Embed Size (px)
Citation preview

A ‘genome to paddock’ approach
to control plant disease
Barbara Howlett, School of BioSciences
the University of Melbourne

Previous Research Projects
• B.Sc (Hons) Cytochrome C (Melb Uni)
• Research Assistant: Tapeworms (CSIRO, Melb)
• Research Assistant: Influenza (ANU)
• M.Sc: Self incompatibility in plants (ANU)
• Research Assistant: Bacterial chemotaxis (UC Berkeley)
• Ph.D: Grass pollen allergens (Melb Uni)
• Post doc: Rhizobia and nitrogen fixation (Stanford)
• Research Fellow: Phytophthora vignae – cowpea interaction (Melb Uni)
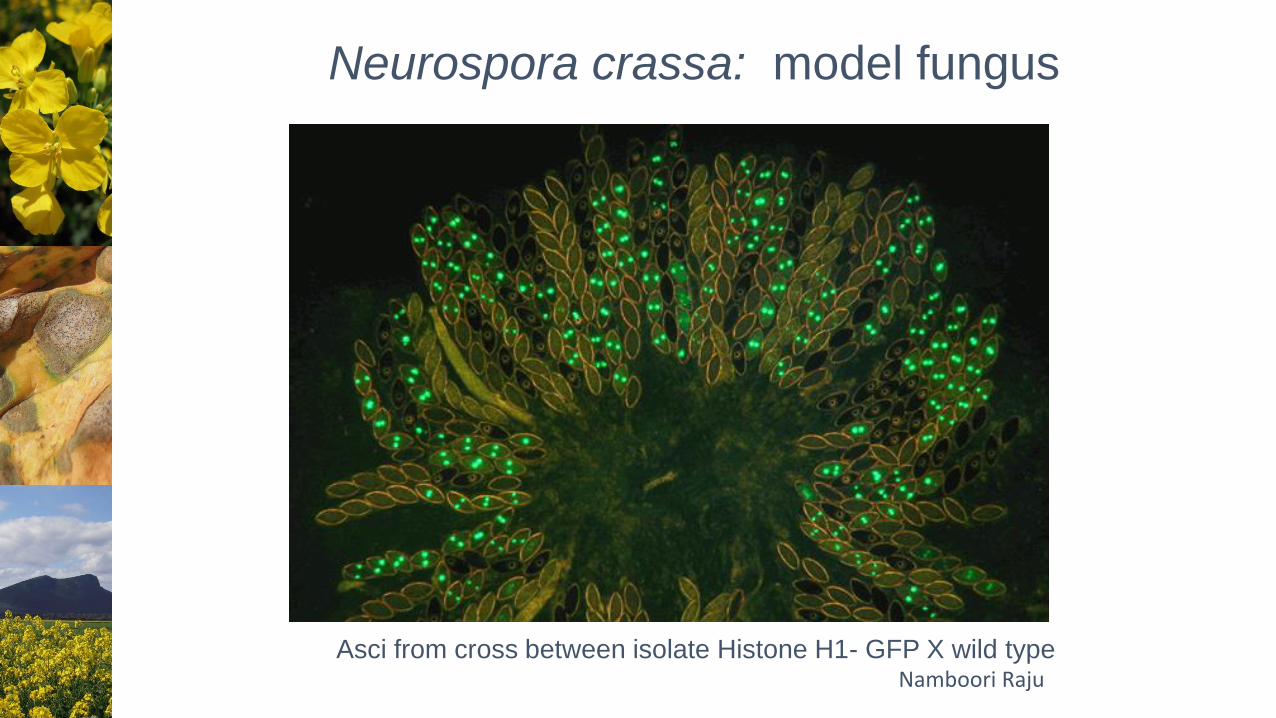
Neurospora crassa: model fungus
Namboori RajuAsci from cross between isolate Histone H1- GFP X wild type
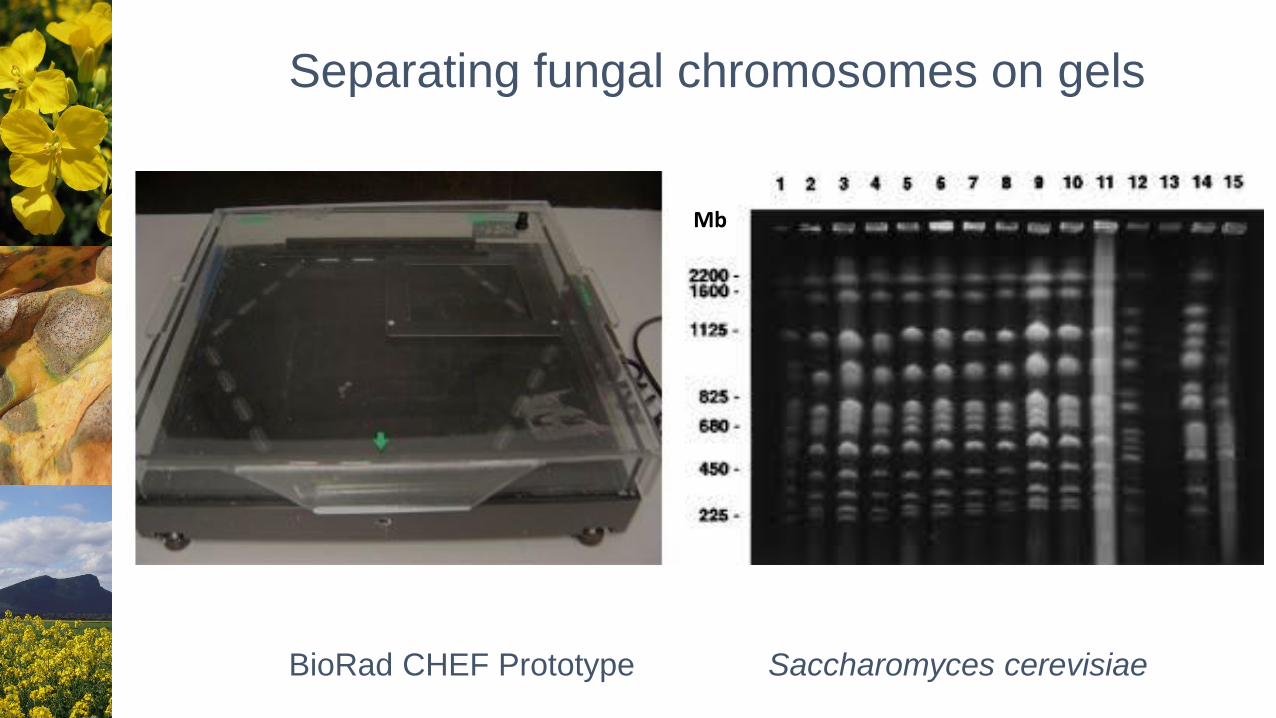
Separating fungal chromosomes on gels
Mb
BioRad CHEF Prototype Saccharomyces cerevisiae

