Embed Size (px)
Citation preview

Large Molecule Therapeutics
A Novel Therapeutic Strategy to Rescue theImmune Effector Function of ProteolyticallyInactivated Cancer Therapeutic AntibodiesXuejun Fan1, Randall J. Brezski2, Hui Deng1, Pooja M. Dhupkar1, Yun Shi1,Anneliese Gonzalez3, Songlin Zhang4, Michael Rycyzyn2,William R. Strohl2,Robert E. Jordan2, Ningyan Zhang1, and Zhiqiang An1
Abstract
Primary and acquired resistance to anticancer antibodyimmunotherapies presents significant clinical challenges. Here,we demonstrate that proteolytic inactivation of cancer-targetingantibodies is an unappreciated contributor to cancer immuneevasion, and the finding presents novel opportunities for ther-apeutic intervention. A single peptide bond cleavage in theIgG1 hinge impairs cancer cell killing due to structural derange-ment of the Fc region. Hinge-cleaved trastuzumab graduallyaccumulated on the surfaces of HER2-expressing cancer celllines in vitro, and was greatly accelerated when the cells wereengineered to express the potent bacterial IgG-degrading pro-teinase (IdeS). Similar to cancer-related matrix metalloprotei-nases (MMP), IdeS exposes a hinge neoepitope that we havedeveloped an antibody, mAb2095-2, to specifically target theepitope. In in vitro studies, mAb2095-2 restored the lost anti-body-dependent cell-mediated cytotoxicity functionality of
cell-bound single-cleaved trastuzumab (scIgG-T). In vivo,mAb2095-2 rescued the impaired Fc-dependent tumor-sup-pressive activity of scIgG-T in a xenograft tumor model andrestored the recruitment of immune effector cells into thetumor microenvironment. More importantly, an Fc-engineeredproteinase-resistant version of mAb2095-2 rescued trastuzu-mab antitumor efficacy in a mouse tumor model with humancancer cells secreting IdeS, whereas trastuzumab alone showedsignificantly reduced antitumor activity in the same model.Consistently, an Fc-engineered proteinase-resistant version oftrastuzumab also greatly improved antitumor efficacy in thexenograft tumor model. Taken together, these findings point toa novel cancer therapeutic strategy to rescue proteolytic damageof antibody effector function by an Fc-engineered mAb againstthe hinge neoepitope and to overcome cancer evasion ofantibody immunity. Mol Cancer Ther; 14(3); 681–91. �2014 AACR.
IntroductionAntibodies are becoming a major drug modality for the treat-
ment of many human diseases, including cancer (1–3). However,development of resistance to antibody therapies is widespread,and an understanding of the resistance mechanisms is a topic ofgreat interest and clinical urgency (4–7). In the case of breastcancer, it is well established that the humanized IgG1 antibodytrastuzumab can inhibit tumor growth by binding via its variable
domain to highly expressed HER2 receptors on cancer cells toinhibit cell signaling (4, 8–10). As an IgG1 antibody, trastuzumabcan also mediate antibody-dependent cell-mediated cytotoxicity(ADCC) by recruiting immune effector cells to HER2-overexpres-sing tumor cells (11). Resistance to trastuzumab was associatedwith substantially increased expression of EGFR and HER3 (9).Upregulation of cMET expression (12), loss of PTEN, or othergenetic mutations in the network (8) were also proposed aspossible resistancemechanisms against trastuzumab. In addition,antibody resistance mechanisms suggestive of an evasion of Fc-mediatedADCChave also beenproposed (11, 13–15).Our grouphas recently reported that a single proteolytic cleavage in the lowerhinge region results in a profound loss of Fc immune effectorfunctions and reduced in vivo efficacy of trastuzumab. Moreover,the results indicated that the lower hinge of trastuzumab can becleaved by matrix metalloproteinases (MMP)—a class of protei-nases associated with the tumor environment (4).
In this study, we demonstrated that an anti-hinge antibodyagainst the site of proteinase cleavage can rescue the loss of Fceffect functions of the single-hinge–cleaved trastuzumab (scIgG-T) in vitro and in vivo. By Fcmolecular engineering, we showed thattrastuzumab itself and the anti-hinge cleavage site antibodymAb2095-2 used in combination with scIgG-T can be renderedresistant to IdeS hinge cleavage by incorporating select mutationsin the lower hinge and adjacent CH2 region. The Fc-engineeredanti-hinge cleavage site antibody restored immune effector func-tions and the antitumor efficacy of hinge cleaved trastuzumab invivo. Thus, the recognition of proteolytic inactivation of IgGs
1TexasTherapeutics Institute, BrownFoundation InstituteofMolecularMedicine, Houston, Texas. 2Biologics Research, Janssen R&D, LLC,Spring House, Pennsylvania. 3Division of Oncology, Department ofInternal Medicine, The University of Texas Health Science Center atHouston, Houston, Texas. 4Department of Pathology and LaboratoryMedicine, The University of Texas Health Science Center at Houston,Houston, Texas.
Note: Supplementary data for this article are available at Molecular CancerTherapeutics Online (http://mct.aacrjournals.org/).
Current address for R.J. Brezski: Antibody Engineering, Genentech, One DNAWay, South San Francisco, CA.
Corresponding Authors: Zhiqiang An, Texas Therapeutics Institute, BrownFoundation Institute of Molecular Medicine, University of Texas Health ScienceCenter at Houston, 1825 Pressler Street, Suite 532, Houston, TX 77030. Phone:713-500-3011; Fax: 713-500-2447; E-mail: [email protected]; andNingyan Zhang, [email protected]
doi: 10.1158/1535-7163.MCT-14-0715
�2014 American Association for Cancer Research.
MolecularCancerTherapeutics
www.aacrjournals.org 681
on June 8, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 31, 2014; DOI: 10.1158/1535-7163.MCT-14-0715

suggests an immune mechanism to restore their functions withinthe tumor microenvironment.
Materials and MethodsCell lines and reagents
MCF7, SKOV-3, SKBR-3, and BT474 cancer cell lines wereobtained from the ATCC in 2010. All cancer cells weremaintainedas previously described (4). Trastuzumab (IgG-T) was purchasedfrom a specialty pharmacy (IDIS; ref. 4). Single-hinge–cleavedtrastuzumab (scIgG-T) was prepared by enzymatic digestion withthe IgG-degrading enzyme S (IdeS) as previously described(4, 16). Anti-hinge cleavage site antibody (mAb2095-2), protein-ase-resistant antibodies, rabbit anti-human cleavage antibodies,and control antibodies were prepared as previously described(4, 17, 18). Recombinant streptococcal IgG-degrading enzyme(IdeS) was expressed in Escherichia coli and purchased fromGenovis AB. Antibodies for pHER2 (Y1248) and pAKT (S473)were purchased from Epitomics (Burlingame).
Detection of MMP expression in cancer cells and antibodyhinge cleavage by cancer cells
MMP expression in cancer cells was detected as describedpreviously (4). To determine the effect of trastuzumab concen-tration on single hinge cleavage, cancer cells were treated withvarying concentrations of trastuzumab (0.1, 1.0, and 10 mg/mL)for 24 hours. To determine the cleavage kinetics over time, cellswere treated with trastuzumab (10 mg/mL) over the period of 0, 4,and 24 hours. Trastuzumab concentration in the cell lysates wasquantified with ELISA using the HER2 ECD protein (Sinobiolo-gicals). Same amount of antibody was used for enrichment basedon the ELISA results from each cancer cell lysates. Enrichment ofsingle hinge–cleaved trastuzumab (scIgG-T) was performed usingProtein A dynabeads followed by SDS-PAGE electrophoresisunder denaturing and reducing conditions and detected by West-ern blotting as described previously (4). Detection of antibodyhinge cleavage by flow cytometry was carried out by anti–poly-cleavage antibodies cocktail followed by anti–rabbit-FITC anti-body (Jackson ImmunoResearch Laboratories). The mean flores-cence intensity (MFI) was measured using a Guava easyCyte HTinstrument (Millipore).
Construction of BT474-IdeS and SKOV-3-IdeS stable cell linesBT474-IdeS and SKOV-3-IdeS stable cell lines were constructed
on the basis of protocols described previously (19), using theRevTet-off system (Clontech Laboratories). The design strategy issummarized in Supplementary Fig. S1. For the detection of IdeSexpression, BT474-IdeS, SKOV-3-IdeS, and vehicle control cellswere cultured in RPMI-1640 media and cell culture supernatantswere collected, and concentrated by centrifugation at 2,000 rpmfor 30minutes at 4�C in AmiconUltra tubes (Millipore). Anti-flagM2 beads (Sigma) were used to pull down the IdeS protein andsubjected to SDS-PAGE separation under denature and nonre-ducing conditions, and the expression of IdeS was detected byWestern blotting.
Quantitation of trastuzumab level in mouse sera and tumortissues by ELISA
HER2 ECD was coated on 96-well plates at 2 mg/mL in PBSbuffer and incubated at 4�C overnight. The plates were thenwashed with 300 mL/well with PBS containing 0.05% Tween-
20 (PBS-T) and blocked with 5% non-fat milk (Bio-Rad Labora-tories) for 1 hour at room temperature (RT). The trastuzumabconcentration wasmeasured in themouse sera and tumor tissues.Goat-anti-human Fc horseradish peroxidase (HRP; Sigma) con-jugate (1:5K dilution) was added and incubated for another1 hour at RT. The plates were developed using the 3,30,5,50–tetramethylbenzidine substrate system (Sigma Life Science) andstopped by the addition of 2MH2SO4. The plates were read at 450nm using Spectra Max M4 (Molecular Devices).
Binding of the anti-hinge cleavage site antibody mAb2095-2 toscIgG-T on cancer cells and antibody-dependent cellularcytotoxicity assay
Specific binding of the anti-hinge cleavage site antibodymAb2095-2 to scIgG-T on SKOV-3 and BT474 cancer cells wasmeasured using a Guava easyCyte HT instrument based on themanufacturer's instructions and as described previously (4). R-PE(phycoerythrin)–conjugated streptavidin was used as a negativecontrol. ADCC activities were assayed as described previously(20), using the xCELLigence instrument (ACEA Biosciences).scIgG-T was added in 3-fold titration starting from 30 nmol/L asreported previously (4), in the presence or absence of mAb-2095-2 at 60 nmol/L concentration. For the detection of antibodyconcentration in mouse sera and tumor tissues, blood samplesand tumor tissues from athymic nude mice (n ¼ 4–5) werecollected 1 day after the last antibody administration. The antibodyconcentrations were determined by ELISA as described previously(4). ADCC activities using SKOV-3-IdeS target cells were examinedby a single (at 0 hour) or multiple addition (0, 24, 48, and 72hours) of trastuzumab, mAb2095-2, cleavage-resistant version oftrastuzumab (PR-trastuzumab) or PR-2095-2 as indicated.
