Embed Size (px)
Citation preview

Plant Physiol. (1990) 93, 857-8630032-0889/90/93/0857/07/$01 .00/0
Received for publication December 26, 1989Accepted February 21, 1990
A Polyethylene Glycol-Mediated Protoplast TransformationSystem for Production of Fertile Transgenic Rice Plants1
Akio Hayashimoto2, Zhijian Li, and Norimoto Murai*
Department of Plant Pathology and Crop Physiology, College of Agriculture, Louisiana State University,Baton Rouge, Louisiana 70803-1720
ABSTRACT
We have established an efficient procedure for protoplasttransformation and regeneration of fertile transgenic plants ofrice (Oryza sativa L.) cultivars Nipponbare and Taipei 309. Proto-plasts were mixed with a plant-expressible hygromycin resist-ance gene and treated with 25% (w/v) polyethylene glycol. Strin-gent selection of transformed colonies was applied to 14-day-oldregenerated protoplasts in the presence of 95 micromolar ofhygromycin B for 12 days. After selection, 450 and 200 resistantcolonies were recovered per million treated Taipei 309 and Nip-ponbare protoplasts, respectively. Southem hybridization analy-sis of hygromycin-resistant cell lines and regenerated plantsindicated that 1 to 10 copies of transferred DNA were integratedat 1 to 4 loci of the rice genome. Southem DNA analysis suggeststhat the introduced plasmid DNA may form concatemers by inter-molecular recombination prior to integration. Four Taipei 309 and39 Nipponbare transgenic rice plants were regenerated andgrown to maturity in the greenhouse. Two Taipei 309 and 35Nipponbare plants set viable seeds. Agronomic traits of Taipei309 transgenic plants and inheritance of the hygromycin resist-ance trait by progeny of the selfed transgenic plants wereanalyzed.
Application of gene transfer procedures for functionalanalysis of eukaryotic genes has greatly facilitated our under-standing of the mechanisms of gene regulation. Plant regula-tory elements involved in transcriptional regulation have beenanalyzed using Agrobacterium tumefaciens-based transfor-mation systems (9, 11, 23).We are interested in studying the regulation ofgene expres-
sion in rice, a monocot plant. Our objective is to develop a
homologous gene transfer system in rice to circumvent thedifficulties encountered in heterologous dicot gene expressionsystems (15). A number of laboratories have already reportedtransformation of rice protoplasts (22, 24) and subsequentregeneration of transgenic plants (21, 25, 26). Production offertile mature plants and their progeny was also recentlydemonstrated (19).We have established an efficient and reproducible proce-
dure for rice protoplast propagation and fertile plant regen-
This work was supported in part by grants from the LouisianaEducation Quality Support Fund (1987-90)-RD-A-6 and from Ishi-hara Sangyo Kaisha, Ltd. to N. M.
2 Present address: Department of Biotechnology, Ishihara SangyoKaisha, Ltd., Kusatsu, Shiga 525, Japan.
857
eration (10). In this procedure, we have optimized parametersin protoplast transformation. Here, we report an efficientprocedure for production of a large number of transgenic riceplants to facilitate quantitative analysis of gene expression.
MATERIALS AND METHODS
Chemicals and Enzymes
Cellulase RS and Macerozyme R-10 were obtained fromYakult Honsha Co. (Tokyo, Japan) and SeaPlaque agarosewas acquired from FMC Co. An oligolabeling kit, mixedsequence hexadeoxynucleotides pd(N)6 and Sephadex G-50(DNA grade) were obtained from Pharmacia. Nytran mem-branes were acquired from Schleicher & Schuell. Thymidine5'-triphosphate (TPP) [methyl-3H] and deoxycytidine 5'-tri-phosphate (dCTP) [a-32P] were purchased from ICNBiochemicals.
Plant Expression Vectors
Plant expression vectors were constructed to express abacterial gene under the control of plant transcription initia-tion and termination signals. The nopaline synthase (nos) andcauliflower mosaic virus (CaMV) 35S promoters were used asconstitutive promoters in expression vectors pTRA104 and105, respectively. The transcription termination and polyad-enylation sites were derived from Agrobacterium tumefaciens.The transcription termination and polyadenylation sites
were isolated as a 292 bp Sau3a fragment from a tumormorphology large (tmi) gene in T-DNA of A. tumefacienspTi 15955. This fragment was located 110 bp downstreamfrom the stop codon ofthe tml gene from nucleotide sequencepositions 10,775 to 11,167 of the T-DNA (2). Sau3a ends ofthe isolated fragment were filled in with Klenow fragmentand ligated with a SacI linker. A 300 bp Sacl fragment wascloned into the SacI site of pUC12, forming pTRA 103.The nos promoter was isolated from plasmid pUC9KN (1),
kindly provided by Dr. M. Bevan, Plant Breeding Institute,Cambridge, U.K. This plasmid was digested with PstI and theresulting 500 bp fragment was cleaved with Sau3a to releasea 270 bp fragment that represents the nucleotide sequencefrom positions -266 to about +35 from the transcriptioninitiation site ofthe nos gene. The Sau3a fragment was clonedinto the BamHI site ofpTRA 103, and the resultant expressionvector was called pTRA 104.The CaMV 35S promoter was isolated from a plasmid
containing the 19S and 35S promoters of the CaMV-S ge-
www.plantphysiol.orgon July 14, 2018 - Published by Downloaded from Copyright © 1990 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 93, 1990
nome, representing the nucleotide sequence from positions7141 to 7666 (7). The plasmid was kindly provided by Drs.H. Hirochika and J. Ikeda, National Institute of Agrobiolog-ical Resources, Tsukuba, Japan. A Bglll site was present atthe 3'-end of the fragment at nucleotide position 7666, 283bp downstream from the transcription initiation site. A BglIH-digested plasmid was treated with exonucleases III and VII togenerate a series of deletions from the BglII site toward thecap site of the 35S transcript. The deleted plasmids werefurther digested with PstI, and fragments 350 to 400 bp inlength were isolated after PAGE. The isolated fragments werecloned into the PstI and HincII sites of the vector M l 3mp 19.The DNA sequence of the isolated fragments was determinedon single strand DNA by the dideoxy method. One clonecarried a 337 bp sequence representing nucleotide positionsfrom -285 to +50 from the transcription initiation site of theCaMV 35S promoter. This sequence was recovered fromM13mpl 9 as a 350 bp HindIII/BamHI fragment and clonedinto pTRA 103, forming the expression vector pTRA 105.
