Embed Size (px)
Citation preview

A STUDY OF THE LOACHES OF THE GENERA COBITIS AND SABANEJEWIA (PISCES, COBITIDAE) OF GREECE,
WITH DESCRIPTION OF SIX NEW TAXA
Trav. Mus. nad. Hist. nat. (~Grigore Antipan
PANOS S. ECONOMIDIS, TEODOR T. NALBANT
Les CobitidCs de Grtce des genres Cobitis et Sabanejewia ont au total neuf esptces, dont huit appartiennent au premier genre et une au second. Quatre espkces nouvelles de Cobitis sont dCcrites (stephanidisi, hellenica, arachthoser~sis et punctilineata), ainsi que deux sous-esptces nouvelles de Sabanejevia arrrata (doiranica et thrakica). L'article contient aussi une ample discution sur I'origine de la faune gr&que de CobitidCs.
Vol. XXXVI
INTRODUCTION
The systematics and phylogeny of the loaches of the Balkan Peninsula, which belong to the genera Cobitis and Sabanejewia, represent one the most complex and interesting of problems. Therefore, the main aim of this contribution is to clarify, as accurately as possible, the position of each species. This seems to be a difficult task, especially because Karaman (1924, 1928, 1929, 1955) has described many taxa of Cobitis very briefly and without figures, excepting ohridana and strumicae. Our difficulties consisted in ascribing our specimens to one or to another of Karaman's taxa. Only for strumicae (see Karaman, 1955: 190-192, Fig. 4) did he gave more details and quite a good figure but no information about the structure of the lamina circularis (Canestrini's scale), an extremely important feature for defining Cobitis species. On the other hand, neither the book of Vukovic & Ivanovic (1971), dealing with the freshwater fishes of Yugoslavia, nor the contribution of Grupce & Dimovski (1976), dedicated to the genus Cobitis in the southern part of this country, give any solution in this problem. Moreover, there is no any mention of Karaman's type material.
As far the situation of Cobitis and Sabanejewia in Greece is concerned, the information is very poor. Stephanidis (1939) refers to Cobitis taenia (now Cobitis hellenica sp. n.) from, the river Louros (western Greece) and more recently (Stephanidis, 1974) he gives a compiete
pp. 295-347 1996

296 P.S. ECONOMIDIS. T.T. NALBANT
description of a new species, Cobitis (Bicanestrinia) trichonica, from lakes Trichonis, Lyssimachia and Ozeros of the western Greek mainland. On the other hand Economidis (1974) noted all morphological variability of various populations of Cobitis taenia (now Cobitis stumicae ) and Cobitis aurata (= Sabanejewia aurata) in their area of distribution, i.e. the aquatic system from the river Evros to the river Strymon in the Greek parts of Thrace and eastern Macedonia. Later, Economidis et al. (1981) and Economidis & Voyadjis (1985) listed Cobitis taenia (Cobitis vardarensis) and Sabanejewia aurata from the Aliakmon and Axios and Gallikos rivers respectively, and Economidis & Sinis (1982) gave a short description of Cobitis taenia (now Cobitis strumicae) from lakes Koronia and Volvi system. Therefore, another aim of this paper is to present a synthesis of all species of Cobitis and Sabanejewia recorded from Greece, on basis of a great number of samples collected at many localities in the country. Besides this, a rich collection of loaches in the Institute of Biology of Bucharest (Romania) is used (see below) as comparative material. Six (4 species and 2 subspecies) taxa are here described as new from different basins of Greece. For more information about the aquatic drainge of Greece see Fig. 1, in which are also presented all Balkan drainges south of the river Danube.
Cobitis ohridana Karaman, 1928, may be present also in the Greek freshwater catchment, since some Cobitis specimens from river Aoos near Konitsa, agree quite well with Karaman's figure (1928: 163, Fig. 5). However, the original description of ohridana is extremely brief and cannot be used for its identification. Therefore, based on our observation of specimens from river Aoos, which have a single lamina circularis, we refer ohridana rather to Cobitis s. str. than to Bicanestrinia.
MATERIAL AND METHODS
Collections. All specimens collected in Greece are mentioned under "material" for each species. Paratypes of the new taxa are also deposited to the following Museums: BMNH, CAS, ISBB, MINB, MNHN, MZUT, NMW, SMF, USNM, ZSM (see abbreviations).
Compara five material. We also used much material from different institutions as follow: ISBB, MINB, MSNT, NMC, RMNH, SMF, ZMB.
In order to understand the value of each charactrer and to discern the relationships between the species, we have used as comparative material the following species (measurements represent the standard lenght in milimeters):
Cobits taenia Linnaeus, 1758: RMNH. AAA-1900999.4949, (7) 55.5-60.8 Sweden, no other indications; RMNH. SBM-1951 365 5031, (1) 80.0, Sweden, Tumba, Uttran; RMNH, AAA-1975999. .4959, (1) 76.0, Sweden without other data; RMNH. AAA-1977999.4958, (2) 52.7 and 61.0,

COBITIDS OF GREECE 297
Sweden, without other data; RMNH. A86-1986213.3605, (1) 47.8 Sweden, Varmdon, Sagsjon; ISBB 2751 (9) 29.7-81.2, Holland (The Netherlands) without other data.
Cobitis danubialis Biicescu, 1993: ISBB uncat. (12) 63.3-96.3, Romania, Transilvania, river Murev, Stinceni, 1991.
Cobitis megaspila Nalbant, 1993: ISBB uncat. (6) 54.6-74.5, Romania, Danube Delta, Caraorman, paratypes.
Cobitis maroccana Pellegrin, 1929: ISBB uncat., (3) 47.0-49.2, Portugal, Tajo drainage, Rio Latas at Coruche; ISBB 3981, (5) 48.3-76.2, Spain, Guadiana drainage, Rio Quejigares, Fontanosas near Ciudad Real. SMF 3231, (2) 65.0 and 66.0, Spain, Silla at Valencia, Aqua Comunero at 1.5 km from Albufera, paratypes of Cobitis haasi Klausewitz, 1955.
Cobitis bilineata Canestrini, 1866: ISBB 3089, (9) 51.3-72.0, Italy, Po drainage, 'Torente Meletta near Carmagnola; ISBB 3090, (15) 54.3-84.0, Italy, Po drainage, Canale Moneta at Carmagnola.
Cobitis narentana Karaman, 1929: ISBB 4577 (3) 50.0-51.0, river Neretva, Croatia, no other data.
Cobitis zanandreai Caviccioli, 1965; ISBB uncat., (1) 63.0, Italy, Lago di Fondi near the Thyrenian Sea, no other data.
Cobitis turcica Hanko, 1925: ISBB uncat., (1) 63.0, Turkey, Anatolia, Iqikli golu; ISBB uncat. (3) 42.3-91.0, Turkey, Kizil Dere, Bagilli-Koyu, 1993; MZUT uncat. (1) 54.0, bearing same data as previous specimens.
Cobitis simplicispina Hanko, 1925; ISBB 3566, (2) 55.2 and 76,2, Turkey, Anatolia, Aci golu, Karahiiyuk; MINB uncat., (1) 66.3, Turkey, Golhisar, 1957.
Cobitis elazigensis Coad & Sarieiyupoglu, 1988: NMC uncat., (1) 108.0, Turkey, Anatolia, Elazig stream near Elazig.
Cobitis linea Heckel, 1840: ISBB uncat., (3) 51.3-84.0, Iran, river Kor near Persepolis, 1800 m altitude, 1979.
Cobitis elongata Heckel & Kner, 1858, ISBB uncat., (I) 146.0. Romania, south Banat, river Nera at Sasca Montana.
Cobitis calderoni Biicescu, 1962: ISBB 2744, (2) 54.0 and 64.2, Spain, Zamora province, Lago de Sandin de Carbaleda.
Cobitis bilseli Battalgil, 1942: MSNT 3948, (1) 195.0, Turkey, Anatolia, Beyshehir golu, Beyshehir; ISBB uncat. (1) 155.0, Beyshehir golu, Beyshehir.
Cobitis macrostigma Dabry, 1872: ISBB uncat., (2) 130.5 and 135.0, China, Hupeh, lake Liang Tse-hu, Yangste drainage.
Cobitis melanoleuca Nichols, 1925: ISBB 3230, (4) 48.6-54.0. China, Shansi, Kwei-hwa, Third Asiatic Expedition; ISBB 2290, (9) 37.0-57.0, Mongolia, Ugij nor, river Nargin Gol, Selenga drainage, Arhangaj Aimak, paratypes of Cobitisgranoei olivai Nalbant, Holcik & Pivnicka, 1970.

298 P.S. ECONOMIDIS. T.T. NALBANT
Cobitis dolichorhynchus Nichols, 1918: ISBB uncat., (1) 79.9, China, Yangtse basin, no other data.
Cobitis sinensis Sauvage & Dabry, 1874: ISBB uncat., (4) 54.3-67.0, South Korea,river Yung San.
Cobitis lutheri Rendahl, 1935: ISBB uncat., (4) 47.4-64.8, South Korea, river Yung San.
Cobitis biwae Jordan and Snyder, 1901: ISBB uncat., (4) 82.2-95.4, Japan, Honsu, river Kamo at Kyoto.
Cobitis striata Okada, 1936: ISBB uncat., (5) 56.7-92.4, Japan, Honsu, Okayama-ken, river Yoshi.
Iksookimia koreensis (Kim, 1976): ISBB uncat., (5) 70.2-80.3, South Korea, Gyonggy-do, river South Han.
Iksookimia longicorpus (Kim, Choi and Nalbant, 1976): ISBB uncat., (4) 92.0-119.0, South Korea, river Seom Jin.
Iksookimia hugowolfeldi Nalbant, 1993: ISBB 4495 (1) holotype 64.5 and ISBB 4496 (2) paratypes 75-93.6, South Korea, river Yung San.
Definitions. In defining a species of the family Cobitidae besides meristic and metric values it very important to take into consideration the following characters.
Scales: shape, size and extent over the body. Rendahl (1943, 1944, 1945, 1948) gave special attention to the shape and size of subdorsal scales. These scales are situated halfway between the dorsal fin and the lateral line or lateral midline of the body, and were used by him for measurements. The shape of these scales is also characteristic for each species. Therefore every mention in the present contribution (or drawing of a scale) refers only to the subdorsal scales.
Colour pattern. In Cobitis species there is a peculiar arrangement of longitudinal rows of spots or bands, already pointed out by Gambetta (1934), and called Gambetta's zones of pigmentation (see Fig. 3 a). However, we found at least six main arrangements and shape of these rows as variations of the initial type (Fig. 3 a-f). On the other hand, at the upper part of base of caudal fin there is a black spot which in different species can be faded to complete absence or, in a few cases, there can be two or even three such spots at the base of caudal. In the closely related genera Sabanejewia and Iksookimia (Nalbant, 1993, 1994) this arrangement of spots is lacking. However, one or two rows of small spots (particularly a more of less lengthy band of vermiculate speckles) are present between the dorsal row and lateral row of large spots.No jet black spot at the upper part of base of caudal is present in Sabanejewia but always there are two dark gray spots sometimes united, at the base of the caudal fin.
The pigmentation of the head is also important. In Cobitinae there is always a dark gray, sometimes blackish, band from the anterior rim of the

