Embed Size (px)
Citation preview

Proc. Indian Acad. Sci. IPlant Sci.), Vol. 92. Number 3, June 1983, pp. 259-264. ~) Printed in India.
A n e w tr iploid race ot Pennisetum orientale R i c h e x h i b i t i n g e h r o m o s o m a l numerica l mosa i c i sm and neo-centr ic activity
Z VISHNU VARDHAN and N LAKSHMI Department of Botany, Nagarjuna University, Nagarjunanagar 522 510, India
MS received 15 July 1982; revised 2 February 1983
Abslract. Ah allotriploid of P. orientale Rich with 2n=27 chromosomes was Iocated. Cytomorphological studie~ revealed that it is a new chromosomal race exhibiting chromosomal numerical mosaicism and neo-centric activity. Heterochromatic knobs present on the chromo6omes are considered as sites of neo-cent romeres. The possible causes of chromosomal mosaicism and neo-centric activity are discussed. Polyploidy coupled with hybridization seems to pl~y a major role in the evolution of this taxon and the aberrant meiotic behaviour is nota barrier for the survival of the species since it is an apomiet.
Keywor Triploid; Pennisetum orientale; chromosomal numefical mosaicism; neo-centdc activity.
1. Introduction
Pennisetum orientale Rich, a valuable pasture is considered to be highly nutritious, palatable, drought resistant and evergreen. The seeds were collected from a park in Hyderabad and raised in the botanic garden of the University. Earlier cytoiogical studies revealed the existence of divergent polyploid and aneuploid taxa with chromosome numbers 2n= 18, 27, ~ and 45, all derived from the same base number x=9 (Krishnaswamy 1941; Parodi 1946; Hrishi 1952; Sharma and De 1956; Patil et al 1962; Mehra et al 1968; Chatterji and Timothy 1969; Sisodia 1972; Mehra and Remanandan 1973). Since the morphology and cyto!ogy of the present material' is different from earlier records detailed cytomorphological studies have been made, the results of which are discussed in this paper.
2. Materials and methods
