Embed Size (px)
Citation preview

A Novel Mechanism of Action for Anti-Thymocyte Globulin:Induction of CD4�CD25�Foxp3� Regulatory T Cells
Marta Lopez, Michael R. Clarkson, Monica Albin, Mohamed H. Sayegh, andNader NajafianTransplantation Research Center, Brigham and Women’s Hospital, and Children’s Hospital Boston, Harvard MedicalSchool, Boston, Massachusetts
T cell–depleting agents are being tested as part of clinical tolerance strategies in humans with autoimmunity and transplan-tation. The immunosuppressive activity of anti-thymocyte globulin (ATG) has been thought to result primarily from depletionof peripheral lymphocytes. Herein is reported for the first time that ATG but not anti-CD52 mAb (alemtuzumab) or the IL-2Rantagonists causes rapid and sustained expansion of CD4�CD25� T cells when cultured with human peripheral bloodlymphocytes. These cells display enhanced expression of the regulatory markers glucocorticoid-induced TNF receptor,cytotoxic T lymphocyte–associated antigen-4 (CTLA-4), and forkhead box P3 and efficiently suppress a direct alloimmuneresponse of the original responder lymphocytes. It is interesting that the cells do not suppress memory responses to the recallantigen mumps. Ex vivo expansion of regulatory T cells is due mainly to conversion of CD4�CD25� into CD4�CD25� T cellsand to a lesser degree to proliferation of natural CD4�CD25� T cells. The induction of regulatory T cells depends onproduction of Th2 cytokines in the generating cultures. These novel data suggest that ATG not only may promote expansion/generation of regulatory T cells but also may be useful in future ex vivo expansion of these cells for cellular therapy inautoimmunity and clinical transplantation.
J Am Soc Nephrol 17: 2844–2853, 2006. doi: 10.1681/ASN.2006050422
P olyclonal anti-thymocyte globulin (ATG) is the purifiedIgG fraction of sera from rabbits, horses, or, morerarely, goats that are immunized with human thymo-
cytes or T cell lines. ATG is used for treatment of variousclinical conditions, including prevention or rescue treatment ofacute rejection in organ transplantation (1), conditioning forhematopoietic stem cell transplantation, treatment of severeaplastic anemia, various autoimmune diseases, and more re-cently graft-versus-host disease (2). The immunosuppressiveactivity of ATG has been thought to result primarily from thedepletion of peripheral lymphocytes from the circulating poolthrough complement-dependent lysis or activation-associatedapoptosis (1,3–5). Other potential mechanisms of action includemodulation of surface adhesion molecules or chemokine recep-tor expression (6).
Regulatory T cells (Treg) are specialized T cell subsets thatplay important roles in maintaining immune homeostasis (7,8).Treg are characterized by the expression of the IL-2 receptor�-chain (CD25) and the transcription factor forkhead box P3(Foxp3) (9). There exists emerging evidence in both rodents andhumans to support important immunoregulatory functions ofCD4�CD25� T Treg in maintaining both self-tolerance and
tolerance toward autoantigens (8) and alloantigens (10). Tregalso may play a role in preventing human renal autoimmunediseases such as Goodpasture’s disease (11). Our group previ-ously demonstrated that active regulation of the alloimmuneresponses by Treg may function to maintain hyporesponsive-ness to alloantigens in renal transplant patients (12,13). In pre-clinical animal models, ex vivo expanded Treg protect micefrom lethal graft-versus-host disease (14). In fact, a clinical trialwas proposed recently to use ex vivo expanded Treg at the timeof hematopoietic stem cell transplantation (15,16). Therefore,understanding the conditions that are required for the genera-tion and propagation of Treg would allow development ofnovel therapeutic strategies for inducing immunologic toler-ance in various immune-mediated diseases.
Here we report the novel finding that ATG-mediated immu-nosuppression is delivered in part via immunologically specificactions involving the generation of Treg, particularlyCD4�CD25highFoxp3� cells. This is due mainly to ATG’sunique ability to convert the CD4�CD25� T cells intoCD4�CD25� T cells. More important, in a mixed lymphocytereaction, the regulatory function is restricted to autologousresponder cells from which Treg originally were generated anddoes not affect the memory response to recall antigens. Ourfindings have relevant clinical implications for designing newimmunomodulatory protocols for immune-mediated diseases.
Materials and MethodsCells and Antibodies
This study was performed with the approval of the InstitutionalReview Board for human investigation at the Brigham and Women’s
Received May 1, 2006. Accepted June 19, 2006.
Published online ahead of print. Publication date available at www.jasn.org.
Address correspondence to: Dr. Nader Najafian, Brigham & Women’s Hospital,Transplantation Research Center, EBRC, 221 Longwood Avenue, 3rd Floor, Bos-ton, MA 02115. Phone: 617-732-5259; Fax: 617-732-5254; E-mail:[email protected]
Copyright © 2006 by the American Society of Nephrology ISSN: 1046-6673/1710-2844

Hospital. Blood from healthy volunteers were obtained in heparinizedtubes, and peripheral blood lymphocytes (PBL) were isolated by stan-dard Ficoll-density-gradient centrifugation. Two types of ATG wereused: A rabbit polyclonal serum raised against human thymocytes(Thymoglobulin; Genzyme, Cambridge MA) or a rabbit polyclonalserum raised against the lymphoblastic Jurkat T cell line (Fresenius,Bad Homburg, Germany). Purified rabbit polyclonal IgG was used ascontrol. To compare the effects of ATG with agents that block the IL-2receptor �-chain (IL-2R�) on the surface of activated T lymphocytes, weused either Basiliximab (Simulect, Novartis, NJ), a chimeric (murine/human) mAb (IgG1k), or Daclizumab (Zenapax, Roche, NJ), a human-ized IgG1 mAb. In addition, we used alemtuzumab (Campath1-H,Genzyme, Cambridge, MA), a recombinant DNA-derived humanizedmAb that is directed against the cell surface glycoprotein CD52, forcomparison studies with ATG.
Generation of TregPBL (1 � 106 per ml) from 10 healthy donors were incubated with
ATG (Treg) or rabbit IgG (TControl) in RPMI 1640 medium (CambrexBioscience, Walkersville, MD) supplemented with 10% heat-inactivatedhuman serum at 37°C with a Pco2 5% for various durations (6 to 96 h).All above agents were used in vitro at concentrations that ranged from1 to 100 �g/ml.
These cultures are called “generating cultures.” For assessment of therole of cytokines in the expansion of Treg, anti–IL-4, anti–IL-13, andanti–IL-10 mAb each at a concentration of 10 �g/ml were addedseparately to the generating cultures (17). All of these antibodies werepurchased from BD Bioscience (San Jose, CA).
In Vitro Suppression AssayTo test the suppression function of cells that were generated under
conditions explained (Treg), we set up a mixed lymphocyte reaction(MLR) assay: 1 � 105 cells that were obtained from the generatingcultures above were co-cultured (1:1 ratio) with fresh responder (au-tologous or third-party PBL) and irradiated stimulator cells in a 96-wellplate (96-well Cell Culture Cluster, round-bottom culture plate;COSTAR, New York, NY) for 120 h. The cultures were labeled with3H-thymidine during the last 8 h of culture (Amersham PharmaciaBiotech, Piscataway, NJ). Cells then were harvested, and radionuclideuptake was measured by a scintillation counting machine. In a similarmanner, we also tested whether Treg could suppress recall responses tomumps antigens.