COBITIDS OF GREECE 299
orbit to the tip of snout (Fig. 2a, b, c), but in different species (see Fig. 2 a, b) other dark gray bands, posterior or inferior the eye, may be present.
Sexual dimorphism. When present in Cobitinae loaches, this is one of the most important characters. The genus Cobitis has peculiar sexual dimorphism: most adult males have in pectoral fins at the base of second ray, sometimes in both first and second rays, a special plate-like process, so called lamina circularis or Canestrini's scale. We follow Rendahl (1930, 1933 a, 1933 b, 1943, 1944, 1945, 1948) in calling this process the lamina circularis instead of Canestrini's scale since the latter term may be confused with a body scale. Although the shape of this process is very variable, in most cases it is round, and we can thus utilize the term for defining this thin osseous plate-like process. This process is characteristic in shape and size for different species (Fig. 4 a, b, c). In the adult males of Sabanejewia this kind of sexual dimporphism is lacking but the males have on the both sides of body two swellings, the larger being before of the dorsal fin and the smaller, behind this fin.
Suborbital spine. This is the movable lateral ethmoid bone which in nearly all Botiinae and Cobitinae is placed on the each side of the head. The different parts of the suborbital spines are treated by Nalbant (1963). Generally the shape and size of the suborbital are characteristic in many species.
Barbels and mental lobes. The structure of the lip in Cobitinae is one of the most important characters in defining species. Rendahl(1933 a, 1944, 1945, 1948) described in detail for each species the structure of mouth and the arrangments of barbels and mental lobes around it. We follow him in this way. Different parts of the mouth structure in Cobitinae were already discussed by Nalbant (1963) and BHnZirescu & Nalbant (1964).
Lateral line. In species of Cobitinae this can be very reduced to a small portion above the pectoral fin or can be completely absent; only in the genus Somileptes from India is the lateral line complete to the base of caudal fin. However, pairs of neuromast pits are present along the sides of the body (Zanandrea et al., 1965).
For terminology of different morphological aspects (mouth and barbels, scales, color pattern, suborbital spine, gas-bladder capsule) see Nalbant (1963).
Counts and measurements. These were made according to BgnZirescu et al. (1972), by the same person in order to avoid individual differences in estimates. All measurements are expressed in percentage of standard length and presented in the tables.
Drawings and maps. Those of species and anatomical parts were made by TTN from whole specimen or after dissections, while the maps were designed by PSE in a Macintosh PC using the Canvas 3.0 program.

300 P.S. ECONOMIDIS. T.T. NALBANT
KEY TO THE SPECIES OF COBITINAE OF GREECE
l(6) Pigmentation of body not disposed in four Gambetta's zones. Caudal fin with 12 branched rays. Sexual dimorphism: adult males without lamina circularis at base of first pectoral rays but with a conspicuous swelling on each side of body before dorsal fin (genus Sabanejewia) . .............................................................................................................. 2
2(5) Lateral spots generaly large and square-like. Only in rivers ................ 3 3(4) 10-18 lateral spots, mostly 13-14 .........................................................
...................................................... Sabanejewia aurata thrakica ssp. nov. 4(3) 7-14 lateral spots, mostly 10-11 ............. Sabanejewia aurata balcanica 5(2) Lateral spots mostly small and round. Only in lake (Doirani) ...............
...................................................... Sabanejewia aurata doiranica ssp. nov.
6(1) Pigmentation of body disposed in four Gambetta's zones. Caudal fin with 14 branched rays. Sexual dimorphism: adult males with lamina circularis at base of first two pectoral ray or only at second one (genus Cobitis) .......................................................................................... 7
7(10) Adult males with one plate-like lamina circularis at base of second pectoral ray (subgenus Cobitis s. str.) Fig. 4c ........................................ 8
8(9) The spots of the fourth Gambetta's zone well separated (as in Fig. 3 a) .................................................................................. Cobitis vardarensis
9(8) The spots of the fourth Gambetta's zone as a continuous band, rarely interrupted (as in Fig. 3 b, c and 6 a) ...... Cobitis stephanidisi sp. nov.
lO(7) Adult males with two plate-like lamina circularis at base of first and second pectoral rays (subgenus Bicanestrinia) Fig. 4 d ...................... 11
ll(12) Body relatively elongate ...................................................................... 17 12(11) Body high and stout ........................................................................... 13 13(16) Lateral spots of fourth Gambetta's zone generally normal in size,
somewhat square-like and not disposed on a black stripe (as in Fig. 3 f) .................................................................................................... 14
14(15) In young specimens the lateral spots of fourth Gambetta's zone are rare or very rare remote enough one another (as in Fig. 12 a -f) ....... ............................................................. Cobitis arachthosensis sp. nov.
15(14) In young specimens the lateral spots of fourth Gambetta's zone are normal in number close enough one another (as in Fig. 10 a, b and 12 g-i) ............................................................. Cobitis hellenica sp. nov.
16(13) Lateral spots of fourth Gambetta's zone reduced in size, rounded, generally contiguous or very close to one another and on a black stripe. (as in Fig. 11 a and 12 j-1) ........... Cobitis punctilineata sp. nov.
17(18) The general pigmentation of body is cloudy, the spots being not well delimited (as in Fig. 8 a) ........................................ Cobitis meridionalis
18(17) The general pigmentation of body and spots are well defined .... 19

COBITIDS OF GREECE 301
19(20) Lateral spots of fourth Gambetta's zone very large and well individualized, on the caudal peduncle being mostly vertical (as in Fig. 9 a) .................................................................. Cobifis trjchonica
20(19) Lateral spots of fourth Gambetta's zone normal or reduced in size, sometimes, close together, having same shape on the whole length of body (as in Fig. 7 a, b) ............................................ Cobitis sfrumicae
RESULTS
In the systematic account all species are presented as follows:
Genus COBITIS Linnaeus, 1758
Subgenus Cobitis s. str.
Cobitis vardarensis Karaman, 1928
(Fig. 5 a-g) Cobitis taenia. Apostolidis. 1892 (partim): 27 (description: an arched black spot in
the base of the upper part of caudal fin: distribution: in many rivers and streams of Thessaly). 1907: 23 (Thessaly): Stephanidis. 1950: 207 (river Pinios in Thessaly), Dimovski & Grupce, 1971: 27 (river Axios drainage: tributary BrCgalnitsa): Economidis, 1973 (partim): 471 (from literature: lake Doirani), 559 (from collections: lake Doirani and rivers & streams Axios, Gallikos, Anthemous): Economidis, Kattoulas & Stephanidis, 1981: 93 (list, lower part of rivers Aliakmon. Edesseos and Itamos): Economidis & Voyadjis, 1985: (list Axios, Doirani, Gallikos).
Cobitis taenia taenia. Karaman, 1924 (partim): 74 (river Axios near Skopje, lake Doirani).
Cobitis taenia vardarensis Karaman. 1928: 162. 174 (short description as new subspecies from river Axios or Vardar): 1929: 173 (short description again); Taler, 1953 (partim): 447 (river Axios or Vardar, lake Doirani): Grupce & Dimovski, 1973: 89 (river Axios system); 1976: 32 (characters, comparisons with other species of South Yugoslavia).
Material: MZUT 1971-123 (28) 24.5-72.8, river Axios catchment, channels near Koufalia, Macedonia, Greece, 16 July 1971, Economidis coll.; MZUT 1971-131 (7) 53.5-76.6, Gallikos stream, ditch near village Mandres Kilkis, Macedonia, Greece, 27 July 1971, Economidis coll.; MZUT 1971-160 (10) 27.5-60.7, Gallikos stream near village Argyroupoli Kilkis, Macedonia, Greece, 29 July 1971, Economidis coll.; MZUT 1977-308 (7) 51.5-78.0, Gallikos stream, near village Petroto Kilkis, Macedonia, Greece, 15 November 1977, Economidis coll.; MZUT 1977-139 (19) 38.8-70.4, river Loudias catchment, near Skydra, Macedonia, Greece, 21 September 1977, Economidis coll.; ISBB uncat., (15) 43.0-69.0, bearing same data as previous; MZUT 1987-55 (13) 30.6-72.1, river Loudias catchment, near village Exaplatanos Aridea, Macedonia, Greece, 10 November 1987, Economidis coll.; MZUT 1971-383 (8) 40.2-82.1, river Aliakmon, down bridge of old national road Thessaloniki-Katerini, Macedonia, Greece,

302 P.S. ECONOMIDIS. T.T. NALBANT
22 September 1971, Stephanidis coll.; MZUT 1986-20 (3) 70.4-87.6, river Pinios catchment, channels near Hasabali, Thessaly, Greece, 10 August 1986, Economidis coll.; MZUT 1989-81 (5) 77.6-83.8, river Pinos catchment, tributary Lithaios within the town Trikala, Thessaly, Greece, 18 October 1989, Economidis and Banarescu coll.; ISBB 4575 (10) 53.4-90.5, bearing same data as previous; ISBB uncat., (12) 40.2-76.3, river Pinios catchment, tr ibutary Xerias at Mati Tyrnavou, Thessaly, Greece, 15 November 1977, Economidis coll.; ISBB uncat., (2) 78.0-78.3, river Axios catchment, at village Gynekokastro Kilkis, Macedonia, Greece, 8 November 1977, Economidis coll.; ISBB 4573, (15) 39.0-75.4, river Loudias drainage, at Aridea, Macedonia, Greece, 11 November 1987, Economidis coll.
Karaman (1928) gave only a short description of the species, and thus a more detailed one needs to be provided here.
Description: Fin ray formula. Generally the number of fin rays is constant, with D I11 7, A 11-111 5, V I 6-1 6, P I 9-1 9, Cn 7 + 7 n, but in a few individuals we found 5 branched rays in both ventrals, 8 branched rays in both pectorals and 8 + 7 or 7 + 6 branched rays in caudal.
Morphology. The body is moderately elongate as in Cobitis taenia with a short caudaLpeduncle (Fig. 5 a, b, c). A very reduced keel above and below of the peduncle, near the base of caudal fin. Ventrals are insered under the second or third dorsal branched rays but in a few specimens the insertion of ventrals is on the same vertical line as the dorsal origin. The head is relatively short and stout, with a blunt snout.
All three pairs of barbels are short. The mental lobes (Fig. 5 d) have folded zone well developed but the median lobes are relatively reduced.
The suborbital spine (Fig. 5 f) is generally straight, with laterocaudal process reduced or very reduced. A relatively small spine is observed in the largest specimens from the rivers Loudias, Gallikos and Pinios.
The scale (Fig. 5 e) are generally circular, wider at base with a small eccentric focal zone. They are embedded and cover the whole body except the head.
Sexual dimorphism: In males there is a plate-like lamina circularis at the base of the second pectoral ray (Fig. 5). In males from the rivers Pinios and Gallikos, the distal margin of the lamina circularis is more rounded than in the males from river Loudias, in which this'process is somewhat more acute.
The colourpattern in life is very similar to that of Cobitis taenia. The whole body is yellowish with olive gray spots and dots. The arrangement and shape of spots and dots is similar in all population from Pinios, Gallikos, Axios and Loudias (Fig. 5a, b, c). The head is pigmented with small speckles. from the antrior part of eye to the tip of the snout a dark stripe. Two other stripes run from the near part of the orbit to the middle of

