Embed Size (px)
Citation preview

Euphytica 67: 35-40, 1993. © 1993 Kluwer Academic Publishers. Printed in the Netherlands.
A gamete abortion locus detected by segregation distortion of isozyme locus Est-9 in wide crosses of rice (Oryza sativa L.)
S.Y. Lin & H. Ikehashi Laboratory of Plant Breeding, Faculty of Horticulture, Chiba University, Matsudo 648, Matsudo, Chiba, Japan 271
Received 3 September 1992; accepted 3 March 1992
Key words: segregation distortion, wide cross, pollen sterility, allele transmission rate, gamete abortion, Oryza sativa rice
Summary
A locus for male gamete abortion in hybrids for Japonica and Indica rice was identified with the aid of marker genes Rc and Est-9 on chromosome 7. In an Indica-Japonica cross, A K A M A I 1/IR50, the Indica allele Est-92 was transmitted via the male gamete with a ratio of 0.29 instead of the normal 0.5, whereas no segregation distortion was observed for the Rc locus. The recombination value (p) for Est-9 and Rc was estimated to be 0.38 by a least square method after adjusting Mendelian segregation ratios with the male transmission ratios of 0.29 (Tr) for Est-92 and 0.71 (1-Tr) for Est-9 ~. The recombination value (q) for the new locus for male gamete abortion, ga-H, and Est-9 was estimated to be 0.23 by using 56 F 3 lines from F 2 plants which were heterozygous for the Est-9 locus. No linkage for Rc and ga-ll was found. Therefore, the two markers and ga-ll were located in the order ofga-ll - Est-9- Rc. Using the estimated recombination value (q), the male transmission rate (k) of ga-H a was estimated to be 0.11 with the F 2 data and - 0.07 with the F 3 line data. Thus, it was apparent that male gametes possessing ga-ll a were frequently aborted in the Indica-Japonica hybrid.
Introduction
Partial sterility of spikelet and pollen in hybrids of wide crosses in rice has been reported by many re- searchers (e.g., Kato et al., 1930; Terao & Mizushi- ma, 1939; Jones & Longley, 1941; Kuang, 1951; Shas- try & Misra, 1961; Kitamura, 1962; Oka, 1974). To date, ten gametophyte genes, ga-1 to ga-lO for segre- gation distortion, have been located on various rice chromosomes (Iwata et al., 1964; Nakagahra, 1972, 1981; Nakagahra et al., 1972; Nakagahra et al., 1974; Mori et al., 1973; Maekawa, 1982; Maekawa & Kita, 1985; Maekawa et al., 1981; Kinoshita & Takamure, 1984).
It was assumed that distortion is caused by differ-
ential fertilization of male gametes carrying differ- ent alleles of one or more ga loci. Spikelet or pollen sterility may also lead to distorted segregation of marker genes. The effect ofga genes has been stud- ied in only a few cases. More recently, many instanc- es of segregation distortion were identified during studies of the inheritance of isozyme phenotypes (Ishikawa et al., 1987a, b; Ranjhan et al., 1988; Wu et al., 1988; Guiderdoni et al., 1989; Lin et al., 1992) or RFLPs (McCouch et al., 1988; Saito et al., 1991).
In our previous studies, segregation distortion via male gametes in Indica-Japonica hybrids in rice was surveyed using one or two marker genes on each of chromosomes 3, 4, 6, 7, 8, 11 and 12 of the 12 rice chromosomes. Marker genes on chromosomes 3, 7,

36
8, 11 and 12 showed clear segregation distortion. The germinabillity of the pollen was less than ten percent in the hybrids, although 45-55 percent of the pollen grains appeared to be morphologically normal (Lin et al., 1992).
Mapping of loci for distorted segregation is diffi- cult because no conventional method is available. However, a method to estimate recombination for a ga locus and a distorted marker was given by Iwata et al. (1964) and Nakagahra (1972, 1981). They test- ed F 3 lines, to distinguish families in which the seg- regation distortion was similar to that found in F 2 from families with different segregation types fol- lowing recombination for the marker and ga gene. Ratios of the different segregation types were used to estimate the recombination value for marker gene and the ga locus.
In this study we report an analysis of a new locus, ga-H, causing distorted segregation of Est-9 on chromosome 7. By determining the recombination value for ga-ll and Est-9, it was possible to estimate the male transmission rates of the respective ga-ll alleles. The results indicated that male gametes car- rying alleles, g a - l l a w e r e aborted. A method to esti- mate the recombination value for a distorted mark- er and a gamete abortion locus is discussed.
