Embed Size (px)
Citation preview

A Public Antigenic Dete, minant in the HLA-B5 Cross-Reacting Group-- A Basis for Cross-Reactivity and a Possible Link with Behcet's Disease
Benjamin D. Schwartz, Lisa K. Luehrman, John Lee, and Glenn E. Rodey
ABSTRACT: Serologic cross-reactiriO, among allelic gene products commonly occurs in the HLA complex. but the molecular basis of these serologfi" phenomena is incompletely characterized. Because of strong cross-reactirity among antigens comprising the B5 cross-reactire group ¢ i.e.. HLA-B5. B15. BIB. and Bw35/, we initiated a study of the chemical basis of cross-reactirity among this gronp of antigens. Using classfi" serologic procedures, an *~';l-Pr6tein A binding assay, and chemical im- munoprecipitation techniques, we hare defined a new antigenfi" determinant, tentatirely desig- nated "Y," which is present on certain HLA-B molecules, By a series of sequential immuno- precipitation experiments, Y was shown to be a "public" antigenic determin.:~nt distinct from the "prirate" determinants B5, BI5, BlS, and Bw35, but present on the same 44.000 dalton glycoprotei~ molecules. Although B5 is most highly associated with Behcet's disease, other members of the B5 cross-reactire group hare aho been associated with Behcet's. albeit to a lesser extent. These associations suggest that determinant Y may p/ay a role in predisposition to Behcet's disease.
ABBREVi ~.TIONS /3,~p. /3~-microglobulin NP-40 Nonidet P-40 CYNAP cytotoxicity negative, absorp- PAGE polyacrylamide gel electro-
tion positive phoresis lgG immunoglobulin G SaCI Staphyloco~'us aureus Cowan ! 2-ME 2.mercaptoethanol strain NHIgG normal human immunogk:b- TBA Tris-buffered saline, pH v.4
ulin G TSN Tris-buffered saline contain- NHS normal human serum ing 0.259f NP-40
I N T R O D U C T I O N
The H L A antigenic determinants have been define~l as a result of the analysis o f the reaction patterns of different alloantisera on panels of human cells [ 1 -3 ] . To date, these m e t h o d s have allowed description of a number of distinct determi-
From The Howard Hughes Medical Institute Laboratory and Departm~,nts of Medicine. Patholog)- and Microbiology, and Immunology, Washington Unirersity School of Medicine, and the Missouri-Illinois Re- gional Red Cross Blood Program, St. Louis, Missouri.
Address req#ests for reprints to: Dr. Benjamin D. Schwartz, Division of Rhe#~u~t~log~. B~x 8045. IF.shington Univ~sity School of Medicine, St, Lo.is, MO 63110.
Reeei~d •979.
Human immtmolo~ 1, 37-~4 (19~0) ~) P,l~,vi~r Nord, Hoflmsd. Inc., 1980 ~2 Vs~led~t A~., New Yock, NY 10017
37 0198-8859/80/01037-1852.25

38 B.D. Schwartz et al.
nants encoded by alleles at each of the recognized HLA loci [4]. However, analysis of the reaction patterns has also indicated that alloantiserum produced by exposure to a given HLA determinant will frequently also react with other HLA determinants as well. This second set of reaction patterns has allowed the organi- zation of particular HLA determinar~ts into cross-reactive groups [5].
The basis for this cross-reactivity has long been sought. Serological and cap- ping studies on cross-reactive HLA molecules suggested that these molecules might possess common antigenic determinants to account for the cross-reactivity [6, 7]. Biochemical confirmation of this postulate, however, has been only re- cently reported [8]. In that previous report, we demonstrated that HLA-B molecules belonging to the B7 cross-reacti~ e group, i.e . HLA-B7, Bw22, B27, and B40, all shared a common alloantigenic determinant, tentatively designated X, which accounted for the cross-reactivity of these m~_:ecules and possibly ac- counted for the finding that all members of this group were associated with the spondyloarthropathies. These results also suggested that similar public antigenic determinants would explain the serologic cross-reactivity of the other groups, and might in part explain other HLA-disease associations.
Behcet's disease was originally described as a triad of recurrent aphthous oral ulcers, genital ulcers, and ocular inflammation. Other systemic features include skin lesions, arthritis, gastrointestinal involvement, thrombophlebitis, arter~tis, and involvement of the central and peripheral nervous system. The disease is particularly prevalent in Japan, the eastern Mediterranean basin, and Italy. Its pathogenesis is unknown. In 1973, 71% of.Japanese patients with Behcet's disease were reported to possess HLA-B5 [91. This association of Behcet's disease with HLA-B5 has been confirmed in other racial groups [10-12]. In addition, other members of the HLA-B5 cross-reactive group (B15, B18, and Bw35) were also repcr~ed to be associated with Behcet's disease [9, 11, 12]; 91% of Japanese patients with the disease possessed one of the B5 cross-reacting antigens [11].
The association of the entire HLA-B5 cross-reactive group with Behcet's dis- ease prompted us to seek the presence of a common antigenic determinant unique to this cross-reactive group. Our present studies demonstrate that HLA-B5, B15, B18, and Bw35 molecules all share a common, but distinc~ an- tigenic determinant.
MATERIALS A N D M E T H O D S
Cells. Peripheral blood cells were obtained from healthy Caucasian volunteers who had previously been typed for HLA-A and HLA-B determinants. Table 1 lists the HLA types of individuals whose cells were used as a source of various HLA-A and HLA-B antigens. No individual possessing HLA-Bw53, another member of the B5 cross-reacting group, was available for our stud),. In collecting cells, approximately 50 mi of blood was immediately heparinized with 0.5 ml ammonium heparin (100/~/mi) (Sherwood Medical Industries, St. Louis, MO). Twenty-five milliliters of heparinized blood in each of two tubes were mixed with 5 ml of dextran in cormal ~line (McGraw Laboratories, Glendale, CA), and allowed to stand for 60 rain at 57°C. At the end of this time, the top 15-20 m! were collected, washed, and used as the source of white cells in subsequent steps [151.
