Embed Size (px)
Citation preview

A petiole-galling insect herbivore decelerates leaf
lamina litter decomposition rates
Christopher J. Frost†,1,2,3,4, Jennifer M. Dean1,4, Erica C. Smyers4, Mark C. Mescher1,4, John
E. Carlson1,2,3,5, Consuelo M. De Moraes1,4 and John F. Tooker*,1,4
1Center for Chemical Ecology, Pennsylvania State University, University Park, Pennsylvania 16802, USA; 2School of
Forest Resources, Pennsylvania State University, University Park, Pennsylvania 16802, USA; 3Schatz Center for Tree
Molecular Genetics, Pennsylvania State University, University Park, Pennsylvania 16802, USA; 4Department of Entomol-
ogy, Pennsylvania State University, University Park, Pennsylvania 16802, USA; and 5Department of Bioenergy Science
and Technology (WCU), Chonnam National University, 333 Yongbongro, Buk-gu, Gwangju 500-757, Korea
Summary
1. Herbivore-mediated changes in leaf-litter chemistry are often considered responsible for alter-
ing litter decomposition rates, but such chemical changes often co-occur with other factors such
as physical alteration of leaf material that also influence decomposition rates. We attempted to
disentangle these effects using the poplar petiole gall moth (Ectoedemia populella Brusk), which
forms galls on petioles at the base of the leaf lamina but does not alter leaf morphology. Thus,
differences in leaf decomposition rates between galled and ungalled leaves should be explained
by gall-mediated changes in leaf chemistry.
2. Petiole galling decelerated leaf lamina litter decomposition in two Populus host species, but in
temporally distinct ways. In Populus granidentata, galling decelerated decomposition by 7%
after 4 months. After 12 and 18 months, Populus tremuloides litter decomposition rates were
12% and 17% lower, respectively, in lamina tissue whose petiole had been galled relative to ung-
alled. On average, the petiole galler increased leaf lamina nitrogen concentrations by 17%,
decreased tannin concentrations from 37% to 53% and decreased tannin-binding capacity by
11% and 37% in P. grandidentata and P. tremuloides, respectively. These changes would be
expected to increase, rather than decrease, decomposition rates.
3. Unlike other insect herbivores guilds that have variable effects on litter decomposition in
direction and magnitude, all gall insects studied to date have decelerated leaf-litter decomposi-
tion. This consistent effect of galling on decomposition provides a framework for deciphering a
fundamental aspect of insect herbivory on a critical ecosystem process.
4. We used a gall-inducing moth with a distinctive natural history to confirm the role of herbi-
vore-mediated litter chemistry in leaf-litter decomposition dynamics. Moreover, we advance the
hypothesis that gall-induced defensive manipulations that protect a host plant from injury by
other herbivores lead to decelerated litter decomposition.
Key-words: Ectoedemia, leaf chemistry, leaf-litter decomposition, plant–herbivore interactions,
Populus
Introduction
The process of decomposing dead plant material facilitates
the recycling of mineral nutrients and organic matter essen-
tial for biological activity in most terrestrial ecosystems
(Parton et al. 2007). Senesced leaf litter is an abundant,
ubiquitous example of such material, and variation in leaf-
litter quality as a substrate can have demonstrable effects
on decomposition processes and thus terrestrial nutrient
availability. As many plant species presumably increase
their Darwinian fitness by altering foliar quality in response
to herbivores (Karban & Baldwin 1997), the potential for
herbivores to indirectly influence litter quality – and thus
decomposition rates – has long been considered plausible
(Choudhury 1988). While a number of studies have shown
clear effects of herbivores on litter decomposition (Findlay
et al. 1996; Belovsky & Slade 2000; Chapman et al. 2003;
Schweitzer et al. 2005b; Chapman, Schweitzer & Whitham
*Correspondence author. E-mail: [email protected]†Present address. Warnell School of Forest Resources, University of
Georgia, Athens, Georgia 30601, USA.
� 2012 The Authors. Functional Ecology � 2012 British Ecological Society
Functional Ecology 2012 doi: 10.1111/j.1365-2435.2012.01986.x

2006; Crutsinger et al. 2008; Frost & Hunter 2008; Kurokawa
& Nakashizuka 2008), constructing a more detailed theoret-
ical framework has been difficult because herbivory can
accelerate or decelerate decomposition depending in part on
whether secondary metabolites are repressed or augmented,
what type of physical damage occurs, the type of herbivory
suffered or the ecosystem in which the decomposition is
measured. Moreover, determining the importance of her-
bivory-induced changes in leaf chemistry for decomposition
rates is often complicated by the co-occurrence of other
herbivore-related factors that can also alter litter decompo-
sition. For example, herbivory may induce changes in leaf-
drop phenology that results in herbivore-affected litter
entering the detrital system in a different condition than
undamaged litter (Chapman et al. 2003). Of equal impor-
tance, most foliar herbivores physically damage leaf tissue
during feeding, and such damage can alter access to the lit-
ter substrate (Findlay et al. 1996; Cornelissen et al. 1999;
Perez-Harguindeguy et al. 2000; Ostertag, Scatena & Silver
2003). Insect herbivore species that influence host-plant
chemistry but only minimally alter other aspects of a host
plant, such as some species of gall insects, offer an ideal
opportunity to explore the effects of herbivore-induced leaf
chemistry on leaf-litter decomposition apart from other
co-occurring effects.
Gall-inducing insects are herbivores that force their host
plants to produce a tumour-like growth that provides the
insect with food and shelter, usually at the expense of plant
growth and ⁄or reproduction. These insects have evolved inti-
mate relationships with their host plants and an unparalleled
ability to influence host-plant morphology and physiology
(Larson 1998; Stone & Schonrogge 2003). Gall insects com-
monly modify host-plant chemistry, often altering concentra-
tions of plant secondary metabolites for their own purposes
(Weis & Abrahamson 1986; Nyman & Julkunen-Tiitto 2000;
Tooker, Koenig & Hanks 2002; Allison & Schultz 2005). For
example, high concentrations of secondary metabolites such
as phenolics tend to be localized in gall exteriors, where they
presumably provide protection against natural enemies, while
the inner nutritive tissue on which the gall insect feeds is lar-
gely or entirely devoid of such potentially toxic compounds
(Nyman & Julkunen-Tiitto 2000; Allison & Schultz 2005).
Moreover, gall insects also variably influence the chemistry of
plant tissue that is not part of the gall itself, although investi-
gations thus far have been confined to neighbouring tissues
(Tooker et al. 2008; Cooper & Rieske 2009); such systemic
effects on chemistry may also influence decomposition
dynamics of those tissues.
A potential consequence of gall insect-mediated alterations
in host-plant chemistry is the modification of rates of decom-
position of senesced or dead plant parts. Such effects have
been demonstrated using the aphid Pemphigus betae and the
midge Rhopalomyia solidaginis, two disparate gall-inducing
taxa; these insect species induced changes that altered rates of
leaf-litter decomposition (Schweitzer et al. 2005b; Crutsinger
et al. 2008). Yet, both of these species alter leaf morphology
to some degree by forming their gall on the leaf lamina
(P. betae) or by radically altering leaf growth patterns
(R. solidaginis) (Fig. S1, Supporting information). As a
result, these systems have shown clearly that gall insects have
ecosystem-level effects, but neither system necessarily disen-
tangles chemical and morphological factors influencing litter
decomposition.