Usual acetocarmine technique was followed for meiotic studies. Fertility of the pollen gmins was determined on the basis of their stainability with 2%Iz Kl.solution.
3. Observations
3.1 Morphology
The present triploid P. orientale is an ascending forro from a stout densely tuIted, bmnched, creeping root stock. Pedicels of the involucels are hispid, bristles 0.8-2.4 cm Iong, all very slender, flexous, purplish and very unequal. The present form is remarkably different from others in its long extremely narrow leaves (0.3 cm broadk high number of tillers (120) and high involucel density t8.78 per cm) (figure 1).
259
260 Z Vishnu Vardhan and N Lakshmi
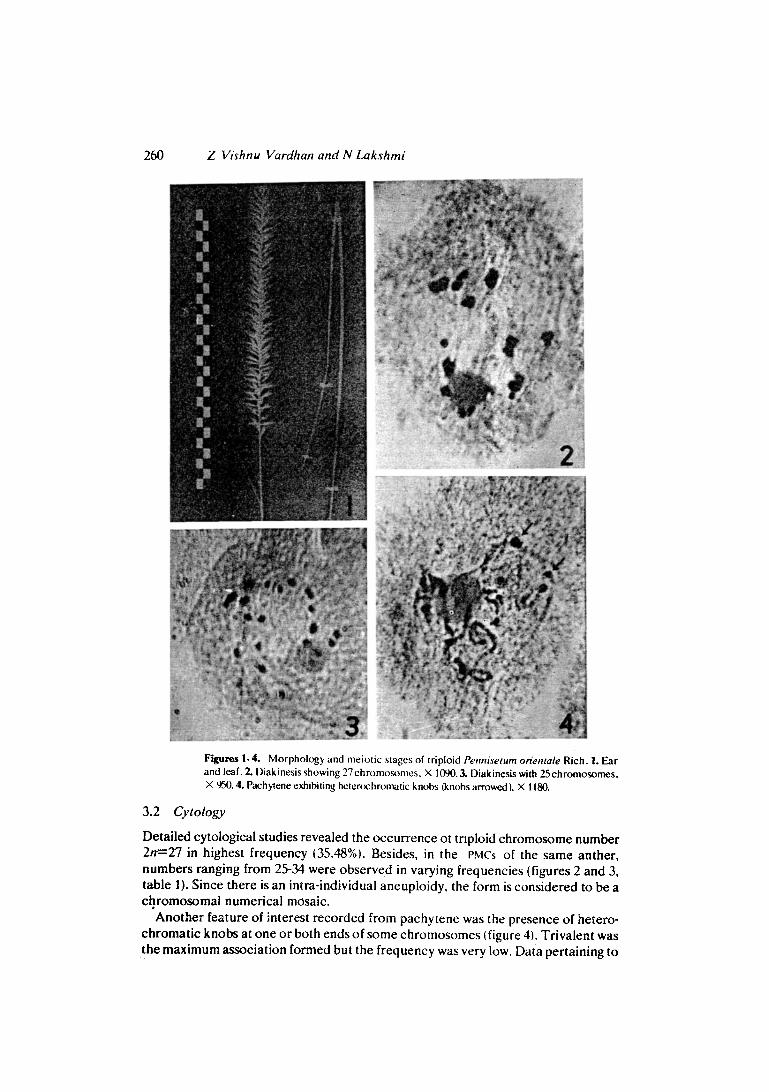
Figures 1.4. Morphology and meiotic stages of triploid Pennisetum otientale Rich. 1. Ear and leaf. 2. Diakinesis showing 27 chromo~mes. • 1090. 3. Diakinesis with 25 chromosomes. • 950. 4. Pachytene exhibiting hetertx:hromatic knobs (knobs arrowed ). X 1180.
3.2 Cytology
Detailed cytological studies revealed the occurrence ot tnploid chromosome number 2n=27 in highest frequency (35.48%). Besides, in the PMCs of the same anther, numbers ranging from 25-34 were observed in varying frequencies (figures 2 and 3, table 1). Since there is an intra-individuai aneuploidy, the form is considered to be a chromosomal numerical mosaic.
Another feature of interest recorded from pachytene was the presence of hetero- chromatic knobs at one or both ends of some chromosomes q 4). Trivalent was the maximum association formed but the frequency was very low. Data pertaining to

A new triploidP, orientale 261
Table !. Percentage of cells with different chromosome numbers and frequency of
trivalents, bivalents and univalents.
Chromosome No. number of cells Percent (PMCs) observed
23 3 4.84 24 I 1.61 25 2 3.23 26 3 4.83 27 22 35.48 28 8 12.9 29 2 3.23 30 14 22.58 32 2 3.23 33 3 4.84 34 2 3.23
Mean values tor trivalents, bivalents and univalents were 1.14, 9.7"2 and 4.35 respectively whil�91 the corresponding range values were 0-6, 5-15 and 1-10.
