Embed Size (px)
Citation preview

490 Jounxel oF THE ArrarnrceN Mosqurro CoNrnol AssocrarroN Vol. 2, No. 4
A MODEL FOR POLYGENIC INHERITANCE OF ABDOMINALTERGAL SCALE PATTERN IN AEDES AEGYPTII
W. KEITH HARTBERG2, CYNTHIA K. MEEKSs and KENNETH R. WILLTAMS.{
ABSTRACT. There is much variation in the amount of white scaling on the abdominal tergites of Aed.esaeglptl.Th3 genetic basisfor^this white scale pattern was investigated in"two.laboratory strains e-stablished bysefection from the CARN .strain of Ae. ae!1pti. These experimental strains were irossed in all possibledirections insingle pair matings. Based on anilysis of their piogeny it is proposed that genes at three.l.p.iu,.Inclependently assorting loci control abdominal tergal scale pattern. Correlation of observed dad andexpected dam was high. Since the abdominal tergal scale pattern'in Ae. aegyfti appears to be controlled bv one
ll1.9,lP:lyg.tic system with modifiers. it.is proposed that it is berter io'coniider differences in ethdlogy,
physrology' and reproductive behavior as the bases for separation of the species into intraspecific groups.
INTRODUCTION
Aedes aegypti (Linn.) imagoes have a black tobrownish cuticle with black, silver, matte-white,or gold scales appearing in an assortment ofpatterns on various body parts. The abdominaltergites have black or brownish backgroundscaling with light (silver or matte-white) scalesappearing in lateral spots and basal bands;sometimes additional white scales extendingfrom the basal bands are present on the tergitEproper. The abdominal tergal scale pattern hasbeen used as the primary bisis for c[assificationof this species into subspecies and varieties(Mattingly 1957).
Based on scaling of abdominal tergites,Matt ingly (1957) recognized three forms oTAe.aegypti. -These are: Ae. aegypti, the type form,with pale scaling on rhe first abdominil tergiteand/or a distinctly paler or browner body t[anthe blackish African subspecies/onnosrrs (Walker);Ae. aegypti formons, which never has any palescales on the first abdominal tergite,
'his a
markedly blackish appearance, and is confinedto Africa south of the Sahara; and Ae. aegyptivariety queenslandenses (Theobald), which-hasincreased white scaling on the abdominaltergites beyond the first, and/or has a lightermesonotal color. Mattingly's (1957) classifica-tion s,ystem stressed the presence of pale scaleson the first abdominal tergite as a gooddiagnostic character of frequent occurrCnce;however, he also noted that it was notcompletely constant.
I This investigation was supported in part by aresearch grant to the first author from the GeorgiaSouthern College Faculty Research Fund.
_ 'Institute of Anhropodology and parasitology,
Department of Piology, Georgia Southern College,Statesboro, GA 30460. Current address: Departme-ntof Biology, Baylor University, Waco. TX 76798.
, r Department of Biology, Georgia Southern Col-
lege, Statesboro, Ga 3046b.
^ a Computer Services, Georgia Southern College,
Statesboro, GA 30460.
A wide range of abdominal tergal scalepattern exists even within distinct populationsof Ae. aegypti. McClelland (1960t examinedabdominal tergal scaling in Ae. aegypti hatchedfrom eggs collected from filter paper-linedwater pots. ,in Kenya. In selected matings,imagoes with abdominal tergal scale patteincorresponding to the nominate form producedprogeny representative of subspecies forrnosusand the nominate form. Parental
-forrnostu
phenotypes produced both forrnons and nomi-nate form offspring. Paler forms, i.e., thoserepresentative of variety queenslandensls, re-sulted in queenslandcwLl pro$eny. These resultswere.substantiated by Hartberg (1969), whoexamined progeny of field-cofected femalesfor abdominal tergal scale pattern and alsofound that femaleJ corresponding to subsoe-ci es form o nu prod uced offs pri n g re-prese.rtatiueof formonu and the nominate form; nominateform females gave offspring of both formostuand nominare form appearance. McClelland(1974) analyzed 74 samples from 69 differentpopulations for abdominal tergal scale pattern.Again, his data showed a wide range ofvariation in abdominal tergal scale pattern.Mogi et al. (1984) also showed a wide ringe ofvariation in abdominal tergal scale patterns ineight populations of Aedes aegypti- from thePhilippines.
Since there is variation in abdominal tergalscale pattern even within the three subdivisionsof Ae. aeglpti, this variation is probably geneticand heritable. The genetic mechanism For theinheritance of abdominal tergal scale color haslong been a subject of speculition. McClelland(1960, 1967, 1974) proposed a mult i factorialmode of inheritance ior bbdominal tergal scalecolor; VandeHey et al. (1978) presenred darathat suggest a polygenic sysrem of inheritance.Other investigators suggested a monofactorialscheme. Johnston and Hartberg (1981) pre-sented a polygenic system of inheritance forvariation in scale pattern of the abdominalsterna of the mosquito Eretrnapodites quinqueait-/atzrs (Theobald).
