Embed Size (px)
Citation preview

Journal of Virological Methods 129 (2005) 47–55
A multiplex polymerase chain reaction method for reliable, sensitiveand simultaneous detection of multiple viruses in citrus trees
Avijit Roy ∗, Amer Fayad, G. Barthe, R.H. BrlanskyUniversity of Florida, IFAS, Citrus Research and Education Center, Plant Pathology,
700 Experiment Station Road, Lake Alfred, FL 33850-2299, USA
Received 4 March 2005; received in revised form 20 April 2005; accepted 9 May 2005Available online 13 June 2005
Abstract
A multiplex polymerase chain reaction (mPCR) assay was developed to detect six RNA and one DNA citrus virus:Citrus leaf rugosevirus (CLRV), Citrus psorosis virus (CPsV),Citrus tatter leaf virus (CTLV), Citrus tristeza virus (CTV), Citrus variegation virus (CVV),Citrus yellow mosaic virus (CYMV), and Indian citrus ringspot virus (ICRSV) from citrus plants. These seven viruses are classified in sixdifferent virus genera. Degenerate primers were designed based on the respective virus isolate sequence data available from the GenBank andw and DNAv l methods forc and werei ion of eachv ing multiplev©
K
1
Mpcvbo1bmica
NAhisood
on ofsys-threel.,and
de et-
tecteddts
0d
ere used for reliable detection of the different viruses by simplex- and mPCR. The sensitive and simultaneous detection of RNAiruses using the mPCR decreases the risk of contamination, saves time and reduces the cost as compared to other conventionaitrus virus detection. Seven different fragments (245–942 bp) specific to the viruses were simultaneously amplified using mPCRdentified on the basis of their molecular sizes. The consistent results of the mPCR were compared with simplex PCR for detectirus pathogen. The mPCR results were confirmed with sequencing analysis. The mPCR provides a useful rapid method for detectiruses in citrus plants that will aid in the production of virus-free citrus plants for certification programs.2005 Elsevier B.V. All rights reserved.
eywords: Citrus virus; Detection; Multiplex; PCR
. Introduction
Citrus is one of the most important fruit crops worldwide.ost of the citrus cultivars are grown as grafted plants. Virusathogens transmitted by grafting as well as insect vectorsan cause economic problems. For the detection of citrusiruses, enzyme-linked immunosorbent assay (ELISA) haseen the most commonly employed procedure used becausef its sensitivity, efficacy and economy (Garnsey and Cambra,991). However, many citrus viruses may be difficult to detectecause of their low titer or uneven distribution. The poly-erase chain reaction (PCR) is a powerful technique, which
s sensitive and highly specific and has been used to detectitrus viruses. Simplex PCR detects one target per reactionnd requires different PCRs to detect multiple targets. Multi-
∗ Corresponding author. Tel.: +1 863 956 1151; fax: +1 863 956 4631.E-mail address: [email protected] (A. Roy).
plex PCR (mPCR) is a useful technique since different Rand DNA viruses may infect a single host like citrus. Tsensitive technique would be very useful in citrus budwcertification programs.
There are several examples of simultaneous detectimultiple targets by mPCR amplification. Several mPCRtems have been developed for the detection of two orplant viruses (Minafra and Hadidi, 1994; Nemichov et a1995; Singh et al., 1996; Jacobi et al., 1998; GriecoGallitelli, 1999; Russo et al., 1999; James, 1999; Saaal., 2000; Sharman et al., 2000). There are only a few examples where more than three plant viruses have been deusing mPCR-based assay (Bariana et al., 1994; Nie anSingh, 2001; Nassuth et al., 2000). Recently, five differentospoviruses (Uga and Tsuda, 2005), five grapevine virusebelonging to two genera (Dovas and Katis, 2003), six RNAviruses from olive trees belonging to four genera (Bertoliniet al., 2001), six citrus viroids and one citrus virus (Ito et al.,
166-0934/$ – see front matter © 2005 Elsevier B.V. All rights reserved.oi:10.1016/j.jviromet.2005.05.008

48 A. Roy et al. / Journal of Virological Methods 129 (2005) 47–55
2002) and seven viroids from four genera (Ragozzino et al.,2004) have been detected using single tube multiplex reversetranscription (RT) PCR reaction.
In this work, we describe the simultaneous detection ofthe seven characterized citrus viruses, which belong to sixdifferent genera:Badnavirus [Citrus yellow mosaic virus(CYMV)], Capillovirus [Citrus tatter leaf virus (CTLV)],Closterovirus [Citrus tristeza virus (CTV)], Ilarvirus [Cit-rus leaf rugose virus (CLRV) and Citrus variegationvirus (CVV)], Mandarivirus [Indian citrus ringspot virus(ICRSV)] andOphiovirus [Citrus psorosis virus (CPsV)].
CLRV and CVV are ilar viruses that affect citrus. Symp-tom differences in red kidney bush bean can be used todistinguish CLRV from CVV. ELISA, nucleic acid hybridiza-tion, and RT-PCR have been developed for CLRV (Ge andScott, 1994; Scott and Ge, 1995) and CVV detection (Davinoand Garnsey, 1984; Bennani et al., 2002).
CPsV is a multicomponent, negative-sense single-stranded RNA (ssRNA) virus. The viral genome is encap-sidated in short and long particles. The virus is a member ofOphiovirus, is graft-transmitted andChenopodium quinoa, isused as a local lesion host for detection. Serological detec-tion of CPsV by ELISA and direct tissue-blot immunoassay(DTBIA) (Martin et al., 2002), and molecular detection byhybridization and RT-PCR (Barthe et al., 1998) has beendeveloped.
i witha hodsi( thEa
e,s tiv-i iono nm olog-i says(i rd eta l.,2
db ef Vg of6 Ah MVd
dia( us6 bout7 buto and
allexi viruses (Rustici et al., 2002). Taxonomically, ICRSVis currently the only member of the new genusMandarivirus(Adams et al., 2004).
In this paper, primer pairs including degenerate primerswere designed using a computer program after aligning allthe GenBank sequences of a particular virus. This allowed thedevelopment of a mPCR assay and facilitated the sequenceanalysis. This included a set of seven primer pairs for two-step PCR amplification enabling the simultaneous detectionof all the seven citrus viruses. To our knowledge, this is thefirst report on detection of seven plant viruses using simul-taneously mPCR. It also demonstrates the feasibility of thismPCR approach for use in diagnostic and certification pro-grams.
2. Materials and methods
2.1. Virus isolates
The citrus viruses used in this study were CLRV, CPsV,CTLV, CTV, CVV, CYMV and ICRSV that are describedabove. Except for CYMV and ICRSV, the other five viruseswere collected from the different citrus orchards and nurs-eries of Florida, USA and maintained in the greenhouse eitherin Mexican lime or Madam Vinous sweet orange seedlings.I oft A.F urceo forI orC ion,B
2
duret ent( andD tedc mgo totalR N,V TheD toi usp
2
andm oatp V;( 8g iono rch
CTLV is the type species of the genusCapillovirus. CTLVs a flexuous filamentous particle, 600–700 nm long
ssRNA genome of 6500 nt. Current detection metnclude biological indexing on Rusk citrange (Citrus sinensisL.) Osb.× Poncirus trifoliata), serological diagnosis wiLISA, and molecular diagnosis with RT-PCR (Miyakawand Ito, 2000).