Alloantiserura. Serum B'~ t was obtained from a multiparous Caucasian female (HLA-Aw30, A - , B12, B - ) whose husband wa~ typed as HLA-A2, A - , BS, B - . When analyzed by microcytotoxicity [14|, this serum reacted with HLA-A2' cells at a titer of 1:2, with HLA-B5 cells at a titer of 1:4, and with HLA-Bw35

A B5-CREG Public Antigen .~3
T A B L E 1 HLA types of individuals us. d as a source of various HLA antigens
HLA antigen HLA-type of individuals
A2 A2, --, B12, B40 B5 All, Aw24, BS, BI7
A2, --, BS, BwS0 Aw24, Aw31, BS, B40
BI5 AI, A9, B12, BI5 AI, Aw32, BIS, B37
BIT AI, A3, BT, B17 BI8 A3, A28, BI4, B18 B21 A2. --, B 12 (Bw44), Bw21 ~B~50)
A2, A3, Bw21 (Bw49), B27 Bw35 A9, Aw30, B8, Bw35 BS, Bw35 A3, Aw31, BS, Bw35 BIS, Bw35 A2, Aw24, BI5, Bw35 Controls A3, Ag, B12, B14
cells at a titer of 1:2 (Table 2), and had previously been shown to react with HLA-Bw53 at a titer of 1:I (J. Lee, unpublished data). In order to separate the various antibody populations within the serum, a series of platelet absorptions was performed. The serum was first twice absorbed with HLA-A2 ÷, B5-, Bw35- platelets [15] (actual typing of platelets used was HLA-A2, A l l , B7, B27) as follows. The platelets from one platelet transfer pack (ca. 7 - 8 x I0 *° platelets; 2-ml packed volume) were used for each absorption. The platclets were thoroughly washed with acid citrate dextrose solution-Formuk; A (Fenwall Laboratories, Deerfield, IL) diluted 1:5 in normal saline. The packe ] platelets were then resuspended in 60 ml of serum BW, and incubated at 37°C for 2 hr. The platelets were removed by centrifugation at 12,000 xg for 15 rain. The once absorbed serum was then mixed with a second 2 ml of fresh packed platelets, and the absorption step repeated. This twice absorbed serum was absorbed twice further with HLA-BS-, Bw35 ~ platelets (actual typing of platelets wa . HLA-A3, Aw24, B12, Bw35) using the procedure described above. This four-times ab- sorbed serum, when retested by microcytotoxicity, retained activity with HLA-B5 cells at a titer of 1:4, but had completely lost its reactivity on HLA- Bw35 cells and had no reactivity on HLAoA2 cells (Table 2). This absorbed
T A B L E 2 Titers of serum BW, anti-Y, and anti-B5 on cells bearing FILA-B Antigens of the B5 cross-reacting group
Cells tested
A2 B5 BI5 B17 Bw21 '~ B18 Bw~5
Serum BW 1:2 1:4 NR ~ NR NR NR 1:2 Anti-Y NR 1:8 l: 1 NR NR NR 1:8 Anti-B5 (absorbed) NR 1:4 NR NR NR NR NR
"Identical results obtaine d for the Bw49 and BwS0 splits of B21. ~NI~ *~ no reaction wkh undiluted serum or antibody.

40 B.D. Schwartz et al.
serum was concentrated 2i/2- to 3-fold by ammonium sulfate precipitation and was used as a functionally monospecific anti-HLA-BS.
The antibody which had reacted with the BS-, Bw35 + platelets in the first absorption step was eluted by resuspending the platelets in 2 ml of physiologic saline, which had been I;rought to pH 3.0 with HC! [151. The platelets were incubated for 3 rain with shaking, and then removed by centrifugation at 12,000 xg for 10 rain. The pH of the eluate was adjusted to 7.0 with NaOH. When retested in a microcytotoxicity assay, the eluted antibody reacted with both HLA-B5 and HLA-Bw35 cells at a titer of 1:8, and had no reaction against HLA-A2 cells ~Table 2). This antibody was tentatively designated anti-Y) Evi- dence of its functional monospecificity in this system is given under Results.
The anti-HLA-Bw35 (Jones) and ~.nti-HLA-B15 (Malotsky) were obtained from the American Red Cross, St. Louis, MO. Each serum is functionally mona- specific so that anti-HLA-Bw35 gives no reaction on HLA-B5, B15, or B18 cells, and the anti-HLA-B15 gives no reaction on HLA-BS, B18, or Bw35 cells. The anti-HLA-B18 (CC186) was the generous gift o f Dr. William E. Braun, Cleveland Clinic, Cleveland, Ohio. It has no reactivity, against BS, B15, or Bw35 cells.
Absorption ofanti-Y with/eukocytes. Fifty microliters ofanti-Y was incubated with 2 x 10 ~ leukocytes bearing B5, B17, B18, Bw21, or Bw35 for 60 min at 37°C, and the cells removed by centrifugation at 1100 xg for 10 rain. This procedure was repeated once, so that in all cases where absorption was necessary, the anti-Y was absorbed twice.
Preparation of an lgG fraction from normal human serum, in cases where the human a2-microglobulin present in normal serum might interfere with the reac- tion of rabbit anti-/~-~-microgiobulin in the test step of sequential immuno- precipitation experiments, a//~-microglobulin-free igG fraction of the serum was used. This fraction was prepared as follows [16]. One volume of serum was incubated with I0 volumes o f a 10% suspension o f Protein A-bearing Staphylococcus aureus Cowan I strain ~SaCI) for 30 rain at 4°C. After thorough washing, the lgG was dissociated by the addition o f one volume of 3.5 M MgCIz for 15 rain at room temperature [17], and the SaC! pellet was removed by centrifugation. The supernate containing the purified igG was dialyzed against Tris-buffered saline (TBS), pH 7.4. This dialysate was used as the igG fraction in subsequent experiments.
Xenoantiserum. Rabbit anti-human B.,-microglobulin was purchased from Accu- rate Scientific Co., Hicksville, N.Y.
Human 3~-microg/obu/in. Human//z-microglobulin was the generous gift of Dr. Nobuyuki Tanigaki, Roswell Park Memorial Institute, Buffalo, NY.
Radiolabeling of cells and preparation of solubi/ized antigens. The procedure used has been previously described [8, 16, 18, 19]. in brief, peripheral blood ieuko- cytes were resuspended at 2 - 5 x l0 T cells/ml in modified Eagle's medium with- out serum in which the sole source of le,,cine was *H-leucine ( 8 0 - 1 1 0 Ci/mM, New England Nuclear Corp, Boston, MA) at a concentration o f 200 t~Ci/mi, and incubated for 4 - 6 hr at 37°C in a humidified 5% CO~ atmosphere. The cells were harvested, washed, and then solubilized in TB$ containing 0.5% of the
~ The designavons of the public determinant as "Y" and the antil:~dy recognizing this determinant as "anti-Y" are used tentatively pending the assignment of a more fi~rmal designation by the Interna- tional Nomenclature Committee.

A B5-CREG Public Antigen 41 .
nonionic detergent Nonidet P-40 (NP-4q; Particle Data Laboratories, Elmhurst, IL). Insoluble debris was removed by ultracentrifugation at 100,000 xg for 60 min. The glycoprotein molecules containing glucose or mannose were then purified 10- to 20-fold by allowing them to bind to an affinity column of /xns culinaris lectin covalently attached to Sepharose 4B (Pharmacia Fine Chemicals, Piscataway, NJ), and eluting them from the column with 0.1 M ~,-methld man- noside [20]. This purified glycoprotein fraction was incubated with 300--500 tzl of packed SaCI for 30 min at 4°C to remove any endogenously radiolabeled lgG. The SaCI were removed by centrifugation, and the supernate was used as the source of solubilized antigen in subsequent steps [19].
lmmunoprecipitation and analysis of solubilized HLA molecules. An aliquot of gly- coprotein fraction containing 100,000-200,000 cpm was incubated with 100 ~.! of alloantiserum (or eluted alloantibody) for 2 hr at 4°C. One milliliter of a lOe~ suspension of SaCI was then added to pellet: any antigen-antibody complexes formed and the incubation was continued an additional 30 min [19]. The pellets were centrifuged at 100 ×g for 10 min, and wa.,;hed three times in TBS containing 0.25% NP-40 (TSN). Radiolabeled antigen was then dissociated from antibody and the staphylococci by boiling it, 100 ~l of 0.0625 M Tris, pH 6.8, containing 2% sodium dodecyl sulfate (SDS) and 2% 2-mercaptoethanol (2-ME) for 1.5 min, and the staphylococci removed by centrifugation. One hundred microliters of 2% 2-ME, 20% glycerol, and 0.004% phenol were added, and the sam.pie analyzed using a modification [18] of the discontinuous SDS-polyacrylamide gel electrophoresis (SDS-PAGE) system originally described by Laemmli [21| and Maizel [22]. Both 10 and 15% polyacrylamide gels were used. The gels were cut into 2-mm slices, each slice incubated overnight at 37°C in 1.5 ml of toluene containing 7% Protosol (New England Nuclear) and 4 g/liter Omnifluor (New England Nuclear) and the radioactivity of each slice determined in a Beckman LS 8000 scintillation counter (Beckman Instruments, Palo Alto, CA). Molecular weights were estimated from the position of marker proteins clectrophoresed at the same time [22].