Mouse xenograft tumor modelsImmunodeficient homogeneous athymic background nude
mice (Charles River Laboratories) were used for mouse xenografttumor studies. These studies were carried out in accordance withthe animal care and use guidelines, and the protocol wasapproved by the Animal Welfare Committee of the University ofTexas Medical School at Houston. BT474 and BT474-IdeS breastcancer cells were implanted s.c. in mice of 6- to 8-weeks-old aspreviously reported (20). When the tumor size reached at 50 to100mm3, mice were randomized into treatment groups with 5 to6 mice per group. Antibody was administered twice weekly i.p. at5 mg/kg for 5 weeks. Tumor size was measured and recorded twotimes every week using a Vernier scale caliper. Xenograft tumortissues were collected and stored at �80�C until further ex vivoanalysis. Mouse spleens were collected in 10 mL of RPMI mediauntil flow-cytometry analysis.
Detection of immune cell infiltration in xenograft tumor tissuesand antibody single hinge cleavage in breast tumor tissues frompatients with high HER2–expressing breast cancer treated withtrastuzumab by immunohistochemistry
The tumor tissues were embedded in optimal cutting temper-ature (Sakura Finetek). Serial 4-mm thick sections were made forIHC staining. The tumor tissue slides were blocked with 5%normal serum in PBS. A drop from the Biotin Blocking Kit (VectorLaboratories) and a drop from the Avidin Blocking Kit (VectorLaboratories) were added and incubated for 30 minutes at RT.aM/CD11b and F4/80 (R&D Systems) were applied as primary
Fan et al.
Mol Cancer Ther; 14(3) March 2015 Molecular Cancer Therapeutics682
on June 8, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 31, 2014; DOI: 10.1158/1535-7163.MCT-14-0715

antibodies and positive staining was visualized using a three-stepstaining procedurewith an Elite ABCkit (Vector Laboratories) andcounterstained with hematoxylin (Vector Laboratories). Threetumor tissue sections (n ¼ 3) were prepared from each treatmentgroup and the entire tumor sections were scanned under �200(magnification ¼ 20 � 10) to identify all positive stained cellsusing a Carl ZEISS microscopy equipped with the CamerasAxioCamMRc5 software (Monument). Positive stained immunecells were counted from eight views of each tumor tissue slide.Average number of immune cell infiltration per tumor tissue slideunder�200magnifications was plotted for each treatment groupand SE was calculated from the three tumor tissue slides in 24images.
Detection of scIgG-T in breast tumor tissues were carried out inaccordance with an Institutional Review Board approved clinicalprotocol (UTHSC-MS-11-0559—protocol no. T-11-102). Thepatients' tumor tissues were excised freshly, fixed in 4% parafor-maldehyde, and embedded in paraffin. Serial 4-mm thick sectionswere prepared for IHC staining. Rabbit anti-human cleavageantibody cocktail (anti-APE, anti-ELLG, and anti-PAP) wasapplied as primary antibody, and positive staining was visualizedusing a three-step staining procedure with the Elite ABC Kit(Vector Laboratories) as described above.
Cell lysate preparation from tumor tissuesMouse xenograft tumor tissues were homogenized using gentle
MACS (MACS Miltenyi Biotec) according to the manufacturer'sprocedure in a cell lysis buffer in the presence of proteinaseinhibitor cocktail (Calbiochem). Protein concentration was mea-sured by Bio-Rad DC Protein Assay (BIO-RAD), and protein wasdetected by Western blotting on SDSPAGE.
Detection of immune cells in dissociated splenocytes by flowcytometry
The mice splenocytes (n ¼ 3) were minced to single cells with apair of tweezers and filtered through cell strainer (70 mm; BDFalcon). Harvested cells were centrifuged at 2,000 rpm for 8minutes at 4�C. The supernatant was discarded and 1 mL ACKlysis buffer (Gibco Life Technologies)was added to each sample for5 minutes at RT. The splenocytes were washed with PBS (pH 7.4)and cells (1� 106/tube) were stainedwith four labeled antibodies:FITC-conjugated anti-Mouse CD11b (BD Pharmingen), PE-conju-gated anti-Mouse CD49b (BD Pharmingen), Peridinin–chloro-phyll proteins (PerCP) conjugated anti-Mouse F4/80 (BioLegend),and allophycocyanin-conjugated anti-Mouse Ly-6G (BD Pharmin-gen) for 30 minutes. The stained cells were washed with 1% BSA–PBS.Cellswere then suspended in 2%BSA–PBS buffer and positivestained cells were analyzed using a Guava easyCyte HT instrumentaccording to the manufacturer's manual.
Statistical analysisStatistical analysis was performed using paired a Student t test.
AP value of less than 0.05between treatment groups is consideredsignificantly different.
ResultsTrastuzumab hinge cleavage occurs upon antibody binding onhigh HER2-expressing cancer cells
Cancer cells produce many types of MMPs and their expressioncan be detected in both culture media and cell lysates (Supple-
mentary Fig. S2). To test whether MMPs expressed by cancer cellscan mediate antibody single hinge cleavage, we examined thecleavage of the trastuzumabhinge using apanel of cancer cell lineswith low or high HER2 expression. Cancer cells were cultured inthe presence of trastuzumab for 24 hours; trastuzumab wasenriched by Protein A bead adsorption from the culture mediaand cell lysate, respectively; and single hinge cleavage of trastu-zumab was measured by Western blotting. Molecular weights ofthe different antibody types and fragments are shown in Fig. 1A.Measurable quantities of single hinge cleavage were detectedwhen trastuzumab was enriched from cell lysate of the three highHER2-expressing cancer cell lines (SKOV-3, SKBR-3, and BT474),but no singlehinge cleavagewasdetected in trastuzumab enrichedfrom cell lysates of the low HER2–expressing MCF7 cancer cellline (Fig. 1B), suggesting that single hinge cleavage takes placewhen trastuzumab is bound to the cell surface. In support of thisconjecture, no single hinge cleavage was detected in the culturemedia collected from high HER2 expression cancer cells (Sup-plementary Fig. S3). For the high HER2–expressing cell lines,trastuzumab cleavage was barely detectable in the cell lysate after4 hours of incubation, but cleavage was readily detectable after 24hours of incubation (Fig. 1C–E). Collectively, these results suggestthat proteolytic hinge cleavage requires the antibody binding tothe cell surface and the cleavage process is accumulative and timedependent.
To investigate whether single hinge cleavage of trastuzumaboccurs in human breast tumors, we stained the surgical breasttumor tissues from 2 patients with HER2-overexpressing breastcancer. The 2 patient surgical breast tumor tissues were sampledafter 6 months of adjuvant treatment with trastuzumab andchemotherapy. Compared with the negative IHC staining con-trol (Fig. 1F), strong singe hinge cleaved antibodies weredetected to be associated with tumor tissues from the 2 patients(Fig. 1G and H). These results demonstrate that the cleavage oftrastuzumab and generation of scIgG-T occur in the tumormicroenvironment.