CTV is a member ofClosterovirus genus, monopartitsRNA virus of approximately 19.3 kb. The biological acty of a CTV isolate is usually determined by inoculatf indicator hosts (Garnsey et al., 1987). Various detectioethods have been used for CTV. These include ser
cal methods such as ELISA, dot-immunobinding asDIBA) (Permar et al., 1990; Rocha-Pena et al., 1991) andmmunofluorescence (Brlansky et al., 1988) and moleculaiagnosis like cDNA probe (Rosner et al., 1986; Narvaezl., 2000) and RT-PCR (Mehta et al., 1997; Huang et a004).
CYMV is a member ofBadnavirus with nonenvelopeacilliform virions, measuring 130 nm× 30 nm that ar
ound in the cytoplasm of infected plants. The CYMenome is a circular, double-stranded DNA molecule.6 kb. ELISA, immuno specific microscopy, DNA–DNybridization and PCR have been developed for CYetection (Ahlawat et al., 1996).
ICRSV is a serious disease of Kinnow mandarin in InRustici et al., 2000). The virions are flexuous filamento50 nm particles in length, with an ssRNA genome of a500 nt. It is similar in some aspects of potex virusesverall genome organization of ICRSV is similar to carla
CRSV and CYMV were reported from India are parthe Exotic Citrus Disease Collection, Beltsville, MD, USlorida isolates 6b1, CVV-1 and CLRV-1 were used as sof CPsV, CVV and CLRV, respectively. The isolates B168
CRSV, B178 for CYMV, B153 for CTLV, B165 and B192 fTV were collected from Exotic Citrus Disease Collecteltsville and used as source isolates for this study.
.2. Nucleic acid extraction
Two methods were evaluated to select a reliable proceo prepare virus RNA and DNA targets: (1) TRIzol reagInvitrogen, Cat. No. 15596-026) was used for total RNANA extraction from 100 mg of healthy and virus infecitrus tissue following the manufacture’s protocol. (2) 100f virus-infected and healthy citrus tissue was used forNA extraction using the RNeasy plant mini kit (QIAGEalencia, CA) according to the manufacturer’s protocol.Neasy plant mini kit (QIAGEN, Valencia, CA) was used
solate total DNA from healthy and CYMV infected citrlants.
.3. Primer design
Seven sets of primers were designed for simplexPCR amplification of: (i) conserved areas within the crotein (CP) gene of CPsV, CTLV, CVV, and ICRSii) the partial polyprotein gene of CYMV; (iii) the p1ene of CTV; and (iv) the gene encoding partial regf RNA-3 of CLRV. The Nucleotide Sequence Sea
A. Roy et al. / Journal of Virological Methods 129 (2005) 47–55 49
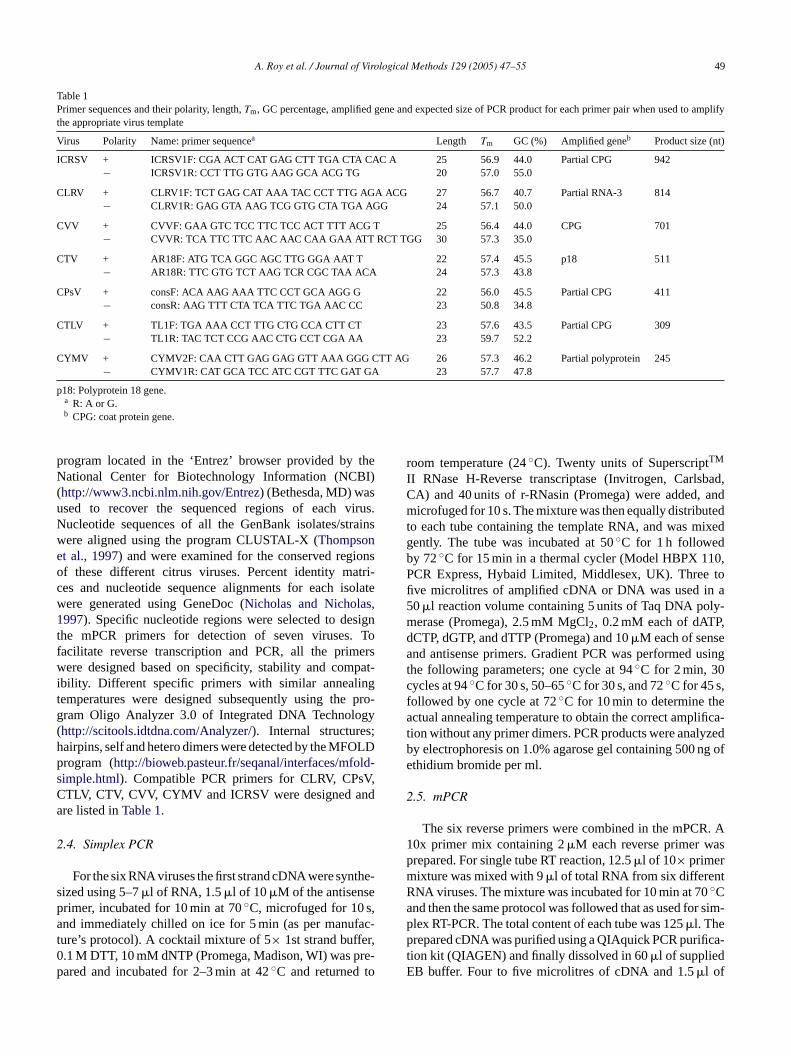
Table 1Primer sequences and their polarity, length,Tm, GC percentage, amplified gene and expected size of PCR product for each primer pair when used to amplifythe appropriate virus template
Virus Polarity Name: primer sequencea Length Tm GC (%) Amplified geneb Product size (nt)
ICRSV + ICRSV1F: CGA ACT CAT GAG CTT TGA CTA CAC A 25 56.9 44.0 Partial CPG 942− ICRSV1R: CCT TTG GTG AAG GCA ACG TG 20 57.0 55.0
CLRV + CLRV1F: TCT GAG CAT AAA TAC CCT TTG AGA ACG 27 56.7 40.7 Partial RNA-3 814− CLRV1R: GAG GTA AAG TCG GTG CTA TGA AGG 24 57.1 50.0
CVV + CVVF: GAA GTC TCC TTC TCC ACT TTT ACG T 25 56.4 44.0 CPG 701− CVVR: TCA TTC TTC AAC AAC CAA GAA ATT RCT TGG 30 57.3 35.0
CTV + AR18F: ATG TCA GGC AGC TTG GGA AAT T 22 57.4 45.5 p18 511− AR18R: TTC GTG TCT AAG TCR CGC TAA ACA 24 57.3 43.8
CPsV + consF: ACA AAG AAA TTC CCT GCA AGG G 22 56.0 45.5 Partial CPG 411− consR: AAG TTT CTA TCA TTC TGA AAC CC 23 50.8 34.8
CTLV + TL1F: TGA AAA CCT TTG CTG CCA CTT CT 23 57.6 43.5 Partial CPG 309− TL1R: TAC TCT CCG AAC CTG CCT CGA AA 23 59.7 52.2
CYMV + CYMV2F: CAA CTT GAG GAG GTT AAA GGG CTT AG 26 57.3 46.2 Partial polyprotein 245− CYMV1R: CAT GCA TCC ATC CGT TTC GAT GA 23 57.7 47.8
p18: Polyprotein 18 gene.a R: A or G.b CPG: coat protein gene.