COBITIDS OF GREECE 303
the opercular margin and another obliquely below the eye down towards the mouth. The median row of dorsal spots is prsent in all specimens.These spots are large enough before the dorsal fin and large and square-like in the post-dorsal space. All four Gambetta's pigmentary zones are well developed. The median lateral row of spots (Gambetta's fourth zone) is formed by a large number of round spots (13-24). The iris of the eye is golden-yellow.
A jet black spot in the superior part of caudal base is normally well marked in all populations but may be lacking in a few individuals.
Dorsal and caudal fins with 4-5 rows of dots, the rest of the fin being pale.
The intramuscular septum along the lateral midline is visible as a longitudinal black stripe.
In all preserved specimens (ethanol 70%) the ground colour becomes bright yellowish-brown or yellowish-white with gray or grayish-brown spots.
Habitat: The species is typically a still water fish, preferring better death branch of river with more or less deep water (more than 50-100 cm) where it burrows into a muddy or silty bottom, especially among the roots of plants. However, it also can be observed in shallows and, rarely, in slowly running water.
Range (Fig. 17-11): The area of this species is confined to the following water systems: Pinios (Thessaly), lower Aliakmon, Loudias, Axios and Gallikos (central Macedonia).
Remarks: This species differs enough from Cobitis taenia in its combination of characters: a relatively high body, numerous and small lateral spots and by the presence of a large jet black spot at superior base of caudal fin.
Cobitis stephanidisi sp. nov.
(Fig. 6 a -e)
Cobitis taenia, Apostolidis, 1892 (partim): 27 (description: an arched black spot in the base of the upper part of the caudal fin, and a series of 12 to 18 black along the side of the body; distribution: karstic spring Kefalovriso in Velestino village): Panagiotopoulos, 1916: 576 (list: lake Karla).
Material: Holotype: MZUT 1992-40, 56.9, lake Karla catchment, spring within Velestino village, Thessaly, Greece, 27 August 1992, Economidis coll.
Paratypes: MZUT 1988-30 (9) 44.8-83.5, lake Karla catchment, spring within the village Velestino, Thessaly, Greece, 14 April 1988, Economidis coll.; MZUT 1989-113 (56) 22.0-71.9, same locality as previous, 2 May 1989, Economidis coll.; MZUT 1989-35 (26) 28.1-83.0, same locality as previous, 27 August 1989, Economidis coll.; MZUT

304 P.S. ECONOMIDIS. T.T. NALBANT
1992-37 (23) 51.9-85.0, same locality as previous, 27 August 1992, Economidis coll.; USNM 330137 (I), same locality as previous, 27 August 1992, Economidis coll.; SMF 27234 (4), bearing same data as previous; MNHN 1994-516 (2), bearing same data as previous; NMW 92822 (1 & 2), bearing same dataas previous.
Diagnosis: A long bodied species of Cobitis with a continuous black or dark gray, fourth Gambetta's zone, along the lateral midline of the body, a conspicuous jet black dot at the upper base of the caudal fin.
Description: Fin ray formula. D 111 7, A 11-111 5, V I 6-1 6, P I 9-1 9, Cn 7 + 7 n. There is some variation in the number of dorsal rays, in few specimens there are 6 branched rays and in a single individual 5 branched rays. In the caudal, fin, generally there is some variation in number of branched rays; two individuals had 7 + 6 branched rays, in one 7 + 5 and another 7 + 8, but in this last case. the uppermost branched ray was possibly divided to the base.
Morphology. Body long, slightly compressed (Fig. 6 a). Head normally large with a pointed snout. Eyes moderately large situated somewhat near the tip of the snout, rather than the near margin of the operculum. The fins moderately large. The dorsal is inserted a little in the second half of the body. Ventrals insertion is situated on the same line of dorsal or a slightly in advance. Caudal peduncle long enough and well compressed laterally.
Mouth (Fig. 6 b) well arched. Upper (anterior) lip moderately thick laterally, and is thickened in its anterior part. Fine transverse grooves on the surface of this lip can be observed. Lower (posterior) lip fleshy, well folded with conspicuous mental lobes having pointed tips. Barbels small.
Suborbital spine (Fig. 6 d) is straight but with well curved caudal processes (thorns). The thorns are not divergent, the smaller being about half the length of the longer.
Scales (Fig. 6 c) nearly rectangular with rounded corners having a very small eccentric focal zone. They cover whole body except the head.
Sexual dimorphism. In stephanidisi the males have a large rounded lamina circularis (Fig. 6 e) at the base of the pectoral ray, which is more thickened than the first. This is covered by soft tissue and skin. The part which connects the ray with the plate-like process, is large enough. Such a part is lacking in the species of Bicanestrinia or somewhat reduced in other species of Cobitis s. str. as in vardarensis (compare the Figures of lamina circularis of vardarensis and stephanidisi).
CoJour pattern (based on preserved specimens). General ground colour is yellowish-white and the spots and dots dark-gray or bluish-dark gray. On the back there are 4-7, usually 5-6, relatively large predorsal spots, while under the dorsal there are only two. Postdorsal spots generally 6-7, rarely fewer or more. All four Gambetta's zones are prksent. First and

COBITIDS OF GREECE 305
third zones, composed of small dots, have a normal extension. The second zone is formed by long lines. The fourth zone is also formed by lines, or very long spots or a nearly continuous band. In the last quarter of the body the first, second and third zones mix and form a network of dots and reticulations. Head with numerous small dots and a dark-gray band between the anterior rim of orbit and tip of snout. Dorsal and caudal fin with numerous gray dots. Other fins pale each having one row of minute dots. On the upper part of the base of the caudal fin a large jet black spot.
Habitat: It is a typical still water living species, frequently all depths from very shallow to deeper places, especially where aquatic plants are present. From the general aspect of its type locality (Velestino spring) it can be assumed that the species does not avoid clean and slow running water. However, silty bottoms, into which it normally burrows, are the more favorite habitats.
Range (Fig. 17-12): To this date the species has been found only in a very limited area, in the Kefalovriso karstic spring at Velestino (Feres) village in eastern Thessaly. This spring belongs to drainage of lake Karla (now dried up), from which the species was listed by Panagiotopoulos (1916) as Cobitis taenia. The species may still survive in the canals passing through the basin of the former lake.
Remarks: This is an unusual species in its combination of characters of shape of body, squamation, suborbital spine, lamina circularis and especially in pigmentation. It is noteworthy that some specimens with somewhat similar colour pattern have been observed in a few Romanian rivers (see Bgngrescu & Nalbant, 1957).
Etymology: The name of this species is given in memory of Dr. Alexander I. Stephanidis (1911-1990), the first modern explorer of the Greek freshwater fish fauna.
Subgenus Bicanestrinia Bscescu, 1962
Cobitis strumicae Karaman, 1955 (Fig. 7 a-f)
Cobitis taenia, Kovatchev, 1921a: 88 (river Strymon), 1921 b. 93 (river Evros); Drensky. 1928: 167. fig. 4.5.1 (description, distribution: rivers Strymon. Nestos, Evros), 1930 (partim): 679 (list: rivers Strymon, Nestos, Evros), 1951 (partim): 124, fig. 80 (description; distribution: river basins of Mediterranean); Berg, 1932 (partim): 155. 1949 (partim): 455 (from literature: rivers Evros to Strymon); Michajlowa. 1865a: 66 (Strymon). 1965b: 280 (Evros); Russev, 1966: 268 (Evros); Economidis, 1973 (partim): 471 (from literature: rivers Strymon, Nestos, Evros). 559 (from collections: lakes Koronia-Volvi and Vistonis, and rivers Strymon-Aggitis, Filiouris, Loutros and Evros), 1974: 91, map 25, fig. 27 (description, variation. distribution: all waters from Strymon to Evros. except stream Marmaras); Economidis & Sinis. 1982: 305 (lakes Koronia and Volvi).
Cobitis taenia taenia. Drensky, 1929 (partim): 231 (list: rivers Evros, Nestos. Strymon).

306 P.S. ECONOMIDIS. T.T. NALBANT
Cobitis taenia vardarensis, Taler, 1953 (partim): 447 (river Strymon drainge: tributary Strumica).
Cobitis taenia strumicae Karaman, 1955: 190, fig. 4 (description as new sub-species, distribution: tributary Strumica, river Strymon drainage); Grupce & Dimovski, 1976: 34 (characters. comparisons with other species of South Yugoslavia).
Cobitis pechevi Siskov & Dobrobolov, 1984: 1673, fig. 1-3 (description as new species. isoelectric focusing of muscle myogens; may be a subspecies of strumicae).
Material: MZUT 1971-242 (23) 31.3-62.3, river Strymon, downstream of national roads's bridge near village Strymonikon, Macedonia, Greece, 31 August 1971, Economidis coll.; MZUT 1973-14 (11) 27.1-57.8, lake Volvi drainage, stream Rihios (Redina), Macedonia, Greece, 28 March 1978, Economidis coll.; MZUT 1972-130 (9) 30.8-69.7, river Nestos catchment, marsh near village Avramilia Kavalas, Macedonia, Greece, 6 July 1972, Economidis coll.; MZUT 1972-103 (18) 33.8-74.0, lake Vistonis, at shore near village Dialambi, Thrace, Greece, 5 July 1972, Economidis coll.; MZUT 1972-75 (29) 43.9-78.3, stream Filiouris, near village Karydia, Thrace, Greece, 4 July 1972, Economidis coll.; MZUT 1986-25 (241) 21.9-82.7, stream Bospos near Komotini town, Thrace, Greece, 20 August 1986, Economidis coll.; MZUT 1971-611 (15) 34.9-82.1, Stream near village Loutros Alexandroupolis, Thrace, Greece, 5 November 1971, Economidis coll.; MZUT 1986-35 (285) 26.7-88.7, river Evros catchment, tributary Erythropotamos near village Mikro Dereio, Thrace, Greece, 4 July 1972, Economidis coll.; ISBB 3971, (1) 94.0, same data as previous.; ISBB 3972, (20) 55.8-100.8, river Evros catchment, tributary near village Ardanio, Thrace, Greece, 5 October 1977, Economidis coll.; ISBB 3973, (14) 35.4-57.6, lake Volvi, Macedonia, Greece, 20 August 1971, Economidis coll.: ISBB 4579 (30) 36.3-88.8, stream Bospos, near Komotini town, Thrace, Greece, 20 August 1986, Economidis coll.
Description: Fin ray formula. Number of rays generally constant in all fins: D I11 7, A 11-111 5, VI 6-1 6, P I 9-1 9, C n 7 + 7 n. In few specimens there are 6 or 8 branched rays in the dorsal fin or 7 or 8 branched rays in pectorals.
Karaman (1955) described the species without providing complete information on its morphology.
Morphology. Body long, generally isomorphic (Fig. 7 a, b). Head moderately long, with snout relatively longer than postorbital space. Eyes normal in size, a little close to the hind margin of the head. The insertion of the dorsal fin is equidistant between the tip of snout and base of caudal fin. However there are specimens in which the predorsal distance is a little greater. Ventrals generally inserted on the same line of the dorsal fin but, in some specimens, these are inserted a little behind, under the second or third branched ray of dorsal. Caudal fin long enough.
Mouth (Fig. 7 c) generally well arched. Upper (or anterior) lip moderately thick, unfurrowed although fine grooves can be seen

