Embed Size (px)
Citation preview

Aglobal response to sulfur starvation inPseudomonasputida andits relationship to the expressionof low-sulfur-content proteinsColin Scott, Margaret E. Hilton, Christopher W. Coppin, Robyn J. Russell, John G. Oakeshott &Tara D. Sutherland
CSIRO, Entomology, Canberra, ACT, Australia
Correspondence: Colin Scott, CSIRO,
Entomology, GPO Box 1700, Canberra, ACT
2601, Australia. Tel.: 161 2 6246 4090;
fax: 161 2 6246 4173;
e-mail: [email protected]
Received 18 October 2006; revised 2 November
2006; accepted 6 November 2006.
First published online 20 December 2006.
DOI:10.1111/j.1574-6968.2006.00575.x
Editor: Christiane Dahl
Keywords
sulfur assimilation; sulfur starvation;
environmental adaptation.
Abstract
Sulfur is essential for life on Earth, but its availability is limited in many
environments. Here the sulfur-starvation response of the model soil bacterium
Pseudomonas putida KT2440 is shown to be associated with an approximately
fivefold reduction in the total soluble thiol content of the cell. A bioinformatic
survey of the P. putida KT2440 genome identified 646 genes encoding proteins
with a significantly lower than average sulfur content (low sulfur-content proteins,
LSPs), the expression of which may have a role in the global reduction of cellular
thiol content during sulfur starvation. Analysis of the genetic organization of the
LSP-encoding genes showed that 31% were potentially transcriptionally associated
with at least one other gene encoding a protein defined as an LSP. In particular, 55
LSP genes were located in three large clusters, termed low-sulfur islands (LSIs)
here. The predicted identities of the proteins encoded by the LSIs strongly suggest
that the LSIs have a role in acquiring sulfur from organic sulfur sources during
sulfur starvation. This hypothesis was supported by transcription fusion studies on
a limited number of LSP promoters under low-sulfur conditions. In a wider survey
of bacterial species, LSIs were found to be more prevalent in free-living, Gram-
negative bacteria than in Gram-positive or obligately intracellular bacteria.
Introduction
Sulfur is ubiquitously required for life, fulfilling a large and
varied number of roles (Beinert, 2000). It is primarily used
as a component of the amino acids cysteine and methionine
as well as of cellular cofactors including biotin, coenzyme A,
s-adenosylmethionine, thiamine, lipoic acid, molybdopter-
ins and iron–sulfur clusters. Sulfur is also critical for
processes such as redox homeostasis via glutathione and
thioredoxin (Ritz & Beckwith, 2001), transcriptional regula-
tion (Green & Paget, 2004) and translation initiation (via
formyl methionine; Laursen et al., 2005). In addition, the
oxidation of sulfur plays a pivotal role in energy transduc-
tion in many eubacteria and archeae (Friedrich et al., 2005).
In general, the preferred source of sulfur is inorganic
sulfate (Kertesz & Weitek, 2001). However, inorganic sulfur
constitutes only a small proportion (3–6%) of total soil
sulfur reserves (Mclaren et al., 1985). The remainder is
largely in the form of carbon-bound sulfur and ester sulfates
(Fitzgerald, 1976; Scherer, 2001). Soil-dwelling bacteria have
evolved a number of systems that allow the use of these
organic sulfur sources (Kertesz, 1999; Mirleau et al., 2005).
Previous attempts to elucidate the proteins or genes in-
volved in these systems have used both proteomic and
transposon-mutagenesis approaches. Although these have
been somewhat successful, many proteins are below the
limits of detection by 2D gel electrophoresis (Beil et al.,
1995) and substrate redundancy has restricted the usefulness
of transposon mutagenesis. For example, the msu, ssu and
tau operons act on an overlapping range of sulfonates (van
der Ploeg et al., 1996; Kertesz et al., 1999; van der Ploeg et al.,
1999; Kahnert et al., 2000). Therefore, there is limited
genetic information regarding many of the systems involved
in the metabolism of organosulfur compounds. For in-
stance, sulfatase activity has been used for many years as a
diagnostic tool for Mycobacterium identification (Mougous
et al., 2002), yet the genes involved are unknown.
Sulfur-acquisition genes in the soil-dwelling bacterium
Pseudomonas putida have been identified either directly or
by homology with known genes from other organisms.
Characterized directly are the ssu operon, which encodes
proteins involved in transport and desulfurization of aryl
sulfates and sulfate esters (Kahnert et al., 2000), and the asf
operon responsible for the import and liberation of sulfur
FEMS Microbiol Lett 267 (2007) 184–193Journal compilation c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Australian government works

from the aryl sulfonates (Vermeij & Kertesz, 1999). Homo-
logs of the Pseudomonas aeruginosa msu (Kertesz et al.,
1999) and Rhodococcus erythropolis dsz operons (Denome
et al., 1994; Piddington et al., 1995) have been detected.
These operons encode monooxygenases and ABC transport
systems that allow the liberation and use of sulfur from
methanesulfonate (msu) and dibenzothiophene (dsz). Also
implicated are the snfECR and snfFG gene products, which
allow the utilization of dimethylsulfone as a sulfur source
(Endoh et al., 2003a, b, 2005), and proteins encoded by the
tau operon, which confers the ability to release sulfur from
taurine, a naturally occurring sulfonate (Eichhorn et al.,
1997; Vermeij et al., 1999).
Here the response of P. putida to sulfur deprivation was
shown to involve a significant reduction in overall thiol
content, possibly involving protein thiols. Along with ob-
servations by other investigators that the proteins required
for sulfur acquisition from nonpreferred sulfur sources often
contain lower than average proportions of sulfur-containing
amino acids (Kertesz, 2001; van der Ploeg et al., 2001), this
result suggests that the sulfur content of gene products could
be used to identify genes and proteins that might be
involved in the sulfur-starvation response in bacteria. The
validation of such an in silico approach in P. putida and the
results in other bacteria from a range of different environ-
ments are described here.
Materials and methods
Bacterial strains, media and plasmids
Pseudomonas putida KT2440 and Escherichia coli JM109
were routinely grown in L medium (Lennox, 1955) at 28
and 37 1C, respectively. Sulfur-free medium (SFM) with
0.2% glucose as carbon source was produced as described
in Sutherland et al. (2000). Media were supplemented with
200 mg mL�1 ampicillin and sodium sulfate as appropriate.
Ion chromatography was used to quantify the residual
sulfur/sulfate content of SFM, and indicated that 4.6 mM
sulfate was present. A Waters 2695 separations module and
Waters IC-Pack Anion HR 4.6� 75 mm column were used
to separate the anions in the medium with a lithium borate/
gluconate mobile phase. Anions were detected using a
Waters 432 conductivity detector. The data were analysed
using Waters EMPOWER 2 chromatography software.