Hygromycin Resistance Gene
A hygromycin resistance gene was used as a selectablemarker for rice protoplast transformation. The coding se-quence of the Escherichia coli hygromycin B phosphotrans-ferase (hph) from the plasmid pLG90 was kindly provided byDr. Linda Gritz, Applied Biotechnology, Cambridge, MA (8).The entire hph coding sequence had been cleaved from thevector as a 1.1 kbp BamHI fragment. The initiation andtermination codons ofthe hph gene resided 14 bp downstreamfrom the 5'-end, and 100 bp upstream from the 3'-end of thefragment, respectively. The hph sequence was inserted intothe BamHI site of the expression vector pTRA 104, so thatthe hph sequence was downstream from the nos gene pro-moter, resulting in pTRAl 31. The hph coding region was alsoplaced downstream from the 35S promoter of the expressionvector pTRA 105, resulting in pTRA 132.
Plasmid DNA Isolation
Plasmid DNA for protoplast transformation was isolatedby the modified method of Hansen and Olsen (4). PlasmidDNA was further purified by CsCl density gradientcentrifugation.
Susnension and Protoplast Culture
Rice (Oryza sativa L.) cultivars Nipponbare and Taipei 309were used for this study. The procedure for callus induction,suspension callus culture, protoplast isolation, and protoplastculture has been described in detail in the previous report(10).
Protoplast Transformation after PEG Treatment
The procedure for PEG-mediated transformation of riceprotoplasts was essentially as described by Negrutiu et al. (13).After the final KMC wash (5), isolated protoplasts were resus-pended at a density of4 x 106 protoplasts per mL in a MaMgsolution containing 0.4 M mannitol, 15 mM MgCl2, 1% (w/v)
Mes (pH 5.6). Ethanol-precipitated plasmid DNA was dis-solved at a density of 10 ,ug per 20 ,uL of 10 mm Tris-HCl (pH7.8), and 1 mM EDTA (TE).3 Ten ,ug of the plasmid DNAwere applied to one million of protoplasts in 0.25 mL ofMaMg solution. Protoplast/DNA mixtures were incubatedfor 10 min at room temperature. Forty % (w/v) PEG (mol wt8,000) in a given amount ofMaMg solution was gently appliedto the protoplast/DNA mixture to achieve the desired PEGconcentration. After 15 min of incubation at room tempera-ture, protoplasts were suspended in 40 mL ofKMC solutionand centrifuged at 1 30g for 8 min. Two million PEG-treatedprotoplasts were resuspended in 1 mL of General protoplastmedium. The protoplast suspension was then mixed with anequal volume of prewarmed protoplast medium containing2.5% (w/v) Seaplaque agarose. The agarose mixture waspoured into a Petri dish (60 x 15 mm) (18). The solidifiedagarose sheet was cut into 10 mm square blocks and wascultured in 5 mL of liquid protoplast medium with nurse cellsas described elsewhere (10).
Oligolabeling
Oligolabeling was performed according to the protocol pro-vided in the Pharmacia oligolabeling kit. Probe DNA frag-ments were isolated by electroelution or glass-powder elution.Plasmid DNA (50-200 ng) was denatured by boiling or al-kaline treatment, and mixed with 1 ,g of random primerpd(N)6 in a solution containing 0.5 mm each of deoxynucleo-tide triphosphates. The polymerization reaction was initiatedby addition of 2 units of Klenow fragment and 50 ,uCi ofTTP[methyl-3H] or dCTP [a-32p], and continued for 3 h at roomtemperature. At the end of the reaction, 43 to 71% of totalradioactivity was incorporated into DNA. The specific activityof the DNA probes thus prepared ranged from 1 to 7 x 107cpm per utg of 3H-labeled DNA, and from 0.4 to 1.5 x 109cpm per ,ug of 32P-labeled DNA.
3H-Labeled Plasmid DNA Transfer to Protoplasts
3H-labeled DNA was used to monitor DNA transfer toprotoplasts after PEG treatment or electroporation. Ten ,ug of3H-labeled plasmid DNA were applied to one million proto-plasts in 0.5 mL of MaMg solution. After 10 min of incuba-tion at room temperature, protoplasts were treated with PEGin a series of concentrations. Protoplasts were washed threetimes in 14 mL of KMC solution and then lysed in 1 mL ofdistilled water. Radioactivity ofprotoplast lysates was countedfor 10 min in a Beckman LS1801 scintillation counter. Back-ground counts for each PEG concentration were obtainedfrom solutions treated exactly in the same manner as theprotoplast samples, except without protoplasts. Transfer ofradioactivity to protoplasts was estimated by subtracting thebackground counts from counts in the protoplast lysates.