COBITIDS OF GREECE 307
sometimes. Lower (posterior) lip fleshy, wide enough and well folded. Mental lobes are shorter than the rest of the lip but moderately large and pointed. Barbels short.
The suborbital spine (Fig. 7 e) is straight, with the longer thorn curved and the smaller about one third of the longer.
The subdorsal scales (Fig. 7 d) are generally rounded somewhat wider at base and with a small eccentric focal zone. The lateral line is reduced, and does not exceed the length of pectoral fins. As in all species of Cobitinae the lateral line is superimposed in each side of the body on a small swelling which is probably in connection with the gas-bladder capsule.
Sexual dimorphism (Fig. 7 f). Males with two laminae circulares, at base of first (unbranched) and second (branched) pectoral rays.
Colourpattern (of live specimens or freshly preserved ones). Ground colour yellowish-white, laterally with an golden-green iridescent band from the tip of the snout to the caudal base. The dorsal part is light olive with 9-15 dark gray blotches. The four Gambetta's zones are complete and usually as in Fig. 3 b, e, rarely as in Fig. 3 a. In strumicae from the Evros basin, from the river Nestos and from the stream Bospos, the lateral spots or blotches are elongated or sometimes two round spots are united to form one elongate blotch. Generally the specimens of strumicae from lake Volvi and from river Strymon system have well defined spots along the sides, round or square-like. The upper part of base of caudal fin without a jet black dot or this dot is gray and reduced to a small point. Dorsal and caudal fins with small dark brown or gray dots. Other fins pale.
Habitat: Over its very wide area of distribution this species presents a remarkable adaptation to different type of habitats with soft bottom. It can be observed from still water of lakes and river's death branch, with more or less deep water and rich vegetation, to slowly running and shallow water of streams. In these places the bottom change from muddy to silty or to sandy respectively, the vegetation being scarce or laking in the last case. In all these biotopes, the typical behavior of the fish, i.e. burrowing in the bottom, is always apparent, especially during summer when streams and death branch retain little water in shallows.
Range: (Fig. 17-7): In Greece this species is dispersed in eastern Macedonia: river Strymon basin, lakes Volvi and Koronia, systems of the Thracian rivers Nestos, Bospos, Filiouris and Evros. Outside Greece Cobitis strumicae is present in the Strymon system (Yugoslavia and Bulgaria) and in the river Evros drainage of Bulgaria (as Cobitis pechevi) and probably in Turkey. It is also present in a few rivers in Bulgaria, tributaries of the Black Sea, but until now has not been found in the basin of the river Danube.
Remarks: Obviously Cobitis strumicae in its elongated body and colour pattern is close to trichonica-meridionalis group, rather than to the

308 P.S. ECONOMIDIS. T.T. NALBANT
helenica-arachthosensis-punctilineata complex. However, strumicae has the scales with a reduced focal zone, and is therefore quite different from the western species meridionalis and trichonica.
Cobitis meridionalis Karaman, 1924 (Fig. 8 a-e)
Cobitis taenia var. meridionalis Karaman, 1924: 75 (short description as new var., distribution: lake Prespa); Stephanidis, 1950: 208 (Vrigiis or Prespa lakes system).
Cobitis taenia meridionalis. Taler. 1953: 447 (lake Prespa); Poljakov et a]., 1958: 103 (lake Prespa): Grupce & Dimovski, 1976: 30 (characters, comparison with other species of South Yugoslavia).
Cobitis meridionalis, Economidis. 1993: 23 (full species rank).
Material: MZUT 1988-104 (16) 34.3-78.9, lake Megali (Great) Prespa at village Psarades, Macedonia, Greece, 9 September 1988, Economidis coll.; ISBB 4568 (16) 49.5-81.0, bearing same data.
Description: Fin ray formula. D I11 7, A I1 5, V I 6-1 6, P I 9-1 9, C n 7 + 7 n, in a few specimens the number of dorsal branched rays is 6 and pectoral rays 8.
Morphology. Body elongate, head long (Fig. 8 a). Mouth arched with the lips a little furrowed, especially the inferior lip which is folded (Fig. 8 b).
Mental lobes well marked, in some specimens pointed. Suborbital spine straight with latero-caudal process very reduced
(Fig. 8 d). Scales are subcircular (Fig. 8 c) with a large focal area. They are
embedded and cover the whole body except the head. Sexual dimorphism: In males expresed as a thickened process
(Fig. 8 e) at the base of the first an second pectorl rays, both being covered by skin. Generally the pectoral fins are longer in males than in females.
The colour pattern is very characteristic in meridionalis (Fig. 8 a). The whole body is yellowish-white, with cloudy dark gray or olive blotches and dots. Back with 7-12 spots, generally rectangular. The second Gambetta's zone is formed by isolated spots, from the upper rear part of the operculum to the superior part of the caudal base. First and third Gambetta's zone is formed each by one row of speckles. The fourth Gambetta's zone is formed by 11-15 rounded, relatively small spots. Lateral median intramuscular septum blackish, very visible. Upper half of head with small dots, without any dusky band. The dot in upper part of caudal base is very reduced and grayish. Dorsal fin with 3-4 rows of gray dots. Caudal fin with 4-5 irregular rows of graysh dots. Sometimes on the anal, ventral and pectoral fins there is one row of grayish dots. Iris golden.
Habitat: The species is a typical still water fish, frequenting mainly lakes. It has also been observed in some short, stream-like still or slowly

COBITIDS OF GREECE 309
running water bodies, which over the year retain water intermittently, especially near to the mouths. However, as in the other close related species, this fish prefers habitats near the lake shore where muddy to silty bottoms and some vegetation are present.
Range (Fig. 17-5): To date, Cobitis meridionalis is known only as endemic to lake Prespa basin.
Remarks: Due to its pigmentation Cobitis meridionalis is well separated from the other members of Bicanestrinia. However, in its elongate body, lack of black spot at caudal base, presence of the black stripe on the intramuscular septum along the lateral midline of the body, this species appeas quite similar to Cobitis trichonica, a Bicanestrinia species with a western range in Balkan Peninsula too (see below). Both species may be holophyletic.
Cobitis trichonica Stephanidis, 1974 (Fig. 9 a-3)
Cobitis (Bicanestrinia) trichonica Stephanidis, 1974: 227, figs 1-3 (description as new species. distribution: lakes Trichonis and Lyssimachia).
Material: MZUT 1988-85, (10) 48.8-68.0, lake Trichonis catchment, tributary at village Panetolio near Malamatas fish farm, Etolia, Greece, 3 August 1988, Economidis coll.; ISBB 4578 (12) 38.2-78.3, bearing same data as previous; MZUT 1988-94, (10) 41.1-62.0, lake Trichonis catchement, tributary at village Myrtia, Etolia, Greece, 4 August 1988, Economidis coll.; MZUT 1988-124, (11) 43.0-67.5, lake Trichonis, Etolia, Greece, 15 September 1988, Economidis coll.; ISBB uncat., (13) 29.7-60.0, lake Ozeros catchment, channel to river Acheloos, Akarnania, Greece, 24 Sept. 1986, Economidis coll.; MZUT 1992-9, (1) 28.6, river Acheloos catchment, spring at village Lesini, Etolia, Greece, 22 April 1992, Economidis coll.; MZUT 1992-10, (3) 39.4-42.8, river Acheloos catchment, spring Agios Dimitrios Lesini, Etolia, Greece, 22 April 1992, Economidis coll.
Stephanidis (1974) gave a good description of the species. In the following are added some details.
Description: Fin ray formula. On most specimens it is as follows: D 11-111 7, A I1 5, V I 6-1 6, P 18-1 8, C n 7+7 n but a few individuals there are 8 branched rays in the dorsal and 7 or 9 branched rays in the pectoral.
Morphology. Body relatively long, but in a few specimens, both males and females, it is short and stout. Head relatively short with an arched upper profile (Fig. 9 a). Eye moderately small and placed about midway between tip of snout and hind margin of operculum. Dorsal and ventrals generally equidistant between tip of snot and base of caudal.

310 P.S. ECONOMIDIS. T.T. NALBANT
Mouth (Fig. 9 b) is arched enough with upper lip less furrowed but lower (or posterior) lip well folded with developed mental lobes, their tips being rounded rather than pointed. Barbels short.
Suborbital spine (Fig. 9 d) is straight with well diverging caudal processes (thorms) but these being reduced enough.
Scales (Fig. 9 c) are somewhat trapezoidal in shape with a large and eccentric focal zone. They cover entirely the body except the head. The lateral line, as in all Cobitinae loaches, is very reduced, not longer than the pectoral fins.
Sexual dimorphism. In all males there are two laminae circulares at the base of the first (unbranched) and second (branched) pectoral rays, both processes being covered by a soft tissue and skin. In males pectcrals and ventrals are longer than in females.
Colour pattern, in live specimens: the ground colour is yellowish- white, sides of head and body having an iridescent or golden band. Dorsum olive with 10-15 brown squareshaped spots. The fourth Gambetta's zones normally expressed as in Fig. 3 a. The colour of the dots and spots is dark brown or dark gray. In many specimens the fourth Gambetta's zone is rectangular and disposed vertically at least on caudal peduncle. The head is covered by dark gray or dark brown speckles. Generally the pigmentation of the head is similar to that shown in Fig. 2 c. At the base of the caudal fin there are two dark-gray spots. The inferior part of the caudal peduncle is sometimes pigmented with dark brown or grayish brown speckles. Dorsal and caudal fins with irregularly rows of dark brown dots, the rest of fins being whitish with generally one row of dark-brown dots. In many specimens the ground colour of the caudal fin is dull orange even reddish.
Habitat: The species present a rather clear preference for still water, given that in the area of its distribution, i.e. the aquatic system of river Acheloos, it is more frequent in lakes. However, it has also been observed in slow to moderately fast running water, as in the Panetolion stream, flowing into lake Trichonis, and the canal uniting lake Ozeros and river Acheloos. In all cases the habitats are characterized by clean water of different depths, a more or less rich aquatic vegetation and a muddy to silty bottom in which the fish burrows.
Range (Fig. 17-6): lake Trichonis and the adjacent lakes, Lysimachia, Ozeros, Amvrakia and also in the river Acheloos in Akarnania and Etolia, western Greece. Stephanidis (1974), did not found the species in lake Amvrakia. Nevertheless, the species has been observed once by one of us (PSE) near the shore of the lake, swimming among aquatic plants.
Remarks: This species is very distinct in its pigmentary features. However, apparently it is close to strumicae and possibly meridionalis in its relatively long body and colour pattern.