Putative sulfur responsive promoter regions from P.
putida KT2440 (upstream of the PP5106, PP0173, PP3217,
PP2763, PP0169, PP0174, PP3213, PP2762, PP3223, PP5904
and lsfA genes; see below), corresponding to approx. 800 bp
upstream and 200 bp downstream from predicted initiator
methionine codons, were amplified by PCR and fragments
were cloned into the P. putida/E. coli shuttle plasmid pQF50
(Farinha & Kropinski, 1990; kindly provided by Dr Max
Schobert, Technische Universitat Braunschweig) to produce
b-galactosidase reporter plasmids. The lsfA promoter was
also cloned into pQF50 as a lacZ : fusion as a positive
control, as expression of the lsfA gene is known to be up-
regulated by sulfur starvation (Kahnert et al., 2000). Pseu-
domonas putida was transformed by electroporation (Cho
et al., 1995).
b-Galactosidase, protein and thiol assays
The b-galactosidase assays were carried out according to the
method of Miller (1972) with three independent biological
replicates for each sample. Recombinant P. putida cultures
were grown in L medium, and cells were washed in SFM and
used to inoculate (equivalent to 1 : 200 inoculum) SFM
supplemented with 10 mM or 10 mM sodium sulfate. Cul-
tures were grown to an OD600 nm of 0.2–0.3. Cell-free
extracts were obtained by sonication using a Branson
Sonifier 250, and cell debris was removed by centrifugation.
Protein concentrations were estimated using the Bio-Rad
protein assay dye (Bio-Rad). Total soluble cell sulfhydryl
concentrations (including protein thiols, glutathione, cy-
steine, homocysteine and all other free SH-groups) were
estimated according to Thelander (1973).
Bioinformatics
The percentage and distribution of sulfur in proteins
predicted from the genome sequences of the following 31
bacterial species (27 genera and 19 families; GenBank) were
calculated: Agrobacterium tumefaciens str. C58 (Wood et al.,
2001), Bacillus anthracis str. Ames (Read et al., 2003),
Bacillus subtilis ssp. subtilis 168 (Kunst et al., 1997), Bifido-
bacterium longum NCC2705 (Schell et al., 2002), Buchnera
aphidicola str. Bp (Baizongia pistaciae) (Van Ham et al.,
2003), Campylobacter jejuni ssp. jejuni NCTC 11168 (Park-
hill et al., 2000), Chlamydophila pneumoniae AR39 (Read
et al., 2000), Corynebacterium efficiens YS-314 (Nishio et al.,
2003), E. coli O157 : H7 EDL933 (Perna et al., 2001),
Helicobacter hepaticus ATCC 51449 (Suerbaum et al., 2003),
Lactobacillus plantarum WCFS1 (Kleerebezem et al., 2003),
Mycobacterium tuberculosis CDC1551 (Fleischmann et al.,
2002), Mesorhizobium loti (Kaneko et al., 2000), Mycoplasma
genitalium (Fraser et al., 1995), P. aeruginosa PA01 (Stover
et al., 2000), P. putida KT2440 (Nelson et al., 2002),
Pseudomonas syringae pv. tomato str. DC3000 (Buell et al.,
2003), Ralstonia solanacearum (Salanoubat et al., 2002),
Salmonella enterica serovar Typhi (Deng et al., 2003),
Shigella flexneri 2a str. 301 (Jin et al., 2002), Sinorhizobium
meliloti (Galibert et al., 2001), Staphylococcus aureus subsp
aureus MW2 (Baba et al., 2002), Streptomyces avermitilis
MA-4680 (Omura et al., 2001), Tropheryma whipplei str
twist (Raoult et al., 2002), Vibrio parahaemolyticus RIMD
2210633 (Makino et al., 2003), Vibrio vulnificus CMCP6
FEMS Microbiol Lett 267 (2007) 184–193 Journal compilation c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Australian government works
185Sulfur content of Pseudomonas putida proteins

(Kim et al., 2003), Wigglesworthia glossinidia (endosymbiont
of Glossina brevipalpis; Akman et al., 2002), Wolinella
succinogenes (Baar et al., 2003), Xanthomonas campestris pv.
campestris str. ATCC 33913 (de Silva et al., 2002), Xylella
fastidiosa strain Temecula1 (Van Sluys et al., 2003) and
Yersinia pestis KIM (Deng et al., 2002). Analysis of P. putida
gene function was conducted using the comprehensive
microbial resource (CMR) database at the Institute for
Genomic Research (TIGR) (Peterson et al., 2001).
Results and discussion
Pseudomonas putida responds to sulfurstarvation by reducing cellular thiolconcentration
To explore the relationship between sulfur availability and
free-thiol concentration on a whole organism scale, P. putida
was grown in a medium replete or restricted in inorganic
sulfate; the total soluble thiol concentration of each sample
was then determined. The cellular thiol concentration was
corrected for cell density by comparing it with the cellular
protein concentration. The [SH] : [protein] ratio differed by
approximately fivefold in medium that had been supple-
mented with between 1mM and 10 mM sulfate (Fig. 1). This
suggests that part of the response to reduced sulfate avail-
ability involves a reduction in total cellular thiol concentra-
tion. Although changes to the concentrations of other
cellular thiols (e.g. glutathione) undoubtedly influence the
adaptive reduction in total thiol concentration, it seems
likely that the sulfur content of the proteome plays a
significant role in this response to sulfur starvation. Indeed,
it has been observed that proteins involved in the acquisition
and utilization of low-preference sulfur sources generally
contain disproportionately low levels of sulfur-containing
amino acids (Kertesz, 2001; van der Ploeg et al., 2001).
The P. putida genome encodes many low-sulfurproteins
Low-sulfur proteins (LSPs) encoded by P. putida genes were
sought by in silico analysis of the P. putida KT2440 genome.
LSPs were defined as proteins that have a sulfur-containing
amino acid content one SD or more lower than the average
after removal of the initiating methionine (as the vast
majority of encoded proteins contain a formylmethionine
initiation codon, it was considered that it was not subject to
the same selective pressures as noninitiating methionines).
The sulfur-containing amino acid contents of predicted
P. putida KT2440 proteins (of greater than 50 amino acid
residues) are distributed normally about a mean of 3.13%
sulfur-containing amino acids per protein, with a SD of
1.34% (supplementary Appendix S1). Of the 5421 predicted
proteins, 11.9% (646 predicted proteins, fully listed in
Appendix S1) were defined as LSPs.
Over 50% of the LSPs had no known functions (Table 1),
in comparison with 30% for the whole P. putida KT2440
proteome (www.tigr.org). Thirty-four of the LSPs were
among the 88 predicted proteins so far annotated as being
involved in sulfur metabolism. Among these LSPs were the
0
5
10
15
20
25
30
[Sulphate] (M)
[SH
]:[P
rote
in] (
µmol
µg−1
)
10−7 10−6 10−5 10−4 10−3 10−2 10−1
Fig. 1. The sulfhydryl : protein ratio of Pseudomonas putida total soluble
cell-free extract grown in SFM supplemented with 1 mM–10 mM sodium
sulfate. These results are the average of three independent samples;
error bars indicate SDs.