Here, we describe how herbivory by larvae of the poplar
petiole gall moth (Ectoedemia populella Busck.) influences
leaf-litter chemistry and decomposition in two species of Pop-
ulus. Ectoedemia are monotrysian, nepticulid moths that
include some of the smallest known lepidopterans; adults of
E. populella have c. 6-mm wingspans (Wilkinson & Scoble
1979). The family consists primarily of leaf miners; only a few
Ectoedemia species consume bark, buds, or – in the case of
E. populella – induce galls (Wilkinson & Scoble 1979). Leaf-
mining Ectoedemia trace at least to the mid-Cretaceous era
97 million years ago (Labandeira et al. 1994); gall-inducing
Ectoedemia on Populus trace to the Miocene era, some 5–
17 million years ago (Madler 1936). Importantly,E. populella
form galls at the junction of the petiole and the lamina caus-
ing no observable morphological difference in the lamina tis-
sue itself (Fig. S2, Supporting information). Moreover, the
phenology of the moth is offset with its hosts such that the
adults emerge and oviposit in May after leaves of their hosts
have fully expanded foliage (Wilkinson& Scoble 1979). Thus,
E. populella galls have limited or no influence on lamina
growth and development, although they may influence lam-
ina quality. Larvae emerge and pupate in October and over-
winter in the soil, so larvae do not reside in decomposing
galled leaf litter. Little else is known about the ecology of
E. populella, including the direct effects of the gall on leaf
lamina metabolite profiles associated with the galled petiole.
However, there are at least two reasons to hypothesize that
E. populella will influence decomposition possibly by altering
laminar chemistry. First, the gall envelopes the petiole, forc-
ing all assimilates and other transported materials through
vasculature that has been altered by the gall. Second, E. pop-
ulella deposits its frass inside the gall in a tight packet that
remains in the gall after leaf senescence; frass, which is nutri-
ent rich, is known to influence ecosystem processes in other
systems (Frost & Hunter 2007; Madritch, Donaldson &
Lindroth 2007).
Throughout its range, E. populella forms galls on big-
toothed aspen (Populus grandidentata) and quaking aspen
(Populus tremuloides) (Wilkinson & Scoble 1979). This
makes the Populus ⁄Ectoedemia system well suited to explore
the independent effects of host species and galling. As plant
diversity at different levels can affect decomposition rates
(Madritch & Hunter 2002; Ball et al. 2008) – and the effects
of herbivores on these rates (Schweitzer et al. 2005b) – we
tested the hypothesis that E. populella galling influences leaf-
litter decomposition dynamics using naturally senesced
leaves from P. grandidentata and P. tremuloides separately.
This allowed us to examine variation of E. populella-
mediated changes to leaf-litter decomposition rates, chemis-
try and nutrient release dynamics between host species over
the course of 18 months.
� 2012 The Authors. Functional Ecology � 2012 British Ecological Society, Functional Ecology
2 C. J. Frost et al.

Materials and methods
Leaf litter from P. grandidentata and P. tremuloides was collected
separately from the ground during natural leaf senescence (October
2006) from three separate locations in Centre County, PA and pooled
by plant species. The two aspen species tend to drop their leaves over
a window of generally <10 days (Fig. S3, Supporting information).
We did not collect leaves until they naturally dehisced from the trees
to ensure natural levels of leaf senescence. However, we also used
leaves that were freshly fallen and sitting on top of other leaf litter to
ensure that they had no contact with soil, to minimize any potential
‘preprocessing’ of litters prior to the experiment. We observed no
apparent difference in leaf-drop phenology; we confirmed this by
marking a number of leaves and monitoring their drop phenology,
and there was no difference in leaf-drop timing between galled and
ungalled leaves (Fig. S3, Supporting information). Litter from each
species was sorted based on the presence of galls. Leaves were selected
that had little or no visible damage from chewing herbivores to the
leaf tissue itself, as such damage has been shown to affect decomposi-
tion dynamics in Populus and other woody plant species (Findlay
et al. 1996; Belovsky & Slade 2000; Chapman et al. 2003; Chapman,
Schweitzer & Whitham 2006; Frost & Hunter 2008). However, we
cannot exclude the possibility that piercing ⁄ sucking herbivores such
as aphids may have fed on the foliage during the growing season,
although nonewas observed during sampling.
Litters were dried separately in the laboratory at room tempera-
ture, and then, subsamples were selected haphazardly, weighed and
sealed in 15 · 15 cm screen bags (1 · 1-mm mesh-size) as previously
described (Frost & Hunter 2008). We used c. 3 g of litter for each lit-
terbag. Based on the amount of material available, we filled 55–57 lit-
terbags with each of the P. grandidentata galled and ungalled litters
and P. tremuloides ungalled litters; 20 litterbags were made with
P. tremuloides galled litter owing to a limited amount of material.
Following previous convention (Schweitzer et al. 2005b; Crutsinger
et al. 2008), we removed petioles from all galled and ungalled laminas
prior to filling the litterbags but did not remove galls from the lamina
litter, and the Ectoedemia gall body was c. 8% of the total litter mass
in galled samples. Importantly, different decomposition patterns
between the two Populus species and the magnitude of the observed
effects (see Results) suggest that the differences between the mass of
galled and ungalled litters were not artifacts of the gall body itself.
We then established a field plot in January 2007 in the Scotia
Range near State College, PA (40�47¢0Æ56¢¢N; 77�57¢20Æ56¢¢W). The
site was a mixed-hardwood stand containingmatureP. grandidentata
and P. tremuloides. The field site consisted of 15 replicated plots
(0Æ5 · 2 m per plot). We assigned randomly one litterbag per treat-
ment per collection date to each plot. Thus, fifteen replicates per date
were established for each of the P. grandidentata galled and ungalled
and P. tremuloides ungalled litters. Five replicates per date were
established for the P. tremuloides galled litters. For the initial sam-
ples, there were 9–12 bags forP. grandidentata galled and ungalled lit-
ters and P. tremuloides ungalled litters and five bags for the
P. tremuloides galled litter.
One set of litterbags was collected immediately to measure initial
litter quality, while the remaining litterbags were collected after 4, 12
and 18 months in the field. Litterbags containing initial litter samples
were brought to the field, established at the site and then immediately
collected and returned to the laboratory. During collections, the stak-
ing flag was carefully removed, debris on the outside of each collected
bag was brushed from the surface and care was taken not to disturb
remaining litterbags. Litterbags were transported to the laboratory at
ambient temperatures and then dried at 65 �C for 48 h. Samples were
then weighed, and the lamina litter was separated from the gall body
in the samples that were galled. The lamina litter tissue only was then
ground to a fine powder in a ball mill, and the resulting powder was
analysed for total C (mg g)1), total N (mg g)1), C ⁄N ratios, tannins
(mg g)1) and fibre content (lignin, cellulose, hemicellulose; mg g)1).
Subsamples were ashed (550 �C, 5 h), and data are reported relative
to ash-free dry mass (AFDM). Thus, the chemical analyses represent
only the ungalled lamina portions of the leaf-litter tissue.