Flow CytometryCells that were harvested from generating cultures (Treg or TCon-
trol) were analyzed using flow cytometric analysis. A total of 2 � 105
cells per sample were stained with anti-human CD4-allophycocyanin(APC), CD25-phycoerythrin (PE), and glucocorticoid-induced TNF re-ceptor (GITR)-FITC, CD8-APC (BD Bioscience; eBioscience, San Diego,CA). For the intracellular cytotoxic T lymphocyte–associated antigen-4(CTLA-4) staining, cells were permeabilized with Perm Buffer (BDBiosciences) for 20 min at 4°C and labeled with CTLA-4 for 30 min at4°C. For flow cytometric analysis of Foxp3, 1 � 106 cells first werestained with anti-human CD4-APC and CD25-PE. After washing, cellswere resuspended in 1 ml of cold Fix/Perm Buffer (eBioscience) andincubated at 4°C overnight in the dark. After washing twice with 2 mlof permeabilization buffer, cells were blocked with 2% normal ratserum for 15 min. Anti-human Foxp3-FITC (PCH101; eBioscience) thenwas added, and cells were incubated at 4°C for another 30 min in thedark. Finally, cells were washed with 2 ml of permeabilization bufferand analyzed on a FACSCalibur flow cytometer using CellQuest soft-
ware (Becton Dickinson, San Jose, CA). For evaluation of the inductionof apoptosis/death of CD4�CD25� and CD4�CD25� T cells, PBL thatwere incubated with ATG or rabbit IgG were stained with mAb againstCD4, CD25, annexin-V, and 7-amino-actinomycin D as per the manu-facturer’s instructions (BD Bioscience).
ELISPOT AssayTo evaluate the frequency of cytokine-producing cells in the gener-
ating culture, we made use of the ELISPOT assay as described previ-ously (12). The resulting spots were counted on a computer-assistedELISASpot Image Analyzer (Cellular Technology Limited, Cleveland,OH). Cells were tested in triplicate wells. The frequencies then wereexpressed as the number of spots per million PBL.
Luminex AssaySupernatants from generating cultures (serum-free medium) were
tested for the presence of TGF-� by Luminex 100 system (LuminexCorp., Austin, TX). Beadlyte Human Multi-Cytokine Beadmaster Kitand Beadlyte Human TGF-�1/�2 Detection system (Upstate, Char-lottesville, VA) were used as per protocol provided by the manufac-turer. Plates were analyzed using Multiplex Data Analysis softwareversion 1 (Luminex, Austin, TX).
Carboxy-Fluorescein Diacetates Succinimidyl Ester StainingTo test whether the expansion of Treg by ATG is due to proliferation
of preexisting naturally occurring CD4�CD25� T cells, we incubatedPBL with carboxy-fluorescein diacetates succinimidyl ester (CFSE) inthe form of 5 mM stock solution in DMSO at a final concentration of 1�M for 6 min at room temperature. CFSE-labeled cells were cultured invitro with phytohemagglutinin (positive control), ATG, and rabbit IgGfor 72 h at 37°C. Cells then were stained with anti-human CD4-APC,CD8-PE, and CD25-PE. 7-Amino-actinomycin D was used to excludedeath cells.
Conversion of CD4�CD25� to CD4�CD25� T CellsFor testing whether Treg that are generated by ATG are induced
from CD4�CD25� T cells, PBL were depleted of CD25 by magnetic cellsorting using MACS columns and MACS separators (Miltenyi Biotec,Auburn, CA). CD25-depleted CD4� T cells then were incubated withATG or rabbit IgG for 24 h. The cells then were harvested and stainedfor CD25 and regulatory markers. Similarly, their suppressor activitywas assessed as described previously.
Statistical AnalysesThe t test was used for comparison of means between experimental
groups examined by flow cytometry and ELISPOT assay. Differenceswere considered to be significant at P � 0.05.
ResultsATG Expands CD4�CD25�Foxp3� T Cells Ex Vivo
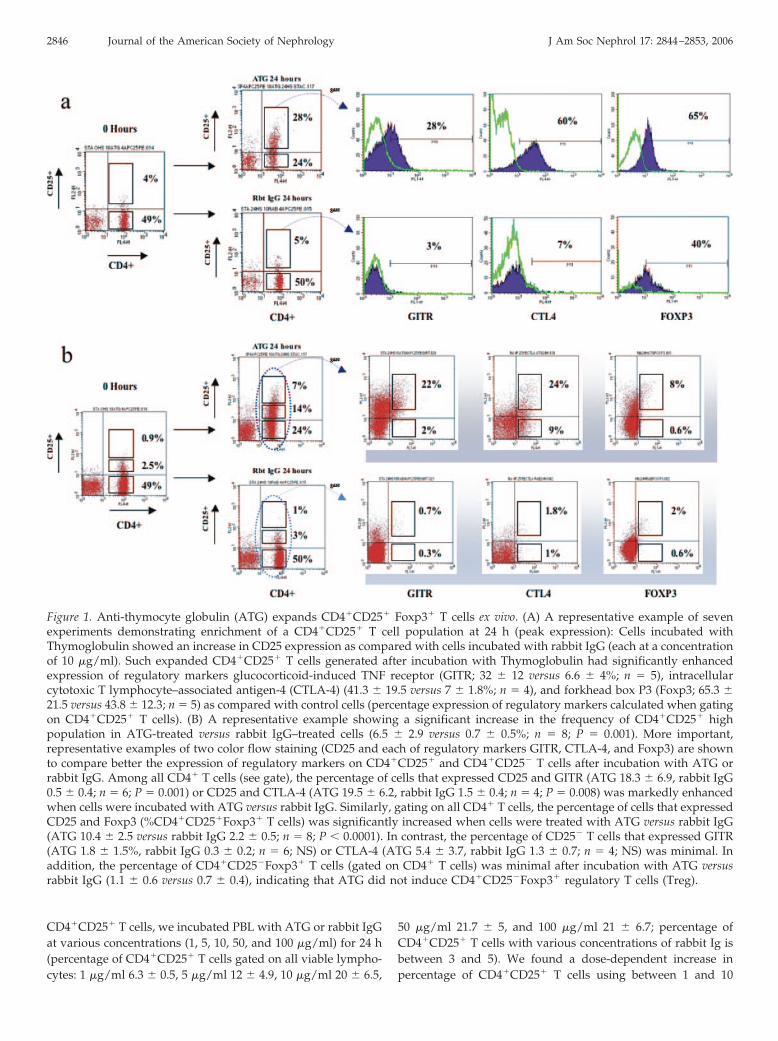
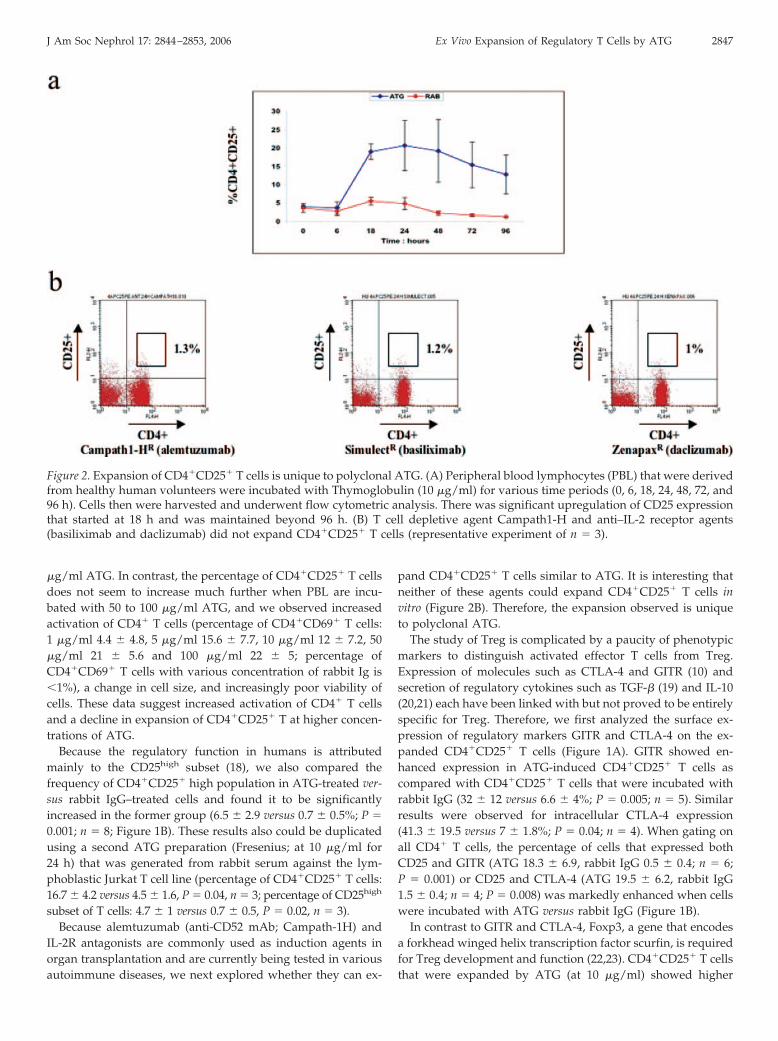
PBL that were derived from healthy volunteers were incu-bated with Thymoglobulin or rabbit IgG (10 �g/ml) for varioustime periods (0, 6, 18, 24, 48, 72, and 96 h). Flow cytometricanalysis of harvested cells demonstrated a significant upregu-lation of CD25 expression that began at 18 h and was main-tained beyond 96 h, with peak expression of CD25 at 24 h(percentage of CD4�CD25� T cells gated on all viable lympho-cytes 20.5 � 7.8 versus 4.5 � 1.6; P � 0.002; n � 7; Figures 1Aand 2A).
To evaluate the dose-response of ATG and expansion of
J Am Soc Nephrol 17: 2844–2853, 2006 Ex Vivo Expansion of Regulatory T Cells by ATG 2845
CD4�CD25� T cells, we incubated PBL with ATG or rabbit IgGat various concentrations (1, 5, 10, 50, and 100 �g/ml) for 24 h(percentage of CD4�CD25� T cells gated on all viable lympho-cytes: 1 �g/ml 6.3 � 0.5, 5 �g/ml 12 � 4.9, 10 �g/ml 20 � 6.5,
