Embed Size (px)
Citation preview

BRUCE D. CLARKSON 59
DSIR Land Resources, C/- FRI, Private Bag 3020, Rotorua, New Zealand.
A REVIEW OF VEGETATION DEVELOPMENT FOLLOWINGRECENT (<450 YEARS) VOLCANIC DISTURBANCE INNORTH ISLAND, NEW ZEALANDSummary: Volcanic activity has damaged or destroyed at least 20,000 ha of indigenous vegetation in theNorth Island in the last 450 years. The primary and secondary successions initiated are discussed andconsidered in relation to some recently proposed models of succession. Most of the variation relates to thescale and intensity of disturbance, and the type and heterogeneity of the substrate emplaced. Wheredisturbance has been extensive and severe, elements of slow 'classical' successions, from lichens andmosses to flowering plants, and facilitation by nitrogen-fixers such as Coriaria spp., are evident. Incontrast, minor disturbance results in vegetative regrowth or regeneration of surviving species.Heterogeneity of substrate enables many taxonomic plant groups to establish more or less concurrently.Partial disturbance successions are not easily categorised and may contain elements of several successionalmodels. The resource-ratio hypothesis of succession can be used to explain the general pattern of thevolcanic successions outlined. Insufficient information, of the type suggested necessary by recent authors,is available to make detailed accurate predictions in respect of future volcanically-initiated successions.Recent volcanism has had a major influence on species distribution and vegetation patterns in the NorthIsland.
Keywords: Succession; volcanic disturbance; vegetation; flora; New Zealand.
IntroductionThis paper reviews vegetation developmentfollowing disturbance by recent (<450 years)volcanic activity in the North Island, and discussesthe applicability of various models of plantsuccession. Eruptions from major volcanic ventshave occurred on seven volcanoes (Cole and Nairn,1975): Rangitoto Island, White Island (Whakaari),Mt Tarawera, Mt Ngauruhoe, Mt Tongariro, MtRuapehu and Mt Taranaki (Mt Egmont) (Fig. 1).A range of vegetation disturbance from minordamage to complete destruction has occurred,spanning altitudes from sea level to more than 2000m. Disturbance is here used in the sense of Sousa(1984), that is, a discrete, punctuated killing,displacement, or damaging of one or moreindividuals (or colonies) that directly or indirectlycreates an opportunity for new individuals (orcolonies) to become established.
Succession and successional models areconsidered first, followed by a select history ofvolcanic disturbance and vegetation development oflocations which have been studied by the author.In the third section, an ecological interpretation ofthe successions outlined is provided. Fourth, thesuccessional models described earlier are applied tosome examples of vegetation developmentfollowing volcanic disturbance. Concludingcomments concern the chances of predictingsuccessional trends after volcanic disturbance.
Models of successionFor the purposes of this review succession isconsidered a directional change with time in speciescomposition and vegetation physiognomy of asingle site where climate remains effectivelyconstant (Finegan, 1984). Primary succession is thecommunity formation process that begins onsubstrates that have never before supportedvegetation whereas secondary succession occurswhere residual soil and plants survive adisturbance. However, the distinction between thetwo is not always clear (Miles, 1979).
Clements (1916) restricted the concept ofsuccession to changes resulting from biologicalreaction causing replacement of one plantcommunity by another until, within a givenclimatic region, a climax was established. Thiscame to be known as facilitation and relay floristics(Miles, 1979). Facilitation is also synonymous withautogenic change (Miles, 1979), that is,environmental changes occurring during successionas a consequence of the presence of plants, asopposed to allogenic change, which is caused byfactors external to the vegetation.
Clements' model of succession was rejected bymany researchers as inapplicable or not whollyapplicable to observed vegetation change. Egler(1954) showed that an important factor in old fieldvegetation development was initial floristiccomposition. In contrast to the relay floristics
New Zealand Journal of Ecology 14:©New Zealand Ecological Society

60 NEW ZEALAND JOURNAL OF ECOLOGY, VOL. 14, 1990
emphasised by Clements, seed banks provide all thespecies destined to predominate at the start of thesuccession. In the early stages, the later dominantsare small and inconspicuous. One succeeding groupafter another dominates the physiognomy, oftenweakening or killing preceding dominants.
Connell and Slatyer (1977) classifiedsuccessional theories and outlined three alternativemodels of mechanisms that bring aboutsuccessional change after a perturbation, assumingno further changes in the abiotic environment. Inthe 'facilitation' (relay floristics) model, the early-succession species modify the environment so thatit is more suitable for later succession species toinvade and grow to maturity. This sequencecontinues until the resident species no longermodifies the site in ways that facilitate the invasionand growth of different species. In the tolerancemodel, the modifications wrought on theenvironment by earlier colonists neither increasenor reduce the rates of recruitment and growth tomaturity of later colonists. Species that appear laterare simply those that arrived either at the verybeginning or later and then grew slowly. Thesequence of species is determined solely by their lifehistory characteristics and tolerance ofenvironmental factors (e.g., shade tolerance). Theinhibition model holds that once earlier colonistssecure space and/or resources they inhibit invasion
of subsequent colonists or suppress the growth ofthose already present. The latter invade or growonly when the dominating residents are damaged orkilled, thus releasing resources.
Connell and Slatyer (1977) suggested there wasevidence for the first model mainly in relation toprimary successions, that there was little evidencepresently available to support the tolerance model,and a considerable body of evidence supporting theinhibition model. More recent work (e.g., Hils andVankat, 1982) has suggested that the tolerancemodel was only disfavoured because the right kindsof experiments had never been undertaken. Themajor dichotomy between the alternative models ofsuccession occurs between relay floristics and initialfloristic composition. Up to this point all themodels agree that certain species will usuallyappear first because they have evolved 'colonising'characteristics but they differ in the mechanismswhich determine how new species appear later inthe sequence.
Finegan (1984) noted that the different modelsof succession were not mutually exclusive. Heconsidered that neither the reductionist (e.g., initialfloristic composition) nor holistic (e.g., facilitation)theories of succession have produced models whichexplain field observations and that a syntheticapproach should be sought. Thus facilitation,tolerance, inhibition and allogenesis are
Figure 1: North Island showing localities mentioned in text.

CLARKSON: VEGETATION AFTER VOLCANIC DISTURBANCE
interdependent mechanisms and may affect thesame individual successively or simultaneouslyduring its life cycle. Further, the relativeimportance of various mechanisms is likely to varywidely between environments and attempts toclassify succession on the basis of a singleunderlying mechanism only obscure understandingand inhibit progress.
Tilman's (1985) attempt at an alternativesimple theory of succession focussed on resourcechanges during succession. According to hisresource-ratio hypothesis, succession results from agradient through time in the relative availabilitiesof limiting resources, in particular the inverserelationship between soil nutrient status and lightavailable at the soil surface. Each species is asuperior competitor at different points along thetemporal gradient. In addition, succession shouldbe directional or repeatable only to the extent thatthe resource supply trajectory is repeatable ordirectional.