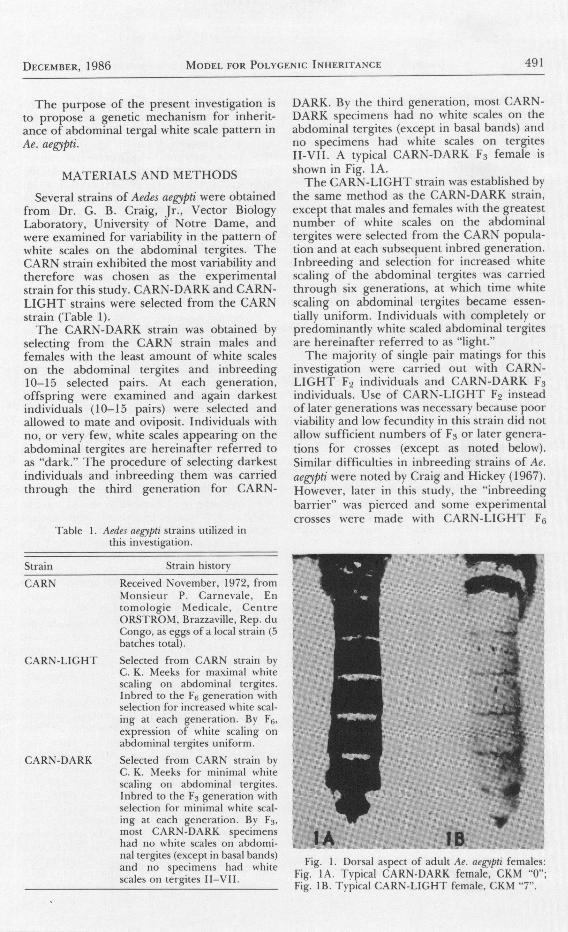

492 JounNel oF THE AurnrcaN Mosqurro coNrnol AssocrarroN vor-. 2. No, 4
individuals (Fig. lB). No significant differenceswere noted between these crosses using CARN-LIGHT F6 and those using CARN-LICHT Fz.Both sexes of the F6 CARN-LIGHT srrain areessentially identical to each other with regardto abdominal tergal scale pattern.
Rearing methods were essentially those de-scribed by Craig and VandeHey (1962) forgenetic research with Aedes aeglpti. All mcsqui-toes were reared in an insectary wittr atemperature of 26' + 3oC and ambient relativehumidity. Larvae were fed on a suspension ofliver powder in-rap water (12 ccllitbr). pupaewere segr:egated by sex and placed in emer-gence cups. After emergence adults wererechecked for sex to ensure that females usedin crosses were virgins.
For single pair matings, etherized virginadults were examined with a stereo-dissectingmicroscope and appropriate males and female-sselected. Each single pair was placed in a cagemade from a pint-sized cardboird container.-Ashell vial lined with absorbent brown papertoweling and 3A filled with warer was injeriedin the cage to provide an oviposition site an<iprevent desiccation. A sugar crrbe *as providedas a carbohydrate source. Females weri offereda blood meal from an anestherized whitemouse 4-b days after emerging. Ovipositionusually occurred 4-5 days after the blood meal.
Adult mosquitoes to be examined for abdom-inal tergal scale pattern were removed fromemergence cups within 24 hours postemer_gence to prevenr loss of scales due tb rubbinsand scraping against each other and/or aeainsithe emergence cup. Adults were anesth;tizedwith ether and transferred to an examinationcontainer for classification. An Olympus stereo-dissecting microscope with a hish-intensitvlight source and heat filter was u*sed in theexamination of all mosquitoes. To eliminateclassification error, it was necessary to orient allimagoes in the same direct ion, since individualsimproperly oriented reflected light differently,and thus obscured the tergal scale pattern.Two pairs of Inox No. 5 watchmaker's-forcepswere used-to separate the wings and expose theabdominal tergites for examination.
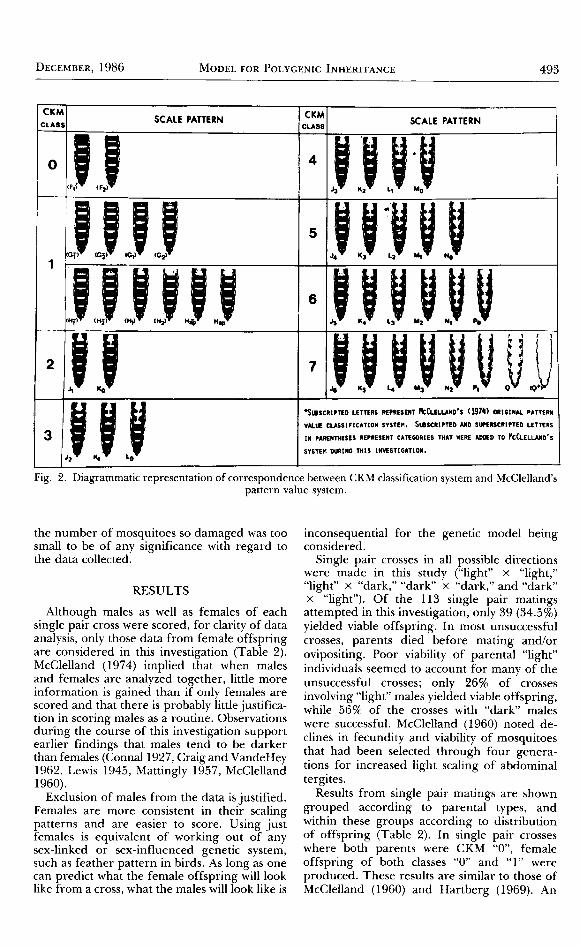
McClelland (1960, 1974) described a svstemfor classifying Ae. aegypti according ro abdomi-nal tergal scale pattern based on the number oftergites that have a continuous medial band ofwhite scales from the apical ro the distal end.The number of rhe mosi posterior tergite withsuch scales is used to d6note the dJgree ofpaleness; in general, all tergites anterioito thistergite are as pale or paler than it is, and thoseposterior to ir, especially the adjacent one, havescattered pale scales (McClelland 1960). McClel-land arbitrari ly assigned a lerrer (F. G, H, J. K,
L, M, N, O, P, or Q) to denore eleven colorgrades, "F" representing a mosquito that hadno white scales except in basal bands (as insubspecies /o rrno su,s, throu gh "Q, " representin gthe most extreme expression of varietyqueenslandensei, where only speckled black scales
?Ppeared on the tergites (McClelland 1974).Since there was so much variation in tergalscale pattern, McClelland (1974) expanded iiresystem to cover 30 basic patterns by subscript-ing each letter with a digit to indicate thenumber of tergites with white scales. Thus,McClelland in his later work incorporated boththe amount of white scaling and the number oftergites with white scales to form a ,,pattern
value" method for mosquito classification.It became apparent early in our investigation
that the number of tergites with white icales,rather than the amounr of white scaling on thetergites, was a better crirerion for classiication.This prompted the formularion of the CKMsystem of classification. In the CKM method,only the number of tergites with white scales(except in basal bands) is considered regardlessof the number of white scales on the iergites.Since abdominal segments VIII-X are riodi-fied as genitalia in mosquitoes, only segmentsI-VII were considered for classificaiion]Wher,white scales were present on any given tergite,they always appeared on all tergitis anterio"r rothe given tergite. Additionally, the amount ofwhite scaling usually decreased from anteriorto posterior end of the abdomen.