50 �g/ml 21.7 � 5, and 100 �g/ml 21 � 6.7; percentage ofCD4�CD25� T cells with various concentrations of rabbit Ig isbetween 3 and 5). We found a dose-dependent increase inpercentage of CD4�CD25� T cells using between 1 and 10
Figure 1. Anti-thymocyte globulin (ATG) expands CD4�CD25� Foxp3� T cells ex vivo. (A) A representative example of sevenexperiments demonstrating enrichment of a CD4�CD25� T cell population at 24 h (peak expression): Cells incubated withThymoglobulin showed an increase in CD25 expression as compared with cells incubated with rabbit IgG (each at a concentrationof 10 �g/ml). Such expanded CD4�CD25� T cells generated after incubation with Thymoglobulin had significantly enhancedexpression of regulatory markers glucocorticoid-induced TNF receptor (GITR; 32 � 12 versus 6.6 � 4%; n � 5), intracellularcytotoxic T lymphocyte–associated antigen-4 (CTLA-4) (41.3 � 19.5 versus 7 � 1.8%; n � 4), and forkhead box P3 (Foxp3; 65.3 �21.5 versus 43.8 � 12.3; n � 5) as compared with control cells (percentage expression of regulatory markers calculated when gatingon CD4�CD25� T cells). (B) A representative example showing a significant increase in the frequency of CD4�CD25� highpopulation in ATG-treated versus rabbit IgG–treated cells (6.5 � 2.9 versus 0.7 � 0.5%; n � 8; P � 0.001). More important,representative examples of two color flow staining (CD25 and each of regulatory markers GITR, CTLA-4, and Foxp3) are shownto compare better the expression of regulatory markers on CD4�CD25� and CD4�CD25� T cells after incubation with ATG orrabbit IgG. Among all CD4� T cells (see gate), the percentage of cells that expressed CD25 and GITR (ATG 18.3 � 6.9, rabbit IgG0.5 � 0.4; n � 6; P � 0.001) or CD25 and CTLA-4 (ATG 19.5 � 6.2, rabbit IgG 1.5 � 0.4; n � 4; P � 0.008) was markedly enhancedwhen cells were incubated with ATG versus rabbit IgG. Similarly, gating on all CD4� T cells, the percentage of cells that expressedCD25 and Foxp3 (%CD4�CD25�Foxp3� T cells) was significantly increased when cells were treated with ATG versus rabbit IgG(ATG 10.4 � 2.5 versus rabbit IgG 2.2 � 0.5; n � 8; P � 0.0001). In contrast, the percentage of CD25� T cells that expressed GITR(ATG 1.8 � 1.5%, rabbit IgG 0.3 � 0.2; n � 6; NS) or CTLA-4 (ATG 5.4 � 3.7, rabbit IgG 1.3 � 0.7; n � 4; NS) was minimal. Inaddition, the percentage of CD4�CD25�Foxp3� T cells (gated on CD4� T cells) was minimal after incubation with ATG versusrabbit IgG (1.1 � 0.6 versus 0.7 � 0.4), indicating that ATG did not induce CD4�CD25�Foxp3� regulatory T cells (Treg).
2846 Journal of the American Society of Nephrology J Am Soc Nephrol 17: 2844–2853, 2006
�g/ml ATG. In contrast, the percentage of CD4�CD25� T cellsdoes not seem to increase much further when PBL are incu-bated with 50 to 100 �g/ml ATG, and we observed increasedactivation of CD4� T cells (percentage of CD4�CD69� T cells:1 �g/ml 4.4 � 4.8, 5 �g/ml 15.6 � 7.7, 10 �g/ml 12 � 7.2, 50�g/ml 21 � 5.6 and 100 �g/ml 22 � 5; percentage ofCD4�CD69� T cells with various concentration of rabbit Ig is�1%), a change in cell size, and increasingly poor viability ofcells. These data suggest increased activation of CD4� T cellsand a decline in expansion of CD4�CD25� T at higher concen-trations of ATG.
Because the regulatory function in humans is attributedmainly to the CD25high subset (18), we also compared thefrequency of CD4�CD25� high population in ATG-treated ver-sus rabbit IgG–treated cells and found it to be significantlyincreased in the former group (6.5 � 2.9 versus 0.7 � 0.5%; P �
0.001; n � 8; Figure 1B). These results also could be duplicatedusing a second ATG preparation (Fresenius; at 10 �g/ml for24 h) that was generated from rabbit serum against the lym-phoblastic Jurkat T cell line (percentage of CD4�CD25� T cells:16.7 � 4.2 versus 4.5 � 1.6, P � 0.04, n � 3; percentage of CD25high
subset of T cells: 4.7 � 1 versus 0.7 � 0.5, P � 0.02, n � 3).Because alemtuzumab (anti-CD52 mAb; Campath-1H) and
IL-2R antagonists are commonly used as induction agents inorgan transplantation and are currently being tested in variousautoimmune diseases, we next explored whether they can ex-
pand CD4�CD25� T cells similar to ATG. It is interesting thatneither of these agents could expand CD4�CD25� T cells invitro (Figure 2B). Therefore, the expansion observed is uniqueto polyclonal ATG.
