Embed Size (px)
Citation preview

A systematic evaluation of basidiospore symmetry and tegumentation in hypogeous and gasteroid Russulales
STEVEN L. MILLER Botany Department, Universiiy of Wyoming, Lararne, WY 82071, U.S.A.
Received September 8, 1987
MILLER, S. L. 1988. A systematic evaluation of basidiospore symmetry and tegumentation in hypogeous and gasteroid Rus- sulales. Can. J. Bot. 66: 2561 -2573.
Spore wall architecture and ontogeny of ornamentation in several genera and species of hypogeous and gasteroid Russulales are similar to those described previously for agaricoid Lactarius lignyotellus. Spore walls are composed of four layers, each differing in thickness and electron density. Layer 2 is electron transparent and corresponds to a dark blue, amyloid layer when mounted in Melzer's iodine reagent and viewed with the light microscope. Establishment of spore symmetry may be regulated by the hilar appendix body, which is a poorly differentiated cytoplasmic region in the hilar appendix of asymmetric spores of Macowanites luteolus, Elasmomyces russuloides, and Zelleromyces versicaulus but which is absent in symmetric spores of Z. sculptisporus, Martellia subochracea, and Gymnomyces yubaensis. A continuum in spore morphology from truly symmetric to asymmetric is evident in spores from individual sporocarps of many species of the Russulales. The variation in spore symmetry and spore surface ornamentation has clouded taxonomic concepts in the Russulales. Systematically, development of orthotropic and heterotropic spores has been regarded as two distinct end points of evolution, when they are likely terms describing degrees of the same phenomenon. The current circumscription of families and genera in the Russulales based on spore symmetry, therefore, appears to be artificial.
MILLER, S. L. 1988. A systematic evaluation of basidiospore symmetry and tegumentation in hypogeous and gasteroid Rus- sulales. Can. J. Bot. 66 : 2561 -2573.
Chez plusieurs genres et espbces de Russulales hypogees et gastkroides, l'architecture de la paroi sporale et 1'ontogCnbse de l'ornementation sont identiques B ce qui a dCjB CtC dCcrit pour l'espbce agaricoide, Lactarius lignyotellus. Les parois sporales sont constituCes de quatre couches, chacune diffkrente par son Cpaisseur et sa densit6 aux Clectrons. La couche 2 est transparente aux Clectrons; elle correspond B une couche amyloide qui se colore en bleu foncC, lorsque montCe dans le rCactif de Melzer iodC et observCe en microscopie photonique. L'Ctablissement de la symCtrie sporale pourrait &tre rCglC par le corps de l'appendice hilaire, une region cytoplasmique faiblement diffCrenciCe de l'appendice hilaire, present chez les spores asymktriques de Macowanites luteolus, Elasmomyces russuloides, et Zelleromyces versicaulus, mais absent dans les spores symCtriques de Z. sculptisporus, Martellia subochracea et Gymnomyces yubaensis. Chez plusieurs espbces de Rus- sulales, les spores provenant de sporocarpes individuels montrent un continuum Cvident dans la morphologie sporale B partir de la symCtrie complbte jusqu'i I'asymCtrie. La variation de la symCtrie et de l'ornementation sporales rend confus les con- cepts taxonomiques sur les Russulales. Sur le plan systkmatique, les dCveloppements de spores orthotropiques et hCtCro- tropiques ont CtC considCrCs comme deux termes Cvolutifs distincts alors que ce sont vraisemblablement deux termes qui dkcrivent des degrCs du m&me phCnombne. I1 semble donc que, chez les Russulales, la disposition actuelle des familles et des genres basei sur la symCtrie sporale soit artificielle.
[Traduit par la revue]
Attempts at formulating natural classifications for Basidio- mycetes by combining hypogeous, gasteroid, and agaricoid taxa into "evolutionary series" have met with difficulty. Although affinities between agarics or boletes and particular gasteroid taxa have been generally agreed upon, the direction of evolution has been hotly argued. Singer (1938, 1971, 1975) and Smith (1971) suggested that agaricoid genera were derived from gasteromycetous ancestors. However, Heim (1931, 1936a, 19366, 1937a, 19376, 1938, 1943, 1948, 1971), Malen~on (1931a), Thiers (1971, 1984), and Singer and Smith (1960) proposed that gasteroid Basidiomycetes have evolved from agaricoid forms through a retrogressive or simplification process. The proper systematic placement of gasteroid Basid- iomycetes is a complex problem and one not easily resolved with current knowledge (Donk 1971).
The Russulales sensu Pegler and Young (1979, 1981) and Beaton et al. (1984) comprises a homogeneous series of agari- coid, gasteroid and hypogeous Basidiomycetes. Taxa in the Russulales share similarities in basidiocarp architecture, tissue differentiation, and spore ornamentation, even though some genera possess an entirely open hymenium at maturity, while in others the hymenium never becomes exposed. Pegler and Young (1979, 1981) proposed delimitation of two families in the Russulales. The Russulaceae, containing Russula, Lac- tarius, Arcangeliella, Cystangium, and Macowanites, are
agaricoid to gasteroid, with spores that are asymmetric (heterotropic) and forcibly or nonforcibly discharged. The Elasmomycetaceae, containing Elasmomyces, Martellia, Gymnomyces, and Zelleromyces, are hypogeous or gasteroid, with spores that are symmetric (orthotropic), and putatively lack forcible discharge. Basidiospore morphology and spore discharge have been emphasized as fundamental characteris- tics in classification of families and genera in the Russulales.
Taxonomic study of hypogeous and gasteroid species in the Russulales has revealed that basidiocarp architecture, tissue differentiation, and spore morphology can vary considerably between species and between different basidiocarps of the same species (Pegler and Young 1979, 1981; Beaton et al. 1984; Miller and Miller 1986). With such variation in impor- tant, but poorly understood, taxonomic characteristics, natural family and generic boundaries are difficult to establish. This study evaluates spore symmetry and wall tegumentation dur- ing basidiospore formation in hypogeous and gasteroid Basid- iomycetes, and their significance in the classification of the Russulales.
Materials and methods
Fresh glebal tissues from Macowanites luteolus Smith & Trappe (OKM 20973, VPI), Elasmomyces russuloides Setchell (HS 1973,
Prmmd in Canada 1 lrnprirni au Canada
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/3
0/14
For
pers
onal
use
onl
y.

2562 CAN. J. BOT. VOL. 66. 1988
SFSC), Gymnomyces yubaensis H. Saylor nom. prov. (HS 1676, SFSC), Zelleromyces versicaulus S. L. Miller nom. prov. (OKM 20862, VPI), Z. sculptisporus S. L. Miller nom. prov. (SLM 764, VPI), Z. cinnabarinus Sing. & Smith (SLM 721, VPI), Z. gilkeyae Sing. & Smith (T7746, OSC), and Martellia subochracea Smith (T 7467, OSC) were cut into 1- to 3-mm radial sections and fixed in 2% glutaraldehyde in 0.2 M phosphate buffer, pH 6.8, for 12 h at room temperature. Herbarium names are abbreviated accord- ing to Holmgren and Keuken (1974). Tissues were then washed three times in the same buffer and postfixed in buffered 1 % osmium tet- roxide for 1 h. Specimens were dehydrated in a graded ethanol series, using 30-min intervals, three changes of 100% EtOH, followed by two changes of propylene oxide. Infiltration was in propylene oxide - polylbed 812 resin (5050, vlv) (Shearer and Hunsicker 1980) overnight, followed by 100% resin for 4 h. Tissues were then flat embedded in 100% polylbed 8 12 resin and allowed to polymerize for 48 h at 60°C. Selected blocks were initially thick sectioned between 0.5 and 1 pm on a Sorvall Porter Blum MT-1 ultramicro- tome to obtain longitudinal orientation of basidia and spores. Thick sections were stained with 0.05% toluidine blue in 0.5% sodium borate, pH 9.5, and examined with light microscopy. Selected blocks were retrimmed to the desired area and serial sectioned with a diamond knife using an American Optical Ultracut microtome. Thin sections were mounted on uncoated 200-mesh "slim-bar" grids (Structure Probe Inc.) and stained with 1 % uranyl acetate for 15 min, followed by 1 % lead citrate for 5 min. Transmission electron micros- copy (TEM) of specimens was on a JEOL 100-C. Spores for light microscopy were mounted in Melzer's iodine reagent modified from Melzer (1924). An attempt was made to model stages in basidiospore development and terminology on McLaughlin (1977) and Yoon and McLaughlin (1979, 1984). Wall teguments were numbered from the spore surface inward based on maximum number of layers observed and not on ontogeny. Spore size includes the hilar appendix to pro- vide a standard point of reference for each spore; sizes were calcu- lated from calibrated electron micrographs at each magnification and with light microscopy using a calibrated ocular micrometer. The maximum number of layers were not always present in each spore wall and apparent fusion of individual layers made characterization difficult. Figures are TEM micrographs unless otherwise indicated.