There are eight possible classes in the CKMsystem: "0" if no white scales appear on thetergites (except in basal bands), .ili' if only thefirst tergite has white scales, "2" if the firsi twotergites have white scales, up to,,7" if all seven'tergites being considered have white scales.The eight classes of the CKM systenr areequated to McClelland's partern value classes inFig. 2. All mosquiroes examined in this studywere classified according to both McClellandtoriginal system and the CKM.system developedduring the present investigation.
According to McClelland (lgj4\, some sub_jectivity in determining rhe partern value ofindividuals scored is unavoidable since all typesof intermediates exist, but he .tut",
'tir"t
subjectivity is unlikely to cause an error ofmore than one pattern value unit. This error isavoided in the CKM merhod, since one caneasily determine whether white scales arepresent or absent on any given tergite. Onepossible source of error in classificati,on in thepresenr study might have been the loss of whitescales due to mosquitoes rubbing against eachother^and/or rubbing against the imergencecup. Since imagoes were examined as qi icklyas possible after emergence, it is believed thal
Dncaunrn, 1986 Moorr ron PolycrNrc IturnrreNcr 493
CKMC L A S S
SCATE PATIERN CKMcLASS
SCAIE PATTENN
oLFF 4 ,H-H.H"H.t F.F..F "F 5 F$H$PtFF"H,HFF6 F$ H"H-H$
2 FF 7 FF$$$Itij !3 F$F
'Suscir?TED LETTER! r€ftEs:lt kCLELrrrD's (1974) ontotul prrrtmr
ylltf cLAstftclttoi svstEr, SttscRtttED ttD SttEl3ctt?lED LEtrcrs
lx patcitxEsEs rEpt€3a{t cArE@RlEs txAt |ERE aDGD to raCLELLITD'S
syslElr DURILG rxls l l t t tsr lGAtl0n'
Fig. 2. Diagrammatic representation of correspondence between CKM classification system and McClelland'spartern value system.
the number of mosquitoes so damaged was toosmall to be of any significance with regard tothe data collected.
RESULTS
Although males as well as females of eachsingle pair cross were scored, for clarity of dataanalysis, only those data from female offspringare considered in this investigation (Table 2).McClelland (1974) implied that when malesand females are analyzed together, little moreinformation is gained than if only females arescored and that there is probably little justifica-tion in scoring males as a routine. Observationsduring the course of this investigation supportearlier findings that males tend to be darkerthan females (Connal 1927, Craig and VandeHey1962, Lewis 1945, Matt ingly 1957, McClel land1960).
Exclusion of males from the data is iustified.Females are more consistent in theii scalingpatterns and are easier to score. Using justfemales is equivalent of working out of anysex-linked or sex-influenced genetic system,such as feather pattern in birds. As long as onecan predict what the female offspring will looklike from a cross. what the males will look like is
inconsequential for the genetic model beingconsidered.
Single pair crosses in all possible directionswere made in this study ("light" x "light,""light" x "dark," "dark" x "dark," and "dark"
x "light"). Of the ll3 single pair matingsattempted in this investigation, only 39 (34.5Vo)yielded viable offspring. In most unsuccessfulcrosses, parents died before mating and/orovipositing. Poor viability of parental "light"
individuals seemed to account for many of theunsuccessful crosses; only 26% of crossesinvolving "light" males yielded viable offspring,while 56% of the crosses with "dark" maleswere successful. McClelland (1960) noted de-clines in fecundity and viability of mosquitoesthat had been selected through four genera-tions for increased light scaling of abdominaltergites.
Results from single pair matings are showngrouped according to parental types, andwithin these groups according to distributionof offspring (Table 2). In single pair crosseswhere both parents were CKM "0", femaleoffspring of both classes "0" and "1" wereproduced. These results are similar to those ofMcClel land (1960) and Hartberg (1969). An

494
bD
h
q,
bo
bo
F
o
, q
a
o
()
U
a!
0
!o
N I9 l
a l> l| ( I. r I
- lo lq r l
; l0 l
o l9 l
O I
0 lF I
U I. l
c \ lI
{ ) l
F I
JounNar- oF THE AunnrceN Mosqurro Cournol AssocrarroN Vor,. 2, No. 4
Io otr .sa 4
l'IIl 'I
* 5 |.*d IE E I: U I
9 t r 1
_ t ; l
z e l
It -IIl a
l'Il.Il '
16 l
x * |; 'L I
c - X l $
0 t r 1
, ! 3 13 ! l -; > |9 V Ic v I
2 l
l-It-Il o
' 6 A d
F . 9 Iv u o
- Q $ - @ O 6 @ N r - N O @ ! Q N @ @ s O O S d O d N O i d o @ o @ r o 6 d 6 or o i l t s o - d i + - N 6 @ d d o $ + - * R ; 6 6 d t s r d 6 i c i - 6 : : o ; 6
i ^ ! - @ - 6 H N @ O
R 6 w s 6 - N < $
s s i 6 € N r o H d o @ o @ 6 $ e o @ ri d d H 6 o N t s i N d 6 t s i r r <
d N
6 $ r - $ o+ < i 6 6 -
- @ O @ - N 6 O 6 6 + d O O or 6 6 r @ i d - N - o
q O $ r A - O - N 6 $T O N T 6 i N F ! o 9 $ $ i ! ! ) $ d Q o i - o @ o 6 6 $ o o @ t s
i d F d d - O @ d d i N 6 t s ; F t s t
_ N
v 6 i
6
O g N - s O N 6 ! d O O ov r : n 6 - N - d F o
- @ O @ - N @ Or 6 6 N € -
- * J O O C C O @ 6 @ 6 6 cF F F F 6 6 d 6 ; F 6 5 5 5 5 5 5 5 5 5 5 5 E 3 4 4 5 5 = = E S A E A E A ; ;
ooooooooo€aa6 'oo_o_oo66 '6 o6 '6 '6 '6 6 '6 '6 6 '66 ,a ' ,6 6 ,a6 6 6 -F F F F F 5 5 5 5 Y : 5 5 5 5 5 5 F F F F F F F F F F F F F F F F F F F = F F
1 3 F F R 9 S - - E E S ; $ 5 9 ! ! s o o * R g - R X R R 3 S g 6 9 g g s 5 9
6l
bb
o
boo
0
(,
a
C)
(.)
