Embed Size (px)
Citation preview

Plant Cell Reports (1989) 8:399-402 Plant Cell Reports © Springer-Verlag 1989
A microtest system for the serial assay of phytotoxic compounds using photoautotrophic cell suspension cultures of Chenopodium rubrum
Jutta Thiemann, Arnold Nieswandt, and Wolfgang Barz
Lehrstuhl ffir Biochemie der Pflanzen, Westf/ilische Wilhelms-Universit/it, D-4400 Mtinster, Federal Republic of Germany
Received August 11, 1989 - Communicated by E. W. Weiler
ABSTRACT
Chenopodium rubrum photoautotrophic ce l l suspensions were grown In ~ c t issue cul ture dishes under pho- toautotrophic condi t ions. Growth was monitored by measuring cel l number, packed ce l l volume, chlorophyl l content and oxygen production. Such mic ro t i te r dishes are su i tab le systems for the ser ia l assay of growth i n h i b i t i o n and various physiological ef fects ( i . e . chlorophyl l f luorescence, cel l v i a b i l i t y , oxygen pro- duction) of photoautotrophic ce l ls as caused by her- bicides and fungal phytotoxins. The a p p l i c a b i l i t y of the test system is discussed.
Abbreviations
pcv = packed cel l volume; f r .w . = fresh weight; rpm = revol , per minute; DMSO = dimethyl su l fox ide; PMS = phenazine methosulfate; NBT = n i t ro -b lue te t ra - zolium chlor ide;
INTRODUCTION Heterotrophic plant ce l l cul tures are sui table sys-
tems for tes t ing the b io log ica l a c t i v i t y and the cel- l u l a r metabolism of nu t r ien ts , tox ins, or environmen- tal and agrochemical compounds (Gressel 1984; Gross- mann et a l . 1982; Mumma and Davidonis 1983; Sandermann et a l . 1984). Several assay systems have been devised al lowing the ser ia l monitoring of numerous compounds or d i f f e ren t concentrations in small ce l l cu l ture vol- umes in a short period of time (Potrykus et ai.1979; Grossmann 1988).
Photoautotrophic plant ce l l suspension cul tures which great ly resemble mesophyll ce l ls (HUsemann 1985) can well be used for the assay of photosynthesis- inhi - b i t i ng herbicides (Sato et a l . 1987). Such ce l l s are rou t ine ly grown in Erlenmeyer or double- t ier f lasks (HUsemann and Barz !977) so that ser ia l assays require considerable e f f o r t in time, labou~and shaker space. To fu r ther promote the a p p l i c a b i l i t y of these green ce l ls for the ser ia l determination of phytotoxic com- pounds we have now established a microtest system using Chenopodium rubrum photoautotrophic ce l l sus- pensions kept in transparent m ic ro t i te r dishes. Our invest igat ions were designed
I ) to demonstrate the e f f i c i e n t growth of the pho- toautotrophic ce l ls in small volumes as monitored by cel l number, pcv, chlorophyl l content, and oxygen production, and
2) to measure the i n h i b i t o r y ef fects of various her- bicides and a fungal phytotoxin on c e l l u l a r growth and several physiological parameters of the green ce l l s .
Offprint requests to." W. Barz
F ig . l : Photograph of transparent m ic ro t i te r dishes used as a microtest system for photoautotrophic growth of Chenopodium rubrum cel l suspensions; dishes are shown withouTTTd
MATERIAL AND METHODS
Cell cu l ture. Photoautotrophic Chenopodium rubrum cel l suspension cul tures (HUsemann and Barz 1977) were used at the ear ly stage of s tat ionary growth. Cells (100 mg f r .w . x 1.5 ml - I ) were suspended in fresh cul ture medium.