Results Macowanites luteolus, Elasmomyces russuloides and Zellero-
myces versicaulus Mature hymenia contained basidia and nonsynchronous
spores at various developmental stages. Sterigmata developed to their full length before basidiospore initiation began.
Stage 1 , inception Initiation of spore development was characterized by swell-
ing of the sterigmal apex to form a spherical apophysis (Figs. 1-3, 6, 39, 40). The diameter of spore initials ranged from 0.8 to 1.3 um (Table 1). A thick electron-dense outer layer and a thin election-transparent inner layer, correspond- ing to layers 3 and 4 present during later spore development, were distinct in the sterigmata (Fig. 3) and some develop- ing spores. Electron-dense layer 1 and electron-transparent layer 2 were distinguishable in some spores (Fig. 2).
A hilar appendix body was evident in the three species as a poorly differentiated, pyrimidal, cytoplasmic region at the adaxial wall near the junction of the basidiospore initial and sterigmal apex (Figs. 1-3, 6). It was granular in texture and was electron dense or similar in electron density to the sur- rounding cytoplasm, but with an electron-dense boundary.
Stage 2 , asymmetric growth Spores grew asymmetrically toward the abaxial side. Spore
shape in the three species was subcylindrical during early
stage 2, as shown for 2. versicaulus in Fig. 4 , with the spore enlarging abaxially and adaxially late in stage 2 (Figs. 39,40). The spores had enlarged three to five times their size by stage 2 (Table 1). Wall layers 3 and 4 were prominent in the hilar appendix region of most spores at stage 2. Layer 2 became noticeable in small patches or as a continuous layer in late stage 2 spores of E. russuloides (Fig. 7). Spore cytoplasm was dominated by large vesicles with dense inclusions (Fig. 4).
The hilar appendix body present in stage 2 spores was situ- ated near the adaxial wall of the hilar appendix and resembled the hilar appendix body of stage 1 spores in size, shape, and location for respective species, as shown for E. russuloides in Fig. 7. The electron density of the hilar appendix body was somewhat greater than in stage 1.
Stage 3 , equal enlargement Spores enlarged abaxially, adaxially, and apically (Figs. 39,
40; Table l) , and initiation of spore surface ornamentation began. Mitochondria, as well as a number of vacuoles, were abundant in the spore cytoplasm (Figs. 5, 8). Large vesicles with dense inclusions observed in stage 2 were scattered throughout the cytoplasm in early stage 3 but were located near the periphery of the cytoplasm or nearly absent by late stage 3 (Fig. 5).
Four wall layers were present in stage 3 spores (Fig. 5). Layers 1 ,2 , and 4 increased in thickness compared with previ- ous stages, while layer 3 remained the same. In early stage 3, layer 2 protruded through layer 1 and by late stage 3, layers 1 and 2 had formed heterogeneous mounds of electron-dense and electron-transparent ornamentation on the surface of the spore (Figs. 5, 8).
The hilar appendix body was prominent during early stage 3 in M. luteolus (Fig. 5); it was less evident in E. russuloides and 2. versicaulus. A vague granular region with a moderately electron dense boundary, corresponding to the size, shape, and location of the hilar appendix body in stage 2, was appar- ent near the adaxial wall of E. russuloides spores (Figs. 8, 9).
Stage 4, elongation Maximum spore dimensions were reached during this stage
in all three species (Table l ) , and the greatest increase was in length (Figs. 39, 40). Stage 4 cytoplasm was characterized by numerous mitochondria, sheets or segments of endoplasmic reticulum (ER), lipid droplets, and the accumulation of glyco- gen (Fig. 10).
Differentiation in spore wall layers was pronounced during stage 4, but the spore wall still consisted of four layers of recognizable materials. Layer 4 thickened unevenly in an acropetal manner from the lateral abaxial and adaxial region above the hilar appendix. Layer 3 remained as a moderately electron dense zone between layers 4 and 2. Increases in orna- mentation occurred from - accumulations of electron-trans- parent material, similar to that found in layer 2, which was surrounded by electron-dense layer 1. No hilar appendix body was observed during stage 4 in any of the three species.
Martiella subochracea, Zelleromyces sculptisporus and Gym- nomyces y ubaensis
The hymenium contained basidia and nonsynchronous basidiospores at various developmental stages. Isolated patches of synchronous basidiospore development were observed in some basidiocarps. Sterigmata developed to their maximum length before basidiospore initiation began. A weakly differ-
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/3
0/14
For
pers
onal
use
onl
y.

MILLER
TABLE 1. Spore dimensions, shape, and number of spores examined for each species at each developmental stage
No. of spores Species and stage examined" Shape" Length and width (pm)"
Macowanites lureolus 1 2 3 . 4
Zelleromyces versicaulus 1 2 3 4
Elaslnolnyces russuloides 1 2 3 4
Gymnomyces yubaensis 1 2 3 4
Marrellia subochracea 1 2 3 4
Zelleromyces sculprisporus 1
Spherical Subcylindrical to obpyriform Broadly elliptical Broadly elliptical
Spherical Subcylindrical to obpyriform Elliptical to broadly elliptical Elliptical to broadly elliptical
Spherical Subcylindrical to obpyriform Subglobose to broadly elliptical Subglobose to broadly elliptical
Spherical Cylindrical to obpyriform Subglobose to obpyriform Subglobose to globose
Spherical Subglobose Subglobose Subglobose to globose
Spherical Cylindrical to obpyriform Globose to subglobose Globose to subglobose
"Transmission electron microscopy. " ~ i ~ h t and transmission electron microscopy
entiated region of cytoplasm was observed near the apex of continuous electron-transparent mounds between layers 3 and some developing sterigmata of G. yubaensis (Figs. 11, 12). 1, and often protruded beyond layer 1 (Figs. 14, 15, 17). Light The region was adherent to the adaxial wall of the sterigma microscopy of stage 2 Z. sculptisporus spores mounted in and was electron dense or moderately electron dense with an Melzer's reagent revealed that layer 2 was amyloid, turning electron-dense boundary. dark blue (Fig. 16). In late stage 2 G. yubaensis spores,
Stage 1 , inception The basidiospore initial first appeared as a spherical to
slightly cylindrical apophysis at the sterigmal apex (Figs. 41, 42; Table 1). In G. yubaensis the spore wall consisted of two layers: a thick electron-dense, outer layer 3 and a thin, elec- tron-transparent inner layer 4, and was continuous with simi- lar layers in the sterigma and basidial apex.
Stage 2 , elongation and symmetrical enlargement Spores of Z. sculptisporus and G. yubaensis increased in
length and were subglobose to subcylindrical throughout early stage 2 (Table I), and enlarged laterally in late stage 2 (Figs. 13, 14, 41, 42). Spores of Martellia subochracea were subglobose from inception (Figs. 15, 41, 42; Table 1). Large vesicles with dense inclusions were abundant throughout the spore cytoplasm (Fig. 15). Small lipid droplets and small seg- ments of ER were present (Fig. 15) and mitochondria were observed in some late stage 2 spores.