>.
oE
Oh
a
!
@
3
oId
(!
OPO
d
0
!0
I
o
q)
00q r '!
o!o
l.l '
o!o
I
v(JEO
**
UI
0
-c)
;. H
I
I
o
k0
I
q)
I()h
oo-0k ,
o
U
dI
IU
oo
I
I
lk
*

Dtcnnrsnn, 1986 Moorr ron PolycrNlc Iusrx.rraxcr 495
individual scored as CKM "0" is representativeof Ae. aeglptiformosu,s, while one scored as CKM"1" corresponds to the type form. Althoughmale data are not included in this report, themajority of males that resulted from thesecrosses were classified as CKM "0": onlv a fewCKM "1" males were observed from "dark" x"dark" crosses. Of the twenty "dark" x "dark"
crosses attempted, six (30%) were successful."Light" x "dark" matings were rhe most
successful of all types of crosses made in thisstudy. Thirty such single pair matings wereattempted; sixteen (53%) of these producedoffspring. Although these parents appearedphenotypically alike (i.e., all females "light" andall males "dark"), variation was exhibited by theoffspring of the various crosses. Three of thecrosses (cross numbers 6, 7, and l0) yieldedoffspring ranging from a CKM "1" to a CKM"7." Four crosses (cross number g, 22,27, and60) gave some dark offspring (CKM "1") andsome light offspring (CKM "6" or "7"). Theremainder of these crosses resulted in lightprogeny (CKM "5" - "7").
Variability in progeny classes was also ob-served in matings of "dark" x "light." In crossnumbers l, 3, 42, and 49, offspring werephenotypically light. Cross numbers 66 and 67differed slightly from the former crosses'parental types, and produced both light anddark offspring. Of the thirty "dark" x "light"
crosses attempted, only six (20%) of theseyielded offspring. Decreased success in thesecrosses was thought to be due to poor viabilityof oarental CARN-LIGHT individuals.
fwo kinds of "light" x "light" crosses wereattempted. The first group (cross numbers 12,20, 55,73, and 77) involved parental CARN-LIGHT and resulted in all CKM "7" progeny.The other group (cross numbers 106-113),involved F1 individuals obtained from a singlepair "light" x "dark" (cross number 90), inwhich all 29 female progeny were CKM "7"
and all 27 male progeny were CKM "6." Thesix successful crosses of these eight attemptedresulted in both dark (CKM "0" and "1") andlight (CKM "7") female offspring. Althoughthe parents in these crosses (cross numbers106-113) were all phenotypically light, bothlight and dark offspring appeared in the F2generation. The overall success of both kindsof "light" x "light" crosses was 33Va. However,there was a great difference in the two groupsof "lighC' x "light" crosses when each groupwas considered separately. The success rate forthe first group was only 20Vc while 75% ofthe crosses in the second group (F2 individualsfrom single pair 90) were successful. Thestriking success of this second group of crosses
may be an exemplification of some type ofheterosis.
Based on results of the single pair matings, itis proposed that genes at three separateindependently assorting loci control the patternof white scaling on the abdominal tergites inAe. aeglpti. The first of these loci determineswhether white scales can or cannot appear onthe tergites, and is designated "tergite-white"
(Tw). In the homozygous recessive state(Tw-Tw*), this locus is epistatic to genes at theother two loci and prevents the appearance ofwhite scales on the tergites. In the homozygousdominant (TuTw) and heterozygous (Tu,,Tza+)forms, this locus allows for white scaling of thetergites, but does not contribute to or influencethe amount of white scaling. The second locusis occupied by a series of multiple alleles whichIargely determine the amount of white scalingon the tergites. These genes are termed"white-scaling" and are symbolized by L1, L2,and 1. 11 produces more white scaling thandoes 12; 1 in a homozyous state produces nowhite scaling. At the third locus appears a genepair that intensifies the effect of the series of Lalleles, producing increased white scaling ofthe tergites except in the homozygous recessiveform. This is the "white-intensifier" locus. 1. Asummary of these postulated gene pairs, theircomputer symbol, the computer model valueassigned to each, and the effect ofeach pair onabdominal tergal white scaling is given in Table3 .
When the "white-intensifier" genes (II or Ii)appear with the genes for "white scaling"(LtLb hL2, L2L2, Lt l , or L21) and where theepistatic genes for "tergite-white" are notpresent to block their effect, the result is amosquito with increased white scaling on theabdominal tergites. But when the "white-
intensifier" genes (1/ or 1i) appear with therecessive alleles for "white-scaling" (1 1), nointensification of white scaling results, as the 11alleles produce no white scales. Therefore, itwould be possible to have "white-intensifier"
genes present in a parent generation, and tohave these genes passed to progeny, eventhough the parent generation had no genes forwhite scaling of the tergites.
From the combinations of the two differentalleles at the Tu locus, the two different allelesat the 1 locus, and the three different alleles atthe l, locus fifty-four possible genotypes (3 x 3x 6) were generated; "Computer ModelValues" were assigned to each genotype (Table4), for use in the computer analysis describedbelow.
From the 54 possible genotypes proposed inthe genetic model, 2,916 hypothetical matings(54 x 54) could occur. However, a number of

496 JounNar, oF THE AunnrceN Mosqurro CoNrnor Assocrerrorrl Vor-. 2, No. 4
Table 3. Postulated gene pairs, their computer symbol, their computer model value, and their effect onabdominal tergal white scale pattern.