Loading of m ic ro t i te r dishes. Plast ic m ic ro t i te r dishes (8.5 x 12.5 cm, Greiner, NUrtingen, FRG) con- ta in ing 24 wells with 3 ml volume each were loaded under aseptic condit ions with 1.5 ml ce l l suspension per we l l . Pr ior to the appl icat ion of ce l ls to the dishes the cel l suspensions were mixed with the herbi- cides and fusar ic acid which were applied in s t e r i l e - f i l t e r e d methanol solut ions so that the organic sol- vent in the ce l l cul ture medium was set at 0.25 % (v /v) . The dishes fu r ther contain 15 addit ional com- partments (volume I ml) which were each f i l l e d with 750 pl K2CO:/KHCO~-buffer generating a 2% CO 9 par t ia l pressure (H~seman~ and Barz 1977) in the gas"sphere of the dishes. Four such m ic ro t i t e r dishes t i g h t l y sealed with parafi lm were incubated on a MTS4 shaker (Jahnke und Kunkel, Staufen, FRG) at 350 rpm under continuous white l i g h t (80pE.m-2.s - I ) at 25oc.
400
Determination of growth parameters. For the determina- t ion of ce l lu la r growth in the micro t i te r dishes the cel l suspensions from an appropriate number of wells were removed under s t e r i l e conditions a f te r various time in terva ls . For each value the cel ls of four wells were separately measured and the standard deviation calculated.
PCV was determined by centr i fugat ion (1400 rpm/10 minT-Sigma 2KD-Centrifuge) of ce l ls in cal ibrated, conical special tubes (volume 2 ml, Fa. Schlee, Witten, FRG).
Cell number was determined by resuspending the cel ls from the pcv assays and a 200 pl a l iquot was with- drawn. The cel ls were mixed with 4.8 ml chromic acid (10% v /v ) , incubated at 70°C for 10 min and counted in a Fuchs-Rosenthal chamber under a microscope.
Chlorophyll content was measured photometrically ~ iegler and Egle 1965) using acetone extracts of the residual cel l suspensions from the pcv assays.
Production of oxygen of the cel ls was assayed with a Hansatech (Bachofer, Reutlingen, FRG) oxygen meter equipped with a Clark electrode. After a short prein- cubation period and determination of the dark respira- t ion the net oxygen production was measured using the cel ls from one wel l . Light in tens i ty was set at 1050 pE x m "2 x sec - I .
Assay of herbicide tolerance. The inh ib i to ry ef fect of herbicides on the photoautotrophic ce l ls was measured af ter 8 days of incubation. For each herbicide concen- t ra t ion one dish was used; the cel ls from 12 wells were assayed for measuring pcv and chlorophyll content. The cel ls from the other 12 wells were separately taken for recording oxygen production, chlorophyll fluorescence and cel l v i a b i l i t y ; the standard devia- tions were calculated.
Chlorophyll fluorescence was recorded using the same cel l batch as used for the measurements of oxygen pro- duction. White l i gh t was f i l t e r e d through a Corning 4-96 f i l t e r to give blue act in ic l i gh t . The f i l t e r (Wratten 88A) used in the detector probe transmits red l i gh t (X>700 nm). The cel ls were kept in darkness for 5 min, i l luminated for I min and during th is time pe- riod fluorescence in tens i ty was recorded. The i n i t i a l maximum signal (Fp) and the terminal signal (FT) of fluorescence were used to calculate the Rfd-Values ac- cording to the equation Rfd =(Fp-FT~F T.
Cell v i a b i l i t y is represented by the a b i l i t y of ce l ls to generate reducing equivalents as measured by the reduction of NBT. I ml cel l suspension from the determination of the oxygen production was thoroughly mixed with 1500 pl ethylacetate, 10 pl DMSO and 200 pl test reagent (I mM NAD +, I mM PMS, I mM NBT, 50 mM TRIS-HCl, pH 7.5) on a vibromixer for 30 sec. After I hour at 20°C the v io le t ethylacetate phase was pho- tometr ica l ly assayed at 520 nm against an ethylacetate blank. This procedure had previously been checked for l i n e a r i t y of pigment production with d i f fe ren t concen- t rat ions of NADH from a standard solution (data not shown).