Selection for Hygromycin Resistance
Hygromycin selection for resistant colonies was performedon 14-d-old regenerated protoplasts. Protoplasts were em-
3 Abbreviation: TE, 10 mM Tris-HCI (pH7.4), 1 mM EDTA.
858 HAYASHIMOTO ET AL.-
www.plantphysiol.orgon July 14, 2018 - Published by Downloaded from Copyright © 1990 American Society of Plant Biologists. All rights reserved.
RICE PROTOPLAST TRANSFORMATION
bedded in agarose blocks (10 x 10 x 0.7 mm) after isolationand all subsequent treatments were done as a bead-typeculture ( 18). Protoplasts were grown in nurse culture for 10 dand in liquid protoplast culture for an additional 4 d. Regen-erated protoplasts were then selected for 12 d in Generalprotoplast medium containing 95 Mm hygromycin B. Agaroseblocks were transferred to General medium containing 0.25%(w/v) Sigma type I agarose and 3% (w/v) sucrose and weremaintained for 2 to 4 weeks or until the colony size reached2 mm in diameter. The number of hygromycin-resistantcolonies was counted in at least 10 agarose blocks after 2weeks of soft agarose culture. The apparent transformationefficiency of protoplasts was defined as a percentage of thenumber of hygromycin-resistant colonies per total number ofprotoplasts originally plated in agarose blocks.
Transgenic Plant Regeneration
Transgenic plants were regenerated from hygromycin-re-sistant protoplast colonies in Murashige and Skoog basalmedium (12) containing 555 gM inositol, 500 mg/L caseinhydrolysate, 2.9 gM indole-3-acetic acid, and 3.6 Mm 6-ben-zylaminopurine (3). The cultures were maintained from 40to 50 d at 27°C under constant light (56 ,umol m-2 s-').Subcultures were repeated every 20 to 25 d.
Progeny Analysis of Hygromycin Resistance
Inheritance of the hygromycin-resistance trait was analyzedin seeds of selfed transgenic plants. Seed were dehulled usinga mortor and pestle. Dehulled seeds were sterilized in 70%(v/v) aqueous ethanol for 1.5 min then 2.6% (v/v) sodiumhypochlorite-water for 35 min with vigorous agitation, fol-lowed by three rinses with sterile distilled water. Sterilizedseeds were plated in MS medium containing 190 Mm ofhygromycin B and cultured for 15 d at 27 °C under light.
Rice DNA Isolation
Rice DNA was isolated from suspension cells or leaves oftransgenic plants by Rogers and Bendich's procedure withminor modifications (16). The DNA solution was adjustedwith CsCl of the density to 1.55 g/mL and was incubated at20°C for 1 h. After removal of cell debris by low-speedcentrifugation, the total DNA solution was centrifuged at1 87,000g for 40 h at 20 'C. A single DNA band thus formedwas extracted with 20 x SSC-saturated isopropanol seventimes and was dialyzed three times against 2 L of TE bufferat 4 'C. The yield ofDNA ranged between 50 and 100 ,ug per10 g tissue.
Southern Blot Hybridization
Southern blot hybridization was performed essentially asdescribed by Southern (20) and by Schleicher & Schuell'sprotocol. Five Mg of rice DNA was digested with BamHI orHindIll. Digested DNA samples were fractionated by 0.8%(w/v) agarose gel electrophoresis. The fractionated DNA wastransferred to a Nytran membrane and hybridized to a 3p_labeled 1.1 kb BamHI fragment corresponding to the coding