Materials and methods
Plant materials and marker genes
Three single crosses between Indicas and Japonicas showed a significant decrease of the Indica allele at the Est-9 locus in F2 populations (Lin et al., 1992). An F 2 population from one of these, A K A M A I 1/IR 50, was chosen to study a locus causing segregation distortion. This locus is hereafter designated as ga-ll. The segregation distortion was limited to the male gamete (Lin et al., 1992). A K A M A I 1 is a Ja- ponica cultivar possessing Rc/Rc (brown pericarp and seed coat) and Est-9~/Est-91 on chromosome 7. IR 50, a typical Indica cultivar released in the Phi- lippines, possesses Rc+/Rc ÷ and Est-9e/Est-92.
In the F 2 population, the segregations of Est-9 and Rc genotypes were examined. Of the 98 F 2 plants showing heterozygosity at Est-9, 56 random plants were scored for Est-9 genotype in the F 3 seed- lings. The segregation data in the F2 population and in F 3 lines were analyzed to estimate the recombina- tion values among ga-11, Est-9 and Rc. The Rc geno- type was scored visually in F 2. The Est-9 isozyme was analyzed by starch gel electrophoresis accord- ing to the method by Ishikawa et al. (1989).
Table 1. Theoretical and observed frequencies of each genotype in the F 2 population of A K A M A I 1/IR50
Genotype a Theoretical frequency Modified frequency No. of plants Tr = 0.29 b
p = 0.5 Modified
Tr = 0.29
p = 0.5
Observed
Expected with Tr =
0.29
p = 0.38
Rc/ - , Est-9 n (1-Tr + Trp2-p2)/2 0.267 58.4 72 66.5 Rc/- , Est-912 (1-p + p2)/2 0.375 82.1 71 83.7
Rc/- , Est-922 (2Trp-Trp2)/2 0.108 23.8 18 19.5 Rc+IRc +, Est-9 ~ (pZ-Trp2)/2 0.089 19.5 14 11.2
Rc÷lRc +, Est-9 ~2 p(l-p)/2 0.125 27.4 27 25.8 Rc÷IRc +, Est-922 (Tr-2Trp + TrpZ)/2 0.036 7.9 17 12.2 Total 1 1.00 219.0 219 218.9 ~z probability < 0.01 0.3~).5
a ESt_911 Est_9~2 and Est-9 e2 indicate Est-91/Est/9 ~, Est/9tEst/9 e and Est-9elEst-92, respectively.
b Tr: Transmission rate of Est-9 e via male gametes, by Tr = (18 + 17)/(18 + 17 + 72 + 14) (n = 0.29). p: Recombination value.
37
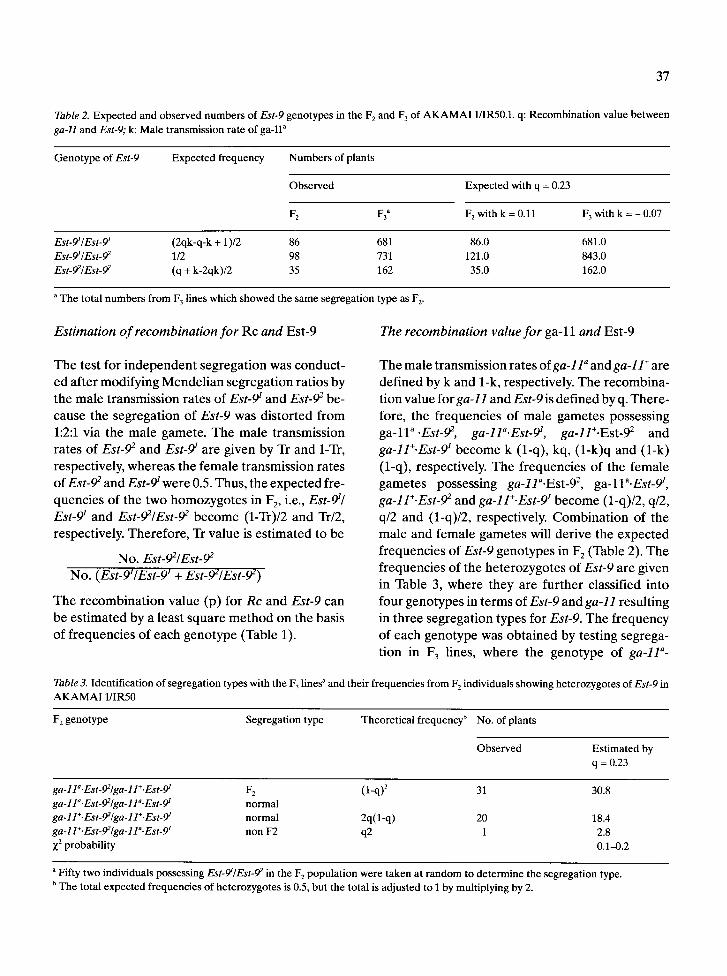
Table 2. Expected and observed numbers of Est-9 genotypes in the F 2 and F 3 of AKAMAI 1/IR50.1. q: Recombination value between ga-ll and Est-9; k: Male transmission rate of ga-ll a
Genotype of Est-9 Expected frequency Numbers of plants
Observed Expected with q = 0.23
F z F3 a F 2 with k = 0.11 F 3 with k = - 0.07
Est-ff/Est-9 ~ (2qk-q-k + 1 )/2 86 681 86.0 681.0 Est-9~/Est-92 1/2 98 731 121.0 843.0 Est-92/Est-92 (q + k-2qk)/2 35 162 35.0 162.0
a The total numbers from F 3 lines which showed the same segregation type as F 2.
Est imat ion o f recombinat ion f o r R c and Est-9 The recombinat ion value f o r g a - l l and Est-9
T h e tes t for i n d e p e n d e n t s eg rega t i on was conduc t -