Gene pairComputer
SymbolComputer
model value Effects of tergal scaling
TwTwTwTu+Tw+Tu+
LtLt
LtLzL2L2, L1I
Lzl
1 1IIJi
ii
LL
LMMM,LN
MN
NNII,IJ
JJ
SSSTTT
000
Allows white scaling.Allows white scaling.Epistatic to L1, L2, and 1; allows no
white scaling.Produces extreme amount of white
scaling.Produces large amount of white scaling.Produces intermediate amount of white
scaling.Produces minimal amount of white
scaling.Produces no white scaling.Intensifies white scaling if Tw and L1 or
l,2 are present.No intensification of white scaling.
3*2x
l *
02*
0* These values are only used when the gene pairs TwTu or TuTu+ are present in the genotype. In the
Presence of the gene pair Tu+Tw+ these vilues are nottsed and the computer model value io"ld 6. 0, sinceTw*Tu* is epistatic to these genes.
these crosses are reciprocal crosses, and thusyield no additional information since thesecharacters do not appear to be influenced bysex-linkage. The number of distinct possiblecrosses, eliminating reciprocal crosses, is 1,485[n(n+l)/2], where n equals the number ofparental genotypes. These crosses can berepresented as follows:
Cross parent #3 with parents #3 through#54
:Cross parent #54 with parent #54.
This crossing scheme eliminates reciprocalcrosses so that each cross is unique.
These 1,485 crosses can produce 64 differ-ent gametic combinations (progeny), sinceindividuals with three pairs of independentlyassorting genes produce eight types of gametes
Cross parent #l with parents#54Cross parent #2 with parenrs#54
#l through
#2 through
Table 4. Possible genotypes and their computer model values (C.M.V.).
C.M.V.* Genot c. .vv000000II2
2.l
3n
3
Genot .v.v44,A
4
444
6
55
6666
C.Tw*Tw* I l i iTw"Tw* I l I iT w * T w * l l I ITw*Tw"L2 l i iTw 'Tw*L2 l I iTw*Tw*L2 l I ITw*Tw*L2L2i iTw+Tw+L2l-z i iTw+Tw+L2L2IITw"Tw*L1 l i iT w * T w * L 1 l l iTw*Tw*L1 I ITw+Tw+L1L2 i iTw*Tw 'L1L2 I iTw*Tw*L1L2 I ITw"Tw*L1L1 i iTw*Tw*L1L1 I iTw+Tw+LrL l I I
TwTw+ I l i iTwTw+ I l l iTwTw" l l I ITwTwIl i iTwTwl l I iTwTwl l I ITwTw"L2li iTwTwl2IiiTwTw*L2L2iiTwTw+L1 l i iTwTwL2L2iiTwTwLll i iTwTw+L2 l I iTwTw+L2 l I ITwTw*L1L2iiTwTwL2lI iTwTwL2lIITwTwLlL2ii
TwTw*L2L2I iTwTw+L2L2IITwTw+L1 l I iTwTw+L1 l I ITwTw+L1L1i iTwTwL2L2IiTwTwL2L2IITwTwLl l I iTwTwL l l I ITwTwLlLl i iTwTw+L1L2I iTwTw+L1L2IITwTwLlL2I iTwTwLIL2I ITwTw+L1L1 I iTwTw*L1L1 I ITwTwLlLl I iTwTwLILI I I
000000000000000000
*Note that Tw+Tw+ is epistatic to L1, L2, and land allows no white scaling; therefore C.M.V. is 0 whenTu+Tw+ is in the genotype.

Drcnnrsrn, 1986 Moonl ron Porycnxrc INnrnrraNcs 497
with equal frequencies. Crossing of two suchindividuals yields 64 (8 x 8) possible combina-tions of these gametes. These gametic combina-tions (progeny) may or may not be genotypi-cally distinct. Those genotypes producing similarphenotypes are grouped by the computer sothat phenotypic ratios for each mating arereadily apparent.
The obvious plethora of possible matings(1,485) and progeny (95,040) necessitatedcomputer analysis. The computer program waswritten so that grouping of crosses by expectedphenotypic distribution of offspring was possi-ble. This allowed the rapid identification ofhypothetical crosses which would produceresults similar to those observed, thus facilitat-ing comparison of observed data to theoreticaldata. The computer program was written inFORTRAN EXTENDED for a Control DataCYBER 70 model 74. Since the program usesextensive character manipulation, it is highlydependent on the extended features of ControlData FORTRAN.
The basic design of the computer programinvolves the creation ofthree tables: (l) A tableconsisting of six character patterns represent-ing the fifty-four possible genotypes. This tableis created one time only. (2) A table ofcorresponding "Computer Model I Values" as-signed to the 54 genotypes. This table iscreated one time only. (3) A table of frequen-cies, i.e., each entry is a count of the number oftimes the corresponding genotype would occurwhen all 64 possible offspring from a givencross are examined. This table is created andresults are printed for each of the 1,485possible crosses. As stated in the previousparagraph, the crosses are grouped by ex-pected distribution of offspring, thus makingcomparison ofobserved to expected results foreach cross easier to complete.
Following are a few selected crosses from thecomputer printout to illustrate how the ex-pected results are displayed. To the right of thecross the Comouter Model Value for theparents crossed-is in parentheses. The geno-types of the parents and offspring are given intheir computer symbols (see Table 3 for genesymbols). The expected frequency of theoffspring for each Computer Model Value islisted under each category. It should be notedthat the crosses between phenotypically similar,but genotypically different.. parents can givedivergent results.
In order to quantify the degree of whitescaling on the tergites, numerical values wereassigned to the proposed alleles. These numer-ical values are referred to as "Computer ModelValues" (Tables 2, 3, and 4). Since the Tzu locusonly determines whether white scales can or
cannot appear on the abdominal tergites anddoes not contribute to the amount of whitescaling, this gene pair was assigned a computermodel value of zero. The series of I genes andthe 1 genes were assigned computer modelvalues as shown (Table 3). Genotypes with acomputer model value of 0 or I would appeardark, with few if any white scales except inbasal bands, which are always present ontergites II-VII and may or may not be presenton tergite I. Intermediate genotypes, i.e., withamount of white scaling between dark andlight, have computer model values rangingfrom 2 to 4. The light genotypes, where whitescaling was maximal, have either a 5 or 6computer model value. Often in these lattergenotypes dark scaling was limited to a fewblack scales speckled along the lateral part ofthe tergites.