sequence of the hph gene (2-7 x 10' cpm). Two control riceDNA samples were reconstructed by adding unlabeled probeDNA to concentrations corresponding to 2 and 10 copies oftransformed DNA per diploid rice genome (0.60 pg). Theintensities of the bands in x-ray films were measured by aBiorad Video Densitometer model 620. The copy number oftransformed DNA was estimated by comparing the bandintensities between transformed DNA and reconstructedsamples.
RESULTS AND DISCUSSION
Selection for Hygromycin Resistance
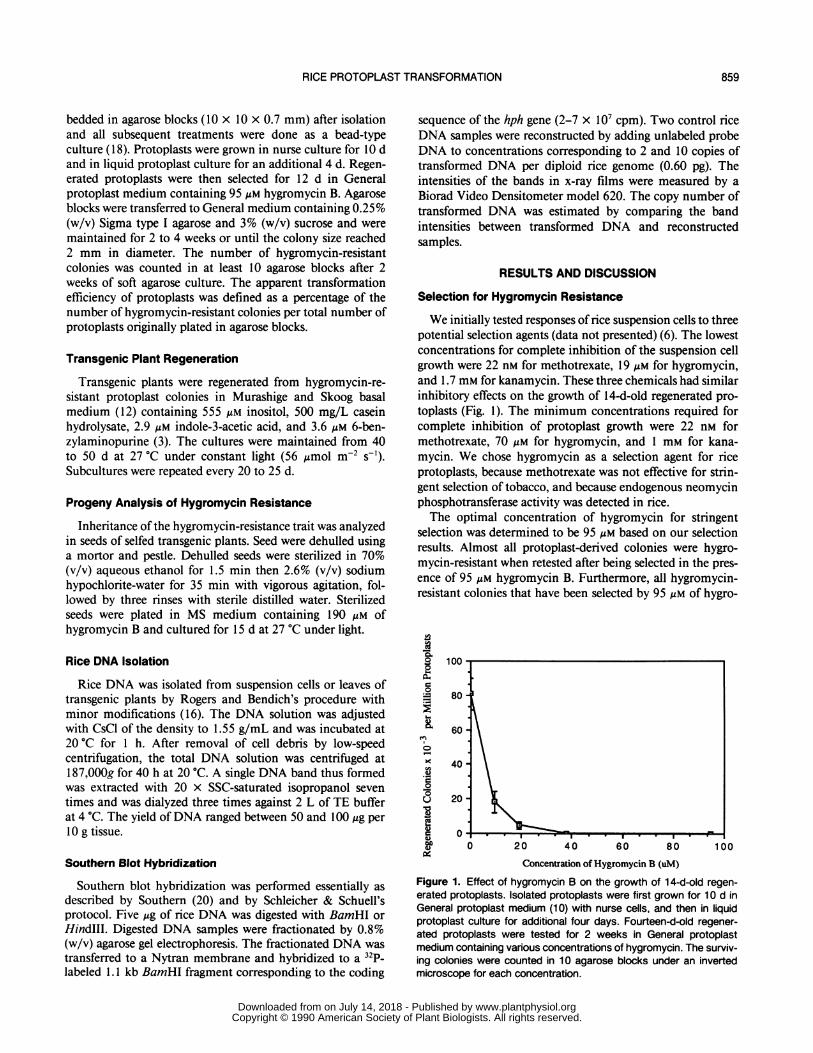
We initially tested responses of rice suspension cells to threepotential selection agents (data not presented) (6). The lowestconcentrations for complete inhibition of the suspension cellgrowth were 22 nm for methotrexate, 19 Mm for hygromycin,and 1.7 mm for kanamycin. These three chemicals had similarinhibitory effects on the growth of 14-d-old regenerated pro-toplasts (Fig. 1). The minimum concentrations required forcomplete inhibition of protoplast growth were 22 nm formethotrexate, 70 uM for hygromycin, and 1 mM for kana-mycin. We chose hygromycin as a selection agent for riceprotoplasts, because methotrexate was not effective for strin-gent selection of tobacco, and because endogenous neomycinphosphotransferase activity was detected in rice.The optimal concentration of hygromycin for stringent
selection was determined to be 95 uM based on our selectionresults. Almost all protoplast-derived colonies were hygro-mycin-resistant when retested after being selected in the pres-ence of 95 uM hygromycin B. Furthermore, all hygromycin-resistant colonies that have been selected by 95 Mm of hygro-
q
0z0
ctr_0
*a0
u
4)
4)
100
80
60
40
20
00 20 40 60 80 1 00
Concentration of Hygromycin B (uM)
Figure 1. Effect of hygromycin B on the growth of 14-d-old regen-erated protoplasts. Isolated protoplasts were first grown for 10 d inGeneral protoplast medium (10) with nurse cells, and then in liquidprotoplast culture for additional four days. Fourteen-d-old regener-ated protoplasts were tested for 2 weeks in General protoplastmedium containing various concentrations of hygromycin. The surviv-ing colonies were counted in 10 agarose blocks under an invertedmicroscope for each concentration.
859
www.plantphysiol.orgon July 14, 2018 - Published by Downloaded from Copyright © 1990 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 93, 1990
200
9 180-
160
140 -
X 120e~ 100-00Ke 80-
6060
= 40
.,l 20
00 5 10 15 20 25 30 35
Final PEG concentration (%)
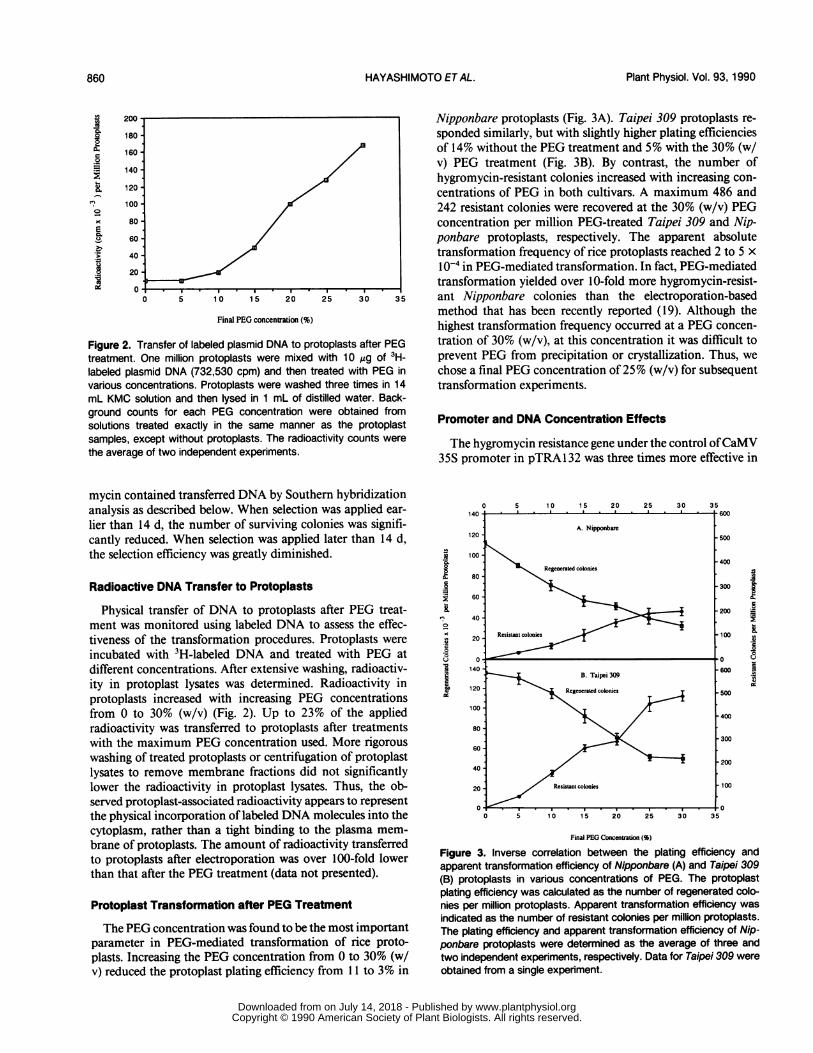
Figure 2. Transfer of labeled plasmid DNA to protoplasts after PEGtreatment. One million protoplasts were mixed with 10 ug of 3H-labeled plasmid DNA (732,530 cpm) and then treated with PEG invarious concentrations. Protoplasts were washed three times in 14mL KMC solution and then lysed in 1 mL of distilled water. Back-ground counts for each PEG concentration were obtained fromsolutions treated exactly in the same manner as the protoplastsamples, except without protoplasts. The radioactivity counts werethe average of two independent experiments.