ed a f t e r mod i fy ing M e n d e l i a n s eg rega t i on ra t ios by
the ma le t r ansmis s ion ra tes of Est-9 ~ and Est-92 be-
cause the s e g r e g a t i o n o f Est-9 was d i s t o r t ed f rom
1:2:1 via the ma le gamete . T h e ma le t r ansmis s ion
ra t e s of Est-92 and Est- f f are g iven by Tr and 1-Tr,
respect ive ly , w h e r e a s the f ema le t r ansmiss ion ra tes
of Est-92 and Est-9 ~ were 0.5. Thus , the e x p e c t e d fre-
quenc ies of the two h o m o z y g o t e s in F2, i.e., Est-9~/ Est-9 ~ and Est-92/Est-92 b e c o m e (1-Tr)/2 and Tr/2,
respect ive ly . The re fo r e , Tr va lue is e s t i m a t e d to be
No. Est-92/Est-92 No. (Est-9~/Est-ff + Est-92/Est-92)
T h e r e c o m b i n a t i o n va lue (p) for Rc and Est-9 can
be e s t i m a t e d by a leas t squa re m e t h o d on the basis
of f r equenc ies of each g e n o t y p e (Table 1).
T h e ma le t r ansmis s ion ra tes o f g a - l l a and g a - l l ÷ are
de f ined by k and l - k , respect ively . T h e r e c o m b i n a -
t ion va lue for ga-11 and Est-9 is de f i ne d by q. T h e r e -
fore, the f r equenc ies of ma le ga me te s possess ing g a - l l a-Est-92, ga-lla'Est-91, ga-ll+.Est-92 and
ga-l l+.Est- f f b e c o m e k ( l - q ) , kq, (1-k)q and ( l - k )
( l - q ) , respect ively . The f r equenc ies of the f ema le
g a m e t e s possess ing ga-11a.Est-92, ga-l la 'Est- f f , ga-11*.Est-92 and ga-l l÷.Est-9 ~ b e c o m e ( l -q) /2 , q/2,
q/2 and ( l -q ) /2 , respect ively . C o m b i n a t i o n of the
ma le and f ema le g a m e t e s will de r ive the e x p e c t e d
f requenc ies of Est-9 g e n o t y p e s in F 2 (Table 2). T h e
f requenc ies of the h e t e r o z y g o t e s of Est-9 are given
in Table 3, w h e r e t hey a re fu r the r c lass i f ied into
fou r g e n o t y p e s in t e rms of Est-9 and ga-11 resu l t ing
in t h r e e s eg rega t ion types for Est-9. T h e f r equency
of each g e n o t y p e was o b t a i n e d by tes t ing segrega-
t ion in F 3 l ines, w h e r e the g e n o t y p e of g a - l P -
Table 3. Identification of segregation types with the F 3 lines ~ and their frequencies from F 2 individuals showing heterozygotes of Est-9 in AKAMAI 1/IR50
F 2 genotype Segregation type Theoretical frequency b No. of plants
Observed Estimated by q = 0.23
ga-lla'Est-92/ga-ll+'Est-9~ F 2 (l-q) 2 31 30.8 ga- l la. Est-92 /ga- l la. Est- gt normal ga-11+.Est-92/ga-ll+.Est-9~ normal 2q(1-q) 20 18.4 ga-11+.Est-92/ga-1P.Est-9~ non F2 q2 1 2.8 X 2 probability 0.1-0.2
a Fifty two individuals possessing Est-9~/Est-9 2 in the F 2 population were taken at random to determine the segregation type. b The total expected frequencies of heterozygotes is 0.5, but the total is adjusted to 1 by multiplying by 2.