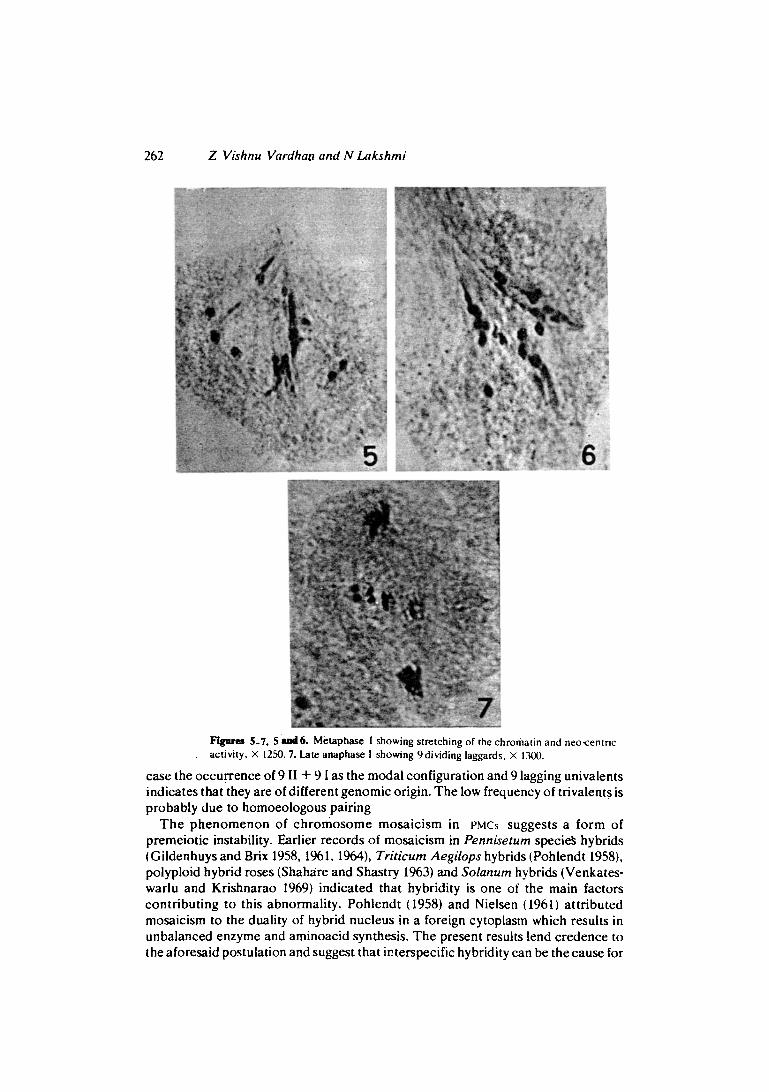
the frequeney of different ehromosome numbers and assoeiations are presented in rabie 1. Another interesting abnormal i ty eneoun te r ed in this material whieh contributes to meiotie drive and unasual segregations is neo-eentrie aetivity. This abnormality manifested itself at the metaphase I stage. Enormous stretehing of the ehromatin is seen at plaees other than the eentromere (figure 5). The bivalents elearly displayed eentrie aetivity in regions other than the eentromeres (figures 5 and 6). The strength of the neo-cent romere fur ther inereased at anaphase and it dominated the aetivity of the original eentromere. The original eentromere is seen to reo¡ and start eo-orientation with the neo-eentromeres of the same ehromosome. A s a result, preeoeious movement of some ehromosomes to poles was seen while others lagged (tab~.e 2). Sometimes this movement was seen towards more than two poles a s a result of eonstitution of multipolar spindle. Henee anaphase I was highly irregular with laggards and bridges (table 2).
In eells with 27 ehromosomes, 9 ehromosomes tend to lag in the hfiddle while the others move to poles (figure 7). The number of lagging ehromosomes was more in eells having high ehromosome numbers. The laggards either divide or move to one of the poles. Majority of the eells at anaphase I showed laggards (64.29%). Seeond divisions were also irregular. Pollen fertility was moderate (60.4%) and seeA set was very poor (4.95%).
4. Diseussion
Formation of tnvalents as the highest possible assoeiation and 27 as the frequent ehromosome number suggest that the taxon is at the triploid level. This apl~ears to be a new ehromosomal race, sineb its morphologieal eharaeteristies are signifieandy different from earlier reports. The meiotie behaviour of this raee indieates that ir is formed a s a result of hybridization between a tetraploid P. orientale a n d a diploid grass of unknown origin with base number x =9 . Levels of intergenomie pairing (multivalent formation) are often used to estimate whether a polyploid is of hybrid origin or not (Kihara and Orto 1926; Darlington 1932; Stebbins 1947). In the present
262 Z Vishnu Vardhal7 and N Lakshmi
�9 - ' ' , ' ~" ' : ~ ' : ! :7"+"'- ~ - ! " ~ , � 9 1 " . : ' , + ' " ~ . . . . . . . . . . . �9 . . . . . : " , ~ . - - - . .