Preparation of~"l-labeled Protein A, Protein A derived from SaC1 (Pharmacia Fine Chemicals) was iodinated using r':'l Bolton-Hunter reagent [23] (New England Nuclear).
12:'l-Protein A binding assay [241. Anti-Y and normal huma~ immunoglobulin G (NHIgG) were ultracentrifuged immediately prior to use to remove material > 10S [25]. Fifty microliters of anti-Y (an amount known to be in excess) or normal human immunoglobulin were incubated with 1,000,000 leukocytes for 30 min at 4°C. The cells were washed three times with Hank's balanced salt solution con- taining 2% ovalbumin, 1,000,000 cpm of v-'~l-Protein A were added, and the incubation continued for 30 rain at 4°C. The cell pellets were then washed three times, and the radioactivity bound to each cell pellet was determined. Each test was done in triplicate, and average values and standard deviations are reported. Identical results were obtained when cells were prepared using Ficoll-Hypaque in place of dextran sedimentation.
RESULTS
S~rum BW Contains Three Funct ional ly Defined Ant ibody Populat ions
The reactions of serum BW with A2 cells and B5 cells would be expected from the HLA types of BW and her husband. However, the additional reaction of this serum with HLA-Bw35 cells, particularly at a different titer from the reaction

42 B.D. Schwartz et al.
TABLE 3 Absorption ofanti-Y by B5 and Bw35 cells
Undiluted anti-Y absorbed twice with cells bearing
Cells staining with eosin (%)
B5 Bw35
No absorption >90 >90 B7, B40 (control) >90 >90 B5 <10 <:10 Bw55 <10 <10
with HLA-B5 cells, suggested that serum BW might contain an antibody to the B5 cross-reactive group. To dissect out the various antibody populations, serum BW was subjected to a series of platelet absorptions. The first set of platelet absorptions with HLA-A2*, BS-, B35- platelets was done to remove the anti- HLA-A2 antibody. The absorbed serum was then twice absorbed with B5-, Bw35 + platelets. This absorbed serum was demonstrated to react only with B5 cells (Table 2). In contrast, the antibody(ies) eluted from the B5-, Bw35 + platelets reacted equally well with both B5 and Bw35 cells, suggesting that a single antibody population designated anti-Y was responsible for both reactions.
To determine if we were in fact dealing with a single antibody population, a series of absorptions was performed. Anti-Y was twice absorbed with ~.-ILA-B5 cells, HLA-Bw35 cells, or control cells, and then tested for residual activity against B5 and Bw35 cells, in both cases, absorption of anti-Y with either B5 or Bw35 cells completely removed activity for both cell types (Table 3). This result indicated that anti-Y contained only a functionally monospecific .~.ntibody popu- lation reactive with both B5 and Bw35 cells.
Anti-Y Reacts with Cells Bearing HLA-BS, HLA-BIS, HLA-BIS, and HLA-Bw35
It was apparent from the initial microcytotoxicity assays that anti-Y reacted with HLA-B5 and HLA-Bw35 cells as measured by direct cytotoxicity. Because of the possibility that anti-Y might react with other members of the B5 cross-reactive group, anti-Y was tested by direct microcytotoxicity on HLA-BI5 cells, B17 cells, BI8 cells, and Bw21 cells. Undiluted anti-Y caused direct cytotoxicity of HLA-B15 cells, but had no reaction against the B18, the B17, or the Bw21 cells (Table 2). However, because of the well-known cytotoxicity negative, absorp- tion positive (CYNAP) phenomenon [26], it was possible that anti-Y was reacting with these cells, but could not induce cytotoxicity. Anti-Y was there- fore absorbed with B17, B18, or Bw21 cells to ascertain if any of these absorptions would remove the cytotoxic activity of anti-Y for Bw35 cells (Table 4). B18 cells completely removed this cytotoxic activity, whereas absorption with control cells, B17 or Bw21 cells had absolutely no effect. We therefore concluded that anti-Y reacted with cells bearing HLA-BS, B15, B18, and Bw35, but not those bearing B17 or Bw21.
12sl-Protein A Binding
The direct microcytotcxicity results and the absorption results suggested that there might be a quantitative difference in the number of available anti-Y binding sites on the various cells. To confirm this contention, we performed an ~ i -
A B5-CREG Public Antigen 4~
T A B L E 4 Absorption of anti-Y by B 17, B* 8, and Bw21 cells
Bw35 cells staining with eosin (%) Serum I~ilution
Anti-Y absorbed with cells bearing Neat 1:2 1:4 1:8
No absorption >90 >90 60-80 40-50 B7/B40 (control) >90 >90 60-80 40-50 B 17 ~>90 >90 60-80 40-50 BI8 <10 <10 <10 <10 Bw21 ~ >90 >90 60-80 40-50
"Identical results obtained for the Bw49 and Bw50 splits of Bw21.
Protein A binding experiment. Table 5 contains the results of one representative experiment. When comparing the binding of one antibody on a number of differ- ent cells, the amount of '"nl bound is a reflection of the number of available antigenic sites i24]. It appeared that there was a hierarchy of available Y antigenic sites depending on the HLA-B antigen which the cell bore, as follows: Bw35 > B5 > B15 > B18. In addition, the binding of cells bearing both B5 and Bw35 approximated the sum of binding by B5 and Bw35 cells, respectively, and the binding of cells bearing both B15 and Bw35 approximated the sum of binding by B15 and Bw35 cells, respectively. Thus, the v':'l-Protein A binding assay con- firmed the results of the microcytotoxicity and C Y N A P experiments, and al- lowed a degree of quantitation.
An t i -Y Reacts w i th an HLA- l ike Molecu le
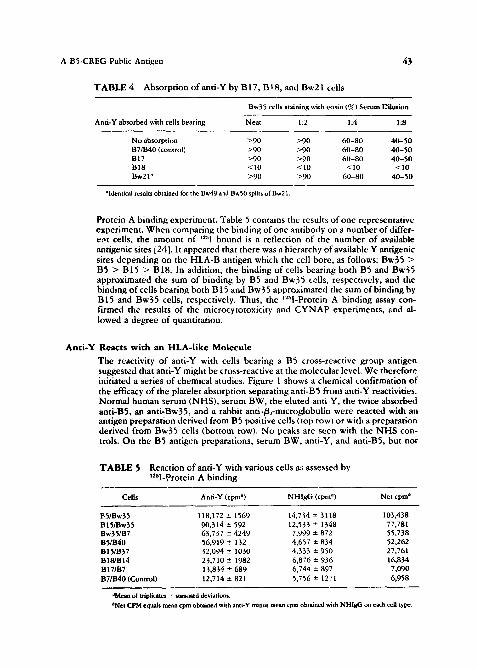
The reactivity of anti-Y with cells bearing a B5 cross-reactive group antigen suggested that anti-Y might be cross-reactive at the molecular level. We therefore initiated a series of chemical studies. Figure 1 shows a chemical confirmation of the efficacy of the platelet absorption separating anti-B5 from anti-Y reactivities. Normal human serum (NHS), serum BW, the eluted anti-Y, the twice absorbed anti-BS, an anti-Bw35, and a rabbit anti-/~..,-microglobulin were reacted with an antigen preparation derived from B5 positive cells (top row) or with a preparation derived from Bw35 cells (bottom row). No peaks are seen with the N H S con- trols. On the B5 antigen preparations, serum BW, anti-Y, and anti-BS, but not
T A B L E 5 Reaction of anti-Y with various cells as assessed by t251-Protein A binding
Cells Anti-Y (cpm a) NHIgG (cpm ~) Net cpm b
135/Bw35 118,172 +- 1569 14,734 +-- 3118 103,438 BIS/Bw35 90,314 -+ 592 12,533 -+ 1348 77,781 Bw35/B7 63,737 -+ 4249 7,999 - 872 55,738 BS/B40 56,919 -+ 132 4,657 -+ 834 52,262 BIS/B37 32,094 -+ 1030 4,333 +- 950 27.761 BIS/B14 23,710 - 1982 6,876 -+ 936 16,834 BIT/B7 13,834 - 689 6,744 +- 897 7,090 BT/B40 (Control) 12,714 +- 82 i 5,756 -- 1271 6,958
°Mean of triplicates :: standatd deviations. ~'Net CPM equah mean cpm obtained with anti-Y minus mean cpm obtained with NHigG on each cell type.