Blackleg caused by Leptosphaeria maculans
• Canola industry established in 1970s but severe blackleg epidemics
• Resistance breeding programs
• Industry re-established in 1990s
• Ascomycete fungus
• Grows on defined media
• Reproducible inoculation protocols
• Genetics: sexual crossing

Canola and blackleg disease in Australia
stubble sexual fruiting bodies windborne ascospores
stem
canker
cotyledon infectionroot rotlodged
plants
SUMMER AUTUMN
SPRING WINTER

Canola in Australia: minimum tillage, long
growing season, temperate climate, high
intensity cropping
2.5 m ha canola sown in 2016; large amount of
blackleg-infested stubble inoculum in 2017

Controlling Blackleg Disease
• Rotation with cereals & pulses
• Fungicide applied to seed, ‘in furrow’ and/or spray
• Resistance breeding; seedling (major gene) & adult plant (minor gene) resistance
No resistance
= disease
Adult (minor gene)
resistance
= some disease
Seedling (major
gene) resistance
= no disease

‘Gene for gene’ interaction
• >10 major resistance genes mapped (LepR3, Rlm2 cloned)
• Avirulence genes cloned (Rouxel INRA, Van de Wouw, Melb Uni)
• Typical effectors: small secreted cysteine-rich proteins
Avirulence gene Resistance gene
AvrLm1 Rlm1
AvrLm6 Rlm6
AvrLm4-7 Rlm4 & Rlm7
AvrLm11 Rlm11
AvrLm5 Rlm5
AvrLm2 Rlm2
AvrLm3 Rlm3
AvrLepR1 LepR1
AvrLepR2 LepR2
AvrLepR3 LepR3

‘Breakdown’ of resistance in 2003
• in 2000 ‘Sylvestris’ cultivars released with resistance gene, LepR3
• Sown extensively on Eyre Peninsula, SA
Eyre Peninsula

2001 and 2002 2003
‘Sylvestris’ cultivars (LepR3)

Beacon
90% yield losses ($30 m) in Eyre Peninsula
Seed withdrawn from sale
2003: Sylvestris

Weekly TimesThe Land

2004: Beacon Sylvestris

2002 2003 2006
Sylvestris resistance
2002 2003 2006
Non-Sylvestris resistance

• Prolific asexual & sexual reproduction - recombination. Large populations of windborne ascospores (inoculum)
• Increased frequency of isolates attacking cultivars with Sylvestris-resistance, not new strain:– isolates collected before 1988 caused disease– breakdown other areas where Sylvestris cultivars sown extensively
• Extensive sowing of cultivars with major gene resistance results in strong selection pressure towards isolates virulent to that resistance gene. Frequency of virulent isolates in populations increases
• What features of the fungal genome allow virulence to evolve so rapidly in field populations?
• What changes in avirulence genes are responsible for breakdown of Sylvestris resistance?
Leptosphaeria maculans - high risk
pathogen for ‘overcoming’ resistance
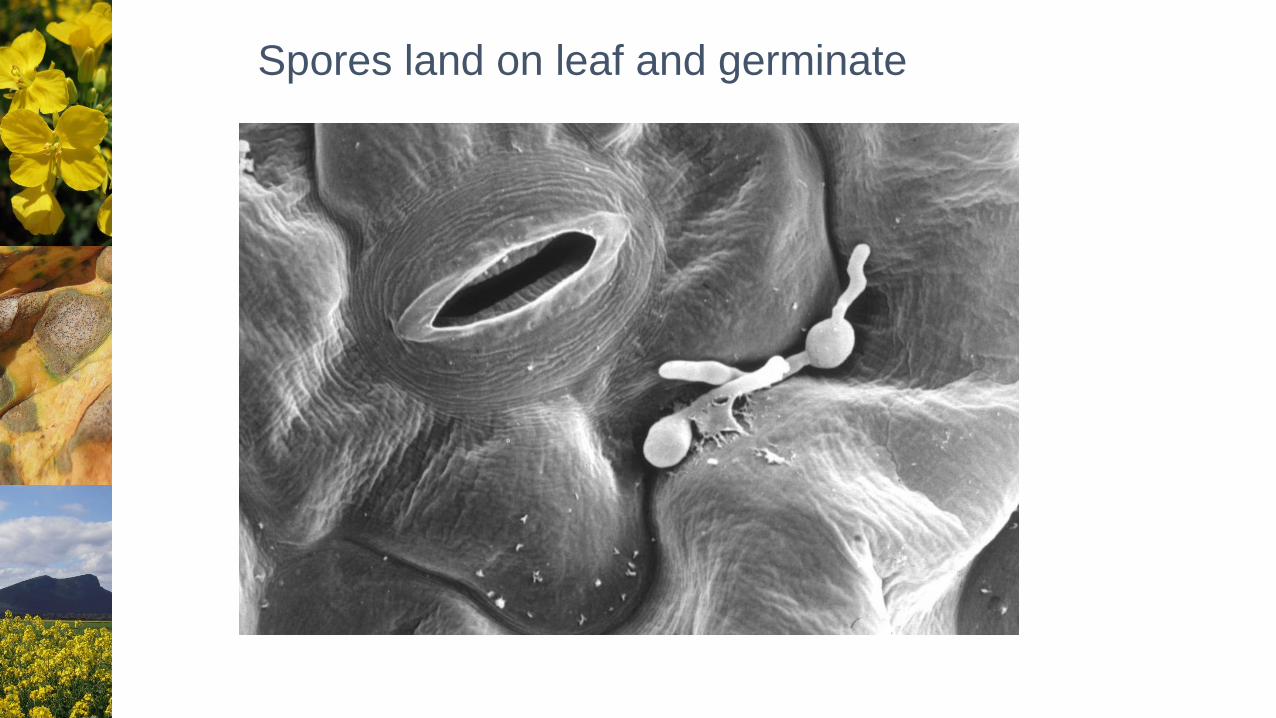
Spores land on leaf and germinate