COBITIDS OF GREECE 311
Cobitis hellenica sp. nov. (Fig. 10 a-f and 12 g-i)
Cobitis taenia. Stephanidis, 1939: 32 (river Louros); Schmidt-Ries, 1943: 325 (from literature. river Louros).
Material: Holotype: MZUT 1989-115, (1) 69.3, river Louros catchment at Barbanakos spring near village Stephani, Epirus, Greece, 7 September 1989, Economidis coll.
Paratypes: MZUT 1963-5 (22) 48.9-85.0, river Louros catchment at Barbanakos spring near village Stephani, Epirus, Greece, 1 August 1963, Economidis coll.; ISBB 3967, (13) 45.0-81.2, same locality as previous, 17 August 1963, Economidis coll.; MZUT 1977-232 (5) 49.2-76.7, same locality as previous, 19 October 1977, Economidis coll.; MZUT 1987-42 (25) 39.6-60.7, same data as previous; RMNH, uncat, (4) 44.7-67.2, same locality as previous, 25 August 1987, Economidis coll.; ZSM, uncat. (2) 47.7-54.0, same data as previous; CAS, uncat. (6) 48.0-72.0, same data as previous; ISBB 4577, (33) 21.4-66.0, same locality as previous, 23 September 1985, Economidis coll.; ISBB 4570, (11) 43.0-66.0 same locality as previous, 28 August 1987, Economidis coll.; USNM 330135, (2) same locality as previous, 25 August 1987, Economidis coll.; SMF 27232 (2), bearing same data as previous; MNHN 1994-514, bearing same data as previous; NMW 92820 (1 & 2), bearing same data as previous; MZUT 1989-41 (34) 31.5-85.9, same locality as previous, 7 September 1989, Economidis coll.; MZUT 1979-22 (29) 43.0-70.3, river Louros catchment near village Philippiada, Epirus, Greece, 13 November 1979, Economidis coll.; ISBB 4569, (10) 45.0-74.0, river Louros catchment, near Hanopoulo village, Epirus, Greece, 7 August 1988, Economidis coll.; MZUT 1988-95 (15) 23.7-52.3, same locality as previous, 4 August 1988; MZUT 1989-43 (4) 38.7-79.5, river Louros at springs Agios Georgios, Epirus, Greece, 8 Sept. 1989, Economidis coll.; SMF 22318 (5) and MINB uncat., (2) all with same data as previous.
Diagnosis: A stout bodied species, laterally compressed, and a high caudal peduncle. Colour based on five pigmentary zones.
Description: Fin ray formula. D I11 7, A 11-111 5, V I 6-1 6, P I 9-1 9, C n 7 + 7 n. On a few specimens we found D 111 6 and even D 111 5 (ISBB specimens from Barbanakos spring, possibly mutilated). A reduced number of rays in the pectoral, 8 and 7, are also present in some specimens all of which are males.
Morphology. Body stout, very high in comparison to other species of Cobitis and well compressed (Fig. 10 a, b). Generally the body profile is isomorphic and are not taper from head to tail. The caudal peduncle is strongly laterally compressed. Head stout with moderate eyes which are sitdated generally close to snout than to the hind margin of head. Only in a

312 P.S. ECONOMIDIS. T.T. NALBANT
few specimens are they situated in the middle of the head. Dorsal fin inserted a little behind the midpoint from tip of snout to caudal base. Ventrals generally inserted under second or third dorsal branched ray. Caudal enough, with rounded margin. Paired fins are short in adults.
Mouth (Fig. 10 c) arched, with moderately thick and unfurrowed upper lip. Sometimes a few fine grooves can be seen on the surface of this lip. Lower lip fleshy, folded with well developed mental lobes. Barbels long enough.
Suborbital spine (Fig. 10 e) relatively straight, the two thorns (caudal processes) being curved, the smaller about half the length of the other.
Scales (Fig. 10 d) round, with a large focal zone, cover whole body excepting the head.
The lateral line, being very reduced, does not exceed the length of the pectorals.
Sexual dimorphism (Fig. 10 f). Males with two laminae circulares, one at base of first (unbranched) pectoral ray and another at the base of the second (branched) pectoral ray.
Colour pattern. In preserved specimens, very characteristic of this species, the ground colour is yellow-brown or orange-brown. On the dorsum there are 4-6 large dark brown spots before the dorsal fin. In juvenile specimens these spots are smaller. Under the dorsal insertion there are 2, rarely 3, dark brown spots. In post dorsal there are 4-6 large dark brown spots rarely less or more. The first zone of Gambetta is formed by small and irregular brown speckles. The second row is formed by round brown dots. The third row, like the first, is also formed by brown spekles. All three rows of dots and speckles in the last fourth part of the body mix and form an area with reticulations or vermiculations. The fourth zone consists of dark brown spots, generally square-like or triangular. The upper part of this row is superimposed on a black or a deep bluish stripe. In most specimens, under the third row there is another row of very small brown dots, generally interrupted in the mid part of the body. Head with a reticulation of gray-brown spekles or minute dots. The dark band before the anterior rim of orbit is hardly discernible. Dorsal and caudal with irregular rows of dark brown spots. Other fins usually pale, but sometimes these are pigmented with small brown dots. Upper base of caudal with a very small dark gray dots.
Habitat: As with many other loaches this species prefers still to moderate running water, better those of death branch of river, large ditches and springs. It frequents more or less deep water, where normally it burrows into the muddy to silty bottom. However, it can also be observed in shallow water but near aquatic plants, as in the Barbanakos spring.
Range: (Fig. 17-1). This species is found only in western Greece in the river Louros tributary in Amvrakikos Gulf, to the Ionian Sea.

COBITIDS OF GREECE 313
Remarks: Of course, hellenica and closely related species, possessing this strange combination of characters (a Bicanestrinia group with deep and compressed body and well pigmented) represent an interesting presence in the freshwater of Greece. It is very difficult to refer the hellenica complex to another group of species in southern Europe, especially from the Balkan Peninsula. In the circum-mediterranean region there are another four deepbodied species of Cobitis, vettonica and maroccana from the waters of Spain and Morocco, zanandreai from southern half of Italy and turcica, in the Central part of Turkey.The first three species have only one lamina circularis, i.e. belong to Cobitis s. str. But turcica deserves special attention. The examination of many specimens from central Turkey (see above in Material and Methods) revealed that these series agrees with the equivocal original description by Hanko (1925: 154, fig. 8). But he does not give some inlormation about sexual dimorphism (if he had some males). The two turcica types from "Eregli", were lost by fire in October 1956 in the National Museum of Natural History in Budapest. However, Hanko gave interesting information about colour pattern of his turcica, concerning the presence of a supplementary row of speckles under the fourth row of Gambetta. He states that: "under the lateral rows of spots, between the pectorals and ventrals, on the yellow ground colour 5, there are some dark fine row of dots" (in original: "Unter den Seitenstreifen zwischen den Pectoralen und Ventralen auf gelber Grundfarbe 5, aus feinen und weningen dunklen Punkten bestehende Nebelflecke"). If turcica, from Eregli belongs to the subgenus Bicanestrinia, it is obvious that it represent the closest related species to the hellenica complex.
Etymology: The name hellenica is derived from the name of the country (Hellas).
Cobifis arachthosensis sp. nov. (Fig. 12 a-f)
Material: Holotype: MZUT 1988-137, 41.9, river Arachthos death branch near Akropotamia village Epirus, Greece, 4 August 1988, Economidis coll.
Paratypes: MZUT 1977-356, (2) 52.1-78.9, river Arachthos at Arta, Epirus, Greece, 20 October 1977, Economidis coll.; ISBB 4571, (2) 49.0-53.0, bearing same data as previous; MZUT 1988-96, (5) 32.4-76.4, river Arachthos death branch near Akropotamia village Epirus, Greece, 4 August 1988, Economidis coll.; MZUT 1988-97, (46) 25.3-76.1, bearing same data as previous; ISBB 4574, (10) 25.042.0, bearing same as previous; USNM 330136, (1) bearing same data as previous; SMF 27233 (I), bearing same data as previous; MNHN 1994-515, (2) bearing same data as previous; NMW 92821 (I), bearing same data as previous.

314 P.S. ECONOMIDIS. T.T. NALBANT
Diagnosis: Similar in many respects to C. hellenica excepting young specimens in which the fourth zone of Gambetta is formed by round deep brown spots very remote from one another. The row with small dots of third zone of Gambetta is lacking.
Description: Fin ray formula. D I11 7, A I11 5, V I 6-1 6, P I 9-1 9, C n 7 + 7 n. Same variation in the number of dorsal and pectoral fin rays as in C. hellenica was observed.
Morphology. The shape of the body and other characters as mouth, suborbital spine. scales, sexual dimorphism are exactly as in hellenica. Howevcr. in many adults the lateral dark brown spots (fourth Gambetta's zone) are well spaced and round. In all juveniles of arachthosensis the lateral spots are few (9-15, mainly 11-12) and remote from one another. Thc third Gambetta's zone is completely lacking.
Habitat: As in the other loaches this is a typically benthic fish, presenting a remarkable adaption from still to moderately running water, and from clean to slightly turbid. It has been observed in the main bed of the river Arachthos, in a place with a more or less fast current, and in some of its death branches with a rather still water. Obviously it prefers biotopes with rich aquatic vegetation, but it may also found out of these. it also shows obvious variation of substrate preference, normally frequenting silty to muddy bottoms but also sandy or even gravely ones. The same variation has been observed in water depth, the fish living well in very shallow to deeper water.
Range (Fig. 17-2): The area of this taxon covers the Arachthos basin, a flowing also as Louros, to the Amvrakikos Gulf and then to Ionian sea.
Remarks: The main difference between arachthosensis and hellenica is in the pigmentation of juvenile specimens (compare Fig. 12 a-f with 12 g-i). This character is extremely important and can separate as good species arachthosensis from hellenica. The reduction of pigmentation represent an apomorphy. On the colour pattern this loach somewhat resembles another species of the subgenus Bicanestrinia, Cobitis linea, from the systems of rivers Kor and Kul, basin of the Arabian Sea, Iran (Bianco & Nalbant, 1980: 904, fig. 1 A).
A recently discovered population from the river Thyamis (about 100 km northwestward), which is now under scrutiny, seems to agree well enough with those from river Arachthos. Initially both populations are regarded as conspecific.
Etymology: The name was given after the river Arachthos in which this loach was found.
Cobitis punctilineata sp. nov. (Fig. 11, 12 1-11
Cobitis taenia. Economidis. 1974 (partim): 91 (river Strymon drainage: Tributary Aggitis. color pattern and other features).