Table 1. Functional classification of LSP genes of Pseudomonas putida
Cellular function
No. of
genes
Putative/unknown function 339 (52.4)
Transport 98 (15.1)
Transcription/transcription regulation 47 (7.3)
Central metabolism/energy
transduction/cofactor biosynthesis
39 (6.0)
Cell envelope 34 (5.3)
DNA/nucleotide metabolism 28 (4.3)
Protein fate 11 (1.7)
Taxis/mobility 11 (1.7)
Cellular processes 16 (2.5)
Amino acid metabolism 10 (1.5)
Protein synthesis 8 (1.2)
Phage 5 (0.8)
Total 646 (99.8)
The percentage of genes in each functional category is indicated in
parentheses. Functional categories are based on the annotations in the
TIGR-CMR database.
FEMS Microbiol Lett 267 (2007) 184–193Journal compilation c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Australian government works
186 C. Scott et al.

products of msu, ssu, tau and dsz gene operons, cystine-
specific ABC transporters, GST transferase, and sulfate and
thiosulfate transporters (Table 2). Interestingly, five of the
predicted LSPs are annotated as uncharacterized monoox-
ygenases (PP1929, 3218, 3219, 3529 and 3573), which could
potentially be involved in currently undescribed organosul-
fur utilization systems.
Known sulfur-metabolizing proteins that are not LSPs
include genes involved in cysteine and methionine biosynth-
esis and several previously characterized sulfatases. Many of
these are expected to have roles outside the sulfur-starvation
response. Biosynthesis of the sulfur-containing amino acids
is not regulated by sulfur abundance per se, but rather by the
abundance of those amino acids (Sekowska et al., 2000).
Sulfatases play many roles aside from scavenging of sulfur
during sulfur starvation. For example, there is a high-sulfur-
content sulfatase from a Pseudomonas species involved in
carbon assimilation from the sulfate ester SDS (Davison
et al., 1992). Sulfatases have also been proposed to act in
regulatory roles. In particular, sulfatases from Rhizobia and
Mycobacteria are thought to be involved in the modulation
of extracellular processes such as cell adhesion and receptor/
ligand binding that mediate host–pathogen interactions
(Mougous et al., 2006).
Although the low cysteine and methionine content of
LSPs is consistent with a role of the proteins during sulfur
starvation, there could be unrelated functional constraints
upon their sulfur contents. For example, a quarter of the
LSPs identified as transporters imported metal ions includ-
ing iron, manganese, nickel, zinc, cadmium and cobalt, and
Table 2. Genes encoding proteins with a known role in sulfur metabolism in Pseudomonas putida identified as LSPs in this study
Gene Encoded protein
PP0169� Dioxygenase, TauD/TfdA family (Pseudomonas entomophila, 98%)
PP0222� Monooxygenase, DszA family (Pseudomonas entomophila, 89%)
PP0223� Monooxygenase, DszC family (Pseudomonas entomophila, 84%)
PP0224� Monooxygenase, DszC family (Pseudomonas fluorescens Pf-5, 78%)
PP0225� Cystine ABC transporter, ATP-binding protein, putative (Pseudomonas entomophila L48, 96%)
PP0227� Cystine ABC transporter, periplasmic cysteine-binding protein, putative (Pseudomonas entomophila L48, 94%)
PP0228� Serine O-acetyltransferase, putative (Pseudomonas syringae pv. phaseolicola 1448A, 89%)
tauD Alpha-ketoglutarate-dependent taurine dioxygenase (P. putida DS1, 98%)
tauC Taurine ABC transporter, permease protein (P. putida DS1, 93%)
tauB Taurine ABC transporter, ATP-binding protein (P. putida DS1, 98%)
tauA Taurine ABC transporter, periplasmic taurine-binding protein (Pseudomonas fluorescens Pf-5, 85%)
lsfA Antioxidant protein LsfA (Pseudomonas entomophila L48, 97%)
ssuA Sulfonate ABC transporter, periplasmic sulfonate-binding protein SsuA (P. putida DS1, 97%)
ssuE FMN reductase (P. putida DS1, 93%)
ssuD Organosulfonate monooxygenase (P. putida DS1, 98%)
ssuC Sulfonate ABC transporter, permease protein SsuC (P. putida DS1, 98%)
ssuB Sulfonate ABC transporter, ATP-binding subunit SsuB (P. putida DS1, 94%)
selB Selenocysteine-specific translation elongation factor (Pseudomonas entomophila L48, 86%)
PP1162� Glutathione S-transferase family protein (Pseudomonas entomophila L48, 87%)
csdA� Cysteine sulfinate desulfinase (Pseudomonas entomophila L48, 89%)
PP2474� Glutathione S-transferase family protein (Erwinia carotovora ssp. atroseptica SCRI1043, 60%)
msuE� NADH-dependent FMN reductase MsuE (P. putida F1, 99%)
msuD� Sulfonate monooxygenase MsuD, putative (P. putida F1, 99%)
PP2933� Glutathione S-transferase family protein (P. putida F1, 98%)
PP3136� Serine O-acetyltransferase (P. putida F1, 98%)
PP3217� Periplasmic aliphatic sulfonate-binding protein, putative
PP3219� Alkansulfonate monooxygenase, putative (Azotobacter vinelandii AvOP, 72%)
PP3228� Periplasmic aliphatic sulfonate-binding protein, putative (Azotobacter vinelandii AvOP, 72%)
PP3229� Periplasmic aliphatic sulfonate-binding protein, putative (Pseudomonas aeruginosa PACS2, 73%)
tpx Thiol peroxidase (P. putida F1, 99%)
PP3637� Sulfonate ABC transporter, ATP-binding protein (P. putida F1, 98%)
PP4305� Periplasmic thiosulfate-binding protein (P. putida F1, 98%)
PP4637� 5-Methyltetrahydropteroyltriglutamate-homocysteine S-methyltransferase family protein (P. putida F1, 96%)
rhdA-2 Thiosulfate sulfurtransferase (P. putida F1, 96%)
serB Phosphoserine phosphatase (P. putida F1, 98%)
PP5118� Thiosulfate sulfurtransferase, putative (P. putida F1, 97%)
cysP Sulfate ABC transporter, periplasmic sulfate-binding protein (P. putida F1, 98%)
�Annotation is based on homology only. The closest homolog identified using a BLAST search, and its identity score are indicated.