Total C and N were measured with a Carlo Erba 1500N total
CHN analyzer (Carlo Erba Instrumentazione, Milan, Italy). Fibre
concentrations were determined by sequential acid digestions using
an ANKOM A200 Fiber Analyzer according to manufacturer’s
instructions (ANKOM Technology, Macedon, NY, USA). Tannin
concentrations and protein-binding capacity were assayed colorimet-
rically following well-established methods (Bate-Smith 1977a,b) after
three rounds of initial extraction with 70 ⁄ 30% acetone ⁄water with1 mM ascorbic acid and subsequent removal of the acetone under vac-
uum evaporation (Frost & Hunter 2008). Condensed tannin concen-
trations were assayed following acid depolymerization in a polar
solvent (N-butanol) at 100 �C (Hagerman & Butler 1980; Hagerman,
Rice & Ritchard 1998), and hydrolyzable tannin concentrations were
measured using the standard potassium iodate assay (Rossiter,
Schultz & Baldwin 1988). We also measured protein-binding capacity
of the leaf tannin extracts using the protein precipitatable phenolics
assay, which provides a realistic assessment of the biological activity
of the tannins. Briefly, protein-binding tannins form a precipitate
complex with the substrate protein bovine serum albumin; the precip-
itate is re-dissolved in triethanolamine ⁄ sodium dodecyl sulphate and
tannins quantified (mg protein-binding tannin per g dry weight) spec-
trophotometrically after adding ferric chloride (Hagerman & Butler
1978). Standards for all phenolic assays were generated from a pooled
mix of undecomposed leaf litter from all the individual experimental
trees that was extracted exhaustively with 70 ⁄ 30% acetone ⁄waterwith 1 mM ascorbic acid similar to the individual samples (Hagerman
& Butler 1989; Hall et al. 2006; Ball et al. 2008; Frost & Hunter
2008). This pooled aqueous extract was then frozen and lyophilized,
and an weighed aliquot of the resulting powder was resuspended in
water and used to generate standard curves for all tannin assays
(Madritch & Hunter 2002, 2004; Hunter & Forkner 2004; Hall et al.
2006; Ball et al. 2008; Frost &Hunter 2008).
Data were analysed using PROC MIXED in SAS (Littell, Stroup &
Freund 2002) using tree species and gall presence as main effects and
plot as a random effect. Data reported on litter AFDM remaining
were arcsine square root transformed, and all other data were log
transformed prior to analysis to satisfy assumptions of ANOVA.
Results
Presence of the petiole gall reduced decomposition rates of
the big-toothed aspen and quaking aspen leaf litters, but in
temporally distinct ways (Fig. 1). Both galling and tree spe-
cies had significant main effects on rates of AFDM loss (spe-
cies F1,163 = 12Æ90, P < 0Æ001; gall F1,163 = 7Æ62,P = 0Æ006); however, only galling influenced AFDM loss
over time (date*gall F3,149 = 3Æ05, P = 0Æ030; date*speciesF3,150 = 2Æ19, P = 0Æ091). Importantly, galling affected
decomposition differently between tree species (date*spe-
cies*gall F3,149 = 2Æ89, P = 0Æ038): galling decelerated the
‘early’ stages of mass loss in P. grandidentata litter and the
‘late’ stages in P. tremuloides litter. After 4 months, AFDM
� 2012 The Authors. Functional Ecology � 2012 British Ecological Society, Functional Ecology
Galling effects on leaf-litter decomposition 3
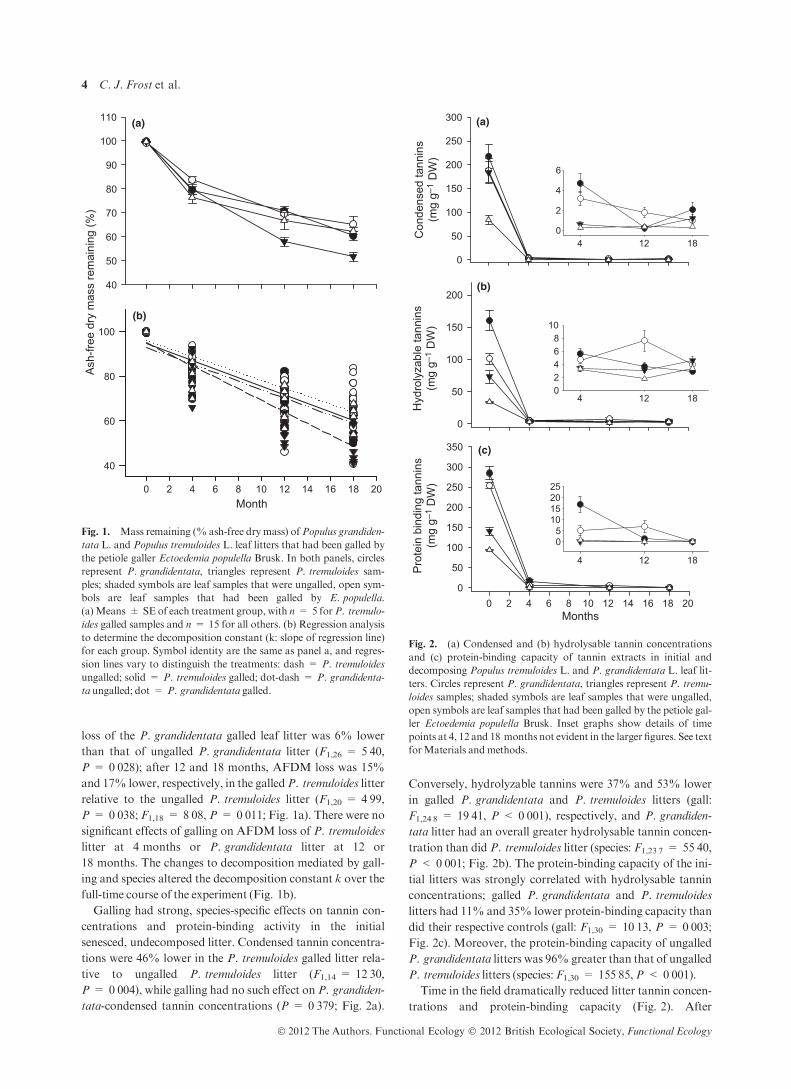
loss of the P. grandidentata galled leaf litter was 6% lower
than that of ungalled P. grandidentata litter (F1,26 = 5Æ40,P = 0Æ028); after 12 and 18 months, AFDM loss was 15%
and 17% lower, respectively, in the galledP. tremuloides litter
relative to the ungalled P. tremuloides litter (F1,20 = 4Æ99,P = 0Æ038; F1,18 = 8Æ08, P = 0Æ011; Fig. 1a). There were nosignificant effects of galling on AFDM loss of P. tremuloides
litter at 4 months or P. grandidentata litter at 12 or
18 months. The changes to decomposition mediated by gall-
ing and species altered the decomposition constant k over the
full-time course of the experiment (Fig. 1b).
Galling had strong, species-specific effects on tannin con-
centrations and protein-binding activity in the initial
senesced, undecomposed litter. Condensed tannin concentra-
tions were 46% lower in the P. tremuloides galled litter rela-
tive to ungalled P. tremuloides litter (F1,14 = 12Æ30,P = 0Æ004), while galling had no such effect on P. grandiden-
tata-condensed tannin concentrations (P = 0Æ379; Fig. 2a).
Conversely, hydrolyzable tannins were 37% and 53% lower
in galled P. grandidentata and P. tremuloides litters (gall:
F1,24Æ8 = 19Æ41, P < 0Æ001), respectively, and P. grandiden-
tata litter had an overall greater hydrolysable tannin concen-
tration than did P. tremuloides litter (species: F1,23Æ7 = 55Æ40,P < 0Æ001; Fig. 2b). The protein-binding capacity of the ini-
tial litters was strongly correlated with hydrolysable tannin
concentrations; galled P. grandidentata and P. tremuloides
litters had 11% and 35% lower protein-binding capacity than
did their respective controls (gall: F1,30 = 10Æ13, P = 0Æ003;Fig. 2c). Moreover, the protein-binding capacity of ungalled
P. grandidentata litters was 96% greater than that of ungalled
P. tremuloides litters (species: F1,30 = 155Æ85,P < 0Æ001).Time in the field dramatically reduced litter tannin concen-
trations and protein-binding capacity (Fig. 2). After
Ash
-free
dry
mas
s re
mai
ning
(%)
40
50
60
70
80
90
100
110
Month0 2 4 6 8 10 12 14 16 18 20
40
60
80
100
(a)
(b)
Fig. 1. Mass remaining (% ash-free dry mass) ofPopulus grandiden-
tata L. and Populus tremuloides L. leaf litters that had been galled by
the petiole galler Ectoedemia populella Brusk. In both panels, circles
represent P. grandidentata, triangles represent P. tremuloides sam-
ples; shaded symbols are leaf samples that were ungalled, open sym-
bols are leaf samples that had been galled by E. populella.