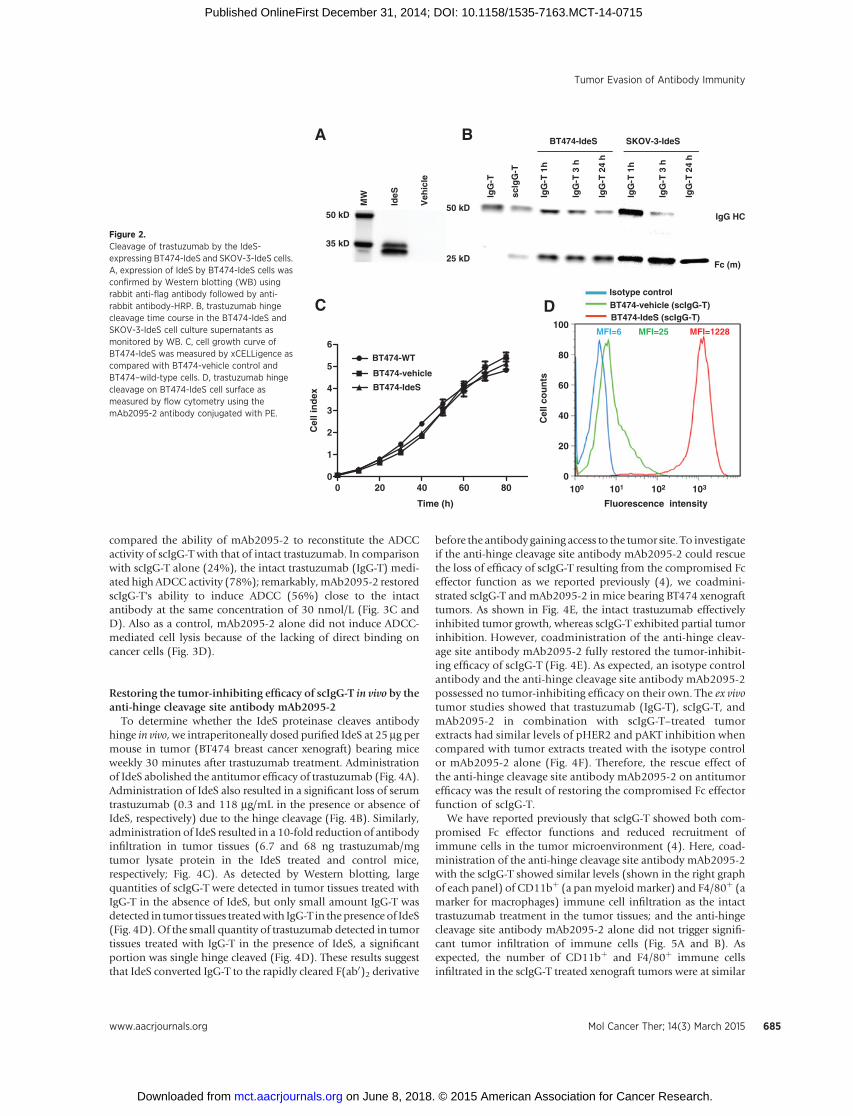
Overexpression of IdeS in cancer cells mediates strong singlehinge cleavage of trastuzumab in vitro and in vivo
As high HER2–expressing cancer cells such as BT474 showedtrastuzumab single hinge cleavage upon antibody binding on thecell surface (Fig. 1B and E), the antibody cleavage sites and theproteinases that mediated the cleavage have yet to be established.Moreover, the enzymatic cleavage kinetics mediated by the nat-ural cancer cells is relatively slow (Fig. 1C–E). To generate a robustmodel of the proteinase-mediated antibody hinge cleavage in vitroand in vivo, we constructedBT474andSKOV-3 cell lines that stablyexpressed the IdeS proteinase (Supplementary Fig. S1). IdeScleaves the hinge of IgG antibody with high specificity at the—LLG236!G237PS—site and has shown strong single cleavage ofthe antibodies in our previous studies (4, 16). The expressedprotein was readily detected in the IdeS-expressing cancer cellsusing an anti-flag antibody, but not in the vehicle transfectedcontrol cells (Fig. 2A). Overexpression of IdeS in both BT474 andSKOV-3 cancer cells showed rapid antibody single hinge cleavagewhen trastuzumab was added in the cultures (Fig. 2B). In addi-tion, overexpression of IdeS showed no impact on proliferation ofcancer cells (Fig. 2C). To further confirm that the single hingecleaved trastuzumab (scIgG-T) were bound to cancer cell surfaces;we detected cell surface bound scIgG-T using flow cytometry afterincubation of trastuzumab with BT474-IdeS cells for 24 hours. A
Tumor Evasion of Antibody Immunity
www.aacrjournals.org Mol Cancer Ther; 14(3) March 2015 683
on June 8, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 31, 2014; DOI: 10.1158/1535-7163.MCT-14-0715
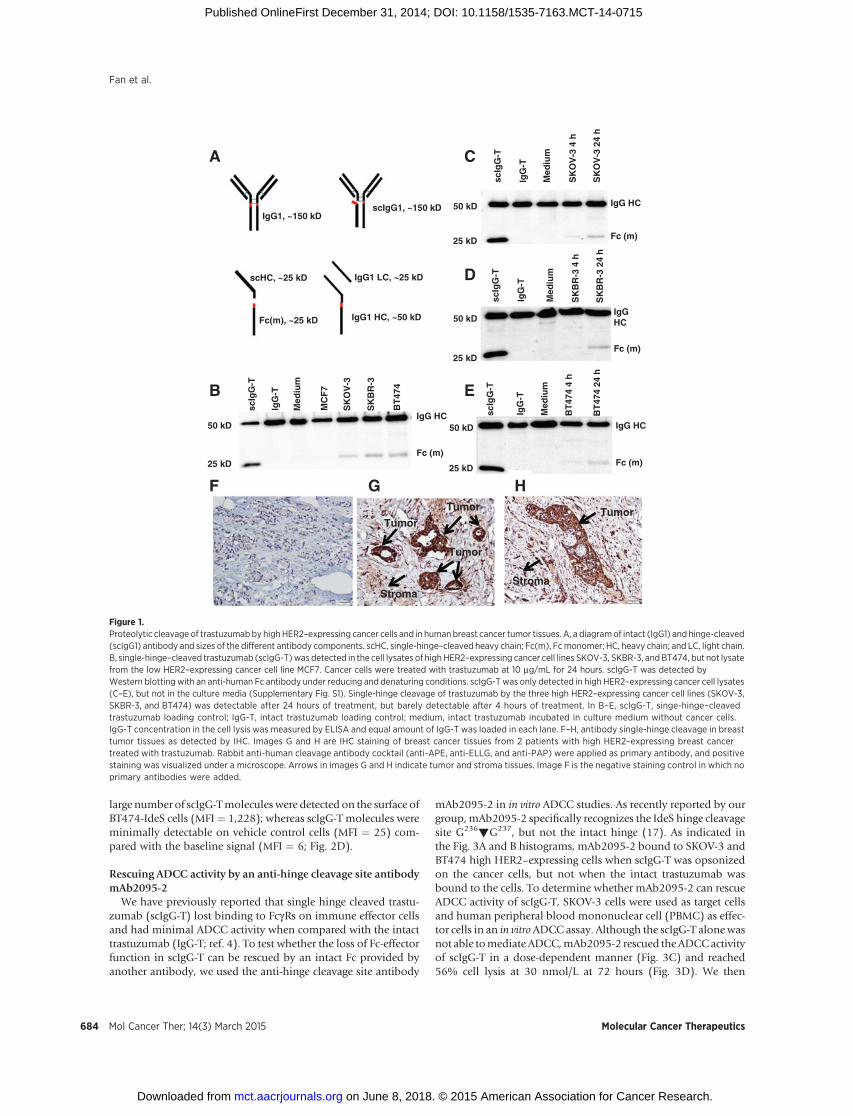
large number of scIgG-Tmolecules were detected on the surface ofBT474-IdeS cells (MFI¼ 1,228); whereas scIgG-T molecules wereminimally detectable on vehicle control cells (MFI ¼ 25) com-pared with the baseline signal (MFI ¼ 6; Fig. 2D).
Rescuing ADCC activity by an anti-hinge cleavage site antibodymAb2095-2
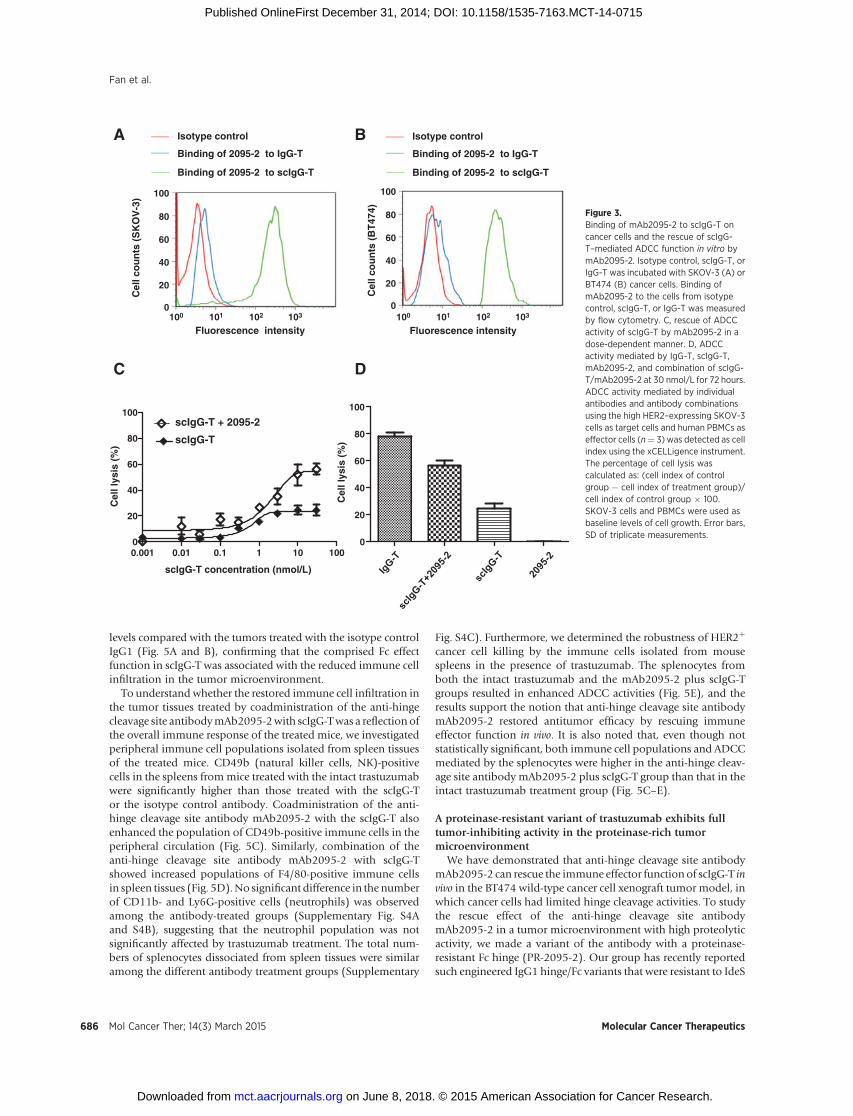
We have previously reported that single hinge cleaved trastu-zumab (scIgG-T) lost binding to FcgRs on immune effector cellsand had minimal ADCC activity when compared with the intacttrastuzumab (IgG-T; ref. 4). To test whether the loss of Fc-effectorfunction in scIgG-T can be rescued by an intact Fc provided byanother antibody, we used the anti-hinge cleavage site antibody
mAb2095-2 in in vitro ADCC studies. As recently reported by ourgroup,mAb2095-2 specifically recognizes the IdeS hinge cleavagesite G236!G237, but not the intact hinge (17). As indicated inthe Fig. 3A and B histograms, mAb2095-2 bound to SKOV-3 andBT474 high HER2–expressing cells when scIgG-T was opsonizedon the cancer cells, but not when the intact trastuzumab wasbound to the cells. To determine whether mAb2095-2 can rescueADCC activity of scIgG-T, SKOV-3 cells were used as target cellsand human peripheral blood mononuclear cell (PBMC) as effec-tor cells in an in vitroADCC assay. Although the scIgG-T alonewasnot able tomediate ADCC,mAb2095-2 rescued theADCCactivityof scIgG-T in a dose-dependent manner (Fig. 3C) and reached56% cell lysis at 30 nmol/L at 72 hours (Fig. 3D). We then
B
D
scIg
G-T
Med
ium
SK
BR
-3 4
h
SK
BR
-3 2
4 h
IgG
-T
IgG
-T
scIg
G-T
Med
ium
SK
OV
-3
BT
474
MC
F7
SK
BR
-3
E
scIg
G-T
Med
ium
SK
OV
-3 4
h
SK
OV
-3 2
4 h
C
IgG
-T
A
scIg
G-T
Med
ium
BT
474
4 h
BT
474
24 h
IgG
-T
25 kD
50 kD
25 kD
50 kD
25 kD
50 kD
IgG HC
Fc (m)
IgG HC
Fc (m)
IgGHC
Fc (m)
IgG HC
Fc (m)25 kD
50 kD
IgG1, ~150 kDscIgG1, ~150 kD
scHC, ~25 kD
Fc(m), ~25 kD
IgG1 LC, ~25 kD
F G H
TumorTumor
StromaStroma
Tumor
Tumor
IgG1 HC, ~50 kD
Figure 1.Proteolytic cleavage of trastuzumabby highHER2–expressing cancer cells and in humanbreast cancer tumor tissues. A, a diagramof intact (IgG1) and hinge-cleaved(scIgG1) antibody and sizes of the different antibody components. scHC, single-hinge–cleaved heavy chain; Fc(m), Fcmonomer; HC, heavy chain; and LC, light chain.B, single-hinge–cleaved trastuzumab (scIgG-T)was detected in the cell lysates of high HER2–expressing cancer cell lines SKOV-3, SKBR-3, and BT474, but not lysatefrom the low HER2–expressing cancer cell line MCF7. Cancer cells were treated with trastuzumab at 10 mg/mL for 24 hours. scIgG-T was detected byWestern blotting with an anti-human Fc antibody under reducing and denaturing conditions. scIgG-T was only detected in high HER2–expressing cancer cell lysates(C–E), but not in the culture media (Supplementary Fig. S1). Single-hinge cleavage of trastuzumab by the three high HER2–expressing cancer cell lines (SKOV-3,SKBR-3, and BT474) was detectable after 24 hours of treatment, but barely detectable after 4 hours of treatment. In B–E, scIgG-T, singe-hinge–cleavedtrastuzumab loading control; IgG-T, intact trastuzumab loading control; medium, intact trastuzumab incubated in culture medium without cancer cells.IgG-T concentration in the cell lysis was measured by ELISA and equal amount of IgG-T was loaded in each lane. F–H, antibody single-hinge cleavage in breasttumor tissues as detected by IHC. Images G and H are IHC staining of breast cancer tissues from 2 patients with high HER2–expressing breast cancertreated with trastuzumab. Rabbit anti-human cleavage antibody cocktail (anti-APE, anti-ELLG, and anti-PAP) were applied as primary antibody, and positivestaining was visualized under a microscope. Arrows in images G and H indicate tumor and stroma tissues. Image F is the negative staining control in which noprimary antibodies were added.