Chemicals. Herbicides were obtained from Bayer AG (Leverkusen), fusar ic acid came from Sigma Corp., and NBT and PMS were purchased from Serva.
RESULTS
Growth of photoautotrophic cel ls in microt i ter dishes The growth of photoautotrophic Chenopodium rubrum
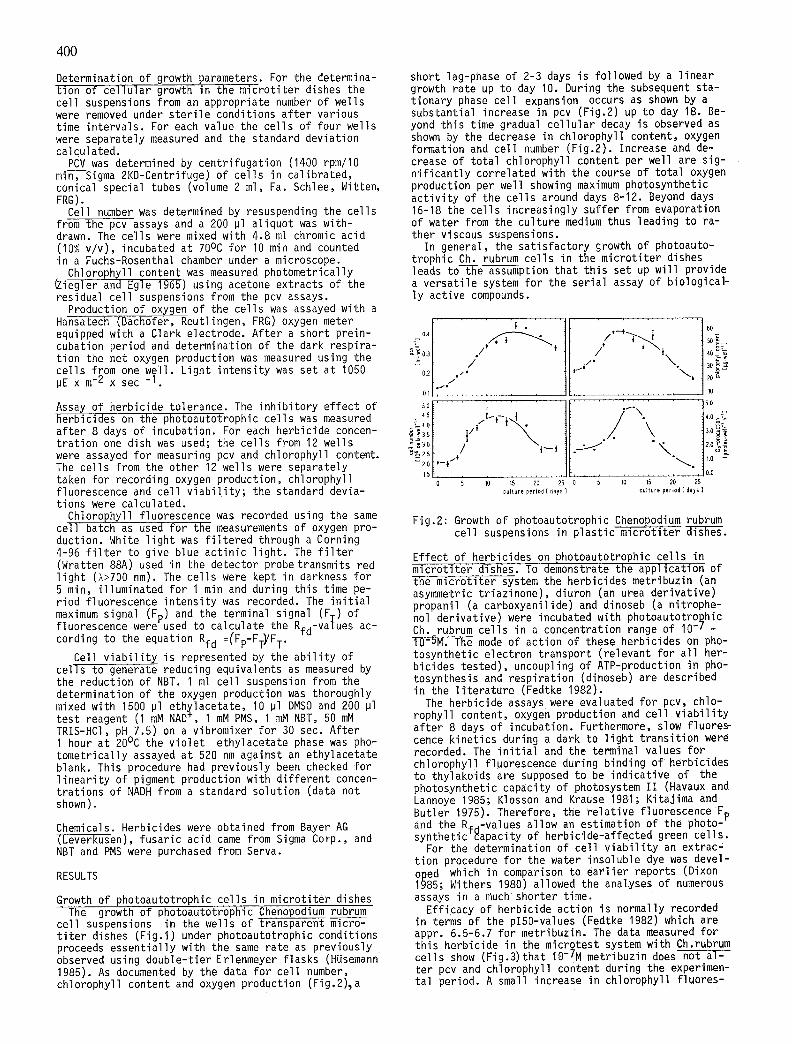
cel l suspensions in the wells of transparent mcro- t i t e r dishes (F ig . l ) under photoautotrophic conditions proceeds essent ia l ly with the same rate as previously observed using double-t ier Erlenmeyer f lasks (HUsemann 1985). As documented by the data for cel l number, chlorophyll content and oxygen production (Fig.2)~a
short lag-phase of 2-3 days is followed by a l inear growth rate up to day 10. During the subsequent sta- t ionary phase cel l expansion occurs as shown by a substantial increase in pcv (Fig.2) up to day 18. Be- yond th is time gradual ce l lu la r decay is observed as shown by the decrease in chlorophyll content, oxygen formation and cel l number (Fig.2). Increase and de- crease of to ta l chlorophyll content per well are sig- n i f i can t l y correlated with the course of tota l oxygen production per well showing maximum photosynthetic a c t i v i t y of the cel ls around days 8-12. Beyond days 16-18 the cel ls increasingly suffer from evaporation of water from the culture medium thus leading to ra- ther viscous suspensions.