FEMS Microbiol Lett 267 (2007) 184–193 Journal compilation c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Australian government works
187Sulfur content of Pseudomonas putida proteins

a low cysteine content may be essential for correctly co-
ordinating the relevant ions. Equally, there are 40 LSP
transcriptional regulators and 10 antioxidant LSPs (the
majority of the 16 LSPs annotated as having a role in
‘cellular processes’; Table 1) that may require a lower than
average cysteine content to prevent miscoordination of
ligands or oxidative inactivation of the proteins. However,
these arguments fail to explain the lower than average
methionine content of these proteins, or the presence of
functionally equivalent proteins in E. coli that do not fulfil
the low sulfur requirements of an LSP (the zinc transporter
ZntA and the peroxide stress regulator OxyR, for example;
Rensing et al., 1997, Paget & Buttner, 2003). This suggests
that many of the LSPs may be expressed during sulfur
starvation, and that the sulfur-starvation response impacts
upon many cellular processes.
Many of the LSPs in P. putida are geneticallyassociated
Overall, 201 LSPs (31%) could potentially be transcribed
with at least one neighboring LSP gene encoded on the same
strand and therefore could belong to the same operon
(Appendix S1). Three larger clusters of genes encoding LSPs
were also found. These three ‘low-sulfur islands’ (LSIs),
defined as runs of at least eight out of 10 genes encoding
proteins with low-sulfur content, account for 8.8% of the P.
putida LSPs (57 genes). The composition of the LSIs is
consistent with a direct involvement in acquisition of sulfur
from nonpreferred sulfur sources. They are comprised of
transporters and catalytic enzymes – 21 of which have
previously been implicated in the assimilation of sulfur from
organosulfate/sulfonates (Table 3). The clustering of genes
encoding proteins involved in the metabolism of low-
preference sulfur sources could facilitate a coordinated
response to sulfur starvation.
LSI1 is homologous to the tauD-ssuF region of P. putida
DS1 which encodes proteins of the tau, dsz and ssu operons
(Endoh et al, 2003a, b). LSI2 encodes proteins with homol-
ogy to the msu operon of P. aeruginosa PAO1 (Kertesz et al.,
1999) in addition to a number of other genes with functions
consistent with organosulfur utilization. LSI3 is yet to be
characterized, but is predicted to encode proteins consistent
with the liberation of sulfur from organic sulfonates, in-
cluding transporter proteins with homology to sulfonate
transporters (PP3217, 3220, 3221, 3222, 3223, 3228 and
3229), a monooxygenase of the NtaA/SnaA/SoxA family
(PP3218), a predicted alkanesulfonate monooxygenase
(PP3219), an acyl-coA dehydrogenase somewhat related to
the P. putida DS1 DszC protein (28%) and a LysR-type
regulator (PP3227). Islands of genes involved in sulfur
metabolism have previously been noted in E. coli (Rocha
et al., 2000), although the proteins encoded by those ‘sulfur
islands’ were not observed to deviate significantly in sulfur
composition from the average for E. coli. It seems likely then
that LSIs are a subset of these sulfur islands that have a
specialized role in obtaining sulfur from nonpreferred
sources.
LSPs are generally expressed in response tosulfur starvation
To test the hypothesis that LSPs identified by the in silico
analysis of the P. putida KT2440 genome were subject to
sulfur-mediated regulation, 10 LSP promoters were selected
randomly (without prior reference to gene names or anno-
tations) and tested for activity during sulfur starvation. The
promoter regions were cloned into the broad-host range
reporter plasmid pQF50 (Farinha & Kropinski, 1990) to
drive expression of b-galactosidase. As a positive control, the
lsfA promoter (Kahnert et al., 2000) was cloned into the
same vector. The activity of the promoters was then assessed
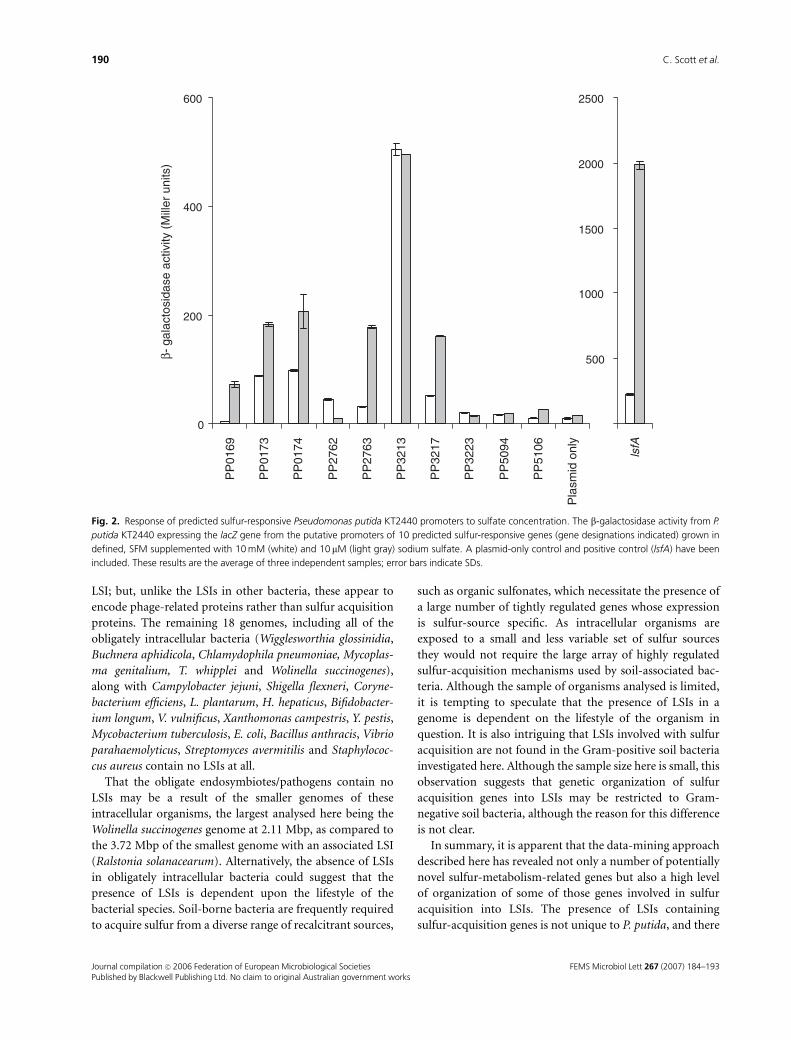
at replete and limited sulfate concentration (Fig. 2). Un-
supplemented SFM was unable to support sufficient growth
of P. putida (data not shown); therefore, SFM was supple-
mented with 10 mM sodium sulfate to produce sulfate-
limited medium and 10 mM for sulfate-replete medium.
Three of the promoters were inactive under the conditions
tested. Of the seven other promoters, six demonstrated a
differential response to the concentration of sulfate, all with
greater activity at the lower sulfate concentration. As ex-
pected, the lsfA promoter also showed such a differential
response dependent upon the sulfate content of the med-
ium.