Fan et al.
Mol Cancer Ther; 14(3) March 2015 Molecular Cancer Therapeutics684
on June 8, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 31, 2014; DOI: 10.1158/1535-7163.MCT-14-0715
compared the ability of mAb2095-2 to reconstitute the ADCCactivity of scIgG-T with that of intact trastuzumab. In comparisonwith scIgG-T alone (24%), the intact trastuzumab (IgG-T) medi-ated high ADCC activity (78%); remarkably,mAb2095-2 restoredscIgG-T's ability to induce ADCC (56%) close to the intactantibody at the same concentration of 30 nmol/L (Fig. 3C andD). Also as a control, mAb2095-2 alone did not induce ADCC-mediated cell lysis because of the lacking of direct binding oncancer cells (Fig. 3D).
Restoring the tumor-inhibiting efficacy of scIgG-T in vivo by theanti-hinge cleavage site antibody mAb2095-2
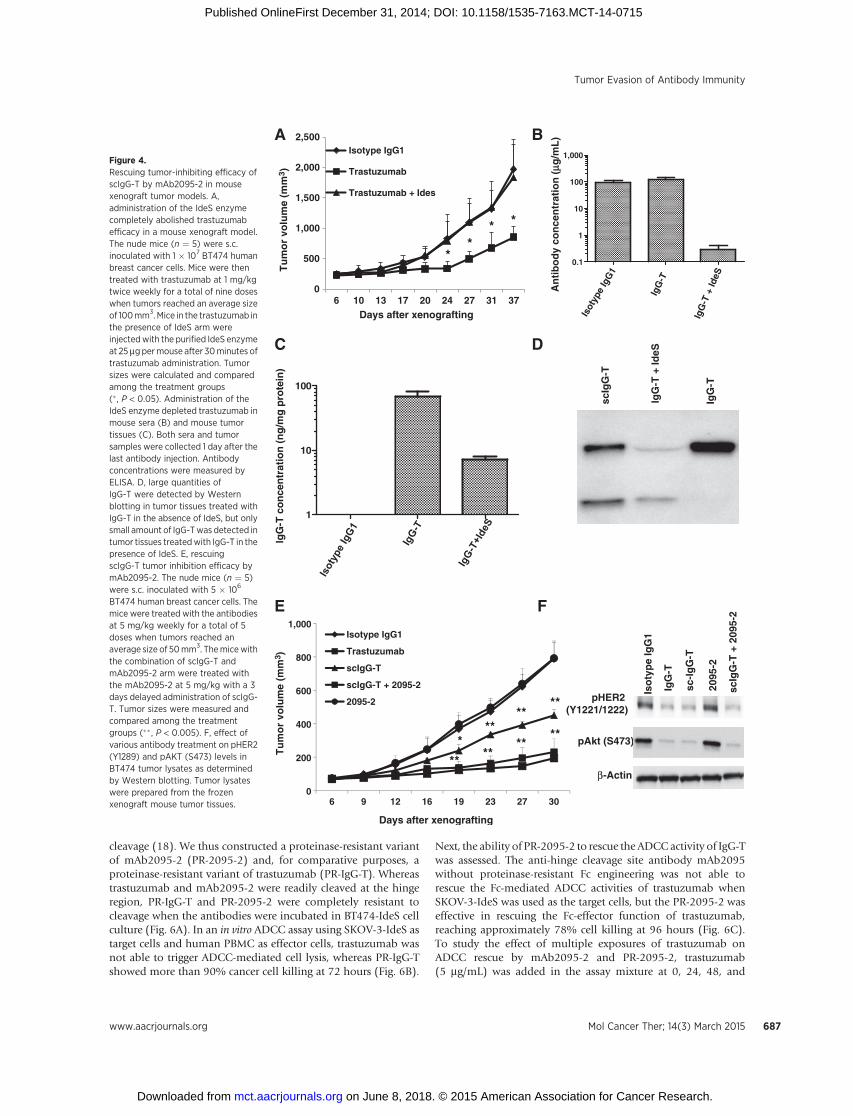
To determine whether the IdeS proteinase cleaves antibodyhinge in vivo, we intraperitoneally dosed purified IdeS at 25 mg permouse in tumor (BT474 breast cancer xenograft) bearing miceweekly 30 minutes after trastuzumab treatment. Administrationof IdeS abolished the antitumor efficacy of trastuzumab (Fig. 4A).Administration of IdeS also resulted in a significant loss of serumtrastuzumab (0.3 and 118 mg/mL in the presence or absence ofIdeS, respectively) due to the hinge cleavage (Fig. 4B). Similarly,administration of IdeS resulted in a 10-fold reduction of antibodyinfiltration in tumor tissues (6.7 and 68 ng trastuzumab/mgtumor lysate protein in the IdeS treated and control mice,respectively; Fig. 4C). As detected by Western blotting, largequantities of scIgG-T were detected in tumor tissues treated withIgG-T in the absence of IdeS, but only small amount IgG-T wasdetected in tumor tissues treatedwith IgG-T in the presence of IdeS(Fig. 4D). Of the small quantity of trastuzumab detected in tumortissues treated with IgG-T in the presence of IdeS, a significantportion was single hinge cleaved (Fig. 4D). These results suggestthat IdeS converted IgG-T to the rapidly cleared F(ab0)2 derivative
before the antibody gaining access to the tumor site. To investigateif the anti-hinge cleavage site antibody mAb2095-2 could rescuethe loss of efficacy of scIgG-T resulting from the compromised Fceffector function as we reported previously (4), we coadmini-strated scIgG-T and mAb2095-2 in mice bearing BT474 xenografttumors. As shown in Fig. 4E, the intact trastuzumab effectivelyinhibited tumor growth, whereas scIgG-T exhibited partial tumorinhibition. However, coadministration of the anti-hinge cleav-age site antibody mAb2095-2 fully restored the tumor-inhibit-ing efficacy of scIgG-T (Fig. 4E). As expected, an isotype controlantibody and the anti-hinge cleavage site antibody mAb2095-2possessed no tumor-inhibiting efficacy on their own. The ex vivotumor studies showed that trastuzumab (IgG-T), scIgG-T, andmAb2095-2 in combination with scIgG-T–treated tumorextracts had similar levels of pHER2 and pAKT inhibition whencompared with tumor extracts treated with the isotype controlor mAb2095-2 alone (Fig. 4F). Therefore, the rescue effect ofthe anti-hinge cleavage site antibody mAb2095-2 on antitumorefficacy was the result of restoring the compromised Fc effectorfunction of scIgG-T.
