Embed Size (px)
Citation preview

1
A DIACHRONIC ASSESSMENT OF HEALTH AND DISEASE FROM THE ADULT DENTITION OF THE NATON BEACH BURIAL COMPLEX IN TUMON BAY, GUAM
By
NICOLETTE M. PARR
A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY
UNIVERSITY OF FLORIDA
2012

2
© 2012 Nicolette Maria Luney Parr

3
To my mother and father for always providing me with endless support and
encouragement

4
ACKNOWLEDGMENTS
First and foremost I would like to thank my committee members, Mike Warren,
John Krigbaum, Dave Daegling, and David Steadman for your tireless advice and
patience and Chris Schmidt for introducing and instilling in me a love for teeth. Drs.
Warren and Krigbaum have been my mentors and friends since my undergraduate
years and never hesited to give me guidance and encouragement along the way. They
took the gamble to accept me back as a Ph.D. student – for that, I will be forever
grateful.
This study would have never been conducted had it not been for the opportunities
provided to me by Patrick O’Day, Nicole Vernon, Mike Desilites, and Garcia and
Associates. I want to express my deepest gratitude to Pat and Nicole for introducing
me to Guam and the Pacific region and for the countless dinners, drinks, birthday
celebrations, holidays, and hours of laughter we shared together. Your kindness and
generosity will never be forgotten. Am am also lucky to have made wonderful friends in
Guam: Jamey, Cyrus, Marie, Justin, and Patrick, to whom I am thankful for so many
great adventures and for reminding me to explore the island, eat good food, and spend
time underwater when the going got tough.
I am particularly thankful to Dave DeFant for access to the Naton Beach skeletal
collection, as well as Cherie Walth, Sandy Yee, Lynn Leon-Guerro, and Michelle Christy,
from SWCA, for providing me with endless amounts of information regarding the Naton
Beach site, access to their library, and workspace in which I spent many months. I am
also grateful to a number of people who have worked in Guam for many years and have
provided me with valuable information, feedback, and thoughtful conversation regarding
my research: Gary Heathcote, Rona Ikehara-Quebral, Judith Amesbury, Rosanna

5
Barcinas, Boyd Dixon, Jonn Peterson, Anne Stodder, Lawrence Cunningham, Lon
Bulgrin, Michael Pietrusewsky, Bruce Anderson, Vince Sava, Michele Douglas, and
Joanne Eakin.
I would like to thank my fellow grad students from the Pound Lab: Laurel, Katie,
Traci, Caroline, Sarah, Allysha, Carrie, and Kristina. You have been with me through
many highs and lows, have endured my caffeine-induced hysteria, and listened to my
usually pointless stories. I am so grateful for all you have done for me, from reading a
myriad of grant proposals to giving me statistical advice, which does not even begin to
enumerate it all. A special thanks goes out to Carlos Zambrano for throughoughly
reviewing this entire document and for putting up with me for six more years and then
some – here’s to ugly babies!
To Lulu, Kim, Viviana, Amber, and Leila: much of who I am today I owe to having
had you in my life; thank you for always being there for me, no matter the distance
between us. I am deeply indebted to my many family members who supported me even
when they thought I would always be a perpetual student, particularly Sonia, Casper,
Piedad, and Jennifer. Last but not least, I would like to give my most heartfelt thanks to
my mother and father who continuously provided me with unconditional love and
support. You have always encouraged me to never stop learning and for teaching me
how to travel, eat good food, and to love life – I could not have done this without you.
This dissertation was supported in part by the William R. Maples Memorial
Scholarship, O. Ruth McQuown Supplemental Award, Wentworth Foundation William M.
Goza Fellowship, Delores Auzenne Minority Dissertation Award, Ellis R. Kerley Forensic
Sciences Foundation Scholarship, and the CA Pound Human Identification Laboratory.

6
TABLE OF CONTENTS page
ACKNOWLEDGMENTS .................................................................................................. 4
LIST OF TABLES ............................................................................................................ 9
LIST OF FIGURES ........................................................................................................ 14
LIST OF ABBREVIATIONS ........................................................................................... 15
ABSTRACT ................................................................................................................... 17
CHAPTER
1 INTRODUCTION .................................................................................................... 19
Theoretical Framework ........................................................................................... 21
Biocultural Approach to Bioarchaeology ........................................................... 22 Stress Models ................................................................................................... 23
Purpose and Research Objectives ......................................................................... 27
Objectives and Hypotheses .................................................................................... 28 Chapter Organization .............................................................................................. 30
2 NATURAL AND CULTURAL ENVIRONMENT ....................................................... 33
Study Location ........................................................................................................ 33
Natural Environment ............................................................................................... 33 Biogeographical Divides ................................................................................... 34 Paleogeography ............................................................................................... 35
Paleoenvironment ............................................................................................ 37 Paleofauna ....................................................................................................... 39
Settlement History .................................................................................................. 40 Colonization of the Pacific Region .................................................................... 40
Archaeology ............................................................................................... 40
Linguistics .................................................................................................. 41 Biology ....................................................................................................... 42
Colonization of the Mariana Islands ................................................................. 44 Linguistics .................................................................................................. 45
Archaeology ............................................................................................... 46 Genetics ..................................................................................................... 48 Bioarchaeology .......................................................................................... 49
Settlement Summary ........................................................................................ 51 Marianas Chronological Sequence ......................................................................... 51
Pre-Latte Period ............................................................................................... 53 Diet ............................................................................................................ 54
The Early Unai phase (3500-3000 BP) ...................................................... 56

7
The Middle Unai period (3000-2500 BP) .................................................... 57
The Late Unai period (2500-2400 BP) ....................................................... 58 The Transitional (Huyong) period (400-1000 CE) ...................................... 59
Latte Period (1000-1668 CE) ............................................................................ 59 Latte architecture ....................................................................................... 60 Cooking and food processing ..................................................................... 61 Diet ............................................................................................................ 62
Post-Contact Era .............................................................................................. 65
The Spanish Colonial Period (1521-1898 CE) ........................................... 66 The First American Period (1898-1941 CE) ............................................... 68 The Japanese World War II Period (1941-1944 CE) .................................. 68 The American World War II Period (1944-1948 CE) .................................. 69 The Second American Period (1945-Present) ........................................... 69
3 MATERIALS ........................................................................................................... 77
Archaeological Sample ........................................................................................... 77 Taphonomic Bias .................................................................................................... 78
Sample Population .................................................................................................. 80 Pre-Latte Demographics................................................................................... 80 Latte Demographics ......................................................................................... 80
4 DENTAL REDUCTION ........................................................................................... 85
Background ............................................................................................................. 85
Current Study .......................................................................................................... 86 Expected Results .................................................................................................... 88
Methods .................................................................................................................. 90 Results .................................................................................................................... 92
Pre-Latte and Latte Differences ........................................................................ 94
Male and Female Differences ........................................................................... 95 Interaction between Time and Sex ................................................................... 95
Discussion .............................................................................................................. 96 Tooth Summaries and Rate of Change ............................................................ 96 Hypothesis Testing ........................................................................................... 98
Time Period Differences ............................................................................ 98 Comparison of dental, craniofacial, and postcranial changes across
time ......................................................................................................... 99
Carious lesions ........................................................................................ 101
Mechanism for Dental Reduction .......................................................................... 102 Probable Mutation Effect ................................................................................ 102 Increasing Population Density Effect .............................................................. 103 Selective Compromise Effect ......................................................................... 104 Masticatory Functional Hypothesis ................................................................. 105
Conclusions .......................................................................................................... 105
5 DEVELOPMENTAL INSTABILITY: ENAMEL HYPOPLASIAS ............................. 110

8
Enamel Hypoplasias ............................................................................................. 110
Health and Disease in the Shift to Agriculture ....................................................... 112 Current Study ........................................................................................................ 114
Materials and Methods.......................................................................................... 117 Results .................................................................................................................. 119
Age Differences .............................................................................................. 120 Sex Differences .............................................................................................. 120 Time Period Differences ................................................................................. 121
Discussion ............................................................................................................ 122 Age Differences .............................................................................................. 122 Sex Differences .............................................................................................. 123 Time Period Differences ................................................................................. 124
Broader Implications ............................................................................................. 127
Conclusions .......................................................................................................... 129
6 CARIOUS LESIONS ............................................................................................. 139
Formation of Carious Lesions ............................................................................... 140
Carious Lesions and Agricultural Intensification ................................................... 140 Culture History of Guam ....................................................................................... 142 Materials and Methods.......................................................................................... 145
Results .................................................................................................................. 146 Tooth Position ................................................................................................ 146
Age Differences .............................................................................................. 147 Sex Differences .............................................................................................. 147
Time Period Differences ................................................................................. 148 Discussion ............................................................................................................ 148
Tooth Position ................................................................................................ 148
Age Differences .............................................................................................. 150 Sex Differences .............................................................................................. 150
Time Period Differences ................................................................................. 151 The effects of diet .................................................................................... 152 The effects of betel-nut chewing .............................................................. 156
Conclusions .......................................................................................................... 158
7 SUMMARY ........................................................................................................... 165
APPENDIX
A DENTAL METRICS............................................................................................... 170
B ANALYSES OF VARIANCE TESTS FOR DENTAL MEASUREMENTS .............. 206
LIST OF REFERENCES ............................................................................................. 249
BIOGRAPHICAL SKETCH .......................................................................................... 279

9
LIST OF TABLES
Table page 2-1 Archaeological and historical chronological sequences of the Marianas
Islands ................................................................................................................ 71
2-2 Activities and artifact assemblage composition of latte sets ............................... 71
3-1 Radiocarbon dates from Naton Beach site, Tumon, Guam. ............................... 82
3-2 Pre-Latte vs. Latte sample distribution ............................................................... 83
3-3 Okura dental sex distributions ............................................................................ 83
3-4 Okura dental age distributions ............................................................................ 83
3-5 Pre-Latte dental sample by age and sex ............................................................ 83
3-6 Latte dental sample by age and sex ................................................................... 84
4-1 Tooth summary data ......................................................................................... 107
4-2 Tooth summaries of prehistoric Chamorro populations from Guam ................. 107
4-3 Tooth summaries of Pacific and circum-Pacific samples .................................. 108
4-4 Mean cranial and mandibular measurements associated with masticatory apparatus ......................................................................................................... 109
5-1 Percentage of teeth with one or more linear enamel hypoplasia ...................... 132
5-2 Individual occurrence of Pre-Latte and Latte linear enamel hypoplasias by age grouping..................................................................................................... 132
5-3 Individual occurrence of linear enamel hypoplasias ......................................... 132
5-4 Tooth count of linear enamel hypoplasias ........................................................ 133
5-5 LEH frequencies in comparative populations.................................................... 134
5-6 Age differences of leh using a Pearson Chi-Square Test ................................. 135
5-7 Sex differences of LEH using a Pearson Chi-Square Test ............................... 135
5-8 Pre-Latte and Latte differences in LEH using a Pearson Chi-Square Test ....... 135
6-1 Individual occurrence carious lesion frequencies ............................................. 160

10
6-2 Tooth count of carious lesion frequencies ........................................................ 160
6-3 Individual occurrence carious lesions by age ................................................... 160
6-4 Frequency of carious lesions by tooth class ..................................................... 161
6-5 Frequency of carious lesions by tooth position ................................................. 161
6-6 Pearson Chi-Square Test on carious lesion expression ................................... 162
A-1 Descriptive statistics of dental measurements .................................................. 170
A-2 Descriptive statistics of dental measurements by time period .......................... 172
A-3 Descriptive statistics of dental measurements by time period and sex. ............ 176
A-4 Descriptive statistics of cross-sectional area by time period ............................. 189
A-5 Descriptive statistics of cross-sectional area by time period and sex ............... 190
A-6 Group comparisons of dental measurements ................................................... 193
A-7 Kolmogorov-Smirnova Test for normality .......................................................... 200
A-8 Levene's Test of Homogeneity of Variance based on the mean ....................... 204
B-1 Two-Way Factorial ANOVA for LMax I1 MD ..................................................... 207
B-2 Two-Way Factorial ANOVA for LMax I1 BL ...................................................... 207
B-3 Two-Way Factorial ANOVA for LMax I2 MD ..................................................... 208
B-4 Two-Way Factorial ANOVA for LMax I2 BL ...................................................... 209
B-5 Two-Way Factorial ANOVA for LMax C MD ..................................................... 210
B-6 Two-Way Factorial ANOVA for LMax C BL ...................................................... 211
B-7 Two-Way Factorial ANOVA for LMax P3 MD ................................................... 212
B-8 Two-Way Factorial ANOVA for LMax P3 BL..................................................... 213
B-9 Two-Way Factorial ANOVA for LMax P4 MD ................................................... 214
B-10 Two-Way Factorial ANOVA for LMax P4 BL..................................................... 215
B-11 Two-Way Factorial ANOVA for LMax M1 MD ................................................... 216
B-12 Two-Way Factorial ANOVA for LMax M1 BL .................................................... 217

11
B-13 Two-Way Factorial ANOVA for LMax M2 MD ................................................... 217
B-14 Two-Way Factorial ANOVA for LMax M2 BL .................................................... 218
B-15 Two-Way Factorial ANOVA for LMax M3 MD ................................................... 218
B-16 Two-Way Factorial ANOVA for LMax M3 BL .................................................... 219
B-17 Two-Way Factorial ANOVA for RMax I1 MD .................................................... 219
B-18 Two-Way Factorial ANOVA for RMax I1 BL ..................................................... 220
B-19 Two-Way Factorial ANOVA for RMax I2 MD .................................................... 220
B-20 Two-Way Factorial ANOVA for RMax I2 BL ..................................................... 221
B-21 Two-Way Factorial ANOVA for RMax C MD..................................................... 222
B-22 Two-Way Factorial ANOVA for RMax C BL ...................................................... 223
B-23 Two-Way Factorial ANOVA for RMax P3 MD ................................................... 223
B-24 Two-Way Factorial ANOVA for RMax P3 BL .................................................... 224
B-25 Two-Way Factorial ANOVA for RMax P4 MD ................................................... 224
B-26 Two-Way Factorial ANOVA for RMax P4 BL .................................................... 225
B-27 Two-Way Factorial ANOVA for RMax M1 MD .................................................. 225
B-28 Two-Way Factorial ANOVA for RMax M1 BL ................................................... 226
B-29 Two-Way Factorial ANOVA for RMax M2 MD .................................................. 226
B-30 Two-Way Factorial ANOVA for RMax M2 BL ................................................... 227
B-31 Two-Way Factorial ANOVA for RMax M3 MD .................................................. 227
B-32 Two-Way Factorial ANOVA for RMax M3 BL ................................................... 228
B-33 Two-Way Factorial ANOVA for LMand I1 MD................................................... 228
B-34 Two-Way Factorial ANOVA for LMand I1 BL .................................................... 229
B-35 Two-Way Factorial ANOVA for LMand I2 MD................................................... 229
B-36 Two-Way Factorial ANOVA for LMand I2 BL .................................................... 230
B-37 Two-Way Factorial ANOVA for LMand C MD ................................................... 230

12
B-38 Two-Way Factorial ANOVA for LMand C BL .................................................... 231
B-39 Two-Way Factorial ANOVA for LMand P3 MD ................................................. 232
B-40 Two-Way Factorial ANOVA for LMand P3 BL .................................................. 232
B-42 Two-Way Factorial ANOVA for LMand P4 BL .................................................. 234
B-43 Two-Way Factorial ANOVA for LMand M1 MD ................................................. 235
B-44 Two-Way Factorial ANOVA for LMand M1 BL .................................................. 235
B-45 Two-Way Factorial ANOVA for LMand M2 MD ................................................. 236
B-46 Two-Way Factorial ANOVA for LMand M2 BL. ................................................. 236
B-47 Two-Way Factorial ANOVA for LMand M3 MD ................................................. 237
B-48 Two-Way Factorial ANOVA for LMand M3 BL .................................................. 238
B-49 Two-Way Factorial ANOVA for RMand I1 MD .................................................. 238
B-50 Two-Way Factorial ANOVA for RMand I1 BL ................................................... 239
B-51 Two-Way Factorial ANOVA for RMand I2 MD .................................................. 239
B-52 Two-Way Factorial ANOVA for RMand I2 BL ................................................... 240
B-53 Two-Way Factorial ANOVA for RMand C MD .................................................. 240
B-54 Two-Way Factorial ANOVA for RMand C BL.................................................... 241
B-55 Two-Way Factorial ANOVA for RMand P3 MD ................................................. 242
B-56 Two-Way Factorial ANOVA for RMand P3 BL .................................................. 242
B-57 Two-Way Factorial ANOVA for RMand P4 MD ................................................. 243
B-58 Two-Way Factorial ANOVA for RMand P4 BL .................................................. 244
B-59 Two-Way Factorial ANOVA for RMand M1 MD ................................................ 245
B-60 Two-Way Factorial ANOVA for RMand M1 BL ................................................. 246
B-61 Two-Way Factorial ANOVA for RMand M2 MD ................................................ 246
B-62 Two-Way Factorial ANOVA for RMand M2 BL ................................................. 247
B-63 Two-Way Factorial ANOVA for RMand M3 MD ................................................ 248

13
B-64 Two-Way Factorial ANOVA for RMand M3 BL ................................................. 248

14
LIST OF FIGURES
Figure page 1-1 Model for interpreting stress in skeletal populations ........................................... 31
1-2 The island of Guam and Naton Beach Site location ........................................... 32
2-1 The Mariana Island Chain .................................................................................. 72
2-2 Paleoshoreline notches along the western coast of Guam. ................................ 73
2-3 Reconstructed Latte set from Reinman’s Talofofo River Valley Site. .................. 73
2-4 Examples of Pre-Latte Period pottery ................................................................. 74
2-5 Island of Guam depicting Latte Set density and distribution, following Hornbostel’s original 1920s survey ..................................................................... 75
2-6 Examples of Latte Period pottery ........................................................................ 76
5-1 Single linear enamel hyoplasia in the mandibular lateral incisor, canine, and third premolar. .................................................................................................. 136
5-2 Multiple linear enamel hypoplasias in a single tooth. ........................................ 136
5-3 Labial abrasion in a Pre-Latte Period individual................................................ 137
5-4 Betel-nut staining in a Latte Period individual. .................................................. 137
5-5 Dental incising in a Latte Period Individual. ...................................................... 138
5-6 Frequency of linear enamel hypoplasias by tooth type .................................... 138
6-1 Pre-Latte carious lesions in the anterior dentition. ............................................ 164
6-2 Pre-Latte dental crowding. ................................................................................ 164
6-3 Betel-nut with piper leaf and slacked lime. ....................................................... 164
15
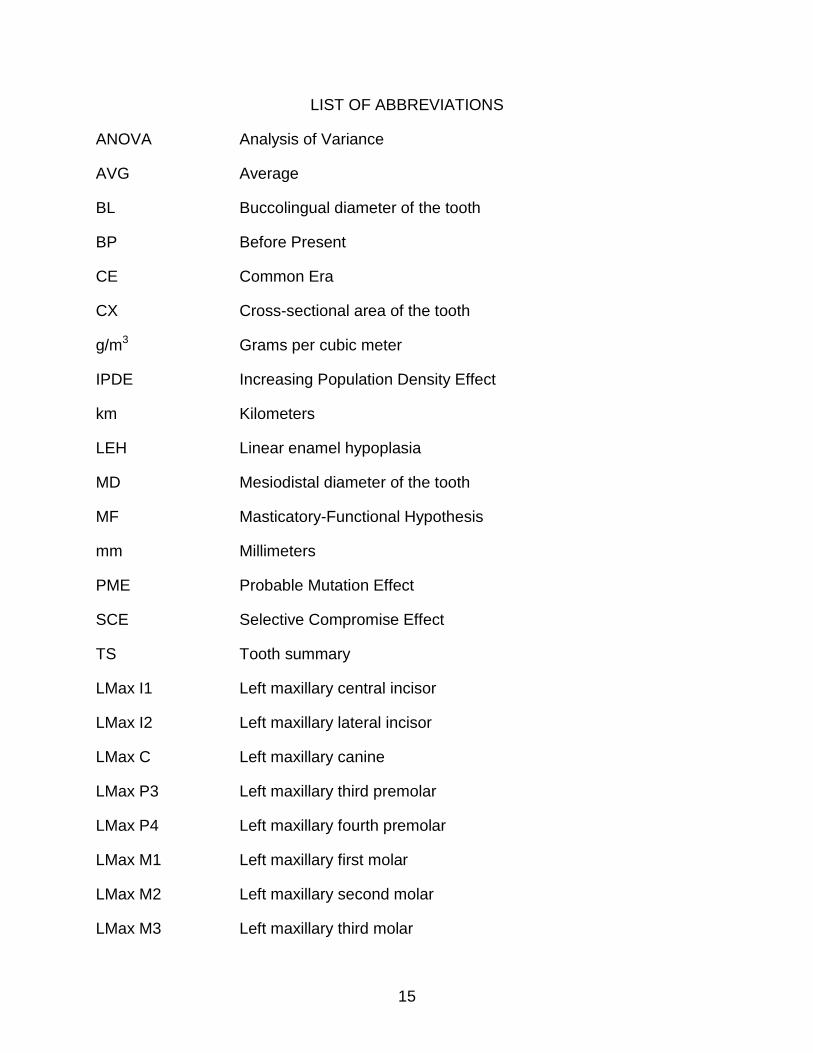
LIST OF ABBREVIATIONS
ANOVA Analysis of Variance
AVG Average
BL Buccolingual diameter of the tooth
BP Before Present
CE Common Era
CX Cross-sectional area of the tooth
g/m3 Grams per cubic meter
IPDE Increasing Population Density Effect
km Kilometers
LEH Linear enamel hypoplasia
MD Mesiodistal diameter of the tooth
MF Masticatory-Functional Hypothesis
mm Millimeters
PME Probable Mutation Effect
SCE Selective Compromise Effect
TS Tooth summary
LMax I1 Left maxillary central incisor
LMax I2 Left maxillary lateral incisor
LMax C Left maxillary canine
LMax P3 Left maxillary third premolar
LMax P4 Left maxillary fourth premolar
LMax M1 Left maxillary first molar
LMax M2 Left maxillary second molar
LMax M3 Left maxillary third molar

16
RMax I1 Right maxillary central incisor
RMax I2 Right maxillary lateral incisor
RMax C Right maxillary canine
RMax P3 Right maxillary third premolar
RMax P4 Right maxillary fourth premolar
RMax M1 Right maxillary first molar
RMax M2 Right maxillary second molar
RMax M3 Right maxillary third molar
LMand I1 Left mandibular central incisor
LMand I2 Left mandibular lateral incisor
LMand C Left mandibular canine
LMand P3 Left mandibular third premolar
LMand P4 Left mandibular fourth premolar
LMand M1 Left mandibular first molar
LMand M2 Left mandibular second molar
LMand M3 Left mandibular third molar
RMand I1 Right mandibular central incisor
RMand I2 Right mandibular lateral incisor
RMand C Right mandibular canine
RMand P3 Right mandibular third premolar
RMand P4 Right mandibular fourth premolar
RMand M1 Right mandibular first molar
RMand M2 Right mandibular second molar
RMand M3 Right mandibular third molar

17
Abstract of Dissertation Presented to the Graduate School of the University of Florida in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy
A DIACHRONIC ASSESSMENT OF HEALTH AND DISEASE FROM THE ADULT
DENTITION OF THE NATON BEACH BURIAL COMPLEX IN TUMON BAY, GUAM
By
Nicolette M.Parr August 2012
Chair: Michael Warren Major: Anthropology
The current study is an investigation of the prehistoric Chamorro in Guam to
assess health and disease patterns over time. The transition from the Pre-Latte to Latte
periods displays a shift from horticultural to early agricultural practices; accompanying
changes include increased population size and technologically advanced food
processing and preparation techniques. These changes occur concomitantly with large-
scale environmental and climatic fluctuations. It is predicted that the cultural and
environmental shifts will be accompanied by biological ones, due to increased stress
levels associated with malnutrition, limited access to resources, and increased
prevalence of disease.
Analyses of odontometrics, linear enamel hypoplasias, and carious lesions were
performed and analyzed in concert with skeletal data collected by other researchers to
construct a health profile of the prehistoric populations in Guam. Expected results
include dental reduction over time coupled with an increase in linear enamel
hypoplasias and carious lesions.
The dentition display an 8% decrease in size from the Pre-Latte to Latte periods.
Increased reliance on starchy crops would have led to selection for smaller dentition to

18
minimize carious lesions. Additionally, sophistication in food processing techniques
decreases the force necessary to break down tough food, leading to reduced functional
demands of the masticatory apparatus. Thus, this finding is best explained by a
combination of the Selective Compromise Effect and Masticatory-Functional
Hypothesis.
Significant differences in linear enamel hypoplasia expression are noted with an
increase over time. While not significant, the data suggests that there may have been
differential access to resources as a result of gender roles associated with food
procurement, where the females in the Latte period were much more highly susceptible
to physiological stress than the males.
Carious lesions are significantly different over time; however, these findings do not
follow the predicted pattern. Caries frequency in the Latte period decrease over time
likely due to the cultural practice of betel-nut chewing, which has cariostatic properties.
This study expands on the current knowledge of prehistoric health in Guam by
demonstrating an overall decrease in health over time as a result of climatic instability
and subsequent dietary transitions.

19
CHAPTER 1 INTRODUCTION
In these Proes1…these people sail in those seas from Island to Island for several hundred Leagues, the Sun serving them for a compass by day and the Moon and Stars by night. When this comes to be prov’d we Shall be no longer at a loss to know how the Islands lying those Seas came to be people’d.
—Captain James Cook, The Journals of Captain James Cook on His Voyages of Discovery (in Beaglehole, 1955)
Since the dawn of European exploration into the Pacific, Captain James Cook, and
others, pondered how the earliest peoples came to inhabit the most remote of the
Pacific islands. This journey across thousands of kilometers of ocean to reach small
landmasses represents the last migration into uncharted territory (Spate, 1979) and is a
remarkable feat of ‘maritime discovery, colonization, and adaptation’ (Russell, 1998:
67).
The relative isolation of the Pacific Islands makes their populations ideal subjects
for evolutionary studies (Howells, 1973). Small populations, as are often found on
islands, are more sensitive to random genetic changes in comparison to large
populations (Turner, 1987). Thus, populations on small, isolated islands are more likely
to undergo more noticeable phenotypic modification due to environmental changes and
subsequent selective pressures (Houghton, 1991a). Additionally, the diversity of
climate and environment throughout the Pacific Islands provide “the possibility of
investigating on a broad comparative scale the effect of environment on phenotypic
development” (Goodenough, 1957: 154).
1Cook is referring to proas, maritime sailing vessels used in the Pacific.

20
The Pacific is the focus of much bioarchaeological research; however, Guam and
its people, the Chamorro, are underrepresented in the scientific literature, in comparison
with other island groups. As Howells noted in 1973: ‘many books have been written
about where the Polynesians came from but nobody cares a straw where the
Guamanians came from. And yet it is probable that they can tell at least as much about
the peopling of the Pacific as can the Polynesians’ (p. 248). Almost 40 years later, the
paucity of skeletal research in Guam still remains, regardless of evidence that places
the earliest colonization of Remote Oceania in the Marianas Islands (Carson, 2008;
Clarke et al., 2010; Carson, 2010).
Most research in Guam is often contracted out to specialists by cultural resource
management firms (Howells, 1973; 1989; Pietrusewsky, 1990; Houghton, 1996), and
many studies combine collections from different time periods while overlooking
important differences that may be gleaned from a more thorough diachronic study of its
populations. Exposure of the remains to a tropical environment over a long period of
time leads to poor preservation and high fragmentation of the skeletal remains, thus
explaining the dearth of skeletal research in Micronesia (Hanson and Butler, 1997).
Burial reports, often difficult to procure, contain the majority of the details, discussions,
conclusions, and raw data regarding these remains (e.g. Graves and Moore, 1985;
Hanson, 1991; Heathcote, 1991; Douglas and Ikehara, 1992; Heathcote, 1994;
Pietrusewsky and Ikehara- Quebral, 1994; Ikehara-Quebral, 1998; Pietrusewsky, 1988;
Ikehara-Quebral, 1999; Heathcote, 2006).
Few published articles on bioarchaeological research in the Marianas are
available, however and most are restricted to a single published volume (vol. 104, 1997)

21
in The American Journal of Physical Anthropology (Ambrose et al., 1997; Arriaza, 1997;
Douglas et al., 1997; Hanihara, 1997; Hanson and Butler, 1997; Hanson and
Pietrusewsky, 1997; Ikehara-Quebral and Douglas, 1997; Ishida and Dodo, 1997;
Pietrusewsky et al., 1997; Stodder, 1997); while others are scattered throughout
additional peer-reviewed journals (e.g. Leigh, 1930; Rothschild and Heathcote, 1993;
Rothschild and Heathcote, 1995; Rothschild and Rothschild, 1995) or in small Pacific-
based publications (e.g. Underwood, 1973, 1976; Houghton, 1991b; Heathcote et al.,
1996; Suzuki, 1986; Pietrusewsky, 1990; Turner, 1990; DeFant, 2008). Furthermore,
the majority of these studies focus primarily on craniometric data for population history
and health and disease of the Chamorro population. Few studies concentrate on the
dentition (Leigh, 1930; Brace et al., 1981; Brace et al., 1990; Hanihara,1990; Turner,
1990; Turner, 1992, Heathcote, 1994), even though the teeth are often the most highly
preserved portion of human skeletal remains, in pre-historic and archaeological
contexts, making them an excellent repository of biological information (Brace et al.,
1987; Kieser, 1990; Hillson, 1996).
Previous dental studies address biological distance between geographical groups
of the Pacific region (Brace et al., 1981; Brace et al., 1990; Hanihara and Ishida, 2005).
However, these studies combine disparate time periods in their population analyses,
thus obscuring important differences that may be inferred through a more fine-grained
analysis of temporal variation.
Theoretical Framework
This research adopts a biocultural approach, which combines biological, cultural,
and archaeological data to analyze adaptations associated with subsistence patterns
and health status in prehistoric populations (Armegalos and Van Gerven, 2003).