These computer model values for the pro-posed alleles were then equated to the CKMmethod of classification. where classification isbased solely on the number of tergites havingwhite scales (Fig. 3). It will be noted that anyone computer model value may be representa-tive of two or three CKM categories. Thisoverlap of categories may be attributable tovarious factors: several modifier genes mayaffect expression of white scaling; differentkinds of gene interaction may result in differ-ent phenotypic expression of alleles; and, thecomputer model values are only numericalestimates of the amount of white scaling thatcan be expected. Thus, any one category ofclassification may contain a very few individu-als of the category immediately adjacent to iton either side. It is believed. however. that themajority of individuals fell in the propercategory of classification, with a few extremesfalling into adjacent categories. For example, amosquito with a genotype resulting in acomputer model value of zero may in fact havea few white scales on the first tergite, appearingto be a CKM "1" instead of a "0". Or, due to adifferent effect of modifier genes or differentgene interaction, a genotype with a computermodel value of "5" may appear to be a CKM"6. "
It is felt that this overlap of categories isjustifiable since, as Mather (1942) has pointedout, the interaction of the genes of organismsand nonheritable agencies can contribute to thelack of completely unambiguous phenotypicclasses for polygenic traits.
Data from female offspring of the 39successful single pair matings are grouped inview of this proposed overlap of categories(Table 2). From these grouped data, theexpected computer model ratio of offspringphenotypes was determined (Table 5). Chi-

498 JounNal oF THE Aurnrceu Mosqurro CoNrnor AssocrerroN Vor,. 2, No. 4
TI!,!NIJ x TII|NII (0 x 0)
0 L 2 3ffiit-T
_ ! L 2 3 4 sffiffi'ffirTc-- ffiT'SSNNJJ 8 SSMNJJ 16 SStO{^u 8
TINNII 8rI},INIJ 16T1[r{NII 15TIT.'MIJ 8TII'IMII 8
SI?{NJJ x SSMNJJ
TTLNJJ X STLNJJ
TTT.NJJ 16TIT,TJJ 8SINNJJ 8
TTI.'NIJ x SSLMIJ
( 1 x l )
( 0 x 2 )
( 0 x s )
0 L 2 3 4mffi6' ffitrG'r:- ffitir
6
1 2 3 4 5ffia- mffi-r ffiffi ffiirT sEffi
STLNJJ 4 SITi,INII 4 S$.IMII 4 STIMII 4SIEMJJ 4 STLNIJ 8
STLNII 4
TITNJJ x STLMIJ ( 0 x 5 )
0 L 2 3 4 5 6mffiffi4- ffirz- ffiJt- sr!or-i- ffiiir-? srmii-- sr-E#-TTIT.INIJ 4 SIEI{'J 4 SfLIJJ 4TIT.NJJ 4TIT,NIJ 4TTEMJJ 4TTEMIJ 4TI'LTJJ 4TTLLIJ 4
S"ILLII x SfLLII (5 x G)
0 L 2 3 4 5 6ffiI6 -
SSLLII 16
SSLUJ x SSLLII ( 4 x 6 )
2 3 4 6Sffifr-A
I

Dncrprnrn. 1986 Moorl FoR PoLYcENrc lNnrnrraNcr 499
&tourrn}ODEL
I, r l- t
III
cre,l 6+pFz---
II
r-li{ Fg-+r *-s-+
Fig. 3. Overlap of categories of classification: computer model and CKM systems
square analysis of deviation of observed datafrom expected ratios was Derformed for eachcross. Tlie Chi-square value and the probabilitythat deviations from the expected are due tochance alone are given (Table 5).
Several times during the course of this study,a light-colored cuticle and light backgroundscaling were observed. The lightness of thiscuticle and scale color was striking whencompared to the normal black or brownish-black cuticle of most Ae. aegypti. A descriptionof the light cuticle and background scales waspenned at the time as "light brown" or "very
light" brown-almost yellowish." This phenom-enon was noted in later selections of theCARN-LIGHT strain and in heavily white-scaled progeny of some single pair matingswhere parental CARN-LIGHT individuals wereused.
Observations during the course of this studysubstantiate earlier findings that males appeardarker than females (Connal 1927, Craig andVandeHey 1962, Lewis 1945, Matt ingly 1957,McClelland 1960). Males generally fell a fullcategory of classification below (darker than)females except in crosses where all progenywere extreme darks or extreme lights. Lightestmales observed in this study were classified asCKM "7," but no male with white scalescovering the entire abdomen was ever ob-served. The first six tergites of the males inpoint were completely clad in ,matte-whitescales, the white pattern ending abruptly withthe white apical band of tergite VII. This is insharp contrast to the white scale pattern inCKM "7" females, where white scales usuallyextended without interruption to the distal endof the tergite VII. In males, randomly scat-tered white scales appeared on the seventhtergite, the number of white scales beingextremely variable. The white scales of thistergite were never observed to form anypattern. Neither were they concentrated onany particular area of the tergite. In theCARN-LIGHT F6 population males differfrom each other only in the number and
random arrangement of white scales on theseventh abdominal tergite.
CARN-LIGHT females also differ from eachother in scaling of the seventh tergite. Gener-ally this tergite, like all those anterior to it, iscompletely covered in matte-white scales. Inmany specimens, however, black scales invarying numbers are present on the apicalportion of tergite VII. When present, blackscales form rough triangles on either side ofthe tergite. These triangles of black scalesbegin at the apex of the tergite and meet thelateral margin of the tergite at about one-thirdthe length of the tergite. The black scales donot fuse in the center of the tergite, so that theappearance of these two small black antero-lateral triangles results. Females vary less inregard to scaling of tergite VII than do males.
DISCUSSION
All experimental crosses except one (crossnumber 7) support the proposed geneticmechanism at the 0.05 or greater level ofsignificance. This exceptional cross yielded asmall number of individuals; thus, small samplesize and/or human error in choosing parentaltypes or in scoring progeny may be responsiblefor failure of this cross to support the proposedgenetic model.