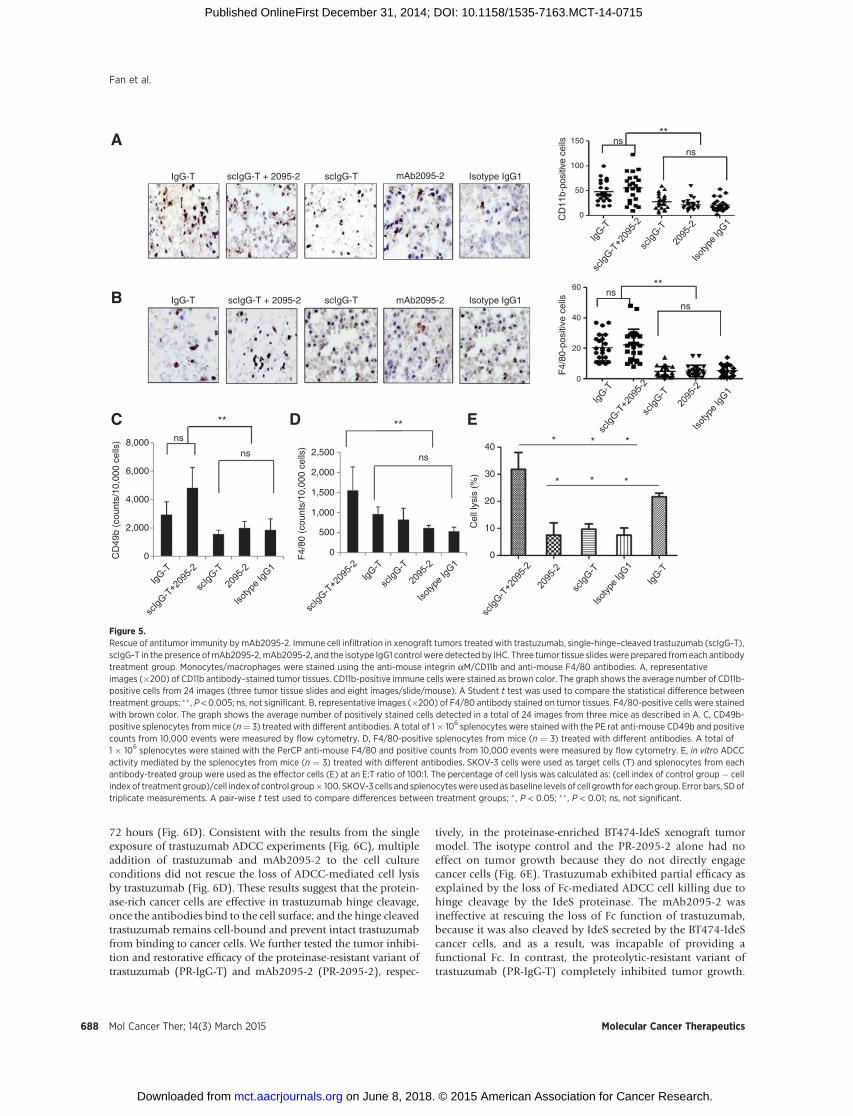
We have reported previously that scIgG-T showed both com-promised Fc effector functions and reduced recruitment ofimmune cells in the tumor microenvironment (4). Here, coad-ministration of the anti-hinge cleavage site antibody mAb2095-2with the scIgG-T showed similar levels (shown in the right graphof each panel) of CD11bþ (a panmyeloid marker) and F4/80þ (amarker for macrophages) immune cell infiltration as the intacttrastuzumab treatment in the tumor tissues; and the anti-hingecleavage site antibody mAb2095-2 alone did not trigger signifi-cant tumor infiltration of immune cells (Fig. 5A and B). Asexpected, the number of CD11bþ and F4/80þ immune cellsinfiltrated in the scIgG-T treated xenograft tumors were at similar
A
C
MW
Veh
icle
IdeS
B
D
Fluorescence intensity
0 20 40 60 800
1
2
3
4
5
6
Time (h)
BT474-WT
BT474-vehicle
BT474-IdeS
Cel
l in
dex
Cel
l co
un
ts
35 kD
50 kD
IgG
-T
scIg
G-T
IgG
-T 1
h
IgG
-T 2
4 h
IgG
-T 3
h
Isotype controlBT474-vehicle (scIgG-T)BT474-IdeS (scIgG-T)
MFI=25 MFI=1228MFI=6
50 kD
25 kD
IgG
-T 1
h
IgG
-T 2
4 h
IgG
-T 3
h
IgG HC
Fc (m)
BT474-IdeS SKOV-3-IdeS
100
80
60
40
20
0100 101 102 103
Figure 2.Cleavage of trastuzumab by the IdeS-expressing BT474-IdeS and SKOV-3-IdeS cells.A, expression of IdeS by BT474-IdeS cells wasconfirmed by Western blotting (WB) usingrabbit anti-flag antibody followed by anti-rabbit antibody-HRP. B, trastuzumab hingecleavage time course in the BT474-IdeS andSKOV-3-IdeS cell culture supernatants asmonitored by WB. C, cell growth curve ofBT474-IdeS was measured by xCELLigence ascompared with BT474-vehicle control andBT474–wild-type cells. D, trastuzumab hingecleavage on BT474-IdeS cell surface asmeasured by flow cytometry using themAb2095-2 antibody conjugated with PE.
Tumor Evasion of Antibody Immunity
www.aacrjournals.org Mol Cancer Ther; 14(3) March 2015 685
on June 8, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 31, 2014; DOI: 10.1158/1535-7163.MCT-14-0715
levels compared with the tumors treated with the isotype controlIgG1 (Fig. 5A and B), confirming that the comprised Fc effectfunction in scIgG-T was associated with the reduced immune cellinfiltration in the tumor microenvironment.
To understand whether the restored immune cell infiltration inthe tumor tissues treated by coadministration of the anti-hingecleavage site antibodymAb2095-2with scIgG-Twas a reflection ofthe overall immune response of the treated mice, we investigatedperipheral immune cell populations isolated from spleen tissuesof the treated mice. CD49b (natural killer cells, NK)-positivecells in the spleens frommice treated with the intact trastuzumabwere significantly higher than those treated with the scIgG-Tor the isotype control antibody. Coadministration of the anti-hinge cleavage site antibody mAb2095-2 with the scIgG-T alsoenhanced the population of CD49b-positive immune cells in theperipheral circulation (Fig. 5C). Similarly, combination of theanti-hinge cleavage site antibody mAb2095-2 with scIgG-Tshowed increased populations of F4/80-positive immune cellsin spleen tissues (Fig. 5D).No significant difference in the numberof CD11b- and Ly6G-positive cells (neutrophils) was observedamong the antibody-treated groups (Supplementary Fig. S4Aand S4B), suggesting that the neutrophil population was notsignificantly affected by trastuzumab treatment. The total num-bers of splenocytes dissociated from spleen tissues were similaramong the different antibody treatment groups (Supplementary
Fig. S4C). Furthermore, we determined the robustness of HER2þ
cancer cell killing by the immune cells isolated from mousespleens in the presence of trastuzumab. The splenocytes fromboth the intact trastuzumab and the mAb2095-2 plus scIgG-Tgroups resulted in enhanced ADCC activities (Fig. 5E), and theresults support the notion that anti-hinge cleavage site antibodymAb2095-2 restored antitumor efficacy by rescuing immuneeffector function in vivo. It is also noted that, even though notstatistically significant, both immune cell populations and ADCCmediated by the splenocytes were higher in the anti-hinge cleav-age site antibody mAb2095-2 plus scIgG-T group than that in theintact trastuzumab treatment group (Fig. 5C–E).
A proteinase-resistant variant of trastuzumab exhibits fulltumor-inhibiting activity in the proteinase-rich tumormicroenvironment
We have demonstrated that anti-hinge cleavage site antibodymAb2095-2 can rescue the immune effector function of scIgG-T invivo in the BT474 wild-type cancer cell xenograft tumor model, inwhich cancer cells had limited hinge cleavage activities. To studythe rescue effect of the anti-hinge cleavage site antibodymAb2095-2 in a tumor microenvironment with high proteolyticactivity, we made a variant of the antibody with a proteinase-resistant Fc hinge (PR-2095-2). Our group has recently reportedsuch engineered IgG1 hinge/Fc variants that were resistant to IdeS
0.001 0.01 0.1 1 10 1000
20
40
60
80
100
BA
Fluorescence intensity
Isotype control
Binding of 2095-2 to scIgG-T___
Binding of 2095-2 to IgG-T___
___
Fluorescence intensity
Cel
l co
un
ts (
BT
474)
scIgG-T concentration (nmol/L)
DC
Isotype control
Binding of 2095-2 to scIgG-T___
Binding of 2095-2 to IgG-T___
___
0
20
40
60
80
100
Cel
l lys
is (
%)scIgG-T
scIgG-T + 2095-2
Cel
l co
un
ts (
SK
OV
-3)
Cel
l lys
is (
%)
0
20
40
60
80
100
0
20
40
60
80
100
100 101 102 103 100 101 102 103
Figure 3.Binding of mAb2095-2 to scIgG-T oncancer cells and the rescue of scIgG-T–mediated ADCC function in vitro bymAb2095-2. Isotype control, scIgG-T, orIgG-T was incubated with SKOV-3 (A) orBT474 (B) cancer cells. Binding ofmAb2095-2 to the cells from isotypecontrol, scIgG-T, or IgG-T was measuredby flow cytometry. C, rescue of ADCCactivity of scIgG-T by mAb2095-2 in adose-dependent manner. D, ADCCactivity mediated by IgG-T, scIgG-T,mAb2095-2, and combination of scIgG-T/mAb2095-2 at 30 nmol/L for 72 hours.ADCC activity mediated by individualantibodies and antibody combinationsusing the high HER2–expressing SKOV-3cells as target cells and human PBMCs aseffector cells (n¼ 3) was detected as cellindex using the xCELLigence instrument.The percentage of cell lysis wascalculated as: (cell index of controlgroup � cell index of treatment group)/cell index of control group � 100.SKOV-3 cells and PBMCs were used asbaseline levels of cell growth. Error bars,SD of triplicate measurements.
Fan et al.
Mol Cancer Ther; 14(3) March 2015 Molecular Cancer Therapeutics686
on June 8, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 31, 2014; DOI: 10.1158/1535-7163.MCT-14-0715
cleavage (18). We thus constructed a proteinase-resistant variantof mAb2095-2 (PR-2095-2) and, for comparative purposes, aproteinase-resistant variant of trastuzumab (PR-IgG-T). Whereastrastuzumab and mAb2095-2 were readily cleaved at the hingeregion, PR-IgG-T and PR-2095-2 were completely resistant tocleavage when the antibodies were incubated in BT474-IdeS cellculture (Fig. 6A). In an in vitro ADCC assay using SKOV-3-IdeS astarget cells and human PBMC as effector cells, trastuzumab wasnot able to trigger ADCC-mediated cell lysis, whereas PR-IgG-Tshowed more than 90% cancer cell killing at 72 hours (Fig. 6B).