. : . . . . . . . ..., ~ . . . , . ~ . . . . . . . . .- + t~ ~ " . . . . .,.~,~
i
~:;+c ~.dlp~~-.+2+_.,+'�91 + ~ + ~., ~,._,' .- .~ :,,Sl .,,+ +. ~ I Ÿ ~ ,,+: . +::,.+a I I ' I I I ~ ~ , + ~ r + + + + ~ I I + , 1 ] ~ L �9 ~'+., . +. . .Y ,~ ' , ' , : ." +-,,..,, ~ L +kr + �9 p + + , ~ , + . ~ ; + + , , . . t Ÿ . . ~ : . + �9 + . . ,,,+,~. + . , . ~ w ~ ~ ~ ~ ~ . . . , . . . . +++ +.-"+ ~++++~, ~ + ,..+.,,.~ ,
mr..++ ~ + ' + ~ ~ @ + + ~ i '.-+..,;+,~.,~.,~+~~+::.,k+.'+ ,.~kl, N i : , ~ ~ + + , , , + ,. ,+.~~+ .-+, , , : ++~~P,-,,,.,~+, :2 +'+r ~: �9
~+";',-~.~ ~~..:_~"~Jl..'l~-"++ '~+~,+,~.+.+~~,., . . . . . . . ~++~. ~ % . ~ ~ ~ . I = " + ~ ' ' " . . , - ' - ,. ~,.~_ ~. ~~,~ ~ , + + i + . ; . . - - , . ~ + , + , + +.' c +Ÿ +~+.+ ~ ' - + <, .+ . , ',,.,.,.e - ~ ~ < + ' ? ' + + +~+:+~ :+ :i:+.Ÿ + -:s.+~.+-++ : :0 +
5-7. 5 ~~16. Metaphase I showing stretching of the chromatin and neo-cenmc activity, X 1250. 7. Late anaphase ! showing 9dividing laggards, • 1300.
case the occurrence of 9 II + 9 1 as the modal configuration and 9 lagging univalents indicates that they are of different genomic origin. The low frequency of trivalent.s is probably due to homoeologous pairing
T h e p h e n o m e n o n of e h r o m o s o m e mosaicism in PMCs suggests a form of premeiotic instability. Earlier records of mosaicism in Pennisetum specieg hybrids IGildenhuys and Brix 1958, 1961, 1964), Triticum Aegilops hybrids (Pohlendt 1958), polyploid hybrid roses (Shahare and Shastry 1963) and Solanum hybrids (Venkates- warlu and Krishnarao 1969) indicated that hybridity is one of the main factors contributing to this abnormality. Pohlendt (1958) and Nielsen (1961) attr ibuted mosaicism to the duality of hybrid nucleus in a foreign cytoplasm which results in unbalanced enzyme and aminoacid synthesis. The present results lend credence to the aforesaid postulation and suggest that interspecific hybridity can be the cause for

A new triploid P. orienta& 263
TLble 2. Analysis of anaphase I.
Number Character of cells Percentage
observed
Cells without la,..:_. ,rds 30 35.71 1-10 laggards 23 27.38
More tha, 10 laggard~ 12 14.28 Cells with only chromatin bridges 17 20.24 Bridges and laggards 2 2.38
gene-controlled spindle abnorn~a lities in the premeiotic mitoses which are passed on to the progeny.
Neo-centric activity, although a rare feature, was recorded earlier in rye (Prakken and Muntzing 1942) and maize (Rhoades 1952). In maize usually chromosomes with he terochromat ic knobs show neo-centric activity in the presence of abnormal chromosome 10 having heterochromatic knob. In the present case although i t i s difficult to identify the individual chromosomes with knobs because of mosaicism, it can be suggested, based on the meiotic behaviour as well as earlier reports on neo- centric activity that the terminal heterochromatic knobs ac t a s sites of neo-cent¡ ac t iv i ty here also.