Hyphae grow across leaf and enter stomata

Successful invasion Unsuccessful invasion
Chen and Howlett (1996) Physiol. Mol. Plant Path. 48: 73-81

Successful invasion
leaf
stem
root
Hyphae (green) grow in leaf into stem then into root Sprague, Watt, Kirkegaard, Howlett (2007) New Phytol 176: 211-22.

Manipulating the blackleg fungus in the
lab
Kim Plummer
Robin Williams

Genome structure: isolates have different-sized
chromosomes
# bands (chromosomes) =16
add sizes of each band (genome size) = 38 Mb
Plummer and Howlett (1993) Curr Genet 24: 107-13

Progeny can have different sized homologous
chromosomes to those of parents
P1 P3P2 P4 P1 P3P2 P4
Same size as parents
Different size to parents
progeny progeny progeny progeny
Plummer and Howlett (1995) Mol Gen Genet 247: 416-22
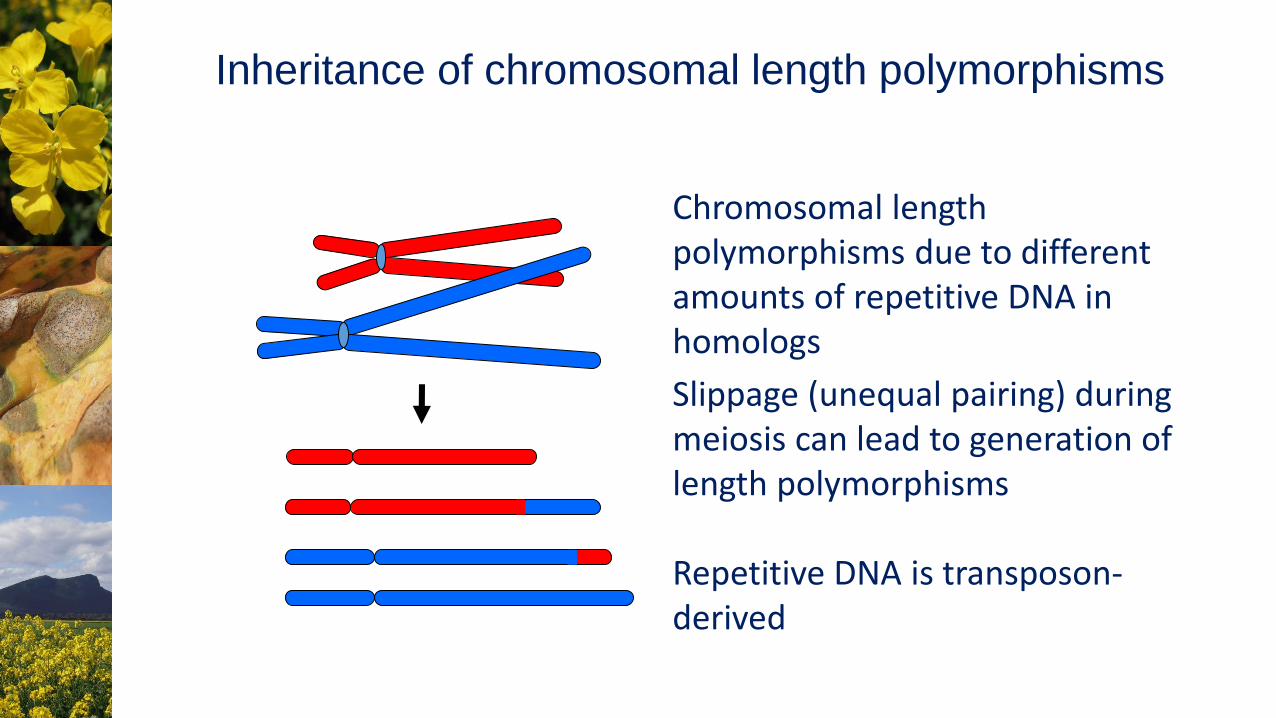
Chromosomal length polymorphisms due to different amounts of repetitive DNA in homologs
Slippage (unequal pairing) during meiosis can lead to generation of length polymorphisms
Repetitive DNA is transposon-derived
Inheritance of chromosomal length polymorphisms