44
200
~ I0~ ~ ~.- ~3 t.¢
Z u 200
,...1 |
u 100 u ' ~
Eluted Absorbed NHS Serum BW Anti-Y Anti-B5 Anti-B35 Anti-~v
~4o~
1400
0 . . . . . . . 1 ~ L ~ . ~ _ ~ - ~
2 4 6 810 24 6 810 24 681024 6 81024 6 810 24 6 810 CM OF GEL
FIGURE 1 Discontinuous SDS-IO'?; PAGE patterns of the molecules solubilizcd t~om :~H-leucine iabeled HLA-B5 positive cells {top~ or HLA-Bw35 positive cells {bottcmL and reactive ~'ith normal human serum, serum BW. the elumd attti-Y, the twice absorbed anti-B5, an anti-Bw35, and r~bit a n t i - ~ . ~ molecules detected ~i mi~me with an apparent raolecular weight of 44,000 daltons. B2~ it~lf cannot be identified on IIY~ polyacryJamide gels.
anti-Bw.~5, react with a molecule(s) which migrate(s) with an apparent molecular weight of 44,000 (/~2-microglobulin cannot be resolved on a 10°~, polyacr,}lamide gel in this system---see below), in contrast, on the Bw39 antigen preps.ration, serum BW, anti-Y, and an anti-Bw35, but not anti-BS, react with such a molecule, showing the completeness o f the initial absorpdon in yielding a func- tionally monospecific and-Bg. Rabbit anti-/~-microglobulin reacted equally well
.with both antigen preparations. The molecular weight estimate o f 44,000 was in agreement with the known
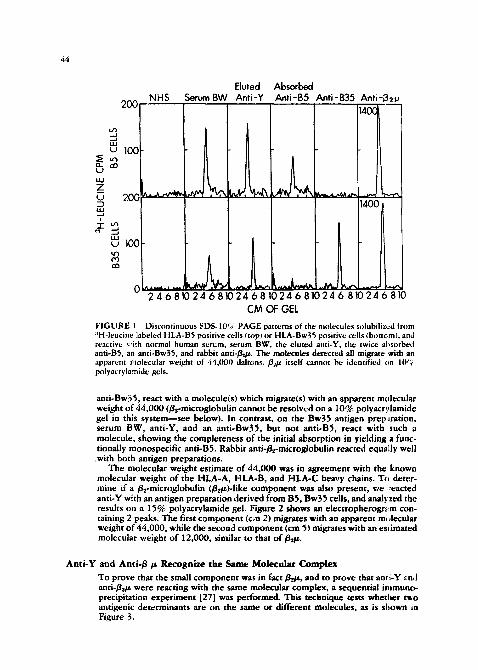
molecular weight of the HLA-A, HLA-B, and HLA-C heavy chains. To deter- mine if a ~e-microglobulin (/~et~)-Iike component was also present, we ~eacted anti-Y with an antigen preparation derived from BS, Bw39 cells, and anal?zeal the results on a 15% polyacrylamide gel. Figure 2 shows an electropherogr.-~m con- taining 2 peaks. The first component (cm 2) migrates with an apparent m,,lecuiar weight of" 44,000, while the secona component (cm 9) migrates with an es*imated molecular weight of 12,000, similar to that o f # ~ .
Ant i -Y and A n t i - g / t Recognize the Same Molecular Complex
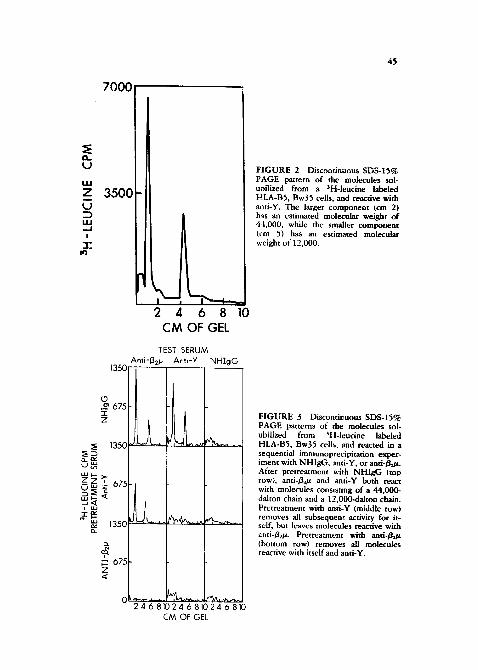
To prove that the small component was in fact gent, and to prove that ami-Y and anti-g~/t were reacting with the same molecular complex, a ~equendal immuno- precipitation experiment [27] was performed. This technique tests whether t~-o antigenic determinants are on the same or different molecules, as is shown in Figure 3.
u
Z
!
7000
3500
2 4 6 8 10 CM OF GEL
1350
O ,_~ 675 ..r Z
~ 1350
~ "- ~75
~ ~ 1350
TEST SERUM Anti-.13~ Anti-Y NHIgG
~ 675 z .~
24 6 81024 6 81024 6 810 CM OF GEL
4~
F I G U R E 2 Discontinuous SDS-15% PAGE pattern of the molecules sob mfilized from a SH-leucine labeled HLA-BS, Bw35 cells, and reactive with anti-Y. The larger component (cm 2) has an estimated molecular weight of 44,000, while the smaller compotient (cm 5) has an esdmared molecular weight of 12,000.
F IGURE 3 Discoatinuous SDS-15% PAGE patterns of the molecules sob ubilized from SH-leucine labeled HLA-B5, Bw35 cells, ,and reacted in a sequential immunoprecipitadon exper- iment with NHIgG, anti-Y, or and-~g,_tz. After pretreatment with NHIgG (top row), and-/g2~ and anti-Y both react with molecules consisting of a 44,000- dalton chain and a 12,000-dalton chain. Pretreatment with and-Y (middle row) removes all subsequen~ activity for it- sell; but leaves molecules reacdve with ~,ati-B.~t~. Pretreatment with anti-Bo.~ (bottom row) removes all molecules reactive with itself and anti-Y.