(a)Means ± SE of each treatment group, with n = 5 forP. tremulo-
ides galled samples and n = 15 for all others. (b) Regression analysis
to determine the decomposition constant (k: slope of regression line)
for each group. Symbol identity are the same as panel a, and regres-
sion lines vary to distinguish the treatments: dash = P. tremuloides
ungalled; solid = P. tremuloides galled; dot-dash = P. grandidenta-
ta ungalled; dot = P. grandidentata galled.
Months0 2 4 6 8 10 12 14 16 18 20
Pro
tein
bin
ding
tann
ins
(mg
g–1
DW
)
0
50
100
150
200
250
300
350
Con
dens
ed ta
nnin
s(m
g g–
1 D
W)
0
50
100
150
200
250
300
Hyd
roly
zabl
e ta
nnin
s(m
g g–
1 D
W)
0
50
100
150
200
(a)
(b)
(c)
0
2
4
6
4 12 18
4 12 1802468
10
4 12 18
05
10152025
Fig. 2. (a) Condensed and (b) hydrolysable tannin concentrations
and (c) protein-binding capacity of tannin extracts in initial and
decomposing Populus tremuloides L. and P. grandidentata L. leaf lit-
ters. Circles represent P. grandidentata, triangles represent P. tremu-
loides samples; shaded symbols are leaf samples that were ungalled,
open symbols are leaf samples that had been galled by the petiole gal-
ler Ectoedemia populella Brusk. Inset graphs show details of time
points at 4, 12 and 18 months not evident in the larger figures. See text
forMaterials andmethods.
� 2012 The Authors. Functional Ecology � 2012 British Ecological Society, Functional Ecology
4 C. J. Frost et al.
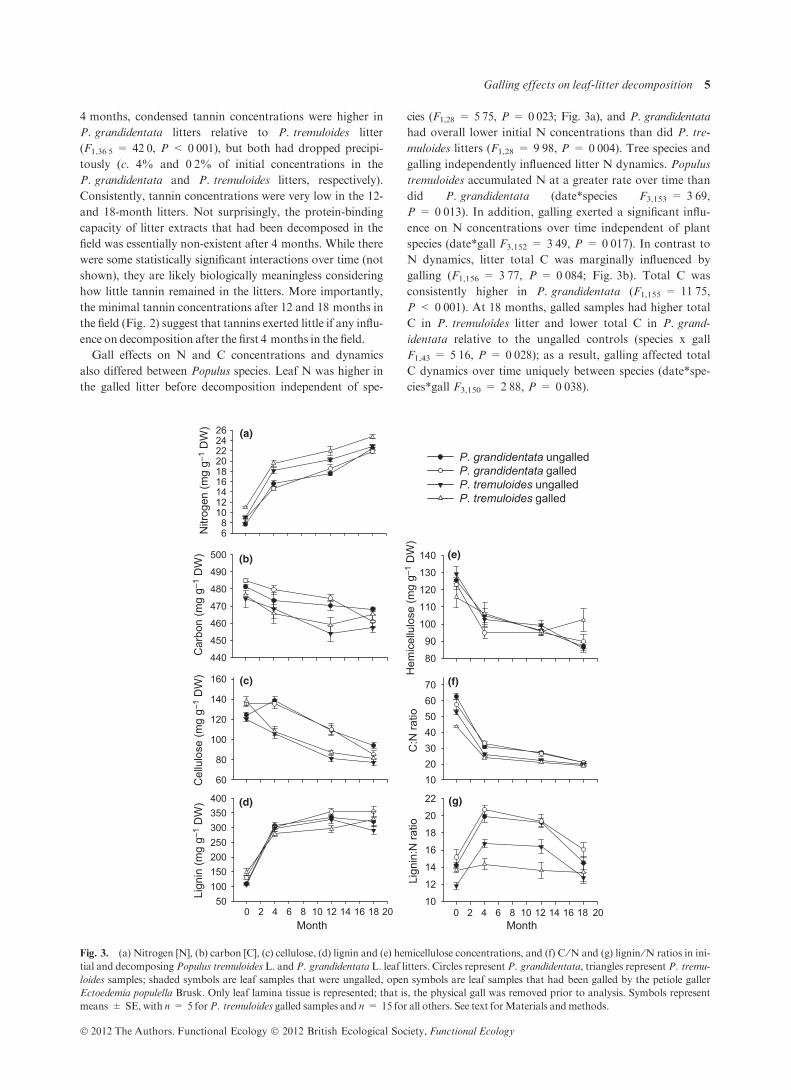
4 months, condensed tannin concentrations were higher in
P. grandidentata litters relative to P. tremuloides litter
(F1,36Æ5 = 42Æ0, P < 0Æ001), but both had dropped precipi-
tously (c. 4% and 0Æ2% of initial concentrations in the
P. grandidentata and P. tremuloides litters, respectively).
Consistently, tannin concentrations were very low in the 12-
and 18-month litters. Not surprisingly, the protein-binding
capacity of litter extracts that had been decomposed in the
field was essentially non-existent after 4 months. While there
were some statistically significant interactions over time (not
shown), they are likely biologically meaningless considering
how little tannin remained in the litters. More importantly,
the minimal tannin concentrations after 12 and 18 months in
the field (Fig. 2) suggest that tannins exerted little if any influ-
ence on decomposition after the first 4 months in the field.
Gall effects on N and C concentrations and dynamics
also differed between Populus species. Leaf N was higher in
the galled litter before decomposition independent of spe-
cies (F1,28 = 5Æ75, P = 0Æ023; Fig. 3a), and P. grandidentata
had overall lower initial N concentrations than did P. tre-
muloides litters (F1,28 = 9Æ98, P = 0Æ004). Tree species and
galling independently influenced litter N dynamics. Populus
tremuloides accumulated N at a greater rate over time than
did P. grandidentata (date*species F3,153 = 3Æ69,P = 0Æ013). In addition, galling exerted a significant influ-
ence on N concentrations over time independent of plant
species (date*gall F3,152 = 3Æ49, P = 0Æ017). In contrast to
N dynamics, litter total C was marginally influenced by
galling (F1,156 = 3Æ77, P = 0Æ084; Fig. 3b). Total C was
consistently higher in P. grandidentata (F1,155 = 11Æ75,P < 0Æ001). At 18 months, galled samples had higher total
C in P. tremuloides litter and lower total C in P. grand-
identata relative to the ungalled controls (species x gall
F1,43 = 5Æ16, P = 0Æ028); as a result, galling affected total
C dynamics over time uniquely between species (date*spe-
cies*gall F3,150 = 2Æ88, P = 0Æ038).