Johnstone (1986) in reviewing invasion successpointed out that time was not a cause ofsuccession, as inferred by many studies. Rather,succession should be seen as the expression of aninteraction of species in an assemblage as a resultof invasion, maintenance and decline. Theseprocesses are stochastic in the sense that the causesfor the probability is system dependent, andbecause of the dynamic interaction of processes,inherently difficult to predict.
Pickett, Collins and Armesto (1987) clarifiedand expanded Connell and Slatyer's models byelaborating the mechanisms of succession involvedin each. Facilitation was seen as operating throughenhanced invasion, amelioration of environmentalstress or increase in resource availability. Tolerancewas shown to be able to be interpreted in twoways, active and passive. An example of activetolerance is the ability of an organism to endurelow resource levels. An example of passivetolerance is contrasting life histories (e.g., life spanleading to replacement of short-lived faster growingspecies by longer-lived slow growing species).Inhibition was also shown to be complex, gradinginto tolerance depending on whether the interactionis seen from the viewpoint of the incumbent or thechallenger. In attempting to generalise about themechanisms of replacement, Pickett et al. (1987)noted that the models were only useful whenrestricted to describing particular replacements ofspecies in a succession.
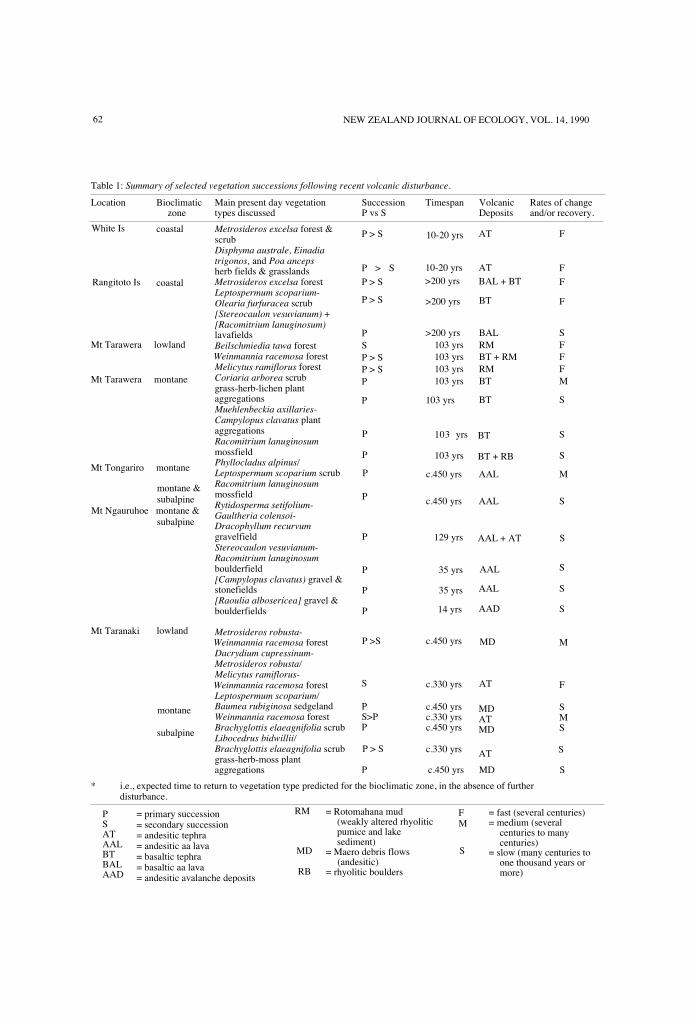
Select history of volcanic disturbanceand vegetation developmentSix areas are described as summarised in Table 1.Vegetation and substrate type names follow the
61
naming system of Atkinson (1985) but underliningto indicate cover classes of the leading dominants isnot used. Species names follow Allan (1961),Moore and Edgar (1970), Allison and Child (1971),Galloway (1985) and Connor and Edgar (1987).
White IslandWhite Island, a 238 ha andesite cone in the Bay ofPlenty, has been in a state of continuous solfataricactivity, with intermittent small steam and tephraeruptions since 1826 when the first recordedlanding was made by Europeans (Nairn and Wood,1986). Between December 1976 and December 1981the largest and longest eruptive episode on theisland in historic times occurred (Houghton et al.,1983), killing large areas of Metrosideros excelsaforests (B.J. Scott, pers. comm.). Prior to thisthere are reports of only minor vegetation damage,for example, loss of leaves by Metrosiderosfollowed by rapid recovery (parham, 1973).
The extant vegetation on the island comprisesa simple forest and scrub type entirely dominatedby Metrosideros excelsa, and herb fields andgrasslands associated with gannetries in which thedominant species are Disphyma australe subsp.australe, Einadia trigonos subsp. trigonos, and Poaanceps subsp. anceps.
A survey in 1986 (Clarkson, Clarkson andSmale 1989) showed that the extent of theMetrosideros forest and scrub (c. 48 ha) had beenreduced by more than one half by the recenteruptions. Only c. 5% of the Metrosideros trees inthe area badly affected by the eruptions survived,the result of resprouting from epicormic buds (Fig.2). A few carried seed capsules, evidently formedduring the December 1985 flowering season but noseedling or sapling Metrosideros were noted in thevicinity. At a site where all the Metrosideros treeswere dead, a single individual of the fernHistiopteris incisa had apparently survived. Theactual cause of death of the Metrosideros trees andshrubs was difficult to determine so long after theeruptions. However, as no evidence of blast orburn was seen, and ultimate branches remainedintact on the majority of individuals, the mostlikely causes were toxic fumes, wet ash coatingleaves and interfering with vital processes, and'acid rain'. Fumes were identified by earlierworkers (Oliver, 1915; Hamilton, 1959) as the mostlikely factor controlling the distribution and extentof Metrosideros forest and scrub. The vascularflora in the last 50 years has numbered at most 13species (Oliver, 1915; Hamilton, 1959; Wilcox,1967) but a recent survey (Clarkson et al., 1989)recorded only 7 species. I have found no publishedrecords of bryophytes, lichens or terrestrial algae.Species lost from the eruption-affected area were
62 NEW ZEALAND JOURNAL OF ECOLOGY, VOL. 14, 1990
Table 1: Summary of selected vegetation successions following recent volcanic disturbance.