The promoters that exhibited a sulfate concentration-
dependent response were associated with three hypothetical
proteins (PP5106, PP2763 and PP0174; www.tigr.org), a
putative transcriptional regulator (PP0173), a putative sul-
fonate-binding protein (PP3217) and a predicted TauD-like
dioxygenase (PP0169) (Fig. 2). Only PP3217 and PP0169
have previously been implicated as having a function related
to sulfur metabolism. PP3213, the one which was not sulfate
responsive, was predicted to drive the expression of a
sulfonate-specific ABC-type transport protein.
Two of the six sulfate-responsive promoters were asso-
ciated with LSIs; promoter PP2763 with LSI2 and promoter
PP3217 with LSI3.
LSIs in other species
A survey of LSPs in an additional 30 bacterial species
revealed LSIs: of similar composition, but diverse genetic
organization, in six of the 24 free-living bacterial species
(Agrobacterium tumefaciens, P. aeruginosa, P. syringae, Ral-
stonia solanacearum, Mesorhizobium loti and Sinorhizobium
meliloti; Table 3). Two other genomes from free-living
species (Bacillus subtilis and Salmonella enterica) contain an
FEMS Microbiol Lett 267 (2007) 184–193Journal compilation c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Australian government works
188 C. Scott et al.

Table 3. Organization of LSIs in seven bacterial species
Bacterium LSI
Pseudomonas putida (LSI1)w PP_0220(� , ABC transporter), 0221(� , ABC transporter), 0222(� , DszA�), 0223(� , acyl-coA dehydrogenase�),
0224(� , acyl-coA dehydrogenase�), 0225(� , ABC transporter�), 0226(� , ABC transporter�), 0227(� , ABC
transporter�), 0228(1, serine O-acetyltransferase), 0229(1, transport protein), 0230(� , dioxygenase�), 0231
(� , TauC�), 0232(� , TauB�), 0233(� , TauA�), 0234(1, porin), 0235(1, LsfA), 0236(1, oxidoreductase�), 0237
(1, SsuA�), 0238(1, SsuD�), 0239(1, SsuC�), 0240(1, SsuB�)
Pseudomonas putida (LSI2) PP_2751(1, ABC transporter), 2752(1, ABC transporter), 2753(1, ABC transporter), 2754(1, porin), 2755
(1, acyl-coA dehydrogenase), 2756(1, unknown), 2757(1, ABC transporter), 2758(1, ABC transporter), 2759
(1, ABC transporter), 2760(1, ABC transporter), 2761(1, ABC transporter), 2762(� , acyl-coA dehydrogenase),
2763(1, unknown), 2764(1, MsuE�), 2765(1, MsuD�), 2766(1, long-chain-fatty-acid–CoA ligase), 2767
(1, ABC transporter), 2768(1, ABC transporter), 2769(1, ABC transporter), 2770(1, ABC transporter), 2771
(� , transcriptional regulator), 2772(� , monooxygenase)
Pseudomonas putida (LSI3) PP_3216(� , unkown), 3217(1, periplasmic binding-protein�), 3218(1, monooxygenase�), 3219(� ,
monooxygenase�), 3220(� , ABC transporter), 3221(� , ABC transporter), 3222(� , ABC transporter), 3223
(� , ABC transporter), 3224(� , aldolase), 3225(� , dehydrogenase), 3226(� , acyl-coA dehydrogenase), 3227
(� , transcriptional regulator�), 3228(1, periplasmic-binding protein�), 3229(1, periplasmic-binding protein�)
Pseudomonas aeruginosa PA2594(� , ABC-transpoter�), 2595(� , ABC-transpoter�), 2596(� , ABC-transpoter�), 2597(� , acyl-CoA
dehydrogenase), 2598(� , monooxygenase�), 2599(� , monooxygenase�), 2600(� , unkown�), 2601
(� , transcriptional regulator�), 2602(1, dioxygenase), 2603(1, thiosulfate sulfurtransferase�)
Sinorhizobium meliloti
(plasmid pSymA)
Sma2049(1, transcriptional regulator), 2051(1, lipid desaturase), 2053(1, MocE), 2055(1, unknown), 2067
(� , sulfonate-binding protein), 2069(� , ABC transporter), 2071(� , unknown), 2073(� , acyl-coA dehydrogenase ),
2075(� , unknown), 2077(� , oxidoreductase), 2079(� , ABC transporter), 2081(� , ABC transporter), 2082
(� , ABC transporter�), 2085(� , ABC transporter�), 2087(1, TRAP transporter�), 2089(1, acyl-CoA
dehydrogenase�), 2091(1,acyl-CoA dehydrogenase�), 2093(1, ABC transporter), 2095(1, isoflavone reductase),
2097(1, monooxygenase), 2099(1, NADH-flavin reductase�), 2101(1, monoxygenase)
Mesorhizobium loti Mlr5216(1, MsuD�), 5217(1, unknown), 5217(1, unkown�), 5218(1, monooxygenase), 5219(1, ABC transporter),
5220(1, ABC transporter), 5222(1, ABC transporter), 5223(1, ABC transporter), 5224(1, ABC transporter), 5225
(1, acyl-coA dehydrogenase�), 5227(1, MsuE�)
Ralstonia solanacearum Rsc1336(1, ABC transporter), 1337(1, oxidoreductase), 1338(1, periplasmic-binding protein), 1339(1,
monooxygenase) 1340(1, ABC transporter), 1341(1, ABC transporter), 1342(1, SsuF), 1343(1, unknown), 1344
(1, esterase), 1345(1, ABC transporter), 1346(1, ABC transporter), 1347(1, ABC transporter), 1348(1, cysB)
Pseudomonas syringae PSPTO5178(� , serine O-acetyltransferase), 5179(1, deaminase), 5180(1, ABC transporter�), 5181(1, ABC
transporter), 5182(1, ABC transporter), 5183(1, unknown), 5184(1, acyl-CoA dehydrogenase), 5185(1, acyl-CoA
dehydrogenase�), 5186(1, monooxygenase�), 5187(1, ABC transporter), 5188(1, ABC transporter), P5189(1, ABC
transporter), 5190(1, unknown), 5191(� , RND transporter), 5192(� , RND transporter), 5193(� , RND transporter),
5194(� , unknown), 5195(� , ABC transporter�), 5196(� , ABC transporter�), 5197(� , ABC transporter�), 5918
(� , TauD�)
Agrobacterium tumefaciens Atu3415(� , ABC transporter), 3416(� , ABC transporter), 3417(� , ABC transporter), 3418(� , acetyltransferase),
3419(� , hydrolase), 3420(� , monooyxygenase�), 3421(� , monooxygenase), 3422(1, unknown), 3423(1,
unkown), 3424(� , acyl-coA dehydrogenase ), 3425(� , monooxygenase), 3426(� , SsuD�), 3427(1, unknown),
3428(� , unkown), A3429(1, unknown), 3430(1, SsuD�), 3431(� , acyl-coA dehydrogenase), 3432(� , TauA�),
3433(� , ABC transporter), 3434(1, ABC transporter), 3435(1, ABC transporter), 3436(1, ABC transporter), 3437
(1, ABC transporter), 3438(1, oxidoreductase�), 3439(1, acyl-coA dehydrogenase�), 3440(1, transcriptional
regulator), 3441(� , ABC transporter), 3442(� , ABC transporter), 3443(1, unknown), 3444(� , ABC transporter),
3445(� , hydrolase), 3446(� , monooxygenase), 3447(� , isoflavone reductase), 3448(� , dszB�), 3449
(� , unknown), 3450(1, monooxygenase�), 3451(� , monooxygenase)
Agrobacterium tumefaciens
(plasmid AT)
Atu5518(� , monooxygenase), 5519(1, unkown), 5520(� , acyl-coA dehydrogenase), 5521(� , ABC transporter),
5522(� , transport protein), 5523(� , ABC transporter), 5524(� , ABC transporter), 5525(� , ABC transporter),
5526(� , monooxygenase�), 5527(� , oxidoreductase�)
Agrobacterium tumefaciens
(plasmid Ti)
Atu6077(� , monooxygenase�), 6078(� , monooxygenase�), 6079(� , monooxygenase�), 6080(1, transcriptional
regulator�), 6081(1, unknown), 6082(1, AAA-ATPase), 6083(1, transcriptional regulator�), 6084
(1, monooxygenase�),6085(1, ABC transporter�)
�Gene products with implied function in sulfur assimilation from organosulfate/sulfonates.wHomologous to the tauD-ssuF region of Pseudomonas putida DS1.