38
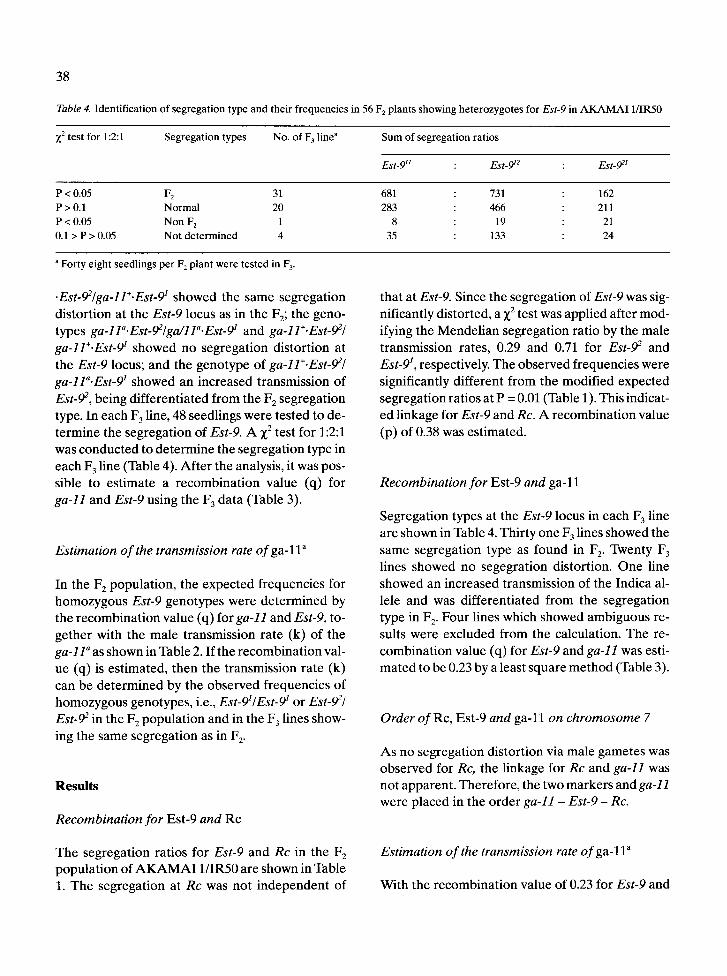
Table 4. Identification of segregation type and their frequencies in 56 F 2 plants showing heterozygotes for Est-9 in AKAMAI 1/IR50
X z test for 1 : 2 : 1 Segregation types No. of F 3 line a Sum of segregation ratios
Est- 9 I I : Est-912 : Est-921
P < 0.05 F 2 31 681 : 731 : 162 P > 0.1 Normal 20 283 : 466 : 211 P < 0.05 Non F 2 1 8 : 19 : 21 0.1 > P > 0.05 Not determined 4 35 : 133 : 24
a Forty eight seedlings per F 2 plant were tested in F 3.
• Est-92/ga-ll+.Est-9 ~ showed the same segregat ion
dis tor t ion at the Est-9 locus as in the F2; the geno- types ga-lla'Est-92/ga/lla'Est-9 l and ga-11+.Est-92/
ga-ll+.Est-9 ~ showed no segregat ion distort ion at the Est-9 locus; and the geno type of ga-11+.Est-92/ ga-11a'Est-91 showed an increased t ransmission of
Est-92, being different ia ted f rom the F z segregat ion type. In each F 3 line, 48 seedlings were tested to de- termine the segregat ion of Est-9. A X 2 test for 1:2:1 was conduc ted to de te rmine the segregat ion type in each F 3 line (Table 4). Af t e r the analysis, it was pos- sible to est imate a r ecombina t ion value (q) for ga-11 and Est-9 using the F 3 data (Table 3).