46 B.D. Schwartz et ai.
An antigen preparation prepared from HLA-BS, Bw35 cells was divided into three aliquots. The first aliquot was pretreated with normal human immunoglob- ulin G (NHIgG) as a control, the second aliquot was pretreated with anti-Y to remove all molecules bearing determinant Y, and the third aliquot was pretreated with anti-Bz/~ to remove all molecules having / / ~ . Each of these pretreated samples was in turn divided into three aliquots, and tested with anti-/~e/z for the presence of/~z/z molecules, with anti-Y for the presence of Y-bearing molecules, or with normal human IgG as a control. After pretreatment with N H I g G (top row), both anti-Bz/* and anti-Y react with molecules consisting of a 44,000- and 12,000-dalton chain. Pre t reatment with anti-Y (second row) removes all molecules reactive with itself, but does leave molecules reactive with anti-/izV.. This, o f course is the expected result, since anti-/~az reacts with HLA-A and HLA-C molecules as well as HLA-B molecules, in contrast, pretreatment with anti-Bz/z (bottom row) completely clears all molecules reactive with itself or with anti-Y. We therefore concluded that anti-Y was reacting with an HLA-like molecule consisting of a heavy chain and Bz/z.
Ant i -Y Reacts wi th a De te rminan t on the Heavy Chain
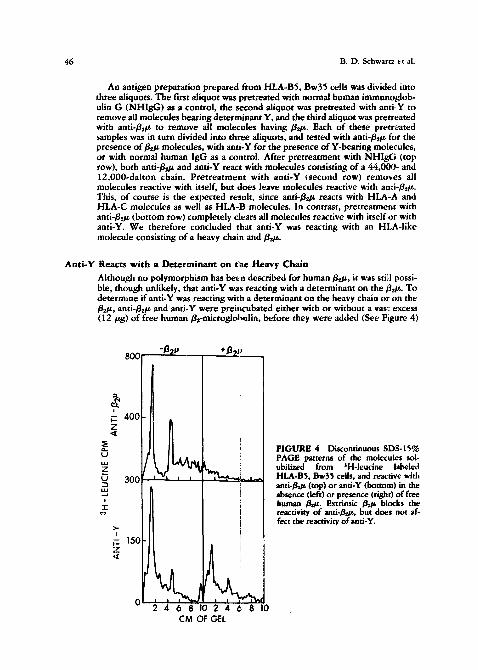
Although no polymorphism has been described for human/~z/z, it was still possi- ble, though unlikely, that anti-Y was reacting with a determinant on the//zt~. To determine if anti-Y was reacting with a determinant on the heavy chain or on the //z/z, anti-/ie/z and anti-Y were prei,cubated either with or without a vas~ excess (12/~g) o f free human/~-microglohttlin, before they were added (See Figure 4)
800 - j 3 2 p ..
!
~ 400 z ,~
i~_
~ 300 ~_1 ._1 .~ ~.~
,,- ~- 15~ :~ ,I
' I
÷~3~1~ ' _
,:
t ! i ~ ' _ _| _~
I i !
2 4 6 8 10 2 4 6 8 I0 CM OF GEL
FIGURE 4 Discontinuous SDS-15~ PAGE patterns of the molecules sol- ubilized from 3H-leucine labeled HLA-BS, Bw35 cells, and reactive with ant i - /~ (top) or anti-Y (bottom) in the absence (left) or presence (right) of free human /JOt. Extrinsic / ~ blocks the reactivity of anti-/!h~, but does not af- fect the reactivity of anti-Y.
A B5-CREG Public Antigen 47
to a B5,Bw35 antigen preparation. In the absence of gz-~nicroglobulin (left panels), both anti-/~z/z and anti-Y react with the HLA-like complex. Preincuba- tion with /~z/~ (right panels) completely blocks the reactivity of and-~gs/g but leaves the activity o f anti-Y virtually undiminished. This res~lt indicated that Y was a determinant on the heavy chain of these molecules.
A n t i - Y Reacts w i th Molecules Bea r ing HLA-B5, HLA-B15, HLA-B18, and HLA-Bw35
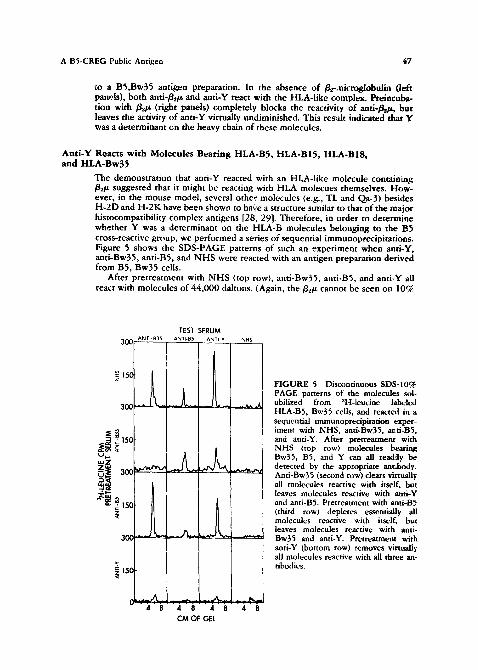
The demonstration that anti-Y reacted with an HLA-like molecule containing g~/z suggested that it might be reacting with HLA molecues themselves. How- ever, in the mouse model, several other molecules (e.g., TL and Qa-3) besides H-2D and H-2K have been shown to have a structure similar to that of the major histocompatibility complex antigens [28, 29]. Therefore, in order to determine whether Y was a determinant on the HLA-B molecules belonging to the E~5 cross-reactive group, we performed a series of sequential immunoprecipitations. Figure 5 shows the SDS-PAGE patterns of such an experiment when and-Y, anti-Bw35, anti-BS, and N H S were reacted with an antigen preparation derived from B5, Bw35 cells.
After pretreatment with N H S (top row), anti-Bw35, anti-BS, and anti-Y all react with molecules of 44,000 daltons. (Again, the g.,/z cannot be seen on 10%
TEST SERUM 30C ANTb B35 ANTI-BS ANTI-Y NHS
~ ~5c
30C
~ ~ ~5c ~ .~ U~.. ~'~
~ ~ 1 ~
3~'
~ ,50
. ~ ~,,~. __~1,~. ~
~ ~ -_ ~,,~ 4 8 4 8 4 8 4 8
CM OF GEL
FIGURE 5 Discontinuous SDS-10% PAGE patterns of the molecules sol- ubilized from aH-leucine labeled HLA-B5, Bw35 cells, and reacted in a sequential immunoprecipitation exper- iment with NHS, and-Bw35, anti-B5, and anti-Y. After pretreatment with NHS (top row) molecules bearing Bw35, BS, and Y can all read,h/ be detected by the appropriate antibody. Anti-Bw35 (second row) clears virtually all molecules reactive with itself, but leaves molecules reactive with anti-Y and anti-BS. Pretreatment with and-B5 (third row) depletes essentially all molecules reactive with itself, but leaves molecules reactive with anti- Bw35 and anti-Y. Pretreatment with anti-Y (bottom row) removes virtually, all molecules reactive with all three an- tibodies.