Four wall layers were present in stage 2 spores. Layer 1 appeared in stage 2 as an electron-dense layer outside of layer 3 and capped the spore, terminating near the sterigmal apex (Fig. 14). Layer 2 appeared as a distinct layer or as dis-
layer 2 was evident as a continuous layer around the spore, terminating at the sterigmal apex near the lowermost extension of layer 1 (Fig. 17). Layers 1 and 2 formed heterogeneous mounds or layers of electron-dense and electron-transparent material in late stage 2, as shown for Martellia subochracea in Figs. 25 and 26.
Spore symmetry varied among the three species and from spore to spore (Figs. 41 -42). Zelleromyces sculptisporus, and G. yubaensis spores examined with TEM were mostly sym- metrical about the axis of the sterigma, as shown for Z. sculp- tisporus in Figs. 13 and 14,. although asymmetric spores of G. yubaensis were observed with light microscopy. Spores of Martellia subochracea were observed both with TEM and light microscopy to be symmetric to slightly asymmetric, with the asymmetry most obvious in the angle of the hilar appendix and growth of the abaxial side (Figs. 18, 41, 42).
Stage 3, midenlargement Spores of the three species enlarged primarily in length, but
also adaxially and abaxially during this stage (Table I), and surface ornamentation continued to develop. Many spores at early stage 3 were partially collapsed or somewhat distorted in
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/3
0/14
For
pers
onal
use
onl
y.
CAN. J. BOT. VOL. 66. 1988
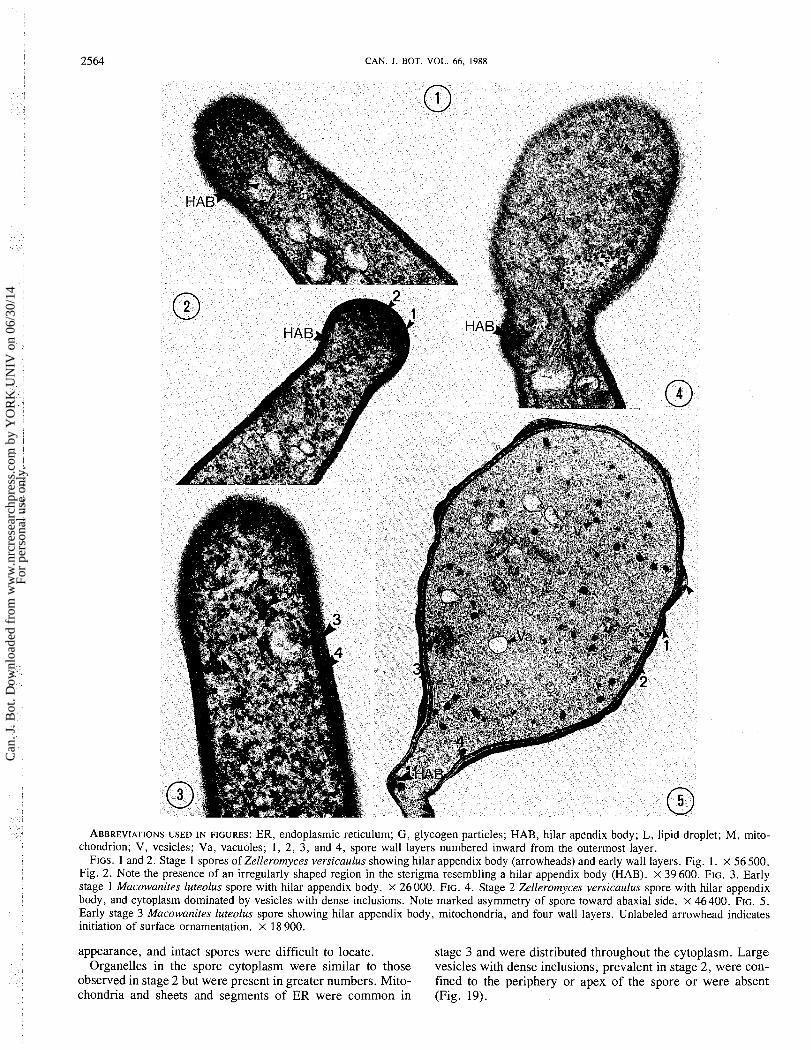
ABBREVIATIONS USED IN FIGURES: ER, endoplasmic reticulum; G , glycogen particles; HAB, hilar apendix body; L, lipid droplet; M, mito- chondrion; V, vesicles; Va, vacuoles; 1, 2, 3, and 4, spore wall layers numbered inward from the outermost layer.
FIGS. 1 and 2. Stage 1 spores of Zelleromyces versicaulus showing hilar appendix body (arrowheads) and early wall layers. Fig. 1. x 56 500. Fig. 2. Note the presence of an irregularly shaped region in the sterigma resembling a hilar appendix body (HAB). X 39 600. FIG. 3. Early stage 1 Macowanites luteolus spore with hilar appendix body. X 26000. FIG. 4. Stage 2 Zelleromyces versicaulus spore with hilar appendix body, and cytoplasm dominated by vesicles with dense inclusions. Note marked asymmetry of spore toward abaxial side. X 46400. FIG. 5. Early stage 3 Macowanites luteolus spore showing hilar appendix body, mitochondria, and four wall layers. Unlabeled arrowhead indicates initiation of surface ornamentation. x 18 900.
appearance, and intact spores were difficult to locate. stage 3 and were distributed throughout the cytoplasm. Large Organelles in the spore cytoplasm were similar to those vesicles with dense inclusions, prevalent in stage 2, were con-
observed in stage 2 but were present in greater numbers. Mito- fined to the periphery or apex of the spore o r were absent chondria and sheets and segments of ER were common in (Fig. 19).
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/3
0/14
For
pers
onal
use
onl
y.
MILLER
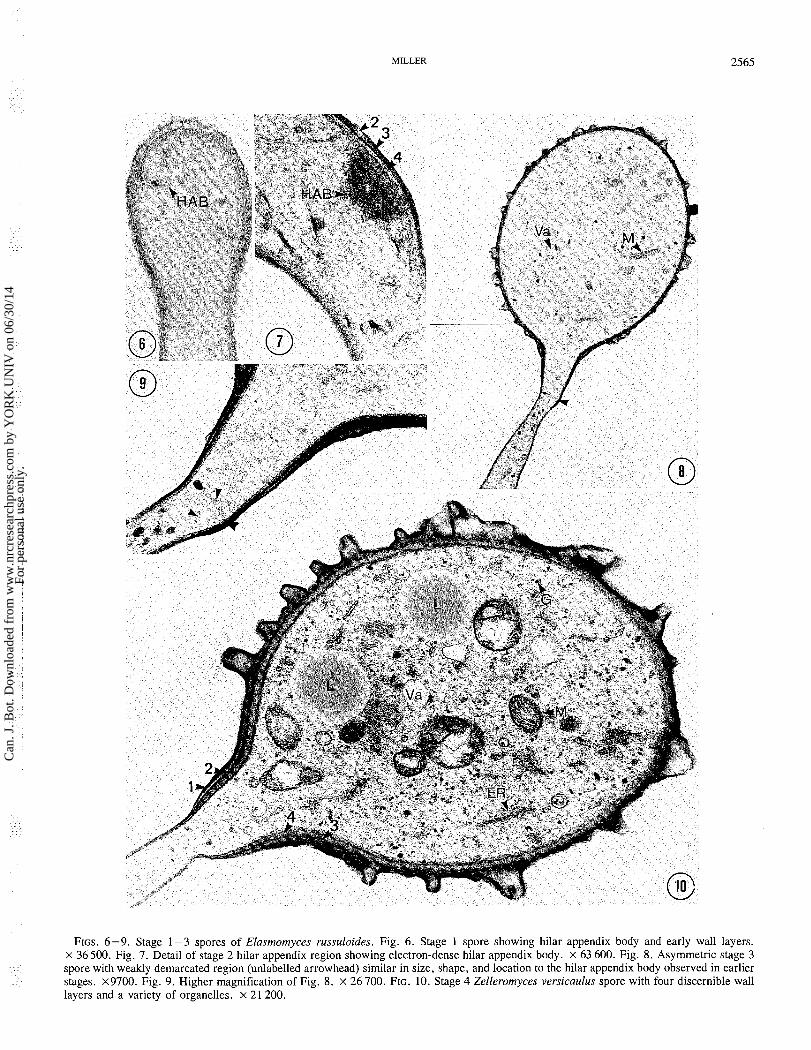
FIGS. 6-9. Stage 1-3 spores of Elasmomyces russuloides. Fig. 6. Stage 1 spore showing hilar appendix body and early wall layers. x 36500. Fig. 7. Detail of stage 2 hilar appendix region showing electron-dense hilar appendix body. x 63 600. Fig. 8. Asymmetric stage 3 spore with weakly demarcated region (unlabelled arrowhead) similar in size, shape, and location to the hilar appendix body observed in earlier stages. x9700. Fig. 9. Higher magnification of Fig. 8. x 26 700. FIG. 10. Stage 4 Zelleromyces versicaulus spore with four discernible wall layers and a variety of organelles. x 21 200.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/3
0/14
For
pers
onal
use
onl
y.