P o l y p l o i d y c o u p l e d with hybr id i za t ion p l a y e d a m a j o r ro le in the evo lu t ion of this species. A p o m i x i s is also r epor t ed in this spc ies (Narayan 1962; Chat ter j i and T i m o t h y 1969). Thus the abe r r an t meio t ic b e h a v i o u r resul t ing f rom interspecif ic hybr id iza t ion , mosa i c i sm and neo-cen t r i c act ivi ty may no t a e t a s a ba r r i e r for the survival of this species .
Acknowledgements
T h e au tho r s a re gra teful to ProI . A S Rao , for faci l i t ies and to Shri N B Rao, D e p a r t m e n t of Botany, V.S.R. Col lege , T e n a l i for t echn ica l a s i s t a n c e and sugges- t ions. One of the au tho r s (ZV) is t hankfu l to the UGC for a fel lowship.
References
Chatterji A K and Timothy D H 1969 Apomixis and tetraploidy in Penniserum orienta& Rich ; Crop Sci. 9 796-799
Darlington C D 1932 Recent advances in Cytology; (London : Churchill) p. 156 Gildenhuys P and Brix K 1958 Cytological abnormalities in Pennisetum dubium ;Heredity 12 441-452 Gildenhuys P and Brix K 1961 Genetic control of a•euploidy in Pennisetum; Heredity 16 358-363 Gildenhuys P and Brix K 1964 Genieally controiled variability of chromosome number in Pennisetum
hybrids; Heredity 19 533-542 H rishi N J 1952 Studies on the eytogenetics of six speeies of Pennisetum and their comparative morphology
and anatomy; Genetica 26 280-356 Kihara H and Ono T 1926 Chromosomenzahlcn und systematische Gruppierung der Rumex-Arten;
Zeitschri/t /uf Zell/orschung und mikroskopische Anatomie" 4 475-481 Krishnaswamy N 1941 Untersuchen zur Cytologie und Systematic der Gramineen; (B.B.C.) Beihe/te zum
Botanischen Centralblatt 611 1-56 Mehra P N and Remanandan P 19'73 Cytological investigations on W. Himalayan Panicoideae; Cytologia
38 25%270 Mehra P N, Khosla P K Kohli B L and Koonar J S 1968 Cytologieal studies in the North lndian Grasses
(Part !); Res. Bull. Punjab Unir. 19 (Part 1-II) 157-230

264 Z Vishnu Vardhan and N Lakshmi
Narayan N K 1962 Apomixis in some spr of Pennisetum and in Panicum antidotale: In Plant embryology. A xymposium: CSIR New Delhi 55-61
Nielsen E L 1961 Cytology and fertility of Agroelymus turneri; Cytologia 26 137-154 Parodi L R 1946 G ramineas bonaHenses. Cia ve para la determinacion de las gineros y enumeracion de las
especies, 4~h edn Patil B D. Hardas M W. O'Connor K Fand Vohra S K 1962 Polyploidy in Pennisetum orientale Rich; Curr.
Sci. 31 161-162 Pohlendt G 1958 Variabilitat der chromoso nenzahlen und andere kernpathologien in Aegilops trunciah~~
X Triticum aestivum; Basterden. Zeitschr. Vererbungsl. 89 170-188 Prakken R and Muntzing A 1942 A meiotic peculiarity in rye simulting a terminal centromere; Heredita.~
28441-481 Rhoades M M 1952 Preferential segregation in maize (ed) J W Gowen; Heterosis (Ames: Iowa State
College Press) 66-80 Shahare M L and Shastry S V S 1963 Meiosis in garden roses; Chromosoma 13 702-724 Sharma A K and De D N 1956 Cytology of some of the miUets; Caryologia $ 294-308 Sisodia K P S 1972 Cytomorphological studies in Pennisetum oHentale Rich; Cytologia 37 309-316 Stebbins G L 1947 Types of polyploids: Their classification and significance; Adv. Genet. i 403.429 Venkateswarlu J and Krishnarao M 1969 Chromosome numerical mosaicism in some hybrids of the
Solanum nigrum complex; Genetica 40 400-406