22
Central to the biocultural analysis of human health and disease is Goodman and
Armelagos’ (1989) model for investigating stress in prehistoric skeletal assemblages,
which will be discussed in further detail below.
Biocultural Approach to Bioarchaeology
Biocultural studies focus on the population, rather than individual typological traits,
as the unit of evolutionary change within the environment (Wellin, 1978). This method
uses integrative thinking to explain the adaptation process as a ‘mechanism for
responding to environmental stressors’ (McElroy, 1990: 246), while creating testable
models to understand interrelatedness of biological, cultural, and environmental
variables, as well as behavioral patterning occurring within a population (Blakely, 1977).
Thus, evaluating information gathered from biological, archaeological, and cultural
contexts could provide much needed insights to dynamic evolutionary processes and
adaptations in prehistory.
In the 1970s the term bioarchaeology was coined independently within the United
Kingdom (U.K.) and the United States (U.S.), each with different definitions. Whereas in
the U.K. the term was originally associated with the study of faunal remains (Clark,
1972), bioarchaeology as defined in the U.S., was coined by Buikstra in an edited
volume by Blakely (1977a), Biocultural Adaptation in Prehistoric America.
Bioarchaeology as it is known in the U.S., and now in other parts of the world, stems
from the emergence of Binford’s New Archaeology (1962), with its focus on a synthetic
approach to population-based, ecological research (Buikstra, 1997; Buikstra and Beck,
2006). Bioarchaeology can be defined as a contextual study of human skeletal remains
from archaeological contexts, which integrates biology, culture, and environment to
better understand health, disease, and demography in prehistoric human populations

23
(Buikstra; 1977; Larsen, 1997; Buikstra and Beck, 2006). Unlike osteological
investigations where skeletal data were largely typological and descriptive, the
bioarchaeological approach emphasizes problem-oriented research to reconstruct
prehistoric lifeways through the integration of biological and archaeological data
(Buikstra, 1977).
Bioarchaeologists often take a biocultural approach to understanding evolutionary
dynamics allowing for a holistic analysis of prehistoric populations and their interactions
with the surrounding environment through a synthesis of biology and culture (Blakely,
1977). Blakely (1977b: 1) states that ‘humans survive not through cultural adaptation
nor biological adaptation, but through biocultural adaptation’ (emphasis his). Recent
studies emphasize a biocultural approach by combining archaeological and osteological
research to answer significant questions about adaptation and the forces that drive
biological change such as weaning and dietary shifts in Guatemala (Wright and
Schwarcz, 1999); demographic collapse in Spanish Florida (Griffin and colleagues,
2001; Stojanowski, 2003; 2005); climatic variability in Japan (Temple, 2007; Temple and
Larsen, 2007; Oxenham and Matsumura, 2008; Temple, 2010); ecological and
demographic pressure in the Nile Valley (Starling and Stock, 2007), and mobility in
Northern Africa (Stojanowski and Knudson, 2011).
Stress Models
Early approaches to the study of stress focused on how external and
environmental parameters place strain on a given organism (Goodman et al., 1988). In
the middle of the twentieth century, Hans Selye (1936, 1956, 1973) introduced a new
perspective on the study of stress where the focus shifted from strain as a result of
environmental stressors to looking at physiological change (i.e., health and

24
development) due to a ‘state of stress’ (Goodman et al., 1988). Selye (1973: 692)
defines stress as ‘the nonspecific response of the body to any demand made upon it’
where alterations of the environment alters the normal and steady state of an organism.
Central to the Selyean concept of stress is the general adaptation syndrome, a defense
mechanism in which homeostatic mechanisms are activated to alleviate a long and
continued stress event (Selye, 1956). Classic laboratory studies showed that increased
exposure to stressors (i.e., noise, cold, and heat) lead to developmental disturbances in
rodents (Siegel and Smookler 1973; Siegel and Doyle 1975a,b; Siegel et al. 1977;
Doyle et al. 1977; Sciulli et al. 1979). Recently rodent studies have demonstrated that
stress stimuli impairs memory (Luine et al., 1994; Conrad et al., 1996); alters
cardiovascular activity (Rudyk et al., 2001), and affects sexual maturity and weight
(Rodriguez et al., 2007). In terms of human skeletal and dental remains, nonspecific
indicators of stress represent an adaptive response to a stressor, which occurred during
the development of an individual (Roberts and Manchester, 2005).
In Goodman and Armelagos’ (1984) seminal volume Paleopathology at the Origins
of Agriculture, the authors present a model for stress applicable to skeletal populations
in which health is the fundamental variable in examining the adaptive processes of an
individual or population (Larsen, 1997; Goodman and Martin, 2002). Subsequently, the
model has been reworked by Goodman and colleagues to include feedback systems
and indicators of stress in the skeleton and provides a systematic framework to analyze
the effects of a physiological disruption (Goodman 1991; Goodman and Armelagos,
1989; Goodman et al. 1988; Goodman and Martin, 2002).

25
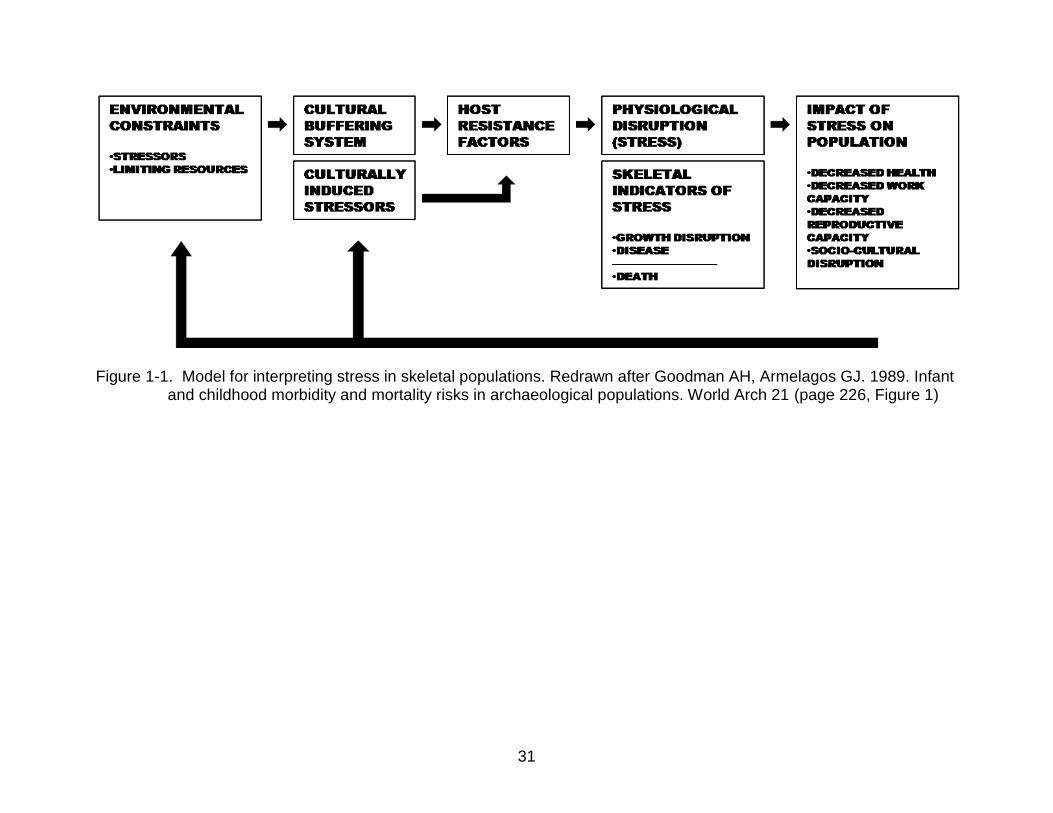
The model presented in Figure 1-1 begins with the environment, which provides
certain necessary resources (e.g., food, shelter, and water) as well as stressors (e.g.,
extreme temperatures, parasites, predators) that may affect the health of a population
(Goodman and Armelagos, 1989; Goodman and Martin, 2002). Technological
advances in culture, such as clothing and shelter, may often provide a buffer to
stressors. In cases where the cultural system does not adequately buffer the
environmental constraints, the stressors will reach the individual or population.
Adaptation to a stressor depends on each individual’s host resistance. No biological
impact will be noted in individuals who can combat any given stressor, however, some
individuals may be unable to resist the stressor, due to genetic susceptibility, disease,
malnutrition, age, sex, and/or resiliency (Goodman et al., 1988; Goodman and Martin,
2002).
The severity of and duration of the stress response may be viewed as a function of the degree of cultural and environmental constraints and stressors, balanced against the adequacy of the cultural buffering system and individual resistance resources (Goodman and Martin, 2002: 18).
If an individual fails to fight a stressor, a physiological disruption or biological stress
response may occur, resulting in permanent and visible changes in the body (Goodman
et al., 1988; Goodman and Armelagos, 1989; Larsen, 1997; Goodman and Martin,
2002). Soft tissue responds more quickly to stress and disease than the skeleton.
Therefore, a stress event must be severe or endure for a prolonged period before the
bone or teeth are affected (Goodman et al., 1988).
Interpreting of skeletal markers of stress is difficult and while some diseases,
such as tuberculosis, syphilis, and leprosy, leave diagnostic lesions on the skeleton,
many other pathogens elicit the same response for a number of given stressors

26
(Goodman et al., 1998). For example, periostitis is one of the most common diseases
indicative of trauma or infection; however, periosteal bone formation occurs in a number
of other infectious diseases (Ortner, 2003). As such, identification of an infection in the
skeleton may seem simple; however, diagnosis of its etiology proves quite difficult.
Additionally, some diseases, such as influenza and other viruses that may result in
decreased health and in some cases death, leave no evidence in the skeleton, which
confounds the interpretation of health and disease from skeletal remains (Goodman et
al., 1998).
In what is now known as the Osteological Paradox, Wood and colleagues (1992)
criticized the conclusions that stemmed from Goodman and Armelagos’ (1984) volume
regarding population health in the transition to agriculture. The authors identified key
conceptual problems that complicate interpretations of health from skeletal remains.
Most pertinent to the current study is the concept of hidden heterogeneity, which refers
to the amount of frailty of any given individual – in essence, how prone an individual is
to disease and death. Individuals who are more are most frail may succumb quickly to
external stressors, leaving no markers of bony response of the disease process on the
skeleton, while others who are exposed to moderate stressors, may survive through the
stress event and thus elicit skeletal markers of stress.
Some studies, however, have shown correlations between disadvantaged
populations and disease. For example, studies in living human and non-human primate
populations have shown that individuals with a higher prevalence of linear enamel
hypoplasias are not at a greater advantage than those without (Zhou and Corruccini,
1998; Guitelli-Steinberg and Benderlioglu, 2006). Nonetheless, since its publications,

27
researchers have suggested several methods to correct for issues related to the
Osteological Paradox. Goodman (1993) suggests that an analysis of multiple stressors,
instead of an individual trait, would reduce the likelihood of misinterpreting health status.
Ideal stress indicators are those associated for use with the health index as proposed
by Steckel and Rose (2002): stature, hypoplasias, anemia, dental health, skeletal
infections, degenerative joint disease, and trauma (Goodman and Martin, 2002).
Furthermore, an interdisciplinary approach to health status may prove helpful in an
interpretation of skeletal lesions in archaeological populations. Larsen (1997: 337)
states that in order to get a clear evaluation of population health, biological indicators of
disease need to be evaluated with “other lines of evidence, including subsistence and
settlement, environmental context, cultural context, and population structure.” Thus, an
analysis of the cultural and environmental factors in association with biological
indicators of stress will provide biocultural understanding of adaptation in archaeological
skeletal samples.
Purpose and Research Objectives
The current study investigates evolutionary dynamics of the prehistoric Chamorro
across time to see how they relate to biocultural and environmental changes in
prehistoric society. Between the Pre-Latte and Latte time periods in Guam, there are
changes in population size and subsistence strategies (Hunter-Anderson and Butler,
1991). Likewise, they changed many of their food procurement and preparation
strategies (Amesbury, 1999; Moore, 2005; Amesbury, 2007). These transitions occur
concomitantly with large-scale environmental and climatic fluctuations such as sea-level
decline and increased storminess, aridity, and drought (Hunter-Anderson and Butler,
1991); Nunn, 2007, Hunter-Anderson, 2010). It is predicted that these cultural and

28
environmental modifications will be accompanied by biological ones. For example,
increase in stress levels associated with malnutrition, limited access to resources, and
increased prevalence of disease, may be evidenced in the dentition as dental reduction,
linear enamel hypoplasias, and carious lesions.
In this study, I adopt a diachronic approach to assess change in the dentition as it
correlates to environmental and cultural changes over time. A diachronic analysis of the
dentition allows for an investigation into biological processes that can lead to biological
change in human populations. An analysis such as this may uncover very small
changes that occur between the two time periods that are often lost in broader studies
that do not take temporal differences into account. Bellwood (1989: 4) emphasizes the
need to evaluate processes that lead to cultural and linguistic diversification such as
“invention, environmental limitation or encouragement, founder influence on small island
cultures, drift through isolation or communication decline, and diffusion/borrowing.”
These temporal changes in Guam represent the type of transitions that Bellwood feels
are needed to analyze biological processes.
Objectives and Hypotheses
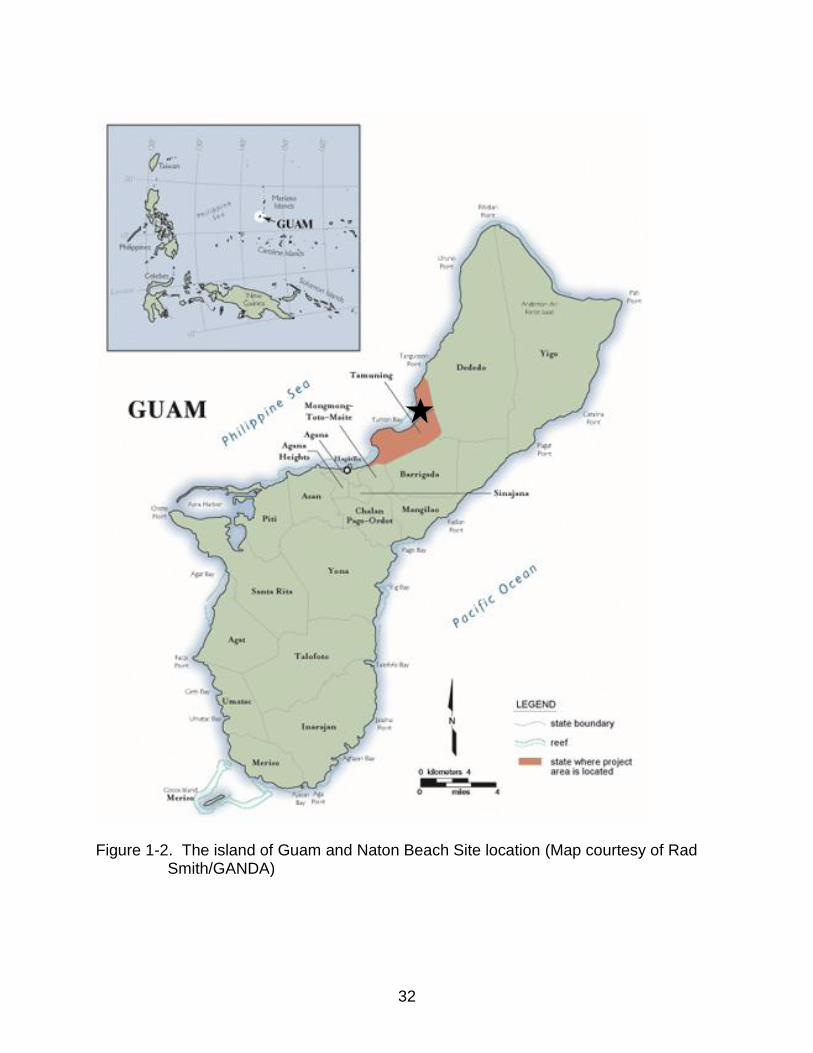
The current study focuses on the Naton Beach mortuary sample, which was
excavated in response to cultural resource management litigation.2 This sample
includes both the Pre-Latte (n = 103) and Latte periods (n = 112) and is located on the
west coast of Guam, in the Western Pacific, in Northern Tumon Bay at the Naton Beach
location (Figures 1-2). Four 14C dates were obtained from conus shell bead necklaces
associated with the earliest burials and range from 2790 to 2330 BP (DeFant, 2008).
2The Naton Beach site is often referred to colloquially as the Okura site in response to its location at the
previous Okura Hotel, which was subsequently renovated to become the Guam Aurora Villas & Spa.

29
This sample represents the largest Pre-Latte skeletal assemblage to be excavated on
Guam and one of the oldest and largest in the remote Western Pacific. Dates from the
Chelechol ra Orrak cemetery in Palau (also in the Western Pacific) range from 3000 BP
to 200 CE (Fitzpatrick, 2003; Fitzpatrick and Nelson, 2008) placing it either slightly
earlier or contemporary to the Naton Beach site; however, the sample size is limited to
26 individuals (Nelson and Fitzpatrick, 2006; Fitzpatrick and Nelson, 2011).
An overall review of the known settlement history of the Pacific and the Marianas
Islands will be presented using four independent lines of evidence: linguistics,
archaeology, genetics, and bioarchaeology. Evidence of cultural change between the
Pre-Latte and Latte time Periods will be evaluated in the archaeological record in an
attempt to define triggers that may have led to change. These cultural changes will be
analyzed in concert with the dental data to determine if biological change has occurred
following cultural shifts. The hypotheses are as follows:
Hn1: There is no significant difference in the dental dimensions between the Pre-Latte and Latte time periods.
Hn2: There is no significant difference in the frequency of linear enamel hypoplasias between the Pre-Latte and Latte time periods.
Hn3: There is no significant difference in the frequency of carious lesions between the Pre-Latte and Latte time periods.
A focus on temporal differences in Guam will help clarify health and disease patterns in
the prehistoric Chamorro, which until recently, was only known for the late prehistoric
peoples. Analysis of the dental data are combined with other indicators of disease from
the postcranial skeleton, gleaned from published and unpublished reports, and
evaluated within the broader frame of the changing ecosystems that coincides with the
Pre-Latte and Latte transition. Additionally, subsistence adaptations are analyzed to

30
determine the impact of agricultural intensification on the oral health of the Chamorro. If
dental reduction, linear enamel hypoplasias, and carious lesion differences are found,
this study will delineate causative factors associated with these changes through further
analysis of the archaeological record. In conclusion, an analysis of dental changes in
the prehistoric Chamorro will elucidate a shift in not only within-island phenomena but
also provide a framework for interpreting biocultural interactions of the Chamorro and
the dynamic environment in which they lived.
Chapter Organization
This dissertation is organized into seven chapters. The current chapter includes
an introduction to the study, outlines the theoretical goals, and provides a brief overview
of the site location. It also outlines the research problem and presents three key
hypotheses to be tested using the recovered skeletal remains. The second chapter
details the natural and cultural history of Guam, beginning with the study location,
paleogeography/environment/fauna, terminating with a review of the settlement history
of the Pacific and the Marianas Islands. In Chapter 3, the sample materials are
described with a focus on taphonomic biases and population demography. The fourth,
fifth, and six chapters report the background, data collection methods, including
statistical procedures, results, and discussion of the research. In Chapter 4,
odontometric analyses are performed to elucidate differences between the populations
with a focus on mechanisms for dental reduction over time. Physiological stress, as
evidenced by linear enamel hypoplasias, is analyzed in Chapter 5 and highlights trends
associated with climatic variability between the populations. The seventh and final
chapter discusses the study as a whole and identifies future studies that can be
conducted for a more holistic interpretation of the lifestyles of the prehistoric Chamorro.
31
Figure 1-1. Model for interpreting stress in skeletal populations. Redrawn after Goodman AH, Armelagos GJ. 1989. Infant and childhood morbidity and mortality risks in archaeological populations. World Arch 21 (page 226, Figure 1)
32
Figure 1-2. The island of Guam and Naton Beach Site location (Map courtesy of Rad Smith/GANDA)

33
CHAPTER 2 NATURAL AND CULTURAL ENVIRONMENT
Study Location
The Mariana Islands are an archipelago in Micronesia that forms a chain of 15
islands extending north-south between 13° and 20° N latitude. These islands are
located approximately 2200 km southeast of Japan and approximately 6000 km west of
Hawai’i (Thompson, 1932) (Figure 2-1). The Marianas lie west of the Marianas Trench
Subduction Zone, where the Pacific and Philippine tectonic plates meet. The larger
Pacific Plate is subsumed beneath the Philippine plate (Rainbird, 1994; Steadman,
2006). The five southern islands (Guam, Rota, Aguiguan, Tinian, and Saipan) are the
oldest and largest of the chain and are composed primarily of raised limestone, while
the ten northern islands are volcanic in nature, eight of which are still active (Steadman,
2006).
Guam is the southern-most and largest of the islands forming the Marianas chain
and is approximately 50 km long and ranges between 6 and 19 km wide with an area of
approximately 554 square km (Thompson, 1932; Karolle, 1993; Mylroie et al., 2001;
Gingerich, 2003). Geologically, Guam is divided by the Pago-Adelup fault line, which
separates the northern low relief limestone plateau and southern volcanic cuesta with
an uplifted limestone component on the eastern coast (Tabrosi et al., 2005).
Natural Environment
A basic understanding of the natural environment of Guam is necessary to
elucidate the complexity of human cultural adaptations over time, particularly when
evaluating the archaeological record. This overview provides insight into environmental
factors that may have led to biocultural changes in the Chamorro.

34
Biogeographical Divides
In the 17th century, the French voyager, Dumont d’Urville (1832) created a tripartite
classification system for the peoples and islands of the Pacific (Melanesia, Micronesia,
and Polynesia) based on typological physical characteristics of incredibly diverse and
heterogeneous peoples (Green 1991; Kirch, 2000; 2010). d’Urville’s partitions have no
utility in terms of biological, cultural, and historical processes, save for Polynesia which
has proven more homogeneous, both culturally and historically (Kirch, 2000; 2010);
nonetheless, his classification system has become ingrained in Western thought and
the terms continue to be utilized today (Thomas, 1989). As with most broad
geographical groups throughout the world, the populations of the Pacific are not
linguistically, culturally, and biologically homogeneous; instead variation across the
Pacific displays clinal trends rather than sharp boundaries (Thomas 1989; Terrell 1986;
Bellwood 1989).
While not particularly useful from a biological standpoint, d’Urville’s classification
scheme is geographically significant. Melanesia, ‘the dark islands’, contains the largest
land masses in the Pacific and includes New Guinea, the Solomons, Vanuatu
(previously known as New Hebrides), New Caledonia, and Fiji (d’Urville,1832; Kirch,
2010). Micronesia, ‘the little islands’, is composed of approximately 21,000 islands and
encompasses five main archipelagos: Palau (Belau), the Marianas, the Carolines, the
Marshalls, and Kiribati (formerly the Gilbert Islands) (d’Urville, 1832; Kirch 2010).
Despite the vast number of islands, the totality of Micronesia’s land mass is only ~2,700
km2, with Guam being the largest of the islands (582 km2) (Kirch, 2010). Lastly,
Polynesia, ‘many islands’, encorporates the far eastern Pacific islands, including

35
Hawai’i, Rapa Nui (Easter Island), Tahiti, and New Zealand (d’Urville, 1832; Kirch
2010).
Green (1991) suggests replacing d’Urville’s terms with Near Oceania, to include
New Guinea, the Bismarck Archipelago, and the Solomon Islands, and Remote
Oceania, which encompasses the entirety of the Pacific islands east of the Solomon
Islands. This terminology is heavily rooted in archaeological findings that have
demonstrated the antiquity of human settlement in Near Oceania extending into the
Pleistocene, compared to Remote Oceania with earliest colonization ranging from 4,000
years ago, in the Western Pacific, to 1000 years ago in the Eastern Pacific (Green,
1991; Kirch, 2000; 2010). Thus, the suggested and continued use of Green’s
classification scheme is in better standing with current linguistic, biological, and
archaeological data.
Paleogeography
Guam has undergone dramatic changes in coastline due to changes in sea level
and bioturbation from storm and wave activity (Bath, 1986; Kurashina and Clayshulte,
1983; Dickinson, 2000; 2003; Carson, 2011). During the mid-Holocene highstand
(~6000 and 4000 BP) sea level elevations ranged between 1.6 m to 2.6 m above
modern day sea levels in the tropical Pacific Islands (including Mariana and eastern
Caroline Islands, Samoa, Fiji, Tonga, and Molokai) (Dickinson, 2001; 2003). Atolls,
barrier reefs and most land masses in the Pacific Ocean were completely submerged
except for the highest volcanic ridges (Dickinson, 2001; Amesbury and Hunter-
Anderson, 2008). After the mid-Holocene, sea level decrease began in 2200 BP, in the
northwest and southwest pacific; however in the eastern boundaries of the Pacific, sea

36
levels did not begin to decline until ~0 CE, and in some cases as late as 800 CE
(Dickinson, 2003).
In Guam, significant sea level changes can be measured directly from the wave-
cut notches of exposed limestone faces and emergent mid-Holocene reef flats (see
Figure 2-2), indicating that between 5400 and 3050 BP the sea level was over 1.8 m
higher than in the present day (Easton et al., 1978; Dickinson, 2000; 2003; Kayanne et
al., 1993). This finding is in accordance with the mid-Holocene highstand estimate
(Dickenson, 2003). Archaeological findings have placed arrival of the earliest colonizers
~3500 BP (see expanded settlement discussion below), which coincides with the mid-
Holocene highstand. Thus, settlers encountered high sea level paleoshorelines with
fringing reefs, coastal flats, mangrove-lined lagoons, stable islets, and estuaries
(Dickinson, 2003; Amesbury and Hunter-Anderson, 2008). Approximately 300 years
after settlement, in 3200 BP, the post-mid-Holocene sea level decline changed the
appearance of the coastline by expanding preferable habitation areas into wide sandy
beaches along the coast and also allowed for more widespread dispersal of inhabitants
throughout the island (Amesbury et al., 1996; Dickinson, 2003). Archaeological studies
in Tumon Bay have found evidence of coastal progradation associated with sea level
decline (Graves and Moore, 1985; Bath, 1986; Olmo, 1997; Magnuson et al., 2000).
Additionally, this sea-level decline necessitated cultural adaptations of the
population to new environmental conditions (Carson, 2011). Understanding sea level
and ecosystem changes in Guam helps clarify spatial differences in the locations of Pre-
Latte and Latte sites as well as variation of shellfish exploitation between the periods.
Prograding coastlines and sea-level drawdown explains why earlier Pre-Latte sites are

37
usually found more inland in comparison with the later Latte sites: the sandy beach
areas, closest to the present shoreline, had not yet emerged in the Pre-Latte times
(Bath, 1986; Graves and Moore, 1985; Amesbury, 1999; Carson, 2011). Additionally,
the shift from bivalve to gastropod consumption may be an indirect result and
subsequent cultural adaptation due to altered ecosystems and the disruption of
mangrove swamps, which are the preferred habitat of bivalves (Amesbury, 1999).
Thus, data on sea level changes, coupled with archaeological data from prehistoric
habitation sites, has shed light on the relationship between the dynamic alterations of
paleocoastlines and associated cultural changes (Carson, 2011).
Paleoenvironment
The paleoenvironmental record has proven difficult to interpret, however, several
researchers provide valuable data to better understand prehistoric environmental
conditions (Athens and Ward, 1993; 1995; 1999; 2004; Ward, 1994; 1995; Nunn, 1999;
Nunn, 2007; Nunn et al., 2007). The mid-Holocene highstand corresponds with the
Holocene Climatic Optimum (HCO) in the Pacific region, which occurred between 6000
and 3000 BP, where higher sea levels, temperatures, and an abundance of organisms
allowed for increased diversity in habitat (Nunn, 1999). Since the end of the HCO
‘cooling, sea-level fall and, in places, a fall in precipitation and loss of biodiversity’
attributed to climate change (Nunn, 2007: 2). Cool temperatures remained stable until
~AD 750, during the Little Climatic Optimum (LCO - also known as the Medieval Warm
Period), when temperatures began to rise slowly, rainfall decreased, and sea levels
once again rose, until approximately AD 1300 (Nunn, 2007; Nunn et al., 2007).
The transitional phase from AD 750 to 1300 is known as the ‘AD 1300 Event’,
where rapid cooling temperatures, decline in sea levels, and increased storminess

38
resulted in greater climatic variability, during this Little Ice Age (Bridgman, 1983; Nunn
and Britton, 2001; Nunn, 2007; Nunn et al., 2007). The AD 1300 Event may well be the
‘most rapid period of climate change to have occurred within the past several millennia’
(Nunn, 2007: 1) and is associated with societal disruption, subsistence change, and
movement to inland habitation areas (Nunn, 2000; Nunn and Britton, 2001; Nunn,
2007). This pattern is observed throughout the Pacific Basin (Nunn, 2000; Nunn and
Britton, 2001; Nunn, 2007). Further, the dramatic ecosystem fluctuations between the
LCO and the Little Ice Age, associated with the AD 1300 Event, correspond with the
shift from the Pre-Latte to Latte time period and concomitant cultural modifications.
Wetland sedimentary cores conducted at various locations throughout Guam have
generated a continuous record of paleoclimate and vegetation changes of the island
landscape that predates human settlement (Ward, 1994; 1995; Athens and Ward, 1993;
1995; 1999; 2004). Pollen analysis from the IARII Laguas Core (Athens and Ward,
1999; 2004), which is the most detailed and complete paleoenvironmental record from
Guam, indicates that this island was largely forested during the early Holocene.
Between 4405 and 2956 cal. BP, forest and swamp/mangrove taxa begin to decline in
conjunction with the arrival of the first human settlers. Likewise, Lycopodium and
Gleichenia ferns begin to appear, circa 3,900 cal. BP, indicating possible gardening and
resource collecting. By 2900 cal. BP, ferns, grasses, and charcoal are in abundance,
suggesting a shift to a savannah-like habitat of open areas with grass cover, possibly
augmented by both intentional and unintentional fires. By 2300 cal. BP, very little of the
native forest persisted on the island; instead the majority of the island had been
converted to the savanna landscape typical of modern day Guam.