Estimation o f the transmission rate o f g a - l l a
In the F2 popula t ion, the expected frequencies for h o m o z y g o u s Est-9 genotypes were de te rmined by the r ecombina t ion value (q) for g a - l l and Est-9, to- ge ther with the male transmission rate (k) of the g a - l l a as shown in Table 2. If the r ecombina t ion val- ue (q) is est imated, then the t ransmission rate (k) can be de te rmined by the observed f requencies of h o m o z y g o u s genotypes , i.e., Est-9~/Est-9 ~ or Est-92/
Est-92 in the F 2 popula t ion and in the F 3 lines show-
ing the same segregat ion as in F 2.
Results
Recombinat ion f o r Est-9 and Rc
The segregat ion ratios for Est-9 and Rc in the F 2 popula t ion of A K A M A I 1/IR50 are shown in Table 1. The segregat ion at Rc was no t independen t of
that at Est-9. Since the segregat ion of Est-9 was sig- nificantly distorted, a 22 test was applied after mod- ifying the Mendel ian segregat ion ratio by the male t ransmission rates, 0.29 and 0.71 for Est-92 and Est-91, respectively. The observed frequencies were significantly different f rom the modif ied expected segregat ion ratios at P = 0.01 (Table 1). This indicat- ed l inkage for Est-9 and Rc. A recombina t ion value
(p) of 0.38 was est imated.
Recombinat ion for Est-9 and ga-11
Segregat ion types at the Est-9 locus in each F 3 line are shown in Table 4. Thir ty one F 3 lines showed the same segregat ion type as found in F 2. Twenty F 3 lines showed no segegrat ion distortion. One line showed an increased t ransmission of the Indica al- lele and was different iated f rom the segregat ion type in F 2. Four lines which showed ambiguous re- suits were excluded f rom the calculation. The re- combina t ion value (q) for Est-9 and ga-11 was esti- ma ted to be 0.23 by a least square m e t h o d (Table 3).
Order o f R c , Est-9 and ga-11 on chromosome 7
As no segregat ion dis tort ion via male gametes was observed for Rc, the l inkage for Rc and g a - l l was not apparent . Therefore , the two markers and g a - l l were placed in the order ga- l l - Est-9 - Rc.
Estimation o f the transmission rate o f g a - l l a
With the recombina t ion value of 0.23 for Est-9 and

g a - l l , the transmission rate (k) of g a - l l ~ was esti- mated to be 0.11 with the F 2 data a n d - 0.07 with the data of F3 lines (Table 2).
Discussion
Wu et al. (1988) and Ranjhan et al. (1988) located Est-9 on chromosome 7 using trisomics, but the lo- cus has not been mapped precisely. In this report, recombination for Rc and Est-9 was estimated to be 0.38 by a least square method after the expected segregation ratio was modified using male trans- mission rates for Est-9 ~ and Est-92. Recently, Yana- gihara et al. (1992) estimated this value to be 0.35 using a cross showing no segregation distortion. This confirmed the validity of the value obtained here. This method of linkage analysis will be useful to estimate recombination values for a non-distort- ed marker and a distorted marker.
To date, ten loci (ga-1 to ga-lO) for distorted seg- regation have been located on various chromo- somes (Iwata et al., 1964; Nakagahra, 1972, 1981; Nakagahra et al., 1972; Nakagahra et al., 1974; Mori et al., 1973; Maekawa et al., 1981; Maekawa, 1982; Maekawa & Kita, 1985). Whether the distorted seg- regation was caused by selective fertilization or by gamete abortion was not critically studied.
To estimate male transmission rates of ga alleles, it is necessary to evaluate the recombination value for a ga and its marker. In this study, we used a co- dominant marker, Est-9, to locate g a - l l , and eval- uated the male transmission rate of the g a - l l a allele. The recombination value (q) for Est-9 and ga-11 on chromosome 7 was estimated to be 0.23. Thus, the transmission rate (k) of g a - l l ~ was estimated to be 0.11 with F2 data and - 0.07 with F 3 data. These re- sults indicated that almost all male gametes pos- sessing ga-11 a were excluded from fertilization. In an analysis of a locus, Fs, for pollen sterility, Kitam- ura (1962) showed that pollen carrying allele Fs ~ was completely sterile when produced by a Fs~/Fs ' geno- type. Iwata et al. (1964) showed that the ga-1 a allele was male transmitted with a frequency of 0.047. Na- kagahra (1981) showed that a ga-8 ~ allele was not transmitted in competit ion with ga-8 + indicating a transmission rate of - 0.124 for ga-8L Most of the
39
data available so far indicate that ga genes causing segregation distortion of linked markers were al- most or completely excluded from fertilization.