In general, the sat is factory growth of photoauto- trophic Ch. rubrum cel ls in the micro t i te r dishes leads to the assumption that th is set up w i l l provide a versat i le system for the ser ia l assay of biological- l y active compounds.
i . . . . . . . I I ' ,_; i I,o
0.2 / = i I i "~ i 70 ~
. . . . . . . . tl . . . . . . . .
'° 1I . . . . . . r ° C'Jo[ -'f'*-i~{i. ""\. ,.o =7
' " ° ' - - - - ' i f " " " I " I.Sf . . . . . . tt . . . . . . . . . . . . . 10.0 o s )'o ",'~ ~o 25 o s ,o ,s 20 2s
cullure period [ days ] culture period [ dais
Fig.2: Growth of photoautotrophic Chenopodium rubrum cel l suspensions in p las t i c m i c r o t i t e r ~ .
Effect of herbicides on photoautotrophic ce l ls in micro t i te r dishes. To demonstrate the appl icat ion of the microt i ter system the herbicides metribuzin (an asymmetric t r iaz inone), diuron (an urea der ivat ive) propanil (a carboxyanil ide) and dinoseb (a nitrophe- nol der ivat ive) were incubated with photoautotro~hic Ch. rubrum cel ls in a concentration range of I0 - I - 10-5M. The mode of action of these herbicides on pho- tosynthetic electron transport (relevant for a l l her- bicides tested), uncoupling of ATP-production in pho- tosynthesis and respirat ion (dinoseb) are described in the l i t e ra tu re (Fedtke 1982).
The herbicide assayS were evaluated for pcv, chlo- rophyll content, oxygen production and cel l v i a b i l i t y af ter 8 days of incubation. Furthermore, slow f luore~ cence kinet ics during a dark to l i gh t t rans i t ion were recorded. The i n i t i a l and the terminal values for chlorophyll fluorescence during binding of herbicides to thylakoids are supposed to be indicat ive of the photosynthetic capacity of photosystem I I (Havaux and Lannoye i985; Klosson and Krause 1981; Kitajima and Butler 1975). Therefore, the re la t i ve fluorescence Fp and the Rfd-Values allow an estimation of the photo-" synthetic capacity of herbicide-affected green ce l ls .
For the determination of cel l v i a b i l i t y an extrac" t ion procedure for the water insoluble dye was devel- oped which in comparison to ear l i e r reports (Dixon 1985; Withers 1980) allowed the analyses of numerous assays in a much shorter time.
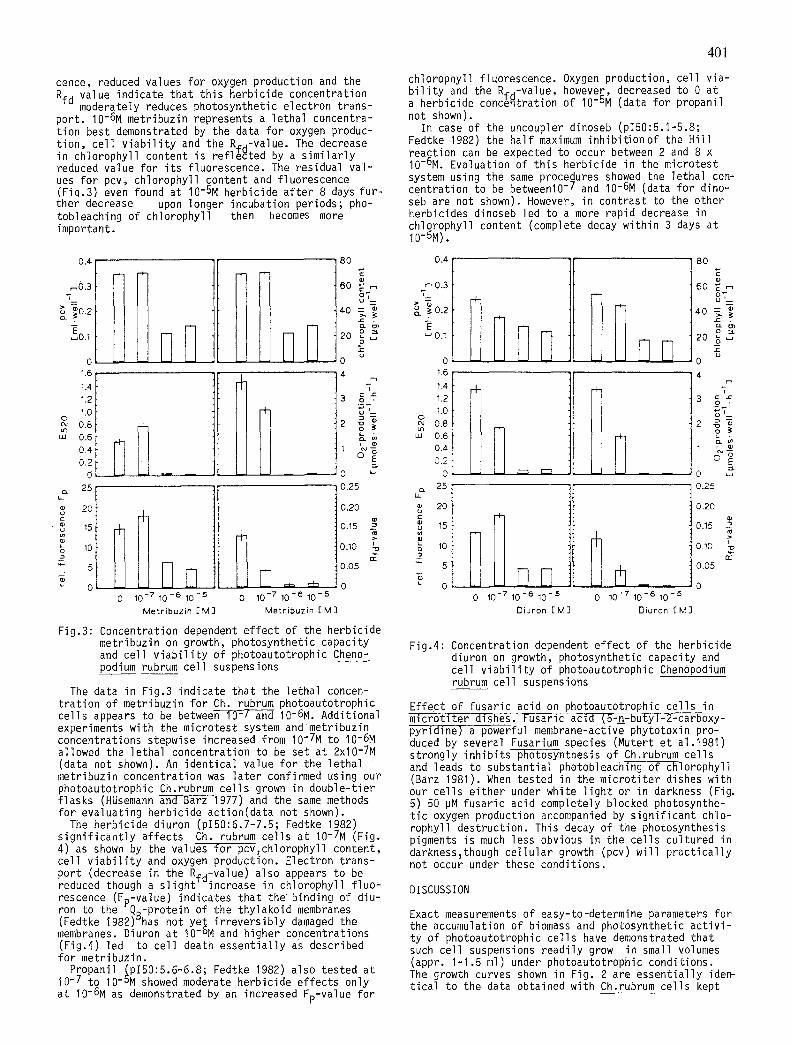
Eff icacy of herbicide action is normally recorded in terms of the pl50-values (Fedtke 1982) which are appr. 6.5-6.7 for metribuzin. The data measured for th is herbicide in the microtest system with Ch.rubrum cel ls show (Fig.3)that 10-7M metribuzin does not a l - ter pcv and chlorophyll content during the experimen- ta l period. A small increase in chlorophyll f luores-