39
Paleofauna
Research by Pregill and Steadman (2009) has provided valuable information on
the prehistoric fossil record in Guam and documents the extensive faunal loss due to
human colonization and the resulting habitat destruction, human predation, and
introduction of exotic predatory animals. Terrestrial vertebrates were collected from two
caves, Ritidian Cave 1 and Gotham Cave, both located on the northernmost area of
Guam. Ten native reptiles were identified as well as two prehistorically introduced
species: the mangrove monitor lizard (Varanus indicus), introduced around 2900 BP
(Liston et al., 1996; Wiles et al., 1989), and blind snakes (Ramphotyphlops sp.). Of the
10 native reptiles identified, the gekkonid lizard (Gekkonidae new sp.) is extinct.
Seventeen species of bird were identified, of which five are currently extinct (Duck:
Anas oustaletii; Rail: Porzana undescribed sp.; Parrot: cf. Vini undescribed sp.; White-
eye: Zosteropidae new sp.), two are extirpated, and eight lost in historic times. Pregill
and Steadman (2009) also found evidence for the introduction of the rat, Rattus rattus,
circa 800 to 1000 CE, approximately 2000 years after human colonization and
corresponding to the shift between the Pre-Latte and Latte time periods. However, no
chicken, dog, or pig remains were found in the prehistoric skeletal assemblage in
Guam, although they have been found in nearly all other Pacific Islands (Wickler, 2004),
Far more lizard species have survived into modern times in comparison to birds.
Extinction of native lizard species occurred after European contact due to habitat
destruction, competition for resources, and predation by rats, the brown tree snake
(Boiga irregularis), and other animals. There are currently only five (of twenty-four
documented) extant species of birds (herons - two species, swifts, starlings, and crows)

40
left on Guam. This extreme decline in birds is attributed to the introduction of the brown
tree snake around World War II (Pregill and Steadman, 2009).
Settlement History
In order to fully understand colonization in Guam and the Mariana islands, an
overview of peopling of the Pacific region will first be presented, followed by a more
detailed overview of settlement history of Guam.
Colonization of the Pacific Region
The peopling of the Pacific is an area that has received a great deal of attention
since the European discovery of the Pacific Islands. Over the past several decades,
there has been much dispute as to the actual origins of the Pacific Islanders, including
one hypothesis of settlement from South America (Heyerdahl, 1952), which has not
been substantiated. Currently researchers are much more united in their ideas on the
regions and dates of colonization and typically believe that the wide amount of
population variation seen in the Pacific region is due primarily to regional processes of
diversification (Bellwood, 1989).
Several independent lines of data have recently come together for a more unified
theory of the colonization of the Pacific. While there are still some debatable issues,
approaches from linguistic, archaeological, and biological perspectives have shed light
on the peopling of the Pacific. A temporal framework for the colonization of the Pacific
Islands is followed by a review of the archaeological, linguistic, and biological evidence.
Archaeology
The Pacific is characterized by two major colonization events (Thomas, 1999).
The first settlement of the Pacific occurred in Near Oceania around 40,000 to 30,000 BP
(Kirch, 1997). The second settlement event began around 3,500 BP with the rapid

41
spread of the Lapita Cultural Complex and the Austronesian languages throughout
Remote Oceania (Green, 1979; Kirch, 1997; Pawley, 1999; 2002). More specifically,
colonization dates, based on archaeological data for the geographical areas of the
Pacific are as follows: Mariana Islands circa 3,500 BP; Eastern Melanesia (Santa Cruz
region through Vanuatu and New Caledonia) circa 3,300 to 3,200 BP; eastern
Micronesia circa 2,000 BP; Polynesia circa 2,000 BP; and the last colonized areas are
New Zealand circa 1,000 BP, and Chatham Islands circa 500 BP (Sutton, 1980;
Davidson, 1984; Bonhomme and Craib, 1987; Kirch and Hunt, 1988; Green 1991b;
Craib, 1993; Butler 1994; Anderson, 1991; 1996).
Linguistics
The three major language groupings in the Pacific are Australian, Papuan, and
Austronesian (Bellwood, 1989). Populations from the region of interest for the current
study are all members of the Austronesian language family, with approximately 1,200
modern languages, thus this family will be discussed in more detail (Kirch, 2010).
Pawley and Green (1984) advocate a dialect chain model in contrast to many of the
hierarchical family tree models often discussed in reference to Pacific language
groupings (e.g. Terrell, 1986; Terrell et al., 1997). The Proto-Austronesian homeland is
believed to have originated in Taiwan approximately 6000-5000 BP before spreading
across the Pacific region (Bellwood, 1991; Bellwood, 1997; Pawley, 1999). As outlined
in Bellwood (2000: 7) the spread of the Austronesian language is as follows: a subgroup
of the Proto-Austronesian subgroup developed into the Proto-Malayo-Polynesian family
with colonization of the Philippines circa 4500 BP; rapid movement through island South
East Asia and western Micronesia permitted the spread of the Malayo-Polynesian
subgroups between 4000 and 3000 BP; finally, the Proto-Oceanic group developed in

42
the Bismark Archipelago, followed by the spread of the Lapita cultural complex which
occurred alongside the spread of the Oceanic languages throughout western Polynesia
between 33000 and 2800 BP. Most languages of island Melanesia, Micronesia, and
Polynesia (except for those of Western New Guinea, Palau, and Guam) stem from
Oceanic subgroup of the Austronesian family (Bellwood, 1989; Pawley, 1972). The
spread of Austronesian languages throughout the Pacific region has been shown to
integrate well with settlement patterns as demonstrated archaeologically (Green, 1999;
Spriggs, 1999; Bellwood, 2000)
Biology
Biologically, colonization studies can be subdivided into anthropometric, genetic,
and biodistance studies (Shapiro and Buck, 1936; Howells, 1970; Brace and Hinton,
1981; Serjeantson, 1985; 1989; Brace and Hunt, 1990; Brace et al., 1990; 1991;
Pietrusewsky, 1990a; 1990b; Houghton, 1991b; 1996; Hanihara, 1992; Turner, 1990a;
1990b; Scott and Turner, 1997; Hurles et al., 2002; Lum et al., 2002; Stephan and
Chapman, 2003). For the purpose of this study, evidence will be paid primarily to the
genetic, skeletal, and dental evidence of colonization of the Pacific.
Genetic: Genetic analyses grouped the human leukocyte antigen (HLA) into two
clusters: island Melanesian and Australian, and western Melanesian (Serjeantson,
1985; 1989). These studies demonstrated that proto-Polynesians likely traveled along
the northern coast of New Guinea before arrival into Polynesia. Y-chromosome studies
have suggested Island Southeast Asia as the ancestral group to both Near and Remote
Oceania (Hurles et al., 2002; Lum et al., 2002). Pietrusewsky (2006) outlines three
conclusions from the genetic studies regarding peopling of the Pacific. First, common
origins for Remote Oceania is likely from a region extending from Island Southeast Asia,

43
the Bismarck Archipelago, and the northern New Guinea coast. Second, admixture
between indigenous groups and Austronesian migrants likely occurred during eastward
expansion across the Pacific. Lastly, differential settlement or gene flow patterns are
postulated between males and females due to the diversity between mtDNA and Y-
chromosome evidence.
Cranial: The majority of multivariate cranial analyses have shown a close
association between Polynesians and Micronesians and have suggested a homeland
origin of Island Southeast Asia. Additionally, the Polynesians and Micronesians are
differentiated from the closely related Australians and Melanesians. Likewise, the
Melanesian and Australian groups display more heterogeneity and variation than what
is found within the more homogeneous Polynesian groups (Pietrusewsky, 1990a;
1990b; Hanihara, 1992; Stephan and Chapman, 2003). These findings are consistent
with two colonization events of the Pacific (Pietrusewsky, 2006). Brace and coworkers
(Brace et al., 1990; 1991) provided evidence of Japan as the homeland for Pacific
Island groups; however, no other studies have corroborated this finding.
Dental: Odontometric studies have shown that Australians and Melanesians have
some of the largest teeth in the world, followed by the smaller Micronesian and
Polynesian teeth (Brace and Hinton, 1981; Brace et al., 1990; 1991). Non-metric dental
variation displays two contrasting patterns between the Sinodont dentition of
Polynesians and Micronesians and the Sundadont pattern found in Southeast Asia.
While Australians and Melanesians do not fit either pattern, they are closest to the
Southeast Asia Sundadont pattern (Turner, 1990a; 1990b; Scott and Turner, 1997).
Likewise Hanihara (1992) found that Polynesians and western Micronesians had a

44
closer affinity with Southeast Asians than with Melanesians. These dental studies also
support a two-part settlement strategy for colonization of the Pacific.
Archaeological, linguistic, and biological studies have already shown that there
appears to be a two-prong settlement pattern in the Pacific. However colonization of
the Marianas is still unclear (Irwin 1992). The following section will outline the known
evidence of settlement patterns in this region.
Colonization of the Mariana Islands
Lowered sea levels following the mid-Holocene in the Pacific Ocean resulted in the
appearance of attractive coastal environments, which subsequently allowed for rapid
dispersal and initial human colonization in the western parts of Remote Oceania
(Dickinson, 2001; Carson, 2011). Most researchers agree that the islands were
colonized by horticulturalists, with sophisticated sea-faring technology, from Island
Southeast Asia (Bellwood, 1975; 1979; Hanson and Butler, 1997; Kirch, 2000).
Intentional and planned migration is supported by analysis of pottery, language, and
DNA (Hanson and Butler, 1997; Lum and Cann, 1998; 2000; Callaghan and Fitzpatrick,
2008). Osborne (1958) posited a ‘stepping-stone’ colonization model of Micronesia
through Palau and Yap, however, this theory has not been supported by the linguistic
and archaeological data (Anderson ; 2005; Clark, 2005). Most recently, Hung and
colleagues (2010) suggest the Northern Philippines as the most likely point of origin for
the earliest colonizers based on shared cultural similarities. Archaeological and
paleoenvironmental studies date initial settlement to approximately 3,500 BP (Spoehr,
1957; Craib, 1993; Butler, 1994; Carson, 2008, 2010; Clark et al., 2010); however one
study, based on sedimentary cores, suggests an earlier date of 4,300 BP (Athens and
Ward, 1999; Athens and Ward, 2004). If these theories are correct, Guam was

45
colonized after an open sea voyage of 2,600 km, which is three times the previously
reported longest ocean-crossing in prehistory (900 km, between Vanuatu and Fiji) by
the Lapita people (Green, 1979; Keegan and Diamond, 1987). Additionally, this voyage
would have occurred 500 years prior to the Lapita expansion (Craib, 1999; Spriggs,
1999), making Guam the earliest colonized island in Remote Oceania. A four-prong
approach, focusing on linguistics, archaeology, genetics, and skeletal data, is utilized to
review colonization patterns of the Marianas.
Linguistics
A substantial amount of debate has occurred over the linguistic position of the
indigenous Chamorro language (Blust, 2000; Reid 2002). Three viewpoints summarize
the theories regarding the Chamorro language. The first suggests that Chamorro is
closest to Philippine languages (Safford, 1909; Topping et al., 1975); the second theory
supports that Chamorro is most closely related to Indonesian languages (Zobel, 2002),
and the last viewpoint, and currently accepted interpretation, suggests that Chamorro is
not closely related to any subgroupings of the larger Austronesian language family
(Dyen, 1965; Blust, 2000; 2009; Reid, 2002).
The current assertion about the position of the Chamorro language places it in the
Western Malayo-Polynesian (WMP) group, which is spoken in the Marianas, Palau,
Philipines, Malaysia, parts of Indonesia, coastal southern Vietnam, and Madagascar.
The WMP group, along with the extra-Formosan Austronesian languages, is part of the
reconstructed Proto-Malayo-Polynesian language, which began to differentiate in
northern Southeast Asia (Ross et al., 1998; Blust, 2000; 2009; Pawley, 2002). The
Chamorro language has a distinctly separate geographic source from the Lapita Proto-
Oceanic group, and historical linguistics suggest direct colonization from Island

46
Southeast Asia (Kirch, 2010), and more specifically, from the central or northern
Philippines (Blust, 2000; Reid, 2000).
Archaeology
The earliest occupation sites in the Marianas are found along the shorelines
associated with dynamic and biologically diverse marine ecosystems (Hanson and
Butler, 1997). Radiocarbon analysis on materials recovered from sites in the Marianas,
including Achugao on Saipan (Butler 1994; 1995), Unai Chulu on Tinian (Craib, 1993;
Craib 1998; Haun et al., 1999), and Unai Bapot on Saipan (Bonhomme and Craib,
1987; Carson, 2008; Carson and Welch, 2005; Clark et al., 2010), provide the earliest
dates of settlement for the Mariana Islands and correspond to the Early Unai Phase.
The Achugao site on Saipan provides the earliest radiocarbon dates for the
Marianas from charcoal samples, 3470 ± 120 BP and 3120 ± 50 BP, and is associated
with a large assemblage of Marianas Red slipped pottery, some with curvilinear or
straight lines and lime filled decorations, termed Achugao Incised, that serves as the
archetype for ceramics associated with early period settlement sites (Butler, 1994;
1995).
Thirteen radiocarbon dates from charcoal samples at the Unai Chulu site on Tinian
place occupation between 3400 and 2900 BP. Like the Achugao site, the Unai Chulu
site is also associated with Marianas Red incised sherds, similar to the Achugao Incised
type (Craib, 1993; Craib, 1998; Haun et al., 1999).
The Unai Bapot site on Saipan is associated with a diverse assemblage of early
decorative redware and blackware, with slipped surfaces, and has been established as
one of the earliest sites in the Marianas, with 31 radiocarbon dates, from charcoal, shell,
and wood, placing settlement between 3400 to 3200 cal. BP (Bonhomme and

47
Craib,1987; Carson, 2008; Clark et al., 2010). Carson’s radiocarbon findings (2008)
pushes occupation even further back to 3600 to 3420 BP, which pre-dates Lapita
expansion into Remote Oceania.
Most of the earliest sites in Guam have questionable dates. Matapang Beach
Park, in Tumon Bay, has been dated as early as 5260 ± 100 BP, however, this sample
is from a scattered charcoal deposit with no clear provenience. Dates with more secure
stratigraphic contexts from the same site, were obtained from fire pit features and range
from 3880 ± 90 BP to 3170 ± 70 BP (Bath, 1986). The Huchunao site is associated with
Achugao Incised pottery has been dated more securely to 3690 to 2830 BP (Dilli et al.,
1998). Most recently, the Ritidian site in northern Guam, dates from the beginning and
end of the Early Unai Period. The early settlement dates to 3547 to 3323 BP and is
associated with very thin, redware pottery. The slightly younger site, with thicker and
coarser pottery, dates to 3056 to 2842 BP (Carson, 2010). This site illustrates
chronological change in pottery sequence over time with solid radiocarbon dates.
Decorated pottery in the Mariana Islands is most similar to ‘red-slipped, circle- and
punctate-stamped pottery from several sites in the Cagayan Valley on Luzon’ in the
Philippines (Hung et al., 2010: 913). The Neolithic and Iron age site of Nagsabaran,
which is located within the Cagayan Valley, dates to 4000 and 3300 BP; thus
encompassing the earliest settlement dates from the Mariana Islands. While decoration
of pottery sherds is rare in Nagsabaran, the motifs are similar to those found in the Early
Unai Phase; additionally the incised designs are often filled with lime (Hung et al.,
2010). Thus, the archaeological evidence supports the linguistic findings of settlement
from the Philippines.

48
Genetics
Lum and Cann (1998) analyzed five mtDNA region V length polymorphisms from
over 800 individuals. This polymorphism is frequent in Island Southeast Asia,
Micronesian, and Polynesia, suggesting an Island Southeast Asia origin for the latter
two groups. Further, the data were compared with linguistic and geographic
interactions spheres and revealed that there was extensive amount of gene flow in
much of Micronesia, except for in the Marianas and in Palau (the two non-Oceanic
speaking populations), suggesting that these regions may have been isolated from
other Micronesian groups.
A subsequent study (Lum and Cann, 2000) sought to elucidate the relationship
between Micronesians and Polynesians, who share a number of cultural (e.g. kava
drinking – only in Central and not Western Micronesia) and biological (craniofacial
measurements) traits (Pietrusewsky, 1990 a,b; Lum and Cann, 2000). The study found
that 89% of Micronesians and Polynesians had shared mtDNA control region
sequences. Additionally, Micronesians and Polynesians have been clustered into the
same lineage group (Lineage group I.1), along with Indonesians. A nodal sequence
(L22) of another lineage group (Lineage group I.2), is also shared by Micronesians and
Polynesians, as well as populations from the Philippines and Borneo. However,
regional differences in shared lineages have been found between Western and Central-
Eastern Micronesia. A chart plotting multidimensional chord distances between
populations groups the central Micronesian islands (Nauru, Pohnpei, Kiribati, Kosrae,
and Marshalls) more closely with Polynesian islands. The western Micronesian islands
(Marianas, Palau) are further removed, with the exception of Yap. Looking specifically
at the at the Marianas sequence it is obvious that it remains isolated and lies in between

49
Near Oceania and Remote Oceania groups, in concordance with its geographical
position. Further, sequences of the Western Micronesia groups suggest that they were
each settled independently, but directly from Island Southeast Asia. Post-settlement
gene flow between the Marianas and central-eastern Micronesia is posited due to the
low frequency and diversity of its group I sequence. These data are further evidence
against the ‘stepping-stone’ hypothesis of settlement of the Marianas through Palau and
Yap.
Bioarchaeology
Howells (1970; 1973; 1977; 1979) was influential in collecting and publishing
anthropometric and cranial data on large numbers of living Pacific populations.
Multivariate analysis of anthropometric data grouped Micronesians closely with
Melanesians suggesting gene flow and intermarriage between those populations
(1970). Craniometric analyses, however, closely link Polynesians and Micronesians as
distinct from Australians and Melanesians who share similar features in skull form
(1977).
Pietrusewsky’s research (1990a; 1990b; 1994; 1995; 1996; 2000; 2005; 2008)
over the past four decades confirms Howell’s findings in regards to the position of
Micronesia populations in comparison to the rest of Remote Oceania, Near Oceania,
Australia, Island Southeast Asia, mainland Southeast Asia, east Asia, and North Asia.
Multivariate craniometric analyses (27 measurements in 63 cranial series) on 2,805
male crania suggest that the crania from Guam (n = 46) are most closely related to the
Polynesian series (Tonga-Samoa, Hawaii, Rapa Nui, Gambier Islands, Marquesas
Islands, Society Islands, Tuamotu Archipelago, and Chatham Islands) (Pietrusewsky,
2005). A dendogram of Mahalanobis D2 shows that the Polynesian and Guam grouping

50
connects with a larger cluster formed by East Asian, North Asian, and Southeast Asian
crania which are closest to cranial series from Island Southeast Asia. Thus, the
Polynesian and Guam populations have likely originated from Island Southeast Asia.
Ishida and Dodo (1997) analyzed 22 non-metric cranial traits from the Marianas
populations and compared them with twelve other groups across the Pacific, mainland
Southeast Asia, East Asia, mainland Asia, and North America. Like Pietruwsewsky’s
and Howells’ results, Ishida and Dodo cluster the Chamorro and Polynesian populations
with Southeast Asian and East Asian populations.
One problem with the interpretation of Pietrusewsky’s and Ishida and Dodo’s
findings on Chamorro settlement patterns is that their samples are derived from the
Hornbostel-Thompson Collection1 of skeletal material from the Mariana Islands
originally housed in the Bishop Museum in Hawai’i. These series of skeletons were
collected by Hans Hornbostel and J.C. Thompson in the 1920s from various locations
across Guam, Saipan, and Tinian (Ishida and Dodo, 1997). Most of these skeletons
date to the pre-contact Latte period and were found in association with Latte sets;
however, the remains are poorly provenienced and the result of selective recovery for
the most well preserved elements (Hanson and Butler, 1977). Thus, these skeletal
remains are not representative of a typical mortuary population. While their research
represents the largest undertakings in understanding the Chamorro settlement patterns
and relationships to other island groups, a similar study on a Pre-Latte sample would
likely provide complementary information regarding early Chamorro populations.
1 The Hornbostle collection has since been repatriated to Guam and is currently being curated in the
Guam Museum’s collections.

51
Unfortunately, as discussed above, Pre-Latte samples are typically very small and
extremely fragmentary, thus precluding this type of analysis.
Settlement Summary
The linguistic, archaeological, genetic, and skeletal findings are consistent in their
findings of settlement of the Marianas from Island Southeast Asia. Most recently,
archaeology and genetic advances have been able to pinpoint a more precise location
for area of origin – the Philippines. As Rainbird (2003: 85) notes, a voyage from the
Philippines to the Marianas ‘would constitute the longest sea-crossing undertaken by
that time in human history’. Thus, the peopling of the Marianas is not only interesting in
terms of population history and initial expansion into Remote Oceania, but also
represents development of new technological advances in precise navigation and sea-
faring skills.
Marianas Chronological Sequence
Spoehr (1954, 1957) was the first to place colonization of the Mariana Island chain
at 3500 BP, based on radiocarbon dates of 3,479 ± 200 years BP from oyster shells at
the Chalan Piao site on the island of Saipan, located approximately 220 km north of
Guam (Figure 2-1). Subsequently, recalibration of Spoehr’s dates yielded a more
recent age dating to 1,700 years later (Cloud et al., 1956). Nonetheless, the majority of
the earliest radiocarbon dates across the Marianas cluster around 3500 to 3000 BP,
therefore substantiating Spoehr’s claim and making Guam the earliest colonized island
of Micronesia and Remote Oceania (Reinman 1977; Kurashina and Clayshulte 1983;
Butler 1995; Carson 2008; Carson 2010; Clark et al. 2010; Hung et al. 2011). Athens
and Ward (2004) push this chronology as far back as 4,300 BP, based on the

52
appearance of charcoal particles in the Laguas paleocore, however, this date has not
been supported by archaeological findings.
The history of the Marianas can be divided into the pre-contact and post-contact
eras, each of which may be further subdivided based on archaeological and historical
periods. While various chronological sequences have been suggested, based primarily
on different temper types found in ceramics, the standard terminology is adapted from
that of by Spoehr (1957) and Moore (1983). Table 2-1 provides an overview of the
standardized chronological sequences used in the Mariana Islands.
Spoehr (1957) proposed the chronology of the Marianas dividing the prehistory
into two periods: the Pre-Latte (3500 BP – 1000 CE) and Latte Periods (1000 – 1521
CE). Named after megalithic architecture, the latte are composed of two parts: coral
limestone pillars, the haligi, which are topped by a hemispherical stone cap, the tasa
(Figure 2-3). This monumental architecture is only associated with the Latte Period.
Grouped latte stones are known as latte sets and were arranged in parallel rows
designed to support wooden structures and are thought to be the foundation for
prehistoric houses and meeting halls (Thompson, 1932; Thompson, 1940; Morgan
1988). Spoehr’s division also distinguished between ceramic types with the Pre-Latte
Marianas Redware and the Latte Period Marianas Plainware.
Moore’s (1983) chronological sequence builds upon Sphoer’s original scheme and
is based on analyses of ceramic sequences from Tarague Beach in Northwestern
Guam. Her classification was later refined into the current standard which has replaced
the term ‘Pre-Latte’ with ‘Unai’, which means beach in Chamorro, and ‘Transitional’ with
‘Huyong’, which translates to ‘going out’ (Moore, 2002) (Table 2-1). The following

53
overview of the chronological sequence is based on a combination of Spoehr (1957)
and Moore’s (2002) divisions.
Pre-Latte Period
The peopling of Micronesia was accomplished by people with sophisticated ocean-
faring technology who already had horticultural capabilities (Bellwood 1979) (see more
detailed discussion below). The Pre-Latte period (3500 BP – 1000 CE) in the Marianas
begins with initial human colonization around 3,500 years ago and concludes with the
onset of latte construction in the first millennium CE. Very few Pre-Latte sites have
been found in the Marianas as older sites are more prone to erosional and depositional
disturbances due to frequent storms. Additionally, wave activity removes the original
soil horizon and deposits materials further inland, causing major alterations to
archaeological deposits. Thus, many Pre-Latte sites have not been preserved intact
and have often been reworked, mixed, moved, or eradicated by storms (Kurashina and
Clayshulte, 1983; Hunter-Anderson and Butler, 1991).
The Pre-Latte period is characterized by small populations with low population
densities and semi-permanent habitation sites situated near the coastal margins
(Hunter-Anderson and Butler, 1991). However, Moore and colleagues (1988) suggest
that the Pre-Latte were not sedentary and moved seasonally in accordance with
resource procurement strategies.
There has been little research on terrestrial animal exploitation of the Pre-Latte
period in Guam and most information comes from the northern Marianas Islands. At
Unai Chulu on the island of Tinian, located approximately 160 km north of Guam, bones
from the fruit bat and rails were found (Haun et al., 1999). However, additional analysis
to interpret food processing, cooking, or breakage patterns were not conducted

54
(Amesbury and Hunter-Anderson, 2003). On the islands of Tinian and Aguiguan,
Steadman (1999) found burned animal bone indicative of human cookery dating to the
Pre-Latte period. In the Railhunter Rockshelter on Tinian, dating to approximately 2400
to 2200 BP, Steadman (1999) found fish, lizard, snake, and several bird species
restricted to a layer that is associated with human activity. Likewise, the Pisonia
Rockshelter on Aguiguan, also contained burned remains of fish, lizard, Rallidae sp.
(rails), and Gallicolumba xanthonura (white throated ground dove) (Steadman, 1999).
Diet
The majority of the Pre-Latte diet came from bivalves, shellfish, and reef fishing
(Amesbury et al., 1991). Archaeological studies on Saipan have shown that the Pre-
Latte period has a much higher percentage of fishing gear and related fishing
production debris per temporal unit in comparison to the Latte Period (44% and 14%,
respectively, of total shell artifacts per unit) (Butler 1995). Studies have shown that
bivalves, specifically Andara antiquate (Blood clam), Gafrarium tumidum (Tumid venus
clam), and Gafrarium pectinatum (Comb venus clam), were found in abundance in Pre-
Latte deposits in Ypao Beach on Guam, as well as on Chalan Piao in Saipan
(Leidemann, 1980; Amesbury et al., 1996). Likewise, Graves and Moore (1985) found
that bivalves, specifically Arcidae (Ark clam) comprised between 61% and 71% of the
bivalve assemblage in the Pre-Latte units from Tumon Bay.
In the Tarague cultural sequence, Kurashina and Clayshulte (1993) found
Tridacna cf. gigas (Giant clam) in the oldest stratigraphic layer, with radiocarbon dates
of 3435 ± 70 BP, placing it in the Pre-Latte period. This finding is rare, as T. gigas is not
known to have existed in the Marianas during the Holocene. Thus, presence of T. gigas

55
may represent goods brought by the early island colonizers or may be the result of trade
or communication networks with other Pacific populations.
Zooarchaeological assemblages have also shown that a significant proportion of
consumed foods came from fish in the Pre-Latte period. Archaeological excavations
conducted in Pagat (Craib, 1986) on the east coast of Guam yielded higher
concentrations of pelagic fish bone in the Pre-Latte Layer (density of 378.37 grams per
cubic meter) in comparison to the Latte layer (187.20 g/m3).
Leach and Davidson (2006a) report on approximately 20 pelagic fish species
excavated from the Mangilao Golf Course site, on the east coast of Guam, and
determined that there are differences in their frequency over time. While the number of
species exploited between Pre-Latte and Latte periods increase, the percent of some
species represented in the archaeological samples decrease throughout prehistory. For
example, the most common fish in the archaeological assemblage, parrotfish
(Scaridae), decreases in frequency from 60%, in the Early Pre-Latte, to 27% in the
Intermediate/Transitional Pre-Latte, and then increases to 43% in the Latte period.
Emperorfish (Lethrinidae) and wrasses/tuskfish (Coridae/Labridae) decrease in
frequency over time, while other pelagics, mahimahi (Coryphaenidae) and
swordfish/marlin (Istiophoridae/Xiphiidae), increase in the Latte period.
McGovern and Wilson (1996) similarly found elevated 15N values in a small Pre-
Latte sample from Saipan, suggesting increased exploitation of pelagic species, rather
than dependence upon coastal and marine habitats. Evidence from Ylig Bay, on the
east coast of Guam, showed no significant differences from frequencies of fish remains

56
between the Transitional Pre-Latte and Latte/Historic phases (Leach and Davidson,
2006b).
The impact of coastal deposition due to eroding hillsides suggests that the
landscape was extensively altered and likely utilized for horticultural practices around
2,400 BP (Athens and Ward, 2004). This evidence of landscape modification suggests
that horticultural practices and foreign cultigens were likely brought with the early
colonizing populations (Hunter-Anderson, Butler, 2001).
The appearance of Lycopodium and Gleichenia ferns early on in
paleoenvironmental sediment cores suggests small-scale gardening and resource
collecting by initial settlers (Athens and Ward, 2004). Breadfruit (Artocarpus) and taro
pollen (Colocasia esculenta) are also noted early in the coring record, and around 1100
BP. Further evidence of taro comes from identified taro starch grains from Pre-Latte
pottery sherds, where Cordyline (Cabbage tree), fish and, shellfish, were also found,
suggesting their importance and use among the foods cooked in the Pre-Latte (Loy,
2001a,b; 2002; Loy and Crowther, 2002).
The Pre-Latte phase is further subdivided into four phases based on differences in
pottery sequences from Tarague Beach on the west coast of Guam: the Early, Middle,
and Late Unai, and the Transitional/Huyong (Moore, 2002). These phases will be
discussed in terms of pottery characteristics and cultural materials associated with each
phase. Known sites dated to these periods will also be addressed.
The Early Unai phase (3500-3000 BP)
The first 500 years following initial human settlement constitute the Early Unai
Phase, which is characterized by non-decorated Marianas Redware (Moore, 2002).
These vessels have thin walls, restricted openings, everted rims, and slipped exteriors

57
(Moore, 2002). Other pottery types, such as Achugao, with curvilinear or straight lines
and dentate infilling, and San Roque, characterized by curvilinear lines with stamped
circles, have also been found in small frequencies in Saipan (Butler, 1995; Moore,
2002) (Figure 2-4). Coastal locations with access to marine resources were likely
habitation areas for the earliest settlers (Hunter-Anderson and Butler, 1991). Cultural
materials associated with this phase include a high proportion of bivalve remains, stone
and shell tools, fishing equipment, and shell ornaments (Bath, 1986; Graves and Moore,
1995). Few sites of this earliest phase are known for Guam, but exceptions include
Huchunao in Mangilao; Ypao and Matapang in Tumon Bay; and Ritidian in the north
(Leidemann, 1980; Bath, 1986; Dilli et al., 1998; Carson 2010).
The Middle Unai period (3000-2500 BP)
The Middle Unai Phase spans the next 500 years following the Early Unai Phase.
Archaeological assemblages are very similar to the Early Unai Phase, with the
exception of pottery. Marianas Redware usage is continued in this phase with everted
or flared unthickened rims. Many of the vessels have calcareous or volcanic sand
tempering. As seen in the Early Unai, many of the surfaces are plain or slipped,
however, new surface treatments also emerged, characterized by polishing or
burnishing, in the Middle Unai (Moore, 2002).
By the Middle Unai Period, the Achugao and San Roque designs were abandoned
and replaced by Ipao Stamped, after Craib (1990), with bold-lime-filled designs with
combinations of straight lines, circles, half circles and chevrons (Moore 2002: 8). Over
30 different types of band designs have been reported from across the Marianas
islands.