If the transmission rate of ga-11 a (k) is zero, the segregation of genotypes can be simplified. When k is zero (Table 2), the frequencies of Est-99Est-91 and Est-92/Est-92 become (l-q)/2 and q/2, respec- tively. On the other hand, the male transmission rates of Est-9 ~ and Est-92 are defined by 1-Tr and Tr, respectively, while the comparable female rates are 0.5. Thus, the expected frequencies of the two ho- mozygotes in F 2, i.e., Est-9J/Est-9 ~ and Est-92/Est-92,
become (1-Tr)/2 and Tr/2, respectively. Because Tr is equal to q, a recombination value for a ga locus and its marker can be estimated directly by a male transmission rate of the marker linked with an aborted allele. In this study, Tr was 0.29 and q was 0.23. Lin et al. (1992) reported an average Tr value of 0.25 for six estimates. Hence, Tr was approxi- mately equal to q, indicating the validity of the as- sumption.
In a study on female gamete abortion in rice, Ike- hashi & Araki (1986, 1988) reported locus S-5 on chromosome 6 where the alleles S-5 i and S-Y were present in Indica and Japonica cultivars, respective- ly. An abortion-neutral allele S-5 n was present in wide compatibility genotypes. In the heterozygote S-Y/S-5~ female gametes possessing S - Y aborted, but in the heterozygote S-Y/S-5 n and S-Y/S-5% fe- male gametes were normally transmitted. A similar genetic mechanism can be assumed for male ga- mete abortion. While hybrid sterility expressed in male gametes may be a barrier in wide crosses, there is a possibility of finding gamete-abortion- neutral alleles which mitigate the male sterility as indicated by Lin et al. (1992).
References
Guiderdoni, E., B. Corutois & J.C. Glaszmann, 1989. Use of iso- zyme markers to monitor recombination and assess gametic selection among anther culture derivatives of remote crosses of rice (Oryza sativa L.). In: A. Mujeeb-Kazi & L.A. Sitch (Eds). Review of advances in plant biotechnology, 1985-88: 2nd International Symposium on Genetic Manipulation in Crops. Mexico, D.E, Mexico, and Manila, Philippines: CIM- MYT and IRRI. pp. 43-55.

40
Ikehashi, H. & H. Araki, 1986. Genetics of F 1 sterility in remote crosses of rice. Rice Genetics, p. 119-130. IRRI, Los Banos, Philippines.
Ikehashi, H. & H. Araki, 1988. Multiple alleles controlling F 1 sterility in remote crosses of rice (Oryza sativa L.). Jpn. J. Breed. 38: 283-291.
Ishikawa, R., T.I. Sato & H. Morishima, 1987a. Abnormal segre- gation of isozyme genes in the Indica x Japonica crosses of the common rice. Jpn. J. Breed. 37 (Suppl. 2): 188-189 (In Japa- nese).
Ishikawa, R., T. Kinoshita & H. Morishima, 1987b. Trisomic analysis of genes for isozymes: Location of Cat-l, Acp-1 and Pox-2 on chromosomes. Rice Genet. Newslett. 4: 75-76.
Ishikawa, R., H. Morishima, K. Mori & T. Kinoshita, 1989. Chro- mosomal analysis of isozyme loci and the allelic expression at cellular level in rice. J. Fac. Agric. Hokkaido Univ. (Japan) 64: 85-98.
lwata, N., T. Nagamatsu & J. Omura, 1964. Abnormal segrega- tion of waxy and apiculus coloration by a gametophyte gene belonging to the first linkage group in rice. Jpn. J. Breed. 14: 33-39.
Jones, J.W. & A.E. Longley, 1941. Sterility and aberrant chromo- some numbers in Caloro and other varieties of rice. J. Agr. Rec. 62: 381-399.
Kato, S., 1930. On the affinity of the cultivated varieties of rice plants Oryza sativa L. J. Dep. Agr. Kyushu Imp. Univ. 2: 241- 276.