401
cence, reduced values for oxygen production and the Rfd value indicate that th is herbicide concentration
moderately reduces photosynthetic electron trans- port. I0-6M metribuzin represents a lethal concentra- t ion best demonstrated by the data for oxygen produc- t ion, cel l v i a b i l i t y and the R#d-Value. The decrease in chlorophyll content is ref l~cted by a s im i l a r l y reduced value for i t s fluorescence. The residual va l - ues for pcv, chlorophyll content and fluorescence (Fiq.3) even found at I0-5M herbicide a f ter 8 days fu r - ther decrease upon longer incubation periods; pho- tobleaching of chlorophyll then becomes more important.
chlorophyll fluorescence. Oxygen production, cel l v ia- b i l i t y and the Rfa-value, however, decreased to 0 at a herbicide conc6~tration of I0-5M (data for propanil not shown).
In case of the uncoupler dinoseb (p150:5.1-5.8; Fedtke 1982) the hal f maximum inhibi t ion of the H i l l reaction can be expected to occur between 2 and 8 x I0-6M. Evaluation of th is herbicide in the microtest system using the same procedures showed the lethal con- centration to be between10 -7 and I0-6M (data for dino- seb are not shown). However, in contrast to the other herbicides dinoseb led to a more rapid decrease in chlorophyll content (complete decay within 3 days at I0-5M).
0 . 4
,-,0.3 7
o.2
~0.1
o 1.6 1.4 1.2 1.0 o
cu 0.8 ~n uJ 0.6
0.4 0.2
25
20
15
10
~- 5 _j, e~
11 IItW
[ fl
/
0 10 -7 10 - 6 10 - 5
M e t r i b u z i n [ M ]
80 0.4
60 ~ ~ ~ 0.3
4-0 ..~>,-~. ~ ..:-~ 0.2 c~ E
o -~ ~ 0 . 1 20 ~
o o 4 1.6 r-1
1.4
3 g =, 1.2
0 ~q
0.4i
0.25 25
0.20 g 20
o.15 ~ ~ 15
O.lO % ~ lO m
0.05 5
o } o 0 10 - 7 10 - 6 10 - 5
M e t r i b u z i n [ M ]
Fig.3: Concentration dependent e f fect of the herbicide metribuzin on growth, photosynthetic capacity and cel l v i a b i l i t y of photoautotrophic Cheno- podium rubrum cel l suspensions
The data in Fig.3 indicate that the lethal concen- t ra t ion of metribuzin for Ch rubrum photoautotrophic cel ls appears to be between i0-7 and I0-6M. Additional experiments with the microtest system and metribuzin concentrations stepwise increased from I0-7M to I0-6M allowed the lethal concentration to be set at 2x10-7M (data not shown). An ident ical value for the lethal metribuzin concentration was la ter confirmed using our photoautotrophic Ch.rubrum ce l ls grown in double-t ier f lasks (HUsemann and Barz 1977) and the same methods for evaluating herbicide action(data not shown).