58
Ray (1981) reports another pottery type characteristic to the Middle Unai, know as
Tarague Striated, which has scrape marks and impressions parallel to the rim seen on
both the inside and outside of the vessel. Other observed surface treatments during
this time period include fingernail impressions, dots, and ridges (Ray, 1981; De Roo and
Goodfellow, 1998; Moore, 2002).
Known sites dating to the Middle Unai increase in abundance in comparison to the
Early period. These sites are primarily located along the coast and in caves and
rockshelters along the shoreline. Some sites associated with the Middle Unai featuring
Ipao Stamped pottery are located in Tumon Bay: Ypao (Leidemann, 1980; Olmo and
Goodman, 1994), Kallingal Property (Moore et al., 2001), and at the site of the current
study, Naton Beach (Defant, 2008).
The Late Unai period (2500-2400 BP)
The Late Unai phase begins around 2,500 years ago and lasts 100 years. This
period is characterized by a decrease in complexity in design of vessel forms (Moore
and Hunter-Anderson, 1999). Ipao Stamped designs are found on a small percentage
of bowls, however, the impressions are limited to the rim of the vessel (DeRoo and
Goodfellow, 1998; Ray, 1981; Moore, 2002).
Thick flat-bottomed pans become common with wide openings and shorter heights
(Moore, 1983). Matt-impressions are also noted, primarily on the base exterior,
however they are sometimes observed on the interior as well (DeRoo and Goodfellow,
1998). The matt impressions are similar to a weaving style found on contemporary
sleeping mats from the Caroline Islands made from the pandanus tree (Pandanus
tectorius) (Safford, 1905; Hunter-Anderson et al., 1998). This finding suggests that the
pan was placed on the mat while drying to prevent collapse prior to firing (Moore, 2002).

59
Quartz inclusions originating from Saipan have been found on a small number of
sherds from this time period suggesting that ceramics were brought to Guam from
Saipan (Dixon et al., 1999; Moore, 2000; Hunter-Anderson et al., 1998). These foreign
sherds are found primarily on the west coast of Guam, including in Tumon Bay,
indicating that inter-island contact occurred but was restricted to the western portion of
the island. This theory is plausible since the high cliff lines along the east coast would
make access to the shoreline much more difficult by canoe (Moore, 2002).
The Transitional (Huyong) period (400-1000 CE)
In the period following AD 400, there is a decline in the use of flat-bottomed pans
and a change in vessel form with thin walls, round bases, and slightly incurvate rims.
Various tempers, including calcareous sand, mixed sands, and volcanic sands, are
common with plain or polished/burnished surface treatments. Decorated rims are
accompanied by wall perforations, likely for practical and not decorative purposes, are
also seen in this time period (Moore, 2002).
Change between vessel form, surface treatment, and tempers were gradual and
did not occur concurrently across sites. There are numerous sites dating to this time
period, which are found along rivers and the coast and include both open-air sites and
rockshelters (Tomonari-Tuggle and Tuggle, 2003). Most notable is Tarague Beach, just
north of Tumon Bay. Coastal and river sites of this period have an abundance of
cultural materials in comparison to the more inland sites (Moore, 2002).
Latte Period (1000-1668 CE)
An increase of charcoal within paleoenvironmental sediment cores around 1,800
BP is suggestive of an intensification of land use and corresponds to the shift from the
Pre-Latte to Latte periods (Athens and Ward, 2004). Unlike Pre-Latte sites, the Latte

60
deposits did not experience extensive disturbances from the long-term effects of storm
events, wave action, and sea-level changes. Thus, most of the current knowledge of
Guam’s prehistory comes from the better preserved Latte period sites (Hunter-Anderson
and Butler, 1991). While ideal settlement areas remained in sandy coastal locations,
populations also began to move inland to more marginal environments, such as the
interior uplands (Figure 2-5).
Latte architecture
The Latte Period begins around AD 1000 and is characterized by the construction
of latte sets and Marianas Plainware pottery (Spoehr, 1957; Graves, 1986) (Figure 2-6).
The construction of the latte sets began at approximately AD 1000 and by AD 1325 had
spread to the northern Mariana Islands, including the northernmost populated island,
Pagan (Graves, 1986). Latte use and construction continued throughout the Latte
period and into the first part of the Spanish Colonial period (Tomonari-Tuggle et al.,
2005). The exact end date for latte construction is unknown, however, it is believed that
the disbanding of indigenous settlements resulted in cessation of latte production
(Moore, 2002).
Several early hypotheses speculate on the function and significance of latte sets
as specialized structures attributed as ceremonial centers, meeting houses, men’s
houses, canoe sheds, and residential structures for the elite (Thompson, 1940;
Thompson, 1945; Spoer, 1957; Reinman, 1977). A survey of cultural material in and
surrounding latte sets in Guam, Saipan, Tinian, and Rota revealed varied artifact
categories and assemblages that indicating that a wide range of domestic activities
were associated with the latte structures and they were not, as previously thought,
specialized to one function. Artifacts associated with cooking, food preparation, tool

61
manufacture and maintenance, fishing, and warfare were found homogenously in all the
latte sets analyzed (Table 2-2). Additionally, human burials are uniformly found within
or around Latte structures (Graves 1986).
Most latte sets are arranged parallel to the coastline, however, some are
perpendicular to the coast (Thompson, 1940). Multiple latte sets are distributed in a
linear pattern, if located along a steep cliff, or clustered along long stretches of sandy
beach or limestone terraces. In areas where several latte sets are clustered together,
the largest latte set is centralized with the smaller sets along the periphery (Graves,
1986). Archaeological evidence points to a stratified society with chiefs who organized
the labor for construction of the latte sets. It has been hypothesized that latte were
symbols of corporate landholdings and a kin group’s claim to land through linage
(Hunter-Anderson, 1989). Further, the practice of burying family members within the
latte set further legitimizes the claim through a direct link to the ancestors (Hanson and
Gordon, 1980). At the time of contact, settlements consisted of groups of latte houses
nucleated into villages (Tomonari-Tuggle et al., 2005).
Cooking and food processing
Marianas Plainware is thick-walled with no slip and little decoration. Pots were
typically hemispherical or globular with restricted mouths, thick, incurving rims and
rounded bottoms (Moore, 2002). Some vessels displayed grooves perpendicular to the
rim, suggestive of being secured with ropes, possibly for suspension, handle for
gripping/pouring, or to secure a lid (Reinman, 1977; Davis et al., 1992; Wickler, 1993;
Moore and Hunter-Anderson, 2001; Moore, 2002). Many of these potsherds, primarily
those with plain and wiped or brushed surfaces, from the Latte Period have charred

62
residue on their interior surfaces that is likely associated with cooking (Loy, 2002;
Moore, 2002).
The wide array of food processing artifacts in the Latte Period are indicative of a
more sedentary population with an increased reliance on agriculture (Hunter-Anderson
and Butler, 1991; Moore, 2005). Stone mortars (Chamorro: lusong) and pounders are
found ubiquitously throughout Guam and date almost exclusively to the Latte Period.
They were likely used to ‘grind, pulverize, and de-husk plant products, such as rice,
cycads, and arrowroot tubers’ (Moore, 2005:100). The presence of scrapers, knives
and blades, usually made from local shells, are associated with digging into the soil and
for scraping, peeling, and cutting tubers, roots, and plant stalks (Moore, 2005).
Additionally, archaeological investigations in Saipan showed a 35% increase of adze
production from the Pre-Latte to Latte periods (Butler, 1988).
Diet
Starch grain analysis: Analysis of starch grain residues on 35 Latte-period
potsherds identifies taro, Cordyline (type of palm – common name Cabbage tree), rice,
sugarcane, and possibly shellfish and fowl, indicative of cooking these foods. Taro was
identified on more than half (n = 18) of these potsherds (Loy, 2001a,b; Loy and
Crowther, 2002; Crowther et al. 2003), suggesting preference for and possibly
cultivation of the introduced tuber. Radiocarbon dates associated with the
archaeological sites from which the potsherds originated, indicate that taro was cooked
in clay pots as early as 2000 years BP (Hunter-Anderson et al., 2001; Moore, 2005).
Likewise, analysis of pollen in sediment cores indicates that taro was present just prior
to and during the early Latte Period (Cummings and Puseman, 1998; Athens and Ward,
1999).

63
Pollen and phytolyth analysis: Pollen data from paleoenvironmental sediment
cores have identified a wide range of cultivable plant foods including betel palm,
breadfruit, coconut, pandanus, Cordyline, Ipomoea (morning glory), and aroids
(Alocasia, Colocasia [taro], and Cyrtosperma) (Athens and Ward, 1995; Athens and
Ward, 1999; Cummings and Puseman, 1998; Dixon et al., 1999; Ward, 2000;
Cummings, 2002; Athens and Ward, 2004). Likewise, phytolith analyses from soil
samples and sediment cores have also found evidence of breadfruit and betel palm, in
addition to arrowroot, rice, bananas, canna, and Job’s tears (Hunter-Anderson, 1994;
Pearsall and Collins 2000; Collins and Pearsall, 2001a; Collins and Pearsall 2001b).
Lastly, analysis of charred wood samples from southwestern Guam identified coconut,
breadfruit, and pandanus (Murakami, 2000).
Stable isotope analysis: Studies focusing on stable isotopes for prehistoric diet
reconstruction in the Marianas are limited in number and generally have very small
sample sizes. However, research has been conducted in Guam, Rota, and Saipan to
estimate the proportion of marine versus terrestrial foods in the prehistoric diet of
populations inhabiting the Mariana Islands (Hanson, 1989; Quinn, 1990; McGovern-
Wilson and Quinn, 1996; Ambrose et al., 1997; Pate et al., 2001).
In an analysis of four individuals from the Duty Free site on Saipan, Hanson (1989)
found consistently low 13C values indicating a significant dependence on terrestrial
foods. The 15N values are more variable and suggestive of a combination of different
marine resources in the diet with a higher emphasis on lagoon resources. Nonetheless,
the isotope signatures are reflective of a diet comprised much more of terrestrial foods
than marine foods, which is surprising given the proximity to the ocean.

64
Quinn’s analysis (1990) of two individuals from different locations in Saipan
revealed one individual with a high proportion of open water foods and no reef foods in
his diet. Likewise, another male with no access to reef or lagoon areas had a high
proportion of the diet comprised of open water marine foods. McGovern-Wilson and
Quinn (1996) found that marine resources contributed 30% (based on mean 15N value)
and 40% (based on mean 13C value) of the diet in Saipan, while these values are still
low, they are higher than expected given the faunal record. Exploitation of pelagic fish
is also revealed by the isotope analysis. Additionally, McGovern-Wilson and Quinn
found no age or sex differences in access to differential resources.
Ambrose and colleague’s (1997) data for Saipan reveal large discrepancies
between diet reconstruction from collagen versus carbonate carbon isotopes. Bone
collagen suggests a 17% marine component to the diet, while the carbonate data
indicates a much higher estimate of 60%. Nonetheless, the very high apatite carbonate
13C values suggest a large component of the Saipan diet came from C4 or marine
plants with very low protein component, such as sugar cane and seaweeds. Small
amounts of marine foods, likely reef and lagoon fish and/or shellfish, contributed to the
diet (Ambrose et al., 1997).
Data from Rota are similar, yet more diverse than what was noted in Saipan. Pate
and colleagues (2001) found much variability between individuals and determined that
22% (ranging from 10% to 41%) of the diet in Rota was derived from marine foods.
Some individuals have elevated 15N values suggesting exploitation of pelagic fish,
while others have negative 13C and 15N values indicative of a diet dominated by
terrestrial foods, suggesting differential access to resources between individuals.

65
Likewise, Ambrose and colleagues (1997) found that marine resources contributed
between 21% and 40% to the diet in Rota. High 15N values suggest a significant
contribution of pelagic fish to the diet in some individuals, while other individuals have
low 13C values suggestive of a predominately terrestrial diet.
Stable isotopes in Guam reveal a similar signature as those reported for Rota,
however, the isotopic analysis is limited to only five individuals. Marine foods contribute
between 27% to 35% to the diet. Collagen 15N and 13C values are similar to what is
seen in Rota, however, the 15N values in Guam are slightly lower than those from Rota.
This suggests that the diet was comprised primarily of terrestrial C3 resources, such as
rice, root crops, and vegetables, and some marine protein, but little reliance on
seaweeds or C4 plants (Ambrose et al., 1997).
Shellfish consumption: A transition from bivalve to gastropod consumption
occurs in the Latte period (Amesbury et al., 1996; Amesbury, 2007). Leidemann’s study
(1980) at Ypao Beach, in Tumon Bay, found that Strombus gibberulus gibbosus
(gibbose conch) far outnumbered any other family. Graves and Moore (1985) also
noted the overabundance of gastropods in comparison with bivalves in Tumon Bay, with
Strombidae (conchs) comprising 90% of the gastropod assemblage. This shift is
attributed to changes in sea level and concomitant changes in local ecosystems where
preferable mangrove environments dissipate once in sea levels decrease (Amesbury,
1999; Graves and Moore, 1985).
Post-Contact Era
The arrival of Ferdinand Magellan and his crew in 1521 on the southern coast of
Guam marks the beginning of the post-contact era as well as the first contact between

66
Europeans and any indigenous Micronesian populations (Alkire, 1977; Lévesque,
1995). The post-contact area is commonly divided into the Spanish Colonial, First
American, Japanese World War II, American World War II, and Second American
periods.
The Spanish Colonial Period (1521-1898 CE)
At the time of Spanish contact, the indigenous peoples of the Mariana Islands were
described as a single population with shared language, culture, and customs (Driver,
1983). In 1565 Miguel Lopez de Legazpi landed in Guam and the Marianas from Spain.
This led to the start of the Manila galleon route where ships sailed from the Philippines
to Acapulco and upon the return voyage, stopped in Rota or Guam where they bartered
iron for fish, fruit, coconut, and rice, with the locals, before the final return back to the
Philippines (Schurz, 1939; Driver, 1983). Contact between Europeans and the
Chamorro remained limited until 1668 when the Spanish missionaries settled in Guam
to convert the Chamorro to Christianity. By this time, the missionaries characterized
Guam as a thriving culture, with 180 villages, the largest of which had 150 houses
(Lévesque, 1995).
Juan Pobre, a Franciscan brother who lived on Rota for seven months in 1602,
provides the most detailed description of the Marianas in the post-contact period. While
descriptions of agricultural practices were not reported, he noted that the Chamorro
used wooden sticks to dig into the soil. Taro, yams, rice and a type of sweet potato
were grown inland and these agricultural products were traded with villagers living along
the coast for fish (Driver, 1989).
Local population size for the island at the start of the 17th century is estimated at
20,000 but by the end of that century diminished to approximately 1,600 individuals due

67
to widespread epidemic disease and rebellion after European contact (Russell and
Fleming, 1990; Tomonari-Tuggle et al., 2005). The Spanish instituted the Reducción
which consolidated the entire Chamorro population of the Mariana Islands into seven
mission centers, six of which were located in Guam and one in Rota which were under
rigid control by the Spanish missionaries and military officers (Rogers, 1995; Tomonari-
Tuggle et al. 2005). Latte sets were eventually replaced by lanchos, small, elevated
thached houses, which served as subsistence farms (Carano and Sanchez, 1964;
Rogers, 1995).
Rice became much more frequently utilized and introduced maize gained dietary
importance over the previous staples of taro and breadfruit (Rogers, 1995). Other
introductions included mango, pineapple, papaya, citrus, hot peppers, and cassava
(Carano and Sanchez, 1964; Rogers, 1995). The Spanish also imported farm animals,
such as carabao (subspecies of the water buffalo indigenous to Southeast Asia), cattle,
horses, deer, pigs, and goats (Farrell, 1991; Hunter-Anderson et al., 2001). The
presence of alternative animal protein contributed to an overall decline in fishing in favor
of farming and raising livestock (Russell, 1988). The village of “Tumhan” is illustrated
on the 17th century map from the Jesuit Charles Le Gobien (1700), yet, according to
Kurishina et al. (1987; 1988), little activity is known to have occurred in the area during
this period due to the limited number of Spanish artifacts found during archaeological
testing (Kurashina 1987). However, Tumon Bay was used by the Spanish for fishing
and likely contributed to subsistence practices of both the Chamorro and Spaniards
(Burtchard 1991).

68
The First American Period (1898-1941 CE)
During the Spanish-American War, the United States took control over Guam in
1898, after Captain Henry Glass attacked the Spanish Forts in Apra Harbor and
demanded that the Spanish surrender Guam to the US. However, American rule was
not firmly set until the 1900s when a centralized, yet unfortified, military presence was
established at Orote Peninsula (Rogers, 1995). In 1922, all coastal defenses were
removed from Guam with the signing of the Naval Limitations Treaty, leaving Guam
unprotected and vulnerable.
According to Safford (1903), the local residents resided in six towns and were
primarily farmers who traveled to the lanchos for subsistence farming, while fishing
practices were greatly reduced. Population estimates for the six villages were: Agafia
with 6,400 inhabitants; Sumai, 9oo; Ynarajan, 550; Agate, 400; Merizo, 300; and Umata,
200. No population records for the Tumon Bay area are available for this time period,
however, hamlets along the coast composed of few houses are described.
The Japanese World War II Period (1941-1944 CE)
On the morning of December 8, 1941 (UTC/GMT +10 hours), the same day of the
December 7th attack on Pearl Harbor, Hawai’i, the Japanese attacked Guam. Two days
of bombing by Japanese air forces culminated with a three-pronged ground attack at
Tumon Bay, Agaña Bay, and Merizo, and the forced surrender of Guam to the
Japanese by the American governor (Rogers, 1995). The majority of the local
population fled to the countryside in an effort to escape the Japanese regime who
attempted to enculturate the Chamorro population with forced language change in the
school system. During this period, the Chamorro reverted back to traditional practices

69
of fishing and agriculture, particularly in more remote areas (Russel et al., 1993;
Rogers, 1995).
With the intensification of the war and Japan’s decreasing control in the Pacific
Theater, the Chamorro were placed into forced labor camps (Sanchez, 1979). The
defense strategy included fortifications along the entire western coast of Guam,
particularly Agat and Asan, but in Tumon as well (Craib and Yoklavich, 1996). Pillbox
fortifications and gun emplacements were scattered along Tumon Bay and some can
still be seen there today.
The American World War II Period (1944-1948 CE)
On July 8, 1944 the US began air attacks on Guam and landed on Agat and
Asan on July 21 of the same year. After the Orote Peninsula was succeeded to the
Americans on July 27th, the Japanese abandoned the southern portion of the island and
fled to the north for the last hold out. In August, the Americans had officially re-captured
Guam, although fighting continued until the last of the Japanese forces surrendered in
September 1945 (Gailey, 1988; Denfeld, 1997).
The Second American Period (1945-Present)
A massive military build-up by the US armed forces began prior to the end of WWII
to assist in continued bombardment of Japan. In 1948, control of Guam was transferred
from the US Navy to the Department of the Interior and the Organic Act of 1950 gave
the Chamorro US citizenship. In 1970, the locals elected their first governor and today
Guam is a US territory with a continued US military presence (Welch et al., 2005).
Extensive damage to Guam occurred due to both massive bombing and post-war
modification of the island landscape through bulldozer clearing and leveling by the US
Navy Seabees and extensive construction. The area of Tumon Bay is currently a

70
popular tourist area and has undergone significant alterations due to post-war
construction of resorts and parks for recreational use (Welch et al., 2005).

71
Table 2-1. Archaeological and historical chronological sequences of the Marianas Islands
Dates Spoehr (1957) Craib (1990) Moore and Hunter-Anderson (1999)
Hunter-Anderson and Moore (2001)
0 BP
AD 1521 Historic (Protohistoric) Latte AD 1521 AD 1521 500 BP
1000 BP Latte Latte Mochong Latte Latte
1500 BP
Pre-Latte
Transitional Ypao
Transitional Huyong
2000 BP
Intermediate Pre-Latte Late Unai 2500 BP Intermediate Pre-Latte
3000 BP
Tarague Early Pre-Latte
Middle Unai
3500 BP Early Unai
Table 2-2. Activities and artifact assemblage composition of latte setsa
Cooking Tool Manufacture/Maintenance Fishing Warfare
Prepared coral floors Shell and stone debitage Stone and shell net sinkers Slingstones
Ovens/cooking areas Unfinished fish hooks Stone and shell line sinkers Bone spear points
Fire-cracked rock and charcoal Tridacna shell blanks Bone and shell fishhooks
Pottery Chert or basalt cores Fish gorges Storage and cooking vessels Hammerstones
Mortars and pestles Adzes Stone drills/perforators Shell/coral files Bone awls and needles
a. Compiled from Graves (1986) survey

72
Figure 2-1. The Mariana Island Chain (Source: http://commons.wikimedia.org/
wiki/File:Casta_Marianas.jpg. Accessed on 22 July 2012)

73
Figure 2-2. Paleoshoreline notches along the western coast of Guam (Photo by author)
Figure 2-3. Reconstructed Latte set from Reinman’s Talofofo River Valley Site (Photo
by author)

74
Figure 2-4. Examples of Pre-Latte Period pottery. Photo taken at Lina’la’ Chamorro
Cultural Park Museum (Photo by author)

75
Figure 2-5. Island of Guam depicting Latte Set density and distribution, following
Hornbostel’s original 1920s survey (Map courtesy of Rad Smith/GANDA)

76
Figure 2-6. Examples of Latte Period pottery. Photo taken at Lina’la’ Chamorro
Cultural Park Museum (Photo by author)

77
CHAPTER 3 MATERIALS
Archaeological Sample
From 2006 to 2008, 376 prehistoric Chamorro burials were excavated by Paul H.
Rosendahl, PhD, Inc., a cultural resource management firm, which was later
incorporated into SWCA© Environmental Consultants, during an archaeological
mitigation project for the renovation of the Guam Aurora Villas & Spa in the Naton
Beach Site on the northern end of Tumon Bay, Guam. Of these, approximately 177 are
associated with the Pre-Latte Period and 190 belong to the Latte Period. The
determination of the burials’ age and cultural affiliation was based on the stratigraphic
location, associated artifacts, and radiocarbon dates (DeFant, 2008). Dating to roughly
2,500 BP, the Pre-Latte group represents some of the earliest settlers in Guam and is
the largest Pre-Latte mortuary sample discovered to date (Table 3-1). There are no
radiocarbon dates for the Latte sample; however, archaeological materials associated
with the Latte remains place the remains in the Latte period between AD 1000 to 1521.
A comprehensive osteological analysis was conducted by Ms. Cherie Walth, of
SWCA. All demographic information, as well as information on cranial and postcranial
metrics and non-dental pathological conditions, cited within this dissertation has been
provided by Ms. Walth and SWCA. A complete archaeological and osteological report
on the Naton Beach Burial Complex, including preliminary results from the current
study, is being prepared for submission as an unpublished manuscript to the Guam
Historic Preservation Office.

78
Taphonomic Bias
Bioarchaeological analysis in the Marianas is constrained by a number of
taphonomic factors. As is the case in many island environments, poor bone
preservation is the result of fluctuating temperatures, differential exposure to wet and
dry environments, and soil factors such as limestone-based soil matrices (Hanson and
Butler, 1997). Coastal skeletal assemblages are further perturbed by the effects of
changing sea levels (Dickinson, 2003), the mechanical effects of the waves leading to
soil and burial disturbance (Graves and Moore, 1985; Butler, 1995) and commingling of
neighboring burials.
A more prolonged exposure of Pre-Latte skeletal remains to the natural
environment, changing ecosystem, and human interaction may explain the dearth of
mortuary assemblages from the earliest inhabitants of Guam. Additionally, the Pre-
Latte settlements and burials are located much deeper in the ground and thus are not
as frequently encountered (Butler, 1995; Hunter-Anderson and Butler, 1991). Thus, an
overwhelming majority of skeletal studies in Guam focus on the Latte Period as Pre-
Latte remains were rarely recovered or too fragmented to provide any information on
the biological variation of Guam’s earliest prehistoric peoples.
Disturbances due to human activity also affect bone preservation, as
archaeological studies have noted that in many cases, skulls and long bones are often
missing from primary interments (Thompson, 1932; Spoehr, 1957; Reinman, 1977).
Ethnohistoric accounts note that the Latte believed that their ancestors’ skeletal remains
possessed power, which could be called upon by descendants for supernatural
purposes (Thompson, 1940; Driver, 1983). Thus, skeletal remains were buried beneath
the Latte sets, as a means for the familial social unit to demonstrate their loyalty and

79
identify with a particular lineage as well as to show strength through close proximity to
their ancestors (Thompson, 1945; Graves, 1986).
Further, Latte burials were often placed in shallow pits, close to the surface,
allowing for easy access and selective removal of skeletal elements: skulls were most
often removed from male burials, while long bones were often taken from female burials
(Spoehr, 1957; Yawata, 1961; Reinman, 1977; Hanson and Gordon, 1989). A
Franciscan priest, Fray Juan Pobre, reported: ‘the one thing for which they have high
regard are the skulls of the ancestors, especially those of their parents and
grandparents’ (Driver, 1983: 214), which were often used in ritualistic activities. Long
bones, on the other hand, were more utilitarian in purpose and were used to make tools
and weapons, such as spearpoints, harpoons, and awls (Hanson and Gordon, 1989).
Prehistoric habitation sites along Tumon Bay have also been impacted by modern
modifications to the landscape. During the 1930s the Tumon coastline was altered
during the occupation of the Japanese who built commercial establishments and
extensive fortifications around the bay (Defant 2002). Post-war modification of the
entire island landscape occurred through bulldozer clearing and leveling performed by
U.S. Navy Seabees after the American take-over (Fulmer et al., 1999). Additionally, in
recent times, Guam has become an increasingly popular destination for tourists over the
past two decades. The increase in tourism brought with it rapidly expanding economic
growth (Hanson and Butler, 1997). Thus, new developments such as hotels, strip malls,
and parks are being built along the bay leading to the accidental disinterment of skeletal
assemblages. Effects of building and construction, using large machinery can result in
fragmentation and commingling of skeletal elements.

80
Sample Population
A total of 215 individuals dentitions were analyzed from the Naton Beach mortuary
sample, 103 from the Pre-Latte Period and 112 from the Latte Period (Table 3-2).
Females, n = 77, were slightly more represented than males, n = 62; however, 76 were
of undetermined sex (Table 3-3). Age cutoffs for the current study are as follows: child
< 10 years of age; juvenile = 10 to 18 years of age; young adult = 18 to 25; middle-aged
adult = 30 to 45; older adult > 40 years of age. The majority of the assemblage is
composed of adult individuals (n = 179). Only juveniles with permanent dentition (n =
36) were analyzed are included in this analysis. The majority of the sample is
composed of young adults (n = 107), followed by middle-aged adults (n = 55), and old-
adults (n = 4). Thirteen individuals were placed into the ‘adult’ category without any
further delineation of age grouping (Table 3-4).
Pre-Latte Demographics
The Pre-Latte sample is represented by 103 individuals, 38 females and 33 males.
Sex could not be determined for 32 individuals. All but 11 individuals are adults (n =
93). More than half of the Pre-Latte assemblage is composed of young adult individuals
(n = 64; 62.1%), followed by middle-aged adults (n = 23; 22.3%). No old-adults were
analyzed. For 5 individuals, only a category of adult age was possible (Table 3-5).
Latte Demographics
The Latte sample is represented by 112 individuals. Males (n = 29) are
underrepresented in the sample in comparison with females (n = 39). Sex could not be
determined for 44 individuals. The majority of the Latte Period individuals are adults (n
= 87), with only 25 subadults represented in the sample. Young adults (n = 43; 38.4%)
make up the majority of the population, followed by middle-aged adults (n = 32; 28.6%),

81
and lastly old-adults (n = 4; 3.6%). Eight individuals are placed in the ‘adult’ category
due to lack of other skeletal indicators to further delineate age (Table 3-6).