48 B.D. Schwartz et al.
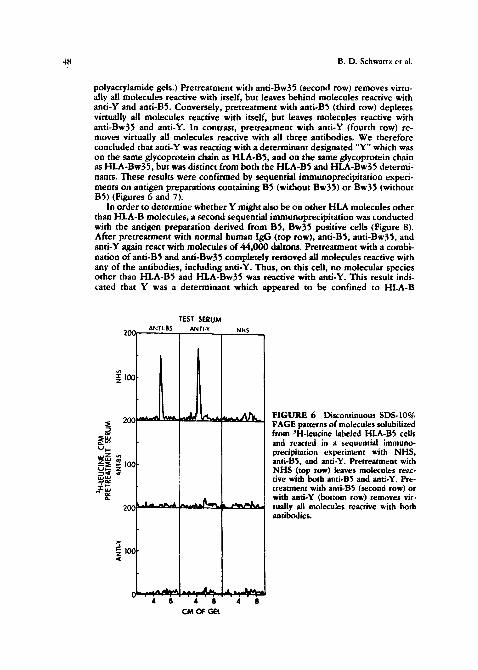
polyacrylamide gels.) Pretreatment with anti-Bw35 (second row) removes virtu- ally all molecules reactive with itself, but leaves behind molecules reactive with anti-Y and anti-BS. Conversely, pretreatment with anti-B5 (third row) depletes virtually all molecules reactive with i~self, but leaves molecules reactive with anti-Bw35 and anti-Y. In contrast, pretreatment with anti-Y (fourth row) re- moves virtually all molecules reactive with all three antibodies. We therefore concluded that anti-Y was reacting with a determinant designated "Y" which was on the same glycoprotein chain as HLA-BS, and on the same glycoprotein chain as HLA-Bw35, but was distinct from both the HLA-B5 and HLA-Bw35 determi- nants. These results were confirmed by sequential immunoprecipitat ion experi- ments on antigen preparations containing B5 (without Bw35) or Bw35 (without BS) (Figures 6 and 7).
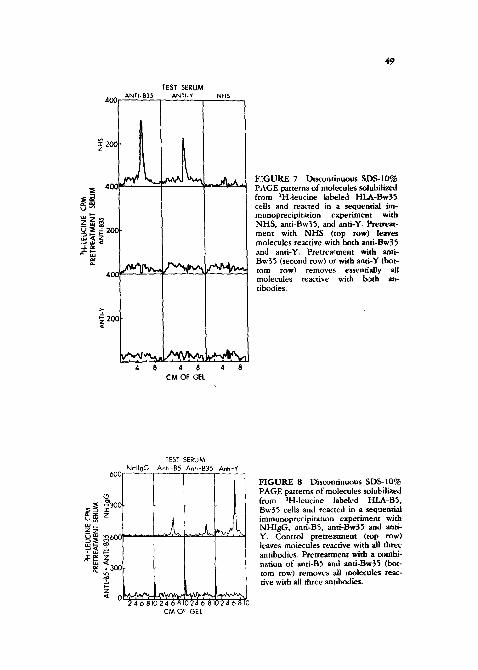
In order to determine whether Y might also be on other HLA molecules other than HLA-B molecules, a ~ c o n d sequential immunoprecipitat ion was conducted with the antigen preparation derived from BS, Bw35 positive cells (Figure 8). After pretreatment with normal hmnan IgG (top row), anti-B5, anti-Bw35, and anti-Y again react with molecules o f 44,000 daltons. Pretreatment with a combi- nation of anti-B5 and anti-Bw35 completely removed all molecules reactive with any of the antibodies, including anti-Y. Thus, on this ceil, no molecular species other than HLA-B5 and HLA-Bw35 was reactive with anti-Y. This result indi- cated that Y was a determinant which appeared to be confined to HLA-B
TEST SERUM 200 ANTI.B5 ANTI.Y NHS __
~ 2o~
~ ~ - ~.,Z, . _z~ ~ ~ ~ IOC ~ ~.H~r~ ~ - ,-N ~,~
~ c i
~- ~ ~ ,~
L ~ . a_m~ ~ _ ~ J . ~ . t , .Jh~,~ 4 8 ~ 8 4 8
CM OF C~L
FIGURE 6 Dis~:ominuou$ SDS-IO% PAGE patterns of molecules ~olubilized from ;H-leucine labeled HLA-B5 cells and reacted in a sequential immuno- precipitation experiment with NHS, anti-BS, and anti-Y. Pretreatment with NHS (top row) leaves molecules re~c- rive with both anti-B5 and anti-Y. Pre- treatment with anti-B5 (second row) or with anti-Y (bottom row) removes vir.. tually all molecules reactive with both antibodies.
400
TEST SERUM ANTI- B35 ANTI- Y NHS
L
~ 40C
~ ~ O ~ ~20( @~ .~,1 ~ .,1~ ;,x~
e ~ ~ ~ ~
~ 2o0
./-~..~ . ~ , ~ ~ / ' ~ 4 8 4 8 4 8
CM OF GEL
F~GURE 7 Discontinuous SDS-IO% PAGE patterns of molecules solubilized from ~H-leucine labeled FILA-Bw35 cells and reacted in a sequential im- munoprecipitation experiment with NHS, anti-Bw35, and anti-Y. Pretreat- ment with NHS (top row) leaves molecules reactive with both and-Bw35 and anti-Y. Pretreatment with and- Bw35 (second row) or with and-Y (bot- tom row) removes essentially all molecules reactive with both an- tibodies.
TEST SERUM NHIgG Ant,-B5 Ant,-B35 Anti-Y
0600 . . . . .
~ ~°°l ~
~
~ o ~ ~ 240810246810246 810246810
CM OF GEL
F I G U R E 8 Discontinuous SD$-IO% PAGE patterns of molecules solubilized from aH-leucine labeled HLA-B5, Bw35 cells and reacted in a sequential immunoprecipita~ion experiment with NHIgG, anti-B5, anti-Bw35 and anti- Y. Control pretreatment (top row) leaves molecules reactive with all three antibodies. Pretreatment with a combi- nation of anti-B5 and anti-Bw35 (bot- tom row) removes all molecules reac- tive with all three antibodies.
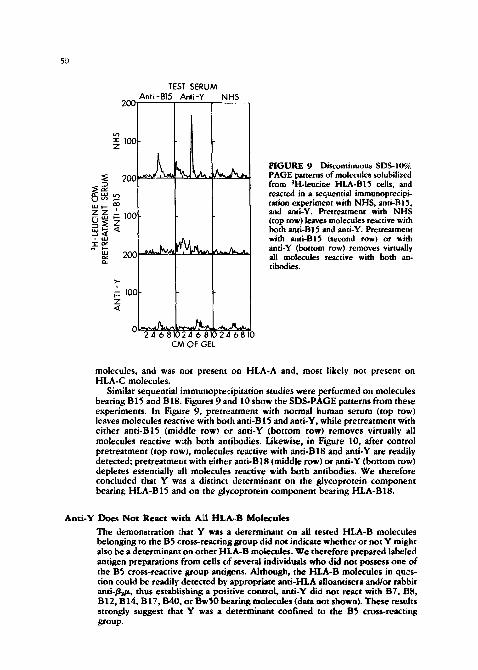
50
200
~ 100 z
~ 200 ~ ~ _ u..I t . ~ u ' ~
~ - ~ , z ~ ~ ~ ~ 100
~ ~ ~o¢ ¢.~
~_ 10C 7 ,¢
TEST SERUM Anti -B15 Anti-Y NHS
~_~k~ @ ~,../k~ ,
,~..A~ ~,,~-.,,. _.,~,~. 24 68~24 6 8~24 6810
CM OF GEL
I:IGURE 9 Discontinuous SDS-10% PAGE patterns of molecules solubilized from 3H-leucine HLA-B15 cells, and reacted in a secluential immunoprecipi- tation experiment with NHS, anti-BlS, and anti-Y. Pretreatment with NHS (top row) leaves molecules reactive with both anti-Bl5 and anti-Y. Pretreatment with anti-Bl5 (second row) or with anti-Y (bottom row) removes virtually all molecules reactive with both an- tibodies.
molecules, and was not present on HLA-A and, most likely not present on HLA-C molecules.