The study of Treg is complicated by a paucity of phenotypicmarkers to distinguish activated effector T cells from Treg.Expression of molecules such as CTLA-4 and GITR (10) andsecretion of regulatory cytokines such as TGF-� (19) and IL-10(20,21) each have been linked with but not proved to be entirelyspecific for Treg. Therefore, we first analyzed the surface ex-pression of regulatory markers GITR and CTLA-4 on the ex-panded CD4�CD25� T cells (Figure 1A). GITR showed en-hanced expression in ATG-induced CD4�CD25� T cells ascompared with CD4�CD25� T cells that were incubated withrabbit IgG (32 � 12 versus 6.6 � 4%; P � 0.005; n � 5). Similarresults were observed for intracellular CTLA-4 expression(41.3 � 19.5 versus 7 � 1.8%; P � 0.04; n � 4). When gating onall CD4� T cells, the percentage of cells that expressed bothCD25 and GITR (ATG 18.3 � 6.9, rabbit IgG 0.5 � 0.4; n � 6;P � 0.001) or CD25 and CTLA-4 (ATG 19.5 � 6.2, rabbit IgG1.5 � 0.4; n � 4; P � 0.008) was markedly enhanced when cellswere incubated with ATG versus rabbit IgG (Figure 1B).
In contrast to GITR and CTLA-4, Foxp3, a gene that encodesa forkhead winged helix transcription factor scurfin, is requiredfor Treg development and function (22,23). CD4�CD25� T cellsthat were expanded by ATG (at 10 �g/ml) showed higher
Figure 2. Expansion of CD4�CD25� T cells is unique to polyclonal ATG. (A) Peripheral blood lymphocytes (PBL) that were derivedfrom healthy human volunteers were incubated with Thymoglobulin (10 �g/ml) for various time periods (0, 6, 18, 24, 48, 72, and96 h). Cells then were harvested and underwent flow cytometric analysis. There was significant upregulation of CD25 expressionthat started at 18 h and was maintained beyond 96 h. (B) T cell depletive agent Campath1-H and anti–IL-2 receptor agents(basiliximab and daclizumab) did not expand CD4�CD25� T cells (representative experiment of n � 3).
J Am Soc Nephrol 17: 2844–2853, 2006 Ex Vivo Expansion of Regulatory T Cells by ATG 2847

expression of Foxp3 as compared with those that were incu-bated with rabbit IgG (63.4 � 12 versus 47 � 8.3%; P � 0.03; n �
5; Figure 1A). It is interesting that contrary to CD69 expression(see above), the Foxp3 expression declined with increasingdosage of ATG in the generating culture (percentage of Foxp3expression in CD4�CD25� T cells: 50 �g/ml 13 � 4.2, 100�g/ml 15.2 � 0.35). Importantly, gating on all CD4� T cells, thepercentage of cells that expressed both CD25 and Foxp3 (per-centage of CD4�CD25�Foxp3� T cells) was significantly in-creased when cells were treated with ATG versus rabbit IgG(ATG 10.4 � 2.5 versus rabbit IgG 2.2 � 0.5; P � 0.0001; n � 8;Figure 1B). In addition, expression of all of the regulatorymarkers was even markedly more enhanced in theCD4�CD25� high population that was expanded after incuba-tion with ATG (GITR 49.4 � 15.9%, n � 7; CTLA-4 55 � 24.4%,n � 6; Foxp3 71 � 14.7%, n � 5).
Previous work indicated that Foxp3 also can be induced inCD4�CD25� T cells and that these cells can function as Treg(24). However, in contrast to CD4�CD25� T cells, CD4�CD25�
T cells that were incubated with ATG or rabbit IgG showedonly minimal increase in GITR (5.6 � 4.4 versus 0) and CTLA-4(11.5 � 4.2 versus 0). Similarly, gating on all CD4� T cells, thepercentage of CD25� T cells that expressed GITR (ATG 1.8 �
1.5% rabbit IgG 0.3 � 0.2; n � 6; NS) or CTLA-4 (ATG 5.4 � 3.7,rabbit IgG 1.3 � 0.7; n � 4; NS) was minimal (Figure 1B). Inaddition, the percentage of CD4�CD25�Foxp3� T cells (gatedon CD4� T cells) was minimal after incubation with ATG versusrabbit IgG (1.1 � 0.6 versus 0.7 � 0.4), indicating that afterincubation with ATG, CD4�CD25� T cells remain Foxp3� (Fig-ure 1B). Although overall there was a slight decrease in theCD8� T cells after incubation with ATG as compared withrabbit IgG (21.96 � 4.5 versus 25.6 � 5.1; n � 8; P � 0.02), wefound no significant difference in the percentage ofCD8�CD28� T cells (11.3 � 5.6 versus 14.9 � 6; n � 5). Inaddition, we found no evidence of CD8�Foxp3� T cells beforeor after treatment with ATG or rabbit IgG. Taken together,these data do not indicate expansion of other previously de-scribed Treg populations in our model.
Suppressor Function of CD4�CD25� T Cells Generated byATG Is Restricted to Responder Cells
After demonstrating that CD4�CD25� T cells that are ex-panded by ATG can maintain their regulatory phenotype, wenext set out to study the actual suppressor function of thesecells in vitro. The in vitro suppressive activity of the cells wasevaluated by examination of their ability to suppress an MLR todonor alloantigens. We found significant suppression of directalloimmune responses of the original responders (PBL fromwhich the Treg initially were generated) to stimulator cells byTreg but not by TControl (1:1 ratio, 61.3 � 7.4 inhibition versus20.2 � 18.4%; n � 4; P � 0.01; Figure 3A). It is interesting thatthe same Treg were unable to suppress an MLR of respondercells that were not autologous to the original responder cells(Figure 3B). We next tested the ability of these Treg to suppressa recall memory response of original responders to mumpsantigen. It is interesting that the proliferative response to recallantigen mumps was not inhibited by Treg, indicating that the
expanded Treg did not block memory cells to the antigen(mumps alone 4848 � 969, Treg 5214 � 900, control 9595 �
1231; Figure 3C).
Conversion of CD4�CD25� into CD4�CD25� T Cells Isthe Main Mechanism of Expansion of Treg by ATG
The absolute number of CD4�CD25� T cells (mean values ofcell numbers counted in wells) that were incubated with ATG(before 75,833 � 31,051, after 639,167 � 249,448; n � 6; P �
0.002) but not rabbit IgG was dramatically increased (before90,833 � 33,229, after 113,333 � 54,283; NS) after 24 h ofincubation. In contrast, there was much more decrease in thenumber of CD4�CD25� T cells that were treated with ATG(before 1878,300 � 322,020, after 1082,500 � 301,027; P � 0.005)as compared with rabbit IgG (1991,700 � 379,548 versus1861,666 � 238 362; NS). On the basis of these findings, wehypothesized that the expansion of Treg by ATG may resultfrom one or more mutually nonexclusive and possibly comple-mentary mechanisms: First, ATG may preferentially promotedeath/apoptosis of CD4�CD25� T cells as compared withCD4�CD25� T cells, creating a new balance in favor of thelatter cells. Second, ATG may promote the proliferation ofalready existing, naturally occurring CD4�CD25� T cells.Third, ATG may be able to convert CD4�CD25� T cells intoCD4�CD25� T cells with regulatory functions (induction ofTreg). The first possibility is supported by published data dem-onstrating that ATG in fact can bind to multiple epitopes on theT cell surface and induce apoptosis in T lymphocytes throughFas-L (CD95L) (3,5). Nevertheless, we found no difference (gat-ing on all lymphocytes without excluding the dead cells) inapoptosis of CD4�CD25� T cells and CD4�CD25� T cells thatwere incubated for 24 h with 10 �g/ml ATG (6.7 � 3.1 versus5 � 4.7%) or rabbit IgG (5 � 3 versus 3.2 � 2.5%). In fact, theoverall viability of the cells was �95% regardless of incubationwith ATG or rabbit IgG.