82
Table 3-1. Radiocarbon dates from Naton Beach site, Tumon, Guam.a
Sample No. Provenience Material
Measured radiocarbon age
Conventional radiocarbon age
Calibrated 2σ age range
Beta- 238482 Feature 2 Soil 1700±40 BP 1680±40 BP AD 250–430
Beta- 238483 Burial 173b Conus shell beads 2490±40 BP 2940±40 BP 770–400 BC
Beta- 238484 Burial 156b Conus shell beads 2330±40 BP 2790±40 BP 590–330 BC
Beta- 238485 Burial 273b Conus shell beads 2490±40 BP 2860±40 BP 720–360 BC
Beta- 238486 Burial 286b Conus shell beads 2640±40 BP 2970±40 BP 790–490 BC a Table recreated from Defant (2008:150). b All burials are from the Pre-Latte assemblage.

83
Table 3-2. Pre-Latte vs. Latte sample distribution
Time Period N
Pre-Latte 103
Latte 112
Total 215
Table 3-3. Okura dental sex distributions
Frequency Percent
M 62 28.8
F 77 35.8
INDT 76 35.3
Total 215 100
Table 3-4. Okura dental age distributions
Frequency Percent
CLD 18 8.4
JUV 18 8.4
YA 107 49.8
MA 55 25.6
OA 4 1.9
ADT 13 6
Total 215 100
Table 3-5. Pre-Latte dental sample by age and sex
Pre-Latte Male Pre-Latte Female
Pre-Latte Indet.
Total
n % n % n % n %
CLD 0 0.0 0 0.0 6 18.8 6 5.8
JUV 0 0.0 2 5.3 3 9.4 5 4.9
YA 19 57.6 30 78.9 15 46.9 64 62.1
MA 13 39.4 6 15.8 4 12.5 23 22.3
OA 0 0.0 0 0.0 0 0.0 0 0.0
Adult 1 3.0 0 0.0 4 12.5 5 4.9
TOTAL 33 100.0 38 100.0 32 100.0 103 100.0
% is shown as percent of female/male/sex indeterminate quantity except for totals which are based on percent of total sample.
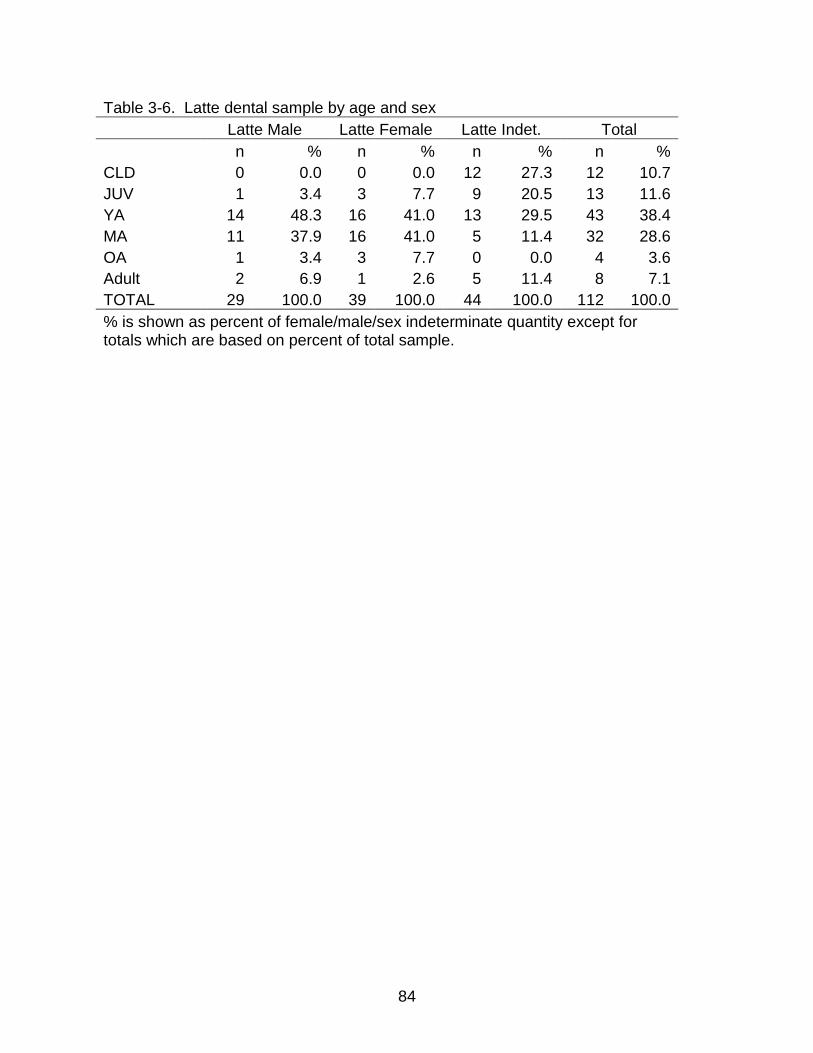
84
Table 3-6. Latte dental sample by age and sex
Latte Male Latte Female Latte Indet. Total
n % n % n % n %
CLD 0 0.0 0 0.0 12 27.3 12 10.7
JUV 1 3.4 3 7.7 9 20.5 13 11.6
YA 14 48.3 16 41.0 13 29.5 43 38.4
MA 11 37.9 16 41.0 5 11.4 32 28.6
OA 1 3.4 3 7.7 0 0.0 4 3.6
Adult 2 6.9 1 2.6 5 11.4 8 7.1
TOTAL 29 100.0 39 100.0 44 100.0 112 100.0
% is shown as percent of female/male/sex indeterminate quantity except for totals which are based on percent of total sample.

85
CHAPTER 4 DENTAL REDUCTION
Over time, and throughout the world, there has been an overall decrease in tooth
size in the human dentition (Brace et al., 1987). The majority of the studies looking at
dental reduction focus on dental trends over relatively long periods of time, from
prehistoric Pleistocene populations to contemporary populations (Dahlberg, 1960;
Greene, 1972; Carlson and van Gerven, 1977; van Gerven et al., 1977; Smith et al.,
1984; Calcagno, 1986; Smith et al., 1986; Brace et al., 1987; Calcagno, 1989; y’Edynak,
1989). Others have focused on microevolutionary trends of dental reduction across
shorter periods of time. For example, Christensen (1998) performed an odontometric
analysis in a prehistoric Oaxaca Valley population spanning from 2600 BP to 1521 CE.
He found a dramatic 4.4% reduction in size between the earliest and the latest
temporally dispersed groups. Pinhasi and colleagues (2008) found uniform reduction in
the buccolingual dimensions of the dentitions of Southern Levant populations ranging
from 12,000 to 7,000 BP. Brace and colleagues (1987) suggest that dental reduction
began in humans during the Late Pleistocene at a rate of 1% over 2,000 years.
However, the rates of reduction have increased to 1% every thousand years, from the
Post-Pleistocene to the current day (Brace et al., 1987).
Background
The exact mechanism for decrease in tooth size is unknown, however, four main
theories aim to explain this phenomenon. The Probable Mutation Effect (PME), as
proposed by Brace and colleagues (Brace 1963; Brace and Mahler 1971; Brace and
Hinton 1981; Brace 1987), suggests that mutations are the primary forces acting on
dental reduction. Whereby, a relaxation of selective forces due to a change in food

86
processing techniques, may allow for an accumulation of mutations, which lead to
decreased tooth size. Calcagno (1986, 1989) introduced the Selective Compromise
Effect (SCE), suggesting that decreased surface area and tooth complexity results in
fewer carious lesions, thus increasing an individual’s fitness. As such, selection for
smaller teeth is the result of an overall decrease of dental dimensions leading to dental
crowding and increased potential for cariogenic disease. The Increasing Population
Density Effect (IPDE) suggests that selection for smaller body size, due to increased
population density, sedentism, and reduced nutritional requirements, results in a
reduction of tooth size (Macchiarelli and Bondionli 1986). Lastly, Carlson and Van
Gerven (1977) adopt a biomechanical approach in their Masticatory-Functional
Hypothesis (MFH) and suggest that shifts in subsistence patterns, such as consuming
softer and more processed foods, led to selection for smaller teeth. They propose that
a shortening of the craniofacial complex, as a result of decreased functional demands of
mastication, led to a compensatory reduction in the size of the dentition.
Current Study
The current study investigates evolutionary dynamics of the prehistoric Chamorro
population to see how they relate to biocultural and environmental changes in
prehistoric society. The Pre-Latte (3500 BP – 1000 CE) and Latte (1000 – 1521 CE)
periods in Guam convey distinctions, not only in population size, but also in diet and
subsistence strategies. The Pre-Latte are characterized by small populations, with semi-
permanent settlements, that subsisted on bivalve shellfish, reef fishing, minimal
agriculture, and limited terrestrial resources consisting of birds, crabs, and bats
(Amesbury et al., 1991; Hunter-Anderson and Butler, 1991; Pietrusewsky et al., 1997).

87
Thin pottery, associated with cooking, has also been found in association with Pre-Latte
sites (Amesbury et al., 1991).
A significant increase in population numbers and densities occurred during the
Latte period ~1000 AD, coupled with an intensification of agriculture (Hunter-Anderson
and Butler, 2001). While archaeological records indicate that there was a shift from
bivalves towards pelagic fish and gastropods (Graves and Moore, 1985), exploitation of
marine resources declined in the Latte Period (Butler, 1988; Ambrose et al., 1997;
Pietrusewsky et al., 1997). Material artifacts associated with food procurement and
cooking include mortars and pestles for food processing, thick pottery, bone spear
points, and composite fish hooks (Ambrose et al., 1997, Amesbury et al., 1991).
Previous investigations show that the Chamorro dentition is some of the largest in
the world, intermediate between the larger Melanesian dentition and the smaller
Polynesia dentition (Brace et al., 1981; Brace et al., 1990; Hanihara and Ishida, 2005).
However, these studies pool samples of varying time periods and island samples
together in their investigations. The large Naton Beach skeletal assemblage allows for
within site diachronic comparisons to be made of the earlier Pre-Latte settlers to the
later prehistoric inhabitants. This study therefore represents the first large within-site
diachronic study of dental size reduction in Guam.
While the aforementioned accounts have shown that dental changes can occur
over short evolutionary time spans, the Pre-Latte and Latte periods span an even
narrower time range, around 1500 years, than those previously investigated. However,
two studies comparing immigrant dental dimensions in contemporary populations
showed significant differences between parents and offspring dental sizes due to

88
differential access to nutritional resources (Goose, 1967; Goose and Lee, 1973). Thus,
change in tooth size can occur over relatively short periods of time.
The null hypothesis is as follows:
Ho: There is no significant reduction in the dental dimensions between the Pre-Latte to Latte time periods.
I hypothesize that there will be dental reduction between the Pre-Latte and Latte due to
selective pressures during odontogenesis as a result of an increase in population size
(followed by decrease in health and competition for food resources) and change in diet
and food procurement strategies.
Expected Results
If a change is found between the Pre-Latte and Latte samples, the trends should
follow one of the proposed models for dental reduction and its associated assumptions
(Pinhasi et al., 2008).
With the PME, a change in cultural practices specifically related to food
preparation, such as tools and techniques for cooking, allow for a relaxation of selection
pressures in maintaining large teeth. Given the state of relaxed selection, mutations
begin to accumulate (Brace, 1964; 1967; 1978; Brace and Mahler, 1971; Wolpoff, 1971;
Wolpoff, 1975; McKee, 1984). Following Sewell Wright’s (1931; 1964) theory of
mutation pressure, which suggests that mutations lead to a reduction in structures,
Brace and colleagues posit that dental size decreases following selection relaxation and
accumulation of mutations. If the PME is the model by which reduction occurs,
reduction should occur equally in both the mesiodistal and buccolingual dimensions
from one time period to the next (Christensen, 1998; Pinhasi et al., 2008).

89
In SCE, there is a compromise between selection for large, complex teeth with
thick enamel and small teeth with thin enamel and less morphological complexity.
Large complex teeth provide a larger surface area for carious lesions and can contribute
to dental crowding. However, a large surface area and thick enamel are well adapted to
populations ingesting a coarse diet. Smaller teeth are more resistant to dental caries
and crowding but are less resistant against the high biomechanical demands of a
course diet (Calcagno, 1986; 1989). Additionally, untreated carious lesions would have
been problematic and may have lead to decreased health in the population. Thus, in
populations who have smaller forces placed on the dentition and are more prone to
dental caries due to a softer diet, selection would be for smaller teeth and an overall
reduction in the masticatory apparatus (Calcagno, 1986; 1989). If SCE is at work in the
temporally dispersed Chamorro populations, the buccolingual and mesiodistal
dimensions and tooth types should be differentially affected, with a constant amount of
variation between the time periods (Christensen, 1998; Pinhasi et al., 2008). Premolars
and molars are most likely to show greater dental reduction due to increased complexity
and higher prevalence for caries.
The MF model falls under the SCE theory (Calcagno, 1989). It suggests, in
accordance with Wolff’s Law of Transformation (Wolff, 1892), that the mandible and
maxilla will undergo apposition or resorption due to the biomechanical stressors placed
on them. Since teeth are more genetically controlled, a decrease on functional
demands placed on the masticatory apparatus would result in a decrease in jaw size but
not tooth size, thus causing dental crowding. Selective pressures would then restore
harmony by reducing the size of the dentition for a better fit within the maxilla and

90
mandible (Calcagno, 1989). If dental reduction follows the MF model, similar results as
what is expected from the SCE model would be expected. The varying effect on the
two dimensions does not suggest that a full sweep reduction of the masticatory
apparatus is at work. Instead, the dental dimensions are affected differently due to
changing functional demands placed on the skull and are not the result of overall
decrease of body size. Tooth classes are expected to be affected equally as a result of
overall dental reduction.
The IPDE model suggests that tooth reduction is a byproduct of smaller body size
as a result of an increase of population density. Thus, new adaptive pressures, such as
environmental stress due to a decrease in nutritional resources, trigger selection for a
reduction of body size and as such, dentition also becomes smaller (Machiarelli and
Bondioli, 1986; Pinhasi et al., 2008). If IPDE is at work, all teeth and both dimensions
should be affected equally. Further, dental reduction should correspond to increase in
population size and carious lesions (or other pathological indicators) (Christensen,
1998; Pinhasi et al., 2008).
The data presented in this study will be analyzed to determine if there is a trend in
dental reduction. If reduction is observed, the aforementioned models will be examined
to determine which best fits the data of the current study.
Methods
Two standardized measurements were taken on each available tooth using
Mitutoyo Digital Extended Pointed Jaw calipers following Moorees (1957) and Mayhall
(1992). These fine-tipped calipers allow for precise measurements of both isolated
teeth and those within the alveolar process. The mesiodistal diameter (MD), or the
length of the crown, was obtained by measuring the greatest distance between the

91
mesial and distal portion of the tooth, as expected in proper anatomical position. The
buccolingual diameter (BL), or the width of the crown, was obtained by measuring the
width of the tooth, perpendicular to the mesiodistal plane. All measurements were
taken to the closest 0.01 mm and were not taken on teeth with moderate to extreme
amounts of wear.
Following Brace (1979, 1980), the mesiodistal and buccolingual diameters were
multiplied to attain the cross-sectional area (CX) of each tooth class. The left and right
antimeres of each tooth are representative of the same genotype, thus two sides were
averaged to best express the cross-sectional area of that tooth class (Brace, 1990).
Tooth summaries (TS) were also calculated following Brace (1978). The tooth
summary is the sum of the upper and lower mean cross-sectional areas of each tooth
category. This number allows for a quick comparison of mean tooth size between
groups and represents an approximation of the total occlusal area of the population.
The data were tested statistically and visually for normality using the Kolmogorov-
Smirnov test and normal Q-Q plots on each dental measurement. Levene’s test of
homogeneity of variance was also performed on each measurement.
A two-way factorial Analysis of Variance (ANOVA) was performed to evaluate the
effect of time period and sex on each measurement using the following model:
Tooth measurement = Time Period + Sex + Time*Sex + error
where the tooth measurement is the dependent variable, time period and sex are the
independent variables (fixed factors), and Time*Sex is the interaction between those
two variables.

92
In order to prevent Type I error, in where a null hypothesis is rejected when in
actuality, it should not be rejected, a sensitive post hoc Bonferonni correction was
applied to prevent inflation of the alpha level when conducting multiple tests (Abdi,
2007). The Bonferonni corrected alpha was calculated by taking the desired alpha
(0.05) divided by the number of ANOVAs run (n = 65). Thus, the Bonferonni alpha is
0.00078. Significance levels of the models were analyzed using the Bonferonni alpha to
determine which measurements are significant. Once these measurements were
identified, significance levels of the time period, sex, and interaction between the two
were evaluated using a 0.05 alpha level.
Results
A total of 215 individuals permanent dentitions were analyzed. From these, 1242
individual teeth were measured. Appendix A presents the overall descriptive statistics
of the dental metrics and the cross-sectional areas of the teeth divided by sex and
population.
The Pre-Latte sample is composed of 633 teeth. Female teeth (n = 299) are
represented more frequently than male teeth (n = 189). The Latte sample is composed
of 609 teeth. Again, female teeth (n = 243) are represented more often than male
dentition (n = 177).
The tooth summary for the Pre-Latte population is very large, 1423.2 mm (Table 4-
1). As expected the male dentition is relatively larger than the female teeth (TS =
1470.9 mm and 1383.6 mm, respectively). The largest measurement of the dental
arcade is the mesiodistal diameter of the mandibular first molars in both males and
females (male: L = 12.7 mm; R = 12.9 mm and female: L = 12.5 mm; R = 12.3 mm) (see
Appendix A, Table A-3). However, in terms of overall cross-sectional area, maxillary

93
first molar is the largest tooth in both males (AVG CX1 = 147.5 mm1) and females (AVG
CX1 = 138.2 mm2) (see Appendix A, Table A-5).
The overall tooth summary for the Latte sample is large, 1307.1 mm, however, it is
significantly smaller than the Pre-Latte tooth summary size. As expected, the tooth
summary for the male dentition (TS = 1377.0 mm) is larger than that of the female
dentition (TS = 1250.2 mm). Following the trend seen in the Pre-Latte dentition, both
males’ and females’ mesiodistal diameter of the mandibular first molar is the largest
measurement (males: L = 12.7 mm; R = 12.6 mm and females: L = 12.1 mm; R = 12.1
mm) (see Appendix A, Table A-3). In males, the largest tooth is the mandibular first
molar with a cross-sectional area of 139.1 mm2. However, the maxillary first molar is
the largest tooth in females with a cross-sectional area of 128.1 mm2 (see Appendix A,
Table A-5).
A comparison of group measurements and cross sectional area shows the
direction of the differences between the groups (Appendix A-6). All of the variables
except LMax C BL and LMax C CX have a larger mean value in the Pre-Latte group
versus the Latte, showing reduction has occurred in the dentition. The percent of
change shows which teeth have the highest and lowest rates of reduction. Overall, the
percent of change in cross sectional areas are larger than in the buccolingual than in
the mesiodistal diameters. The biggest difference is seen in the cross-sectional area of
the mandibular third molars: R mandibular molar (20%) and L mandibular molar (17%).
The following teeth display a 10% or more decrease in cross-sectional areas: left and
1 This cross-sectional area is the average measurement of the left and right sides.

94
right maxillary molar (14%), right fourth premolar (11%), left mandibular second molar
(10%) and, left and right mandibular first incisors (10%).
Kolmogorov-Smirnov tests of normality showed that the majority (52 of 64) of
measurements were normally distributed (Appendix A-7). Likewise, the data are tested
for homoscedasticity using Levene’s test and all but three have equal variance
(Appendix A-8).
A two-way factorial ANOVA was run on each of the 64 measurements (Appendix
B, Tables B-1 through B-64). Thirty-nine of the 64 models were found to be statistically
significant using the Bonferonni corrected alpha of 0.00078 (labeled red in Appendix B,
Tables B-1 through B-64). Of these significant models, 28 have statistically significantly
differences between the time periods, 29 had significant sex differences, and four had a
significant interaction between time and sex, at a 0.05 alpha level (labeled green in
Appendix B, Tables B-1 through B-64).
Pre-Latte and Latte Differences
Very few measurements of the anterior dentition (incisors and canines) are
significantly different between the two time periods. The only anterior teeth with
statistically significant differences are the left maxillary incisor BL, left mandibular incisor
MD, and left mandibular canine BL. The remaining 25 measurements with significant
differences are in the premolars and molars.
Comparing the maxillary and mandibular dentition shows that a little less than a
quarter of the measurements with significant differences are located in the upper jaw
(15 of 64; 23.4%) and half are from the lower jaw (13 of 64; 20.3%), with the maxillary
measurements represented at a slightly higher rate.

95
The number of significant differences of the buccolingual measurements far
outweighs the mesiodistal measurements. Seventeen buccolingual measurements
(26.6%) are significantly different between the time periods while only eleven of 64
(17.2%) of the mesiodistal measurements are significantly different between the time
periods.
Male and Female Differences
As was seen in the time period differences, the majority of the measurements that
display significant differences between the sexes come from the posterior dentition.
Only seven anterior teeth display significant differences between the sexes and only
one of these measurements comes from the left maxillary lateral incisor BL. The left
and right maxillary MD as well as both the buccolingual and mesiodistal measurements
for the right and left mandibular canines are also significantly different between the
sexes. The remaining 22 measurements with significant differences are on the
premolars and molars.
Significant sex differences between the maxillary and mandibular dentition are
found in approximately 20% of the upper and lower teeth (21.9% and 23.4%,
respectively). Sex differences between the buccolingual and mesiodistal
measurements are not as pronounced as was seen when comparing time periods.
Significant differences in dental dimensions are nearly equal, with 25.0% (16 of 64) of
the buccolingual measurements and 20.3% (13 of 64) mesiodistal measurements being
significantly different.
Interaction between Time and Sex
Only six measurements have a significant interaction between time period and
sex: left maxillary lateral incisor BL, left mandibular fourth premolar MD, right

96
mandibular canine BL, right mandibular second molar MD, right mandibular fourth
premolar BL, and right mandibular first molar MD. Two of the six measurements are in
the anterior dentition, while the remaining measurements belong to the posterior teeth.
All measurements, except the upper incisor dimension, come from the mandible. The
buccolingual and mesiodistal measurements are represented equally.
Discussion
Tooth Summaries and Rate of Change
In both samples, the male tooth summaries are larger than the female tooth
summaries, indicating some degree of dimorphism between males and females.
However, there is a slightly larger discrepancy in male to female tooth size in the Latte
sample (1377 mm and 1250 mm, respectively) than in the Pre-Latte sample (1471 mm
and 1384 mm, respectively). Thus, the Latte sample displays a higher degree of sexual
dimorphism than the Pre-Latte sample, with a 10% difference between sexes compared
to a Pre-Latte difference of 6%.
Overall, the Pre-Latte tooth summary is much larger than the Latte tooth summary
(1423 mm vs. 1307 mm). Both overall tooth summaries, and those separated by sex,
the Pre-Latte tooth summaries are much larger than the Latte tooth summaries. These
data points to a small amount of dental reduction over time. Overall, the dentition has
reduced in size by 8% between the Pre-Latte and Latte time periods.
Previous studies in the prehistoric Chamorro populations in Guam, as shown in
Table 4-2 (see table for references), have shown tooth summaries ranging from 1034
mm to 1487 mm (Bath, 1986; Pietrusewsky, 1986; Douglas and Ikehara, 1992; Trembly,
1999; Pietrusewsky et al., 2003). The Leo Palace sample, recovered from Tumon Bay,
south of the current Naton Beach sample, represents the smallest tooth size in the

97
range. The largest tooth summary comes from the Fujita sample, which were also
recovered from Tumon Bay. The Naton Beach tooth summaries from both time periods
fall within this range. The Pre-Latte sample summaries fall on the high end of the
range, closest to the Fujita sample with a tooth summary of 1423 mm. Interestingly, the
Fujita sample is the only other Pre-Latte sample available for comparison.
The Latte Period sample from Naton Beach has the largest tooth summary size
(1307 mm) in comparison to the rest of the dentition from the Latte Period and is
followed closely by the San Vitores, Right-of-Way sample (1294 mm). In the remaining
comparative samples, the tooth summaries fall far below 1300 mm. A comparison of
the Pre-Latte and Latte samples show that the Pre-Latte teeth, from both Naton Beach
and Fujita, are much larger than all of the Latte groups analyzed.
When comparing the Chamorro dentition to other samples in the Asia-Pacific
region, the Pre-Latte Fujita sample (1487 mm) has the largest tooth summary and is
even larger than the oft-reported largest Australian (1486 mm) and Tasmanian (1429
mm) dentitions (Table 4-3, see table for references). The tooth summary size of the
Pre-Latte Naton Beach sample of the current study (1423 mm) follows closely behind
the Tasmanian tooth summary size. The Latte Naton Beach sample (1307 mm) is
relatively smaller and falls between the Hornbostle-Thompson Collection in Guam (1309
mm) and the Vanuatu sample (1295 mm).
Analysis of the group comparison of mean measurements and cross-sectional
areas shows the difference in mean measurements over time as well as the percent of
change. In all of the variables, except for two, the measurements and cross-sectional
areas show a reduction in mean size over time. Two variables left maxillary canine, BL

98
and CX, show a slight increase in size from Pre-Latte to Latte time periods. The
buccolingual measurement shows a rate of change of -0.62% while the cross-sectional
area variable has a -1.26% of change. Regardless of direction of change, these
numbers are quite small and may not have any biological significance or may simply
attest to the evolutionary importance of large canine size and it’s genetic stability. The
largest amount of dental reduction occurred in the lower third molars followed by the
upper second molars, upper right fourth premolar, and lower first incisors.
Hypothesis Testing
When looking at the Pre-Latte tooth summaries from both Naton Beach and Fujita,
it is apparent that the Pre-Latte teeth are larger than all other reported tooth sizes from
the Latte Period. Hypothesis testing, using two-way factorial ANOVA on 64 variables,
was performed to determine the significance of these observed differences. Of the
variables, 61% of these models are significant. Tooth size is significantly affected by
time period in 44% of the cases, by sex in 45% of the cases, and the interaction
between time period and sex in only 6% of the cases. While sex seems to have a
slightly higher effect on the size of the dentition, there is not a very large interaction
between time period and sex. Thus, there does appear to be a significant decrease in
size between the Pre-Latte and Latte samples in some measurements and not others.
The null hypothesis of no difference in tooth size between the time periods can be
rejected.
Time Period Differences
The majority of the significant differences between the Pre-Latte and Latte are
relegated to the buccal dentition with only 5% of the significant differences belonging to

99
the incisors and canines. Thus, over time the premolars and molars are reducing at a
significant rate while the anterior teeth are much more stable between the time periods.
There does not appear to be a large discrepancy in change in tooth size between
the maxillary and mandibular arcades. Approximately 20% of the significant
measurements are found in both the upper and the lower jaw, with the maxillary
dentition reducing at a 3% higher rate. The data show that both the maxillary and
mandibular dentition are reducing at approximately the same rate over time.
The buccolingual measurements reduce at a much higher rate than do the
mesiodistal measurements. Approximately one quarter, 27%, of the buccolingual
variables are found to be significantly different between the time periods, while only
17% of the mesiodistal measurements are affected by time. This finding is line with the
results from craniofacial data2, which show a higher decrease in width measurements,
in both the cranium and mandible, in comparison with length measurements over time
(see detailed discussion below) (Walth, pers. comm).
Comparison of dental, craniofacial, and postcranial changes across time2
In the following section, caution must be taken due to the fragmentary nature of
the skeletal remains and small sample sizes. Very few intact crania and long bones
were found and many of the estimates are taken on reconstructed bone. Thus, the data
presented may not represent the amount of variation seen in the population.
Nonetheless, limited data were collected and analyzed in an attempt to better
understand the differences in cranial size and height between the time periods.
2 All cranial and postcranial measurements were taken on the Naton Beach sample by Cherie Walth. Her
data, in conjunction with the data from the current study, were collected as part of a larger study on the Naton Beach Burial Complex, which is currently being compiled for submission to the Guam Historic Preservation Office.

100
Table 4-4 presents the cranial and mandibular measurements collected by Walth
(pers. comm.). Cranial length, cranial breadth, bizygomatic breadth, bimaxillary
breadth, and maximum alveolar breadth all show a decrease over time (Walth, pers.
comm.). The largest difference between the samples is seen in width measurements:
cranial breadth and bimaxillary breadth. Both males (Pre-Latte: 144.3 mm; Latte: 133.7
mm) and females (Pre-Latte: 136.0 mm; Latte: 128.8 mm) display a decrease of nearly
10 mm in cranial breadth (Walth, pers. comm.). While there are no data on bimaxillary
breadth in the Pre-Latte females, the males show a large reduction of maxilla width
(Pre-Latte: 123.0 mm; Latte: 106.0 mm) (Walth, pers. comm.).
Likewise, mandibular width has also undergone a decrease in size over time,
particularly in the bigonial and bicondylar measurements. Bigonial breadth in females is
unchanging over time (approximately 97 mm in both time periods); however, males
show a large decrease from the Pre-Latte (114.8 mm) to the Latte periods (106.5 mm)
(Walth, pers. comm.). The bicondylar breadth also displays differences between the
time periods in both males (Pre-Latte: 136.0 mm; Latte: 126.1 mm) and females (Pre-
Latte: 122.5 mm; Latte: 113.9 mm) (Walth, pers. comm.).
The cranial length trends differ between males and females. While the male
length decreases slightly over time (Pre-Latte: 187.3 mm; Latte: 183.5 mm), the female
length actually increases, albeit very little, over time (Pre-Latte: 176.3 mm; Latte: 179.0
mm) (Walth, pers. comm.). Comparably, mandibular length values remain fairly
constant in both males and females over time (Pre-Latte: M: 79.5 mm, F: 76.5 mm;
Latte: M: 79.2 mm, F: 75.4 mm) (Walth, pers. comm.).