Location Bioclimatic Main present day vegetation Succession Timespan Volcanic Rates of changezone types discussed P vs S Deposits and/or recovery.
coastal Metrosideros excelsa forest &scrubDisphyma australe, Einadiatrigonos, and Poa ancepsherb fields & grasslandsMetrosideros excelsa forestLeptospermum scoparium-Olearia furfuracea scrub[Stereocaulon vesuvianum) +[Racomitrium lanuginosum)lavafieldsBeilschmiedia tawa forestWeinmannia racemosa forestMelicytus ramiflorus forestCoriaria arborea scrubgrass-herb-lichen plantaggregationsMuehlenbeckia axillaries-Campylopus clavatus plantaggregationsRacomitrium lanuginosummossfieldPhyllocladus alpinus/Leptospermum scoparium scrub PRacomitrium lanuginosummossfieldRytidosperma setifolium-Gaultheria colensoi-Dracophyllum recurvumgravelfieldStereocaulon vesuvianum-Racomitrium lanuginosumboulderfield[Campylopus clavatus) gravel &stonefields[Raoulia albosericea] gravel &boulderfields
Metrosideros robusta-Weinmannia racemosa forestDacrydium cupressinum-Metrosideros robusta/Melicytus ramiflorus-Weinmannia racemosa forestLeptospermum scoparium/Baumea rubiginosa sedgelandWeinmannia racemosa forestBrachyglottis elaeagnifolia scrubLibocedrus bidwillii/Brachyglottis elaeagnifolia scrub P > Sgrass-herb-moss plantaggregations P c.450 yrs MD S
* i.e., expected time to return to vegetation type predicted for the bioclimatic zone, in the absence of furtherdisturbance.
= primary succession= secondary succession= andesitic tephra= andesitic aa lava= basaltic tephra= basaltic aa lava= andesitic avalanche deposits
White Is
Rangitoto Is coastal
Mt Tarawera lowland
Mt Tarawera montane
Mt Tongariro montane
montane &subalpine
Mt Ngauruhoe montane &subalpine
Mt Taranaki lowland
montane
subalpine
PSATAALBTBALAAD
= Rotomahana mud(weakly altered rhyoliticpumice and lakesediment)
= Maero debris flows(andesitic)
= rhyolitic boulders
P > S
P > S
P > S
P > S
PSP > SP > SP
P
P
P
P
P
P
P
P
P >S
S
PS>PP
RM
MD
RB
10-20 yrs
10-20 yrs>200 yrs
>200 yrs
>200 yrs103 yrs103 yrs103 yrs103 yrs
103 yrs
103 yrs
103 yrs
c.450 yrs
c.450 yrs
129 yrs
35 yrs
35 yrs
14 yrs
c.450 yrs
c.330 yrs
c.450 yrsc.330 yrsc.450 yrs
c.330 yrs
AT
ATBAL + BT
BT
BALRMBT + RMRMBT
BT
BT
BT + RB
AAL
AAL
AAL + AT
AAL
AAL
AAD
MD
AT
MDATMD
AT
F
F
F
F
SFFFM
S
S
S
M
S
S
S
S
S
M
F
SMS
S
= fast (several centuries)= medium (several
centuries to manycenturies)
= slow (many centuries toone thousand years ormore)
FM
S

CLARKSON: VEGETATION AFTER VOLCANIC DISTURBANCE 63
Figure 2: Metrosideros excelsa at old campsite Ohauora,
White Island showing dead crowns and recovery by
epicormic resprouts (1986).
mainly dicotyledonous herbs associated withgannetries, and burial by tephra was probably themain cause.
Mt TaraweraMt Tarawera, a series of coalescing low rhyolitedomes and an associated crater lake (LakeRotomahana) erupted in 1886, initiating primarysuccessions over c. 10 km2 and secondarysuccessions over a further c. 20 km2. Eye witnessaccounts of the eruptions and subsequent 100 yearsof vegetation change are available (Thomas, 1888;Aston, 1915; Turner, 1928; Burke, 1964; Nicholls,1963; Dickinson, 1980; Timmins, 1983; Clarksonand Clarkson, 1983).
Rates and patterns of vegetation change havebeen variable. At some of the most inhospitablesites on the peaks, such as those comprising largerhyolite boulders with a matrix of scoria, lichensand mosses have established but not floweringplants. On the unstable ash and lapilli slopes in thevicinity of Ruawahia trig (1111 m) plantaggregations, dominated by Muehlenbeckia axillarisand Campylopus clavatus and assumed to haveestablished within 50 years of the eruption, in a 15year period (1964-1979) doubled from 2.5 m2 to 4.5m2, and taxa increased from 13 to 19 (Clarksonand Clarkson, 1983). At other high altitude sites,life forms ranging from mosses to ferns andflowering plants were all present within 27 years ofthe eruption but in different microsites e.g., rockcrevices, tephra flats, hollows (Aston, 1915). At acrater site ('Crater 7') the total vascular floraincreased from 63 to 74 in fifteen years(1964-1979), with 28 taxa having established and 17being lost (Clarkson and Clarkson, 1983). A
Racomitrium /anuginosum mossfield adjacent tothe crater doubled in extent over the same period(Clarkson and Clarkson, 1983). On the extensivetephra flats of Plateau dome plant aggregationsdominated mainly by grasses, herbs, mosses,lichens and small shrubs with up to 35 taxa,including several hybrids, over the last 25 yearscoalesced into continuous mats of vegetation (Fig.3).
In the last 10 years taller shrubs, especiallyCoriaria arborea, which fIxes nitrogen throughsymbiosis with an actinomycete endophyte(Frankia) (Silvester, 1978), have entered theseaggregations and plants have now coalesced toform continuous scrub. During this transition up to50 taxa were present in some large aggregations butwith the development of continuous scrub and theconsequent loss of light-demanding taxa thenumber dropped to as few as 10. Change has beenmost rapid along a front just beyond the presentforest edge. In 70 years bare surfaces progressed tograss-herb-lichen ring growths (Burke, 1964).Subsequently Coriaria scrub has developed in 15years (1964-1979) (Clarkson and Clarkson, 1983).Seedlings and saplings of the forest treesWeinmannia racemosa and Griselinia littoralis hadestablished in this scrub type before 1979.Unusually severe winter frosts in 1982 caused someCoriaria to die back, releasing some saplings ofWeinmannia and Griselinia the following year. Thesequence described above was considerablycontracted in time closer to the interface between aprimary surface and sites where forest survived theeruption, low Weinmannia or Kunzea ericoidesforest having been in place more than 50 years.
Figure 3: Plant aggregations, comprising mainly mosses,
lichens and grasses with shrubs of Dracophyllumsubulatum, on Plateau dome (944 m a.s.l.), Mt
Tarawera; Conaria arborea shrubs are beginning to
invade (1979).

64 NEW ZEALAND JOURNAL OF ECOLOGY, VOL. 14, 1990
One pocket of forest within 1.5 km of the nearestcrater has trees of six different canopy specieswhich survived the eruption. However, it issurrounded by stands of Weinmannia into whichthere is little or no spread of the other potentialcanopy dominants.