mycin contained transferred DNA by Southern hybridizationanalysis as described below. When selection was applied ear-lier than 14 d, the number of surviving colonies was signifi-cantly reduced. When selection was applied later than 14 d,the selection efficiency was greatly diminished.
Radioactive DNA Transfer to Protoplasts
Physical transfer of DNA to protoplasts after PEG treat-ment was monitored using labeled DNA to assess the effec-tiveness of the transformation procedures. Protoplasts wereincubated with 3H-labeled DNA and treated with PEG atdifferent concentrations. After extensive washing, radioactiv-ity in protoplast lysates was determined. Radioactivity inprotoplasts increased with increasing PEG concentrationsfrom 0 to 30% (w/v) (Fig. 2). Up to 23% of the appliedradioactivity was transferred to protoplasts after treatmentswith the maximum PEG concentration used. More rigorouswashing of treated protoplasts or centrifugation of protoplastlysates to remove membrane fractions did not significantlylower the radioactivity in protoplast lysates. Thus, the ob-served protoplast-associated radioactivity appears to representthe physical incorporation of labeled DNA molecules into thecytoplasm, rather than a tight binding to the plasma mem-brane of protoplasts. The amount of radioactivity transferredto protoplasts after electroporation was over 100-fold lowerthan that after the PEG treatment (data not presented).
Protoplast Transformation after PEG Treatment
The PEG concentration was found to be the most importantparameter in PEG-mediated transformation of rice proto-plasts. Increasing the PEG concentration from 0 to 30% (w/v) reduced the protoplast plating efficiency from 11 to 3% in
Nipponbare protoplasts (Fig. 3A). Taipei 309 protoplasts re-sponded similarly, but with slightly higher plating efficienciesof 14% without the PEG treatment and 5% with the 30% (w/v) PEG treatment (Fig. 3B). By contrast, the number ofhygromycin-resistant colonies increased with increasing con-centrations of PEG in both cultivars. A maximum 486 and242 resistant colonies were recovered at the 30% (w/v) PEGconcentration per million PEG-treated Taipei 309 and Nip-ponbare protoplasts, respectively. The apparent absolutetransformation frequency of rice protoplasts reached 2 to 5 x10-4 in PEG-mediated transformation. In fact, PEG-mediatedtransformation yielded over 10-fold more hygromycin-resist-ant Nipponbare colonies than the electroporation-basedmethod that has been recently reported (19). Although thehighest transformation frequency occurred at a PEG concen-tration of 30% (w/v), at this concentration it was difficult toprevent PEG from precipitation or crystallization. Thus, wechose a final PEG concentration of 25% (w/v) for subsequenttransformation experiments.
Promoter and DNA Concentration Effects
The hygromycin resistance gene under the control ofCaMV35S promoter in pTRA 132 was three times more effective in
0a
a
sU
acn
.280
a
AI.9
a
fiU
Final PE Cocentrati (%)
Figure 3. Inverse correlation between the plating efficiency andapparent transformation efficiency of Nipponbare (A) and Taipei 309(B) protoplasts in various concentrations of PEG. The protoplastplating efficiency was calculated as the number of regenerated colo-nies per million protoplasts. Apparent transformation efficiency wasindicated as the number of resistant colonies per million protoplasts.The plating efficiency and apparent transformation efficiency of Nip-ponbare protoplasts were determined as the average of three andtwo independent experiments, respectively. Data for Taipei 309 wereobtained from a single experiment.
860 HAYASHIMOTO ET AL.
www.plantphysiol.orgon July 14, 2018 - Published by Downloaded from Copyright © 1990 American Society of Plant Biologists. All rights reserved.

RICE PROTOPLAST TRANSFORMATION
producing resistant colonies than the one under the nospromoter in pTRA 131 (Fig. 4). After PEG-mediated transfor-mation with pTRA 132, the numbers of resistant colonieswere 197 and 176 per million Nipponbare protoplasts in thepresence of 95 and 190 ,uM hygromycin, respectively. Thecorresponding colony numbers were 76 and 59 after transfor-mation with pTRA 131. These results were consistent with thefindings that the CaMV 35S promoter was expressed moreeffectively than the nos promoter in tobacco and maize cells(17).The apparent transformation frequency of Taipei 309 pro-
toplasts increased with higher applied concentrations of plas-mid DNA. There were 98, 226, 243, and 492 hygromycinresistant-colonies per million PEG-treated protoplasts aftertransformation with 5, 10, 15, and 20 ,ug of pTRA132, re-spectively. We have not tested more than 20 jig DNA per 20,uL TE buffer because DNA was difficult to dissolve in a smallvolume. We used 10 utg DNA per million protoplasts in allother experiments reported here.