The direction of transcription and predicted function of the gene product for each gene are given in parentheses. LSPs are denoted in italics.
FEMS Microbiol Lett 267 (2007) 184–193 Journal compilation c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Australian government works
189Sulfur content of Pseudomonas putida proteins
LSI; but, unlike the LSIs in other bacteria, these appear to
encode phage-related proteins rather than sulfur acquisition
proteins. The remaining 18 genomes, including all of the
obligately intracellular bacteria (Wigglesworthia glossinidia,
Buchnera aphidicola, Chlamydophila pneumoniae, Mycoplas-
ma genitalium, T. whipplei and Wolinella succinogenes),
along with Campylobacter jejuni, Shigella flexneri, Coryne-
bacterium efficiens, L. plantarum, H. hepaticus, Bifidobacter-
ium longum, V. vulnificus, Xanthomonas campestris, Y. pestis,
Mycobacterium tuberculosis, E. coli, Bacillus anthracis, Vibrio
parahaemolyticus, Streptomyces avermitilis and Staphylococ-
cus aureus contain no LSIs at all.
That the obligate endosymbiotes/pathogens contain no
LSIs may be a result of the smaller genomes of these
intracellular organisms, the largest analysed here being the
Wolinella succinogenes genome at 2.11 Mbp, as compared to
the 3.72 Mbp of the smallest genome with an associated LSI
(Ralstonia solanacearum). Alternatively, the absence of LSIs
in obligately intracellular bacteria could suggest that the
presence of LSIs is dependent upon the lifestyle of the
bacterial species. Soil-borne bacteria are frequently required
to acquire sulfur from a diverse range of recalcitrant sources,
such as organic sulfonates, which necessitate the presence of
a large number of tightly regulated genes whose expression
is sulfur-source specific. As intracellular organisms are
exposed to a small and less variable set of sulfur sources
they would not require the large array of highly regulated
sulfur-acquisition mechanisms used by soil-associated bac-
teria. Although the sample of organisms analysed is limited,
it is tempting to speculate that the presence of LSIs in a
genome is dependent on the lifestyle of the organism in
question. It is also intriguing that LSIs involved with sulfur
acquisition are not found in the Gram-positive soil bacteria
investigated here. Although the sample size here is small, this
observation suggests that genetic organization of sulfur
acquisition genes into LSIs may be restricted to Gram-
negative soil bacteria, although the reason for this difference
is not clear.
In summary, it is apparent that the data-mining approach
described here has revealed not only a number of potentially
novel sulfur-metabolism-related genes but also a high level
of organization of some of those genes involved in sulfur
acquisition into LSIs. The presence of LSIs containing
sulfur-acquisition genes is not unique to P. putida, and there
0
500
1000
1500
2000
2500
lsfA
0
200
400
600
PP
0169
PP
0173
PP
0174
PP
2762
PP
2763
PP
3213
PP
3217
PP
3223
PP
5094
PP
5106
Pla
smid
onl
y
β- g
alac
tosi
dase
act
ivity
(M
iller
uni
ts)
Fig. 2. Response of predicted sulfur-responsive Pseudomonas putida KT2440 promoters to sulfate concentration. The b-galactosidase activity from P.
putida KT2440 expressing the lacZ gene from the putative promoters of 10 predicted sulfur-responsive genes (gene designations indicated) grown in
defined, SFM supplemented with 10 mM (white) and 10 mM (light gray) sodium sulfate. A plasmid-only control and positive control (lsfA) have been
included. These results are the average of three independent samples; error bars indicate SDs.
FEMS Microbiol Lett 267 (2007) 184–193Journal compilation c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Australian government works
190 C. Scott et al.

is evidence to suggest that the presence of LSIs in a genome
may depend on the lifestyle and phylogeny of the bacterium
in question.
Acknowledgements
We thank Dr Max Schobert of Institut fuer Mikrobiologie at
the Technische Universitaet, Braunschweig for kindly pro-
viding the pQF50 reporter plasmid and Dr Anthony Herlt of
the Research School of Chemistry at the Australian National
University, Canberra for his help with the ion chromatogra-
phy of the SFM.
References
Akman L, Yamashita A, Watanabe H et al. (2002) Genome
sequence of the endocellular obligate symbiont of tsetse flies,
Wigglesworthia glossinidia. Nature Genet 32: 402–407.
Baar C, Eppinger M, Raddatz G et al. (2003) Complete genome
sequence and analysis of Wolinella succino genes. Proc Natl
Acad Sci USA 100: 11690–11695.
Baba T, Takeuchi F, Kuroda M et al. (2002) Genome and virulence
determinants of high virulence community-acquired MRSA.
Lancet 359: 1819–1827.
Beil S, Kehrli H, James P, Staudenmann W, Cook AM, Leisinger T
& Kertesz MA (1995) Purification and characterization of the
arylsulfatase synthesized by Pseudomonas aeruginosa PAO
during growth in sulfate-free medium and cloning of the
arylsulfatase gene (atsA). Eur J Biochem 229: 385–394.
Beinert H (2000) A tribute to sulfur. Eur J Biochem 267:
5657–5664.
Buell CR, Joardar V, Lindeberg M et al. (2003) The complete
genome sequence of the Arabidopsis and tomato pathogen
Pseudomonas syringae pv. tomato DC3000. Proc Natl Acad Sci
USA 100: 10181–10186.
Cho JH, Kim EK & So JS (1995) Improved transformation of
Pseudomonas putida KT2440 by electroporation. Biotechnol
Tech 9: 41–44.