program located in the ‘Entrez’ browser provided by theNational Center for Biotechnology Information (NCBI)(http://www3.ncbi.nlm.nih.gov/Entrez) (Bethesda, MD) wasused to recover the sequenced regions of each virus.Nucleotide sequences of all the GenBank isolates/strainswere aligned using the program CLUSTAL-X (Thompsonet al., 1997) and were examined for the conserved regionsof these different citrus viruses. Percent identity matri-ces and nucleotide sequence alignments for each isolatewere generated using GeneDoc (Nicholas and Nicholas,1997). Specific nucleotide regions were selected to designthe mPCR primers for detection of seven viruses. Tofacilitate reverse transcription and PCR, all the primerswere designed based on specificity, stability and compat-ibility. Different specific primers with similar annealingtemperatures were designed subsequently using the pro-gram Oligo Analyzer 3.0 of Integrated DNA Technology(http://scitools.idtdna.com/Analyzer/). Internal structures;hairpins, self and hetero dimers were detected by the MFOLDprogram (http://bioweb.pasteur.fr/seqanal/interfaces/mfold-simple.html). Compatible PCR primers for CLRV, CPsV,CTLV, CTV, CVV, CYMV and ICRSV were designed andare listed inTable 1.
2.4. Simplex PCR
the-s ep ,a fac-t r,0 re-p o
room temperature (24◦C). Twenty units of SuperscriptTM
II RNase H-Reverse transcriptase (Invitrogen, Carlsbad,CA) and 40 units of r-RNasin (Promega) were added, andmicrofuged for 10 s. The mixture was then equally distributedto each tube containing the template RNA, and was mixedgently. The tube was incubated at 50◦C for 1 h followedby 72◦C for 15 min in a thermal cycler (Model HBPX 110,PCR Express, Hybaid Limited, Middlesex, UK). Three tofive microlitres of amplified cDNA or DNA was used in a50�l reaction volume containing 5 units of Taq DNA poly-merase (Promega), 2.5 mM MgCl2, 0.2 mM each of dATP,dCTP, dGTP, and dTTP (Promega) and 10�M each of senseand antisense primers. Gradient PCR was performed usingthe following parameters; one cycle at 94◦C for 2 min, 30cycles at 94◦C for 30 s, 50–65◦C for 30 s, and 72◦C for 45 s,followed by one cycle at 72◦C for 10 min to determine theactual annealing temperature to obtain the correct amplifica-tion without any primer dimers. PCR products were analyzedby electrophoresis on 1.0% agarose gel containing 500 ng ofethidium bromide per ml.
2.5. mPCR
The six reverse primers were combined in the mPCR. A10x primer mix containing 2�M each reverse primer wasprepared. For single tube RT reaction, 12.5�l of 10× primerm tRa sim-pp ca-tE
For the six RNA viruses the first strand cDNA were synized using 5–7�l of RNA, 1.5�l of 10�M of the antisensrimer, incubated for 10 min at 70◦C, microfuged for 10 snd immediately chilled on ice for 5 min (as per manu
ure’s protocol). A cocktail mixture of 5× 1st strand buffe.1 M DTT, 10 mM dNTP (Promega, Madison, WI) was pared and incubated for 2–3 min at 42◦C and returned t
ixture was mixed with 9�l of total RNA from six differenNA viruses. The mixture was incubated for 10 min at 70◦Cnd then the same protocol was followed that as used forlex RT-PCR. The total content of each tube was 125�l. Therepared cDNA was purified using a QIAquick PCR purifi
ion kit (QIAGEN) and finally dissolved in 60�l of suppliedB buffer. Four to five microlitres of cDNA and 1.5�l of

50 A. Roy et al. / Journal of Virological Methods 129 (2005) 47–55
CYMV DNA were added in a 50�l PCR reaction mixture.QIAGEN multiplex PCR kit (Cat. No. 206143) was usedfor mPCR amplification. The mPCR mix contained 25�l ofmPCR master mix, 5�l of 10× primer mix (2�M of eachprimer) and the final volume made up with distilled water.Gradient PCR was performed using the following parame-ters; one cycle at 94◦C for 15 min as heating step of HotStarTaq DNA polymerase, 35 cycles at 94◦C for 30 s, 55–63◦Cfor 90 s, and 72◦C for 90 s, followed by one cycle at 72◦C for10 min to determine the annealing temperature for all sevenviruses amplifications. PCR products were analyzed by elec-trophoresis on a 1.4% agarose gel.
2.6. Sensitivity testing of the simplex and mPCR
Total RNA and DNA preparations from viral infectedplants were adjusted to 200 ng/�l. To compare the sensitiv-ity level of mPCR and simplex PCR, 10-fold serial dilutions(10−1 to 10−6) in nuclease-free water containing the six viralcDNA and one DNA from CYMV infected sample weretested by simplex and mPCR. The simplex and mPCR werecarried out simultaneously for the comparison of the dilutionseries.
2.7. Gel extraction and sequence analysis
singa elp PCRa thei uri-fi nds nta-t
2n
D,U ens.S itrusv oret CRs ates;F ),B Va edi
3
3m
alityD the
DNeasy and RNeasy kits produced more consistent amplifi-cation products as compared to the TRIzol reagent.
3.2. Design of primer pairs for the detection of six RNAand one DNA virus by mPCR
The published primer sets for the detection of CLRV (Geand Scott, 1994), CPsV (Barthe et al., 1998), CTLV (Ito etal., 2002), CTV (Huang et al., 2004), CVV (Bennani et al.,2002), CYMV (Ahlawat et al., 1996) and ICRSV (Rustici etal., 2000, 2002) were not adequate for use with the mPCRtechnique because of primer interference and similar productsizes that overlapped and hindered accurate detection (datanot shown). New primers were designed after alignment ofconserved areas for the seven citrus viruses. The designedprimer pairs are listed inTable 1and displayed similar meltingtemperatures, G + C contents and nucleotide length. Threepairs of primers for each virus were designed and of the three,only one primer pair produced a single band for each virusin simplex as well as in mPCR amplification. This primer setwas used for further detection using mPCR technique.Table 1includes a list of these best suited designed oligonucleotideprimers.