Similar sequential immunopre¢ipitation studies were performed on molecules bearing B15 and B18. Figures 9 and 10 show the SD$-PAGE patterns from these experiments. In Figure 9, pretreatment with normal human serum (top row) leaves molecules reactive with both anti-B 15 and anti-Y, while pretreatment with either anti-B15 (middle row) or anti-Y (bottom row) removes virtually all molecules reactive w~th both antibodies. Likewise, in Figure 10, after control pretreatment (top rowh molecules reactive with anti-Bl8 and anti-Y are readily detected; pretreatment with either anti-Bl8 (middle row) or anti-Y (bottom row) depletes essentially all molecules reactive with both antibodies. We therefore concluded that Y was a distinct determinant on the glycoprotein component bearing HLA-B15 and on the glycoprotein component bearing HLA-B18.
Anti-Y Does Not React with A~I HLA-B Molecules
The demonstration chat Y was a determinant on all tested HLA-B molecules belonging to the B5 cross-reacting group did not indicate whether or not Y might also be a determinant on other HLA-B molecules. We therefore prepared labeled antigen preparations from cells of several individuals who did not possess one of the B5 cross-reactive group antigens. Although, the HLA-B molecules in ques- tion could be readily detected by appropriate anti-HLA alloantisera and/or rabbit anti-/~/~, thus establishing a positive control, anti-Y did not react with B7, B8, B12, B14, B17, B40, or BwS0 bearing molecules (data not shown). These results strongly suggest that Y was a determinant confined to the B5 cross-reacting group.
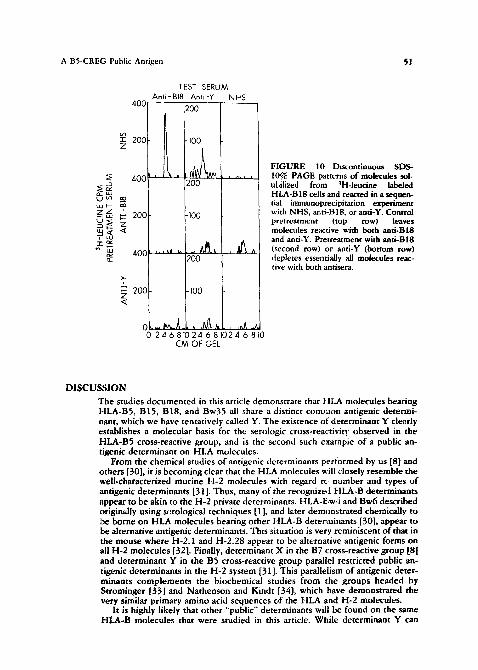
A BS-CREG Public Antigen ~1
400
TEST SERUM Ant i - BI8 Ant, -Y NHS
200
I ~ 2ooi -,oo
~ 4 0 0 ~ :1~. . . . . . I ~ ~ ~ 200 L) UO ~o ~ , ~
~ ~ ~oo ~,oo
~ ~ ~ ~ 400 , . , ~ t u .. ~
~ 2 ~
~ 200 .]oo ,~
0 - ~ d ~ ~ , ~ . ~ . , ~ ~ 0 ~ ' ~ e 8 , 0 ~ e s , 0 ~ e s , 0
CM OF G-;L
FIGURE 10 Discontinuous SDS- 10% PAGE patterns of molecules sol- u|,ilized from 3H-leucine labeled HLA-B18 cells and reacted in a sequen- tial immunoprecipitation experiment with NHS, aati-Bl8, or anti-Y. Control pretreatment (top row) leaves molecules reactive with both anti-BIB and anti-Y. Pretreatment with anti-BIB (second row) or anti-Y (bottom row) depletes essentially all molecules reac- tive with both antisera.
D I S C U S S I O N
The studies documented in this article demonstrate that HLA molecules bearing HLA-B5, B15, B18, and Bw35 all share a distinct comn~on antigenic determi- nant, which we have tentatively called Y. The existence of d ~terminant Y cle~.rly establishes a molecular basis for the serologic cross-reacti'~tity observed in the HLA-B5 cross-reactive group, and is the second such exarapie o f a public an- tigenic determinant on HLA molecules.
From the chemical studies of antigenic determinants performed by us [8] and others [30], it is becoming clear that the HLA molecules will closely resemble the well-characterized routine H-2 molecules with regard to number and types o f antigenic determinants [31]. Thus, many of the recognized HLA-B determinants appear to be akin to the H-2 private determinants. HLA-I/.w4 and Bw6 described originally using serological techniques [1], and later demonstrated chemically to be borne on HLA molecules bearing other HLA-B determinants [30], appear to be alternative antigenic determinants. This situation is very reminiscent of that in the mouse where H-2.1 and H-2.28 appear to be alternative antigenic forms on all H-2 molecules [32]. Finally, determinant X in the B7 cross-reactive group [8] and determinant Y in the B5 cross-reactive group parallel restricted public an- tigenic determinants in the H-2 system [31]. This parallelism of antigenic deter- minants complements the biochemical studies from the groups headed by Strominger [33] and Nathenson and Kindt [34], which have demonstrated the very similar primary amino acid sequences of the HLA and H-2 molecules.
It is highly likely that other "public" determinants will be found on the same HLA-B molecules that were studied in this article. While determinant Y can

52 B.D. Schwartz et al.
account for cross-reactivity between HLA-BS, B15, B18, and Bw35, it clearly cannot account for the cross-reactivity frequendy observed between B15 and B 17 [5]. Thus, it is likely that yet another public antigenic determinant shared by B15 and B17, and distinct from Y will be found on HLA-B15 and B17. Thus, HLA-B15 will bear at least two distinct public antigenic determinants.
The differential binding of anti-Y to the several Y-bearing molecules, as as- sessed by cytotoxicity testing, Protein A binding, and chemical analyses is an interesting phenomenon and is an identical situation to that which we pre.~iously observed with antioX and the B7 cross-reactive group [8]. At that time we made two assumptions, namely that the number of HLA-B molecules borne on the cell surface is independent of the antigenic determinant on those molecules, and that all HLA-B molecules determined by a single haplotype are identical. On the basis of these two assamptions, and the finding of differential reactivity of the anti- public antigen andbody with various cells, we concluded that the recognized HLA-B private determinant on the molecules influenced the availability (and/or affinity) of the public determinant to bind (and/or retain) antibody. Our present data also strongly favor this concept of the private determinant exerting an "al- losteric" effect on the public determinant.
While HLA-B5 has been highly associated with Behcet's disease in several studies [9-12], at least two of these studies found that there was, in addition., a higher than expected incidence of the B5 cross-reactive group antigens in pa- tients with Behcet's [11, 12]. This clustering of the Y-bearing molecules with Behcet's prompts speculation that the public antigenic determinant Y may have a role in disease predisposition. The two most likely hypotheses are that Y may be directly involved in disease predisposition or that the gene determining Y may merely be closely linked with a disease susceptibility gene for Behcet's.
In the first hyFothesis, Y might be viewed as a receptor for an extrinsic etiologic agent that causes Behcet's. This model would hold that the BS-Behcet's association is observed most frequently because B5 exerts the aforemention$,d allosteric effect on Y that allows Y to assume the conformation most conducive for binding the pathogenic agent. Other B5 cross-reactive group private determi- nants do not permit this optimal conformation of the Y receptor site, and thus these private antigens are less often associated with Behcet's.
