiazepam and zolpidem on EEG beta frequencies are behavior-specific in rats,Neuropharmacology 47 (2) (2004) 163–174.
26] C. Guilleminault, Efficacy of agomelatine versus venlafaxine on subjectivesleep of patients with major depressive disorder, Eur.Neuropsychopharmacol. 15 (2005) S419–S420.
27] R.W. Lam, Sleep disturbances and depression: a challenge for antidepressants,Int. Clin. Psychopharmacol. 21 (2006) S25–S29.
28] C. Anaclet, L. Ferrari, E. Arrigoni, C.E. Bass, C.B. Saper, J. Lu, P.M. Fuller, The










![A - Benzodiazepine-Chloride Receptor-Targeted Therapy for ......nisms through GABAA and GABAB receptors [12]. GABA is classified into two main categories: GABAA and GABAB. GABAA and](https://img.pdfslide.us/doc/110x75/60f82a0e0bab2d34196b5ccd/a-benzodiazepine-chloride-receptor-targeted-therapy-for-nisms-through.jpg)


