2566 CAN. 1. BOT. VOL. 66, 1988
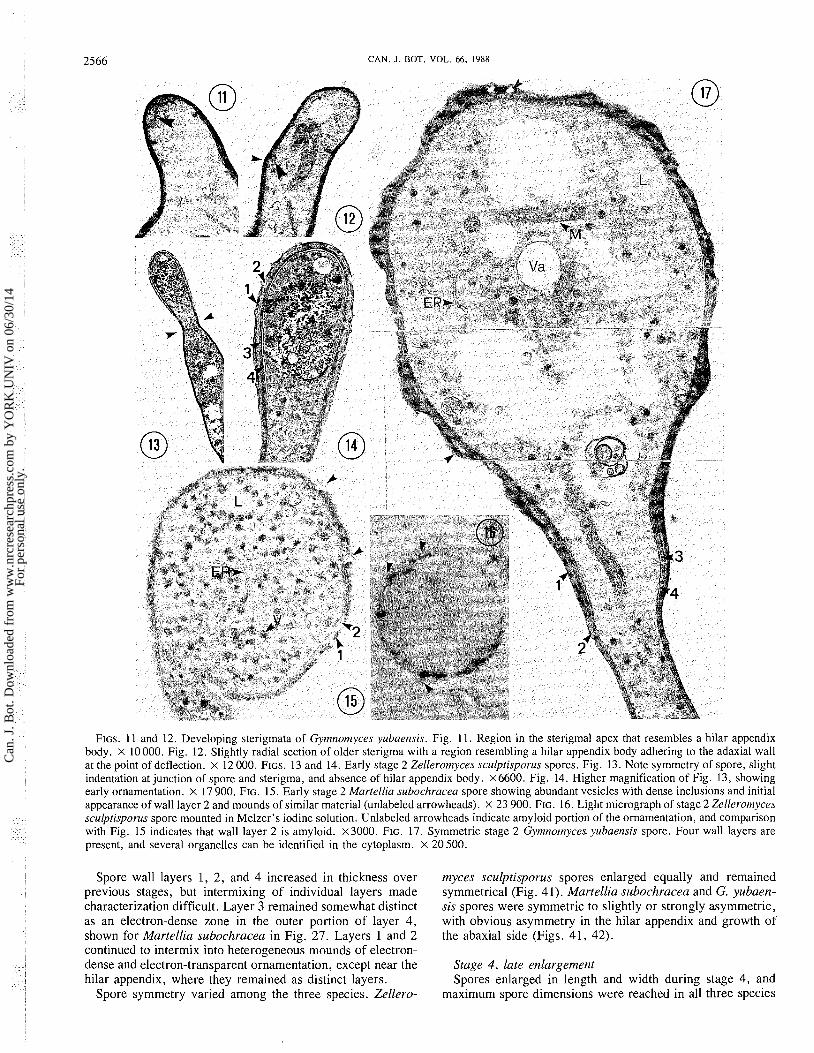
FIGS. 11 and 12. Developing sterigmata of Gymnomyces yubaensis. Fig. 11. Region in the sterigmal apex that resembles a hilar appendix body. X 10000. Fig. 12. Slightly radial section of older sterigma with a region resembling a hilar appendix body adhering to the adaxial wall at the point of deflection. x 12 000. FIGS. 13 and 14. Early stage 2 Zellerotnyces sculptisporus spores. Fig. 13. Note symmetry of spore, slight indentation at junction of spore and sterigma, and absence of hilar appendix body. ~ 6 6 0 0 . Fig. 14. Higher magnification of Fig. 13, showing early ornamentation. x 17 900. FIG. 15. Early stage 2 Martellia subochracea spore showing abundant vesicles with dense inclusions and initial appearance of wall layer 2 and mounds of similar material (unlabeled arrowheads). X 23 900. FIG. 16. Light micrograph of stage 2 Zellerotnyces sculptisporus spore mounted in Melzer's iodine solution. Unlabeled arrowheads indicate amyloid portion of the ornamentation, and comparison with Fig. 15 indicates that wall layer 2 is amyloid. x3000. FIG. 17. Symmetric stage 2 Gytnnotnyces yubaensis spore. Four wall layers are present, and several organelles can be identified in the cytoplasm. X 20 500.
Spore wall layers 1, 2, and 4 increased in thickness over previous stages, but intermixing of individual layers made characterization difficult. Layer 3 remained somewhat distinct as an electron-dense zone in the outer portion of layer 4, shown for Martellia subochracea in Fig. 27. Layers 1 and 2 continued to intermix into heterogeneous mounds of electron- dense and electron-transparent ornamentation, except near the hilar appendix, where they remained as distinct layers.
Spore symmetry varied among the three species. Zellero-
myces sculptisporus spores enlarged equally and remained symmetrical (Fig. 41). Martellia subochracea and G. yubaen- sis spores were symmetric to slightly or strongly asymmetric, with obvious asymmetry in the hilar appendix and growth of the abaxial side (Figs. 41, 42).
Stage 4, late enlargement Spores enlarged in length and width during stage 4, and
maximum spore dimensions were reached in all three species
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/3
0/14
For
pers
onal
use
onl
y.
MILLER
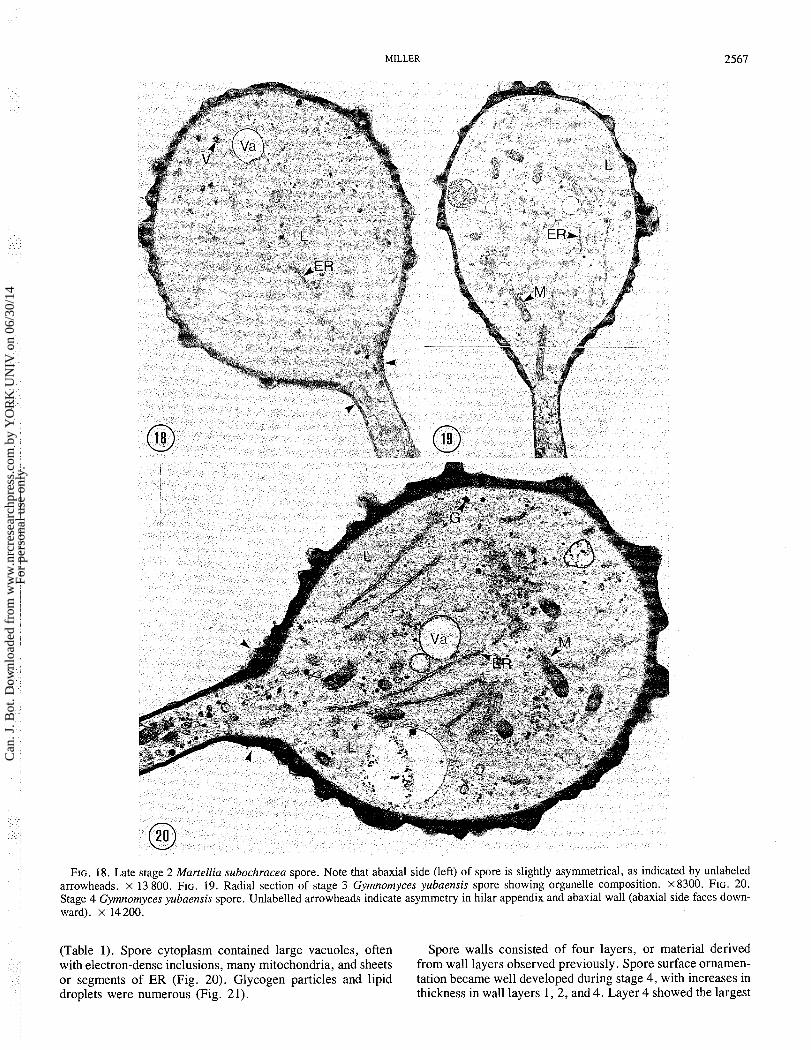
FIG. 18. Late stage 2 Martellia subochracea spore. Note that abaxial side (left) of spore is slightly asymmetrical, as indicated by unlabeled arrowheads. x 13 800. FIG. 19. Radial section of stage 3 Gymnomyces yubaensis spore showing organelle composition. x8300. FIG. 20. Stage 4 Gymnomyces yubaensis spore. Unlabelled arrowheads indicate asymmetry in hilar appendix and abaxial wall (abaxial side faces down- ward). x 14 200.