Kinoshita, T. & I. Takamure, 1984. Inheritance and zebra nebra necrosis in rice - Genetical studies on rice plant, LXXXVIII. J. Fac. Agr. Hokkaido Univ. 61: 445-455.
Kitamura, E., 1962. Genetic studies on sterilities observed in hy- brids between distantly related varieties of rice, Oryza sativa L. Bull. Chugoku Agr. Exp. Sta. Series A, 8,141-205. (In Japa- nese, with English summary).
Kuang, H., 1951. Studies on rice cytology and genetics as well as breeding work in China. Agron. J. 43: 387-397.
Lin, S.Y., H. Ikehashi, S. Yanagihara, K. Kawashima, 1992. Seg- regation distortion via male gametes in hybrids between In- dica and Japonica or wide-compatibility varieties of rice (Ory- za sativa L.). Theor. Appl. Genet. 84: 812-818.
Maekawa, M., 1982. Studies on genetical differentiation in dis- tant crosses of rice. Mem. Fac. Agr. Hokkaido Univ. 13: 147- 177 (In Japanese with English summary).
Maekawa, M., T. Kinoshita & M. Takahashi, 1981. Genetical studies on rice plant. LXXVI. A new gametophyte gene in the second linkage group of rice. J. Fac. Agr. Hokkaido Univ. 60: 107-114.
Maekawa, M. & E Kita, 1985. New gametophyte genes located inthe third linkage group (chromosome 3) of rice. Jpn. J. Breed. 35: 25-31.
McCouch, S.R., G. Kochert, H.Z. Yu, Z.Y. Wang, G.S. Khush, W.R. Coffman & S.D. Tanksley, 1988. Molecular mapping of rice chromosomes. Theor. Appl. Genet. 76: 815-829.
Mori, K., T. Kinoshita & M. Takahashi, 1973. Genetical studies on rice plant. LVIII. Segregation distortion and its causation of an endosperm character in crosses of distantly related rice va- rieties. Mem. Fac. Agr. Hokkaido Univ. 9:74-86 (In Japanese with English summary).
Nakagahra, M., 1972. Genetic mechanism on the distorted segre- gation of marker genes belonging to the eleventh linkage group in cultivated rice. Jpn. J. Breed. 22: 232-238.
Nakagahra, M., 1981. New distorted inheritance of markers lo- cated on chromosome 3 in wide crosses of rice Oryza sativa L. Bull. Nat. Inst. Agr. Sci. D 32:15-44 (In Japanese with English summary).
Nakagahra, M., T. Omura & N. Iwata, 1972. Gametophyte genes and their loci on the eleventh linkage group of cultivated rice. Jpn. J. Breed. 22: 305-312.
Nakagahra, M., T. Omura & N. Iwata, 1974. New certation gene on the first linkage found by inter-subspecific hybridization of cultivated rice. J. Fac. Agr. Kyushu Univ. 18: 157-167.
Oka, H., 1974. Analysis of genes controlling F 1 sterility in rice by the use of isogenic lines. Genetics. 77: 521-534.
Ranjhan, S., J.C. Glaszmann, D.A. Ramirez & G.S. Khush, 1988. Chromosomal localization of four isozyme loci by trisomic analysis in rice (Oryza sativa. L.). Theor. Appl. Genet. 75: 741-745.
Saito, A., E. Shimosaka, Y. Hayano, K. Saito, S. Matuura & M. Yano, 1991. Molecular mapping on chromosome 3 of rice. Jpn. J. Breed. 41 (Suppl. 1): 158-159 (In Japanese).
Shastry, S.V.S. & R.N. Misra, 1961. Pachytene analysis in Oryza II. Sterility in Japonica-lndica hybrids. Chromosome 12: 248- 271.
Terao, H. & U. Mizushima, 1939. Some considerations on the classification of Oryza sativa L. into two subspecies, so-called 'Japonica' and 'Indica'. Jpn. J. Bot. 10: 213-258.
Wu, K.S., J.C. Glaszmann & G.S. Khush, 1988. Chromosomal locations of ten isozyme loci in rice (Oryza sativa L.) through trisomic analysis. Biochemical Genetics 26: 303-320.
Yanagihara, S., H. Kato & H. Ikehashi, 1992. A new locus for multiple alleles causing hybrid sterility between an Aus varie- ty and Javanica varieties in rice (Oryza sativa L.). Jpn. J. Breed. 42: 793-801.





