The herbicide diuron (p150:6.7-7.5; Fedtke 1982) s i gn i f i can t l y affects Ch. rubrum cel ls at I0-7M (Fig. 4) as shown by the values for pcv, chlorophyll content, cel l v i a b i l i t y and oxygen production. Electron trans- port (decrease in the Rfd-Value) also appears to be reduced though a s l i gh t increase in chlorophyll f luo- rescence (Fp-value) indicates that the binding of diu- ron to the QB-protein of the thylakoid membranes (Fedtke 1982) has not yet i r reve rs ib l y damaged the membranes. Diuron at IO-DM and higher concentrations (Fig.4) led to cel l death essent ia l ly as described for metribuzin.
Propanil ~p150:5.6-6.8; Fedtke 1982) also tested at 10 -7 to 10-bM showed moderate herbicide ef fects only at I0-6M as demonstrated by an increased Fp-value for
DD
I
0 10 - 7 10 - 6 10 - 5 0 10 - 7 10 - 6 10 - 5
D i u r o n [ M ] D i u r o n [ M ]
8o
60 ~ o,-_
40 =~ 2 . X ~
20 g ~
u o 4
m i
3 c ..~ o ,
- 0 ~
0.25
0.20
o.15 ? O.lO ~=
0.05
0
Fig.4: Concentration dependent ef fect of the herbicide diuron on growth, photosynthetic capacity and cel l v i a b i l i t y of photoautotrophic Chenopodium rubrum cel l suspensions
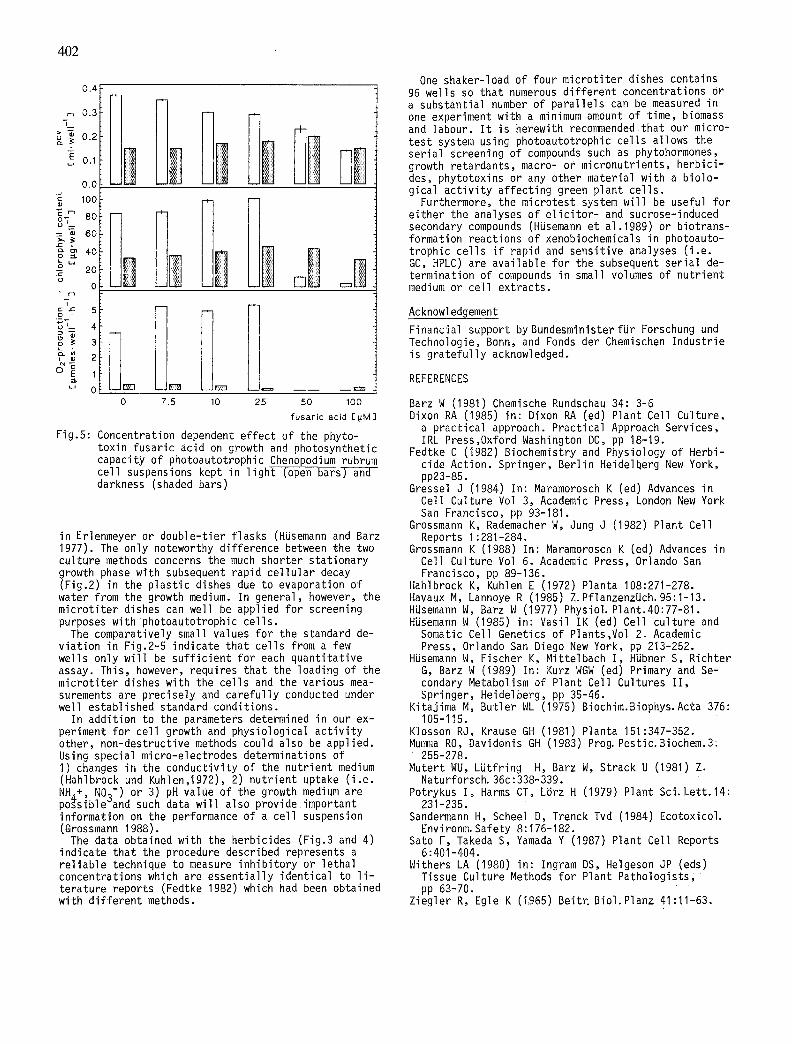
Effect of fusaric acid on photoautotrophic ce l ls in micro t i te r dishes. Fusaric acid (5-~-butyl-2-carboxy- pyridine) a powerful membrane-active phytotoxin pro- duced by several Fusarium species (Mutert et alo1981) strongly inh ib i ts photosynthesis of Ch.rubrum cel ls and leads to substantial photobleaching of chlorophyll (Barz 1981). When tested in the micro t i te r dishes with our ce l ls e i ther under white l i gh t or in darkness (Fig. 5) 50 pM fusar ic acid completely blocked photosynthe- t i c oxygen production accompanied by s ign i f i can t chlo- rophyll destruction. This decay of the photosynthesis pigments is much less obvious in the cel ls cultured in darkness~though ce l l u la r growth (pcv) w i l l p rac t i ca l l y not occur under these conditions.
DISCUSSION