(Table 1). Spore cytoplasm contained large vacuoles, often Spore walls consisted of four layers, or material derived with electron-dense inclusions, many mitochondria, and sheets from wall layers observed previously. Spore surface ornamen- or segments of ER (Fig. 20). Glycogen particles and lipid tation became well developed during stage 4, with increases in droplets were numerous (Fig. 21). thickness in wall layers 1 , 2 , and 4. Layer 4 showed the largest
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/3
0/14
For
pers
onal
use
onl
y.
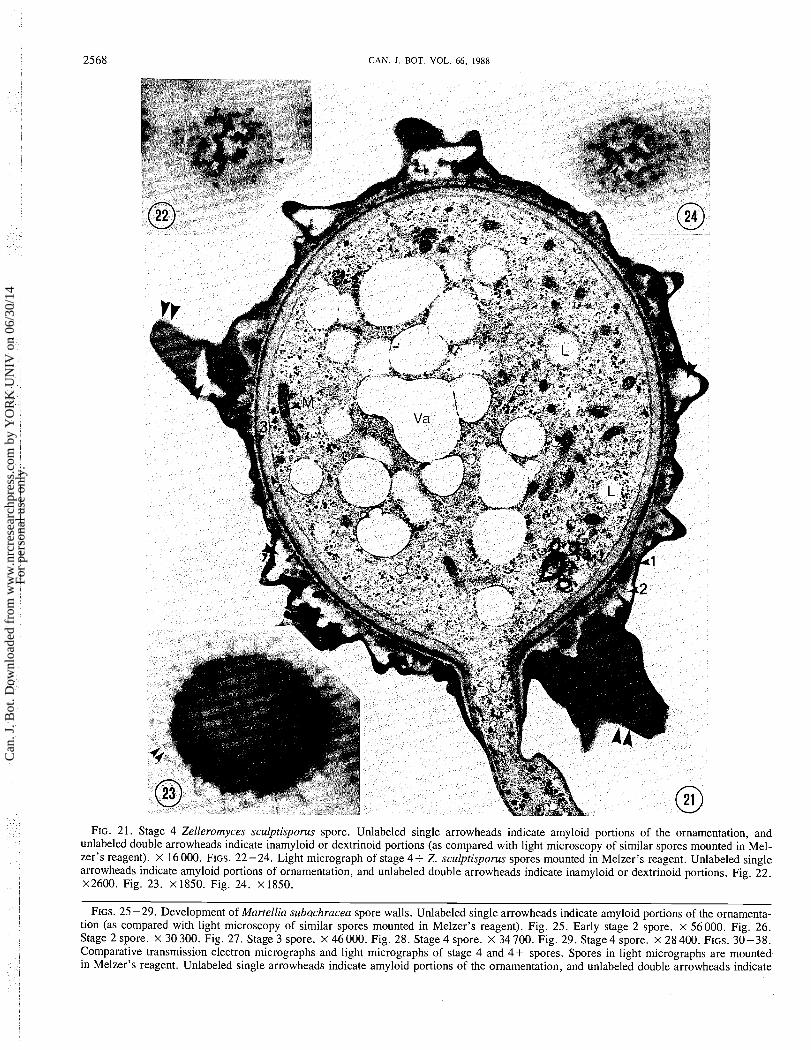
2568 CAN. J. BOT. VOL. 66, 1988
FIG. 21. Stage 4 Zelleromyces sculptisporus spore. Unlabeled single arrowheads indicate amyloid portions of the ornamentation, and unlabeled double arrowheads indicate inamyloid or dextrinoid portions (as compared with light microscopy of similar spores mounted in Mel- zer's reagent). x 16000. FIGS. 22-24. Light micrograph of stage 4+ Z. sculptisporus spores mounted in Melzer's reagent. Unlabeled single arrowheads indicate amyloid portions of ornamentation, and unlabeled double arrowheads indicate inamyloid or dextrinoid portions. Fig. 22. x2600. Fig. 23. x 1850. Fig. 24. x 1850.
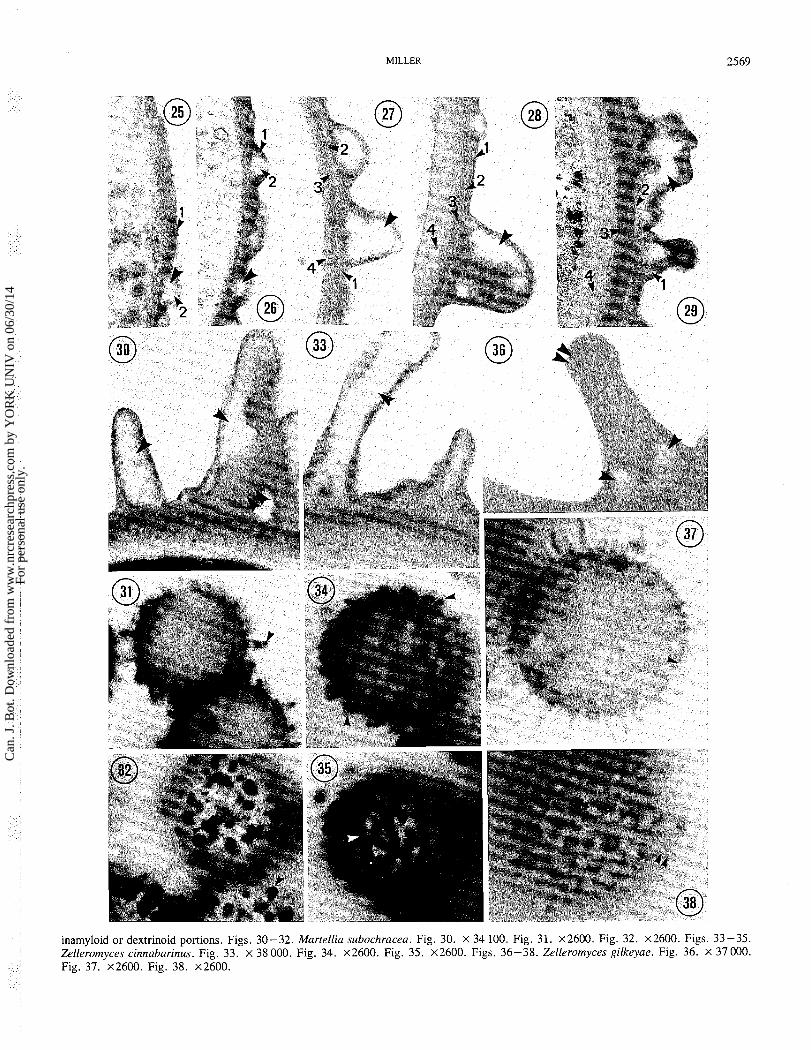
FIGS. 25 -29. Development of Martellia subachracea spore walls. Unlabeled single arrowheads indicate amyloid portions of the ornamenta- tion (as compared with light microscopy of similar spores mounted in Melzer's reagent). Fig. 25. Early stage 2 spore. x 56000. Fig. 26. Stage 2 spore. x 30 300. Fig. 27. Stage 3 spore. x 46 000. Fig. 28. Stage 4 spore. X 34 700. Fig. 29. Stage 4 spore. x 28 400. FIGS. 30-38. Comparative transmission electron micrographs and light micrographs of stage 4 and 4+ spores. Spores in light micrographs are mounted in Melzer's reagent. Unlabeled single arrowheads indicate amyloid portions of the ornamentation, and unlabeled double arrowheads indicate
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/3
0/14
For
pers
onal
use
onl
y.