Nitr
ogen
(mg
g–1
DW
)
68
101214161820222426
Car
bon
(mg
g–1
DW
)
440
450
460
470
480
490
500
(a)
(b)
Cel
lulo
se (m
g g–
1 D
W)
60
80
100
120
140
160
Month
Lign
in (m
g g–
1 D
W)
50100150200250300350400
Hem
icel
lulo
se (m
g g–
1 D
W)
80
90
100
110
120
130
140 (e)
Month0 2 4 6 8 10 12 14 16 18 20 0 2 4 6 8 10 12 14 16 18 20
Lign
in:N
ratio
10
12
14
16
18
20
22
C:N
ratio
10203040506070
P. grandidentata ungalledP. grandidentata galledP. tremuloides ungalledP. tremuloides galled
(g)(d)
(c) (f)
Fig. 3. (a) Nitrogen [N], (b) carbon [C], (c) cellulose, (d) lignin and (e) hemicellulose concentrations, and (f) C ⁄N and (g) lignin ⁄N ratios in ini-
tial and decomposing Populus tremuloides L. and P. grandidentata L. leaf litters. Circles represent P. grandidentata, triangles represent P. tremu-
loides samples; shaded symbols are leaf samples that were ungalled, open symbols are leaf samples that had been galled by the petiole galler
Ectoedemia populella Brusk. Only leaf lamina tissue is represented; that is, the physical gall was removed prior to analysis. Symbols represent
means ± SE, with n = 5 forP. tremuloides galled samples and n = 15 for all others. See text forMaterials andmethods.
� 2012 The Authors. Functional Ecology � 2012 British Ecological Society, Functional Ecology
Galling effects on leaf-litter decomposition 5

Concentrations of cell wall constituents were influenced by
galling and plant species. Cellulose and lignin concentrations
in senesced, undecomposed leaf tissue were 12% and 29%
higher in galled litters relative to ungalled litters, respectively,
independent of plant species (cellulose: F1,28 = 21Æ51,P < 0Æ001; lignin: F1,28 = 22Æ97, P < 0Æ001; Fig. 3c,d).
During decomposition, cellulose concentrations segregated
by plant species (species: F1,158 = 39Æ95, P < 0Æ001) and the
initial effect of galling were lost (gall: F1,159 = 0Æ85,P = 0Æ359). The two species also varied in the rate of cellu-
lose loss (date*species: F3,149 = 4Æ90, P = 0Æ003). In con-
trast, lignin concentrations and their rate of change in the
decomposing litter were influenced by galling (gall:
F1,158 = 8Æ27, P = 0Æ005; date*gall: F3,149 = 6Æ61,P < 0Æ001). The date*gall interactive effect seemed driven by
a significant gall effect at 18 months (F1,32Æ6 = 4Æ27,P = 0Æ047) that was not present at 4 or 12 months. Hemicel-
lulose concentrations were not affected by either tree species
or galling (Fig. 3e).
Nutrient ratios can play an important role in shaping litter
decomposition dynamics. The C ⁄N ratios were driven by N
concentrations and, P. tremuloides had lower C ⁄N ratios
(F1,161 = 99Æ45, P < 0Æ001) and a more rapid reduction in
C ⁄N ratios over time (date*species F3,152 = 3Æ78,P = 0Æ012)than did P. grandidentata (Fig. 3f). In other words, assuming
that C ⁄N ratios are predictive of litter quality, P. tremuloides
was of higher quality to decomposers than was P. grandentata.
Galling had a relatively small effect on C ⁄N ratios. In con-
trast, the influence of galling on lignin ⁄N ratios was pro-
nounced (date*gall F3,147 = 3Æ17, P = 0Æ026; Fig. 3g). Eventhough there was no statistically significant date*spe-
cies*gall interaction (P = 0Æ278), galling clearly affected lig-
nin ⁄N ratios in P. tremuloides more than in
P. grandidentata: there were significant species*gall interac-
tions at 4 and 12 months (4 months F1,45 = 6Æ91,P = 0Æ011; 12 months F1,35Æ4 = 5Æ56, P = 0Æ024). In fact,
lignin ⁄N ratios were unchanged over time in galledP. tremu-
loides litters (Fig. 3g).
Discussion
A long-standing hypothesis in plant–herbivore ecology is that
herbivores influence rates of leaf-litter decomposition via
changes in litter chemical quality (Grace 1986; Choudhury
1988).While abiotic factors such as temperature andmoisture
clearly drive decomposition dynamics across biomes (Trofy-
mow et al. 2002; Wall et al. 2008), the influence of substrate
quality is most pronounced on spatial scales where tempera-
ture and moisture conditions have relatively minimal varia-
tion (Swift, Heal & Anderson 1979). That is, substrate quality
likely matters in most mixed-stand terrestrial forests. Yet,
herbivores can influence litter quality in a number of ways. In
all cases of chewing or piercing ⁄ sucking herbivores, physical
damage to the lamina tissue occurs regardless of herbivore-
induced changes to lamina chemistry and may complicate the
effects of herbivore-induced chemistry per se. As a drastic
example of physical damage, experimental crushing of leaf lit-
ter dramatically increases decomposition rates by facilitating
access to the substrate (Madritch & Hunter 2003). Such
substrate access increases the more a leaf is consumed by a
chewing herbivore, which causes physical damage to the leaf
tissue (Cornelissen et al. 1999; Perez-Harguindeguy et al.
2000). The Ectoedemia ⁄Populus system described here pro-
vides an opportunity to examine the role of herbivore-altered
plant chemistry apart from other factors associated with her-
bivory because the galler does not physically manipulate the
lamina. Our results indicate that an insect herbivore that has
little influence on leaf lamina development and does not
physically damage the lamina can nonetheless clearly
influence litter decomposition and nutrient release rates.
Herbivore-mediated changes in litter chemistry alone are suf-
ficient to alter leaf-litter decomposition rates and, presum-
ably, other ecosystem processes responsive to those rates.
Although decomposition dynamics differed between tree
species, E. populella petiole galling consistently decelerated
leaf lamina litter decomposition. Deceleration of decomposi-
tion theoretically results from low substrate quality; this typi-
cally implies relatively lower concentrations of beneficial
components such as N and relatively higher concentrations of
detrimental components such as tannins and lignin, each of
which has been shown to influence decomposition via effects
on decomposer organisms (Northup, Dahlgren & McColl
1998; Maie et al. 2003; Hattenschwiler & Gasser 2005). This
general model held for the species main effect in our system:
P. grandidentata litter had lower total N, higher tannin con-
centrations and binding activity (but no difference in initial
litter lignin) and decomposed more slowly than did P. tremu-
loides litter. However, the presence of the gall confounded this
relationship. The relatively higher N concentrations in galled
P. tremuloides and relatively lower tannin concentrations and
binding capacity in galled tissues would typically be predicted
to accelerate, not decelerate, decomposition. Evidently some
other metabolite(s) associated with galled lamina trumped
the higher N concentrations. Moreover, the gall-mediated
reduction in initial tannin concentrations and binding capac-
ity suggests that tannins are not involved. One possibility is
that galling altered leaf toughness, which is has been shown to
affect decomposition rates (Cornelissen et al. 1999; Perez-
Harguindeguy et al. 2000). We did not measure leaf tough-
ness directly, but P. grandidentata leaves were obviously
tougher than were P. tremuloides and, as expected, P. grand-
identata decomposed more slowly. There were no such obvi-
ous phenotypic differences in leaf toughness in galled vs.
ungalled litter within either species that would appear to
account for the differential decomposition rates.
The result that Ectoedemia galling decelerated lamina litter
decomposition is consistent with effects from other gall
insects. In fact, gall insects from diverse lineages (i.e. a social
hemipteran, a cecidomyiid dipteran and the microlepidopter-
an studied here) independently decelerate leaf-litter decompo-
sition under field conditions (Schweitzer et al. 2005b;
Crutsinger et al. 2008). These effects presumably result from
modification of foliar chemical composition as our data sug-
gest, but may also include morphological changes in the other
� 2012 The Authors. Functional Ecology � 2012 British Ecological Society, Functional Ecology
6 C. J. Frost et al.

systems. Indeed, gall insects may have a larger relative influ-
ence on decomposition dynamics than other insect–herbivore
guilds because of their intimate host-plant associations and
their ability to manipulate the molecular physiology of their
host (Schweitzer et al. 2005b). However, other herbivore
guilds can also influence litter decomposition. In some cases,
chewing and piercing-sucking herbivores have accelerated
decomposition (Ritchie, Tilman & Knops 1998; Chapman
et al. 2003); in other cases, herbivory did not influence mass
loss despite significant differences in initial litter quality (Hall
et al. 2006; Frost & Hunter 2008). Based on the three galling
systems studied, the magnitude of galling-herbivore effects on
decomposition relative to other insect–herbivore guilds may
not be as important as their direction: galling modifies leaf
quality in a manner that consistently decelerates leaf-litter
decomposition independent of the identity of the gall insect
or host-plant species.