Exact measurements of easy-to-determine parameters for the accumulation of biomass and photosynthetic a c t i v i - ty of photoautotrophic ce l ls have demonstrated that such cel l suspensions readi ly grow in small volumes (appr. I -1.5 ml) under photoautotrophic conditions. The growth curves shown in Fig. 2 are essent ia l ly idem t ica l to the data obtained with Ch,rubrum cel ls kept
402
0 . 4
0 . 3 "T
,E, 0.1
0.0 ¸
1oo C m oo,-__ 80i = ~ 60
o _ ~ 4 0
0 - 2 0 . c 0
0
I
~ f s gL_ 4
0
~ 0 1o Io I 0 7.5 10 25 50 100
fusar i c acid E~M]
Fig.5: Concentration dependent ef fect of the phyto- toxin fusaric acid on growth and photosynthetic capacity of photoautotrophic Chenopodium rubrum cel l suspensions kept in l i gh t (open barsT-a-~-- darkness (shaded bars)
in Erlenmeyer or double-t ier f lasks (HUsemann and Barz 1977). The only noteworthy difference between the two culture methods concerns the much shorter stat ionary growth phase with subsequent rapid ce l l u la r decay (Fig.2) in the p las t ic dishes due to evaporation of water from the growth medium. In general, however, the micro t i te r dishes can well be applied for screening purposes with photoautotrophic ce l ls .
The comparatively small values for the standard de- v ia t ion in Fig.2-5 indicate that ce l ls from a few wells only w i l l be su f f i c ien t for each quant i tat ive assay. This, however, requires that the loading of the micro t i te r dishes with the cel ls and the various mea- surements are precisely and carefu l ly conducted under well established standard conditions.
In addition to the parameters determined in our ex- periment for cel l growth and physiological a c t i v i t y other, non-destructive methods could also be applied. Using special micro-electrodes determinations of I ) changes in the conductivi ty of the nutr ient medium (Hahlbrock und Kuhlen,1972), 2) nutr ient uptake ( i . e . NH~+, NO3-) or 3) pH value of the growth medium are possible and such data w i l l also provide important information on the performance of a cel l suspension (Grossmann 1988).