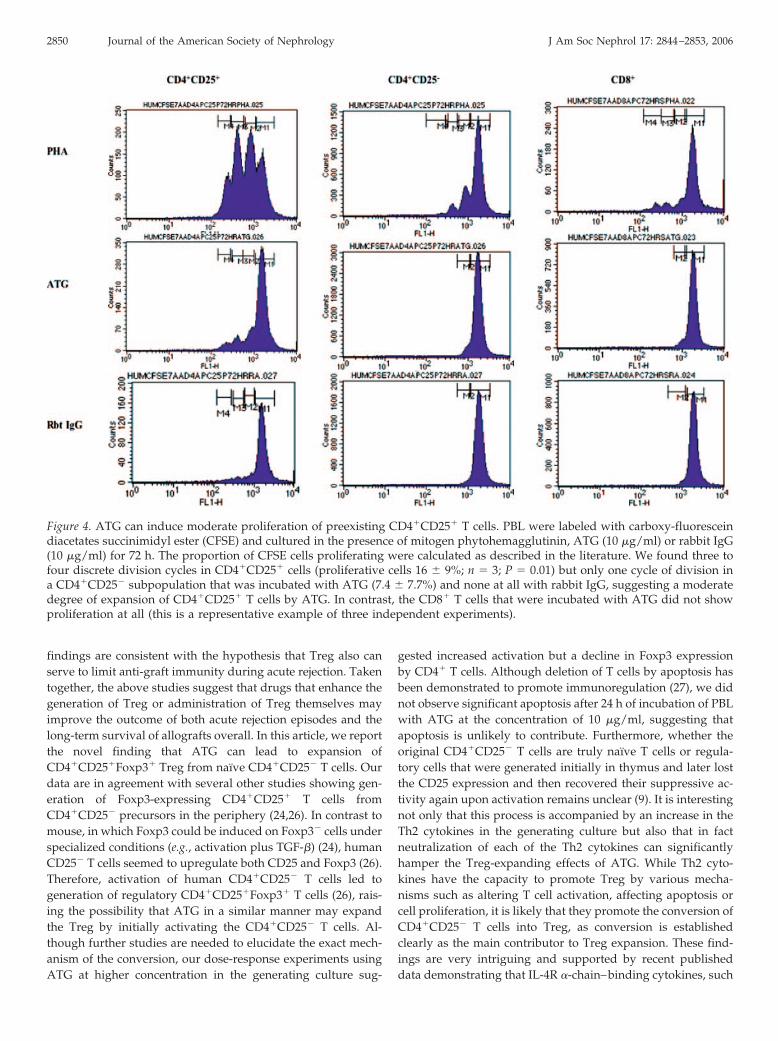
To address the possibility of the proliferative effect of ATGon already existing, naturally occurring Treg, we cultured invitro CFSE-labeled PBL with ATG or rabbit IgG for 72 h (10�g/ml). We found three to four discrete division cycles inCD4�CD25� cells (proliferative cells 16 � 9%; n � 3; P � 0.01)but only one cycle of division in a CD4�CD25� subpopulationthat was incubated with ATG (7.4 � 7.7%) and none at all withrabbit IgG, suggesting a moderate degree of expansion ofCD4�CD25� T cells. It is possible that some of the proliferatingCD4�CD25� T cells simply may be activated T cells rather thanproliferating naturally occurring Treg. Nevertheless, whereasthere is an eight-fold expansion of CD4�CD25� T cells at 24 hwith ATG, proliferating cells make up only approximately 16%of the CD25� population at 72 h. This indicates that prolifera-tion is unlikely to contribute significantly to the expansion ofthe cells. In contrast, the CD8� T cells that were incubated withATG did not show proliferation at all (Figure 4).
To determine whether ATG is capable of convertingCD4�CD25� cells into CD4�CD25� cells with regulatory prop-erties, we first depleted naturally occurring CD25� T cells fromPBL ex vivo with magnetic bead separation (before 5.9 � 2.9versus after depletion 0.6 � 0.9%; Figure 5A). Such CD25-
2848 Journal of the American Society of Nephrology J Am Soc Nephrol 17: 2844–2853, 2006
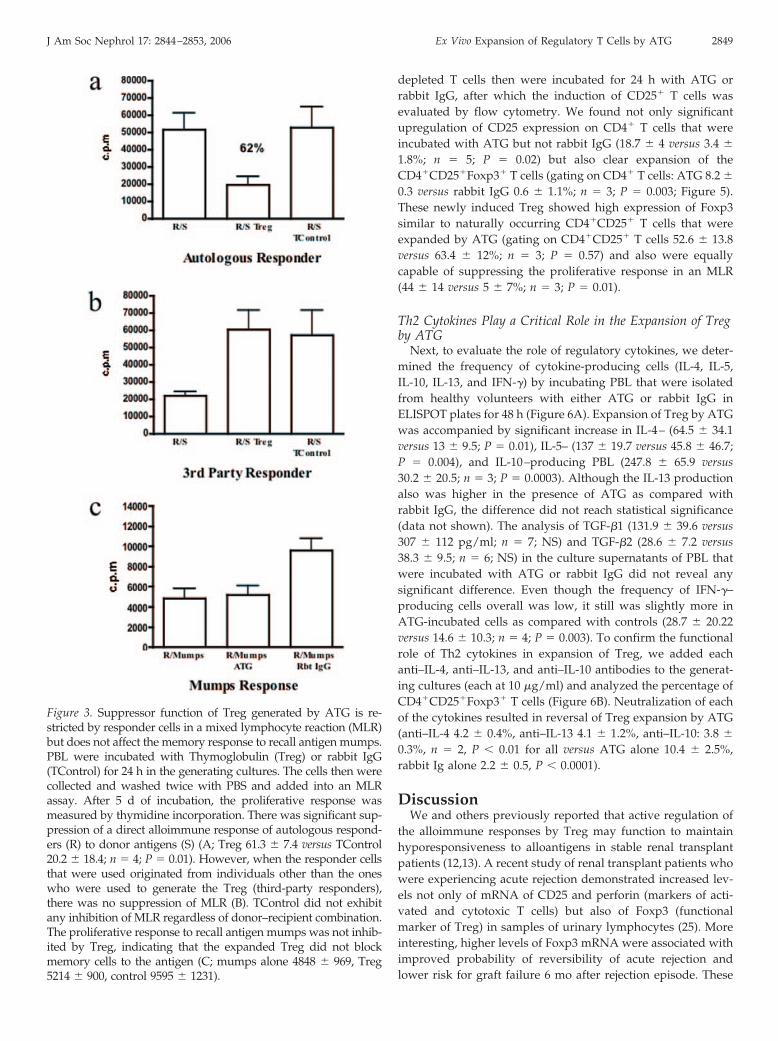
depleted T cells then were incubated for 24 h with ATG orrabbit IgG, after which the induction of CD25� T cells wasevaluated by flow cytometry. We found not only significantupregulation of CD25 expression on CD4� T cells that wereincubated with ATG but not rabbit IgG (18.7 � 4 versus 3.4 �
1.8%; n � 5; P � 0.02) but also clear expansion of theCD4�CD25�Foxp3� T cells (gating on CD4� T cells: ATG 8.2 �
0.3 versus rabbit IgG 0.6 � 1.1%; n � 3; P � 0.003; Figure 5).These newly induced Treg showed high expression of Foxp3similar to naturally occurring CD4�CD25� T cells that wereexpanded by ATG (gating on CD4�CD25� T cells 52.6 � 13.8versus 63.4 � 12%; n � 3; P � 0.57) and also were equallycapable of suppressing the proliferative response in an MLR(44 � 14 versus 5 � 7%; n � 3; P � 0.01).