Attributes that are common to gall insects may provide a
theoretical framework for the consistently observed gall-
mediated deceleration of litter decomposition that is based on
modified litter chemistry. In forcing the host plant to con-
struct an enclosed domicile with a fortified exterior and nutri-
tive interior, many gall insects are less vulnerable to predators
yet have access to an ideal food source during development.
One limitation of galling is that the insect depends on the suc-
cess of the host plant in a manner that is distinct from other
herbivore guilds. While chewing and piercing-sucking herbi-
vores can often search for new suitable host plants, the sessile
nature of gall insects mandates that they have a vested interest
in protecting their host plants. This suggests that gall insects
may manipulate their host plants to be less suitable to other
herbivores while evading host defenses themselves (Tooker &
De Moraes 2007; Stireman & Cipollini 2008; Tooker et al.
2008). Indeed, some gall insect activity can increase concen-
trations of defensive metabolites systemically in ungalled
parts of the plant, which presumably protects the resource
from other forms of herbivory (Pascual-Alvarado et al. 2008;
Cooper & Rieske 2009; Tooker & De Moraes 2009). This
leads to the hypothesis that gall-induced defensive manipula-
tions that protect a host plant from injury by other herbivores
result in decelerated litter decomposition. That is, the same
chemical compounds in foliage that confer resistance to her-
bivores also decelerate decomposition in leaf litter.
Although the life history of E. populella allows us to con-
clude that changes in leaf chemistry play a central role influ-
encing leaf-litter decomposition, gall-mediated differences in
leaf quality that result in the observed decomposition
changes remain unclear. To complicate matters, the effects
of E. populella on leaf-litter decomposition in our study var-
ied between the two host Populus species (Fig. 1), which sug-
gests that the gall-induced chemistry that altered
decomposition dynamics may differ between tree species.
Unfortunately, the gall-induced changes in litter chemistry
in these species are not well known; we therefore focused on
leaf chemistry parameters that are known to affect decompo-
sition (and had demonstrated effects between the two tree
species). Lignin concentrations in our system were 29%
higher initially in galled tissues, and this difference may have
affected the slower decomposition after 4 months in the
P. grandidentata litters. Yet, lignin dynamics over time did
not appear to explain the observed galling effects on decom-
position rates in P. tremuloides litters. In Solidago, lower
total N concentration in the midge-galled litters related to
decelerated decomposition (Crutsinger et al. 2008); Ectoede-
mia-galled litters had higher N concentrations but nonethe-
less exhibited decelerated decomposition. It is possible that
the form of N was different between the galled and ungalled
litters and that this difference was important. In the cotton-
wood system, higher condensed tannin concentrations in
galled litters contributed to decelerated decomposition
(Schweitzer et al. 2005b). In our case, tannin concentrations
and binding capacity were lower in galled tissue and were
essentially lost after 4 months. Although these are com-
monly measured aspects of litter quality, they do not neces-
sarily confer resistance against insect herbivores and do not
appear to explain the decelerating effects of gall-mediated lit-
ter chemistry on litter decomposition. A future line of
enquiry will obviously be needed to explore in both Populus
species the gall-induced changes in chemistry that may be
associated with delayed decomposition.
Other aspects of E. populella life history might also influ-
ence leaf chemistry in ways that alter litter decomposition.
For example, insect frass, which is N rich and remains in the
gall after leaf senescence (See Fig. S1, Supporting informa-
tion) and the higher initial concentration ofN in the galled lit-
ter may have resulted from N absorbed by the plant from the
frass (Frost &Hunter 2007). This frass may therefore provide
a nutrient resource to decomposer microfauna and accelerate
decomposition (Reynolds &Hunter 2001; Fonte & Schowalter
2005; Madritch, Donaldson & Lindroth 2007; Frost & Hun-
ter 2008), although the consistent deceleration of decomposi-
tion in galled tissue does not support this. Nevertheless, our
results do not rule out a role of frass in mediating lamina
chemistry because frass can also contain high levels of plant
defensive compounds (Kopper et al. 2002; Chen et al. 2007).
With E. populella in particular, frass remains within the gall
body and we assume has antimicrobial metabolites to pre-
vent rampant bacterial or fungal growth in the enclosed gall
environment (Tooker and De Moraes 2006). Such metabo-
lites may have extended, indirect effects inhibiting microbial
colonization of leaf litter that presumably would also decel-
erate decomposition.
As a final point, our experimental design isolated litters
from each tree species separately to investigate independent
effects of galling on the two main host tree species, but litters
in natural mixed-stand forests obviously contain a mix of spe-
cies. Litters from different tree species and genotypes decom-
pose at different rates (Fig. 1a), but mixed-species litters can
decompose in an non-additive manner that is not predicted
by the decomposition rates of individual species (Madritch &
Hunter 2003, 2005; Schweitzer et al. 2005a; Ball et al. 2008).
Herbivory may impose an additional factor on mixed-species
litter decomposition dynamics. Herbivory and geno-
typic ⁄ genetic variation are known to interact to affect litter
� 2012 The Authors. Functional Ecology � 2012 British Ecological Society, Functional Ecology
Galling effects on leaf-litter decomposition 7

decomposition (Schweitzer et al. 2005b; Chapman, Schweit-
zer &Whitham 2006), and our results thatEctoedemia galling
differentially affected decomposition dynamics in the two
host Populus species provide further evidence for this. How-
ever, it is unknown how herbivory interacts specifically with
non-additive effects of mixed-species litter decomposition.
So, while the different decomposition dynamics between the
two Populus species in our study is not surprising, the differ-
ential effects of galling on decomposition may be altered
when the two leaf litters are mixed in in ways that may not be
predicted by the galling effects on either species individually.
In summary, we have used a model system with a unique
natural history to demonstrate that insect herbivore-mediated
changes in leaf chemistry can alter leaf-litter decomposition
rates. Further, based on the results from this system and the
other two gall systems, we develop the hypothesis that gall-
mediated deceleration in leaf-litter decomposition results
from induced defensive changes that protect a host plant from
damage from other herbivores, thus increasing gall insect fit-
ness. This hypothesis provides a basis for generalizing the
influence of a large guild of herbivores on a fundamental eco-
system process and should contribute to the theoretical
framework relating species interactions with ecosystem
ecology.
Acknowledgements
We thank Jennifer Schweitzer for insightful comments, Janet Saunders for labo-
ratory assistance, Peter Wilf and Conrad Labandeira for discussions on Ec-
toedemia evolutionary history, Patrick Abbot for Pemphigus photographs
and discussion, Megan Marshall and Tom Richard for use of an ANKOM
fiber analyzer, and Gary Felton and Michelle Peiffer for use of a SpectraMax
plate reader. TomMaddox (Odum School of Ecology Analytical Laboratory)
processed the C ⁄ N samples. The National Research Initiative of the USDA
(#2007-35302-18087 [CJF] and #2006-01823 [JFT]) funded this work.