COBITIDS OF GREECE 315
Material: Holotype: M Z U T 1992-39, 59.0, Strymon-Aggit is catchment, channels near the Kalambaki village, Drama, Macedonia Greece, 19 January 1992, Economidis and Nalbant coll.
Paratypes: MZUT 1971-389 (3) 31.1-65.0, Strymon catchment, tributary Aggitis ditch near Agia Paraskevi village, Drama. Macedonia, greece, 23 September 1971, Economidis coll.; MZUT 1972-200 (2) 62.1-75.5, Strymon-Aggitis catchment spring near Kephalari village, Drama, Macedonia, Greece, 1 October 1972, Economidis coll.; MZUT 1973-22 (6) 27.3-59.8, same locality as previous, 7 September 1973, Economidis coll.; MZUT 1973-30 (1) 62.6, Strymon-Aggitis catchment spring near Mylopotamos village, Drama, Macedonia, Greece, 7 September 1973, Economidis coll.; MZUT 1988-72 (15) 27.1-60.0, Strymon-Aggitis catchment channels near the Philippi village, Kavala, Macedonia, Greece, 20 July 1988, Economidis coll.; MZUT 1989-70 (18) 47.3-95.0, Strymon- Aggitis catchment, channels near the Kalambaki village, Drama, Macedonia Greece, 14 October 1989, Economidis and BTinirescu coll.; ISBB 4576, (3) 50.0-51.0, bearing same data as previous; USNM 330138, (2) bearing same data as previous; SMF 22317 (4) and SMF 27235 (2), bearing same data as previous; MNHN 1994-517 (2), bearing same data as previous; NMW 92823 (2), bearing same data as previous.
Diagnosis: A stout bodied species, with a complete pigmentation of Gambetta's zones, but the fourth one is formed by small round deep brown spots. A supplementary pigmentation is present under this row of lateral spots.
Description: Fin ray formula. D I11 7, A 11-111 5, V I 6-1 6, P I 9-1 9, C n 7 + 7 n. The same limits of variation in the number of fin rays as in hellenica were observed in punctilineata.
Morphology. Stout bodied species, with compressed caudal peduncle (Fig. 11 a). The whole profile of the body is isomorphic. Head stout and blunt. Eyes moderate in size, placed nearer to the tip of the snout than to the hind margin of the opercule. The dorsal fin is situated a little behind the midpoint from the tip of the snout to the base of the caudal fin. Ventrals inserted under the second, rarely under the third, branched ray. The dorsal, anal and caudal fins are moderately long but paired fins are short in adults.
Mouth (Fig. 11 b), arched, upper (anterior) lip being moderately thick with very fine transverse furrows. Lower (posterior) lip fleshy but with very few folds, sometimes only one fold. Mental lobes relatively reduced. Possibly the length of these lobes is related to sexual activity. The barbels are long enough.

316 P.S. ECONOMIDIS. T.T. NALBANT
Suborbital spine (Fig. 11 d) is a little curved its thorns (caudal processes) also slightly curved, the smallest forming a half from the longest in its length.
Scales (Fig. 11 c) are large, elliptical with a large eccentric focal zone. They cover the whole body except the head. The lateral line is very reduced, as in hellenica, arachthosensis and other species.
Sexual dimorphism (Fig. 11 e). There are two laminae circulares at the base of the first (unbranched) and second (branched) pectoral rays in males. Sometimes in full adults very fine and small buttons are observed in both sexes.
Colour pattern. In the live specimens the ground colour is yellowish- white, bright olive on the back. Along the lateral midline of the body a golden. iridescent band is present. On the median part of back, before the dorsal fin , there are 6-8, mostly 7 and rarely 9, round dark gray spots. Under the dorsal fin there are always two spots and on the post-dorsal space 7-9 spots. The first Gambetta's zone consists of very fine speckles. The second zone is formed by relatively small dots. The third zone is exactly similar to the first zone, with very fine speckles. In the last quarter of the body these three zones are merged. The fourth zone is formed by small and rounded dark gray spots, generally more than 40, superimposed on a black or dark blue stripe. A fifth row of fine speckles is present under this. The dorsal and caudal fins have 4-6 rows of brown dots. The head is covered by very fine dark gray speckles. In the upper part of the base of caudal fin there is a faint brown small dot. Other fins pale sometimes with dark dots.
Habitat: This species has evolved in a lenitic area characterized by more or less shallow and still, but normally clean, water. Its typical habitat is the death branches of streams or springs and ditches, with a still water, a moderately depth and sometimes a slight current. Bottoms, in which the fish prefers to burrow, are normally muddly to silty. Aquatic vegetation if present is of preference of fish.
Range (Fig. 17-4): This species is present only in a system of streams and channels, tributaries of the stream Aggitis, of the lower Strymon system.
Remarks: This species is very close to hellenica and arachthosensis, but differs essentially from them in its combination of colour pattern, especially in the great number of lateral spots (see Fig. 12j-1). This species agrees in many respects with Cobitis turcica Hanko (1925). There is a single but major difference between turcica and punctilineata: in the former the scales have an eccentric and very reduced focal zone while in the latter this is very large.
Etymology: The name was given for the great number of dark spots in the middle lateral row.

COBITIDS OF GREECE 317
Genus SABANEJEWIA Vladykov, 1929 Sabanejewia aurata balcanica (Karaman, 1922)
(Fig. 13 a-e and 16 a-c)
Cobitis balcanica Karaman, 1922: 189, fig. 1-2 (description as new species, characters, comparison distribution Vardar at Skopie and Veles); 1924: 75 (short description: river Vardar or Axios near Skopje); 1928: 164,174 (river Axios or Vardar).
Cobitis aurata balcanica. Dimovski & Grupce, 1971: 27 (river Axios drainage: tributary BrCgalnitsa); 1972: 199 (river Axios drainage: tributary Treska); Grupce & Dimovski. 1973: 89 (river Axios system); 1976: 43 (characters, distribution: river Axios drainage).
Subanejawia aurata. Economidis, Kattoulas & Stephanidis. 1981: 93 )list: rivers Edesseos and Itamos); Economidis & Voyadjis, 1985: 92 (list Axios, Doirani, Gallikos).
Cobitis aurata, Economidis, 1973 (partim): 561 (from collections: rivers Axios and Gallikos).
Material: MZUT 1969-3, (1) 41.6, stream Anthemous, near the village of Vasilika, Macedonia, Greece, 10 October 1969, collected by T. Gaitatzis; MZUT 1977-284, (8) 51.2-75.1, stream Gallikos, near Kolchida village, Macedonia, Greece, 8 November 1977, Economidis coll.; MZUT 1977-324. (7) 51.8-77.7, stream Gallikos, near the village of Petroto Kilkis, Macedonia, Greece, 15 November 1977, Economidis coll.; MZUT 1977-312. (1) 56.6, stream Mavroneri, near Milia Katerinis, Macedonia, Greece, 15 November 1977, Economidis coll.; MZUT 1988-24, 861) 31 .l-51.4, river Pinios catchement, tributary Enipeas upstream from the bridge of national road Velestino-Farsala, Thessaly, Greece, 14 April 1988, Economidis coll.
Description: Fin ray formula. D I11 6, A I1 5, V I 6-1 6, P I 8-1 8, C n 6 + 6 n. The variation in the number of branched rays in the dorsal fin is large enough. So that there are some specimens with 5 rays (a very few) or with 7 rays. In ventral fins there are generally 5 rays but always there is a supplementary ray which may be counted as the 6th. In the pectoral fins the number is also variable. There are some specimens with 7 branched rays or 9, even, 10, rays.
Morphology. Body long enough, generally isomorphic, with quite a short caudal peduncle (Fig. 13 a). The head is moderately long with an arched superior profile. Eyes situated closer to the hind margin of the opercule than to the tip of snout. The dorsal insertion is in the middle of the body or before it. However, in a few number of specimens this insertion is a little behind the middle of the body. The ventral insertion is generally placed under first or second branched ray of the dorsal fin. Behind the dorsal fin, along to the caudal peduncle, there is a conspicuous keel. Such a keel is present also on the latter half of the lower part of the caudal peduncle.

318 P.S. ECONOMIDIS. T.T. NALBANT
The mouth (Fig. 13 c) has moderate thick lips. The upper (anterior) lip is strongly furrowed, crossed by deep grooves. The lower (posterior) lip is fleshy and folded enough. The mental lobes are well developed with two or three small excresence or lobules.
The suborbital spine (Fig. 13 d) is straight with very divergent caudal processes (thorns). The scales (Fig. 13 e) are elliptical with large slightly eccentric focal zone. They are slightly embedded and cover the whole body except the head. The lateral line is very short, not exceeding the length of pectorals.
Sexual dimorphism (Fig. 13 b). Adult males have on each side of body, before the dorsal fin, a conspicuous swelling. Behind the dorsal fin there is another one but not as large as the first ones.
Colour pattern (Fig. 13 a, 16 a-c), based on preserved material: Ground colour yellowish or bright orange-brown. On back there are before the dorsal fin 3-6. mostly 4, square-like brown or dark gray blotches. Dorso-lateral pigmentation is based on 7-14, mostly 10-11, large dark brown spots, generally square-like in shape. Head and its sides covered by dark brown small speckles. From the antrior rim of the orbit to the tip of snout there is a blackish stripe. Sometimes this stripe may be very deep blue. At the caudal base there are always two dark-brown distinct spots. All fins arc translucent but the caudal and dorsal have irregular rows of small brownish or graysh dots.
Habitat: The species is a typical stream-living fish, preferring more the main bed of flowing water, in the bottom of which it burrows into the sand. Normally it avoids silty or muddy bottoms, while it can be observed sometimes on gravel. Such places have shallow but clean water with a moderate current, but are almost without any aquatic vegetation.
Range: (Fig. 18-1): The area of this subspecies is a very large one (BSnirescu et al., 1972). i t includes most of the Danubian basin, Vistula basin, and nearly all Axios basin in Yugoslavia and Greece. In Greece it also present i n the basins of rivers Gallikos, Loudias and Pinios, in Macedonia and Thessaly.
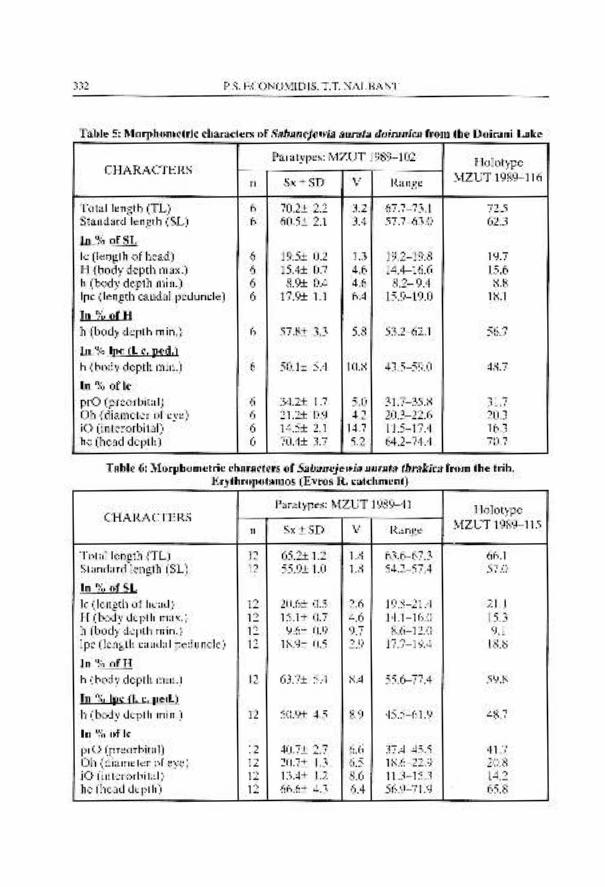
Sabanejewia aurata doiranica ssp. nov. (Fig. 14 a-d and 16 d-f)
Material: Holotype. MZUT 1989-116, 63.0, lake Doirani, Macedonia, Greece, 25 october 1989, Economidis and BBnBrescu coll.
Paratypes. MZUT 1989-102 (5) 57.7-66.0, lake Doirani, Macedonia, greece, 25 october 1989, Economidis and BiinBrescu coll.; ISBB 4580, (16)57.0-65.4, same data as previous.
Diagnosis: A subspecies of Sabanejewia aurata living only in lake Doirani with lateral spots very reduced in size.