MILLER 2569
inamyloid or dextrinoid portions. Figs. 30-32. Martellia subochracea. Fig. 30. X 34 100. Fig. 31. ~ 2 6 0 0 . Fig. 32. ~ 2 6 0 0 . Figs. 33-35. Zelleromyces cinnabarinus. Fig. 33. x 38000. Fig. 34. ~ 2 6 0 0 . Fig. 35. x2600. Figs. 36-38. Zelleromyces gilkeyae. Fig. 36. X 37000. Fig. 37. ~ 2 6 0 0 . Fig. 38. ~ 2 6 0 0 .
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/3
0/14
For
pers
onal
use
onl
y.
CAN. J . BOT. VOL. 66, 1988
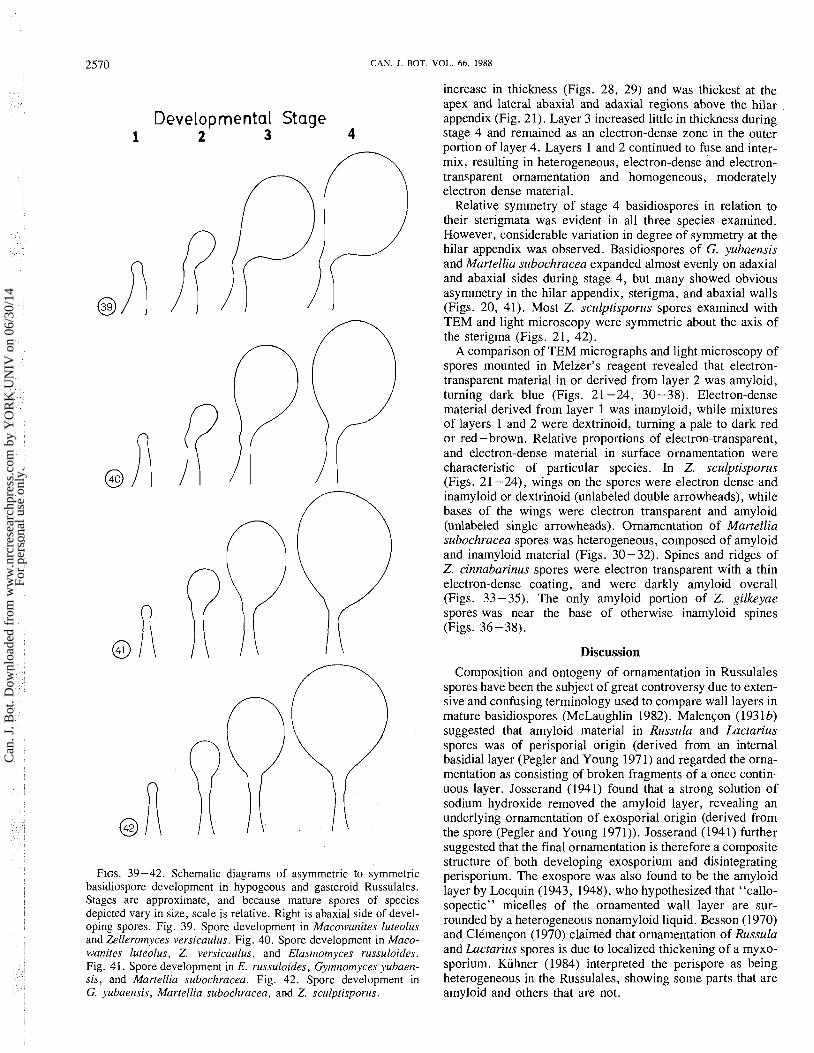
increase in thickness (Figs. 28, 29) and was thickest at the apex and lateral abaxial and adaxial regions above the hilar
Developmental Stage appendix (Fig. 21). Layer 3 increased little in thickness during
1 2 3 4 stage 4 and remained as an electron-dense zone in the outer portion of layer 4. Layers 1 and 2 continued to fuse and inter-
P ! mix, resulting in heterogeneous, electron-dense and electron- transparent ornamentation and homogeneous, moderately electron dense material.
Relative symmetry of stage 4 basidiospores in relation to their sterigmata was evident in all three species examined. However, considerable variation in degree of symmetry at the
A f hilar appendix was observed. Basidiospores of G. yubaensis and Martellia subochracea expanded almost evenly on adaxial and abaxial sides during stage 4, but many showed obvious asymmetry in the hilar appendix, sterigma, and abaxial walls
@ (Figs. 20, 41). Most Z. sculptisporus spores examined with TEM and light microscopy were symmetric about the axis of
PP the sterigma (Figs. 21, 42). A comparison of TEM micrographs and light microscopy of
spores mounted in Melzer's reagent revealed that electron- transparent material in or derived from layer 2 was amyloid, turning dark blue (Figs. 2 1 -24, 30 - 38). Electron-dense material derived from layer 1 was inamyloid, while mixtures
A R' of layers 1 and 2 were dextrinoid, turning a pale to dark red or red - brown. Relative proportions of electron-transparent, and electron-dense material in surface ornamentation were
@ characteristic of particular species. In Z. sculptisporus (Figs. 2 1 -24), wings on the spores were electron dense and inamyloid or dextrinoid (unlabeled double arrowheads), while bases of the wings were electron transparent and amyloid (unlabeled single arrowheads). Ornamentation of Martellia subochracea spores was heterogeneous, composed of amyloid and inamyloid material (Figs. 30-32). Spines and ridges of Z. cinnabarinus spores were electron transparent with a thin electron-dense coating, and were darkly amyloid overall (Figs. 33-35). The only amyloid portion of Z. gilkeyae spores was near the base of otherwise inamyloid spines (Figs. 36-38).
@ Discussion
FIGS. 39-42. Schematic diagrams of asymmetric to symmetric basidiospore development in hypogeous and gasteroid Russulales. Stages are approximate, and because mature spores of species depicted vary in size, scale is relative. Right is abaxial side of devel- oping spores. Fig. 39. Spore development in Macowanites luteolus and Zelleromyces versicaulus. Fig. 40. Spore development in Maco- wanites luteolus, Z. versicaulus, and Elasrnornyces russuloides. Fig. 41. Spore development in E. russuloides, Gyrnrzornyces yubaerz- sis, and Martellia subochracea. Fig. 42. Spore development in G. yubaensis, Martellia subochracea, and Z. sculptisporus.
Composition and ontogeny of ornamentation in Russulales spores have been the subject of great controversy due to exten- sive and confusing terminology used to compare wall layers in mature basidiospores (McLaughlin 1982). Malen~on (1931b) suggested that amyloid material in Russula and Lactarius spores was of perisporial origin (derived from an internal basidial layer (Pegler and Young 1971) and regarded the orna- mentation as consisting of broken fragments of a once contin- uous layer. Josserand (1941) found that a strong solution of sodium hydroxide removed the amyloid layer, revealing an underlying ornamentation of exosporial origin (derived from the spore (Pegler and Young 1971)). Josserand (194 1) further suggested that the final ornamentation is therefore a composite structure of both developing exosporium and disintegrating perisporium. The exospore was also found to be the amyloid layer by Locquin (1943, 1948), who hypothesized that "callo- sopectic" micelles of the ornamented wall layer are sur- rounded by a heterogeneous nonamyloid liquid. Besson (1970) and Clkmen~on (1970) claimed that ornamentation of Russula and Lactarius spores is due to localized thickening of a myxo- sporium. Kiihner (1984) interpreted the perispore as being heterogeneous in the Russulales, showing some parts that are amyloid and others that are not.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/3
0/14
For
pers
onal
use
onl
y.