References
Allison, S.D. & Schultz, J.C. (2005) Biochemical responses of chestnut oak to a
galling cynipid. Journal of Chemical Ecology, 31, 151–166.
Ball, B.A., Hunter, M.D., Kominoski, J.S., Swan, C.M. & Bradford, M.A.
(2008) Consequences of non-random species loss for decomposition dynam-
ics: experimental evidence for additive and non-additive effects. Journal of
Ecology, 96, 303–313.
Bate-Smith, E.C. (1977a) Astringent tannins of Acer species. Phytochemistry,
16, 1421–1426.
Bate-Smith, E.C. (1977b) Detection and determination of ellagitannins. Phyto-
chemistry, 11, 1153–1156.
Belovsky,G.E.& Slade, J.B. (2000) Insect herbivory accelerates nutrient cycling
and increases plant production. Proceedings of the National Academy of Sci-
ences of theUnited States of America, 97, 14412–14417.
Chapman, S.K., Schweitzer, J.A. & Whitham, T.G. (2006) Herbivory differen-
tially alters plant litter dynamics of evergreen and deciduous trees. Oikos,
114, 566–574.
Chapman, S.K., Hart, S.C., Cobb, N.S., Whitham, T.G. & Koch, G.W. (2003)
Insect herbivory increases litter quality and decomposition: an extension of
the acceleration hypothesis.Ecology, 84, 2867–2876.
Chen, H., Gonzales-Vigil, E., Wilkerson, C.G. & Howe, G.A. (2007) Stability
of plant defense proteins in the gut of insect herbivores. Plant Physiology,
143, 1954–1967.
Choudhury, D. (1988) Herbivore induced changes in leaf-litter resource quality:
a neglected aspect of herbivory in ecosystem nutrient dynamics. Oikos, 51,
389–393.
Cooper, W.R. & Rieske, L.K. (2009) Woody stem galls interact with foliage to
affect community associations.Environmental Entomology, 38, 417–424.
Cornelissen, J.H.C., Perez-Harguindeguy, N., Diaz, S., Grime, J.P., Marzano, B.,
Cabido, M., Vendramini, F. & Cerabolini, B. (1999) Leaf structure and
defence control litter decomposition rate across species and life forms in
regional floras on two continents.NewPhytologist, 143, 191–200.
Crutsinger, G.M., Habenicht,M.N., Classen, A.T., Schweitzer, J.A. & Sanders,
N.J. (2008) Galling by Rhopalomyia solidaginis alters Solidago altissima
architecture and litter nutrient dynamics in an old-field ecosystem. Plant and
Soil, 303, 95–103.
Findlay, S., Carreiro, M., Krischik, V. & Jones, C.G. (1996) Effects of
damage to living plants on leaf litter quality. Ecological Applications,
6, 269–275.
Fonte, S.J. & Schowalter, T.D. (2005) The influence of a neotropical herbivore
(Lamponius portoricensis) on nutrient cycling and soil processes.Oecologia,
146, 423–431.
Frost, C.J. & Hunter, M.D. (2007) Recycling of nitrogen in herbivore feces:
plant recovery, herbivore assimilation, soil retention, and leaching losses.
Oecologia, 151, 42–53.
Frost, C.J. &Hunter, M.D. (2008) Insect herbivores and their frass affect Quer-
cus rubra leaf quality and initial stages of subsequent litter decomposition.
Oikos, 117, 13–22.
Grace, J.R. (1986) The influence of gypsymoth on the composition and nutrient
contentof litter fall in aPennsylvania oak forest.ForestScience, 32, 855–870.
Hagerman, A.E. & Butler, L.G. (1978) Protein precipitationmethod for quanti-
tative determination of tannins. Journal of Agricultural and Food Chemistry,
26, 809–812.
Hagerman, A.E. & Butler, L.G. (1980) Condensed tannin purification and
characterization of tannin-associated proteins. Journal of Agricultural and
Food Chemistry, 28, 947–952.
Hagerman, A.E. & Butler, L.G. (1989) Choosing appropriate methods and
standards for assaying tannin. Journal of Chemical Ecology, 15, 1795–1810.
Hagerman, A.E., Rice, M.E. & Ritchard, N.T. (1998) Mechanisms of protein
precipitation for two tannins, pentagalloyl glucose and epicatechin 16 (4–
>8) catechin (procyanidin). Journal of Agricultural and Food Chemistry, 46,
2590–2595.
Hall, M.C., Stiling, P., Moon, D.C., Drake, B.G. & Hunter, M.D. (2006) Ele-
vated CO2 increases the long-term decomposition rate of Quercus myrtifolia
leaf litter.Global Change Biology, 12, 568–577.
Hattenschwiler, S. & Gasser, P. (2005) Soil animals alter plant litter diversity
effects on decomposition. Proceedings of the National Academy of Sciences
of the United States of America, 102, 1519–1524.
Hunter,M.D. &Forkner, R.E. (2004)Hurricane damage influences foliar poly-
phenolics and subsequent herbivory on surviving trees. Ecology, 80, 2676–
2682.
Karban, R. & Baldwin, I.T. (1997) Induced Responses to Herbivory. University
of Chicago Press, Chicago, IL.
Kopper, B.J., Jakobi, V.N., Osier, T.L. & Lindroth, R.L. (2002) Effects of
paper birch condensed tannin on whitemarked tussock moth (Lepidoptera:
Lymantriidae) performance.Environmental Entomology, 31, 10–14.
Kurokawa, H. & Nakashizuka, T. (2008) Leaf herbivory and decomposability
in aMalaysian tropical rain forest.Ecology, 89, 2645–2656.
Labandeira, C.C., Dilcher, D.L., Davis, D.R. & Wagner, D.L. (1994) Ninety-
seven million years of angiosperm-insect association: paleobiological
insights into the meaning of coevolution. Proceedings of the National Acad-
emy of Sciences of the United States of America, 91, 12278–12282.
Larson, K.C. (1998) The impact of two gall-forming arthropods on the photo-
synthetic rates of their hosts.Oecologia, 115, 161–166.
Littell, R.C., Stroup,W.W. & Freund, R.J. (2002) SAS for LinearModels. SAS
Institute, Inc., Cary, NC.
Madler, V.A.K. (1936) Eine Blattgalle an einem vorweltlichn Pappel-Blatt.
Natur und Volk, 66, 271–274.
Madritch, M.D., Donaldson, J.R. & Lindroth, R.L. (2007) Canopy herbivory
can mediate the influence of plant genotype on soil processes through frass
deposition.Soil Biology & Biochemistry, 39, 1192–1201.
Madritch, M.D. & Hunter, M.D. (2002) Phenotypic diversity influences
ecosystem functioning in an oak sandhills community. Ecology, 83,
2084–2090.
Madritch, M.D. & Hunter, M.D. (2003) Intraspecific litter diversity and nitro-
gen deposition affect nutrient dynamics and soil respiration.Oecologia, 136,
124–128.
Madritch,M.D. &Hunter, M.D. (2004) Phenotypic diversity and litter chemis-
try affect nutrient dynamics during litter decomposition in a two species mix.
Oikos, 105, 125–131.
Madritch,M.D.&Hunter,M.D. (2005) Phenotypic variation in oak litter influ-
ences short- and long-term nutrient cycling through litter chemistry. Soil
Biology & Biochemistry, 37, 319–327.
� 2012 The Authors. Functional Ecology � 2012 British Ecological Society, Functional Ecology
8 C. J. Frost et al.

Maie, N., Behrens, A., Knicker, H. & Kogel-Knabner, I. (2003) Changes in the
structure and protein binding capacity of condensed tannins during decom-
position. Soil Biology& Biochemistry, 35, 577–589.