3.3. Optimization of simplex and mPCR
izesw m-p 2 bpf rC forC lifi-c lants(
lifi-c ndM atedt int wn)a ck ofe iruscR theb
getsw -e PCRr omC -e ersepa ) andu andH . Fors -i t the
After electrophoresis, the gel products were purified upurification kit from QIAGEN (Cat. No. 28704). G
urified products were used as templates for furthermplification using virus-specific primer pairs to confirm
dentity of each virus. The PCR products were again ped by QIAquick PCR purification kit (Cat. No. 28104) aequenced using virus-specific primer pairs in both orieions.
.8. Detection of more than one citrus virus fromaturally infected samples by mPCR
The Exotic Citrus Disease Collection at Beltsville, MSA, maintains different kinds of citrus systemic pathogome of plants are infected with one or more than one cirus. Some of the isolates that are known to contain mhan one type of citrus virus were tested using our mPystem. The tested isolates include: two Florida field isolL-1 (CLRV and CVV) and FL-2 (CLRV, CVV and CTV384 and B396 (for CTV and CTLV), B74 and B360 (CTnd CPsV), B172 (ICRSV, CPsV and CTV) to verify mix
nfections.
. Results
.1. Comparison between the DNeasy, RNeasy plantini kit and TRIzol reagent nucleic acid extractions
Both the methods produced a good yield of high quNA and RNA. For mPCR reaction, nucleic acids from
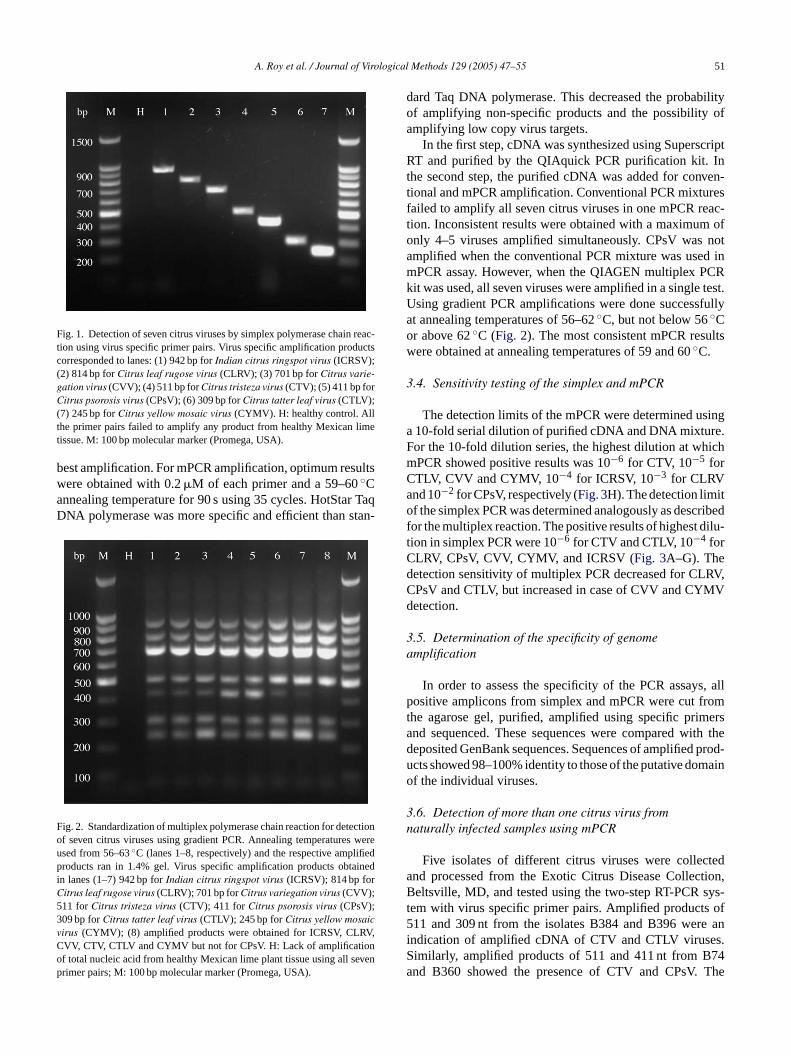
Specific PCR amplification products of the expected sere obtained from all positive RNA and DNA virus sales. Seven amplified products of expected sizes (94
or ICRSV; 814 bp for CLRV; 701 bp for CVV; 511 bp foTV; 411 bp for CPsV; 309 bp for CTLV; and 245 bpYMV) were observed only in infected samples. No ampation products were obtained from extracts of healthy pFig. 1).
During cDNA synthesis of simplex and mPCR ampation, SuperscriptTM II RNase H-Reverse transcriptase a-MLV RT were used and compared. The results indic
hat Superscript RT was more efficient than M-MLV RThe simplex as well as in mPCR reaction (data not shond worked better in the mPCR reactions due to the landonuclease activity and a lower RNase H activity. Voncentration, temperature of RT step (42◦C for M-MLVT and 50◦C for Superscript), and salt concentration ofuffer were also considered for mPCR amplification.
In the simplex and mPCR, single and mixed tarere detected successfully (Figs. 1 and 2). Various paramters were considered to optimize the simplex and meactions for the individual and mixed RNA targets frLRV, CPsV, CTLV, CTV, CVV, CYMV and ICRSV. Paramters such as the concentration of forward and revrimers (0.002–2�M), annealing temperature (50–65◦C)nd annealing time (30–90 s), number of cycles (25–45se of two kinds of Taq DNA polymerases (standardotStar Taq) were evaluated using virus infected plantsimplex PCR, 0.4�M of each primer and a 58◦C anneal
ng temperature for 30 s and 30 cycles was used to ge
A. Roy et al. / Journal of Virological Methods 129 (2005) 47–55 51
Fig. 1. Detection of seven citrus viruses by simplex polymerase chain reac-tion using virus specific primer pairs. Virus specific amplification productscorresponded to lanes: (1) 942 bp forIndian citrus ringspot virus (ICRSV);(2) 814 bp forCitrus leaf rugose virus (CLRV); (3) 701 bp forCitrus varie-gation virus (CVV); (4) 511 bp forCitrus tristeza virus (CTV); (5) 411 bp forCitrus psorosis virus (CPsV); (6) 309 bp forCitrus tatter leaf virus (CTLV);(7) 245 bp forCitrus yellow mosaic virus (CYMV). H: healthy control. Allthe primer pairs failed to amplify any product from healthy Mexican limetissue. M: 100 bp molecular marker (Promega, USA).
best amplification. For mPCR amplification, optimum resultswere obtained with 0.2�M of each primer and a 59–60◦Cannealing temperature for 90 s using 35 cycles. HotStar TaqDNA polymerase was more specific and efficient than stan-
Fig. 2. Standardization of multiplex polymerase chain reaction for detectionof seven citrus viruses using gradient PCR. Annealing temperatures wereused from 56–63◦C (lanes 1–8, respectively) and the respective amplifiedproducts ran in 1.4% gel. Virus specific amplification products obtainedi rC53v V,Cop
dard Taq DNA polymerase. This decreased the probabilityof amplifying non-specific products and the possibility ofamplifying low copy virus targets.