Nearly all taxa present on dome tops haveprobably arrived from sources within the RotoruaLakes District, through propagules dispersed bywind or birds from a range of unforested sites suchas 'frost flats', mire margins or rock outcrops, orfrom vegetation disturbed at an earlier time. Likelyexceptions include Helichrysum sp. unnamed ('H.alpinum' of Cockayne, 1928), Raoulia sp. unnamed(aff. R. australis), Cassinia vauvilliersii, Epilobiumglabellum and E. melanocaulon, all with winddispersed seed and otherwise known only fromlocalities more than 50 km away.
Where Rotomahana mud was thickly depositedon slopes, as in parts of Lake Tarawera ScenicReserve, successions have led to Melicytusramiflorus dominance and Weinmannia is eitherabsent or rare. Where forest was only partlydamaged, trees such as Weinmannia andBeilschmiedia tawa resprouted from epicormic budsand some even flowered within a year of theeruption (Aston, 1915). Outside the forests the fernPteridium esculentum penetrated wherever theRotomahana mud was less than 0.6 m thick(Nicholls, 1963). Remnants of Beilschmiedi-dominant forest, covering 10-30 ha, representingmany survivors of the Tarawera eruption, have anative vascular flora of 60-80 taxa, similar innumber to stands unaffected by the eruption.However, in the volcanically-disturbed forest,Litsea calicaris, Knightia excelsa and Weinmanniaare more common in the canopy, Metrosiderosrobusta and Dacrydium cupressinum less so, andtree height and basal area are significantly lower.
The relatively even-aged Weinmannia forestswhich developed on the southern slopes of MtTarawera after the eruption have the potential forcohort senescence and die back. Assuming alifespan of c. 300 years (Beveridge, 1973), diebackcould be expected to occur about 2200 AD.Perhaps then, potential canopy species, representednow by saplings and seedlings will have theiropportunity to reach the canopy. Thus at higheraltitude there may be a return of Podocarpus halliiWeinmannia forest which existed before theeruption, as evidenced by the many deadPodocarpus spars. And at lower altitude, Nestegislanceolata, Beilschmiedia and Litsea should benefitbut there is only limited indication yet thatDacrydium or other podocarps might becomeprominent (Burke, 1974).
Mt TaranakiOn this west coast North Island andesite strato-volcano (2518 m), tephra eruptions and associateddebris flows and fires over approximately the last450 years have destroyed c. 6700 ha of vegetationand damaged an equivalent area (Druce, 1976).
Pollen analysis (McGlone, Neall and Clarkson,1988) suggests that areas of upper montane forestcompletely destroyed or badly damaged by theBurrell eruptions of 1655 AD (Druce, 1966) haveshown a succession from shrubs, including Coriariaarborea, to Kunzea and Fuchsia excorticata, andthen to Weinmannia forest. However, Libocedrusbidwillii has not, as yet, regained lost ground,being found as scattered survivors over a lowcanopy of broadleaved shrubs such as Brachyglottiselaeagnifolia (Fig. 4). The treeline and upperaltitudinal limits of tree species are generally loweron Mt Taranaki than on the adjacent, less-affectedPouakai Range (Clarkson, 1981). Upper montaneforest stands today are still characterised by theextreme dominance of Weinmannia and a vascularflora of c. 40 taxa (probably fewer than in thetransitional stages some 100-150 years ago). In thelowland forest zone, where damage was less severe,many Weinmannia resprouted from epicormic budsand gained dominance within 50 years of theeruption except in the sector between WaingongoroRiver and Kaupokonui Stream. Here thecombination of minor damage and a dressing ofvery fine ash, raised soil nutrients to a level thatenabled Melicytus to become very prominent(Druce, 1964). A topographic pattern of Melicytusdominance on flat to moderate slopes, where ash isretained or accumulated, and Weinmannia onsteeper slopes, where it has been lost, remains tothis day (Clarkson, 1981).
Figure 4: Pre-Burrell eruption Libocedrus bidwilliiemergent over broadleaved scrub beside StratfordMountain Road (1035 m a.s.l.), Mt Taranaki (1990).

65
On the debris fans formed by Maero debrisflows following the Newall eruptions of c. 1550 AD(Druce, 1966), the forest upper limit is depressedby 300 m. A lowered upper limit of the forest treeBeilschmiedia also occurs. On the lower portion ofthese fans Metrosideros, which establishedterrestrially along with more slender Weinmannia,make up the bulk of the forest. Kunzea isprominent on sites affected by even more recent (c.100 yr) Kahui flows. Above the forest limit thedebris fans have been colonised by moss fields ofRacomitrium lanuginosum and R. ptycophyllum.In these mossfields, circular aggregations of therhizomatous shrubs Coriaria pteridoides and C.plumosa, which have nitrogen-fixing symbionts,along with Chionochloa mbra and up to 15subalpine or alpine grass and herb taxa haveestablished (Fig. 5) (Clarkson, 1986). Similaraggregations of shrub species also occur at loweraltitude on these debris fans; there are places wherethese have coalesced to produce a continuousscrub. Elsewhere 100 year old 'crops' ofBrachyglottis elaeagnifolia occur with no obviousevidence of aggregation. On moderately slopingportions of the fans, where drainage is impeded,simple mire types comprising mainly the shrubLeptospermum scoparium and the sedge Baumeambiginosa have developed.
Figure 5: Chionochloa rubra (centre) and Coriariapteridoides and/or Coriaria plumosa (margins)
aggregations in a matrix of Racomitrium lanuginosummossfield on the western slopes (1066 m a.s.l.) of Mt
Taranaki (1982).
Rangitoto IslandOn Rangitoto Island, a basalt shield volcano, lava
flows and cone building tephra eruptions provided
some 20 km2 of primary surface for plants tocolonise. According to Robertson (1986), the
volcano appears to have been active between 850
AD and 1800 AD with a maximum lava flow
activity around 1300 AD. However, Clark (1989)suggests that it is unlikely any eruption occurredafter 1500 AD. The primary surface is mainlycrusty (aa) basaltic lava but the summit consists ofscoria cones. The last lava flows were followed byan emission of ash which filled most of the crevicesand shallow hollows (Millener, 1979).
Millener (1953; 1979) and Atkinson (1960)have described the vegetation in detail. On themost inhospitable sites only lichens, mosses(including R. lanuginosum) and blue-green algaeare present and forest is not expected to developfor hundreds, probably thousands, of years. Onthe other lava fields this sequence is being by-passed by the progressive establishment ofaggregations of higher vegetation, usually centeredaround the prime rock coloniser, Metrosiderosexcelsa (Fig. 6). In places these have coalesced toform continuous forest. In the more scrubby opengrowth, Leptospermum scoparium, Stypheliafasciculata, Hebe stricta var. stricta, Oleariafurfuracea and Coriaria arborea are prominent. Onthe scoria cones, Leptospermum and Olearia aremost abundant and the presence of emergentMetrosideros excelsa and Knightia excelsa indicatesthe early stages of forest development.