Next, the ability of PR-2095-2 to rescue the ADCCactivity of IgG-Twas assessed. The anti-hinge cleavage site antibody mAb2095without proteinase-resistant Fc engineering was not able torescue the Fc-mediated ADCC activities of trastuzumab whenSKOV-3-IdeS was used as the target cells, but the PR-2095-2 waseffective in rescuing the Fc-effector function of trastuzumab,reaching approximately 78% cell killing at 96 hours (Fig. 6C).To study the effect of multiple exposures of trastuzumab onADCC rescue by mAb2095-2 and PR-2095-2, trastuzumab(5 mg/mL) was added in the assay mixture at 0, 24, 48, and
0
200
400
600
800
1,000
6 9 12 16 19 23 27 30
Isotype IgG1
Trastuzumab
scIgG-T
scIgG-T + 2095-2
2095-2
DC
Tu
mo
r vo
lum
e (m
m3 )
E
A B
An
tib
od
y co
nce
ntr
atio
n (
μμg/m
L)
0.1
1
10
100
1,000
1
10
100
scIg
G-T
IgG
-T +
IdeS
IgG
-T
IgG
-T c
on
cen
trat
ion
(n
g/m
g p
rote
in)
Iso
typ
e Ig
G1
IgG
-T
sc-I
gG
-T
2095
-2
scIg
G-T
+ 2
095-
2
pHER2(Y1221/1222)
β-Actin
pAkt (S473)****
****
**
****
*
Days after xenografting
F
IgG
-T +
IdeS
IgG
-T
Isot
ype
IgG
1
6 10 13 17 20 24 27 31 37
Isotype IgG1
Trastuzumab
Trastuzumab + Ides
Tu
mo
r vo
lum
e (m
m3 )
Days after xenografting
***
*
0
500
1,000
1,500
2,000
2,500
Figure 4.Rescuing tumor-inhibiting efficacy ofscIgG-T by mAb2095-2 in mousexenograft tumor models. A,administration of the IdeS enzymecompletely abolished trastuzumabefficacy in a mouse xenograft model.The nude mice (n ¼ 5) were s.c.inoculated with 1 � 107 BT474 humanbreast cancer cells. Mice were thentreated with trastuzumab at 1 mg/kgtwice weekly for a total of nine doseswhen tumors reached an average sizeof 100mm3.Mice in the trastuzumab inthe presence of IdeS arm wereinjectedwith the purified IdeS enzymeat 25mgpermouse after 30minutes oftrastuzumab administration. Tumorsizes were calculated and comparedamong the treatment groups(� , P < 0.05). Administration of theIdeS enzyme depleted trastuzumab inmouse sera (B) and mouse tumortissues (C). Both sera and tumorsamples were collected 1 day after thelast antibody injection. Antibodyconcentrations were measured byELISA. D, large quantities ofIgG-T were detected by Westernblotting in tumor tissues treated withIgG-T in the absence of IdeS, but onlysmall amount of IgG-Twas detected intumor tissues treatedwith IgG-T in thepresence of IdeS. E, rescuingscIgG-T tumor inhibition efficacy bymAb2095-2. The nude mice (n ¼ 5)were s.c. inoculated with 5 � 106
BT474 human breast cancer cells. Themice were treated with the antibodiesat 5 mg/kg weekly for a total of 5doses when tumors reached anaverage size of 50mm3. Themicewiththe combination of scIgG-T andmAb2095-2 arm were treated withthe mAb2095-2 at 5 mg/kg with a 3days delayed administration of scIgG-T. Tumor sizes were measured andcompared among the treatmentgroups (�� , P < 0.005). F, effect ofvarious antibody treatment on pHER2(Y1289) and pAKT (S473) levels inBT474 tumor lysates as determinedby Western blotting. Tumor lysateswere prepared from the frozenxenograft mouse tumor tissues.
Tumor Evasion of Antibody Immunity
www.aacrjournals.org Mol Cancer Ther; 14(3) March 2015 687
on June 8, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 31, 2014; DOI: 10.1158/1535-7163.MCT-14-0715
72 hours (Fig. 6D). Consistent with the results from the singleexposure of trastuzumab ADCC experiments (Fig. 6C), multipleaddition of trastuzumab and mAb2095-2 to the cell cultureconditions did not rescue the loss of ADCC-mediated cell lysisby trastuzumab (Fig. 6D). These results suggest that the protein-ase-rich cancer cells are effective in trastuzumab hinge cleavage,once the antibodies bind to the cell surface; and the hinge cleavedtrastuzumab remains cell-bound and prevent intact trastuzumabfrom binding to cancer cells. We further tested the tumor inhibi-tion and restorative efficacy of the proteinase-resistant variant oftrastuzumab (PR-IgG-T) and mAb2095-2 (PR-2095-2), respec-
tively, in the proteinase-enriched BT474-IdeS xenograft tumormodel. The isotype control and the PR-2095-2 alone had noeffect on tumor growth because they do not directly engagecancer cells (Fig. 6E). Trastuzumab exhibited partial efficacy asexplained by the loss of Fc-mediated ADCC cell killing due tohinge cleavage by the IdeS proteinase. The mAb2095-2 wasineffective at rescuing the loss of Fc function of trastuzumab,because it was also cleaved by IdeS secreted by the BT474-IdeScancer cells, and as a result, was incapable of providing afunctional Fc. In contrast, the proteolytic-resistant variant oftrastuzumab (PR-IgG-T) completely inhibited tumor growth.
0
500
1,000
1,500
2,000
2,500
A
CD
11b-
posi
tive
cells
**ns
ns
B
IgG-T scIgG-T mAb2095-2 Isotype IgG1scIgG-T + 2095-2
EDC
**ns
ns
F4/
80-p
ositi
ve c
ells
**
ns
0
2,000
4,000
6,000
8,000
**
ns
ns
CD
49b
(cou
nts/
10,0
00 c
ells
)
F4/
80 (
coun
ts/1
0,00
0 ce
lls)
IgG-T scIgG-T mAb2095-2scIgG-T + 2095-2 Isotype IgG1
0
10
20
30
40 ** *
** *C
ell l
ysis
(%
)
150
100
50
0
60
40
20
0
Figure 5.Rescue of antitumor immunity by mAb2095-2. Immune cell infiltration in xenograft tumors treated with trastuzumab, single-hinge–cleaved trastuzumab (scIgG-T),scIgG-T in the presence ofmAb2095-2, mAb2095-2, and the isotype IgG1 control were detected by IHC. Three tumor tissue slideswere prepared from each antibodytreatment group. Monocytes/macrophages were stained using the anti-mouse integrin aM/CD11b and anti-mouse F4/80 antibodies. A, representativeimages (�200) of CD11b antibody–stained tumor tissues. CD11b-positive immune cells were stained as brown color. The graph shows the average number of CD11b-positive cells from 24 images (three tumor tissue slides and eight images/slide/mouse). A Student t test was used to compare the statistical difference betweentreatment groups; �� , P < 0.005; ns, not significant. B, representative images (�200) of F4/80 antibody stained on tumor tissues. F4/80-positive cells were stainedwith brown color. The graph shows the average number of positively stained cells detected in a total of 24 images from three mice as described in A. C, CD49b-positive splenocytes frommice (n¼ 3) treated with different antibodies. A total of 1� 106 splenocytes were stained with the PE rat anti-mouse CD49b and positivecounts from 10,000 events were measured by flow cytometry. D, F4/80-positive splenocytes from mice (n ¼ 3) treated with different antibodies. A total of1 � 106 splenocytes were stained with the PerCP anti-mouse F4/80 and positive counts from 10,000 events were measured by flow cytometry. E, in vitro ADCCactivity mediated by the splenocytes from mice (n ¼ 3) treated with different antibodies. SKOV-3 cells were used as target cells (T) and splenocytes from eachantibody-treated group were used as the effector cells (E) at an E:T ratio of 100:1. The percentage of cell lysis was calculated as: (cell index of control group � cellindex of treatment group)/cell index of control group� 100. SKOV-3 cells and splenocyteswere used as baseline levels of cell growth for eachgroup. Error bars, SDoftriplicate measurements. A pair-wise t test used to compare differences between treatment groups; � , P < 0.05; �� , P < 0.01; ns, not significant.
Fan et al.
Mol Cancer Ther; 14(3) March 2015 Molecular Cancer Therapeutics688
on June 8, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 31, 2014; DOI: 10.1158/1535-7163.MCT-14-0715
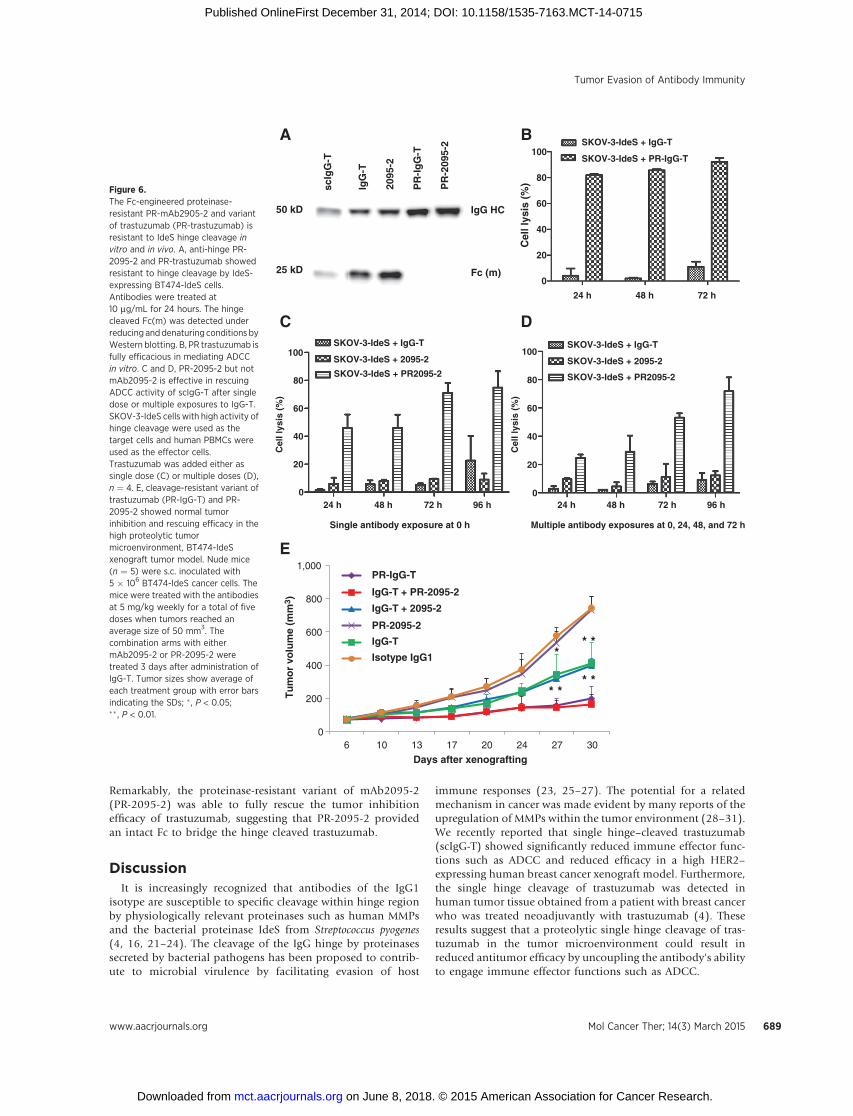
Remarkably, the proteinase-resistant variant of mAb2095-2(PR-2095-2) was able to fully rescue the tumor inhibitionefficacy of trastuzumab, suggesting that PR-2095-2 providedan intact Fc to bridge the hinge cleaved trastuzumab.