The data obtained with the herbicides (Fig.3 and 4) indicate that the procedure described represents a re l iab le technique to measure inh ib i to ry or lethal concentrations which are essent ia l ly ident ical to l i - terature reports (Fedtke 1982) which had been obtained with d i f fe ren t methods.
One shaker-load of four mic ro t i te r dishes contains 96 wells so that numerous d i f fe ren t concentrations Or a substantial number of para l le ls can be measured in one experiment with a minimum amount of time, biomass and labour. I t is herewith recommended that our micro- test system using photoautotrophic cel ls allows the ser ia l screening of compounds such as phytohormones, growth retardants, macro- or micronutrients, herbic i - des, phytotoxins or any other material with a biolo- gical a c t i v i t y af fect ing green plant ce l ls .
Furthermore, the microtest system w i l l be useful for e i ther the analyses of e l i c i t o r - and sucrose-induced secondary compounds (HUsemann et ai.1989) or biotrans- formation reactions of xenobiochemicals in photoauto- trophic cel ls i f rapid and sensi t ive analyses ( i . e . GC, HPLC) are avai lable for the subsequent ser ia l de- termination of compounds in small volumes of nutr ient medium or cel l extracts.
Acknowledgement
Financial support byBundesministerfUr Forschung und Technologie, Bonn, and Fonds der Chemischen Industr ie is g ra te fu l l y acknowledged.
REFERENCES
Barz W (1981) Chemische Rundschau 34:3-6 Dixon RA (1985) in: Dixon RA (ed) Plant Cell Culture,
a pract ical approach. Practical Approach Services, IRL Press,Oxford Washington DC, pp 18-19.
Fedtke C (1982) Biochemistry and Physiology of Herbi- cide Action. Springer, Berl in Heidelberg New York, pp23-85.
Gressel J (1984) In: Maramorosch K (ed) Advances in Cell Culture Vol 3, Academic Press, London New York San Francisco, pp 93-181.
Grossmann K, Rademacher W, Jung J (1982) Plant Cell Reports 1:281-284.
Grossmann K (1988) In: Maramorosch K (ed) Advances in Cell Culture Vol 6. Academic Press, Orlando San Francisco, pp 89-136.
Hahlbrock K, Kuhlen E (1972) Planta 108:271-278. Havaux M, Lannoye R (1985) Z. PflanzenzUch. 95:1-13. HUsemann W, Barz W (1977) Physiol. Plant.40:77-81. HUsemann W (1985) in: Vasil IK (ed) Cell culture and
Somatic Cell Genetics of Plants,Vol 2, Academic Press, Orlando San Diego New York, pp 213-252.
HUsemann W, Fischer K, Mittelbach I , Hfibner S, Richter G, Barz W (1989) In: Kurz WGW (ed) Primary and Se- condary Metabolism of Plant Cell Cultures I I , Springer, Heidelberg, pp 35-46.
Kitajima M, Butler WL (1975) Biochim.Biophys. Acta 376: 105-115.
Klosson RJ, Krause GH (1981) Planta 151:347-352. Mumma RO, Davidonis GH (1983) Prog. Pestic, Biochem.3:
255-278. Mutert WU, LUtfring H, Barz W, Strack U (1981) Z.
Naturforsch. 36c:338-339. Potrykus I , Harms CT, L~rz H (1979) Plant Sci. Lett. 14:
231-235. Sandermann H, Scheel D, Trenck Tvd (1984) Ecotoxicol.
Environm. Safety 8:176-182. Sato F, Takeda S, Yamada Y (1987) Plant Cell Reports
6:401-404. Withers LA (1980) in: Ingram DS, Helgeson JP (eds)
Tissue Culture Methods for Plant Patho log is ts , pp 63-70.
Ziegler R, Egle K (1965) Beit~ Biol. Planz 41:11-63.





