Mutation: Repeat Induced Point (RIP)
• Genome defence against invasion of fungi by transposable
elements?
• Mutates multicopy DNA; occurs in haploid nuclei after
fertilization, prior to mitotic DNA replication in N.crassa
• Transitions from C:G to T:A base pairs; often generates
stop codons
• RIP ‘signature’ deduced by bioinformatics
• Demonstrated experimentally in L.maculans (Alex Idnurm)
• Idnurm & Howlett (2003) Curr Gen 39: 31-7

Leptosphaeria maculans genome sequencing
• Rouxel & Balesdent, INRA & Howlett, UoM;
Genoscope; URGI, France; Oliver (Perth)
• 12,500 genes; 45 Mb -closely related Stagonospora
nodorum (37 Mb)
• 36% genome (9% S. nodorum) is AT-rich repetitive
DNA comprising degenerated transposable elements
• Repeat Induced Point (RIP) mutation in repetitive
DNA
• Unique genome structure

‘Patchwork’ genome: gene–rich (GC) & gene-
poor, repeat –rich repetitive (AT) blocks
Gene-rich blocks(1 gene per 2.4 kb)
Repeat-rich blocks
Gene poor (1 gene/ 30 kb) degenerated transposons only 3.5% of total genes but 20% of effectors
Rouxel et al. Nature Communications (2011) 2: 22


ATTATTATTATTATTATTATTATTATTATTATTATTATTATTA
If repeats align correctly during meiosis
Location of avirulence genes in repetitive regions can lead to gene loss or mutation
Avr
or
Avirulence genes identical in parents & progeny
During sexual reproduction, sister
chromatids align
Crossovers can occur (recombination)
ATTATTATTATTATTATTATTATTATTATTATTATTATTATTA
ATTATTATTATTATTATTATTATTATTATTATTATTATTATTA
ATTATTATTATTATTATTATTATTATTATTATTATTATTATTAAvr
Avr
Avr

If repeats align incorrectly(slippage) during meiosis
Location of avirulence genes in repetitive regions can lead to gene loss or mutation
ATTATTATTATTATTATTATTATTATTATTATTA
Deletion of Avirulence gene
ATTATTATTATTATTATTATTATTATTATTATTATTATTATTATTATTATTAvr
ATTATTATTATTATTATTATTATTATTATTATTATTATT
ATTATTATTATTATTATTATTATTATTATTATTATTATTATTA
Gene duplication
Avr
Avr Avr
RIP mutation

Resistance breakdown and evolution of
virulence
• Why does virulence evolve so rapidly in the blackleg fungus?
• Avirulence genes are located in repetitive regions of genome where RIP mutation and gene loss occur readily
• Sylvestris cultivars have Rlm1 and LepR3
• What changes in AvrLm1 are responsible for breakdown of Sylvestris resistance?