Davison J, Brunel F, Phanopoulos A, Prozzi D & Terpstra P (1992)
Cloning and sequencing of Pseudomonas genes determining
sodium dodecyl sulfate biodegradation. Gene 114: 19–24.
Deng W, Burland V, Plunkett G III et al. (2002) Genome sequence
of Yersinia pestis KIM. J Bacteriol 184: 4601–4611.
Deng W, Liou SR, Plunkett G III et al. (2003) Comparative
genomics of Salmonella enterica serovar Typhi strains Ty2 and
CT18. J Bacteriol 185: 2330–2337.
Denome SA, Oldfield C, Nash LJ & Young KD (1994)
Characterisation of the desulfurization genes from
Rhodococcus sp. strain IGTS8. J Bacteriol 176: 6707–6716.
de Silva AC, Ferro JA, Reinach FC et al. (2002) Comparison of the
genomes of two Xanthomonas pathogens with differing host
specificities. Nature 417: 459–463.
Eichhorn E, van der Ploeg JR, Kertesz MA & Leisinger T (1997)
Characterization of alpha-ketoglutarate-dependent taurine
dioxygenase from Escherichia coli. J Biol Chem 272:
23031–23036.
Endoh T, Kasuga K, Horinouchi M, Yoshida T, Habe H, Nojiri H
& Omori T (2003a) Characterization and identification of
genes essential for dimethyl sulfide utilization in Pseudomonas
putida strain DS1. Appl Microbiol Biotechnol 62: 83–91.
Endoh T, Habe H, Yoshida T, Noriji H & Omori T (2003b) A
CysB-regulated and s54-dependent regulator, SnfR, is essential
for dimethyl sulfone metabolism of Pseudomonas putida strain
DS1. Microbiology 149: 991–1000.
Endoh T, Habe H, Noriji H, Yamane H & Omori T (2005) The
s54-dependent transcriptional activator SfnR regulates the
expressoin of the Pseudomonas putida sfnFG operon
responsible for dimethyl sulfone utilization. Mol Microbiol 55:
897–911.
Farinha MA & Kropinski AM (1990) Construction of broad-host
range plasmid vectors for easy visible selection and analysis of
promoters. J Bacteriol 172: 3496–3499.
Fitzgerald JW (1976) Sulfate ester formation and hydrolysis: a
potentially important yet often ignored aspect of the sulfur
cycle of aerobic soils. Bacteriol Rev 40: 698–721.
Fleischmann RD, Alland D, Eisen JA et al. (2002) Whole genome
comparison of Mycobacterium tuberculosis clinical and
laboratory strains. J Bacteriol 184: 5479–5490.
Fraser CM, Gocayne JD, White O et al. (1995) The minimal gene
complement of Mycoplasma genitalium. Science 270: 397–403.
Friedrich CG, Bardischewsky F, Rother D, Quentmeier A &
Fischer J (2005) Prokaryotic sulfur oxidation. Curr Opin
Microbiol 8: 253–259.
Galibert F, Finan TM, Long SR et al. (2001) The composite
genome of the legume symbiont Sinorhizobium meliloti.
Science 293: 668–672.
Green J & Paget MS (2004) Bacterial redox sensors. Nat Rev
Microbiol 2: 954–966.
Jin Q, Yuan Z, Xu J et al. (2002) Genome sequence of Shigella
flexneri 2a: insights into pathogenicity through comparison
with genomes of Escherichia coli K12 and O157. Nucleic Acids
Res 30: 4432–4441.
Kahnert A, Vermeij P, Weitek C, James P, Leisinger T & Kertesz
MA (2000) The ssu locus plays a key role in organosulfur
metabolism in Pseudomonas putida S-313. J Bacteriol 182:
2869–2878.
Kaneko T, Nakamura Y, Sato S et al. (2000) Complete genome
sequence of the nitrogen fixing symbiotic bacterium
Mesorhizobium loti. DNA Res 6: 331–338.
Kertesz MA (1999) Riding the sulfur cycle-metabolism of
sulfonates and sulfate esters in gram-negative bacteria. FEMS
Microbiol Rev 24: 135–175.
Kertesz MA, Schmidt-Larbig K & Wuest T (1999) A novel
reduced flavin mononucleotide-dependent methanesulfonate
sulfonatase encoded by the sulfur-regulated msu operon of
Pseudomonas aeruginosa. J Bacteriol 181: 1464–1473.
Kertesz MA (2001) Bacterial transporters for sulfate and
organosulfur compounds. Res Microbiol 152: 279–290.
FEMS Microbiol Lett 267 (2007) 184–193 Journal compilation c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Australian government works
191Sulfur content of Pseudomonas putida proteins

Kertesz MA & Weitek C (2001) Desulfurization and
desulfonation: applications of sulfur-controlled gene
expression in bacteria. Appl Microbiol Biotechnol 57: 460–466.
Kim YR, Lee SE, Kim CM et al. (2003) Characterisation and
pathogenic significance of Vibrio vulnificus antigens
preferentially expressed in sepinemic patients. Infect Immun
71: 5461–5471.
Kleerebezem M, Boekhorst J, van Krannenburg R et al. (2003)
Complete genome sequence of Lactobacillus plantarum
WCFS1. Proc Natl Acad Sci USA 100: 1990–1995.
Kunst F, Ogasawara N, Moszer I et al. (1997) The complete
genome sequence of the Gram-positive bacterium Bacillus
subtilis. Nature 390: 249–256.
Laursen BS, S�rensen HP, Mortensen KK & Sperling-Petersen HU
(2005) Initiation of protein synthesis in bacteria. Microbiol
Mol Biol Rev 69: 101–123.
Lennox ES (1955) Transduction of linked genetic characters by of
host by bacteriophage P1. Virology 1: 190–206.
Makino K, Oshima K, Kurokawa K et al. (2003) Genome
sequence of Vibrio parahaemolyticus: a pathogenic mechanism
distinct from V. cholerae. Lancet 361: 743–749.
McLaren RG, Keer JI & Swift RS (1985) Sulfur transformations in
soils using sulfur-35 labelling. Soil Biol Biochem 17: 73–79.
Miller JH (1972) Experiments in Molecular Genetics, Cold Spring
Harbor Laboratory Press, Cold Spring Harbor, NY.
Mirleau P, Wogelius R, Smith A & Kertesz MA (2005) Importance
of organosulfur utilization for the survival of Pseudomonas
putida in soil and rhizosphere. Appl Environ Microbiol 71:
6571–6577.
Mougous JD, Green RE, Williams SJ, Brenner SE & Bertozzi CR
(2002) Sulfotransferases and sulfatases in Mycobacteria. Chem
Biol 9: 767–776.
Mougous JD, Senaratne RH, Petzold CJ et al. (2006) A sulfated
metabolite produced by stf3 negatively regulates the virulence
of Mycobacterium tuberculosis. Proc Natl Acad Sci USA 103:
4258–4263.