Southern Hybridization Analysis
Transformation of hygromycin-resistant colonies was con-firmed by Southern hybridization analysis of Taipei 309genomic DNA. Total DNA was isolated from five independ-ent suspernsion cell lines and leaves of three transgenic plantsderived from these cell lines. Taipei 309 DNA was digestedwith HindIII, then the gel-fractionated DNA was transferredto nylon filters and probed with a 1.1 kbp BamHI fragmentrepresenting the hph coding sequence (Figs. 4B and 5A). Theplasmid pTRA132 contains a unique HindIII site (Fig. 4A).Should individual copies of plasmid DNA be inserted atmultiple loci of the rice genome, the number of variableborder fragments detected will be the lower limit for thenumber of integrated plasmid copies. The size of borderfragments will depend on the integration site in the plasmidmolecule and the location of neighboring HindIII site in therice genome (Fig. 5A). Should tandem copies ofplasmid DNA
be inserted at a single locus, a border and 4.4 kbp internalfragments will be generated. The intensity ratio of 4.4 kbpinternal to border band will indicate the copy number ofintegrated plasmid DNA.BamHI digestions should generate a 1.1 kbp band, indicat-
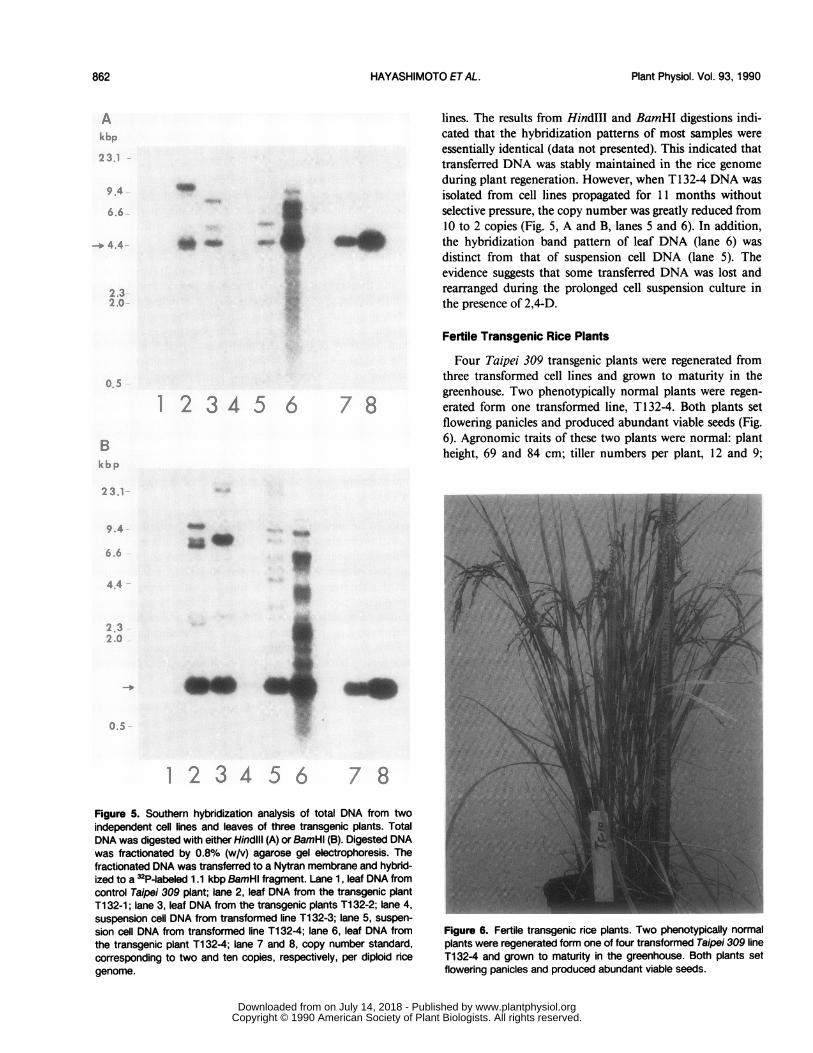
ing the presence of the intact hph coding sequence. Shouldthe hph coding sequences be disrupted in the integration orsubsequent rearrangement event, fragments with variablesizes would be detected (Fig. 5B).The results from HindIII digestion indicated that one
(T132-4c) (Fig. 5A, lane 5), two (T132-1 and 2) (lanes 2 and3) and five copies (Tl 32-4p) (lane 6) of the internal 4.4 kbpband were detected. This suggests that the introduced plasmidDNA might have formed concatemers by intermolecular re-combination prior to integration into the rice genome. Lessthan one copy of the 3 kbp band was detected in T-132-3(Fig. SA, lane 4). Based on the all digestion results, the copynumber of the integrated plasmid was estimated to be lessthan one (T 132-3) (Fig. 5, lane 4), two to three (T1 32-4c)(lane 5), four (T132-1 and -2) (lanes 2 and 3), and 10 copies(Tl 32-4p) (lane 6). A single two-copy concatemer appearedto be present in the rice genome in T 132-4 (lane 5). Two-copy concatemers may have inserted into two locations of thegenome in T 132-1 and 2 (lanes 2 and 3). Several concatemermolecules may be integrated at four loci of the rice genomein T1 32-4p (lane 6).
Similar evidence for plasmid concatenation was previouslyreported in transgenic tobacco plants after protoplast trans-formation by electroporation (14). In tobacco, linear plasmidmolecule appeared to form both head-to-head and head-to-tail concatemers. A tandem repeat of T-DNA was also de-tected after transformation of tobacco with Agrobacteriumtumefaciens nopaline-type Ti plasmid (9, 23). Thus, the inte-gration of transferred DNA after concatemer formation ap-pears to be a general feature of plant cell transformation.DNA from hygromycin-resistant calli and corresponding
transgenic plants were analyzed in three independent cell
A. Hygromycin resistance gene in pTRA132.
Hph CodingSequence
1.1 I 1
Bam Eco Bam
B. Southern Hybridization Analysis.
(1) Probe DNA
(2) Expected fragments
BamHl digestion: 1.1 kbp
HindIlI digestion: X 4.4 kbp
nmi
rerminator pUC12
0.3 2.7Eco Hind Figure 4. A, Physical structure of the hygromy-
cin resistance gene in the plasmid pTRA1 32; B,probe DNA used in southern analysis was a 1.1BamHI fragment corresponding to the codingsequence of the hygromycin resistance gene.Sizes of expected DNA fragments in BamHI andHindlil digestions of genomic DNA.