Th2 Cytokines Play a Critical Role in the Expansion of Tregby ATG
Next, to evaluate the role of regulatory cytokines, we deter-mined the frequency of cytokine-producing cells (IL-4, IL-5,IL-10, IL-13, and IFN-�) by incubating PBL that were isolatedfrom healthy volunteers with either ATG or rabbit IgG inELISPOT plates for 48 h (Figure 6A). Expansion of Treg by ATGwas accompanied by significant increase in IL-4– (64.5 � 34.1versus 13 � 9.5; P � 0.01), IL-5– (137 � 19.7 versus 45.8 � 46.7;P � 0.004), and IL-10–producing PBL (247.8 � 65.9 versus30.2 � 20.5; n � 3; P � 0.0003). Although the IL-13 productionalso was higher in the presence of ATG as compared withrabbit IgG, the difference did not reach statistical significance(data not shown). The analysis of TGF-�1 (131.9 � 39.6 versus307 � 112 pg/ml; n � 7; NS) and TGF-�2 (28.6 � 7.2 versus38.3 � 9.5; n � 6; NS) in the culture supernatants of PBL thatwere incubated with ATG or rabbit IgG did not reveal anysignificant difference. Even though the frequency of IFN-�–producing cells overall was low, it still was slightly more inATG-incubated cells as compared with controls (28.7 � 20.22versus 14.6 � 10.3; n � 4; P � 0.003). To confirm the functionalrole of Th2 cytokines in expansion of Treg, we added eachanti–IL-4, anti–IL-13, and anti–IL-10 antibodies to the generat-ing cultures (each at 10 �g/ml) and analyzed the percentage ofCD4�CD25�Foxp3� T cells (Figure 6B). Neutralization of eachof the cytokines resulted in reversal of Treg expansion by ATG(anti–IL-4 4.2 � 0.4%, anti–IL-13 4.1 � 1.2%, anti–IL-10: 3.8 �
0.3%, n � 2, P � 0.01 for all versus ATG alone 10.4 � 2.5%,rabbit Ig alone 2.2 � 0.5, P � 0.0001).
DiscussionWe and others previously reported that active regulation of
the alloimmune responses by Treg may function to maintainhyporesponsiveness to alloantigens in stable renal transplantpatients (12,13). A recent study of renal transplant patients whowere experiencing acute rejection demonstrated increased lev-els not only of mRNA of CD25 and perforin (markers of acti-vated and cytotoxic T cells) but also of Foxp3 (functionalmarker of Treg) in samples of urinary lymphocytes (25). Moreinteresting, higher levels of Foxp3 mRNA were associated withimproved probability of reversibility of acute rejection andlower risk for graft failure 6 mo after rejection episode. These
Figure 3. Suppressor function of Treg generated by ATG is re-stricted by responder cells in a mixed lymphocyte reaction (MLR)but does not affect the memory response to recall antigen mumps.PBL were incubated with Thymoglobulin (Treg) or rabbit IgG(TControl) for 24 h in the generating cultures. The cells then werecollected and washed twice with PBS and added into an MLRassay. After 5 d of incubation, the proliferative response wasmeasured by thymidine incorporation. There was significant sup-pression of a direct alloimmune response of autologous respond-ers (R) to donor antigens (S) (A; Treg 61.3 � 7.4 versus TControl20.2 � 18.4; n � 4; P � 0.01). However, when the responder cellsthat were used originated from individuals other than the oneswho were used to generate the Treg (third-party responders),there was no suppression of MLR (B). TControl did not exhibitany inhibition of MLR regardless of donor–recipient combination.The proliferative response to recall antigen mumps was not inhib-ited by Treg, indicating that the expanded Treg did not blockmemory cells to the antigen (C; mumps alone 4848 � 969, Treg5214 � 900, control 9595 � 1231).
J Am Soc Nephrol 17: 2844–2853, 2006 Ex Vivo Expansion of Regulatory T Cells by ATG 2849
findings are consistent with the hypothesis that Treg also canserve to limit anti-graft immunity during acute rejection. Takentogether, the above studies suggest that drugs that enhance thegeneration of Treg or administration of Treg themselves mayimprove the outcome of both acute rejection episodes and thelong-term survival of allografts overall. In this article, we reportthe novel finding that ATG can lead to expansion ofCD4�CD25�Foxp3� Treg from naı̈ve CD4�CD25� T cells. Ourdata are in agreement with several other studies showing gen-eration of Foxp3-expressing CD4�CD25� T cells fromCD4�CD25� precursors in the periphery (24,26). In contrast tomouse, in which Foxp3 could be induced on Foxp3� cells underspecialized conditions (e.g., activation plus TGF-�) (24), humanCD25� T cells seemed to upregulate both CD25 and Foxp3 (26).Therefore, activation of human CD4�CD25� T cells led togeneration of regulatory CD4�CD25�Foxp3� T cells (26), rais-ing the possibility that ATG in a similar manner may expandthe Treg by initially activating the CD4�CD25� T cells. Al-though further studies are needed to elucidate the exact mech-anism of the conversion, our dose-response experiments usingATG at higher concentration in the generating culture sug-
gested increased activation but a decline in Foxp3 expressionby CD4� T cells. Although deletion of T cells by apoptosis hasbeen demonstrated to promote immunoregulation (27), we didnot observe significant apoptosis after 24 h of incubation of PBLwith ATG at the concentration of 10 �g/ml, suggesting thatapoptosis is unlikely to contribute. Furthermore, whether theoriginal CD4�CD25� T cells are truly naı̈ve T cells or regula-tory cells that were generated initially in thymus and later lostthe CD25 expression and then recovered their suppressive ac-tivity again upon activation remains unclear (9). It is interestingnot only that this process is accompanied by an increase in theTh2 cytokines in the generating culture but also that in factneutralization of each of the Th2 cytokines can significantlyhamper the Treg-expanding effects of ATG. While Th2 cyto-kines have the capacity to promote Treg by various mecha-nisms such as altering T cell activation, affecting apoptosis orcell proliferation, it is likely that they promote the conversion ofCD4�CD25� T cells into Treg, as conversion is establishedclearly as the main contributor to Treg expansion. These find-ings are very intriguing and supported by recent publisheddata demonstrating that IL-4R �-chain–binding cytokines, such
Figure 4. ATG can induce moderate proliferation of preexisting CD4�CD25� T cells. PBL were labeled with carboxy-fluoresceindiacetates succinimidyl ester (CFSE) and cultured in the presence of mitogen phytohemagglutinin, ATG (10 �g/ml) or rabbit IgG(10 �g/ml) for 72 h. The proportion of CFSE cells proliferating were calculated as described in the literature. We found three tofour discrete division cycles in CD4�CD25� cells (proliferative cells 16 � 9%; n � 3; P � 0.01) but only one cycle of division ina CD4�CD25� subpopulation that was incubated with ATG (7.4 � 7.7%) and none at all with rabbit IgG, suggesting a moderatedegree of expansion of CD4�CD25� T cells by ATG. In contrast, the CD8� T cells that were incubated with ATG did not showproliferation at all (this is a representative example of three independent experiments).