Until rather recently there has been a basicassumption that any quantitative trait must beconrolled by a large number of polygenes.Reexamination of this assumption by Thompson(1975, 1977, 1979) shows that only a smallnumber of loci are involved. The presentinvestigation supports his view since onlythree loci are involved in the genetic systemproposed and supported in this study.
Several mutant genes are known to affectabdominal tergal scale pattern in Ae. aegypti.The mutations white abdomen (I4/) and lateralsilver spot (s), which when present increase theamount of white scaling on the tergites, weredescribed by Craig and VandeHey (1962).Craig and Hickey (1967) list spot (s) and

500 JounNel oF THE AurxrcaN Mosqurro CoNrnor AssocrarroN Vol. 2, No. 4
Table 5. Chi-Square analysis of results of single pair matings.
Expectedcomputer model value ratro Model fitCross
numberClassification
of parents* 12 value
r25 5
l ca 1
2049L 9
I3
6667293 l34J I
39l 0 ll 076I
222760
8262428293084908 l
l l 0108106tt2107l l 3
7(6) 7(6)7(6) 7(6)7(6) 7(6)7(6) 7(6)7(6) 6(6)0(0) 6(6)0(0) 6(6)0(0) 6(6)0(0) 5(6)1(0) 7(6)r(0) 6(6)0(0) 0(0)0(0) 0(0)0(0) 0(0)0(0) 0(0)0(0) 0(0)0(0) 0(0)7(6) 0(0)7(6) 0(0)7(6) 0(0)7(6) 0(0)7(6) 0(0)7(6) 0(0)7(6) 0(0)7(6) 0(0)7(6) 0(0)7(6) 0(0)7(6) 0(0)7(6) 0(0)7(6) 0(0)7(6) 0(0)7(6) 0(0)7(6) 0(0)7(6) 6(6)7(6) 6(6)7(6) 6(6)7(6) 6(6)7(6) 6(6)7(6) 6(6)
IIJ J
3 l3 l5 5
3 ll l/4
IItI
q
333J
3
0 0.990 0.990 '0.990 0.990 0.99
0.47 0.300 0.99
0.89 0.300 0.99
0.43 0.500.09 0.700.26 0.500.24 0.50
0 0.990.82 0.300.54 0.30
0 0.990.43 0.708.22x* 0.014 . r5 0 .100.17** 0.500.10 0.700.84 0.30o.44 0.50
0 0.990 0.990 0.990 0.990 0.990 0.990 0.990 0.990 0.99
0.07 0.700.53 0.700.03 0.70l . l 9 0 .203.44** 0.050.53 0.30
* First number represents cKM classification of individuals crossed; numbercomputer model classification. Female parent listed first.
*x Yates'correction factor used on these data.
i:r parentheses represents
pale-abdomen (pa) as causing increased whitetergal scaling. A gene designated as blacktergite (Bl*)5 (VandeHey et al. 1978) conrrolswhite scal ing of the f irst abdominal tergiret i tsallele, Bt, produces no white scales on the firsttergite. These genes may serve as modifiers tothe presently proposed polygenic system forinheritance of abdominal tergal scale pattern inAe. aegpti.
- A strikingly differenr parrern of scaling ofthe seventh tergire in CARN-LIGHT wasobserved during this investigation. So far ascan be determined, these observations have notpreviously been reported in the literature. Thescale pattern of this tergite may be influencedby a genetic system other than the onecontrolling the scaling of the first six rergites.Such an influence would not be tompletelyunexpected since tergite VII serves as some-what of a housing for the terminalia which inboth sexes are formed from modifications ofsegments VIII-X. Tergite VII may thereforebe influenced by genes which conrol develop-ment of the terminalia in both sexes. as well ls
s Machado-Allison, C. E. 1971. Genetic differencesamong subspecies of Aedes aeg,tpti and their evolution-ary implication. Ph.D. Thesis, University of NotreDame, Notre Dame. IN.

Drcrnrstn, 1986 Moorr ron PorvcnNtc INunnruNcr 501
by those genes which control rotation of theterminalia in males. That the seventh tergiteappears to be more greatly affected in malesthan in females suggests the possibility of asecondary sex characteristic or a sex-influencedcharacteristic. It was beyond the scope of thisinvestigation to determine the genetic systemcontrolling the scale pattern of the seventhtergite in particular; it is not known if suchcontrol is mediated by a number of genes or bya single gene as has been shown for tergite I5(VandeHey et al. 1978)
The present system of Ae. aegypti taxonomyproposed by Mattingly (1957) separates thespecies into two types: mosquitoes withoutwhite scales on the first tergite, and those withwhite scales on at least the first tergite. Aspreviously noted, the scaling of this tergite iscontrolled by the B/ locus. The great variabilityof abdominal tergal scaling led McClelland(1967) to suggest that the species be inter-preted as a polymorphic rather than a polytypicone. McClelland's suggestion was further sup-ported when he observed that a continuousrange of abdominal tergal scale pattern existedin some populations of Ae. aegpti (McClelland1974). Scott and McClelland (1977) proposed amodel to explain polymorphism in Ae. aegypti.Their model included an indoor ecotvpe andan ourdoor ecotype, and three possibl i habi-tats: human, peridomestic and natural. Theindoor ecotype consisted of mosquitoes withincreased white scaling of abdominal tergites,while mosquitoes with predominantly darkscaling on abdominal tergites comprised theoutdoor ecotype. The model indicated thatpolymorphism of Ae. aeglpti in East Africa wasa result of three factors: (l) the presence of adry season during which breeding occurs onlyin the human habitat; (2) greater fitness of theindoor ecotype in the human habitat and of theoutdoor ecotype in the natural habitat; and (3)less than random movement between humanand natural habitats (Scott and McClelland1977). Paterson et al. (1976) proposed that sincethe indoor and outdoor ecotypes existsympatrically and yet remain distinct types,they must be different species. Scott andMcClelland (1977) disagreed with Paterson etal. (1976) on this point. Furthermore, there hasbeen little or no evidence of reproductiveisolation in subspecies of Ae. aegypti (Hartbergand Craig 1968, McClelland 1967, Scott andMcClel land 1975).