In the first step, cDNA was synthesized using SuperscriptRT and purified by the QIAquick PCR purification kit. Inthe second step, the purified cDNA was added for conven-tional and mPCR amplification. Conventional PCR mixturesfailed to amplify all seven citrus viruses in one mPCR reac-tion. Inconsistent results were obtained with a maximum ofonly 4–5 viruses amplified simultaneously. CPsV was notamplified when the conventional PCR mixture was used inmPCR assay. However, when the QIAGEN multiplex PCRkit was used, all seven viruses were amplified in a single test.Using gradient PCR amplifications were done successfullyat annealing temperatures of 56–62◦C, but not below 56◦Cor above 62◦C (Fig. 2). The most consistent mPCR resultswere obtained at annealing temperatures of 59 and 60◦C.
3.4. Sensitivity testing of the simplex and mPCR
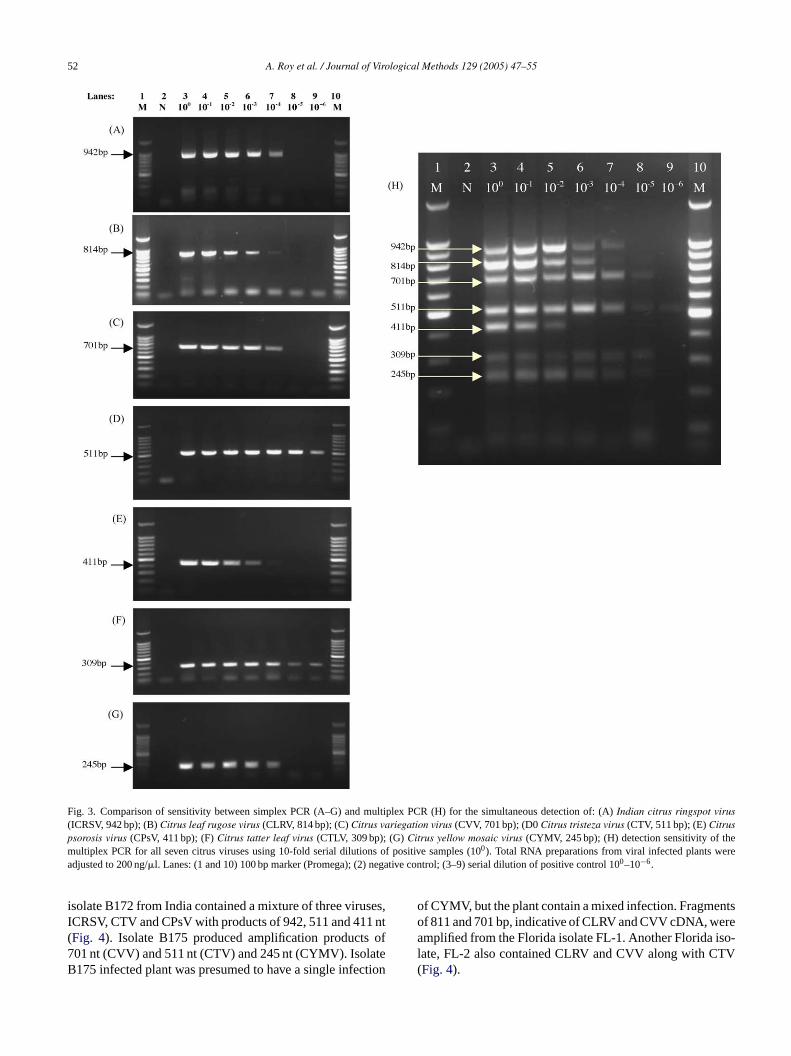
The detection limits of the mPCR were determined usinga 10-fold serial dilution of purified cDNA and DNA mixture.For the 10-fold dilution series, the highest dilution at whichmPCR showed positive results was 10−6 for CTV, 10−5 forCTLV, CVV and CYMV, 10−4 for ICRSV, 10−3 for CLRVand 10−2 for CPsV, respectively (Fig. 3H). The detection limito ribedf ilu-tCd RV,C MVd
3a
s, allp romt ersa ith thed prod-u maino
3n
teda tion,B sys-t of5 e an
n lanes (1–7) 942 bp forIndian citrus ringspot virus (ICRSV); 814 bp foitrus leaf rugose virus (CLRV); 701 bp forCitrus variegation virus (CVV);11 forCitrus tristeza virus (CTV); 411 forCitrus psorosis virus (CPsV);09 bp forCitrus tatter leaf virus (CTLV); 245 bp forCitrus yellow mosaicirus (CYMV); (8) amplified products were obtained for ICRSV, CLR
VV, CTV, CTLV and CYMV but not for CPsV. H: Lack of amplificationf total nucleic acid from healthy Mexican lime plant tissue using all sevenrimer pairs; M: 100 bp molecular marker (Promega, USA).i s.S 74a The
f the simplex PCR was determined analogously as descor the multiplex reaction. The positive results of highest dion in simplex PCR were 10−6 for CTV and CTLV, 10−4 forLRV, CPsV, CVV, CYMV, and ICRSV (Fig. 3A–G). Theetection sensitivity of multiplex PCR decreased for CLPsV and CTLV, but increased in case of CVV and CYetection.
.5. Determination of the specificity of genomemplification
In order to assess the specificity of the PCR assayositive amplicons from simplex and mPCR were cut f
he agarose gel, purified, amplified using specific primnd sequenced. These sequences were compared weposited GenBank sequences. Sequences of amplifiedcts showed 98–100% identity to those of the putative dof the individual viruses.
.6. Detection of more than one citrus virus fromaturally infected samples using mPCR
Five isolates of different citrus viruses were collecnd processed from the Exotic Citrus Disease Colleceltsville, MD, and tested using the two-step RT-PCR
em with virus specific primer pairs. Amplified products11 and 309 nt from the isolates B384 and B396 wer
ndication of amplified cDNA of CTV and CTLV viruseimilarly, amplified products of 511 and 411 nt from Bnd B360 showed the presence of CTV and CPsV.
52 A. Roy et al. / Journal of Virological Methods 129 (2005) 47–55
Fig. 3. Comparison of sensitivity between simplex PCR (A–G) and multiplex PCR (H) for the simultaneous detection of: (A)Indian citrus ringspot virus(ICRSV, 942 bp); (B)Citrus leaf rugose virus (CLRV, 814 bp); (C)Citrus variegation virus (CVV, 701 bp); (D0Citrus tristeza virus (CTV, 511 bp); (E)Citruspsorosis virus (CPsV, 411 bp); (F)Citrus tatter leaf virus (CTLV, 309 bp); (G)Citrus yellow mosaic virus (CYMV, 245 bp); (H) detection sensitivity of themultiplex PCR for all seven citrus viruses using 10-fold serial dilutions of positive samples (100). Total RNA preparations from viral infected plants wereadjusted to 200 ng/�l. Lanes: (1 and 10) 100 bp marker (Promega); (2) negative control; (3–9) serial dilution of positive control 100–10−6.