101
The cranial and dental findings are complementary. Over time, there is an overall
trend in the reduction of the width of the craniofacial complex coupled with a significant
decrease in many buccolingual dental measurements. On the other hand, variables
that correlate with craniofacial length, such as cranial and mandibular length, as well as
mesiodistal diameter, remain fairly stable.
Stature estimates serve as a proxy for overall body size; however, as stated
above, caution must be taken with the following approximations as taphonomic factors
leading to severe fragmentation of the bone resulted in small sample sizes. The Pre-
Latte group, in particular, has a sample size of 12 for males and three for females.
Nonetheless, Walth’s stature data (pers. comm.)2 show very little difference in stature
between the time periods. The Pre-Latte male mean stature is 5’8” while the female
stature averages at 5’3”. The average Latte male height is 5’7” and the female mean
height is 5’2”. Overall, the Latte individuals are shorter than the Pre-Latte group, with
the mean stature differences varying by approximately one inch. While the sample size
for stature estimates is too small to make any definitive conclusions, nonetheless, the
observed decrease in stature is so minimal that it is unlikely that such a drastic
decrease in the size of the dentition is the result in overall body size reduction.
Carious lesions
The data on carious lesions are not discussed in detail in this section, as they will
be fully addressed in Chapter 6. However, a brief overview of the carious lesion data
will be presented to evaluate the modes of dental reduction. The data show a drastic
drop in the overall frequencies of carious lesions between the Pre-Latte and Latte
periods. Looking at carious lesion frequency per individual, the male frequencies drop
from 74% to 28% while the female frequencies decrease from 76% to 25%. However,

102
this drastic decrease in infectious disease may have more to do with a the introduced
cultural practice of betel-nut chewing, which has cariostatic properties, as opposed to an
selection for smaller and less complex teeth. Further, physiological stress, as
expressed by linear enamel hypoplasias, becomes more frequent in the Latte period
(see Chapter 5 for a more detailed discussion), serving as additional evidence that
health status did not between the Pre-Latte and Latte periods.
Mechanism for Dental Reduction
Various mechanisms for dental reduction have been previously discussed. To
reiterate, these theories are:
Probable Mutation Effect (PME) - Relaxation of selective forces due to a change in food processing techniques
Increasing Population Density Effect (IPDE) - Overall reduction of body size (and thus tooth size) due to increased population density, sedentism, and reduced nutritional requirements
Selective Compromise Effect (SCE) - Increase of fitness concomitant with decrease in carious lesions due to reduction of tooth complexity as a result of the reduction of tooth size
Masticatory-Functional Hypothesis (MF) - Decreased functional demands of mastication as a result of a shortened craniofacial complex.
These mechanisms are not necessarily mutually exclusive and it is possible that a
combination of these may be at work and the result of dental reduction over time.
Probable Mutation Effect
Brace and colleagues’ theory of PME (Brace, 1963; Brace and Mahler, 1971;
Brace and Hinton, 1981; Brace, 1987) as an explanation dental reduction is based on
Wright’s (1931; 1964) theory of mutation pressure and accumulation. Relaxation of
selection occurs after the advent of new cultural practices involved with cookery,
followed by an accumulation of mutations. Archaeological data suggest that there are

103
changes in the exploitation of certain foods and also an increased amount of food
processing associated with the Latte period populations (Graves and Moore, 1985;
Hunter-Anderson and Butler, 1991; Moore, 2005).
If PME were to work, it would not be advantageous for one dental dimension to
decrease more than another; thus, both the mesiodistal and buccolingual dimensions
would decrease at the same rate. The data show, however, a greater proportion of
buccolingual measurements rather than mesiodistal dimensions have significant
differences. As such, PME can be rejected as the mechanism of dental reduction,
regardless of the observed differences in food production techniques.
Increasing Population Density Effect
Between the Pre-Latte and Latte periods there was an increase in population
density (Hunter-Anderson and Butler, 1991). The IPDE model suggests that smaller
body size, and as a result smaller tooth size, is concomitant with an increase in
population size. Smaller body size reduces environmental stress due to competition for
nutritional resources (Machiarelli and Bondioli, 1986). All teeth and both dimensions
should be affected equally. Specifically, in an island environment where space is
limited, this seems like a plausible explanation.
However, analysis of height between the Pre-Latte and Latte periods shows little
variation between the two, in both males and females. Further, the data show that each
tooth type is affected differently with the posterior dentition reducing at a greater rate
than the anterior dentition. Likewise, the mesiodistal and buccolingual dimensions are
not affected equally. As such, dental reduction cannot be explained solely by smaller
body size, as hypothesized in the IPDE model.

104
Selective Compromise Effect
Calcagno (1986; 1989) suggests that a highly cariogenic diet will lead towards
selection for smaller dentition, which are more resistant to dental disease and crowding.
The increased dependence on agriculture, particularly taro, yams, and rice, during the
Latte Period would suggest a diet that is highly cariogenic. If SCE is at work, the
mesiodistal and buccolingual diameters should be affected differently, as malocclusion
would lead to selection for smaller mesiodistal diameters; whereas, selection for smaller
buccolingual dimensions would occur with large amounts of carious lesions (Greene,
1970; Sofaer, 1973; Christensen, 1998). Additionally, the posterior dentition would
show greater amount of dental reduction to reduce occlusal complexity and prevalence
for carious lesions.
There is a significant difference in the frequency of carious lesions between the
Pre-Latte and Latte with a drastic decrease in carious lesions in the later population.
This finding could be the result of decreased size and complexity of the dentition as well
as the culturally introduced practice of betel-nut chewing which is known to have
cariostatic properties.
The current study also demonstrates that the buccolingual and mesiodistal
dimensions are not equally affected. A predilection for reduction of the buccolingual
diameter is expected given the high number of carious lesions in the early population.
Further, the anterior dentition shows a much smaller rate of dental reduction, which is
primarily restricted to, and most prominent in the premolars and molars. Given these
findings, the SCE is a likely model to explain the dental reduction observed in the
prehistoric Chamorro.

105
Masticatory Functional Hypothesis
Following Wolff’s Law (1892), the MFH posits that dental reduction occurs as a
result of decreased functional demands placed on the craniofacial apparatus, which in
turn leads to a decrease in the masticatory complex (Carlson and Van Gerven, 1977;
Calcagno, 1989). The archaeological data show significant changes in food processing
techniques, as well as change in diet, associated with an increase in agricultural
processes and cookery that result in softer diets. These cultural changes may have had
such an impact that they led, indirectly, to biological changes, such as a decrease in the
craniofacial complex, which subsequently affected the size of the dentition.
Cranial and mandibular data show a decrease in size over time, particularly in the
width dimensions. As was previously discussed, this finding is not associated with an
overall decrease in body size, as stature remains fairly constant between the time
periods. Thus, it can be assumed that a decrease in the craniofacial complex is
associated with reduced biomechanical demands due to dietary and food processing
shifts. Therefore, the MFH model can also be utilized to explain the decrease in dental
size over time in Guam.
Conclusions
This study analyzed diachronic trends in dental dimensions of the prehistoric
Chamorro population on the island of Guam. An 8% reduction in tooth size is observed
from the Pre-Latte to Latte Periods. The results showed that significant reduction
occurred in 28 of the 64 analyzed dimensions. Reduction occurred most frequently in
the buccolingual dimensions and at a greater rate in the posterior dentition. The maxilla
and mandible were equally affected.

106
These findings may be best explained by a combination of the Selective
Compromise Effect and Masticatory-Functional Hypothesis. An increased reliance on
taro-based agriculture would have led to a highly cariogenic diet. In this situation, small
teeth would have been ideal, greatly reducing the complexity of the occlusal surface and
thus preventing formation of carious lesions. Additionally, increased food processing
techniques, such as the use of mortar and pestles and cooking, minimize the force
necessary to break down tough food, which lead to decreased functional demands of
the masticatory apparatus. These shifts occur over a relatively short period of time
where dynamic transitions in cultural practices may have been the catalyst for biological
change.

107
Table 4-1. Tooth summary data
Time Period Male Female Indet. Total
Pre-Latte 1470.9 1383.6 1463.9 1423.2
(n) (189) (299) (145) (633)
Latte 1377.0 1250.2 1318.8 1307.1
(n) (177) (243) (189) (609)
Table 4-2. Tooth summaries of prehistoric Chamorro populations from Guam
Study Sample Subsample Time Period Male Female Total Sample References
San Vitores Road
Fujita Drainfield Pre-Latte 1412.0 - 1487.0 Bath, 1986; Pietrusewsky, 1986
Naton Beach - Pre-Latte 1470.9 1383.6 1423.2 Current Study
Naton Beach - Latte 1377.0 1250.2 1307.1 Current Study
San Vitores Road
Right-of-Way Latte 1303.0 1281.0 1294.0 Bath, 1986; Pietrusewsky, 1986
Apurguan N/A Latte 1259.5 1236.9 1247.9
Pietrusewsky, Douglas, Ikehara-Quebral, 2003
Hyatt Hotel N/A Latte 1193.8 1138.0 1161.7 Trembly, 1999
Leo Palace N/A Latte 1058.5 985.6 1034.5 Douglas and Ikehara, 1992

108
Table 4-3. Tooth summaries of Pacific and circum-Pacific samples
Study Sample Sample Tooth Summary References
Guam, Fujita 1487 Pietrusewsky, 1986
Australia 1486 Brace, 1980
Tasmania 1429 Brace, 1980
Guam, Naton Beach (Pre-Latte) 1423 Current Study Papua New Guinea, East Highlands 1395 Brace and Hinton, 1981
Tonga 1371 Brace and Hinton, 1981
Bougainville 1359 Brace and Hinton, 1981
Tumon Bay 1347 Pietrusewsky, 1986a
Northern Marianas 1341 Pietrusewsky and Batista, 1980
Fiji 1338 Brace and Hinton, 1981
New Britain 1334 Brace and Hinton, 1981
Papua New Guinea, Sepik River 1321 Brace and Hinton, 1981
Samoa 1311 Brace and Hinton, 1981 Guam, Hornbostle-Thompson Collection 1309 Brace and Hinton, 1981
Guam, Naton Beach (Latte) 1307 Current Study
Vanuatu 1295 Brace and Hinton, 1981
Guam, Right-of-Way 1294 Pietrusewsky, 1986a
Phillippines, Visayas 1288 Brace and Hinton, 1981
Pohnpei, Nan Mandol 1287 Pietrusewsky and Douglas, 1985
Papua New Guinea, North Coast 1286 Brace and Hinton, 1981
New Ireland 1266 Brace and Hinton, 1981
New Caledonia 1256 Brace and Hinton, 1981
Java 1240 Brace and Hinton, 1981
Northern Marianas 1238 Pietrusewsky, 1986b
Thailand 1233 Brace and Hinton, 1981
Marquesas 1204 Brace and Hinton, 1981
Hawaii 1200 Brace and Hinton, 1981
Japan 1200 Brace and Hinton, 1981
Borneo 1190 Brace and Hinton, 1981
Chatham Islands 1181 Brace and Hinton, 1981
China 1157 Brace and Hinton, 1981
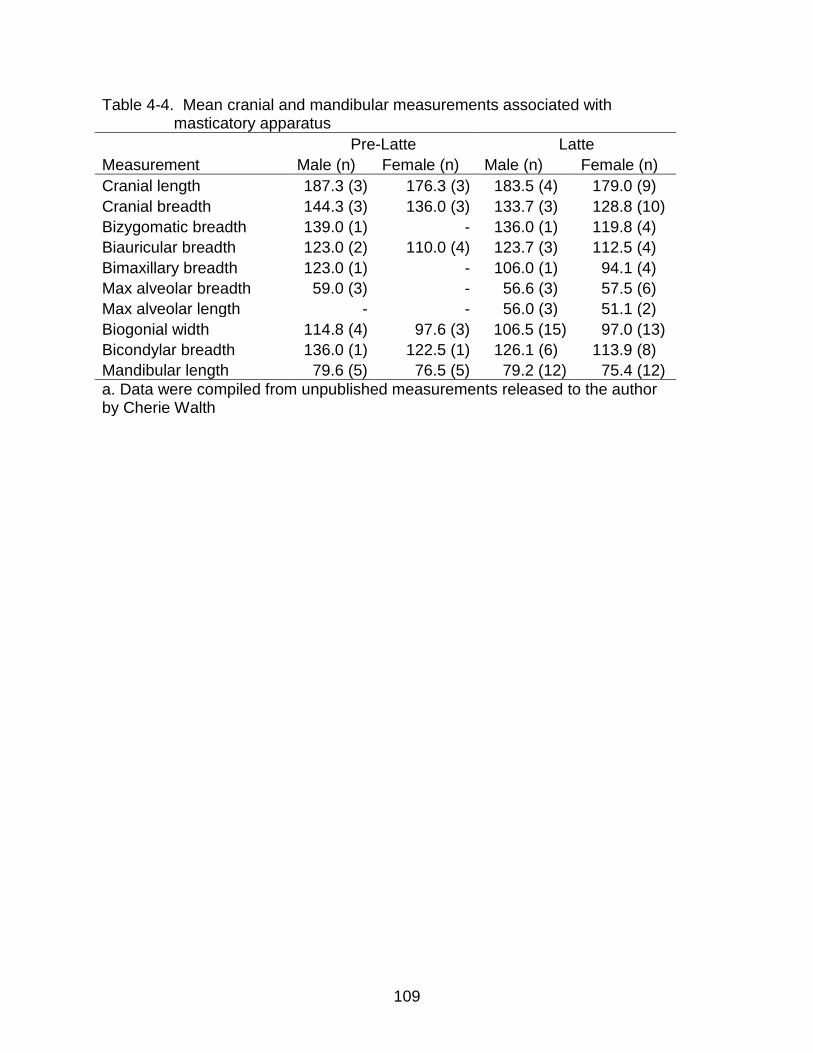
109
Table 4-4. Mean cranial and mandibular measurements associated with masticatory apparatus
Pre-Latte Latte
Measurement Male (n) Female (n) Male (n) Female (n)
Cranial length 187.3 (3) 176.3 (3) 183.5 (4) 179.0 (9)
Cranial breadth 144.3 (3) 136.0 (3) 133.7 (3) 128.8 (10)
Bizygomatic breadth 139.0 (1) - 136.0 (1) 119.8 (4)
Biauricular breadth 123.0 (2) 110.0 (4) 123.7 (3) 112.5 (4)
Bimaxillary breadth 123.0 (1) - 106.0 (1) 94.1 (4)
Max alveolar breadth 59.0 (3) - 56.6 (3) 57.5 (6)
Max alveolar length - - 56.0 (3) 51.1 (2)
Biogonial width 114.8 (4) 97.6 (3) 106.5 (15) 97.0 (13)
Bicondylar breadth 136.0 (1) 122.5 (1) 126.1 (6) 113.9 (8)
Mandibular length 79.6 (5) 76.5 (5) 79.2 (12) 75.4 (12)
a. Data were compiled from unpublished measurements released to the author by Cherie Walth

110
CHAPTER 5 DEVELOPMENTAL INSTABILITY: ENAMEL HYPOPLASIAS
Hans Selye (1973: 692) defines stress as ‘the nonspecific response of the body to
any demand made upon it’ where alterations in the environment change the normal and
steady state of an organism. During periods of environmental stress, an organism
undergoes neural and endocrinal responses to activate homeostatic mechanisms to
alleviate chronic stress events, which can leave lasting and permanent markers in the
body, and specifically in the skeleton (Selye, 1973; Goodman et al., 1988; Goodman
and Armelagos; 1989; Larsen, 1997; Goodman and Martin, 2002).
Goodman and Armelagos (1984) present a stress model applicable to skeletal
populations in which health is the fundamental variable in examining the adaptive
processes of a population (Larsen, 1997; Goodman and Martin, 2002). In this model,
markers of skeletal stress develop when new cultural systems fail to buffer
environmental stressors, such as disease or access to resources, therefore resulting in
physiological disruption in the skeletal remains.
Linear enamel hypoplasias (LEH) can be analyzed to estimate periods of non-
specific stress events in an individual or population. The current study investigates the
use LEH as indicators of physiological disruptions and stress in two temporally
disparate populations from the Western Pacific island of Guam, in an attempt to
understand the relationship between developmental instabilities and subsistence
change in a population undergoing agricultural intensification.
Enamel Hypoplasias
Enamel hypoplasias are deficiencies in enamel thickness that appear as horizontal
linear grooves or pits and are the result of a disruption in amelogenesis (Pindborg,

111
1970; Goodman and Armelagos, 1985). These defects are a permanent record of
developmental disturbances that occurred during prenatal and early childhood
development due to physiological stress and are a direct response to an anomaly in
matrix secretion that arrests ameloblastic activity (Pindborg, 1970; Duray, 1992).
Enamel hypoplasias reflect a nonspecific physiological disruption that occurred during
growth and development of the tooth. Thus, disruption of enamel formation may be
related to physiologic and metabolic stress where activation of the sympathetico-adrenal
medullary and pituitary-adrenal cortical axes induces stress (Rose et al., 1985). These
axes control output of hormones, which when elevated, will decrease the amount of
protein made throughout the body and in turn interrupt the process of amelogenesis
(Rose et al., 1985).
The problem with understanding enamel defects lies not in how the developmental
process of hypoplasias occurs, but in what external factors initiate the disruption
(Goodman and Rose, 1990). Experimental studies in rats have reported hypoplasia
formation as a result of exposure to various stressors such as malnutrition (Becks and
Furata, 1941), infectious agents (Kreshover and Clough, 1953; Kreshover et al., 1953),
and fever (Kreshover and Clough, 1953). These early studies solidified the current
notion that linear enamel hypoplasias are the result of non-specific physiological
disturbances, which could be attributed to many different external stressors. As such,
frequencies of enamel hypoplasias are often used to assess the relative health and
nutritional status of prehistoric, historic, and contemporary human populations (Massler
et al., 1941; Schiulli, 1978; Cook and Buikstra, 1979; Goodman et al., 1980; Cohen and
Armelagos, 1984; Larsen, 1997; Zhou and Corruccini, 1998; Cucina, 2002; Pechenkina

112
et al., 2002; Steckel and Rose, 2002; Hillson, 2005; King et al., 2005; Buzon, 2006;
Pechenkina and Delgado, 2006; Boldsen 2007; Berbesque and Doran, 2008), as well
as in Neandertals (Ogilvie et al., 1989; Guatelli-Steinberg et al., 2004), hominin
ancestors (Skinner, 1996; Guatelli-Steinberg, 2003; Cunha et al., 2004), and non-
human primates (Guatelli-Steinberg and Lukacs, 1999; Lukacs, 1999; Lukacs, 2001;
Skinner and Hopwood, 2004).
Health and Disease in the Shift to Agriculture
The agricultural revolution has long been thought of as one of the major advances
in human culture that led to the rise of civilizations (Braidwood, 1960; Cohen, 1989).
However, research presented in Cohen and Armelagos’s (1984) edited volume
‘Paleopathology at the Rise of Agriculture’ demonstrated that overall rates of disease in
prehistoric populations increased in the shift from hunter-gathering to agriculture. An
increase in infectious skeletal (yaws and tuberculosis) and dental diseases (carious
lesions), non-specific markers of stress (linear enamel hypoplasias) coupled with
decrease in stature, and life expectancies were reported as evidence for declining
health (Cohen and Armelagos, 1984). While Cohen and Armelagos’ book (1984)
changed the paradigm of health and agriculture, it was faulty in that it was skewed
towards North American populations practicing the domestication and intensification of
maize agriculture and it provided an overly simplified interpretation of the agricultural
transition. Additionally, in the mid-1980s, techniques and diagnoses had not yet been
standardized (e.g. Buikstra and Ubelaker, 1994; Steckel and Rose, 2002).
In the last two decades, vast improvements in methodologies have been made
and research linking decreased health with dietary shifts and agricultural intensification
has been conducted on nearly every continent: North America (Larsen et al., 2002),

113
South America (Ubelaker and Newson, 2002), Europe (Cucina, 2002), Africa (Keita and
Boyce, 2001; Keita, 2003; Starling and Stock, 2007), Asia (Yammamoto, 1988; Lukacs,
1992; Lukacs and Walimbe, 1998) Temple and Larsen, 2007; Temple, 2007) and
Australia (Webb, 1984; Seow et al., 1991). These studies have shown that declining
health cannot be explained by a single external stimulus and is multifactorial in nature.
In populations undergoing a shift to agriculture, reduced health is related to a
combination of increased population density and concomitant rise of infectious disease
associated with sedentism and reduced access to nutritional requirements and food
shortage due to drought or animal infestation related to the adoption of agriculture
(Pietrusewsky and Douglas, 2002).
However, the pattern of declining health with agricultural intensification is not
universal may have no effect or may have an opposite trend of improved health
(Hodges, 1987; Neves and Wesolowski, 2002; Pietrusewsky and Douglas, 2002; Eshed
et al., 2004; Douglas and Pietrusewsky, 2007). In some cases, paleodemographic
studies have shown an increase in fertility after agricultural intensification, suggesting
improved health status in the population (Bocquet-Appel and Naji, 2006).
For example, research from South East Asia does not appear to follow the trends
observed in other parts of the world. A skeletal series from northeastern Thailand,
spanning two millennia and including a period of agricultural intensification, shows
mixed results (Pietrusewsky and Douglas, 2002; Douglas and Pietrusewsky, 2007).
The earlier Non Nok Tha (5000 BP – 1700 BP) displays a decrease in linear enamel
hypoplasias and cribra orbitalia and increase in stature despite the reliance on starchy
foods (Douglas and Pietrusewsky, 2007); while the later Ban Chiang population (4100

114
BP – 1800 BP) has a decrease in carious lesions while LEH remain constant,
demonstrating little evidence for health decline (Pietrusewsky and Douglas, 2002;
Douglas and Pietrusewsky, 2007). Other skeletal assemblages in northeast Thailand
demonstrate a similar trend. Domett and Tayles (2007) observe an improvement of
health over time with an increase in height and similar rates of carious lesions in Bronze
Age (Ban Lum Khao: 3400 – 2500 BP) and Iron Age (Noen U-Loke: 2300 – 1600 BP)
individuals. The pattern of health and disease in Thailand is contrary to what is seen
with the North American populations.
Much of these results are attributed to the adoption of rice-based agricultural
practices instead of maize agriculture, which is prevalent in North America (Tayles et
al., 2000). Further, rice, and other starchy crops, may have more nutritional value than
other cultigens. Thus, many factors should be taken into account when attempting to
interpret health differences in the shift to agricultural intensification. Environmental and
genetic patterns may also come into play and affect the way cultural shifts in a
population are expressed biologically. As such, varying regions should be analyzed
independently to observe population-specific trends that accompany intensification of
agriculture.
Current Study
The shift between the Pre-Latte (3500 BP to 1000 CE) and the Latte (1000 to 1521
CE) time periods in Guam involves changes in population size, diet, and food
procurement/preparation strategies (Hunter-Anderson and Butler, 1991; Moore, 2005;
Amesbury, 2007). The Pre-Latte population is a small semi-nomadic, foraging
population that subsisted on bivalves, shellfish, and reef and pelagic fishing (Amesbery
et al., 1991; McGovern and Wilson, 1996). Horticultural practices were likely brought to

115
Guam by its earliest inhabitants from island Southeast Asia (Bellwood, 1978). Data
from paleoenvironmental sediment cores suggest small-scale gardening and resource
collecting by initial settlers (Athens and Ward, 2004), and analyses of pollen and starch
residues on the interior of Pre-Latte pottery found evidence of taro, cabbage tree, fish,
and shellfish (Loy, 2001a,b; Loy, 2002; Loy and Crowther, 2002).
Charcoal becomes increasingly present in paleoenvironmental sediment cores
around 1,800 BP, which corresponds to the Pre-Latte/Latte transition, and is suggestive
of intensified land use (Athens and Ward, 2004). Latte period archaeology has
identified a wide array of food processing artifacts, such as stone mortars and
pounders, scrapers, knives, blades, and adzes, which are indicative of a more
sedentary population with an increased reliance on agriculture (Hunter-Anderson and
Butler, 1991; Moore, 2005). Starch grain residues on Latte period potsherds identified
cabbage tree, rice, sugarcane, and a taro (Loy, 2001a,b; Loy, 2002; Loy and Crowther,
2002). Taro was identified more than any other plant, suggesting a preference for this
introduced tuber. Pollen and phytolith analyses have also found evidence of betel palm,
breadfruit, coconut, bananas, and pandanus (Hunter-Anderson and Butler, 1991;
Cummings and Puseman, 1998; Dixon et al., 1999; Pearsall and Collins, 2000; Ward,
2000; Cummings, 2002; Athens and Ward, 2004).
Stable isotope data reveal that the majority of the Latte diet was composed
primarily of terrestrial C3 resources, such as rice, root crops, and vegetables and that
marine foods are merely a supplement, comprising approximately 30% of the diet
(Hanson, 1991; Ambrose et al., 1997). Higher 13 N values are suggestive of
preferential exploitation of reef and/or lagoon fish over pelagic resources (Hanson,

116
1991; Ambrose, 1997). Ethnohistoric and archaeological evidence points to exploitation
of some larger pelagic fish such as tuna, dolphin, and marlin (Driver, 1993; Freycinet,
1996; Amesbury and Hunter-Anderson, 2008), however, deep-sea fishing is more
dangerous and thus not used as a reliable subsistence base (Russell, 1998). Thus, the
favored reliance of marine-based foods was abandoned for terrestrial resources and
agricultural crops in the Latte period. The stable isotope findings are further supported
by archaeological units demonstrating a greater proportion of fishing gear and fishing-
related debris in the Pre-Latte period in comparison with the Latte Period (Butler, 1995).
Additionally, intensification of agriculture, in the Latte period, would have permitted an
expansion in population size. Population growth is evident with more varied habitation
areas ranging from the preferred coastal locale to more marginal and inland
environments, such as the interior uplands (Hunter-Anderson and Butler, 2001).
Underwood (1973) and Hezel (1982) suggest that the prehistoric Latte population, at its
largest, ranged between 30,000 to 40,000.
The shift to agricultural intensification coupled with newly adapted food processing
tools is likely to be accompanied by biological changes, such as an increase in stress
levels associated with population growth, limited access to resources, malnutrition, and
increased prevalence of disease. This study is restricted to the dentition due to the poor
quality of skeletal preservation that is typical in island and coastal environments such as
Guam (Hanson and Butler, 1997). However, analyses conducted by Cherie Walth over
a year and a half period, will be considered to assist in the construction a better health
profile of the Chamorro population.

117
The current study compares the frequency of linear enamel hypoplasias between
the Pre-Latte and Latte time periods in Guam. Decreased health, as evidenced by
increased linear hypoplasias, is predicted concomitant with agricultural intensification,
increased population size, and increase of infectious disease. The null hypothesis
tested in this study is that there is no significant difference in the frequency of linear
enamel hypoplasias between the Pre-Latte and Latte time periods.
Materials and Methods
From 2006 to 2008, 376 prehistoric Chamorro burials were excavated during an
archaeological mitigation project for the renovation of the Guam Aurora Villas & Spa in
the Naton Beach Site on the northern end of Tumon Bay, Guam (DeFant, 2008). Of
these, approximately 177 are associated with the Pre-Latte Period and 190 belong to
the Latte Period (DeFant, 2008). The determination of the burials’ age and cultural
affiliation was based on the stratigraphic location, associated artifacts, and radiocarbon
dates (DeFant, 2008). Dating to roughly 2,500 BP, the Pre-Latte Naton Beach sample
represents some of the earliest settlers in Guam and is the largest Pre-Latte mortuary
sample discovered to date (DeFant, 2008). There are no radiocarbon dates for the
Latte sample; however, archaeological materials associated the remains are consistent
with those relegated to the Latte period, between AD 1000 to 1521.
This study looked at the horizontal grooves or linear type hypoplasias, most
commonly referred to in the literature as linear enamel hypoplasias (LEH), and
categorized as a ‘Type 4’ defect in the Developmental Defects of Enamel Index (FDI,
1982). The LEH were analyzed on the labial or buccal surfaces of the permanent
dentition in individuals older than 10 years of age. Age estimates were determined
skeletally using standards recommended by Buikstra and Ubelaker (1994) by Cherie

118
Walth1. Frequencies of LEH were examined in 197 individuals, 97 from the Pre-Latte
period and 100 from the Latte period. Distribution of sex is fairly even across the
samples with females being represented more frequently than males in both the Pre-
Latte (38 and 33, respectively) and Latte (39 and 29, respectively) periods.
LEH were scored macroscopically by both individual and tooth count methods, in
all available and unmodified teeth, as suggested by Lukacs (1989). LEH were not
scored in dentitions displaying moderate to extreme wear, which was defined as teeth
missing 75% or more of crown height. Using the individual count method, LEH was
scored as a discrete trait as present or absent. In the tooth count method, LEH were
analyzed in each tooth individually and scored as an ordinal trait using the following
scale: 0 = absence of LEH; 1 = one hypoplastic defect observed in a single tooth; 2 =
more than one LEH in a single tooth (Figures 5-1 & 5-2).
The effects of labial abrasion, betel-nut staining, and dental incising may have
biased the analysis of LEH. The Pre-Latte population displays a unique pattern of
dental modification that is not seen in the Latte period. Abrasion of the labial surface of
adult maxillary teeth, from the central incisors to the fourth premolar, ranges from slight
to extreme, and thus may obliterate evidence of linear enamel hypoplasias (Figure 5-3).
Additionally, central and lateral incisors are modified to a higher extent than the more
posterior teeth (Parr and Walth, 2011), further skewing the data analysis since these
teeth are also most likely to display LEH (Goodman and Armelagos, 1985). The Latte
dentition, on the other hand, displays dark reddish-brown staining on the teeth due to
1 Data analysis on the cranial and postcranial skeletal remains of the Naton Beach sample was conducted
by Cherie Walth. Her data, in conjunction with the data from the current study, were collected as part of a larger study on the Naton Beach Burial Complex, which is currently being compiled for submission to the Guam Historic Preservation Office.