Under the present system of Ae. aegyptitaxonomy, taxonomic characters can be alteredby single genes such as black tergitea or byexpression of alleles at one or two loci whichdetermine lightness according to the polygenicsystem proposed herein. These facts seem to
indicate that reevaluation of the bases for Ae.aegypti taxonomy is needed. It is not disputedthat indoor and outdoor ecotypes existsympatrically, and also as distinct allopatricpopulations. However, it appears that toomuch emphasis has been placed on abdominaltergal scaling patterns as the taxonomic crite-rion. Regarding the presently accepted systemof classification of Ae. aegypti based, on Matt-ingly (1957), Craig and Hickey (1967) state thatit is significant that taxonomic characters canbe altered by single genes. Results of thepresent study support the above statement, asabdominal tergal scale pattern can be influ-enced greatly by any one of the three geneticloci proposed in the genetic model, or probablyeven bv anv one or more of the several listedpossible modifiers. Abdominal tergal scalepattern may be useful as a tool in distinguish-ing populations, but may not be valid as theexclusive taxonomic basis for classification ofthe species.
References Cited
Connal, S. L. M. S. 1927. On the variations occurringin Aedzs argenteus, Pioret, in Lagos, Nigeria. Bull.Entomol. Res. l8:5-12.
Craig, G. 8., Jr. and W. A. Hickey. 1967. Genetics ofAedes aegypti, pp. 67-131. In: J. Wright and R. Pal(Eds.). Genetics of insect vectors of disease.Elsevier Press, Amsterdam.
Craig, G. B., Jr. and R. C. VandeHey. 1962. Geneticvariability in Aedes aegpti (Diprera: Culicidae). I.Mutations affecting color pattern. Ann. Entomol.Soc. Am. 55:58-69.
Hartberg, W. K. 1969. Genetical assessment oftaxonomic characters of Aedes aegypti (L.) inTanzania. WHO/VBC/69. I 52. Mimeographed doc-ument, World Health Organization, Geneva.
Hartberg, W. K. and G. B. Craig, Jr. 1968.Reproductive isolation in Stegornyia mosquitoes. I.Sexual isolation between Aedes aegypti and ,4.mascarensis. Ann. Entomol. Soc. Am. 6 I : 865-870.
Johnston, K. W. and W. K. Hartberg. 1981.Polygenic inheritance of variation in scale patternof the abdominal sterna of Eretmapod,ites quinqueuit-tatus (Diotera: Culicidae). T. Med. Entomol.I 8:300-3 i2.
Lewis, D. J. 1945. Observations on the distributionand taxonomy of Culicidae (Diptera) in the Sudan.Trans. R. Entomol. Soc. Lond. 95:I-24.
Mather, K. 1942. The balance of polygenic combina-t ion. J. Genet. 43:309-336.
Mattingly, P. F. 1957. Genetical aspects of the Aedesaegypti problem. I. Taxonomy and bionomics. Ann.Troo. Med. Parasitol. 5l:392-408.
McCleiland, G. A. H. 1960. A preliminary study onthe genetics of abdominal colour variation in Aefusaegypti (L.) (Diptera: Culicidae). Ann. Trop. Med.Parasitol. 54:305-320.
McClelland, G. A. H. 1967. Speciation and evolutionin Aedes, pp. 277-311. 1n J. Wright and R. Pal

502 JounNer- oF THE ArrlnnrcaN Mosqurro Cournor AssocrarroN Vor, . 2 , No.4
(Eds.). Genetics of insect vectors of disease.Elsevier Press, Amsterdam.
McClelland, G. A. H. 1974. A worldwide survev ofvariation in scale parrern of rhe abdominal tergumot Aedes aegJpti (L.) (Diprera: Culicidae). Trani. R.Ent. Soc. Lond. 126:239-259.
Mogi, M., T. Odazawa, I. Miyagi and L. A. DelasLlagas. 1984. Variation in abdominal color pattern
T ..giglt populations of Aedcs aegypti from thePhilippines. Mosq. News 44:604b.-"
Paterson, H. E., C. A. Green and R. J. Mahon. 1976.Aed,es aegypti complex in Africa. (Letter). Nature259:252.
Scott, J. A. and c. A. H. McClelland. 1975.Electrophoretic differences between symparricecotypes. Nature 256:405-406.
Scott, J. A. and G. A. H. McClelland,. 1977. A modelof polymorphism with several seasons and several
habitats, and its application to the mosquito Aadesaeg)pti. Theor. Pop. Biol. | | :242-Zbb.
Thompson, J. N., j . . 1g75. euanritat ive variat ionand gene number. Nature 258:665-66g.
Thompson, J. N., Jr. 1977. Analysis of gene numberand development in polygenic systemi, pp.63-g2./n: Stadler symposium, Vol. 9. University ofMissouri.
Thompson, .J. N., J.. 1979. Synthesis: polygenicvariat ion in perspective, pp.2g5-301. fr , ' I . N.Thompson, Jr. and J. M. Thoday, (Eds.), erianti-tative genetic variation. Academic press, Incl, NewYork.
VandeHey, R. C., M. G. Leahy and K. S. Booth.1978. Analysis of color variat ion in feral.peridomestic, and domestic populations of Aedzsaegypti (L.) (Diptera: Culicidae). Bull. Entomol.Res. 68:443-453.
THE SOUTH COOK COUNTY MOSOU|TO naniElrEr.rr olSrnrcr155th Street and Dixle Hlghway
P.O. Box 1030, Harvey, lllinois 60426Board of Trustees
LawnsNce P. Gutorra-presidentGeoncs J. CULLEN-JecretaryCr-nnrxcr Bosse-Vrce presidentFneo M,csser---Tre as ure rTupooone Coox-Vice presidentKurrrl K. Lrev, Ph.D.-Manager-Entomologist
The Distrlct has scrved South Cook County Illinois since 1954..





