The indigenous vascular flora of the islandexceeds 210 taxa (R.O. Gardner, 1987, unpublishedspecies list) and of great interest is the range andabundance of Metrosideros excelsa x M. robustahybrids (Cooper, 1954).
Millener (1979) claimed that, as the oldest treeshe aged were <200 years and a radiocarbon dateof wood buried beneath the lava gave 225 years BP± 110, the development of Metrosideros excelsaforest was initially rapid. The first trees establishedin the ash accumulations and quickly spread overthe island last century, slowing down as favourablehabitats became fewer. However, Atkinson (1960)
Figure 6: Metrosideros excelsa aggregations on Rangitoto
Island basalt aa lava (1990).

66 NEW ZEALAND JOURNAL OF ECOLOGY, VOL. 14, 1990
speculated on the possibility that a period of450-500 years may have been available for plantcolonisation following the last eruption and Clark(1989) recently rejected the radiocarbon date citedby Millener, as possibly from a tree-root post-dating the lava. Further research on the vegetationis required to resolve what Atkinson termed theproblem of Rangitoto.
In forest on much older basaltic lava blocks onthe eastern flank of Mt Eden, Metrosideros exce/sashares dominance with Vitex lucens, Litseacalicaris, Dysoxylum spectabile, Alectryon excelsusand Myoporum laetum (Millener, 1979) and thismight indicate future trends.
Mt Ngauruhoe and Mt TongariroLava flows, pyroclastic flows and debris flowsfrom Mt Ngauruhoe (Fig. 7) and Mt Tongariro,andesite central North Island cones reaching 2291m and 1968 m respectively, have overwhelmedsome 12 km2 of vegetation between c. 1550 ADand the present day.
Prehistoric lava flows in the MangatepopoValley support mainly Dracophyllum recurvum-
Rytidosperma setifolium/Campylopus clavatus
shrubland with up to 50 taxa present overall.
Figure 7: Mt Ngauruhoe showing prehistoric lava flows
(foreground), 1949-54 lava flows (middleground) and
1975 pyroclastic flows on upper slopes (1987).
The c. 450 year old Te Mari flow (Topping,1974) on the northern slopes of Tongariro extendsfrom 1400 m to 800 m a.s.l. Adjacent unaffectedvegetation is, in ascending order, Podocarpus halliiforest, Phyllocladus alpinus / Leptospermumscoparium scrub and Chionoch/oarubra/Dracophyllum recurvum tussockland(Atkinson, 1981). The lower one-third of the flowhas returned to tall P. alpinus/L.. scoparium scrubcontaining the probable future forest dominant P.hallii (Atkinson, 1981) while the upper two-thirdssupports only sparse vegetation dominated by
Racomitrium /anuginosum although with morethan 30 other taxa present (I.A.E. Atkinson andC.C. Ogle in Uhe, 1972).
At 1675 m, Ngauruhoe lava flows erupted in1870 support a floristically poor (c. 30 taxa)vegetation with 25-390/0 cover, the main speciesbeing Rytidosperma setijolium, Gaultheria colensoiand D. recurvum. The grasses, herbs and shrubsgrow mainly in ash or gravel overlying the lavawith mosses and lichens present on the lavaboulders. There has been no detectable change incover or composition since Uhe (1972) sampledthese flows in 1966.
Flows erupted in 1949 and 1954 (Gregg, 1960)and extending in a westerly and northwesterlydirection from 2070 m to 1324 m a.s.l., arefloristically poorer with 4 to II taxa, depending onaltitude and the proportions of boulders, stones,gravel and ash present. The most extensive surface,comprising boulder-sized lava blocks, has thepoorest type, Stereocaulon vesuvianum-Racomitriumlanuginosum boulderfield, which has cover valuesexceeding 50%. Sites with higher proportions ofgravel, stones and ash, that accumulated after thelava was emplaced, are mainly [Campylopusclavatus] gravel or stonefields. Although supportingmore species, including Rytidosperma setifoliumand Raoulia albosericea, cover does not exceed33%. In 1966 Uhe (1972) noted only R.lanuginosum and S. vesuvianum on the 1954 lavablocks so there has been a significant increase inthe number of species and cover of vegetation inthe intervening 21 years but rates of change are stillextremely slow.
From the outset the pyroclastic flows of 1975(Nairn and Self, 1978) comprised a greater mixtureof different sized particles, and subsequent debrisflows have continued to add to this. They can beclassified mainly as [Raoulia albosericea] gravel orboulderfields and have 6-10 taxa. The faster rateand concurrent establishment of differenttaxonomic groups, highlights the importance of thenature of the ejecta for plant colonisation andestablishment.
Only on the lower portion of the Te Mari flowhas there been sequential replacement of onespecies group by another. In contrast, on the otherflows, although species have established either atdifferent times or concurrently, albeit on differentmicrosites, there has been a gradual accumulationof species.
InterpretationPlant successions on volcanic substrates areextremely variable both in terms of rates of changeand the sequence of species establishing. Allowingfor bioclimatic zonal differences, the variability is

CLARKSON: VEGETATION AFTER VOLCANIC DISTURBANCE 67
explicable in terms of a gradient of scale andseverity of disturbance, and of suitability of thesubstrate emplaced for plant establishment andgrowth. On relatively homogeneous, excessivelydrained or exposed habitats, resulting fromdisturbance so severe that no plants or dormantpropagules survive, the slow 'classical' successionbeginning with lichens and mosses is evident.
Where the severity is as great but there is lessuniformity in the surface emplaced, with differentmicrosites available, especially ash accumulations,all taxonomic groups of plants establish more orless concurrently. Most of the 'soils' availablefollowing extensive severe disturbance are deficientin nutrients, particularly phosphorus and nitrogen,and have very low water-holding capacity (Uhe,1972).
Nitrogen-fixers, including lichens and Coriariaspp. may significantly alter soils or other aspects ofthe environment, making it more suitable for latersuccessional species to invade or grow to maturity.Plant aggregations that later coalesce intocontinuous cover are usually a feature of thesesuccessions. The species involved are generallyshade-intolerant, tolerant of low nutrient levels orpossess efficient nutrient uptake systems throughmycorrhizal associations (Uhe, 1972). They alsotend to be short lived and have propagules that arereadily dispersed by wind or birds. Dispersal ismainly from nearby open sites or vegetationrecovering from earlier eruptions. The creation ofopen habitat (sensu Grant, 1981) allows abundantoverlap of taxonomically related species, whichrarely or never occurs in later successional stages.Thus hybrids and hybrid swarms are relativelycommon. On soils low in nitrogen but withsufficient phosphorus and bases to allow Coriariaspp. to establish and grow vigorously (Vucetich andWells, 1982), succession rates are faster, e.g.,Rotomahana mud and Burrell tephra, than on soilswhere Coriaria spp. establishment is delayed, e.g.,Tarawera lapilli.