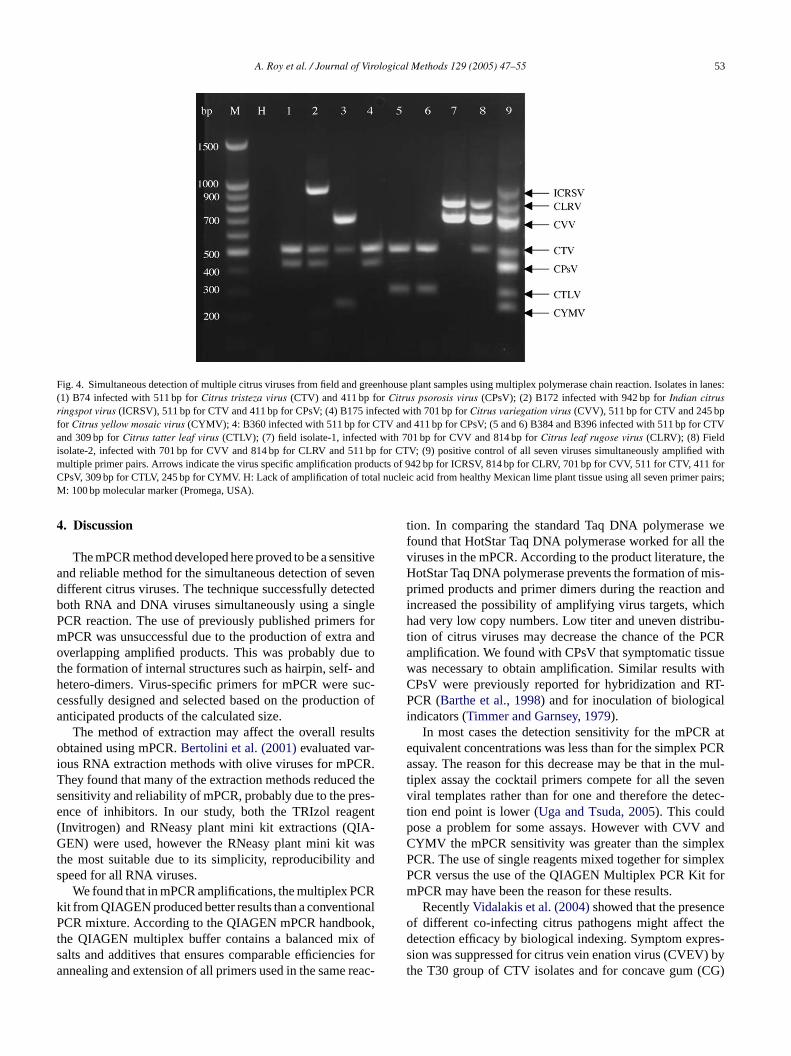
isolate B172 from India contained a mixture of three viruses,ICRSV, CTV and CPsV with products of 942, 511 and 411 nt(Fig. 4). Isolate B175 produced amplification products of701 nt (CVV) and 511 nt (CTV) and 245 nt (CYMV). IsolateB175 infected plant was presumed to have a single infection
of CYMV, but the plant contain a mixed infection. Fragmentsof 811 and 701 bp, indicative of CLRV and CVV cDNA, wereamplified from the Florida isolate FL-1. Another Florida iso-late, FL-2 also contained CLRV and CVV along with CTV(Fig. 4).
A. Roy et al. / Journal of Virological Methods 129 (2005) 47–55 53
Fig. 4. Simultaneous detection of multiple citrus viruses from field and greenhouse plant samples using multiplex polymerase chain reaction. Isolates in lanes:(1) B74 infected with 511 bp forCitrus tristeza virus (CTV) and 411 bp forCitrus psorosis virus (CPsV); (2) B172 infected with 942 bp forIndian citrusringspot virus (ICRSV), 511 bp for CTV and 411 bp for CPsV; (4) B175 infected with 701 bp forCitrus variegation virus (CVV), 511 bp for CTV and 245 bpfor Citrus yellow mosaic virus (CYMV); 4: B360 infected with 511 bp for CTV and 411 bp for CPsV; (5 and 6) B384 and B396 infected with 511 bp for CTVand 309 bp forCitrus tatter leaf virus (CTLV); (7) field isolate-1, infected with 701 bp for CVV and 814 bp forCitrus leaf rugose virus (CLRV); (8) Fieldisolate-2, infected with 701 bp for CVV and 814 bp for CLRV and 511 bp for CTV; (9) positive control of all seven viruses simultaneously amplified withmultiple primer pairs. Arrows indicate the virus specific amplification products of 942 bp for ICRSV, 814 bp for CLRV, 701 bp for CVV, 511 for CTV, 411 forCPsV, 309 bp for CTLV, 245 bp for CYMV. H: Lack of amplification of total nucleic acid from healthy Mexican lime plant tissue using all seven primer pairs;M: 100 bp molecular marker (Promega, USA).
4. Discussion
The mPCR method developed here proved to be a sensitiveand reliable method for the simultaneous detection of sevendifferent citrus viruses. The technique successfully detectedboth RNA and DNA viruses simultaneously using a singlePCR reaction. The use of previously published primers formPCR was unsuccessful due to the production of extra andoverlapping amplified products. This was probably due tothe formation of internal structures such as hairpin, self- andhetero-dimers. Virus-specific primers for mPCR were suc-cessfully designed and selected based on the production ofanticipated products of the calculated size.
The method of extraction may affect the overall resultsobtained using mPCR.Bertolini et al. (2001)evaluated var-ious RNA extraction methods with olive viruses for mPCR.They found that many of the extraction methods reduced thesensitivity and reliability of mPCR, probably due to the pres-ence of inhibitors. In our study, both the TRIzol reagent(Invitrogen) and RNeasy plant mini kit extractions (QIA-GEN) were used, however the RNeasy plant mini kit wasthe most suitable due to its simplicity, reproducibility andspeed for all RNA viruses.
We found that in mPCR amplifications, the multiplex PCRkit from QIAGEN produced better results than a conventionalPCR mixture. According to the QIAGEN mPCR handbook,t ofs s fora reac-
tion. In comparing the standard Taq DNA polymerase wefound that HotStar Taq DNA polymerase worked for all theviruses in the mPCR. According to the product literature, theHotStar Taq DNA polymerase prevents the formation of mis-primed products and primer dimers during the reaction andincreased the possibility of amplifying virus targets, whichhad very low copy numbers. Low titer and uneven distribu-tion of citrus viruses may decrease the chance of the PCRamplification. We found with CPsV that symptomatic tissuewas necessary to obtain amplification. Similar results withCPsV were previously reported for hybridization and RT-PCR (Barthe et al., 1998) and for inoculation of biologicalindicators (Timmer and Garnsey, 1979).