119
the cultural practice of betel-nut chewing (Figure 5-4). This staining made LEH
observation more difficult. Thus, LEH were scored as present if they were a palpable
indention, through use of a fingernail, on the enamel surface. Intentional dental
modification of the anterior dentition, in the form of incising cross-hatched and oblique
patterns on the enamel surface, is also practiced in the Latte period (Ikehara-Quebral
and Douglas, 1997; Parr and Walt, 2012). Thus, LEH was not scored in teeth with
modified surfaces.
Frequency data by tooth and individual are reported. Multivariate statistical testing
was utilized to determine if observed frequency differences were statistically significant.
Differences between adult and subadult frequencies, sex, and time period were tested
using a Pearson Chi-Square test. Goodman and Armelagos (1985) demonstrated that
the maxillary central incisors and mandibular canines are the most hypoplastic teeth in
the dental arcade, as these teeth develop the earliest, tend to be more often affected.
As such, this study compared the LEH frequencies of the maxillary central incisors and
mandibular canines, using a Pearson Chi-Square test, to see if significant differences
exist between the time periods in these teeth.
Results
The mandibular canines and maxillary central incisors displayed the highest
frequencies of linear enamel hypoplasias in both periods (Table 5-1 and Figure-6). In
the Pre-Latte, the mandibular canine has the highest frequency of occurrence (n = 11;
29%), followed by the central maxillary incisor (n = 5;13%). Likewise, in the Latte
period, LEH is highest in the mandibular canine (n = 27; 17%) and the maxillary central
incisor (n = 21; 14%).

120
Age Differences
Frequency differences of LEH in the juvenile and adult populations are apparent
between the periods; however, caution must be taken due to the small sample size of
the juvenile individuals, particularly in the Pre-Latte population (n=5) (Table 5-2). In
both periods, the juveniles have higher frequencies of hypoplastic teeth than the adults.
In the Pre-Latte, 40% (n = 2) of the juveniles display LEH, while 15.2% (n = 14) of the
adults have LEH. The LEH frequencies increase for both the juveniles (n = 8; 61.5%)
and adults (n = 36; 41.4%) in the Latte period.
The LEH frequencies were tested using the Pearson Chi-Square test to determine
if the observed differences between subadults and adults were statistically significant;
significant differences in age were found in both the Pre-Latte or Latte time periods
(Pre-Latte: p = 0.001; Latte: p = 0.017, at the 0.05 alpha level) (Table 5-6).
Sex Differences
Sex specific differences for LEH expression are noted in both the Pre-Latte and
the Latte periods, by individual and tooth count (Table 5-3 and 5-4). Even with the low
frequencies of LEH in the Pre-Latte, there are still major differences between male and
female expression, where males exhibit LEH almost twice as frequently (n = 8; 24.2%)
as females (n = 5; 13.2%). In the Latte period, the opposite trend is observed. Almost
half (n = 19; 48.7%) of the female population displays at least one or more LEH, while
just over a quarter of the males exhibit LEH (n = 8; 27.6%). Looking at tooth count
incidence between the sexes, males from the Pre-Latte period display LEH more
frequently than females (14/634; 2.2% and 13/825; 1.6%; respectively); while in the
Latte period, the female dentition has a slightly higher incidence of LEH (54/670; 8.1%)
when compared to males (33/450; 7.3%).

121
Sex differences (with and without indeterminate sex category inclusion) were
tested to determine significance of linear enamel hypoplasia expression both between
and within each time period. Inclusion of indeterminate sex category did not affect the
outcome, however the results discussed are based on tests on only males versus
females. A Pearson Chi-Square test found no significance sex differences between
male and female LEH occurrence in either the Pre-Latte (p = 0.228) or Latte (p = 0.078)
time periods, at a 0.05 alpha level (Table 5-7).
Time Period Differences
Linear enamel hypoplasia frequencies are discussed both in terms of overall
individual frequency per time period (Table 5-3) as well as tooth count frequency (Table
5-4). Discrepancies are seen with expression of LEH between the time periods. The
Pre-Latte individuals have a low frequency of LEH, with only 16.5% (n = 16) affected
(Table 5-3), while LEH frequencies are much higher in the Latte where nearly half of the
population, 45.0% (n = 45), displays at least one or more LEH (Table 5-3). Likewise, in
terms of tooth count, the Latte dentition (155/1693; 9.2%) is much more highly affected
by LEH than the Pre Latte dentition (38/1925; 2%) (Table 5-4).
A Pearson Chi-Square test was performed to determine if the observed frequency
differences of LEH expression between time periods were statistically significant (Table
5-8). The Pearson Chi-Square test demonstrates that there is a significant difference,
at a 0.05 alpha level (p < 0.001), in expression of linear enamel hypoplasias between
the Pre-Latte and Latte time periods.
Goodman and Armelagos (1985) showed that the maxillary central incisors and
mandibular canines display the highest frequencies of linear enamel hypoplasias. Thus,
these teeth were tested independently across time periods with a Pearson Chi-Square

122
test. Results indicate significant differences in LEH expression in the maxillary central
incisors and mandibular canines between the time periods at a 0.05 alpha level: left I1: p
= 0.008; right I1: p = 0.031; left C1: p = 0.012; right C1: p = 0.038 (Table 5-8).
Discussion
The current study found that the maxillary central incisor and the mandibular
canines display the highest frequency of LEH. These findings are consistent with those
reported in other studies (Goodman and Armelagos, 1985; Hillier and Craig, 1992;
Stodder, 1997).
Age Differences
No significant differences in subadult and adult expression of linear enamel
hypoplasias were found between the time periods; however, when looking at the raw
frequency data both the Pre-Latte and Latte individuals displaying increase of LEH with
age. Both groups display similar rates of LEH increase, with subadults increasing by
22% and adults increasing by 26% between the Pre-Latte and Latte periods. These
findings suggest that there is an increase in LEH expression from the Pre-Latte to the
Latte periods in both subadults and adults and that the Latte population was exposed to
significantly higher amounts of physiological stress than the Pre-Latte. The raw
frequency data show that juveniles have a higher frequency of hypoplastic teeth than
the adults in each time period. In populations with abrasive diets this finding could be
attributed to high rates of attritional wear, however, skeletal populations in the Marianas
display minimal wear until approximately age 40 (Leigh, 1929; Stodder, 1993). Further,
LEH data was not collected on individuals with moderate to extreme amounts of wear.
The disparate hypoplastic rates between juveniles and adults suggest that individuals
surviving to adulthood may have been healthier and less susceptible to physiological

123
stress compared to the greater frequency of individuals with LEH who died as
subadults. While not statistically significant, this finding is similar to others who found
significant differences between subadult and adult LEH frequencies in Guam (Stodder,
1997), Croatia (Slaus, 2000), and North America (Cook and Buikstra, 1979; Duray,
1996) and may indicate a correlation between linear enamel hypoplasias and life
expectancy (Slaus, 2000).
Sex Differences
As was noted in other studies in the Western Hemisphere (Lanphear, 1990; Duray,
1996; Malville, 1997; Berbesque and Doran, 2008), as well as within Guam (Douglas et
al., 1997; Pietrusewsky, 1997) sex is not found to be a statistically significant factor.
However, Guatelli-Steinberg and Lukacs (1999) found that expression of LEH is highly
variable between the sexes. Female frequencies, by individual count (13%), are fairly
low in the Pre-Latte and increase dramatically to 49% in the Latte, whereas male LEH
frequencies increase by a mere 5%. Sex-specific differences were also noted in LEH
occurrence, by tooth count, in the Leo Palace and Hyatt Latte dentition, also located in
Tumon Bay (Douglas and Ikehara, 1992; Trembly, 1999). The largest discrepancy is
seen in the Leo Palace sample where the 38% of the male dentition display LEH, while
only 5% of the female dentition displays LEH. Likewise, in the Hyatt sample, the males
(10.2%) display LEH almost twice as frequently as the female dentition (6.2%).
However, the findings of the Leo Palace (Douglas and Ikehara, 1992) and Hyatt
(Trembly, 1999) sites are more in line with the Pre-Latte sample of the current study,
with females having smaller rates of LEH than men. In the Apurguan study, however,
males have smaller LEH frequencies than females (Pietrusewsky et al., 1997;

124
Pietrusewsky et al., 2003) and do not explain the increase of female LEH in the Latte
population of the current study.
The greater frequency of LEH in the female Latte populations is in contrast to the
findings from the Leo Palace and Hyatt hotel sites, which show more developmental
disturbances in men in comparison to women (Douglas and Ikehara, 1992; Trembly,
1999). While, the sex differences in the Naton Beach sample were statistically
significant, the drastic switch in female susceptibility to physiological stress between the
time periods may indicate a cultural change in women’s access to resources over time.
Studies analyzing division of labor in hunter-gathering and farming populations have
shown significant dietary differences between the sexes (Hill and Hurtado, 1989; Walker
and Hewlett, 1990). Pre-Latte women may have had better access to nutritionally rich
resources. This trend appears to have shifted in the Latte period, as the LEH
frequencies in women greatly exceed those of men. Thus, men in the Latte likely had
better access to resource thus making them less susceptible to physiological stress.
Time Period Differences
Overall, the average of both the Pre-Latte and Latte tooth count frequencies in the
current study (1.9% and 7.7%, respectively) are much lower than all other LEH tooth
count frequencies reported for Guam, which range from 13% in the Hyatt sample
(Trembly, 1999) to 22% in the Leo Palace sample (Douglas and Ikehara, 1992). When
looking solely at the incisors and canines, an even higher frequency of LEH is observed
in Guam. Pietrusewsky and colleagues (1997) surveyed LEH expression in incisors
and canines from six prehistoric Chamorro populations in Guam and report a combined
tooth count frequency of 31%, however, this number combines both Pre-Latte and Latte
groups. The low frequency of the current study’s findings may be exacerbated by the

125
inclusion of all observable teeth in the dental arcade, including third molars.
Additionally, the Pre-Latte cultural practice of labial abrasion (Parr and Walth, 2011),
which is most prominent in the anterior dentition (Figure 5-3), precludes examination of
LEH in a large number of Pre-Latte individuals.
The current study shows an increase of LEH expression over time from 17% to
45% between the Pre-Latte and Latte individuals and is supported by Pearson Chi-
Square test, which demonstrates statistically significant differences in LEH expression
between the time periods. This finding suggests that the Latte population were more
susceptible to external stressors than the Pre-Latte. Analysis of infectious disease
demonstrates that the Latte population also had a greater degree of infectious disease
(Walth, pers. comm.).
Hanson and Butler (1977) report that treponemal infection and non-specific
periostitis are the most common infectious diseases in the Marianas Islands during the
Latte period. In the current sample, evidence of periostitis, endemic yaws, and leprosy
were observed, the majority of which occurred almost entirely in the Latte population
(Walth, pers. comm.). Non-specific periostitis is the most common infectious disease
occurring in primarily the Latte sample and seven Latte individuals displayed periosteal
inflammation and infection indicative of yaws (Walth, pers. comm.). Yaws was also
found in 19% of the adjacent Gongna-Gun Beach population (Rothschild and
Heathcote, 1993) and 20% of the Hyatt Hotel individuals (Trembly, 1999), both
relegated to the Latte period and also located Tumon Bay. The Apurguan sample of
Agana Bay, just south of Tumon Bay, has a much lower incidence, 9%, of treponemal
infection (Pietrusewsky et al., 1997). A survey of remains from Guam, Saipan, and

126
Tinan, housed at the Bishop Museum in Honoulu, reported very high rates of
treponemal infection, 77%, (Suzuki, 1986). However, these remains were collected in
the early 1900s, are not well provenienced, and may have been retained specifically for
interesting anatomical or pathological variants. None of the individuals from the Pre-
Latte sample were observed with yaws-like infections (Walth, pers. comm.). Leprosy
was observed at a lesser extent in both periods, with only one individual from the Pre-
Latte displaying lesions that may have been due to Mycobacterium leprae infection and
two individuals from the Latte display lesions characteristic of leprosy (Walth, pers.
comm.). These findings suggest that the Pre-Latte individuals were much healthier
overall with decreased rates of infectious disease as well as linear enamel hypoplasias,
than their Latte counterparts.
Carious lesions, on the other hand, reveal an opposite trend and are more
prominent in the Pre-Latte period and decrease in frequency in the Latte period. This
finding is opposite of what had been expected and is likely due to the cultural practice of
betel-nut chewing of the Latte adults, which has cariostatic properties. Additionally, the
Latte were dependent on other starchy crops, such as rice, taro, and yams, which may
not be as cariogenic as maize (see Chapter 6 for more detailed discussion).
The current study found a significant increase of LEH frequencies over time, thus,
the null hypothesis of no significant difference in the frequency of linear enamel
hypoplasias between the two time periods can be rejected. It can be inferred that an
increase of LEH over time is correlated with the intensification of agriculture
concomitant with reduced nutritional values, population increase, and subsequent
increase in infectious disease. These findings are consistent with studies, showing a

127
positive correlation between an increase of hypoplastic defects with a shift in
subsistence economies and agricultural intensification. This trend is best illustrated in
the Dickinson Mounds burial complex of Illinois where Goodman and colleagues (1980;
1984) documented an increase in LEH from the semi-sedentary Late Woodland hunter-
gatherers (45%), to the Mississippian Acculturated Late Woodland transitional
population (60%), and Middle Mississippian population (80%) with a large sedentary
population reliant on intensified maize agriculture. Changing rates of LEH over time
associated with dietary transitions were also found in North America with the adoption of
a primarily maize subsistence economy (Sciulli, 1977; Sciulli, 1978; Cook, 1984; Larsen,
1995); as well as in the late Paleolithic to Mesolithic shift to early agriculture in the
Levant (Smith et al., 1984); intensified agriculture in Ecuador (Ubelaker, 1984);
improved food processing technologies associated with intensified agriculture in India
(Lukacs, 1992); shift to protein deficient and carbohydrate rich agricultural diet in the
Early Bronze Age of Italy (Cucina, 2002); transition to early agriculture in the Nile Valley
(Starling and Stock, 2007); and subsistence shifts directed by environmental change in
Jomon foragers (Temple, 2007).
Broader Implications
An increased prevalence of linear enamel hypoplasias between the Pre-Latte and
Latte periods suggest a period of higher levels of physiological stress in the Latte
period. Large-scale environmental oscillations occurred during the Pre-Latte/Latte
transition which corresponds to the shift from the Medieval Warm Period (AD 800 to AD
1350) to the Little Ice Age (AD 1350 to AD 1900), also known as the ‘AD 1300 Event’,
where rapid cooling temperatures, decline in sea levels, and increased storminess lead
to greater climatic variability (Bridgman, 1983; Hanson, 1991; Nunn and Britton, 2001;

128
Nunn, 2007; Nunn et al., 2007; Hunter-Anderson, 2010). During the Little Ice Age,
rainfall in the Western Pacific was more erratic with more frequent and prolonged
droughts (Nunn, 2007). Nunn (2007:1) describes the AD 1300 Event as the ‘most rapid
period of climate change to have occurred within the past several millennia’ and thus,
would have been associated with societal disruption, subsistence change, and
movement to inland habitation areas throughout the Pacific Basin (Nunn, 2000; Nunn
and Britton, 2001; Nunn, 2007).
Archaeological studies have demonstrated the predicted subsistence change with
differential exploitation of marine resources and increased reliance on agriculture. A
shift from bivalve to gastropod consumption, coupled with a shift from pelagic to
reef/lagoon fish exploitation accompanied the Pre-Latte/Latte transition as a result of
ecosystem and sea level changes (Leidemann, 1980; Graves and Moore, 1985;
Hanson, 1991; Amesbury et al., 1996; Ambrose, 1993; Ambrose et al., 1997; Amesbury,
2007). Additionally, reduction of fishing gear (Butler, 1995) and increased diversification
of artifacts associated with farming (Moore, 2005) between the Pre-Latte and Latte
period indicates that there was a decreased emphasis on fishing and an increased
reliance on agricultural crops. Further evidence of agricultural intensification include
technological advances in food preparation techniques with the advent of the lusong,
the Chamorro stone mortar, used to process rice, taro, and yams (Moore, 2005), and
production of thickened pottery for cooking (Loy, 2002; Moore, 2002). The
archaeological data is supported by stable isotope analysis, which demonstrates that
marine resources account for only 30% of the Latte diet with most of the sustenance
derived from terrestrial resources, such as rice, root crops, and vegetables (Hanson,

129
1991; Ambrose et al., 1997), adding further evidence of the importance of agricultural
crops as a mainstay in the Latte period diet. Thus, the cultural and dietary shifts
between the Pre-Latte to Latte period were likely brought on by climatic variation during
the AD 1300 Event, which resulted in a transition from mixed horticultural subsistence to
an increased reliance on agriculture and subsequent a population growth, as well as a
decreased reliance on marine resources, specifically pelagic fish.
The above evidence points to a greater prevalence of developmental instabilities
and infectious disease in the Latte period coupled with climate change and agricultural
intensification, and subsequent malnutrition. Similar studies in the prehistoric North
American Southwest (Stodder et al., 2002) and Japan (Temple, 2007), as well as in
mid-1900s China (Zhou and Corruccini, 1998), have also shown a correlation between
seasonal resource depletion and systemic stress. Specifically, Temple’s analysis
(2007) of carious lesions in the Middle to Late Jomon time periods of Japan suggests:
The presence of a dietary shift after a significant climate change follow the model of culturally induced stress of Goodman and Armelagos (1989), where behavioral decision in response to environmental constraint often carry biological consequences (p. 1043).
The current study predicts a similar scenario in Guam, where environmental
constraints due to climatic variability and instability, lead to dietary transitions and thus
greater levels of stress as evidenced by increased linear enamel hypoplasias over time.
Conclusions
There were no significant differences in the rates of juvenile and adult LEH;
however, in terms of raw frequency data, juveniles displayed higher hypoplastic defects
than adults. This may suggest a correlation between life expectancy and physiological
disruptions. Male and female differences in LEH expression were not significant within

130
each time period, but looking at the raw frequency data provides interesting insight into
differential access to resources. In the Pre-Latte sample, the females were exposed to
less physiological stressors and as such, may have had better access to nutrition-rich
foods compared to the men. However, this trend shifted significantly in the Latte where
women became much more highly susceptible to physiological stressors than men.
Differences in gender status does not explain the gap between male and female LEH
rates, as the Latte peoples followed matrilineal kinship system where the women were
powerful and respected in all aspects of society (Souder, 1992).
Thus, division of labor, with women collecting and gathering versus males hunting
or fishing may be the reason for sex discrepancies in frequencies of physiological
stress. The Pre-Latte peoples were horticulturalists and supplemented food with marine
resources. Women may have gathered food as well as collected bivalves for
consumption along the coast, while the men explored beyond the reefs to collect the
larger pelagic fish. With the intensification of agriculture in the Latte period, more time
would have been spent tending to and cultivating crops. Ethnohistoric accounts
suggest that job was delegated to women (Driver, 1993), who likely had less time to
collect coastal marine foods. Men are reported as being the primary fishermen (Driver,
1993; Russell, 1998) who abandoned dangerous exploitation of the pelagic fish for the
more easily obtainable reef fish and gastropods. In this scenario, men would have had
greater access to protein content than women, and making them less susceptible to
physiological disruptions.
The Pre-Latte people, overall, appear to have been healthier than the Latte
individuals with less frequency of LEH and infectious disease. The increased

131
susceptibility of the Latte population to linear enamel hypoplasias is associated with
seasonal variability, such as typhoons, droughts, and increased aridity, which would
have damaged crops and surrounding reef systems resulting in a reduction of
agricultural productivity, depletion and /or destruction of staple foods (Stodder, 1997;
Hunter-Anderson, 2010), and spread of endemic disease (Pietrusewsky et al., 1997;
Stodder, 1997). Such climatic variability and decreased access to nutritional resources
would have been detrimental to the overall health of the Latte peoples, especially in
developing children when the physiological disturbances that affect enamel formation
are taking place.
The current study provides a diachronic analysis of the Chamorro health profile.
Previous studies have described the Latte people as having higher susceptibility to
linear enamel hypoplasias compared to Hawaiians as a result of nutritional deficiencies
and infectious disease (Pietrusewsky et al., 1997). This study validates that claim and
expands on the current knowledge of prehistoric health in the Chamorro by
demonstrating that the earlier Pre-Latte inhabitants were healthier than the later Latte
population as a result of climatic instability and subsequent dietary transitions.
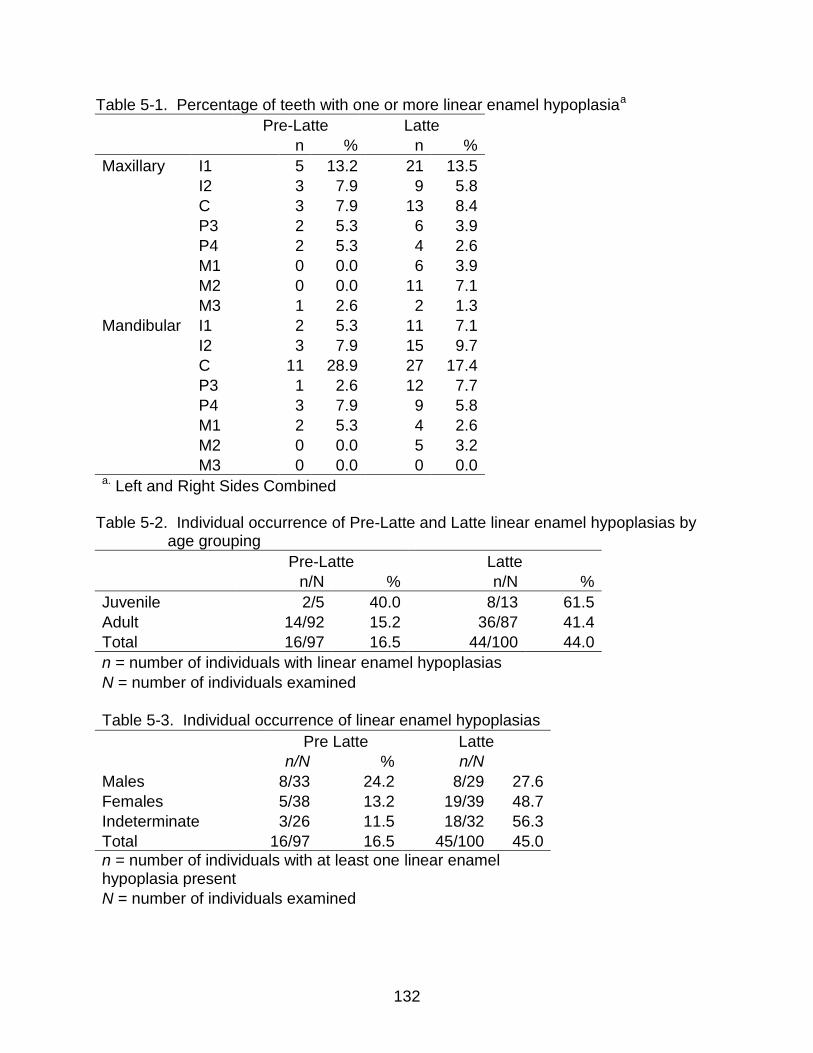
132
Table 5-1. Percentage of teeth with one or more linear enamel hypoplasiaa
Pre-Latte Latte
n % n %
Maxillary I1 5 13.2 21 13.5
I2 3 7.9 9 5.8
C 3 7.9 13 8.4
P3 2 5.3 6 3.9
P4 2 5.3 4 2.6
M1 0 0.0 6 3.9
M2 0 0.0 11 7.1
M3 1 2.6 2 1.3
Mandibular I1 2 5.3 11 7.1
I2 3 7.9 15 9.7
C 11 28.9 27 17.4
P3 1 2.6 12 7.7
P4 3 7.9 9 5.8
M1 2 5.3 4 2.6
M2 0 0.0 5 3.2
M3 0 0.0 0 0.0 a. Left and Right Sides Combined
Table 5-2. Individual occurrence of Pre-Latte and Latte linear enamel hypoplasias by
age grouping
Pre-Latte Latte
n/N % n/N %
Juvenile 2/5 40.0 8/13 61.5
Adult 14/92 15.2 36/87 41.4
Total 16/97 16.5 44/100 44.0
n = number of individuals with linear enamel hypoplasias
N = number of individuals examined
Table 5-3. Individual occurrence of linear enamel hypoplasias
Pre Latte Latte
n/N % n/N %
Males 8/33 24.2 8/29 27.6
Females 5/38 13.2 19/39 48.7
Indeterminate 3/26 11.5 18/32 56.3
Total 16/97 16.5 45/100 45.0
n = number of individuals with at least one linear enamel hypoplasia present
N = number of individuals examined

133
Table 5-4. Tooth count of linear enamel hypoplasias
Pre Latte Latte
n/N % n/N %
Males 14/634 2.2 33/450 7.3
Females 13/825 1.6 54/670 8.1
Indeterminate 11/466 4.1 68/573 11.9
Total 38/1925 2.0 155/1693 9.2
n = number of teeth with at least one linear enamel hypoplasia
N = number of teeth examined

134
Table 5-5. LEH frequencies in comparative populations
Tooth Count Individual Count
Study Sample Subsample Time Period Male %
Female %
Male %
Female % References
Naton Beach Pre-Latte Pre-Latte 2.2 1.6 23.5 13.2 Current Study
San Vitores Road
Fujita Drainfield Pre-Latte 19.5
- -
Bath, 1986; Pietrusewsky, 1986
Naton Beach Latte Latte 7.3 8.1 28.6 48.7 Current Study
Apurguan N/A Latte 20.6 25.1 - - Douglas et al., 1997; Pietrusewsky et al., 2003
Hyatt Hotel N/A Latte 10.2 6.2 - - Trembly, 1999
Leo Palace N/A Latte 38.0 5.0 - - Douglas and Ikehara, 1992
Fiesta Resort N/A Latte - - 11.8 - Defant et al., 2008

135
Table 5-6. Age differences of LEH using a Pearson Chi-Square Test
Value Df Asymp. Sig (2-sided)
Age within Pre-Latte 15.547 3 0.001*
Age within Latte 12.089 4 0.017*
* Indicates statistically significant differences at the 0.05 alpha level
Table 5-7. Sex differences of LEH using a Pearson Chi-Square Test
Value Df Asymp. Sig
(2-sided) Exact Sig. (2-sided)
Exact Sig. (1-sided)
Age within Pre-Latte 1.451 1 0.228 0.357 0.185
Age within Latte 3.102 1 0.078 0.087 0.065 a. Indeterminate sex not included
+ Indicates no statistical relationship at the 0.05 alpha level Table 5-8. Pre-Latte and Latte differences in LEH using a Pearson Chi-Square Test
Value Df Asymp. Sig (2-sided)
Exact Sig. (2-sided)
Exact Sig. (1-sided)
Between Time Periods 18.716 1 0.000 0.000 0.000
LMAX I1 9.785 2 0.008 - -
RMAX I1 8.889 3 0.031 - -
LMAND C 10.894 3 0.012 - -
RMAND C 8.444 3 0.038 - -
* Indicates statistically significant differences at the 0.05 alpha level

136
Figure 5-1. Single linear enamel hypoplasia in the mandibular lateral incisor, canine,
and third premolar (Photo by author)
Figure 5-2. Multiple linear enamel hypoplasias in a single tooth (Photo by author)

137
Figure 5-3. Labial abrasion in a Pre-Latte Period individual (Photo by author)
Figure 5-4. Betel-nut staining in a Latte Period individual (Photo by author)

138
Figure 5-5. Dental incising in a Latte Period Individual (Photo by author)
Figure 5-6. Frequency of linear enamel hypoplasias by tooth type (right and left sides
combined)
0
5
10
15
20
25
30
I1 I2 C P3 P4 M1 M2 M3 I1 I2 C P3 P4 M1 M2 M3
Maxillary38 38 38 38 38 38Mandibular38 38 38 38 38 38 38
LE
H F
req
ue
ncy
Tooth Type
Pre-Latte
Latte

139
CHAPTER 6 CARIOUS LESIONS
Patterns of dental disease have long been studied by anthropologists interested in
the health, diet, and lifestyle of prehistoric populations. Aristotle was the first to record
the correlation between diet and dental disease in antiquity when he observed that
sweet figs adhering to the teeth caused dental caries (Powell, 1985). Carious lesions,
in particular, have been of interest because they are a direct indication of an infectious
disease process and are easily observable in the dentition, which is more likely to
survive in an archaeological setting than other indicators of skeletal infection. As such,
carious lesions are the most common dental disease (Bunting, 1933) and are the most
frequently reported dental pathology found in archaeological populations (Roberts and
Manchester, 1995).
The current study examines carious lesion rates between the horticultural Pre-
Latte (3500 BP to 1000 CE) and early agricultural Latte (1000 to 1521 CE) populations
in Guam to investigate what effect intensified agriculture and increased reliance on
starchy foods such as rice, taro, and yams have on the oral health status of the
population. The excavation of the Naton Beach Burial Complex allows for the first
diachronic assessment health in prehistoric Chamorro population using large sample
sizes. Decreased health, as evidenced by increased carious lesions, is predicted
concomitant with agricultural intensification. This study tests the null hypothesis that
there is no significant difference in the frequency of carious lesions between the Pre-
Latte and Latte time periods.