DiscussionIt is increasingly recognized that antibodies of the IgG1
isotype are susceptible to specific cleavage within hinge regionby physiologically relevant proteinases such as human MMPsand the bacterial proteinase IdeS from Streptococcus pyogenes(4, 16, 21–24). The cleavage of the IgG hinge by proteinasessecreted by bacterial pathogens has been proposed to contrib-ute to microbial virulence by facilitating evasion of host
immune responses (23, 25–27). The potential for a relatedmechanism in cancer was made evident by many reports of theupregulation of MMPs within the tumor environment (28–31).We recently reported that single hinge–cleaved trastuzumab(scIgG-T) showed significantly reduced immune effector func-tions such as ADCC and reduced efficacy in a high HER2–expressing human breast cancer xenograft model. Furthermore,the single hinge cleavage of trastuzumab was detected inhuman tumor tissue obtained from a patient with breast cancerwho was treated neoadjuvantly with trastuzumab (4). Theseresults suggest that a proteolytic single hinge cleavage of tras-tuzumab in the tumor microenvironment could result inreduced antitumor efficacy by uncoupling the antibody's abilityto engage immune effector functions such as ADCC.
B
PR
-Ig
G-T
IgG
-T
2095
-2
25 kD
50 kD
scIg
G-T
PR
-209
5-2
IgG HC
Fc (m)
A
Cel
l lys
is (
%)
0
20
40
60
80
100SKOV-3-IdeS + PR-IgG-T
SKOV-3-IdeS + IgG-T
72 h48 h24 h
0
20
40
60
80
100
Cel
l lys
is (
%)
Cel
l lys
is (
%)
SKOV-3-IdeS + 2095-2
SKOV-3-IdeS + IgG-T
SKOV-3-IdeS + PR2095-2
96 h72 h48 h24 h
DC
0
20
40
60
80
100SKOV-3-IdeS + 2095-2
SKOV-3-IdeS + IgG-T
SKOV-3-IdeS + PR2095-2
Single antibody exposure at 0 h
96 h72 h48 h24 h
Multiple antibody exposures at 0, 24, 48, and 72 h
E
Tu
mo
r vo
lum
e (m
m3 )
0
200
400
600
800
1,000
6 10 13 17 20 24 27 30
Days after xenografting
PR-IgG-T
IgG-T + PR-2095-2
IgG-T + 2095-2
PR-2095-2
IgG-T
Isotype IgG1 ** *
* ** *
Figure 6.The Fc-engineered proteinase-resistant PR-mAb2905-2 and variantof trastuzumab (PR-trastuzumab) isresistant to IdeS hinge cleavage invitro and in vivo. A, anti-hinge PR-2095-2 and PR-trastuzumab showedresistant to hinge cleavage by IdeS-expressing BT474-IdeS cells.Antibodies were treated at10 mg/mL for 24 hours. The hingecleaved Fc(m) was detected underreducing anddenaturing conditions byWestern blotting. B, PR trastuzumab isfully efficacious in mediating ADCCin vitro. C and D, PR-2095-2 but notmAb2095-2 is effective in rescuingADCC activity of scIgG-T after singledose or multiple exposures to IgG-T.SKOV-3-IdeS cells with high activity ofhinge cleavage were used as thetarget cells and human PBMCs wereused as the effector cells.Trastuzumab was added either assingle dose (C) or multiple doses (D),n ¼ 4. E, cleavage-resistant variant oftrastuzumab (PR-IgG-T) and PR-2095-2 showed normal tumorinhibition and rescuing efficacy in thehigh proteolytic tumormicroenvironment, BT474-IdeSxenograft tumor model. Nude mice(n ¼ 5) were s.c. inoculated with5 � 106 BT474-IdeS cancer cells. Themice were treated with the antibodiesat 5 mg/kg weekly for a total of fivedoses when tumors reached anaverage size of 50 mm3. Thecombination arms with eithermAb2095-2 or PR-2095-2 weretreated 3 days after administration ofIgG-T. Tumor sizes show average ofeach treatment group with error barsindicating the SDs; � , P < 0.05;�� , P < 0.01.
Tumor Evasion of Antibody Immunity
www.aacrjournals.org Mol Cancer Ther; 14(3) March 2015 689
on June 8, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 31, 2014; DOI: 10.1158/1535-7163.MCT-14-0715

Members of the MMP family play important roles in malig-nant processes. Their functions have been associated primarilywith degradation of extracellular matrix components in inva-sion and metastasis (29, 32, 33). Until relatively recently, littleattention was given to endogenous human IgG or specificantitumor mAb therapeutics as substrates whose functioncould be impaired by proteinases in the tumor environment(34). Recent studies from our group have demonstrated thattrastuzumab can readily be cleaved by recombinant forms ofMMP3, 7, 12, and 13 in vitro (4). We showed in this study thatMMPs are expressed by the tested cancer cells and demonstrat-ed for the first time that cancer cells possess the ability to cleavetargeting antitumor antibodies and cause impairment of Fc-mediated immune function.
As we reported previously, single hinge–cleaved trastuzumab(scIgG-T) by IdeS abolishes the antibody's ability to induceADCC-mediated cell lysis in vitro and in vivo (4, 16). The loss ofADCC activity of scIgG-T was a result of the compromised Fc-effector function due to reduced interaction with FcgRs (4, 16).Therefore, IdeS-expressing cancer cells provided a unique tool forexamining the ability of the anti-hinge cleavage site antibody toovercome the effects of antibody hinge cleavage in vitro and in vivo.Pertinent to the concept of in vivo proteolysis of IgGs was thefinding that a coadministration of IdeS proteinase with trastuzu-mab totally abolished antitumor efficacy in the BT474 mousexenograft model. This finding verified the potent IgG cleavageactivity of IdeS and the in vivo consequences of IgG functionalimpairment by this proteinase and potentially others with rele-vance to cancer (35).
Proteolytic cleavage of the IgG hinge exposes cryptic epitopesfor antibody recognition. The predominant epitopes are thosewith a free carboxyl terminus; for example, –C–P–A–P–COO–
resulting from MMP3 cleavage (23, 36). To enable the use of theIdeS proteinase as amodel system, we first generated mAb2095-2against a peptide analogue of the neoepitope of the IdeS hingecleavage site, C–P–A–P–E–L–L–G–COO–. MAb2095-2 binds tothis epitope in cleaved IgG1 in a highly specific manner and doesnot interact with intact IgG1 antibody (17). Therefore, the anti-hinge cleavage site antibody mAb2095-2 was a key tool forinvestigating if an anti-hinge cleavage site antibody could restoretherapeutic efficacy to proteolytically damaged antibody such astrastuzumab.
Indeed, the combination of mAb2095-2 with the inactivescIgG-T fully restored Fc-mediated cell-killing functions inADCC assays in vitro. The rescue effect is best explained by thecomplex formed between mAb2095-2 and scIgG-T on thecancer cell surface providing a surrogate intact hinge andfunctional Fc for engagement of immune effector cells. BecausescIgG-T retains its full capacity of binding affinity for the HER2,the binding of mAb2095-2 on the hinge cleavage site of scIgG-Tprovides the link between cancer cells and effective engagementof immune cells. The IdeS-overexpressing BT474 cancer celltumor model brought about rapid proteolytic hinge cleavage oftrastuzumab that greatly impaired the antitumor efficacy ofthe mAb. These data support previous studies with in vitrogenerated scIgG-T that also showed reduced antitumor activityin vivo (4).
In the context of low proteolytic activities against antibodyhinge such as wild-type BT474 cancer cells, coadministration ofmAb2095-2 (IgG1) fully restored scIgG-T to the efficacy level ofintact trastuzumab. This indicates that an administered anti-hinge
cleavage site antibody, in this case mAb2095-2, can reconstituteimmune effector function by the contribution of an intact Fcdomain. Our findings that anti-hinge cleavage site antibodymAb2095-2 restored immune cell populations in tumor tissuesand showed modulation of peripheral immune system in mice,support the notion that anti-hinge antibodies could sufficientlyreconstitute host immunity against cancer cells by providingintact Fc engagement of effector cells.
We further propose that these results, obtained in a modelsystem of trastuzumab hinge cleavage by IdeS, could guide thedesign of new Fc platforms for antibody therapeutics targetingvarious cancer cells in which endogenous proteinases are preva-lent. Because anti-hinge antibodies could recognize hinge-cleavedantibodieswhenever andwherever they are associatedwith cancercells, the anti-hinge cleavage site antibody would restore immuneeffector functions without the need of knowing the targets of theprimary (damaged) antibodies. The present results point to analternative and previously untested strategy to supplement hostimmunity in cancer.
It is of obvious concern that a therapeutic anti-hinge antibodywith the IgG1 hinge would be prone to an analogous hingecleavage in proteinase-rich physiologic conditions. As reportedrecently by our group, an Fc-engineered construct of an anti-CD20antibody showed full immune effector function and proteinaseresistance (18). In this study, a proteinase-resistant variant oftrastuzumab showed its ability to resist IdeS cleavage in vivo andpossessed full antitumor efficacy, whereas unmodified trastuzu-mab that was cleaved by IdeS resulted in the loss of antitumorefficacy in the BT474-IdeS mouse tumor model. More interest-ingly, we used an Fc-engineered anti-hinge cleavage site antibodyresistant to IdeS hinge cleavage that restored the antitumorefficacy of trastuzumab when it was impaired by IdeS in theBT474-IdeS tumor model. This successful rescue of trastuzumabefficacy by the Fc-engineered anti-hinge cleavage site antibodysignals the potential use of such a therapeutic approach toovercome the cleavage of antibodies in tumor tissue and to restorethe impaired Fc immune effector function.
Disclosure of Potential Conflicts of InterestW.R. Strohl received a commercial research grant from Janssen R&D. No
potential conflicts of interest were disclosed by the other authors.