In most cases the detection sensitivity for the mPCR atequivalent concentrations was less than for the simplex PCRassay. The reason for this decrease may be that in the mul-tiplex assay the cocktail primers compete for all the sevenviral templates rather than for one and therefore the detec-tion end point is lower (Uga and Tsuda, 2005). This couldpose a problem for some assays. However with CVV andCYMV the mPCR sensitivity was greater than the simplexPCR. The use of single reagents mixed together for simplexPCR versus the use of the QIAGEN Multiplex PCR Kit formPCR may have been the reason for these results.
RecentlyVidalakis et al. (2004)showed that the presenceof different co-infecting citrus pathogens might affect thed res-s ) byt G)
he QIAGEN multiplex buffer contains a balanced mixalts and additives that ensures comparable efficienciennealing and extension of all primers used in the same
etection efficacy by biological indexing. Symptom expion was suppressed for citrus vein enation virus (CVEVhe T30 group of CTV isolates and for concave gum (C

54 A. Roy et al. / Journal of Virological Methods 129 (2005) 47–55
by CPsV in two citrus hosts. The reason for this symptomsuppression of one pathogen by another is unknown. Thelack of alternative diagnostic protocols for both CVEV andCG further complicates the understanding of the suppressionmechanisms. Titer or distribution of some viruses or virus-like pathogens may be affected in co-infected plants and thusdecrease the efficacy of molecular detection procedures suchas mPCR.Vidalakis et al. (2004)further pointed out that theseexperimental mixed infections may not simulate naturallyoccurring infections where equilibriums between pathogensmay be established and which could differ from recent infec-tions. The use of the mPCR system that we have describedneeds testing with naturally infected materials in conjunctionwith bioindexing methods.
In conclusion, the mPCR assay developed here is a simple,reliable, rapid, sensitive, specific and cost-effective diagnos-tic for multiple citrus viruses. It was successfully used forsimultaneously detecting seven viruses as well as multiplevirus infections in the same plant. It should be useful in con-junction with other diagnostic assays to determine unknownvirus infections and in certification and plant improvementprograms for obtaining virus free planting materials. As newcitrus viruses are characterized and primers are developed,they can be added to this method using the steps outlined inthis work. This method also should be applicable for detec-tion of other plant viruses and where confirmation of otherm
A
Ms.C orp theE ro-v theirl tancef Cit-r loridaA ica-t
R
A e, T.,Thecri-
A orty,saic
B 1994.tiplex.
B truse and537.
Bennani, B., Mendes, C., Zemzami, M., Azeddoug, H., Nolasco, G.,2002. Citrus variegation virus: molecular variability of apportion ofthe RNA-3 containging the coat protein gene and design of the primersfor RT-PCR detection. Eur. J. Plant Path. 108, 155–162.
Bertolini, E., Olmos, A., Carmen Martıı̌nez, M., Marı̌ıa Teresa, G., Cam-bra, M., 2001. Single-step multiplex RT-PCR for simultaneous andcolourimetric detection of six RNA viruses in olive trees. J. Virol.Meth. 96, 33–41.
Brlansky, R.H., Lee, R.F., Garnsey, S.M., 1988. In situ immunofluores-cence for the detection ofCitrus tristeza virus inclusion bodies. PlantDis. 72, 1039–1041.
Davino, M., Garnsey, S. M., 1984. Purification, characterization and serol-ogy of a mild strain of citrus variegation virus from florida. In:Garnsey, S.M., Timmer, L.W., Dodds, J.A. (Eds.), Proceedings ofthe Ninth Conference on International Organization Citrus Virology.IOCV Riverside, CA, pp. 196–203.
Dovas, C.I., Katis, N.I., 2003. A spot nested RT-PCR method for thesimultaneous detection of members of the Vitivirus andFoveavirusgenera in grapevine. J. Virol. Meth. 170, 99–106.
Garnsey, S. M., Cambra, M., 1991. Enzyme-linked immunosorbent assay(ELISA) for citrus pathogens. In: Roistacher, C.N. (Ed.), Graft Trans-missible Diseases of Citrus. Handbook for Detection and Diagnosis.FAO, Rome, pp. 193–216.
Garnsey, S.M., Gumpf, D.J., Roistacher, C.N., Civerolo, E.L., Lee, R.F.,Yokomi, R.K., Bar-Joseph, M., 1987. Toward the standardized evalua-tion of the biological properties of citrus tristeza virus. Phytophylatica19, 151–158.
Ge, X., Scott, S.W., 1994. The nucleotide sequence of citrus leaf rugoseilarvirus RNA-2. J. Gen. Virol. 75, 2841–2846.
Grieco, F., Gallitelli, D., 1999. Multiplex reverse transcriptase chain reac-tion applied to virus detection in globe artichoke. J. Phytopathol. 147,
H andst Dis.
I trusplex106,
J elop-and
Virol.
J n ofssay.
M tionsayand
M poly-.
M irusnd
M .W.,ases,MN,
N no,latesol.
N 000.ulta-
ethods is needed.
cknowledgements
The authors are grateful to Dr. John Hartung andristina Paul, USDA ARS, Fruit Lab., Beltsville, MD, froviding the CYMV and ICRSV infected sources fromxotic Citrus Disease Collection, Beltsville, MD, and for piding laboratory assistance in nucleic acid extraction inaboratory. The authors acknowledge the financial assisrom the USDA Specific Cooperative Agreement Exoticus Disease Grant. This research was supported by the Fgricultural Experiment Station, and approved for publ
ion as Journal Series No. R-10765.
eferences
dams, M.J., Antoniw, J.F., Bar-Joseph, M., Brunt, A.A., CandressFoster, G.D., Martelli, G.P., Milne, R.G., Fauquet, C.M., 2004.new plant virus familyFlexiviridae and assessment of molecularteria for species demarcation. Arch. Virol. 149, 1045–1060.
hlawat, Y.S., Pant, R.P., Lockhart, B.E.L., Srivastava, M., ChakrabN.K., Varma, A., 1996. Association of a badnavirus with citrus modisease in India. Plant Dis. 80, 590–592.
ariana, H.S., Shannon, A.L., Chu, P.W.G., Waterhouse, P.M.,Detection of five seedborn legume viruses in one sensitive mulpolymerase chain reaction test. Phytopathology 84, 1201–1205
arthe, G.A., Ceccardi, T.L., Manjunath, K.L., Derrick, K.S., 1998. Cipsorosis virus: nucleotide sequencing of the coat protein gendetection by hybridization and RT-PCR. J. Gen. Virol. 79, 1531–1
183–185.uang, Z., Rundell, P.A., Guan, X., Powell, C.A., 2004. Detection
isolate differentiation ofCitrus tristeza virus in infected field treebased on reverse transcription polymerase chain reaction. Plan88, 625–629.