,,/ -
CaMV35SPromoter
1 0.3
Hind
r
861
www.plantphysiol.orgon July 14, 2018 - Published by Downloaded from Copyright © 1990 American Society of Plant Biologists. All rights reserved.
862HAYASHIMOTOETAL.~~~~~~~~~~~~~~~~~~~PlantPhysiol. Vol. 93, 1990
Ak bp
4.. 4 eo m0
2.320.
I2 34 5
kB
lines. The results from Hind~ll and BamHI digestions indi-cated that the hybridization patterns of most samples wereessentially identical (data not presented). This indicated thattransferred DNA was stably maintained in the rice genomeduring plant regeneration. However, when T 132-4 DNA wasisolated from cell lines propagated for 11I months withoutselective pressure, the copy number was greatly reduced from10 to 2 copies (Fig. 5, A and B, lanes 5 and 6). In addition,the hybridization band pattern of leaf DNA (lane 6) wasdistinct from that of suspension cell DNA (lane 5). Theevidence suggests that some transferred DNA was lost andrearranged during the prolonged cell suspension culture inthe presence of 2,4-D.
Fertile Transgenic Rice Plants
Four Taipei 309 transgenic plants were regenerated fromthree transformed cell lines and grown to maturity in thegreenhouse. Two phenotypically normal plants were regen-erated form one transformed line, T 132-4. Both plants setflowering panicles and produced abundant viable seeds (Fig.6). Agronomic traits of these two plants were normal: plantheight, 69 and 84 cm; tiller numbers per plant, 12 and 9;
6
6
4~~~)0
Figure 5. Southern hybridization analysis of total DNA from twoindependent cell lines and leaves of three transgenic plants. TotalDNA was digested with either HindlIll (A) or BamHl (B). Digested DNAwas fractionated by 0.8% (w/v) agarose gel electrophoresis. Thefractionated DNA was transferred to a Nytran membrane and hybrid-ized to a 32P-labeled 1.1 kbp BamHl fragment. Lane 1, leaf DNA fromcontrol Taipei 309 plant; lane 2, leaf DNA from the transgenic plantTi 32-1; lane 3, leaf DNA from the transgenic plants Ti 32-2; lane 4,suspension cell DNA from transformed line T132-3; lane 5, suspen-sion cell DNA from transformed line Ti 32-4; lane 6, leaf DNA fromthe transgenic plant Ti 32-4; lane 7 and 8, copy number standard,corresponding to two and ten copies, respectively, per diploid ricegenome.
Figure 6. Fertile transgenic rice plants. Two phenotypically normalplants were regenerated form one of four transformed Taipei 309 lineT132-4 and grown to maturity in the greenhouse. Both plants setflowering panicles and produced abundant viable seeds.
862 HAYASHIMOTO ET AL.
www.plantphysiol.orgon July 14, 2018 - Published by Downloaded from Copyright © 1990 American Society of Plant Biologists. All rights reserved.

RICE PROTOPLAST TRANSFORMATION
average panicle length, 19 and 16 cm; average spikelet num-bers per panicle, 89 and 63; and seed fertility, 62 and 94%.
Inheritance of the hygromycin-resistance trait by progeny
was demonstrated by genetic analysis of seeds of two selfedT 132-4 plants. The segregation ratios of resistant to sensitivetraits in progeny were 48 to 25 and 38 to 17, respectively.Thus, the progeny analysis indicated that the hygromycin-resistance trait is located at one locus or one closely linkedloci of the T 132-4 genome.
Two other Taipei 309 plants regenerated from transformedcell lines T 132-1 and T 132-2 displayed some abnormal phe-notypic characteristics. They were sterile despite persistentattempts to obtain viable seeds in the greenhouse. The plantT132-1 was taller than average, and had wide, thick leaves,increased kernel size and longer awn. These changes are
typical of polyploid or aneuploid plants (3). The architectureof T132-2 plant was abnormal in that the tiller angles were
much wider than the average. These abnormal phenotypeshave been frequently observed among somaclonal rice var-
iants (3).Thirty-nine Nipponbare transgenic plants were grown to
maturity and set flowering panicles in the greenhouse. Four-teen plants were transformed with pTRA131 and 25 plantswith pTRA 132. After selfing, 35 plants produced viable seeds.Analysis of seed fertility and other agronomic traits of thesetransgenic plants and inheritance of the hygromycin-resist-ance gene will be reported in a subsequent publication.
ACKNOWLEDGMENTS
We wish to thank Masahiro Oshima, Masahiro Sakamoto, andKazuyuki Ohira for constructing plant expression vectors pTRA 103,104, and 105; Andreana Lisca and Christian Knaak for their excellenttechnical assistance; Drs. Linda Gritz, Michael W. Bevan, HirohikoHirochika, and Johe Ikeda for providing plasmids; Drs. David J.Longstreth, Mary E. Musgrave, Milton C. Rush, and Mark D. Burowfor critical reading of the manuscript; and all membersof the Plant Molecular Biology Laboratory for their support andencouragement.