Where vegetation is severely damaged but newsurfaces are discontinuous or where new materialforms a thin veneer, recovery can involveestablishment of species ranging from thosecharacteristic of primary successions to those fromexisting 'mature' vegetation. Seed banks, short-distance dispersal, sapling release and vegetativeregrowth may all be involved. Decay of plantskilled by the disturbance may provide an initialnutrient source to early colonisers and, dependingon rooting depths, tephra depths and penetrability,buried soils mayor may not be available. Severalof the tall forest trees have attributes which enablethem to recover from damage rapidly or reoccupybared ground quickly. Weinmannia and
Metrosideros produce abundant, light, wind-dispersed seed from an early age, and can coppiceafter damage. Beilschmiedia also coppices but hasmore limited seed dispersal and is less competitiveon thick tephras. Litsea and Melicytus areapparently intermediate in their ability to cope withvolcanic disturbance, with the latter requiring arelatively fertile substrate.
Finally, in situations where damage is onlyslight, little change may occur in the compositionor structure of the vegetation. Frequently a smallcanopy gap or margin may be closed by vegetativeexpansion or release of an existing juvenile.
Application of successional modelsThe examples of vegetation succession outlinedillustrate many elements of successional models ortheories described earlier. At one extreme someprimary successions, for example, on the dometops of Tarawera, have elements of facilitation,autogenic change, and relay floristics. Changes inthe composition of plant aggregations seem toinvolve a range of models including facilitation andinhibition but the replacement is not deterministic.That is, when dieback occurred of plants within theTarawera Plateau dome aggregations, the resultingspace could be occupied by either new species orspecies already present. Alternatively, new speciesestablished even in the absence of dieback of partsof the plant aggregation.
The tolerance model is applicable whenconsidering early establishment of plantaggregations as the species are able to tolerate lowsubstrate resource levels and full exposure to light.The facilitation model is applicable where theaggregations modify the substrate in any way whichenables new species to colonise. For example,through stabilisation of tephra on slopes ortrapping of 'fines', improving the water holdingcapacity of the substrate or increasing the nitrogenstatus of soils.
The development of Coriaria scrub increasesthe nitrogen status of the soils and changes themicroclimate and light characteristics of the 'soil'surface. The rapid change in the flora which resultsfrom Coriaria dominance and shading out of theplant aggregations is consistent with relay floristics.Seedlings of forest trees Weinmannia and Griseliniaestablish preferentially beneath the Coriaria(facilitation); the change from Coriaria scrub toWeinmannia-Griselinia forest however arises inseveral ways. The checking of Coriaria caused byfrosting and the subsequent release of saplings ofWeinmannia and Griselinia fits the inhibitionmodel. When Weinmannia or Griselinia overtophealthy Coriaria they are able to do so becausethey can, as seedlings or saplings, tolerate the

68 NEW ZEALAND JOURNAL OF ECOLOGY, VOL 14, 1990
degree of shading characterisitic of a Coriariacanopy. When Weinmannia or Griselinia overtop anaturally senescent Coriaria canopy the inhibitionmodel is applicable.
Successions with partial disturbance areespecially difficult to fit into successional models.On Mt Tarawera and Mt Taranaki where forestcanopies were partly killed, neither relay floristicsnor initial floristics encompass the resulting mosaicvegetation pattern. Some existing canopy treesrecovered, some undergrowth species were releasedand other species colonised the sites. Separatingexamples of tolerance and inhibition seems almostimpossible in some instances. On Mt Tarawera andMt Taranaki most canopies dominated by Kunzeaseem to offer little resistance to invasion byWeinmannia. In contrast, Beilschmiedia seedlingsare both suppressed by, and tolerant of, the lowerlight levels under the earlier establishedWeinmannia canopy. In successions on the lavaflows of Rangitoto Island and Mt Ngauruhoe,allogenic change through the accumulation of ashor lapilli after the lava is emplaced significantlyaffects the composition and rate of speciesinvasion. On White Island the very depauperateflora, including the single pioneer tree speciesMetrosideros excelsa, ensures that the relayfloristics model cannot apply at allover the periodstudied. Metrosideros becomes dominant early byrecovery from the disturbance, or later bycolonisation of the new surface. This successioncould be considered as an example of the tolerancemodel but the term 'direct succession' as used byWhittaker and Levin (1977) appears moreappropriate.
In short, these examples support Finegan's(1984) conclusion that facilitation, tolerance,inhibition and aIlogenesis are interdependentmechanisms in succession and may affect thesame individual successively or simultaneouslyduring its life cycle. They also highlight the needto consider specific replacements rather thanwhole pathways as outlined by Pickett et al.(1987).
The recently proposed resource-ratiohypothesis of succession (Tilman, 1985) can beused to explain the general pattern of thesuccessions described without the need to categorisethem. As outlined earlier, succession results from agradient through time of the relative availabilitiesof limiting resources, in particular the inverserelationship between soil nutrient status andirradiance levels at the soil surface. Different scalesor severities of volcanic disturbance initiatedifferent resource supply trajectories and eachspecies is a superior competitor at different pointsalong such temporal gradients. As predicted by thehypothesis, the same qualitative separation and
ordering of species along spatial resource gradientsis apparent. However, Tilman recognises but doesnot incorporate into his scheme other importantprocesses that could be involved such as species-specific herbivory, differential colonisation abilitiesand temperature-dependent growth. Chance factorsmay also intervene. Nevertheless, the model itstands provides a simple way of organising muchvariable information about vegetation changefollowing volcanic disturbance.
Following the recommendations of the authorscited, prediction of volcanic disturbancesuccesssions will require knowledge of resourcesupply trajectories, the potential flora, the lifehistories and attributes of the major species, andquantification of the probabilities of the process ofinvasion, maintenance and decline. Generalisedpredictions for relatively short time frames can bemade at present but detailed predictions are notpossible. No experimentation or manipulation hasbeen carried out to test which are the mostsignificant contributing processes or conditionscausing succession, or to differentiate betweensuccessional models.