Authors' ContributionsConception and design: R.J. Brezski, M. Rycyzyn, W.R. Strohl, N. Zhang, Z. AnDevelopment of methodology: X. Fan, R.J. Brezski, P.M. Dhupkar, R.E. Jordan,N. ZhangAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): P.M.Dhupkar, Y. Shi, H. Deng, A. Gonzalez, N. ZhangAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): X. Fan, R.J. Brezski, H. Deng, S. Zhang, M. Rycyzyn,N. Zhang, Z. AnWriting, review, and/or revision of the manuscript: X. Fan, R.J. Brezski,M. Rycyzyn, R.E. Jordan, N. Zhang, Z. AnAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): R.J. Brezski, N. ZhangStudy supervision: R.J. Brezski, W.R. Strohl, N. Zhang, Z. AnOther (conductedmouse tumor xenograft studies, and in vitro assays): X. FanOther (developed the method and conducted pull-down and Western blot-ting analysis to detect antibody hinge cleavage by cancer cells): P.M. Dhupkar
AcknowledgmentsThe authors thank Bin Yuan for constructing the IdeS expression vector and
Zhao Huang for assistance on Western blotting. The authors also thank Drs.
Fan et al.
Mol Cancer Ther; 14(3) March 2015 Molecular Cancer Therapeutics690
on June 8, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 31, 2014; DOI: 10.1158/1535-7163.MCT-14-0715

Michelle Kinder and Katharine Grugan for critical discussions throughout thestudies.
Grant SupportThis study was partially funded by grants from Janssen R&D, LLC (to
N. Zhang and Z. An), the Texas Emerging Technology Fund (to N. Zhang andZ. An), and the Welch Foundation grant no.AU0042 (to Z. An).
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received August 21, 2014; revised December 11, 2014; accepted December11, 2014; published OnlineFirst December 31, 2014.
References1. An Z. Monoclonal antibodies—a proven and rapidly expanding therapeu-
tic modality for human diseases. Protein Cell 2010;1:319–30.2. Reichert JM. Antibodies to watch in 2013: mid-year update. MAbs
2013;5:513–7.3. Strohl WR. Therapeutic monoclonal antibodies: past, present, and future.
In:An Z, editor. Therapeutic monoclonal antibodies: from bench to clinic.Hoboken, NJ: John Wiley & Sons, Inc.; 2009. p. 889.
4. Fan X, Brezski RJ, Fa M, Deng H, Oberholtzer A, Gonzalez A, et al. A singleproteolytic cleavage within the lower hinge of trastuzumab reducesimmune effector function and in vivo efficacy. Breast Cancer Res 2012;14:R116.
5. Chung A, Cui X, Audeh W, Giuliano A. Current status of anti-humanepidermal growth factor receptor 2 therapies: predicting and overcomingherceptin resistance. Clin Breast Cancer 2013;13:223–32.
6. Ratti M, Tomasello G. Emerging combination therapies to overcomeresistance in EGFR-driven tumors. Anti-cancer Drugs 2014;25:127–39.
7. Thery JC, Spano JP, Azria D, Raymond E, Penault Llorca F. Resistance tohuman epidermal growth factor receptor type 2-targeted therapies. Eur JCancer 2014;50:892–901.
8. Nagata Y, Lan KH, Zhou X, Tan M, Esteva FJ, Sahin AA, et al. PTENactivation contributes to tumor inhibition by trastuzumab, and loss ofPTEN predicts trastuzumab resistance in patients. Cancer Cell 2004;6:117–27.
9. Narayan M, Wilken JA, Harris LN, Baron AT, Kimbler KD, Maihle NJ.Trastuzumab-inducedHER reprogramming in "resistant" breast carcinomacells. Cancer Res 2009;69:2191–4.
10. She QB, Chandarlapaty S, Ye Q, Lobo J, Haskell KM, Leander KR, et al.Breast tumor cells with PI3K mutation or HER2 amplification are selec-tively addicted to Akt signaling. PLoS ONE 2008;3:e3065.
11. Clynes RA, Towers TL, Presta LG, Ravetch JV. Inhibitory Fc receptorsmodulate in vivo cytoxicity against tumor targets. Nat Med 2000;6:443–6.
12. Shattuck DL, Miller JK, Carraway KL III, Sweeney C. Met receptor con-tributes to trastuzumab resistance of Her2-overexpressing breast cancercells. Cancer Res 2008;68:1471–7.
13. Arnould L, Gelly M, Penault-Llorca F, Benoit L, Bonnetain F, Migeon C,et al. Trastuzumab-based treatment of HER2-positive breast cancer: anantibody-dependent cellular cytotoxicity mechanism? Br J Cancer2006;94:259–67.
14. Cooley S, Burns LJ, Repka T, Miller JS. Natural killer cell cytotoxicity ofbreast cancer targets is enhanced by two distinct mechanisms of antibody-dependent cellular cytotoxicity against LFA-3 andHER2/neu. ExpHematol1999;27:1533–41.
15. Mimura K, Kono K, Hanawa M, Kanzaki M, Nakao A, Ooi A, et al.Trastuzumab-mediated antibody-dependent cellular cytotoxicityagainst esophageal squamous cell carcinoma. Clin Cancer Res 2005;11:4898–904.
16. Brezski RJ, Vafa O, Petrone D, Tam SH, Powers G, Ryan MH, et al. Tumor-associated and microbial proteases compromise host IgG effector func-tions by a single cleavage proximal to the hinge. Proc Natl Acad Sci U S A2009;106:17864–9.
17. Malia TJ, TeplyakovA, Brezski RJ, Luo J, KinderM, Sweet RW, et al. Structureand specificity of an antibody targeting a proteolytically-cleaved IgG hinge.Proteins 2014;82:1656–67.
18. Kinder M, Greenplate AR, Grugan KD, Soring KL, Heeringa KA, McCarthySG, et al. Engineered protease-resistant antibodies with selectable cell-killing functions. J Biol Chem 2013;288:30843–54.
19. Huang Z, Choi BK, Mujoo K, Fan X, Fa M, Mukherjee S, et al. The E3ubiquitin ligase NEDD4 negatively regulates HER3/ErbB3 level and sig-naling. Oncogene 2014;56:1–11.
20. Zhang N, Liu L, Dan Dumitru C, Cummings NR, Cukan M, Jiang Y, et al.Glycoengineered Pichia produced anti-HER2 is comparable to trastuzu-mab in preclinical study. MAbs 2011;3:289–98.
21. Porter RR. Separation and isolation of fractions of rabbit gamma-globulincontaining the antibody and antigenic combining sites. Nature 1958;182:670–1.
22. Nisonoff A,Wissler FC, LipmanLN.Properties of themajor component of apeptic digest of rabbit antibody. Science 1960;132:1770–1.
23. Ryan MH, Petrone D, Nemeth JF, Barnathan E, Bjorck L, Jordan RE.Proteolysis of purified IgGs by human and bacterial enzymes in vitro andthe detection of specific proteolytic fragments of endogenous IgG inrheumatoid synovial fluid. Mol Immunol 2008;45:1837–46.
24. Gearing AJ, Thorpe SJ, Miller K, Mangan M, Varley PG, Dudgeon T, et al.Selective cleavage of human IgG by the matrix metalloproteinases, matri-lysin and stromelysin. Immunol Lett 2002;81:41–8.
25. von Pawel-Rammingen U, Johansson BP, Bjorck L. IdeS, a novel strepto-coccal cysteine proteinase with unique specificity for immunoglobulin G.EMBO J 2002;21:1607–15.
26. Mehta SK, Plaut AG, CalvanicoNJ, Tomasi TB Jr. Human immunoglobulinA: production of an Fc fragment by an enteric microbial proteolyticenzyme. J Immunol 1973;111:1274–6.
27. Senior BW, Woof JM. Effect of mutations in the human immunoglobulinA1 (IgA1) hinge on its susceptibility to cleavage by diverse bacterial IgA1proteases. Infect Immun 2005;73:1515–22.
28. Egeblad M, Werb Z. New functions for the matrix metalloproteinases incancer progression. Nat Rev Cancer 2002;2:161–74.
29. Kessenbrock K, Plaks V, Werb Z. Matrix metalloproteinases: regulators ofthe tumor microenvironment. Cell 2010;141:52–67.
30. Ruffell B, Au A, Rugo HS, Esserman LJ, Hwang ES, Coussens LM. BreastCancer Special Feature: leukocyte composition of human breast cancer.Proc Natl Acad Sci U S A 2012;109:2796–801.
31. Rhee JS, Coussens LM. RECKing MMP function: implications for cancerdevelopment. Trends Cell Biol 2002;12:209–11.
32. Radisky ES, Radisky DC. Matrix metalloproteinase-induced epithelial–mesenchymal transition inbreast cancer. JMammaryGlandBiolNeoplasia2010;15:201–12.
33. Hadler-Olsen E, Winberg JO, Uhlin-Hansen L. Matrix metalloproteinasesin cancer: their value as diagnostic and prognostic markers and therapeutictargets. Tumour Biol 2013;34:2041–51.
34. Brezski RJ, Jordan RE. Cleavage of IgGs by proteases associated withinvasive diseases: an evasion tactic against host immunity? MAbs2010;2:212–20.
35. von Pawel-Rammingen U. Streptococcal IdeS and its impact on immuneresponse and inflammation. J Innate Immun 2012;4:132–40.
36. SchmidtA,DeRiggiD, JordanRE,Whitaker B,HeavnerGA, KruszynskiM.Asynthetic peptide approach for elucidating the points of natural auto-antibody reactivity to proteolytic fragments of human IgG. Adv Exp MedBiol 2009;611:411–2.
www.aacrjournals.org Mol Cancer Ther; 14(3) March 2015 691
Tumor Evasion of Antibody Immunity
on June 8, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 31, 2014; DOI: 10.1158/1535-7163.MCT-14-0715

2015;14:681-691. Published OnlineFirst December 31, 2014.Mol Cancer Ther Xuejun Fan, Randall J. Brezski, Hui Deng, et al. AntibodiesFunction of Proteolytically Inactivated Cancer Therapeutic A Novel Therapeutic Strategy to Rescue the Immune Effector
Updated version
10.1158/1535-7163.MCT-14-0715doi:
Access the most recent version of this article at:
Material
Supplementary
http://mct.aacrjournals.org/content/suppl/2014/12/31/1535-7163.MCT-14-0715.DC1
Access the most recent supplemental material at:
Cited articles
http://mct.aacrjournals.org/content/14/3/681.full#ref-list-1
This article cites 35 articles, 10 of which you can access for free at:
Citing articles
http://mct.aacrjournals.org/content/14/3/681.full#related-urls
This article has been cited by 3 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mct.aacrjournals.org/content/14/3/681To request permission to re-use all or part of this article, use this link
on June 8, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 31, 2014; DOI: 10.1158/1535-7163.MCT-14-0715





