LITERATURE CITED
1. Bevan MW, Flavell RB, Chilton MD (1983) A chimaeric anti-biotic resistance gene as a selectable marker for plant celltransformation. Nature 304: 184-187
2. Barker RF, Idler LB, Thompson DV, Kemp JD (1983) Nucleo-tide sequence of the T-DNA region from the Agrobacteriumtumefaciens octopine Ti plasmid pTi 15955. Plant Mol Biol 2:335-350
3. Cao J, Rush MC, Nabors M, Xie QT, Croughan TP, NowickEM (1989) Development and inheritance of somaclonal vari-ation in rice. In Biological nitrogen fixation associated withrice production. Oxford and IBH Publishing, New Deli, India
4. Hansen JB, Olsen RH (1978) Isolation oflarge bacterial plasmidsand characterization of the P2 incompatibility group plasmidspMGl and pMG5. J Bacteriol 135: 227-238
5. Harms CT, Potrykus I (1978) Fractionation of plant protoplasttypes by iso-osmotic density gradient centrifugation. TheorAppl Genet 53: 57-63
6. Hauptmann RM, Vasil V, Ozias-Akins P, Tabaeizadeh Z, RogersSG, Fraley RT, Horsch RB, Vasil IK (1988) Evaluation of
selectable markers for obtaining stable transformation in thegramineae. Plant Physiol 86: 602-606
7. Hirochika H, Takatsuji H, Ubasawa A, Ikeda J (1985) Site-specific deletion in cauliflower mosaic virus DNA: possibleinvolvement ofRNA splicing and reverse transcription. EMBOJ 4: 1673-1680
8. Gritz L, Davies J (1983) Plasmid-encoded hygromycin B resist-ance: the sequence of hygromycin B phosphotransferase geneand its expression in Escherichia coli and Saccharomycescerevisiae. Gene 25: 179-188
9. Kuhlemeier CP, Green J, Chua NH (1987) Regulation of geneexpression in higher plants. Annu Rev Plant Physiol 38: 221-257
10. Li Z, Murai N (1990) General medium for efficient plant regen-eration from rice (Oryza sativa L.) protoplasts. Plant Cell Rep(in press)
11. Murai N, Sutton DW, Murray MJ, Slightom JL, Merlo DL,Reichert NA, Sengupta-Gopalan C, Stock CA, Baker RF,Kemp JD, Hall TC (1983) Phaseolin gene from bean is ex-pressed after transfer to sunflower via tumor-inducing plasmidvectors. Science 222: 476-482
12. Murashige T, Skoog F (1962) A revised medium for rapid growthand bioassays with tobacco tissue cultures. Physiol Plant 15:473-497
13. Negrutiu I, Shillito R, Potrykus I, Biasini G, Sala F (1987)Hybrid genes in the analysis of transformation conditions. I.Setting up a simple method for direct gene transfer in plantprotoplasts. Plant Mol Biol 8: 363-373
14. Riggs CD, Bates GW (1986) Stable transformation of tobaccoby electroporation: evidence for plasmid concatenation. ProcNatl Acad Sci USA 83: 5602-5606
15. Robert LS, Thompson RD, Flavell RB (1989) Tissue-specificexpression of a wheat high molecular weight gene in transgenictobacco. Plant Cell 1: 569-578
16. Rogers SO, Bendich AJ (1985) Extraction of DNA from milli-gram amounts of fresh, herbarium and mummified planttissues. Plant Mol Biol 5: 69-76
17. Sanders PR, Winter JA, Barnason AR, Rogers SG, Fraley RT(1987) Comparison of cauliflower mosaic virus 35S and no-paline synthase promoters in transgenic plants. Nucleic AcidsRes 15: 1543-1558
18. Shillito RD, Paszkowski J, Potrykus I (1983) Agarose platingand a bead type culture technique enable and stimulate devel-opment of protoplast-derived colonies in a number of plantspecies. Plant Cell Report 2: 244-247
19. Shimamoto K, Terada R, Izawa T, Fujimoto H (1989) Fertiletransgenic rice plants regenerated from transformed proto-plasts. Nature 338: 274-276
20. Southern EM (1975) Detection of specific sequences amongDNA fragments separated by gel electrophoresis. J Mol Biol98: 503-517
21. Toriyama KF, Arimoto Y, Uchimiya H, Hinata K (1988) Trans-genic rice plants after direct gene transfer into protoplasts. Bio/technol 6: 1072-1074
22. Uchimiya H, Fushimi T, Hashimoto H, Harada H, Syono K,Sugawara Y (1986) Expression of a foreign gene in callusderived from DNA treated protoplasts of rice (Oryza sativaL.). Mol Gen Genet 204: 204-207
23. Weising 0, Schell J, Kahl G (1988) Foreign genes in plants:transfer, structure, expression, and application. Annu RevGenet 22: 421-477
24. Yang H, Zhang HM, Davey MR, Mulligan BJ, Cocking EC(1988) Production of kanamycin resistant rice tissue followingDNA uptake into protoplasts. Plant Cell Report 7: 421-425
25. Zhang HM, Yang H, Rech EL, Golds TJ, Davis AS, MulliganBJ, Cocking EC, Davey MR (1988) Transgenic rice plantsproduced by electroporation-mediated plasmid uptake intoprotoplasts. Plant Cell Rep 7: 379-384
26. Zhang W, Wu R (1988) Efficient regeneration oftransgenic plantsfrom rice protoplasts and correctly regulated expression of theforeign gene in the plants. Theor Appl Genet 76: 835-840
863
www.plantphysiol.orgon July 14, 2018 - Published by Downloaded from Copyright © 1990 American Society of Plant Biologists. All rights reserved.





