MlLLER 257 1
Spore walls of hypogeous and gasteroid species of Rus- sulales examined in this study were composed of four layers and conformed in many respects to hypotheses forwarded by other workers regarding Russula and Lactarius spores. Layers 1 and 2 are best regarded as the perisporium or myxo- sporium of Besson-Antoine and Kiihner (1972). The exospore or tunica of ClCmen~on (1970) corresponds to layer 3, and layer 4 is the epispore or sclerosporium of Hugueney (1972). Layer 2 is amyloid in Melzer's reagent, usually forms a con- tinuous amyloid layer beginning at stage 2 or 3 in most species, and melds with layer 1 to form heterogeneous non- amyloid, electron-dense and amyloid, electron-transparent ornamentation. Burge (1966), using nonaldehyde fixatives and older TEM techniques, reported five layers in species of Rus- sula, Macowanites, Elasmomyces, and Martellia, while Burge (1979), using similar methodology, found up to six layers in Russula spores.
Early spore asymmetry in heterotropic basidiospores has been hypothesized to be under the control of the hilar appendix body (McLaughlin 1977, 1982; Yoon and McLaughlin 1979, 1984). While the composition and function of the hilar appen- dix body are unknown, McLaughlin (1977) suggested that it is involved in extracellular secretions and perhaps prevents vesicle deposition on the wall adjacent to the hilar appendix. The adaxial wall then becomes rigid, directing growth toward the abaxial side. Although there was some indication that the adaxial hilar appendix wall in Martellia luteolus, Z. versi- caulus, and E. russuloides was more distinct, and possibly more rigid, than the wall at the apex of the spore initial, no confirmation of the function of the hilar appendix body was made. However, the presence of a hilar appendix body in young asymmetric spores of Martellia luteolus, Z. versi- caulus, and E. russuloides and its absence in truly symmetric spores of Martellia subochracea and Z. sculptisporus are fur- ther evidence that the hilar appendix body is responsible for early asymmetric development. As reported previously for Lactarius lignyotellus (Miller 1988), the hilar appendix body in Martellia luteolus, Z. versicaulus, and E. russuloides was a poorly differentiated region of cytoplasm located in the hilar appendix adjacent to the adaxial wall. It was present in the basidiospore primordia before initiation of asymmetric growth and remained visible through stage 3. The hilar appendix body in examined species of Russulales appears to be less differen- tiated and less electron dense than that reported for Coprinus cinereus (Schaeff ex Fr.) S. F. Gray and Boletus rubinellus Peck (McLaughlin 1977, 1982; Yoon and McLaughlin 1979, 1984), but this may be due to differences in fixation. McLaughlin (1977) replorted that the hilar appendix body in C. cinereus was present during stage 4.
Establishing precise limits for each stage, as was accom- plished in the laboratory by McLaughlin (1977) and Yoon and McLaughlin (1979, 1984), was more difficult for the field- collected, nonsynchronous species in this study. However, four approximate stages of symmetric basidiospore formation in Martellia subochracea, G. yubaensis, and Z. sculptisporus could be established from differences in spore wall architec- ture, spore size and morphology, and organelle composition. The "asymmetric growth" phase, used by McLaughlin (1977) to describe stage 2 in heterotropic spore development, was absent or reduced in many Martellia subochracea, G. yubaensis, and Z. sculptisporus spores, and "elongation and symmetrical enlargement," "midenlargement," and "late enlargement" were substituted for stages 2-4, respectively.
Asymmetry observed in some G. yubaensis and Martellia subochracea spores was established early in development, and growth of the spores was approximately equal during later stages of development. Symmetric spores of Martellia sub- ochracea, G. yubaensis, and Z. versicaulus remained essenti- ally in a midenlargement or late enlargement phase throughout development.
Malen~on (193 la) described four developmental phases in asymmetric Russula and Lactarius spores and suggested that spore morphology in gasteroid Russulales represents varying degrees of disorganization or decline of basic Russula and Lactarius spore development. According to Malen~on's con- cept, strong asymmetric development in Macowanites luteolus would be like that in species of Russula; asymmetric develop- ment in many spores of Z. versicaulus, and E. russuloides, like Russula spore development that stops at the third phase; and symmetric development in Z. sculptisporus, like Russula and Lactarius spores that do not pass beyond the first or second phase. Differences in presence, time of appearance, location, and duration of a hilar appendix body or similar cyto- plasmic region in developing spores could prpduce effects on spore symmetry that corrobrate Malen~on's hypothesis. Although no hilar appendix body was observed in slightly asymmetric spores of G. yubaensis or Martellia subochracea, it is possible that it is present briefly during late stage 1 or early stage 2 and only infrequently encountered. Diversity in degree and location of asymmetry, relative number of asym- metric and symmetric spores, and number of species with both symmetric and asymmetric spores indicate that spore morphol- ogy varies along a continuum in the Russulales and suggest that heterotropy and orthotropy are results of varying degrees of the same process.
Variation in spore development and ornamentation in hypo- geous and gasteroid Russulales poses a taxonomic problem. Young spores frequently are amyloid, while older spores from the same basidiocarp may or may not be amyloid. The contin- uous amyloid layer is difficult to demonstrate with light microscopy when the hyaline or pale background color of spores gives the impression that only the ornamentation is amyloid. Ornamentation in young spores is usually not as well developed as in older spores and can look totally different, and many species have spores that are not amyloid at all, only dex- trinoid. Similarly, most species exhibit a range in spore size and symmetry in the same basidiocarp. Young or small spores are often slightly to strongly asymmetric, while mature or large spores are more symmetric.
Variation in spore symmetry has also led to difficulty in determining relationships between hypogeous, gasteroid, and agaricoid taxa within the Russulales. Classification proposed by Pegler and Young (1979, 1981) emphasized spore sym- metry and discharge as family and generic characteristics. Although exact mechanisms of forcible spore discharge have not been elucidated, Miller and Miller (1988) hypothesized that spore release and spore discharge are separate processes in the Russulales, and that a graduation in spore release mech- anisms exists in hypogeous, gasteroid, and agaricoid taxa. Likewise, the present study suggests that spore symmetry in the Russulales is expressed as a continuum, often in spores from the same basidiocarp, and that boundaries are difficult to define. In addition, Heim (1937~) described spores of tropical, lamellate, agaricoid Russula annulata Heim as "globose rather than ovoid, and with central rather than axial sym- metry," similar to symmetric spores of many hypogeous taxa.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/3
0/14
For
pers
onal
use
onl
y.

2572 CAN. J. BOT. VOL. 66, 1988
Following the Pegler and Young classification, R. annulata would not fit in the Russulaceae because of its symmetrical spores, nor in the Elasmomycetaceae because of its forcible spore discharge and agaricoid architecture. The continuum in spore symmetry in the Russulales suggests that taxa with sym- metric and asymmetric spores may be more closely related than is implied by current classification and that circumscrip- tion of genera and families with symmetric versus asymmetric spores is artificial.
Acknowledgements This paper is a portion of a dissertation submitted to the
Graduate School of Virginia Polytechnic Institute and State University, Blacksburg, V A 24061, U.S.A., in partial fulfill- ment of the requirements for the degree of Doctor of Philos- ophy. Special thanks a re extended to Professor Orson K. Miller for serving as an advisor. Thanks a re also extended to Dr. David Stetler and Carol Gassman for their advice and support in electron microscopy, to Bob Fogel for use of a dia- mond knife, and to Herb Saylor and Jim Trappe for sending fixed material. This study was funded in part by National Science Foundation grant BSR-8505972 and the USDA Forest Service, Pacific Northwest Research Station, Corvallis, O R 97331, U.S.A.