2850 Journal of the American Society of Nephrology J Am Soc Nephrol 17: 2844–2853, 2006

as IL-4 and IL-13, play a crucial role in generatingFoxp3�CD25� Treg extrathymically (17). In addition, thesedata point to an intriguing link between the well-establishedimmunoregulatory capacity of Th2 cells and the potentCD4�CD25� regulatory T cells. These results also are impor-tant in light of data showing that cyclosporine and tacrolimus,two commonly used immunosuppressive drugs, both inhibitthe production of IL-2, an essential growth factor for Treg (28).Conversely, murine T cells that were activated in the presenceof rapamycin were highly enriched in Foxp3�CD4�CD25� Tcells in vitro (29). Nevertheless, our findings here are uniquebecause we clearly demonstrate that ATG can rapidly expandhuman Treg by promotion of Th2 cytokine production ex vivo.
Although the identification of the exact component of ATG(Genzyme; generated by using thymocytes as the immunogen)that is responsible for the expansion of Treg will require furtherinvestigation, our ability to generate Treg using a different ATG(Fresenius; produced with purified and activated CD3� lym-phoblastic cells as immunogen) suggests a key role for antibod-ies that are directed against epitopes on T cells. Another uniquefinding of the study is that ATG-induced Treg significantlysuppressed direct alloimmune responses to alloantigens byautologous PBL from which Treg originally were generated. Incontrast, nonautologous responders could not be regulated bythe Treg. Conversely, the memory response to recall antigenmumps is not affected. This may be due to lack of activation ofTreg by mumps antigen or the overall inability of Treg to
inhibit the memory responses. Given the great interest in cel-lular treatment with ex vivo expanded Treg as a means tomodulate alloimmune responses without affecting memory re-sponses to infectious agents, these findings are of crucial im-portance.
Although the in vivo effects of ATG on the induction, expan-sion, and function of Treg in allograft recipients remain to becharacterized fully, recent data demonstrated that CD25� Tcells are spared from anti-lymphocyte serum–mediated dele-tion in mice, suggesting the concomitant presence of both T celldepletion and continuous regulatory T cell activity in an in vivoanimal model (30). Second, in studying the in vivo effects ofATG in humans, the achieved concentrations of the ATG inblood and lymphoid tissues should be taken carefully intoconsideration. ATG is used at a dosage of 1 to 1.5 mg/kg per dfor 3 to 5 d as induction immunosuppression and also fortreatment of steroid-resistant acute rejection, leading to serumlevels of the drug between 50 and 100 �g/ml (2,5). This dosingregimen is based on ATG’s efficacy to deplete T cells in theperipheral blood rather than promotion of Treg. There are nohuman data regarding the achieved concentrations in second-ary lymphoid tissues, a potentially important site for induc-tion/expansion of Treg, although the data in primates suggestgood penetration into lymph nodes and spleen but minimalpenetration into thymus (31). We were able to expand Treg invitro with a concentration of 10 �g/ml, a dosage that is signif-icantly lower than the levels that currently are achieved in
Figure 5. ATG induces Treg from CD4�CD25� T cells. PBL were depleted of CD25 using MACS columns. Such CD25-depleted Tcells then were incubated for 24 h with ATG or rabbit IgG. Flow cytometric analysis showed an increase in CD25 expression onCD4� T cells (representative example of three independent experiments shown). The two-color staining demonstrated also clearexpansion of the CD4�CD25�Foxp3� T cells (ATG 8.2 � 0.3 versus rabbit IgG 0.6 � 1.1%; n � 3; P � 0.003, gated on all CD4� Tcells).
J Am Soc Nephrol 17: 2844–2853, 2006 Ex Vivo Expansion of Regulatory T Cells by ATG 2851

blood. In fact, our in vitro data may suggest that lower dosagesof ATG may lead to expansion of Treg in vivo, but clearlyfurther investigation is needed and is currently ongoing.
ConclusionWe report a novel mechanism of action for ATG, namely its
ability to expand Treg ex vivo, mainly by inducingCD4�CD25�Foxp3� T cells. The regulatory function clearly isrestricted by the responder cells but does not affect the memoryresponse to recall antigen. These findings are clinically impor-tant for two important reasons: First, they may suggest that thetherapeutic effect of ATG may be due to both concomitant T celldeletion and continuous regulatory T cell activity; second, theymay help us to develop strategies to expand these very rareregulatory cells ex vivo for use as cellular treatment in trans-plantation and autoimmunity.
AcknowledgmentsThis work was supported by National Institutes of Health grant PPG
PO1 AI-050157. Assay development was supported by National Insti-tutes of Health grants UO1 AI-055801 and UO1 AI-063623. N.N. is a
recipient of the American Society of Nephrology John Merrill Trans-plant Scholar Grant and the American Heart Association Scientist De-velopment Grant. M.H.S. is a consultant for Genzyme.
References1. Beiras-Fernandez A, Thein E, Hammer C: Induction of
immunosuppression with polyclonal antithymocyte glob-ulins: An overview. Exp Clin Transplant 1: 79–84, 2003
2. Lowsky R, Takahashi T, Liu YP, Dejbakhsh-Jones S, Gru-met FC, Shizuru JA, Laport GG, Stockerl-Goldstein KE,Johnston LJ, Hoppe RT, Bloch DA, Blume KG, Megrin RS,Strober S: Protective conditioning for acute graft-versus-host disease. N Engl J Med 353: 1321–1331, 2005
3. Genestier L, Fournel S, Flacher M, Assossou O, RevillardJP, Bonnefoy-Berard N: Induction of Fas (Apo-1, CD95)-mediated apoptosis of activated lymphocytes by poly-clonal antithymocyte globulins. Blood 91: 2360–2368, 1998
4. Michallet MC, Preville X, Flacher M, Fournel S, GenestierL, Revillard JP: Functional antibodies to leukocyte adhe-sion molecules in antithymocyte globulins. Transplantation75: 657–662, 2003
Figure 6. Th2 cytokines play a key role in the expansion of Treg. (A) Expansion of CD4�CD25� T cells by ATG in the generatingculture is accompanied by Th2 cytokine production: PBL from healthy donors were incubated in an ELISPOT plate in the presenceof ATG or rabbit IgG for 48 h (triplicate). The quantification of spots (mean of at least three independent experiments for eachcytokine) revealed significant increase of IL-4–, IL-5–, and IL-10–producing PBL that were incubated with ATG. (B) Neutralizationof Th2 cytokines decreases expansion of Treg: Anti–IL-4, anti–IL-10, and anti–IL-13 mAb or corresponding isotype controls eachwere added separately to PBL that were incubated with ATG or rabbit IgG (10 �g/ml). Cells then were harvested after 24 h, and%CD4�CD25�Foxp3� T cells (gated on CD4� lymphocytes) were measured by flow cytometry. The neutralization led to asignificant decline in percentage of CD4� T cells that expressed CD25 and Foxp3. Mean values of two independent experimentsare shown here.