to, T., Ieki, H., Ozaki, K., 2002. Simultaneous detection of six civiroids and apple stem grooving virus from citrus plants by multireverse transcription polymerase chain reaction. J. Virol. Meth.235–239.
acobi, V., Bachand, G.D., Hamelin, R.C., Castello, J.D., 1998. Devment of a multiplex immunocapture RTPCR assay for detectiondifferentiation of tomato and tobacco mosaic tobamoviruses. J.Meth. 74, 167–178.
ames, D., 1999. A simple and reliable protocol for the detectioapple stem grooving virus by RT-PCR and in a multiplex PCR aJ. Virol. Meth. 83, 1–9.
artin, S., Alioto, D., Milne, R.G., Guerri, J., Moreno, P., 2002. Detecof Citrus psorosis virus in field trees by direct tissue blot immunoasin comparison with ELISA, symptomatology, biological indexing,cross-protection tests. Plant Pathol. 51, 134–141.
ehta, P., Brlansky, R.H., Gowda, S., 1997. Reverse transcriptionmerase chain reaction detection ofCitrus tristeza virus in aphidsPlant Dis. 81, 1066–1069.
inafra, A., Hadidi, A., 1994. Sensitive detection of grapevine vA, B or leafroll-associated virus III from viruliferous mealybugs ainfected tissue. J. Virol. Meth. 47, 175–187.
iyakawa, T., Ito, T., 2000. Tatter leaf-citrange stunt. In: Timmer, LGarnsey, S.M., Graham, J.H. (Eds.), Compendium of Citrus Disesecond ed. The American Phytopathological Society, St. Paul,p. 60.
arvaez, G., Skander, B.S., Ayllon, M.A., Rubio, L., Guerri, J., MoreP., 2000. A new procedure to differentiate citrus tristeza virus isoby hybridization with digoxigenin-labelled cDNA probes. J. VirMeth. 85, 83–92.
assuth, A., Pollari, E., Helmeczy, K., Stewart, S., Kofalvi, S., 2Improved RNA extraction and one-tube RT-PCR assay for sim

A. Roy et al. / Journal of Virological Methods 129 (2005) 47–55 55
neous detection of control plant RNA plus several viruses in plantextracts. J. Virol. Meth. 90, 37–49.
Nemichov, L., Hadidi, A., Candresse, T., Foster, J.A., Verderevskaya,T., 1995. Sensitive detection of apple chlorotic leaf spot virus frominfected apple or peach tissue using RT-PCR, -IC-RT-PCR or multi-plex IC-RT-PCR. Acta Hort. 386, 51–62.
Nicholas, K.B., Nicholas Jr., H.B., 1997. GeneDoc: a tool for edit-ing and annotating multiple sequence alignments. Available from:www.psc.edu/biomed/genedoc.
Nie, X., Singh, R.P., 2001. A novel usage of random primers for multiplexRT-PCR detection of virus and viroid in aphids, leaves, and tubers. J.Virol. Meth. 91, 37–49.
Permar, T.A., Garnsey, S.M., Gumpf, D.J., Lee, R.F., 1990. A monoclonalantibody that discriminates strains of citrus tristeza virus. Phytopathol-ogy 80, 224–228.
Ragozzino, E., Faggioli, F., Barba, M., 2004. Development of a one tube-one step RT-PCR protocol for the detection of seven viroids in fourgenera: Apscaviroid, Hostuviroid, Pelamoviroid and Pospiviroid. J.Virol. Meth. 121, 25–29.
Rocha-Pena, M.A., Lee, R.F., Permar, T.A., Yokomi, R.K., Garnsey, S.M.,1991. Use of enzyme-linked immunosorbent and dot-immunobindingassays to evaluate two cross protection experiments after challengewith a severe citrus tristeza virus isolate. In: Proceedings of the 11thConference on International Organization Citrus Virology, Riverside,California.
Rosner, A., Lee, R.F., Bar-Joseph, M., 1986. Differential hybridizationwith cloned cDNA sequences for detecting a specific isolate of citrustristeza virus. Phytopathology 76, 820–824.
Russo, P., Miller, L., Singh, R.P., Slack, S.A., 1999. Comparison of PLRVand PVY detection in potato seed samples tested by Florida winterfield inspection and RT-PCR. Am. J. Potato Res. 76, 313–316.
R lne,with
some affinities to potex, carla, fovea, and allexiviruses. Arch. Virol.145, 1895–1908.
Rustici, G., Milne, R.G., Accotto, G.P., 2002. Nucleotide sequence,genome organisation and phylogenetic analysis of Indian citrusringspot virus. Arch. Virol. 147, 2215–2224.
Saade, M., Aparicio, F., Sanchez-Navarro, J.A., Herranz, M.C., Myrta, A.,Di Terlizzi, B., Palla’s, V., 2000. Simultaneous detection of the threecharacterized ilarviruses affecting stone fruit trees by non-isotopicmolecular hybridization and multiplex RT-PCR. Phytopathology 90,1330–1336.
Scott, S.W., Ge, X., 1995. The complete nucleotide sequence of RNA-3of citrus leaf rugose and citrus variegation ilarvirus. J. Gen. Virol. 76,957–963.
Sharman, M., Thomas, J., Dietzgen, R.G., 2000. Development of a multi-plex immunocapture PCR with colourimetric detection for viruses ofbanana. J. Virol. Meth. 89, 75–88.
Singh, R.P., Kurz, J., Boiteau, G., 1996. Detection of stylet borne andcirculative potato viruses in aphids by duplex reverse transcriptionpolymerase chain reaction. J. Virol. Meth. 59, 189–196.
Timmer, L.W., Garnsey, S.M., 1979. Variation in the distribution of citrusringspot and psorosis viruses within citrus hosts. Phytopathology 69,200–203.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., Higgins,D.G., 1997. The Clustal X windows interface: flexible strategies formultiple sequence alignment aided by quality analysis tools. NucleicAcids Res. 24, 4876–4882.
Uga, H., Tsuda, S., 2005. A one-step reverse transcription-polymerasechain reaction system for the simultaneous detection and identi-fication of multiple tospovirus infection. Phytopathology 95, 166–171.
Vidalakis, G., Garnsey, S.M., Bash, J.A., Greer, G.D., Gumpf, D.J., 2004.s in
ustici, G., Accotto, G.P., Noris, E., Masenga, V., Luisoni, E., MiR.G., 2000. Indian citrus ringspot virus: a proposed new species
Efficacy of bioindexing for graft-transmissible citrus pathogenmixed infections. Plant Dis. 88, 1328–1334.












![A Molecular Biology: Open Access · 2020-01-09 · reverse transcription polymerase chain reaction (RT-PCR) [16,17], Multiplex RT-PCR (mRT-PCR) [18] and real-time RT-PCR [19,20]](https://img.pdfslide.us/doc/110x75/5f0cc7037e708231d43714b2/a-molecular-biology-open-access-2020-01-09-reverse-transcription-polymerase-chain.jpg)