Genomic environment of AvrLm1
• 530 kb region: repetitive (AT- rich) DNA, recombination-deficient
• AvrLm1, AvrLm6, LmCys1 and LmCys2 effectors
• Sequenced genes in isolates:
• 137 before breakdown of resistance (before 2004)
• 158 after breakdown (2004 and later)

Features of AvrLm1, AvrLm6, LmCys1 & LmCys2 effectors in 295 isolates
Gene Mutation Virulence
mechanism
Frequencies of
virulence alleles
AvrLm1 Deletion, aa
sub
Deletion only Eight fold increase
post- sylvestris
breakdown
AvrLm6 Deletion, RIP,
aa sub
Deletion
(70%), RIP &
aa sub
Six fold increase
post- sylvestris
breakdown
LmCys1 RIP, aa
substitutions
Not
applicable
Not applicable
LmCys2 Deletion Not
applicable
Not applicable
Van de Wouw et al. (2010) PLoS Path 6: e1001180

Breakdown of ‘sylvestris’ resistance
• First study of evolutionary constraints on multiple genes in large number fungal isolates
• Linked effector genes evolve differently: deletions, Repeat-Induced Point mutations & amino acid substitutions
• Degree of RIP mutation in single copy genes is proportional to proximity to repetitive transposable elements - ‘leakage‘
• Frequency of isolates with virulence alleles of AvrLm1 & AvrLm6 increased after extensive sowing of cultivars with ‘sylvestris’ resistance, even though no Rlm6 cultivars had been sown
• Genomic location (low recombination frequency) led to a selective sweep whereby selection at AvrLm1 affected frequency of virulence alleles of linked AvrLm6 (hitchhiking)
34

Implications for disease management
• Selection imposed by wide-spread deployment of resistance gene affects evolution of complementary avirulence gene and closely linked avirulence genes
• Knowledge of fungal genome important
• Maximise effective lifetime of resistance genes as each resistance gene costs >$4 million to commercialise and release to farmers
• Virulence frequencies of fungal populations can be manipulated by sowing cultivars with different resistance genes, thus minimising risk of resistance breakdown (pot, field plot experiments, Eyre Peninsula crops)
• Is monitoring changes in frequency of virulence then selectively withdrawing cultivars at risk of resistance breakdown an effective disease control strategy?
35

National Canola Pathology Project
NSW DPI:
Kurt Lindbeck
UM:
Alex Idnurm
Barbara Howlett
Angela Van de Wouw
DAFWA:
Ravjit Khangura
SARDI:
Andrew Ware
Jenny Davidson
MGP:
Steve Marcroft
Angela Van de Wouw
Elizabeth Sheedy
Alistair Smith
Buffy Harrison
CSIRO:
Susie Sprague

NSW DPI:
Kurt Lindbeck
UM:
Alex Idnurm
Barbara Howlett
Angela Van de Wouw
DAFWA:
Ravjit Khangura
SARDI:
Andrew Ware
Jenny Davidson
CSIRO:
Susie Sprague
‘Genome to Paddock’ holistic approach
Monitoring fungal popsAvr gene identificationFungicide tolerance?
Monitoring field diseaseBlackleg resistance groups
Blackleg Management Guide
Canola
Resistance
Fungal
virulence
Sustainable control
of blackleg disease
MGP:
Steve Marcroft
Elizabeth Sheedy
Angela Van de Wouw
Alistair Smith
Buffy Harrison

• Rotating resistance genes requires knowledge of which
genes are in which cultivars
• Inoculate with 12 ‘differential’ isolates (different Avirulence genes) to classify all breeding lines according to complement of resistance genes
• Resistance groups (A-H) based on complement of resistance genes
Virulent(susceptible reaction)
Avirulent(resistant reaction)
Resistance genotyping (grouping) of Australian breeding lines and cultivars

eastern Australia
Determine risk of resistance breakdownby monitoring changes in frequency of avirulence in
fungal populations
Survey (Internal Stem Infection)
Western Australia
Collect stubble: high throughput
molecular assay of ascospores for
frequency of avirulence