COBITIDS OF GREECE 319
Description: Fin ray formula. D I11 7, A I1 5, V I 7-1 7, P I 8-1 8, C n 6 + 6 n C, there is some variation in the number of branched rays in dorsal (6 rays), ventrals (6,s and 9), pectorals (9) and caudal (7 + 6).
Morphology. Generally there are no differences in body proportions between doiranica and balcanica. However, in doiranica the specimens appears to be thicker than in balcanica or the next subspecies thrakica (Fig. 14 a).
The mouth (Fig. 14 b) has an upper (anterior) lip, more or less furrowed with fine grooves on its surface. Lower (posterior) lip generally fleshy with well developed mental lobes with two or three excrescence. Barbels somewhat long, although shorter than in balcanica, but this is a very variable character.
The suborbital spine (Fig. 14 d) is straight with well diverging caudal processes (thorns) and with a reduced size comparatively with balcanica.
The scales (Fig. 14 c) are sub-circular, with a large eccentric enough focal zone. They cover whole body excepting head and are slightly embedded. Lateral line very reduced.
The sexual dimorphism is similar to that of the other species of the genus Sabanejewia. In adult males, on each side of the body, before the dorsal fin there is a conspicuous swell.
The colour pattern (Fig. 14 a, Fig. 16 d-f) based on preserved specimens is the same as that in balcanica except for the lateral row of dark gray spots. These spots are very reduced, to rounded or thin vertically short stripes. This unusual pigmentation of Sabanejewia aurata is found only in the specimens of Sabanejewia aurata aralensis (see Berg, 1949: 461, fig. 646) from the lower part of Syr-Daria and Amu- Daria, in a population of Sabanejewia aurata from the lower Mures basin in Romania and "Cobitis" (in fact Sabanejewia) albicoloris of Chichkof (1932) from river Provadishka in Bulgaria.
Habitat: In spite of the rheophilic character of the aurata species this sub-species is typical of still water living fishes, observed up to now only in lake Doirani. The population studied was observed in clean and shallow water, not more than 50 cm. Living specimens have been observed to swim close to the bottom, just before they burrow quickly into the sandy bottom. Aquatic plants, especially Myriophyllum, were very scarce in the area.
Range (Fig. 18-2): This subspecies is confined to the Doirani. It is the single known form which is living in a lake.
Remarks: This subspecies is unique in its pigentation and ecological characters among other subspecies of Sabanejewia aurata. This population is probably a derivative of the populations of Axios basin but isolated in lake Doirani from a long time.
Etymology: The name of this subspecies comes from the name of the lake in which this interesting loach was found.

320 P.S. ECONOMIDIS. T.T. NALBANT
Sabanejewia aurata thrakica ssp. nov. (Fig. 15 and 16 g-i)
Cobitis balcanica. Drensky, 1926: 138 (distribution: river Evros); 1928 (partim): 169. fig. 5.3 (distribution: river Evros); 1929 (partim); 230. 231 (distribution: river Evros); 1930: 679 (list: river Evros
Cobitis aurata. Berg. 1932 (partim): 155 (distribution: river Evros): Berg. 1949 (partim): 459 (distribution: river Evros): Economidis. 1973 (partim): 561 (collection: river Evros drainage: tributary Erythropotamos).
Cobitis aurata balcanica. Drensky. 1948: 44 (distribution: river Evros); 1951: fig. 81 (description. distribution: river Evros); Binirescu, 1960 (partim): 125 (distribution: river Evros).
Sabanejewia aurata. Economidis. 1974: 94. map 26. fig. 28 (description. variation. distribution. river Evros drainage: tributary Erythropotamos).
Material: Holotype. MZUT 1986-54 57.4, river Evros catchment. tributary Erythropotamos, near Mikro Dereio village. Thrace, Greece. 24 August 1986, Economidis coll.
Paratypes. MZUT 1971-654 (1) 34.7. river Evros catchement. tributary Erythropotamos, near village Mikro Dereio. Thrace, Greece, 4 November 1971. Economidis coll.; MZUT 1986-39 (43) 27.5-62.6, river Evros catchment, tributary Erythropotamos, near village Mikro Dereio. Thrace, Greece, 21 August 1986, Economidis coll.: ISBB 4581, (43) 30.9-66.2, same locality as previous, 24 August 1986, Economidis coll.
Diagnosis: A subspecies of Sabanejewia aurata. very close to balcanica in its features but differing from it in pigmentation, having more numerous and larger lateral spots.
Description: Fin ray formula. D I11 7, A I1 5. V I 7-1 7, P I 8-1 8. C n 6 + 6 n. There is some variation in the number of branched rays in the caudal fin 7 + 6 and 6 + 4.
Morphology. The mouth is similar to that of balcanica. The mental lobes are more reduced than in doiranica. Barbels are short or very short sometimes as in Cobitis species.
The suborbital spine (Fig. 15 b) is straight with diverging caudal processes (thorns) is reduced enough comparing with those of balcanica, apparently even more reduced than doiranica.
The scales are sub-circular with a large eccentric focal zone. They cover the entire body, except the head. Lateral line very reduced. not exceeding the length of the pectorals.
The sexual dimorphism is similar to that in other Sabanejewia species and subspecies of aura fa.
The colour pattern (Fig. 15 a and Fig. 16 g-i) in preserved specimens. Ground colour yellowish or orange-brown. The blotches, spots, dots or speckles are deep-brown. In the back there are 4-7 irregular pre-dorsal spots. Some of them are thin or triangular, others square-like and large enough. Under dorsal there are always two spots generally large. Post-

COBITIDS OF GREECE 321
dorsal spots vary in number and shape. The limits of variation in the number of these spots are between 4 and 8. Their shape is quite variable, from rounded to triangular or square-like. The lateral spots are generally rectangular, sometimes triangular and elongated vertically. Rounded spots may be present also but they are rare. The number of lateral spots are quite variable, from 7 to 18, mostly 15-16. The high number and vertically arrangement of these spots is characteristic of this subspecies. Dorso-lateral pigmentation, speckels and dots, occur in the space from the hind margin of the head to the caudal base. Two very dark brown or gray lunate dots are present at the base of caudal fin. Sometimes there is a supplementary row of dark speckles under the lateral spots. The head, except its ventral part, is covered by dark speckles. From the anterior rim of the orbit to the tip of the snout, a nearly black stripe is present. Both dorsal and caudal fins have irregular rows of small dots.
Habitat: The type locality of this fish was examined for the first time (Erythropotamos tributary of Evros, near Dereio) in summer time, is a very shallow area (no more than 50 cm), with a sandy to gravely bottom and without any aquatic vegetation. The water was quite clean and its current not very fast. Obviously, this situation changes to more turbid, deeper and faster running water, during high water when it is raining, especially in late winter and spring.
Range (Fig. 18-3): This subspecies of Sabajewia aurata is confined to the river Evros basin.
Etymology: The name is derivated from the Greek name of region Thraki (Thrace), in which this subspecies was found.
DISCUSSION
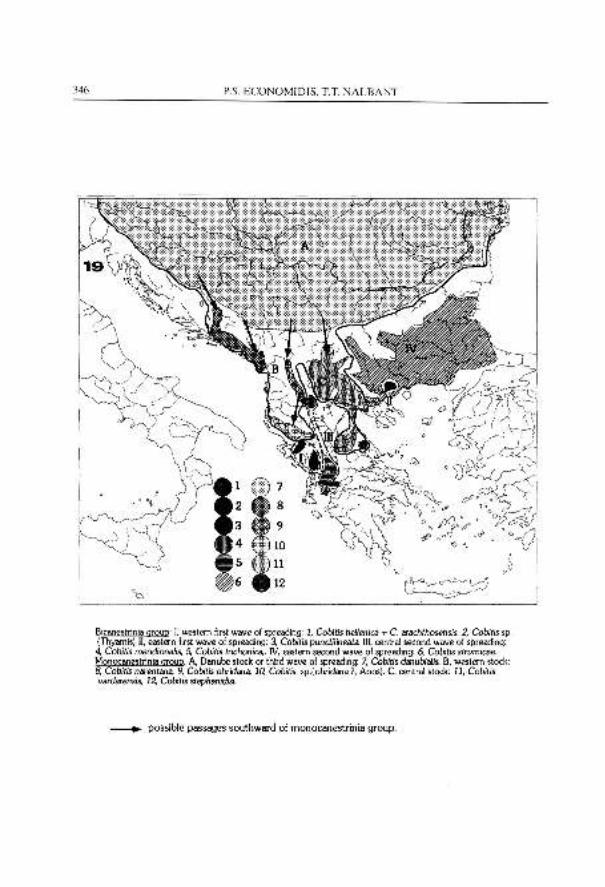
Zoogeographical and phylogenetic aspects of the species of Cobitis and Sabanejewia of Balkan peninsula and Anatolia are one of the most fascinating problem of the family Cobitidae. Therefore, a solution concerning the phylogeny of the genus Cobitis in the Balkan Peninsula cannot be presented without a discussion of Anatolian species as well. However, the phylogenetic and systematic aspects of the Anatolian species will form a subjet of a separate paper (Erkakan, Ekmekci, Balik & Nalbant, in press). In the present contribution we shall try to make an outline of the problem of origin of different species 'of Cobitis in the Balkan Peninsula.
During the Middle Miocene, the Balkan area was populated from the north (i.e. Central Europe) by one or even more species of Cobitis, but the position of Bicanestrinia group deserves a special attention.