Concluding commentsFor the volcanic sites studied, the return period forvolcanic disturbance varies from less than once tomore than ten times per 500 years, that is, less thanthe life span of many late successional tree speciesof the central North Island. Thus, speciesdominant elsewhere in the same region which arecharacteristically absent or severely reduced arethose of late successional vegetation, example ofwhich have been given earlier. Although area forarea, volcanically disturbed vegetation may have alarger flora because of the vegetation mosaicspresent, when particular plant communities ondifferent volcanoes are compared they may besignificantly depauperate or sometimes entirelyabsent. Thus, the Metrosideros forest on WhiteIsland has only a fragment of the flora of the sametype on Rangitoto Island, which in turn isfloristically poorer than the type on Hauturu (LittleBarrier Island) (see Hamilton and Atkinson, 1961).The Chionochloa tussockland and herb field zoneson Mt Taranaki lack many species (Druce, 1974)and these types are all but absent from equivalentaltitudes on Mt Ngauruhoe.
On White Island and Mt Ngauruhoe, at least,the disturbance regime could be viewed asproducing a shifting-mosaic steady-state (sensuSousa, 1984). The patchy and asynchronous natureof the disturbance results in a continuouslychanging mosaic of patches of different sizes andages which average to an equilibrium pattern whenone considers the mean dynamics of an area

69CLARKSON: VEGETATION AFTER VOLCANIC DISTURBANCE
containing such patches. Fleming's (1963) tentativeidentifications of fossil Metrosideros, Histiopterisand Phormium suggest Metrosideros forests havebeen waxing and waning on White Island for manycenturies. In contrast, Weinmannia abundance inMt Taranaki forests apparently coincides only withrecent eruptions (McGlone et al., 1988).
Many genera and species are common to thevolcanic successions outlined but there is noexclusive volcanic succession flora; the sameattributes ensure success in the face of other typesof disturbance, especially fire. Indeed, the rangeand abundance of many of the same species wasconsiderably expanded in periods of Polynesianand later European burning (McGlone, 1983) andfire sometimes is caused by volcanic disturbance(Cooper, 1949; Druce, 1976). Recent volcanicdisturbance has shaped most species distributionaland abundance patterns in a large portion of theNorth Island. Other consequences are the frequentoccurrence of hybrids and possibly the low numberof regionally endemic species.'
Too little is known about many aspects of theprocesses directly and indirectly controllingsuccession. Long term quantitative studies ofvegetation change and quantitative characterisationof associated disturbance events will be needed toenter the realm of detailed prediction (Miles, 1979;Sousa, 1984). Prediction will be probabilistic ratherthan deterministic or mechanistic (Egler, 1942;Johnstone, 1986). Long term monitoring will alsobe necessary to detect the rare event (for example,the frosting of Coriaria oil Mt Tarawera) whichmay shape vegetation pattern for decades orcenturies (Miles, 1987).
Experimental and manipulative studies are nowneeded to test which are the most significantprocesses or conditions causing succession and todifferentiate between successional models.
AcknowledgementsBeverley Clarkson, Paula Broekhuizen and JohnNicholls assisted in the field. Ian Atkinson, ColinBurrows, John Nicholls, Trevor Partridge andPeter Wardle provided valuable comments onearlier drafts. Murray Boase supplied the slides forFigures 3 and 5. Bill Lee and an anonymous refereemade valuable comments on the manuscript.
ReferencesAllan, H.H. 1961. Flora of New Zealand, Vol. 1.
Government Printer, Wellington.Allison, K.W.; Child, J. 1971. The mosses of New
Zealand. University of Otago Press, Dunedin.
Aston, B.C. 1915. The Vegetation of the Taraweramountains, New Zealand. Royal Society ofNew Zealand Transactions and Proceedings48: 304-314.
Atkinson, I.A.E. 1960. Forest vegetation of theinner islands of the Hauraki Gulf. Proceedingsof the New Zealand Ecological Society 7: 29-33.
Atkinson, I.A.E. 1981. Vegetation map ofTongariro National Park, North Island, NewZealand. Scale 1:50,000. Department ofScientific and Industrial Research, Wellington.
Atkinson, I.A.E. 1985. Derivation of vegetationmapping units for an ecological survey ofTongariro National Park, North Island, NewZealand. New Zealand Journal of Botany 23:361-378.
Beveridge, A.E. 1973. Regeneration of podocarpsin a central North Island forest. New ZealandJournal of Forestry 18: 23-35.
Burke, W.D. 1964 (unpublished). A study of thevegetation of a part of Mt Tarawera. M.Sc.Thesis, Victoria University of Wellington,Wellington.
Burke, W.D. 1974. Regeneration of podocarps onMt Tarawera, Rotorua. New Zealand Journalof Botany 12: 9-26.
Clark, R.H. 1989. New Zealand from the road:landforms of the North Island. HeinemannReed, Auckland.
Clarkson, B.D. 1981 (unpublished). Vegetationstudies in the Taranaki Land District, NewZealand. D.Phil. Thesis, University ofWaikato, Hamilton.
Clarkson, B.D. 1986. Vegetation of EgmontNational Park, New Zealand. National ParksScientific Series 5. Science InformationPublishing Centre, Wellington.
Clarkson, B.D.; Clarkson, B.R.; Smale, M.C.1989. Vegetation destruction and recoveryfollowing recent eruptions on White Island(Whakaari), Bay of Plenty, New Zealand. In:New Zealand Geological Survey Bulletin 103,pp. 137-138. New Zealand Geological Survey,Lower Hutt.
Clarkson, B.R.; Clarkson, B.D. ]983. MtTarawera. 2. Rates of change in the vegetationand flora of the high domes. New ZealandJournal of Ecology 6: 107-119.
Clements, F.E. 1916. Plant succession. CarnegieInstitute, Washington.
Cockayne, L. 1928. The vegetation of NewZealand. 2nd ed. Englemann, Leipzig.
Cole, J.W.; Nairn, LA. 1975. Catalogue of theactive volcanoes of the World includingsolfatara fields. Part 22: New Zealand.International Association of Volcanology andChemistry of the Earths Interior, Rome, Italy.

70 NEW ZEALAND JOURNAL OF ECOLOGY, VOL. 14, 1990
Connell, R.H. and Slatyer, R.O. 1977. Mechanismsof succession in natural communities and theirrole in community stability and organization.The American Naturalist 111: 1119-1144.
Connor, H.E.; Edgar, E. 1987. Name changes inthe indigenous New Zealand flora, 1960-1986and Nomina Nova IV, 1983-1986. NewZealand Journal of Botany 25: 115-170.
Cooper, R.C. 1949. The effect of the recenteruption on the plants of Ngauruhoe. Recordsof the Auckland Institute and Museum 3:381-385.
Cooper, R.C. 1954. Pohutukawa x Rata. Variationin Metrosideros (Myrtaceae) on RangitotoIsland, New Zealand. Records of theAuckland Institute and Museum 4: 205-212.
Dickinson, B.R. 1980 (unpublished). Flora andvegetation of the high domes of Mt Tarawera.M.Sc. Thesis, University of Waikato,Hamilton.