Tools: Isolate collection & molecular markers
• Isolates (>4000) since 2000; stubble from 2007 onwards
• Eight avirulence genes cloned
• AvrLm1 = Rlm1; AvrLm4 = Rlm4; AvrLm3 = Rlm3; AvrLm6 = Rlm6 (these R genes in Aust. cultivars)
• Genetic mechanisms conferring virulence identified
• Deletion of entire gene
• Single nucleotide polymorphism (SNP) - amino acid substitution
• Repeat Induced Point mutations
• Molecular markers developed - applied to single isolates & populations
• Allele frequencies change rapidly over time; presence of resistance genes influences avirulence gene frequencies
Van de Wouw et al. (2010) Plant Pathology; (2013) J App Microbiol; (2017) Crop and Pasture Science

• Stubble placed in wind tunnel, ascospores captured on tape
• DNA extracted and analysed by quantitative PCR for presence/absence of band for AvrLm1, AvrLm6
• Pyrosequencing assay for AvrLm4 based on nucleotide polymorphism at base 358 leading to aa change G120 to R120
• Total number spores estimated by PCR of ribosomal DNA region
• Frequency virulence allele measured; risk resistance breakdown determined in each region
• Farmers advised on cultivar choice
High throughput molecular assay to assess virulence alleles in fungal populations

• Guide updated with blackleg
ratings, cultural practices,
resistance groups of cultivars,
fungicide recommendations
• Regional disease warnings based
on risk resistance breakdown
• Encourages farmers to survey their
crops
• Message - Staying Ahead of
Blackleg
• Canola Pathology Workshops for
researchers, breeders, pathologists
and industry
Canola
Communication


2011: Field and pot trials predicted resistance breakdown of Hyola 50 on Eyre Peninsula in 2012
44
Hyola50 infected from Hyola50 stubble
Hyola50 infected from ATR-Cobbler stubble

Breakdown of Hyola 50 ResistanceFeb 2012

Disease severity of cultivars from different
resistance groups: Eyre Peninsula field plots
0
20
40
60
80
100
Hyola444 ATR-Marlin ATR-Stingray CB-Telfer Thumper TT
Perc
enta
ge inte
rnal in
fection
Cultivar
Group
AGroup
CGroup
B
Group
E
October 2012
Group
D
Hyola 50: 90% disease severity

Eyre Peninsula October 2012
Group DGroup E

Wagga Wagga NSW Oct 2012
Group D

Averting yield losses on Eyre Peninsula in 2012
• 60,000 ha sown to canola in 2012
• Assuming 50% yield losses if 30% of area sown to Group D cultivars (eg. Hyola 50) and $500 /tonne for canola (conservative estimates)
• Benefits
• Farmers were saved losses of $18 million
• Group D cultivars still sown in other regions, so breeding companies very supportive of recommendations to avoid resistance breakdown
• Win-win situation for farmers and companies


Current disease issues
• Upper branch canopy infection -
early sowing
• Stacking of resistance genes: many
cultivars have Group A. Few
cultivars to ‘rotate’
• Industry now reliant on fungicides
(triazoles only)
• High throughput screen with
stubble from several locations
shows isolates with tolerance to
fluquinconazole (seed dressing)

• Knocking out L.maculans genes
by CRISPR
• L.maculans genes, toxins
involved in stem cankering
• Determining mechanisms of RIP
mutation in L.maculans
• Cryptococcus – animal pathogen
• Environmental sensing by fungi
Current Lab Activities (Alex Idnurm)


Summary
• Multidisciplinary research can be effective at solving problems – and is extremely rewarding
• Blackleg fungus can rapidly breakdown disease resistance bred into canola cultivars
• Propensity for resistance breakdown due to location of fungal avirulence genes in repetitive (AT-rich) DNA where mutation and deletion readily occurs
• Monitoring changes in disease severity and virulence of fungal populations, then selectively withdrawing cultivars at risk of resistance breakdown minimisesblackleg disease





