Northup, R.R., Dahlgren, R.A. & McColl, J.G. (1998) Polyphenols as regula-
tors of plant-litter-soil interactions in northern California’s pygmy forest: a
positive feedback?Biogeochemistry, 42, 189–220.
Nyman, T. & Julkunen-Tiitto, R. (2000) Manipulation of the phenolic chemis-
try of willows by gall-inducing sawflies.Proceedings of the National Academy
of Sciences of the United States of America, 97, 13184–13187.
Ostertag, R., Scatena, F.N. & Silver, W.L. (2003) Forest floor decomposition
following hurricane litter inputs in several Puerto Rican forests. Ecosystems,
6, 261–273.
Parton,W., Silver,W.L., Burke, I.C., Grassens, L., Harmon,M.E., Currie,W.S.,
King, J.Y., Adair, E.C., Brandt, L.A., Hart, S.C. & Fasth, B. (2007) Global-
scale similarities in nitrogen release patterns during long-term decomposi-
tion.Science, 315, 361–364.
Pascual-Alvarado, E., Cuevas-Reyes, P., Quesada, M. & Oyama, K. (2008)
Interactions between galling insects and leaf-feeding insects: the role of plant
phenolic compounds and their possible interference with herbivores. Journal
of Tropical Ecology, 24, 329–336.
Perez-Harguindeguy, N., Dıaz, S., Cornelissen, J., Vendramini, F., Cabido, M.
& Castellanos, A. (2000) Chemistry and toughness predict leaf litter decom-
position rates over a wide spectrum of functional types and taxa in central
Argentina.Plant and Soil, 218, 21–30.
Reynolds, B.C. &Hunter, M.D. (2001) Responses of soil respiration, soil nutri-
ents, and litter decomposition to inputs form canopy herbivores.Soil Biology
& Biochemistry, 33, 1641–1652.
Ritchie, M.E., Tilman, D. & Knops, J.M.H. (1998) Herbivore effects on plant
and nitrogen dynamics in oak savanna.Ecology, 79, 165–177.
Rossiter,M., Schultz, J.C. & Baldwin, I.T. (1988) Relationships among defolia-
tion, red oak phenolics, and gypsy moth growth and reproduction. Ecology,
69, 267–277.
Schweitzer, J.A., Bailey, J.K., Hart, S.C. & Whitham, T.G. (2005a) Nonaddi-
tive effects of mixing cottonwood genotypes on litter decomposition and
nutrient dynamics.Ecology, 86, 2834–2840.
Schweitzer, J.A., Bailey, J.K., Hart, S.C., Wimp, G.M., Chapman, S.K. &
Whitham, T.G. (2005b) The interaction of plant genotype and herbivory
decelerate leaf litter decomposition and alter nutrient dynamics. Oikos, 110,
133–145.
Stireman, J.O. & Cipollini, D. (2008) Stealth tactics of galling parasites and
their potential indirect effects.New Phytologist, 178, 462–465.
Stone, G.N. & Schonrogge, K. (2003) The adaptive significance of insect gall
morphology.Trends in Ecology & Evolution, 18, 512–522.
Swift, M.J., Heal, O.W. & Anderson, J.M. (1979) Decomposition in Terrestrial
Ecosystems. University of California Press, Berkeley, CA.
Tooker, J.F. & De Moraes, C.M. (2006) Jasmonate in lepidopteran larvae.
Journal of Chemical Ecology, 32, 2321–2326.
Tooker, J.F. & De Moraes, C.M. (2007) Feeding by Hessian fly [Mayetiola
destructor (Say)] larvae does not induce plant indirect defences. Ecological
Entomology, 32, 153–161.
Tooker, J.F. & De Moraes, C.M. (2009) A gall-inducing caterpillar species
increases essential fatty acid content of its host plant without concomitant
increases in phytohormone levels.Molecular Plant-Microbe Interactions, 22,
551–559.
Tooker, J.F., Koenig, W.A. & Hanks, L.M. (2002) Altered host plant volatiles
are proxies for sex pheromones in the gall waspAntistrophus rufus.Proceed-
ings of the National Academy of Sciences of the United States of America, 99,
15486–15491.
Tooker, J.F., Rohr, J.R., Abrahamson, W.G. & De Moraes, C.M. (2008) Gall
insects can avoid and alter indirect plant defenses. New Phytologist, 178,
657–671.
Trofymow, J.A., Moore, T.R., Titus, B.D., Prescott, C., Morrison, I., Siltanen,
M., Smith, S.M., Fyles, J., Wein, R., Camir, C., Duschene, L., Kozak, L.M.,
Kranabetter, M. & Visser, S. (2002) Rates of litter decomposition over
6 years in Canadian forests: influence of litter quality and climate. Canadian
Journal of Forest Research, 32, 789–804.
Wall, D.H., Bradford,M.A., John,M.G.S., Trofymow, J.A., Behan-Pelletier, V.,
Bignell, D.D.E., Dangerfield, J.M., Parton,W.J., Rusek, J., Voigt, W., Wol-
ters, V., Gardel, H.Z., Ayuke, F.O., Bashford, R., Beljakova, O.I., Bohlen,
P.J., Brauman, A., Flemming, S., Henschel, J.R., Johnson, D.L., Jones,
T.H., Kovarova, M., Kranabetter, J.M., Kutny, L., Lin, K.C., Maryati, M.,
Masse, D., Pokarzhevskii, A., Rahman, H., Sabara, M.G., Salamon, J.A.,
Swift, M.J., Varela, A., Vasconcelos, H.L., White, D. & Zou, X.M. (2008)
Global decomposition experiment shows soil animal impacts on decomposi-
tion are climate-dependent.Global Change Biology, 14, 2661–2677.
Weis, A.E. & Abrahamson, W.G. (1986) Evolution of host-plant manipulation
by gall makers – ecological and genetic-factors in the Solidago-Eurosta sys-
tem.AmericanNaturalist, 127, 681–695.
Wilkinson, C. & Scoble, M.J. (1979) The Nepticulidae (Lepidoptera) of
Canada.Memoirs of the Entomological Society of Canada, 111, 1–129.
Received 17 July 2011; accepted 15 February 2012
Handling Editor: DanHare
Supporting Information
Additional supporting information may be found in the online ver-
sion of this article.
Figure S1. Photographs showing morphological and structural
changes to the leaf lamina by (A–B) Rhopalomyia solidaginis and (C)
Pemphigus betae.
Figure S2. Gall induced on the petiole by Ectoedemia populella Brusk
(A) in the field showing no apparent morphological changes to the
leaf lamina, which was fully expanded before galling. (B–C) Close
view of the gall exterior and interior. (D–E) E. populella larvae and
frass pack. Photo credits: Christopher Frost.
Figure S3. Leaf drop timing in Populus grandidentata during active
litter fall.
As a service to our authors and readers, this journal provides Sup-
porting Information supplied by the authors. Such materials may be
re-organized for online delivery, but are not copy-edited or typeset.
Technical support issues arising from Supporting Information (other
thanmissing files) should be addressed to the authors.
� 2012 The Authors. Functional Ecology � 2012 British Ecological Society, Functional Ecology
Galling effects on leaf-litter decomposition 9





![The Barley Uniculme4 Gene Encodes a BLADE-ON- PETIOLE-Like ... · The Barley Uniculme4 Gene Encodes a BLADE-ON-PETIOLE-Like Protein That Controls Tillering and Leaf Patterning1[OPEN]](https://img.pdfslide.us/doc/110x75/5ed141dccd86a73bbf4f388b/the-barley-uniculme4-gene-encodes-a-blade-on-petiole-like-the-barley-uniculme4.jpg)