Druce, A.P. 1964. The vegetation. In: Scanlan,A.B. (Editor), Egmont National ParkHandbook, 2nd ed, pp 51-55. New Plymouth,Egmont National Park Board.
Druce, A.P. 1966. Tree-ring dating of recent ashand lapilli, Mt Egmont. New Zealand Journalof Botany 4: 2-41.
Druce, A.P. 1974. Some notes on the flora of MtEgmont. Auckland Botanical SocietyNewsletter 31: 1-4.
Druce, A.P. 1976. Egmont botanical studies: In:National Parks Series 6, pp 39-50. NationalParks Authority, Wellington.
Egler, F.E. 1942. Vegetation as an object of study.Philosophy of Science 9: 245-260.
Egler, F.E. 1954. Vegetation Science concepts. LInitial floristic composition - a factor in old-field vegetation development. Vegetatio 4:412-417.
Finegan, B. 1984. Forest succession. Nature 312:109-114.
Fleming, C.A. 1963. Plant fossils from WhiteIsland. New Zealand Journal of Geology andGeophysics 6: 705-706.
Galloway, D.J. 1985. Flora of New Zealand:Lichens. Government Printer, Wellington.
Grant, V. 1981. Plant speciation. ed. 2. ColumbiaUniversity Press, New York.
Gregg, D.R. 1960. The geology of Tongariro sub-division. New Zealand Geological SurveyBulletin 40. New Zealand Geological Survey,Wellington.
Hamilton, W.M. 1959. Vegetation. In: Hamilton,W.M.; Baumgart, LL. compilers, WhiteIsland, pp 62-69. D.S.I.R. Bulletin 127.Government Printer, Wellington.
Hamilton, W.M.; Atkinson, I.A. 1961. Vegetation.In: Little Barrier Island (Hauturu). D.S.1.R.Bulletin 137, pp 87-121. Government Printer,Wellington.
Hils, M.H.; Vankat, J.L 1982. Species removalsfrom a first-year old-field plant community.Ecology 63: 705-711.
Houghton, B.F.; Scott, B.J.; Nairn, I.A.; Wood,C.P. 1983. Cyclic variation in eruptionproducts, White Island volcano, New Zealand1976-1979 (Note). New Zealand Journal ofGeology and Geophysics 26: 213-216.
Johnstone, I.M. 1986. Plant invasion windows: atime based classification of invasion potential.Biological Reviews of the CambridgePhilosophical Society 61: 369-394.
McGlone, M.S. 1983. Polynesian deforestation ofNew Zealand: a preliminary synthesis.Archaeology in Oceania 18: 11-25.
McGlone, M.S.; Neall, V.E.; Clarkson, B.D. 1988.The effect of recent volcanic events andclimatic changes on the vegetation of MtEgmont (Mt Taranaki), New Zealand. NewZealand Journal of Botany 26: 123-144.
Miles, J. 1979. Vegetation dynamics. Chapman andHall, London.
Miles, J. 1987. 1. Vegetation succession: past andpresent perceptions. In: Gray, A.J.; Crawley,M.J.; Edwards, P.J. (Editors), Colonisationsuccession and stability. Blackwell ScientificPublishers, Oxford.
Millener, L.M. 1953. How old is the vegetation onRangitoto Island? Report of 2nd New ZealandEcological Society Annual Meeting: 17-18.
Millener, L.M. 1979. Forest, scrub and fresh-water communities. In: Brook, P.J. (Editor),Natural history of Auckland: an introduction,pp. 35-46. Auckland Institute and Museum,Auckland.
Moore, L.B., Edgar, E. 1970. Flora of NewZealand, Vol. 2. Government Printer,Wellington.
Nairn, I.A. and Self, S. 1978. Explosive eruptionsand pyroclastic avalanches from Ngauruhoe inFebruary 1975. Journal of Volcanology andGeothermal Research 3: 39-60.
Nairn, I.A.; Wood, C.P. 1986 (unpublished).International Volcanological Congress, NewZealand, 1986. Tour Guide C6: activevolcanoes of Taupo Volcanic Zone. NewZealand Geological Survey, D.S.I.R.,Rotorua.
Nicholls, J.L 1963. Volcanicity and indigenousvegetation in the Rotorua district. Proceedingsof the New Zealand Ecological Society 10:58-65.

CLARKSON: VEGETATION AFTER VOLCANIC DISTURBANCE 71
Oliver, W.R.B. 1915. The vegetation of WhiteIsland, New Zealand. Journal of the LinnaeanSociety of London, Botany 43: 41-47.
Parham, W.T. 1973. Island volcano: White IslandNew Zealand. Collins, Auckland.Pickett, S.T.A.; Collins, S.L.; Armesto, J.J. 1987.
Models, mechanisms and pathways ofsuccession. The Botanical Review 53: 334-371.
Robertson, D.J. 1986. A paleomagnetic study ofRangitoto Island, Auckland, New Zealand.New Zealand Journal of Geology andGeophysics 29: 405-411.
Silvester, W.B. 1978. Ecological and economicsignificance of the non-legume symbioses. In:Newton, W.E. and Nyman, C.J. (Editors),Nitrogen fixation. Proceedings of the firstinternational symposium, pp. 489-506.Washington State University Press, Pullman,Washington.
Sousa, W.P. 1984. The role of disturbance innatural communities. Annual Review ofEcology and Systematics 15: 353-39l.
Thomas, A.P.W. 1888. Report on the eruption ofTarawera and Rotomahana. New Zealand.Government Printer, Wellington.
Tilman, D. 1985. The resource-ratio hypothesis ofplant succession. The American Naturalist 125:827-852.
Timmins, S.M. 1983. Mt Tarawera. l. Vegetationtypes and successions. New Zealand Journal ofEcology 6: 99-105.
Topping, W.W. 1974 (unpublished). Some aspectsof Quaternary history of Tongariro VolcanicCentre. Ph.D. Thesis, Victoria University ofWellington, Wellington.
Turner, E.P. 1928. A brief account of the re-establishment of vegetation of Taraweramountain since the eruption of 1886. RoyalSociety of New Zealand Transactions andProceedings 59: 60-66.
Uhe, G. 1972 (unpublished). A contribution to theecology of vegetation on recent volcanicdeposits. Ph.D. Thesis, University ofAuckland, Auckland.
Vucetich, C.G. and Wells, N. 1982. Soils,agriculture and forestry of the Waiotapuregion, central North Island, New Zealand.Soil Bureau Bulletin 31. Department ofScientific and Industrial Research, Wellington.
Whittaker, R.H.; Levin, S.A. 1977. The role ofmosaic phenomena in natural communities.Theoretical Population Biology 12: 117-139.
Wilcox, M.D. 1967. Auckland Botanical SocietyNewsletter, April 1967: 6-7.





