322 P.S. ECONOMIDIS. T.T. NALBANT
Possibly, a single and a very simple lamina circularis represents the most primitive condition, a feature met in the genus Iksookimia (Kim et al., 1976; Nalbant, 1993) and in a few species of Cobitis such as C. biwae (see Okada, 1960) and C. takatsuensis (see Mizuno, 1970). The "mono- canestrinia" type was largely spread from East Asia to Europe. Possibly in the Middle Miocene this primitive allelomorph gene which gave these strange aspects of lamina circularis was blocked during the dispersal of Cobitis across the Euro-Asiatic continent. It is also possible, that in the Balkan area the allelomorphic gene of a simply lamina circularis was reactivated and moreover gave two simply plates at the base of first two pectoral rays. Both allelomorph genes were selected but in different territories: "monocanestrinia" (i.e. Cobitis s. str.) in whole Asia and Europe and Bicanestrinia in Balkan peninsula. Lodi (1979, 1980) has already pointed out that the lamina circularis (or Canestrini's organ) can be different mutants within of a population. Of course, the males with two or even three laminae circulares at the base of the pectoral fin are extremely rare in a population of "monocanestrinia" (i.e. Cobitis s. str.) species. However, this explains how a hidden character preserved for a long time in the gene pool of a lineage, under some circumstances, can be expressed and even selected. Possibly the period of first selection of the Bicanestrinia type in the Balkan area was about 15 my during the Middle Miocene. probably this type covered the whole hydrographic network of Balkan Peninsula and then Anatolia. It was represented by one or more species, one of them being the ancestor of the hellenica - punctilineata - turcica lineage. In the Upper Miocene, during the Messinian salinity crisis, a new lineage was formed from the initial stock of Bicanestrinia, as a result of a relatively rapid transformation of the Balkan territory (see Hsii, 1972; Hsii & Giovanoli, 1979; Bianco, 1990; Almaqa, 1990). The evolution of this new lineage (the ancestors of present meridionalis, trichonica, strumicae and levantina group) was gradually extended across the Aegeis land connection to the whole of Anataolia as far as Orontes basin. The new wave of Bicanestrinia was more competitive than the first one so that it transformed the area of the former (hellenica - punctilineata - turcica lineage) to a small relict area. Special attention must be given to the relation between C. strumicae and C, punctilineata, which nowadays are living sympatrically in the river Strymon drainage. As noted above the distribution of punctilineata is restricted only in the upper Aggitis catchment, which collects from the Philippi plain, a former shallow lake, and flows into the river Strymon. According to Vavliakis et al. (1986) the inner Aggitis catchment (Drama basin) was isolated from the river Strymon valley during the Lower to the Middle Miocene and remained isolated until about the Lower Quaternary. So, isolation of the punctilineata stem could be dated

COBITIDS OF GREECE 323
from this time, because geomorphological events taking place later in the Strymon valley did not influence very much this area. As cited above, the strumicae stem is younger in the area, where it was not possible to mix with punctilineata not more than Quaternary period, when speciation of punctilineata was already completed.
The deterioration of the climatic conditions in the Pliocene and then in the Pleistocene, favored the spreading of the northern main stem, those of Cobitis s. str. or the "monocanestrinia" group. The penetration of this group in Balkan peninsula and western Anatolia took place relatively recently, although the ways and periods were different. A progresive cooling climate especially during the last million years (Pleistocene) pushed southward the fauna including freshwater fishes. This displacement to the south was facilitated by a permanent change in the hydrographic network. The former continuous range of Bicanestrinia was cut in many parts by the penetration from the north of the Cobitis s. str. lineage (see Fig. 19). The first route followed was across the Dalmatian corridor (Fig. 19, B) by the ancestors of narentana - dalmatina and a unnamed "monocanestrinia" species. This route was used too by other species like Barbus peloponnesius, Gobio gobio, Alburnoides bipunctatus etc., (see Economidis & B5n?irescu, 1991). Today, the western part of Greece, a relatively long territory (Fig. 19, I), contains the small areas of four Bicanestrinia species: two of them (hellenica and arachthosensis) being from the older stock of Bicanestrinia and often two, one unnamed and the southernmost trichonica belong to the new stock. The penetration of central European (i.e. Danubian) fauna was made along the tributaries of the river Sava (possibly the Kupa and Una basins) which are very close to the Adriatic sea border and by the Lika, Cetina, Neretva and Drin which belong to the Adriatic slope.
A new penetration of Danubian fauna was the result of a connection of this basin with those of Axios (Vardar) through the Morava - Vardar corridor (for other details see Economidis & Bgnilrescu, 1991: 279). By this passage the ancestors of the present Cobitis vardarensis and of Cobitis stephanidisi spread southward. The closest relative of vardarensis is Cobitis danubialis from the Danube basin, possibly a good species different enough from taenia from northern part of Europe. Cobitis vardarensis preserves at least one pleisomorphy of the former stock, a high body. Concerning the scale feature it is difficult to say if a reduced and eccentric focal area represents an apomorphy or a plesiomorphy. This problem will be dis- cussed later. Concerning stephanidisi it appears to be very close to some specimens which were accidentaly observed in different rivers in Romania (see also Bgngrescu & Nalbant, 1957: 285, Fig. 3 and 4). But this allelic morph was selected and fixed only in Greece, in lake Karla basin.

3 24 P.S. ECONOMIDIS. T.T. NALBANT
The penetration of Danubian Cobitis s. str. species, through the Morava - Vardar corridor, spilt again the former Bicanestrinia area, so that it has produced differentiation in a new lineage of this subgenus (i.e. m eridionalis, strumicae) .
The problem of scale structure deserves special attention. This was discussed long ago by Kobayasi (1954, 1957) but until now remains unsolved. Generally a very reduced and eccentric focal area should be considered as a plesiomorphy since it is met in primitive species of Botia, Leptobotia, Misgurnus, Lepidocephalichtys. But almost all species of Cobitis, from Japan to central Europe, have the scales with a large focal area and only a few, from the southern part of Europe and Anatolia (i.e. maroccana), have the scales with a reduced focal zone. Possibly this synplesiomorphy, which units the members of a southern Cobitis, was selected under a few special condition of this area.
Concerning the polytypic species of Sabanejewia aurata, their area covers those of Cobitis vardarensis, i.e. Axios, Loudias, Gallikos and Pinios basins, and after an east-west discontinuity this species reappears in Evros basin (see Fig. 18). As in other parts of its range (see BSnZirescu et al., 1972) this species forms local geographical races.
A phylogeny of the species of Cobitis in Greece was not possible until now due to the polyphyly of lineages beyond the limits of the country and due to the unknown lineages from other parts of the areas.
The authors of this study believe that loach species, especially those which are living in isolated and restricted areas, as in Greece, consist of some of the most important elements of European Nature. Therefore, they need more care and protection and a better understanding concerning their origin and life history. More studies, especially those using modern approaches, are desirable.
ACKNOWLEDGMENTS
Authors extend their thanks to the following persons for their important help during the preparation of this work. in providing material and information: Drs Pier Giorgio Bianco (L'Aquila). Giovanni Delmastro (Carmagnola). Marie-Louise Bauchot (Paris). Erik Ahlander (Stockholm). I k Soo Kim (Chonju), Fusun Erkakan and Giiler Ekmekci (Ankara). Suleyman Balik (Izmir), Milorad Mrakovcic (Zagreb). lgnacio Doadrio (Madrid) and Mihai C. Bacescu (Bucure~ti). Special thanks go to Dr. Peter J. Miller (Bristol) and Dr. Petru M. BSnSrescu (Bucurevti) for discussion. criticism and valuable suggestions. PSE also thanks Greek Ministry of Agriculture (Fishery Service) for financial supporting the study of Greek endemic freshwater fish fauna. and Dr. Marilena Zarfdijan. Dr. Dimitra Bobori and Mr. Vassilis Vogiatzis MS. main staff of Laboratory of Ichthyology of Aristotle University. for any kind of assistance.

COBITIDS OF GREECE 325
INSTITUTION ABBREVIATIONS
BMNH: British Museum (Natural History), London CAS: California Academy of Sciences, San Francisco ISBB: Institutul de Stiinfe Biologice, Bucureqti MINB: Muzeul de Istorie Natural5 ,,Grigore Antipa", Bucureqti MNHN: Museum National d'Histoire Naturelle, Paris MSNT: Museo di Storia Naturale, Torino MZUT: Museum of Zoology of Aristotle University, Thessaloniky NMC: National Museum of Canada, Ottawa, Ontario NMW: Naturhistorisches Museum, Wien RMNH: Royal Museum of Natural History, Stockholm SMF: Senckenberg Museum und Forschungsinstitut, Frankfurt am Main USNM: National Museum of Natural History, Washington D C ZMB: Zoologisches M~lseum, Berlin ZSM: Zoologisches Staatssammlung, Miinchen
STUDIU ASUPRA COBITIDELOR DIN GENURILE COBITIS $1 SABANEJEWIA (PISCES, COBITIDAE) DIN GRECIA
CU DESCRIEREA A SASE TAXONI NO1
REZUMAT
Cobitidele Greciei din genurile Cobitis qi Sabanejewia au in total 9 specii, dintre care 8 aparfin primului gen qi una celui de a1 doilea. Pentru Cobitis sunt descrise 4 specii noi (stephanidisi, hellenica, arachthosensis qi punctilineata) iar pentru Sabanejewia aurata 2 subspecii noi (doiranica qi thrakica). Se face o amp15 discutie asupra originii faunei greceqti de cobitide.
REFERENCES
ALMACA, (C.), 1990 - Neogene Circum-Mediterranean peleogeography and Euro- Mediterranean Biogeography. Arq. Mus. Boc. Nova sCrie 1 (41): 585-61 1.
APOSTOLIDIS, (N.). 1892 - Les poissons d'eau douce de la Thessalie. Athknes, 31 p. (in Greek).
APOSTOLIDIS, (N.), 1907 - La p&che en Grkce. Faune Ichthyologique de Grkce. Athknes. 68 PP.
BANARESCU. (P.M.), 1960 - Einige Fragen zur Herkunft und Verbeitung der Siisswasserfischfauna der Europaischmediterranean Unterregion. Arch. Hydrobiol. 55: 16-134.
BANARESCU. (P) - Pisces-Osteichthyes. Fauna Republicii Populare Romfine. Bucure~ti, 962 pp.