The second theory associating Behcet's with the Y-bearing molecules postu- lates that a linkage disequilibrium originally existed between a Y determiniag gene and a disease-susceptibility gene for Behcet's. Mutations causing and/or affecting the private determinant on the Y-bearing molecule might also affect the degree of lit, kage disequilibrium, in this model, the linkage disequilibrium be- tween Y and Behcet's would be the strongest when B5 was present, and the linkage disequilibria with the other B5 cross-reactive determinants would be weaker, accounting for the presently observed HLA-Behcet's associations.
Although both models are speculative they may nonetheless suggest helpful experiments, which can differentiate between them, and help elucidate the un- derlying mechanisms of HLA-disease associations.
ACKNOWLEDGMENT This work was supported in part by Nati~mal Institutes of Health/:rants A1-13782 ,~nd A1-15353. by grant IM-131 from the American Cancer Society and b~, an Amerkan National Red Cross Research Grant. The authors wish to thank Dr. Susan E. Cullen fo* fruitful discussions and helpful suggestions, and Lorraine Bourisaw and Jud~, Craig t~:r expert preparation of the manuscript.

A B5-CREG Public Antigen 53
REFERENCES
1. van Rood JJ, van Leeuwen A: Leukocyte grouping. A method and its application. J Clin Invest 42:1382, 1963.
2. Payne R, Tripp M, Weigle J, Bodmer W, Bodmer J: A new leukocyte isoantigen system in man. Cold Spring Harbor Symp Quant Biol 29: 285, 1964.
3. Bodmer W, Bodmer J, Adler S, Payne R, Bialek J: Genetics of 4 and LA human leukocTte e, roups. Ann NY Acad Sci 129: 473, 1966.
4. WHO Report: Nomenclature for factors of the HLA s~/stem. EurJ immunol v: 911, 1977 .
5. Joysey VC, Wolf W: HLA-A, -B, and -C, antigens, their serology and cross-reaction. Br Med Bull 3: 217, 1978.
6. Legrand L, Dausset J: The cotnplexity of the HL-A gene product. I. Study of a serum produced against HL-A5 in an HL-A semi-identical situation. Tissue Antigens 4: ~29. 19"/4.
.7. Legrand L, Dausset J: The complexity of the HL-A gent product. 11. Possible evi- dence fi~r a public determinant common to the first and second HL-A series. Trans- plantation 19: 177, 1975.
8. Schwartz BD, Luehrman LK, Rodey GE: A public antigenic determinant on a family of HLA-B molecules. A basis for cross-reactivity and a possible link with disease predisposition. ~ Clin Invest 64: 938, 1979.
9. Ohno S, Aoki K, Sugiura S, Nakayama S, itaku~a K, Aizawa M: HL-A3 and B~hcet's disease. Lancet 2: 1383, 197~.
10. Ryder LP, Anderson E, Swigaatd A: HLA and ~fisease: Third report from the HLA and Disease Registry. Tissue Antigens, in press.
11. Ohno S, Nakayama LE, Sugiura S, Itakura K, Aoki K, Aizawa M: Specific hisc~om- patibility antigens associated with Behcet's disease. Am J Ophthalmo180: 636, 19"~%
12. Rose,clef E, Saudan Y, Jeannet M: Recherche des antigenes HL-A dans la maladie de Behcet. Ophthahnologica (Basel) 172:116, 19"~6.
13. Skoog WA, Beck WS: Studies on the fibrinogen dextran, and phytohemagglutinin methods of isolating leukocytes. Blood 11: 436, 1956.
I.i. Mir.tal KK, Mickey MR, Singal DP, Terasaki PI: Serotyping for homotrans- plantat ion--Refinement of microdroplet lymphocyte cytotoxicity test. Trans- plantation 6: 913, 1968.
15. Rodey GE, Sturm B, Aster RH: Cross-reactive HL-A antibodies. Separation of multi- ple HL-A antibody,' specificities b~, platelet absorption and acid elution. Tissue Anti- gens 3,: 63, 19"3.
16. Schw~'~rtz BD, Cit.:en R, Berggard I, Shevach EM: Guinea pig homologues of TL and Qa-2 antigens. J ~mmunol 121: 835, 19"78.
17. Kessler S: Rapid isolation ot antigens from cells with a staphylococcal Protein A- antibe.dy absorbent: parameters of the interaction of antibody-antigen complexes with Protein A.J Immunol 115:161 ~, 19"75.
18. Schwartz BD, Kask A, Paul W, Shevach E: Structural characteristics of the alloaati- gens determined by the major histocompatibility complex of the guinea pig. J Exp Med 143: 5-il, 19"~6.
19. Cullen SE, Schwartz BD: An improved method fi)r isolation of H-2 and la alloanti- gens using immunoprecipitation induced by Protein-A bearing staphylococci. J lm- munoi 11~': 136, 1976,

54 B.. D. Schwartz et al.
20. Hayman MJ, Crumpton MJ: Isolation of glycoproteins from pig lymphocyte plasma membranes using Lens culinaris phytohemagglutinin. Biochem Biophys Res Commun 47: 923, 1972.
21. Laemmli UK: Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680, 1970.
22. Maizel Jv Jr: Polyacrylamide gel electrophoresis of viral proteins. Methods Virol 5: 179, 1971.
23. Bolton AE, Fiunter WM: The labelling of proteins to high specific radioactivities by conjugation to a 12:'l-containing acylating agent. Application to the radioimmunoassay with .~.n appendix on the preparation method by J Rudinger anti V. Ruegg. Biochem J 133: ~,29, 1973.
24. DorvM G, Welsh KI, Wigzeli H: Labeled Staphylococcal Protein-A as an immunologi- cal probe in the analysis of cell surface markers. Stand J lmmunol 3: 4{}5, 1974.
29. Trautman R, Cowan JM: Preparative and analytical ultracentrifugation, in Methods in Immunology and Immunochemistry, vol. 2 CA Williams, MW Chase, ed. New York, Academic Press, 1968, p. 81.
26. Yunis EJ, Ward FE, Amos DB: Observations of the CYNAP phenomenon, in His- tocompability Testing 197{}. PI Terasaki. ed. Copenhagen, Munksgaard, 1970. p. 351.
27. Cullen SE, Schwartz BD, Nathenson SG: The distribution of alloantigenic spe¢i- ficities of native H-2 products. J Immunol 108: 596, 1972.
28. Vitetta E, Uhr JW, Boyse EA: Isolation and characterization of FI-2 and TL alloanti- gens from the surface of mouse lymphocytes. Cell Immunoi 4: 187, 1972.
29. Micb.aelson J, Flaherty L, Vitetta E, Poulik MD: Molecular similarities between the Qa-2 alloantigen and other gene prt~lucts of the 17th chromosome of the mouse. J E×p Med 145: 1066, 1977.
30. A~'res J, Cresswell P: HLA-B specificities and w4, w6 spe¢ificities are on the same polypepdde. EurJ Immunol 6: 794, 1976.
31. Klein J: Biology of the Mouse Histocompatibility-2 Complex. New York, Springer- Verlag, 1975, p. 124.
32. Snell GD, Cherry M, D~mant P: H-2: its structure and similarity to FIL-A. Transplant Re~ 15: 3, 1973.
33. Orr FIT, Lopez-deCastro JA, Parham P, Ploegh HL, Strominger JL. Comparison of amino acid sequences of two human hlstocompatibility antigens, HLA-A2 and FILA-BT: The location of putative alloantigenlc sites. Proc Natl Acad Sci USA, in press.
34. Coligan JE, Kindt TJ, Ewenstein BM, Uehara H, Martinko JM, Nathenson SG: Further structural analysis of the murine FI-2K" glycoprotein using radiochemical methodology. Mol Immunol 16: 3, 1979.