Nelson KE, Weinel C, Paulsen IT et al. (2002) Complete genome
sequence and comparative analysis of the metabolically
versatile Pseudomonas putida KT2440. Environ Microbiol 4:
799–808.
Nishio Y, Nakamura Y, Kawarabayasi Y et al. (2003) Comparative
complete genome sequence analysis of the amino acid
replacements responsible for the thermostability of
Corynebacterium efficiens. Genome Res 13: 1572–1579.
Omura S, Ikeda H, Ishikwa J et al. (2001) Genome sequence of an
industrial microorganism Streptomyced avermitilis: dedcing
the ability of producing secondary metabolites. Proc Natl Acad
Sci USA 98: 12215–12220.
Paget MSB & Buttner MJ (2003) Thiol-based regulatory switches.
Ann Rev Genet 37: 91–121.
Parkhill J, Wren BW, Mungall K et al. (2000) The genome
sequence of the food-borne pathogen Campylobacter jejuni
reveals hypervariable sequences. Nature 403: 665–668.
Perna NT, Plunkett G III, Burland V et al. (2001) Genome
sequence of the enterohaemorrhagic Escherichia coli
O157 : H7. 409: 529–533.
Peterson JD, Umayam LA, Dickinson TM, Hickey EK & White O
(2001) The comprehensive microbial resource. Nucleic Acids
Res 29: 123–125.
Piddington CS, Kovacevich BR & Rambosek J (1995) Sequence
and molecular characterization of a DNA region encoding the
dibenzothiophene desulfurization operon of Rhodococcus sp.
strain IGTS8. Appl Environ Microbiol 61: 468–475.
Raoult D, Audic S, Robert C et al. Tropheryma whipplei illustrates
the diversity of gene loss patterns in small genome bacterial
pathogens. Unpublished GeneBank submission (as of 31 July
2002).
Read TD, Peterson SN, Tourasse N et al. (2003) The genome
sequence of Bacillus anthracis Ames and comparison to closely
related bacteria. Nature 423: 81–86.
Read TD, Brunham RC, Shen C et al. (2000) Genome sequences
of Chlamydia trachomatis MoPn and Chlamydia pneumoniae
AR39. Nucleic Acids Res 28: 1397–1406.
Rensing C, Mitra B & Rosen BP (1997) The zntA gene of
Escherichia coli encodes a Zn(II)-translocating P-typeATPase.
Proc Natl Acad Sci USA 23: 14326–14331.
Ritz D & Beckwith J (2001) Roles of thiol-redox pathways in
bacteria. Ann Rev Microbiol 55: 21–48.
Rocha EPC, Sekowska A & Danchin A (2000) Sulphur islands in
the Escherichia coli genome: markers of the cell’s architecture?
FEBS Lett 476: 8–11.
Salanoubat M, Genin S, Ariguenave F et al. (2002) Genome
sequence of the plant pathogen Ralstonia solanacearum.
Nature 415: 497–502.
Schell MA, Karmirantzou M, Snel B et al. (2002) The genome
sequence of Bifidobacterium longum reflects its adaptation to
the human gastrointestinal tract. Proc Natl Acad Sci USA 99:
14422–14427.
Scherer HW (2001) Sulfur in crop production – invited paper.
Eur J Agro 14: 81–111.
Sekowska A, Kung HF & Danchin A (2000) Sulfur metabolism in
Escherichia coli and related bacteria: facts and fiction. J Mol
Microbiol Biotechnol 2: 145–177.
Stover CK, Pham XQ, Erwin AL et al. (2000) Complete genome
sequence of Pseudomonas aeruginosa PA01, an opportunistic
pathogen. Nature 406: 959–964.
Suerbaum S, Josenhaus C, Sterzbach T et al. (2003) The complete
genome sequence of the carcinogenic bacterium Helicobacter
hepaticus. Proc Natl Acad Sci USA 100: 7901–7906.
Sutherland TD, Horne I, Lacey MJ, Harcourt RL, Russell RJ &
Oakeshott JG (2000) Enrichment of an endosulfan-degrading
mixed bacterial culture. Appl Environ Microbiol 66: 2822–2828.
Thelander L (1973) Physiochemical characterisation of
ribonucleotide diphosphate reductase from Escherichia coli. J
Biol Chem 248: 4591–4601.
van der Ploeg JR, Weiss MA, Saller E, Nashimoto H, Saito N,
Kertesz MA & Leisinger T (1996) Identification of sulfur
starvation-regulated genes in Escherichia coli: a gene cluster
FEMS Microbiol Lett 267 (2007) 184–193Journal compilation c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Australian government works
192 C. Scott et al.

involved in the utilization of taurine as a sulfur source. J
Bacteriol 178: 5438–5446.
van der Ploeg JR, Iwanicka-Nowicka R, Bykowski T, Hryniewicz
MM & Leisinger T (1999) The Escherichia coli ssuEADCB gene
cluster is required for the utilization of sulfur from aliphatic
sulfonates and is regulated by the transcriptional activator Cbl.
J Biol Chem 274: 29358–29365.
van der Ploeg JR, Eichhorn E & Leisinger T (2001) Sulfonate-
sulfur metabolism and its regulation in Escherichia coli. Arch
Microbiol 176: 1–8.
Van Ham RC, Kamerbeek J, Palacios C et al. (2003) Reductive
genome evolution in Buchnera aphidicola. Proc Natl Acad Sci
USA 100: 581–586.
Van Sluys MA, de Oliveira MC, Monteiro-Vitorello CB et al.
(2003) Comparative analyses of the complete genome
sequences of Pierce’s disease and citrus variegated chlorosis
strains of Xylella fastidiosa. J Bacteriol 185: 1018–1026.
Vermeij P & Kertesz MA (1999) Pathways of assimilative sulfur
metabolism in Pseudomonas putida. J Bacteriol 181:
5833–5837.
Vermeij P, Weitek C, Kahnert A, Wuest T & Kertesz MA (1999)
Genetic organization of sulfur-controlled aryl desulfonation in
Pseudomonas putida S-313. Mol Microbiol 32: 913–926.
Wood DW, Setubal JC, Kaul R et al. (2001) The genome of the
natural genetic engineer Agrobacterium tumefaciens C58.
Science 294: 2317–2323.
Supplementarymaterial
The following supplementary material is available for this
article:
Appendix S1. Bioinformatic analysis of the P. putida
KT2440 proteome.
This material is available as part of the online article
from: http://www.blackwell-synergy.com/doi/abs/10.1111/
j.1574.6968.2006.00575.x (This link will take you to the
article abstract).
Please note: Blackwell Publishing are not responsible
for the content or functionality of any supplementary
materials supplied by the authors. Any queries (other than
missing material) should be directed to the corresponding
author for the article.
FEMS Microbiol Lett 267 (2007) 184–193 Journal compilation c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original Australian government works
193Sulfur content of Pseudomonas putida proteins