2852 Journal of the American Society of Nephrology J Am Soc Nephrol 17: 2844–2853, 2006

5. Zand MS, Vo T, Huggins J, Felgar R, Liesveld J, Pellegrin T,Bozorgzadeh A, Sanz I, Briggs BJ: Polyclonal rabbit anti-thymocyte globulin triggers B-cell and plasma cell apopto-sis by multiple pathways. Transplantation 79: 1507–1515,2005
6. Brennan DC: Faith supported by reason: Mechanistic sup-port for the use of polyclonal antibodies in transplantation.Transplantation 75: 577–578, 2003
7. Fehervari Z, Sakaguchi S: Development and function ofCD25�CD4� regulatory T cells. Curr Opin Immunol 16:203–208, 2004
8. Sakaguchi S, Sakaguchi N: Regulatory T cells in immuno-logic self-tolerance and autoimmune disease. Int Rev Im-munol 24: 211–226, 2005
9. Sakaguchi S: The origin of FOXP3-expressing CD4� regu-latory T cells: Thymus or periphery. J Clin Invest 112:1310–1312, 2003
10. Wood KJ, Sakaguchi S: Regulatory T cells in transplanta-tion tolerance. Nat Rev Immunol 3: 199–210, 2003
11. Salama AD, Chaudhry AN, Holthaus KA, Mosley K, Kal-luri R, Sayegh MH, Lechler RI, Pusey CD, Lightstone L:Regulation by CD25� lymphocytes of autoantigen-specificT-cell responses in Goodpasture’s (anti-GBM) disease. Kid-ney Int 64: 1685–1694, 2003
12. Najafian N, Salama AD, Fedoseyeva EV, Benichou G,Sayegh MH: Enzyme-linked immunosorbent spot assayanalysis of peripheral blood lymphocyte reactivity to do-nor HLA-DR peptides: Potential novel assay for predictionof outcomes for renal transplant recipients. J Am Soc Neph-rol 13: 252–259, 2002
13. Salama AD, Najafian N, Clarkson MR, Harmon WE,Sayegh MH: Regulatory CD25� T cells in human kidneytransplant recipients. J Am Soc Nephrol 14: 1643–1651, 2003
14. Taylor PA, Lees CJ, Blazar BR: The infusion of ex vivoactivated and expanded CD4(�)CD25(�) immune regula-tory cells inhibits graft-versus-host disease lethality. Blood99: 3493–3499, 2002
15. Bluestone JA: Regulatory T-cell therapy: Is it ready for theclinic? Nat Rev Immunol 5: 343–349, 2005
16. Gregori S, Bacchetta R, Hauben E, Battaglia M, RoncaroloMG: Regulatory T cells: Prospective for clinical applicationin hematopoietic stem cell transplantation. Curr Opin He-matol 12: 451–456, 2005
17. Skapenko A, Kalden JR, Lipsky PE, Schulze-Koops H: TheIL-4 receptor alpha-chain-binding cytokines, IL-4 and IL-13, induce forkhead box P3-expressing CD25�CD4� reg-ulatory T cells from CD25-CD4� precursors. J Immunol175: 6107–6116, 2005
18. Baecher-Allan C, Brown JA, Freeman GJ, Hafler DA:
CD4�CD25high regulatory cells in human peripheralblood. J Immunol 167: 1245–1253, 2001
19. Nakamura K, Kitani A, Fuss I, Pedersen A, Harada N,Nawata H, Strober W: TGF-beta 1 plays an important rolein the mechanism of CD4�CD25� regulatory T cell activ-ity in both humans and mice. J Immunol 172: 834–842, 2004
20. Hara M, Kingsley CI, Niimi M, Read S, Turvey SE, BushellAR, Morris PJ, Powrie F, Wood KJ: IL-10 is required forregulatory T cells to mediate tolerance to alloantigens invivo. J Immunol 166: 3789–3796, 2001
21. Kohyama M, Sugahara D, Sugiyama S, Yagita H, OkumuraK, Hozumi N: Inducible costimulator-dependent IL-10production by regulatory T cells specific for self-antigen.Proc Natl Acad Sci U S A 101: 4192–4197, 2004
22. Fontenot JD, Gavin MA, Rudensky AY: Foxp3 programsthe development and function of CD4�CD25� regulatoryT cells. Nat Immunol 4: 330–336, 2003
23. Hori S, Nomura T, Sakaguchi S: Control of regulatory Tcell development by the transcription factor Foxp3. Science299: 1057–1061, 2003
24. Zheng SG, Wang JH, Gray JD, Soucier H, Horwitz DA: Nat-ural and induced CD4�CD25� cells educate CD4�CD25-cells to develop suppressive activity: The role of IL-2, TGF-beta, and IL-10. J Immunol 172: 5213–5221, 2004
25. Muthukumar T, Dadhania D, Ding R, Snopkowski C,Naqvi R, Lee JB, Hartono C, Li B, Sharma VK, Seshan SV,Kapur S, Schwartz JE, Hancock WW, Suthanthiram M:Messenger RNA for FOXP3 in the urine of renal-allograftrecipients. N Engl J Med 353: 2342–2351, 2005
26. Walker MR, Kasprowicz DJ, Gersuk VH, Benard A, VanLandeghen M, Buckner JH, Ziegler SF: Induction of FoxP3and acquisition of T regulatory activity by stimulated hu-man CD4�CD25- T cells. J Clin Invest 112: 1437–1443, 2003
27. Chiffoleau E, Walsh PT, Turka L: Apoptosis and transplan-tation tolerance. Immunol Rev 193: 124–145, 2003
28. Almeida AR, Legrand N, Papiernik M, Freitas AA: Ho-meostasis of peripheral CD4� T cells: IL-2R alpha and IL-2shape a population of regulatory cells that controls CD4�T cell numbers. J Immunol 169: 4850–4860, 2002
29. Battaglia M, Stabilini A, Roncarolo MG: Rapamycin selec-tively expands CD4�CD25�FoxP3� regulatory T cells.Blood 105: 4743–4748, 2005
30. Minamimura K, Gao W, Maki T: CD4� regulatory T cellsare spared from deletion by antilymphocyte serum, a poly-clonal anti-T cell antibody. J Immunol 176: 4125–4132, 2006
31. Preville X, Flacher M, LeMauff B, Beauchard S, Davelu P,Tiollier J, Revillard JP: Mechanisms involved in antithymo-cyte globulin immunosuppressive activity in a nonhumanprimate model. Transplantation 71: 460–468, 2001
See the related editorial, “Immunosuppression and Regulation: Cast in New Light,” on pages 2644–2646.
J Am Soc Nephrol 17: 2844–2853, 2006 Ex Vivo Expansion of Regulatory T Cells by ATG 2853


![Thymoglobulin (anti-thymocyte globulin [rabbit])products.sanofi.ca/en/Thymoglobulin.pdf · 2016-05-03 · sanofi-aventis Canada Inc. 2905 Place Louis-R.-Renaud . Laval, Quebec H7V](https://img.pdfslide.us/doc/110x75/5d16098d88c99312028e0754/thymoglobulin-anti-thymocyte-globulin-rabbit-2016-05-03-sanofi-aventis.jpg)


