BEATON, G., PEGLER, D. N., and YOUNG, T. W. K. 1984. Gasteroid Basidiomycota of Victoria State, Australia 2. Russulales. Kew Bull. 39: 669-698.
BESSON, M. 1970. Ultrastructure de la paroi sporique amyloid et ornCe de quelques HymCnomycetes. C.R. Hebd. Seances Acad. Sci. Ser. D, 271: 964-967.
BESSON-ANTOINE, M., and KUHNER, R. 1972. Les couches de la paroi sporique des RhodophyllacCes (Agaricales). C.R. Hebd. Seances Acad. Sci. Ser. D, 275: 1489 - 1494.
BURGE, H. A. P. 1966. The structure and development of astero- spores. Ph.D. thesis, University of Michigan, Ann Arbor, MI.
1979. Basidiospore structure and development in the genus Russula. Mycologia, 71: 977 -995.
CLEMENCON, H. 1970. Bau der Wande der Basidiosporen und ein Vorschlag zur benennung ihrer Schichten. Z. Pilzkd. 36: 112- 133.
DONK, M. A. 1971. Progress in the study of the classification of the higher Basidiomycetes. In Evolution in the higher Basidiomycetes. Edited by R. H. Petersen. University of Tennessee Press, Knox- ville, TN. pp. 3-25.
HEIM, R. 1931. Les leins phylktiques entre les Agarics ochrosporCs et certains GastCromycktes. C.R. Hebd. Seances Acad. Sci. Ser. D, 192: 291 -293.
1936a. Sur la parent6 entre Lactaires et certains GastCromy- cktes. C.R. Hebd. Seances Acad. Sci. Ser. D, 202: 2101-2103.
1936b. Sur la phylogknie des Lactario-RussulCs. C.R. Hebd. Seances Acad. Sci. Ser. D, 203: 108 - 1 10.
1937a. Observations sur la flore mycologiques malagache. 111-IV. Les Lactario-RussulCs h anneau : ontoghie et phylogCnie. Rev. Mycol. 2: 61 -75.
1937b. Observations sur la flore mycologiques malagache. V-VI. Les Lactario-RussulCs h anneau : ontoghie et phylogCnie. Rev. Mycol. 2: 109- 117.
1938. Prodrome h une flore mycologique de Madagascar et dipendances. I. Les Lactario-RussulCs du domaine oriental de Madagascar. Essai sur la classification et la phylogCnie des AstCro- sporales. Laboratoire de Cryptogamie du Museum National d'His- toire Naturelle, Paris. pp. 1 - 196.
1943. Remarques sur les formes primitives ou dCgradCs de Lactario-RussulCs tropicaux. Boissiera, 7: 266-280.
1948. Phylogeny and natural classification of macro-fungi.
Trans. Br. Mycol. Soc. 30: 161 - 178. 1971. The inter-relationships between the Agaricales and
Gasteromycetales. In Evolution in the higher Basidiornycetes. Edited by R. H. Petersen. University of Tennessee Press, Knox- ville, TN. pp. 505-534.
HOLMGREN, P. K., and KEUKEN, W. 1974. Index Herbariorum. Reg- num Veg. 92: 1-397.
HUGUENEY, R. 1972. Ontogenkse des infrastructures de la paroi spo- rique de Coprinus cineratus QuC1. var. nudisporus Kiihner (Agari- cales). C.R. Hebd. Seances Acad. Sci. Ser. D, 275: 1495 - 1498.
JOSSERAND, M. 1941. Etude su l'ornamentation sporique des Lac- taires et de quelques autres espkces h spores amyloi'des. Bull. Soc. Mycol. Fr. 56: 738.
KUHNER, R. 1984. Some mainlines of classification in the gill fungi. Mycologia, 76: 1059 - 1074.
LOCQUIN, M. 1943. La disposition des micelles callospectiques dans les membranes des spores de quelques Basidiomycktes. Bull. Soc. Mycol. Fr. 59: 44-51.
1948. Morphologie et structure des spores de Russula. Rev. Mycol. 13: 10-29.
MALENCON, G. 193 la . La sCrie des AstCrosporks. Recueil de travaux cryptogamiques dCdiCs h Louis Mangin. Laboratoire de Crypto- gamie du Museum National d'Histoire Naturelle, Paris. pp. 377 - 396.
1931b. Considerations sur les spores des RussulCs et Lac- taires. Bull. Soc. Mycol. Fr. 47: 72-86.
MCLAUGHLIN, D. J. 1977. Basidiospore initiation and early develop- ment in Coprinus cinereus. Am. J. Bot. 65: 1 - 16.
1982. Ultrastructure and cytochemistry of basidial and basid- iospore development. In Basidium and basidiocarp, evolution, cytology, function, and development. Edited by K. Wells and E. K. Wells. Springer-Verlag, New York. pp. 37-74.
MELZER, V. 1924. L'ornamentation des spores des RussulCs. Bull. Soc. Mycol. Fr. 40: 78-81.
MILLER, S. L. 1988. Early spore formation in Lactarius lignyotellus. Mycologia, 80: 99 - 107.
MILLER, S. L., and MILLER, 0 . K. 1986. Zelleromyces stephensii, an interesting member of the gasteroid Russulales from Europe. Mycol. Helvetica, 2: 59-66.
1988. Spore release in hypogeous, gasteroid and agaricoid Russulales. Trans. Br. Mycol. Soc. 90: 513-526.
PEGLER, D. N., and YOUNG, T. W. K. 1971. Basidiospore morphology in the Agaricales. Nova Hedwigia, 35: 1-210.
1979. The gasteroid Russulales. Trans. Br. Mycol. Soc. 72: 353 -388.
1981. World pollen and spore flora. 10. Russulaceae Roze. Edited by S. Nilsson. Almquist and Wiksell Periodical Co., Stock- holm. pp. 1-35.
SHEARER, T. P., and HUNSICKER, L. C. 1980. A rapid method for embedding tissues for electron microscopy using 1,4-dioxane and polylbed 812. J. Histochem. Cytochem. 28: 465-467.
SINGER, R. 1938. Contribution i 1'Ctude des RussulCs. 111. Quelques RussulCs amkricaines et asiatiques. Bull. Soc. Mycol. Fr. 54: 132- 177.
1971. A revision of the genus Melanomphalia as a basis of the phylogeny of the Crepidotaceae. In Evolution in the higher Basidiomycetes. Edited by R. H. Petersen. University of Tennessee Press, Knoxville, TN. pp. 441 -480.
1975. The Agaricales in modern taxonomy. 3rd ed. J. Cramer, Vaduz.
SINGER, R., and SMITH, A. H. 1960. Studies on secotiaceous fungi. IX. The astrogastraceous series. Mem. Torrey Bot. Club, 21: 1 - 122.
SMITH, A. H. 1971. The origin and evolution of the Agaricales. In Evolution in the higher Basidiomycetes. Edited by R. H. Petersen. University of Tennessee Press, Knoxville, TN. pp. 481 -504.
THIERS, H. D. 1971. Some ideas concerning the phylogeny and evolu- tion of the boletes. In Evolution in the higher Basidiomycetes. Edited by R. H. Petersen. University of Tennessee Press, Knox-
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/3
0/14
For
pers
onal
use
onl
y.

MILLER 2573
ville, TN. pp. 423-440. appendix in basidiospores of Boletus rubinellus. Am. J . Bot. 66: 1984. The secotioid syndrome. Presidential address to the 870 - 873.
Mycological Society of America. Mycologia, 76: 1-8. 1984. Basidiosporogenesis in Boletus rubinellus. I. Sterigmal YOON, K. S., and MCLAUGHLIN, D. J. 1979. Formation of the hilar initiation and early spore development. Am. J. Bot. 71: 80-90.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y Y
OR
K U
NIV
on
06/3
0/14
For
pers
onal
use
onl
y.






