140
Formation of Carious Lesions
Dental caries can take on many different forms such as pit and fissure caries,
smooth surface caries, root caries, and deep dentin caries (Newbrun, 1982). Larsen
(1997:65) describes dental caries as ‘a disease process characterized by the focal
demineralization of dental hard tissues by organic acids produced by bacterial
fermentation of dietary carbohydrates, especially sugars.’ The pH balance of plaque is
of particular importance as it varies based on amounts of protein and carbohydrates in
the diet. Lactic acid is produced when plaque bacteria metabolize carbohydrates, while
alkaline waste products are produced by metabolization of protein. Thus, the formation
of a carious lesion occurs when periods of acidity outweigh periods of alkalinity and
mineral destruction of the enamel occurs (Hillson, 1979). The epidemiological literature
has shown that sugar is one of the major factors of carious lesions (Stoppelaar et al.,
1970; Newbrun, 1982) and that foods with high amounts of dietary sucrose, which
correlate to acidic periods in plaque, are more likely to result in carious lesion formation
(Hillson, 1979; Larsen, 1983).
Carious Lesions and Agricultural Intensification
Carious lesion etiology is not fully understood, however, many factors have been
associated with their development, including genetic predisposition, salivary flow and
chemistry, tooth size and morphology, diet, fluoride component in drinking water
(Newbrun, 1982; Rowe, 1982), as well as the physical properties of food and the way it
is prepared (Larsen, 1997).
Turner (1979) reports carious lesion frequencies drawn from populations with
different subsistence patterns from around the world, and suggests that caries
prevalence increase from hunter-gatherer societies to agricultural ones. He further

141
proposes frequency ranges, in terms of tooth count frequencies, for different
subsistence patterns: hunter-gatherers: 0.0% to 5.3%, mixed economies: 0.44% to
10.3%, and agricultural: 2.3% to 26.9%. These values are often used in studies to
assist in reconstructing dietary subsistence patterns (e.g. Delgado-Darias et al., 2005;
Bernal et al., 2007). Cohen and Armelagos’ (1984) benchmark book, Paleopathology at
the Origins of Agriculture, validated Turner’s claim with the consensus that carious
lesions increase in frequency in the shift from hunter-gatherer populations to agricultural
ones. An increase in carious lesion prevalence and agricultural intensification, has been
repeatedly demonstrated world-wide, with various food crops such as maize in North
America (Larsen, 1981; Cook, 1984; Larsen, 1984; Larsen et al.,1991) and Ecuador
(Ubelaker, 1980; Ubelaker, 1984; Ubelaker and Newson, 2002); wheat and barley in
Egypt (Hillson, 1979), the Levant (Smith et al., 1984), and Pakistan (Lukacs, 1992);
millet in North Africa (Martin et al., 1984) and Northern China (Pechenkina et al., 2002);
and rice in Southeast Asia (Krigbaum, 2007) and Japan (Temple, 2007; Temple and
Larsen, 2007).
However, some recent studies have shown no significant relationship between
carious lesion frequency and intensification of agriculture (Oxenham et al., 2000; Tayles
et al., 2000; Pietrusewsky and Douglas, 2002; Eshad et al., 2006; Domett and Tayles,
2007; Douglas and Pietrusewsky, 2007; Lanfranco and Eggers, 2010). Specifically,
trends in Southeast Asia show either homogeneity in carious lesion rate or decline in
caries associated with the intensification of agriculture and reveal little evidence for
decreased health status over time (Oxenham et al., 2000; Tayles et al., 2000;
Pietrusewsky and Douglas, 2002; Oxenham et al., 2006; Douglas and Pietrusewsky,

142
2007; Domett and Tayles, 2007). This geographical trend has been attributed to the
transition to rice-based agriculture, which was not analyzed in Cohen and Armelagos’
(1984) volume. However, other studies have found an increase of carious lesions with
agricultural intensification of rice in present-day Malaysia (Krigbaum, 2007) and Japan
(Temple and Larsen, 2007). Temple and Larsen (2007) suggest, in support of their
findings, that rice, while not as cariogenic as other starches, has more cariogenic
properties than originally proposed by Tayles and colleagues (2000). In a follow-up
article, Tayles and colleagues (2009:163) suggest that the variability in rice cariogenicity
is the result of ‘structure, the method of processing, and the extent to which [it is]
gelatinised during cooking.’ Thus, the decreased frequency of carious lesions in some
groups reliant on rice may be due to minimal processing of the starchy crop (Talyes et
al., 2009).
The above studies depict the difficulty of interpreting carious lesions within the
realm of dietary transitions, specifically in regards to rice agriculture. Thus, the
development of carious lesions within an individual or population must be understood as
a process that is multifactorial in nature and cannot be attributed to a single variable.
As such, analysis of carious lesions must be undertaken with a careful examination into
diet, food processing, and other cultural factors that may be in effect.
Culture History of Guam
The shift between the Pre-Latte (3500 BP to 1000 CE) and the Latte (1000 to 1521
CE) time periods in Guam involves changes in population size, diet, and food
procurement/preparation strategies (Hunter-Anderson and Butler, 1991; Moore, 2005;
Amesbury, 2007). The Pre-Latte population is a small semi-nomadic, foraging
population that subsisted on bivalves, shellfish, and reef and pelagic fishing (Amesbery

143
et al., 1991; McGovern and Wilson, 1996). Horticultural practices were likely brought to
Guam by its earliest inhabitants from island Southeast Asia (Bellwood, 1978). Data
from paleoenvironmental sediment cores suggest small-scale gardening and resource
collecting by initial settlers (Athens and Ward, 2004), and analyses of pollen and starch
residues on the interior of Pre-Latte pottery found evidence of taro, cabbage tree, fish,
and shellfish (Loy, 2001a,b; Loy, 2002; Loy and Crowther, 2002).
Charcoal becomes increasingly present in paleoenvironmental sediment cores
around 1,800 BP, which corresponds to the Pre-Latte/Latte transition, and is suggestive
of intensified land use (Athens and Ward, 2004). Latte period archaeology has
identified a wide array of food processing artifacts, such as stone mortars and
pounders, scrapers, knives, blades, and adzes, which are indicative of a more
sedentary population with an increased reliance on agriculture (Hunter-Anderson and
Butler, 1991; Moore, 2005). Starch grain residues on Latte period potsherds identified
cabbage tree, rice, sugarcane, and a taro (Loy, 2001a,b; Loy, 2002; Loy and Crowther,
2002). Taro was identified more than any other plant, suggesting a preference for this
introduced tuber. Pollen and phytolith analyses have also found evidence of betel palm,
breadfruit, coconut, bananas, and pandanus (Hunter-Anderson and Butler, 1991;
Cummings and Puseman, 1998; Dixon et al., 1999; Pearsall and Collins, 2000; Ward,
2000; Cummings, 2002; Athens and Ward, 2004).
Stable isotope data reveal that the majority of the Latte diet was composed
primarily of terrestrial C3 resources, such as rice, root crops, and vegetables and that
marine foods are merely a supplement, comprising approximately 30% of the diet
(Hanson, 1991; Ambrose et al., 1997). Higher 13 N values are suggestive of

144
preferential exploitation of reef and/or lagoon fish over pelagic resources (Hanson,
1991; Ambrose, 1997). Ethnohistoric and archaeological evidence points to exploitation
of some larger pelagic fish such as tuna, dolphin, and marlin (Driver, 1993; Freycinet,
1996; Amesbury and Hunter-Anderson, 2008), however, deep-sea fishing is more
dangerous and thus not used as a reliable subsistence base (Russell, 1998). Thus, the
favored reliance of marine-based foods was abandoned for terrestrial resources and
agricultural crops in the Latte period. The stable isotope findings are further supported
by archaeological units demonstrating a greater proportion of fishing gear and fishing-
related debris in the Pre-Latte period in comparison with the Latte Period (Butler, 1995).
Additionally, intensification of agriculture, in the Latte period, would have permitted an
expansion in population size. Population growth is evident with more varied habitation
areas ranging from the preferred coastal locale to more marginal and inland
environments, such as the interior uplands (Hunter-Anderson and Butler, 2001).
Underwood (1973) and Hezel (1982) suggest that the prehistoric Latte population, at its
largest, ranged between 30,000 and 40,000.
The cultural modifications associated with intensification of agriculture, such as
food processing, are likely to be accompanied by biological changes, such as an
increase in stress levels associated with population growth, limited access to resources,
malnutrition, and increased prevalence of disease. This study is restricted to the
dentition due to the poor quality of skeletal preservation that is typical in island and
coastal environments such as Guam (Hanson and Butler, 1997). However, analyses
conducted by Cherie Walth, will be utilized to augment data from the current study to
construct a health profile of the Chamorro population.

145
Materials and Methods
From 2006 to 2008, 376 prehistoric Chamorro burials were excavated during an
archaeological mitigation project for the renovation of the Guam Aurora Villas & Spa in
the Naton Beach Site on the northern end of Tumon Bay, Guam (DeFant, 2008). Of
these, approximately 177 are associated with the Pre-Latte Period and 190 belong to
the Latte Period. The determination of the burials’ age and cultural affiliation was based
on the stratigraphic location, associated artifacts, and radiocarbon dates (DeFant,
2008). Dating to roughly 2,500 BP, the Naton Beach Pre-Latte sample represents some
of the earliest settlers in Guam and is the largest Pre-Latte mortuary sample discovered
to date (DeFant, 2008). There are no radiocarbon dates for the Latte sample; however,
archaeological materials associated the remains are consistent with those relegated to
the Latte period, between AD 1000 to 1521.
Carious lesions were analyzed macroscopically on all fully formed permanent
teeth, by individual and by tooth count methods, as recommended by Lukacs (1989).
LEH by individual count was scored as present or absent of carious lesions. Carious
lesions, by tooth count, were scored using an ordinal scale: 0 = absence of carious
lesions; 1 = presence of carious lesions in a single tooth; 2 = presence of one or more
caries in a single tooth.
The Pre-Latte sample consists of 99 individuals, while 108 individuals from the
Latte period were analyzed (Table 6-1). A total of 3,666 teeth were analyzed: 1,930
from the Pre-Latte and 1,736 from the Latte (Table 6-2). Females1 are represented
1 Demographic data, as well as postcranial pathological conditions, were recorded by Cherie Walth. Her
data, in conjunction with the data from the current study, were collected as part of a larger study on the Naton Beach Burial Complex, which is currently being compiled for submission to the Guam Historic Preservation Office.

146
more often than males in both samples; however, sex is distributed fairly evenly in the
Pre-Latte sample (females: n = 38; males: n = 33). The distribution within the Latte
sample is more heterogeneous (females: n = 37; males: n = 29). Age groups were also
analyzed separately, however, the children and juvenile samples are very small,
particularly in the Pre-Latte period (Table 6-3).
Raw frequency data is reported by time period, sex, and age. Inflated carious
rates are often reported in the bioarchaeological literature when tooth class is not taken
into account, as the molars, followed by the premolars, are more susceptible to caries
and are also the best preserved in archaeological conditions and less vulnerable to
postmortem loss (Hillson, 1996). Therefore, carious lesions were also analyzed in
terms of tooth position.
Multivariate statistical testing was performed to determine statistical significance of
differences in carious lesion frequency between groups. A Pearson Chi-Square test
was used to test for significant differences between the time periods, the sexes, and the
age groups.
Results
Tooth Position
As expected, the molars have a greater frequency of carious lesions than rest of
the dentition, with the mandibular second molar displaying the highest frequency of
carious lesions in both groups (Pre-Latte: n = 37; 29.6%, Latte: n = 14; 10.2%) (Table 6-
4). The mandibular incisors and canines of the Pre-Latte had higher than expected
carious lesions prevalence (12% or greater), especially when compared to the Latte,
which had relatively no carious lesions in the mandibular incisors (Mand I1: n = 0; 0%;
Mand I2: n = 1; 0.9%) and only 3.6% (n = 4) in the mandibular canine.

147
When anterior teeth (incisors and canines) and posterior teeth (premolars and
molars) are combined, the data show that the posterior teeth have greater incidence of
carious lesions (Pre-Latte: n = 168; 13.8%; Latte: n = 60; 5.4%) than the anterior teeth
(Pre-Latte: n = 77; 10.7%; Latte: n = 17; 2.4%) (Table 6-5). Again, as was noted earlier,
the Pre-Latte anterior dentition has a greater frequency of carious lesions in comparison
to the Latte period.
Age Differences
Carious lesions were compared between age groups to see if the frequency
increased with age (Table 6-3). However, caution must be taken, as the sample sizes
of both children and juveniles are small. The Pre-Latte population shows an increase of
carious lesion frequencies from 40% (n = 2) in juveniles to 78.1% (n = 50) in young
adults, which then decreased to 73.9% (n = 17) in middle-aged adults. The Latte,
however, display a decrease with increased maturity with 31% (n = 4) of the juveniles
and 25% (n = 10) of the young adults displaying carious lesions; however, with
increasing senility, caries increase in frequency with 31.1% (n = 10) of the middle aged
adults affected. The disparities between the age groups are slight, as is demonstrated
with a Pearson Chi-Square test which found no significant age differences in either the
Pre-Latte (p = 0.093, at the 0.05 alpha level) or Latte the (p = 0.741, at the 0.05 alpha
level) time periods (Table 6-6).
Sex Differences
No sex specific differences are apparent in the distribution of carious lesions in
either the Pre-Latte or Latte individuals (Table 6-1). By individual count, both males and
females, of the Pre-Latte period, display high frequencies of carious lesions (n = 25;
75.8% and n = 29; 76.3%, respectively) with females being slightly more prone to

148
display carious lesions. Similarly, in the Latte period, males and females exhibit low
frequencies of carious lesions (n = 7; 24.1% and n = 11; 29.7%, respectively).
However, this may be due to sampling bias as females are more often represented than
males in both groups. When looking at carious lesion frequency by tooth count
method, males (110/644; 17.1%) have a higher frequency than females (94/858; 11%)
in the Pre-Latte period (Table 6-2). Thus, while more women display slightly more
carious lesions than men, men are more prone to have multiple infected teeth. A
Pearson Chi-Square test found no significant differences between male and female
expression of carious lesions in the Pre-Latte (p = 0.956, at a 0.05 alpha level) or the
Latte (P = 0.613, at a 0.05 alpha level) time periods (Table 6-6).
Time Period Differences
Large differences in carious lesion frequencies are apparent between the Pre-
Latte and Latte populations. Carious lesions, by individual count, were found in the
majority, 72.7% (n = 72) of the Pre-Latte population, while only 24.1% (n = 26) of the
Latte individuals displayed carious lesions (Table 6-1). In terms of tooth count, the Pre-
Latte incidence (246/1930; 12.7%) of carious lesions is triple what is seen in the Latte
period (76/1736; 4.4%) (Table 6-2). These findings are supported using a Pearson Chi-
Square test, which found significant differences (p < 0.000 at a 0.05 alpha level) in
carious lesion frequencies between the Pre-Latte and Latte time periods (Table 6-6).
Discussion
Tooth Position
At first glance, the location of carious lesion, in terms of tooth type, follows the
characteristic pattern of an increasing gradient in frequency from the anterior to
posterior dentition (Watt et al., 1997; Vodanovic et al., 2005; Han et al., 2010; Meng et

149
al., 2011). However, departure for the norm occurs in the Pre-Latte sample with
uncharacteristically high levels of caries in the anterior teeth. The expression of carious
lesions in the anterior dentition of the Pre-Latte is higher than seen the observed rates
in premolars, which are usually more prone to bacterial infection (Hillson, 1996). The
location of carious lesions on the anterior dentition typically occurs at the cemento-
enamel junction, and at times, the root surface was also involved (Figure 1). Similar
findings of high rates of carious lesions in the anterior dentition was noted in one
individual and isolated teeth from the Chelechol ra Orrak cemetery in Palau (Nelson and
Fitzpatrick, 2005) and in the Longshan-period Kangjia of Northern China. Nelson and
Fitzpatrick (2005) suggest the carious lesions may be related to betel-hut chewing;
however, this interpretation seems unlikely given the cariostatic properties of betel-nut.
No attempt was made to explain the anterior caries in Northern China (Pechenkina et
al., 2002).
This type of carious lesion is often associated with periodontal disease where the
receding alveolus exposes the root and lesions develop circumferentially around the
cemento-enamel junction (Hillson, 1996). In a survey of skeletal reports from around
the Marianas, Pietrusewsky and colleagues (1997) note a higher frequency of alveolar
resorption in two Pre-Latte samples, Matapang (44.3%) and Fujita (36.8%), whose rates
of resorption exceed that of all other Latte sites in Guam, except one.
Another factor leading to the higher rate of carious lesions in the anterior teeth is
the extreme dental crowding noted in the Pre-Latte remains (Figure 2), which due to the
fragmentary nature was not systematically analyzed. In the few individuals where
maxillary or mandibular reconstruction was possible, dental crowding was evident in the

150
Pre-Latte dentition and could increase the amount of food trapped interstitially between
the incisors and canines. Dental crowding was not observed in the Latte individuals and
thus did not likely contribute to carious defects in the anterior teeth.
Age Differences
No significant difference in carious lesion frequency was found between the age
groups in either period. The data were re-analyzed grouping the adult sub-groups (i.e.
young adult, middle-aged adult, and old adult) and comparing them to the juveniles;
likewise, adult sub-groups were analyzed separately to see if frequencies varied with
senility; however, no significant difference were found in either case (data not shown).
The raw frequency data indicates a decrease between the young adult and middle-aged
adult categories in the Pre-Latte sample, whereas in the Latte, there is an increase
between those two groups. There were no older adults in the Pre-Latte individuals and
only four available for analysis in the Latte, none of which had carious lesions. The
decrease in caries rate with age, in the Pre-Latte sample, is surprising given the
progressive nature of carious lesion development, which usually occurs at higher
frequencies in older individuals (Thylstrup and Fejerskov, 1994). Dental attrition could
explain decrease in caries with age, where the complex morphology of the occlusal
surface of the dentition is worn away, leaving less possibility for infection. The Latte
pattern, on the other hand, follows the traditional trend with increase of caries
prevalence with age. The effect of age on carious lesion formation has not been
addressed in other studies from Guam.
Sex Differences
The current study found no significant differences in caries frequencies between
males and females, in either the Pre-Latte or the Latte groups, suggesting that both

151
sexes were eating foods with similar levels of cariogenicity. Analysis of carious lesion
frequencies in Taiwan also failed to show significant differences in caries rate between
the sexes (Pietrusewsky and Tsang, 2003). Douglas and colleagues (1997), on the
other hand, found significant sex differences in carious lesion prevalence in the Latte
period Apurguan site, located south of the Naton Beach in Tumon Bay. They report a
greater frequency of carious lesions, by tooth count, in young adult males in comparison
to young adult females and suggest a differential access to sweet or sticky foods.
However, when combining the age groups, males and females of the Apurguan sample
show similar caries frequencies (males: 2.8%, females: 2.1%), which are lower than that
found in the current study (males: 3.5%, females: 6.6%).
While not statistically significant, the Latte females of the Naton Beach sample
have a higher prevalence of carious lesions than males. This finding is more in line with
other studies that have shown sex-specific with higher caries rates in females (Larsen et
al., 1991; Kelley et al., 1991; Lukacs, 1996; Temple and Larsen, 2007). These
difference sex-specific differences have been attributed to differential access to dietary
resources between the sexes due to division of labor. It is likely that this may also be
the case in the current sample and is plausible given the linear enamel hypoplasia
findings, which show greater incidence of hypoplasias in females.
Time Period Differences
The Naton Beach sample displays significant differences in carious lesion
frequencies between the two time periods, with the Pre-Latte displaying higher
prevalence of caries than the Latte period. In their survey of carious lesions frequencies
in various sites in Guam, Pietrusewsky and colleagues (1997) hint at the possibility of
temporal changes in health and disease between the periods with 14.3% of the Pre-

152
Latte Fujita sample displaying carious lesions, which is higher than the Latte groups that
range from 1.3% to 12.3% (Table 6-7). However, Pietrusewsky and colleagues (1997)
resist from making any definitive conclusions until larger sample sizes became
available.
The current study provides a large Pre-Latte sample that can be used for
comparative purposes. The tooth count frequency of carious lesions in the Pre-Latte
Naton Beach sample is 12.7%, which is comparable to the Fujita frequency. Likewise,
the Latte prevalence of 4.4% falls in line with the other Latte samples whose caries
rates range from 1.3% in the Leo Palace Sample to 12.3% in the Right-of-Way sample
(see Table 6-x for references). Overall, the caries prevalence in the Pre-Latte is higher
than the Latte when comparing the various sites across Guam.
A significant difference in carious lesion frequency of 8% was found between the
Pre-Latte and Latte groups; thus, the null hypothesis of no differences in carious lesion
frequencies between time periods can be rejected. This study confirms the hypothesis
of different caries rates between the Pre-Latte and Latte periods; however, the cause of
this disparity needs to be evaluated in concert with the varying dietary and cultural
practices between the populations.
The effects of diet
The above findings are contrary to the expected results of an increase of carious
lesions associated with intensification of agriculture, as was found in many studies
throughout the world (Hillson, 1979; Ubelaker, 1980; Larsen, 1981; Cook, 1984; Larsen,
1984; Martin et al., 1984; Smith et al., 1984; Ubelaker, 1984; Larsen et al.,1991;
Lukacs, 1992; Pechenkina et al., 2002; Ubelaker and Newson, 2002; Pechenkina et al.,
2002; Krigbaum, 2007; Temple, 2007; Temple and Larsen, 2007). In the current study,

153
the opposite trend is demonstrated, thus a re-analysis of the dietary differences
between the groups is warranted.
Pre-Latte: The Pre-Latte population displays much higher rates of carious lesions
than would be expected for a pre-agricultural society, and dental crowding alone may
explain an increase in bacterial infection; however, other causative factors will also be
explored. Turner (1979) proposed increasing range of carious lesion prevalence from
hunter-gatherers (0.0% to 5.3%) to populations with mixed economies (0.44% to 10.3%)
and agricultural populations (2.3% to 26.9%). The combined male and female
frequency of Pre-Latte sample, 13%, falls well within the range of agriculturalists;
however, the rates are unexpectedly high for a horticultural population. Pollen and
starch analyses found evidence of taro on Pre-Latte pottery, which has cariogenic
properties. Yet, the high prevalence of carious lesions in the Pre-Latte would not be
expected unless taro was being intensively cultivated and was relied on as a staple crop
on a broader scale. Charcoal presence, dating to the Pre-Latte in paleoenvironmental
cores is indicative of repeated burning of forest patches (Athens and Ward, 2004). This
practice may have been used as a method to clear land for small-scale gardening.
Thus, it is possible that the Pre-Latte were more reliant on taro than had been
previously expected and may have had participated more in incipient agricultural
activities than has been observed through the archaeological record.
An alternative explanation for the high rates of carious lesions may be sugarcane
consumption, as a positive correlation between long-term sugarcane chewing and
carious lesions has been established (Frencken et al., 1968). Phytolyth analysis of one
Latte period pottery sherd (Hunter-Anderson and Moore, 2002) and stable isotope

154
analyses (Ambrose et al., 1997) indicate that sugarcane may have been an important
part of the diet in the Latte population. While presence of sugarcane in the Pre-Latte
period has not been supported through paleoenvironment sediment cores (Athens and
Ward, 2004) or other archaeological evidence, it is possible that it may have been
brought to Guam by the early colonizers (Moore, 2002), as it was domesticated in the
southwest Pacific prior to expansion into Remote Oceania (Daniels and Daniels, 1993;
Grivet et al., 2004).
Latte: Guam is unique in that it is the only tropical Pacific island to have cultivated
rice prior to European colonization (Hunter-Anderson et al., 1995). Rice impressions
were found on pottery sherds from Tumon Bay that were radiocarbon dated to the Latte
period (640 ± 50 BP; calibrated date range, 2σ: AD 1284-1425) (Moore et al., 1993) and
subsequently, rice-impressed Latte period pottery sherds were also found in other areas
around Guam (Moore, 1994; Moore and Hunter-Anderson, 1994). Presence of rice in
the Latte Period is further established through identification of rice through phytolith
analysis on four potsherds (Loy, 2001a).
Given the archaeological evidence for the presence of rice in the Latte period, the
low prevalence of caries is unexpected. The cariogenicity of rice in prehistoric
populations is currently being debated (e.g. Temple and Larsen, 2007; Tayles et al.,
2000; Talyes et al., 2009). Some studies have shown an increase of carious lesions
with the adoption of rice agriculture (Krigbaum, 2007; Temple and Larsen, 2007), while
others have shown the opposite (Oxenham et al., 2000; Tayles et al., 2000;
Pietrusewsky and Douglas, 2002; Oxenham et al., 2006; Douglas and Pietrusewsky,

155
2007; Domett and Tayles, 2007), and a study in living Thai children found a decrease in
caries rates in children eating a primarily rice-based diet (Kedjarune et al., 1997).
Experimental studies have also produced varied results. Rats fed diets of wheat,
corn, rice, and oats (independently) developed carious lesions, however, wheat and
corn were determined to be the most cariogenic agents (Dodds, 1960). Krasse (1985)
showed that the cariogenic potential of rice is low and Sreebny (1983) found no
correlation of rice consumption and carious lesions. Grenby (1997) and Lingstrom and
colleagues (2000) conclude that the cariogenicity of food starches can be greatly altered
by cooking and food processing, and that a mix of processed starches and sugars are
more cariogenic than starch alone (Grenby, 1997).
Besides rice, the Latte people were known to have consumed other cariogenic
foods. Starch grain residues have identified cabbage tree, sugarcane, and taro (Loy,
2001a,b; Loy, 2002; Loy and Crowther, 2002), while pollen and phytolith analyses found
evidence of betel palm, breadfruit, coconut, bananas and pandanus (Hunter-Anderson
and Butler, 1991; Cummings and Puseman, 1998; Dixon et al., 1999; Pearsall and
Collins, 2000; Ward, 2000; Cummings, 2002; Athens and Ward, 2004). Further, yam
cultivation in the interior of Guam (Moore, 2005) and a preponderance of taro residues
on the interior of clay pots (Loy 2001a, 2001b, Low and Crowther, 2002) suggest a
preference for these tubers. These data indicate that the prehistoric diet of the Latte
period was more diversified than other populations experiencing intensification of
agricultural practices who usually relied on one staple crop.
Hunter-Anderson and colleagues (1995) suggest that rice may have been used
ceremonially as a prestige food and thus, did contribute significantly to the prehistoric

156
diet. Even if this is the case and rice was not a dietary staple, many of the other primary
foods such as sugarcane, breadfruit, bananas, yams, and taro are highly cariogenic. As
Grenby (1997) demonstrated, a diet combining starchy foods, such as rice, taro, and
yams with sugary foods, such as sugarcane, breadfruit, bananas would lead to a highly
cariogenic diet with a prevalence of dental caries. This finding lends further support to
the multifactorial nature of carious etiology that cannot be explained by diet alone.
Thus, the low prevalence of carious lesions in the Latte sample, in light of demonstrated
exploitation of highly cariogenic foods, cannot be explained by dietary choices.
The effects of betel-nut chewing
Another possibility, and the most likely explanation, for the low levels of carious
lesions in the Latte period is betel-nut chewing, which has been practiced in antiquity to
modern times throughout South Asia, Southeast Asia, and the Western Pacific
(Strickland, 2002; Zumbroich, 2007). Paleoenvironmental data from Guam have
established that betel-nut (Areca catechu) is indigenous to the Marianas and pre-dates
human settlement (Athens and Ward, 2004). In the Marianas Islands, the areca nut is
usually combined with the betel leaf (Piper betle) and slaked lime (CaCO3) (Figure 5)
(Hanson and Butler, 1997) and increases stamina, reduces hunger, and creates a
sense of euphoria when chewed (Chu, 2001, 2002). Over time, betel-nut chewing
results in a dark reddish-brown stain on the dentition (see Chapter 5, Figure 5-4).
The cariostatic effects of betel-nut have been demonstrated in the epidemiological
literature, where the high alkalinity of the areca nut neutralizes acid formation in the
mouth, and thus creates an environment unsuitable for dental caries (Chandra and
Desai, 1970; Howden, 1984; Moller et al., 1977; Chatrchaiwiwatana, 2006). Hanson

157
and Butler (1997:280) outline four mechanisms that may lead to cariostasis with betel
chewing:
1. Cleansing of tooth surfaces and thus removing sites of potential carious lesions 2. Appetite reduction which reduces potential intake of cariogenic foods 3. Increase of salivary flow and proteins that inhibit bacterial activity 4. Increase alkalinity of the oral cavity 5. Provides a physical barrier against the spread of cariogenic agents. Thus, the physical and chemical aspects associated with betel-nut chewing have great
potential in reducing the prevalence of carious lesions.
In the Naton Beach sample, betel-nut staining is relegated almost entirely to the
Latte period with only three Pre-Latte individuals displaying betel-nut staining. The
majority of the adult Latte population displays betel-nut staining (64%), while no
juveniles under the age of 18 were found with staining (Parr, 2012). When analysis is
restricted to middle-aged and older adults, the frequency of betel-nut staining rises to
82%. These findings fall within the reported frequency levels reported in the Apurguan
sample, 58.7% (Douglas et al., 1997) and an island-wide survey, 92% (Hanson and
Butler, 1997), both of which also noted low frequencies of carious lesions in the
population. These data, coupled with the paucity of betel-nut chewing in the Pre-Latte
and high prevalence of caries suggest that formation of carious lesions, in Guam, have
an inverse relationship with betel chewing. However, it is interesting to note that the
areca nut tree was present in the Pre-Latte period (Athens and Ward), and whether
betel-chewing was initiated with the purpose of combatting poor dental health is not
known. Nonetheless, the data of the current study indicate that the cultural practice of
betel-nut chewing in the Latte period stymied the effects of a highly cariogenic diet

158
associated with intensification of agricultural practices, resulting in better overall dental
health.
Conclusions
This study investigates patterns of carious lesion frequency across time in the
prehistoric Chamorro and supports and expands on initial findings of temporal change in
relation to dental health reported in an earlier study (Pietrusewsky et al., 1997). Neither
sex-specific nor age-specific changes were noted, indicating uniformity in diet across
the subgroups.
The early Pre-Latte population displays high rates of carious lesions and an
unusual trend of elevated caries in the anterior dentition likely due to dental crowding.
The overall caries rates in the Pre-Latte are more consistent of a population with an
agricultural economy than a horticultural one (Turner, 1979). This finding may be
suggestive increased reliance on taro than was previously expected or sugarcane
consumption; however, neither of these hypotheses are supported by the
archaeological record.
The low levels of caries prevalence in the Latte individuals are surprising given
their carbohydrate-rich and cariogenic diets. This decrease in carious lesion frequency
observed over time is contrary to the expected results as agricultural intensification is
usually associated with a higher degree of carious lesions. This study demonstrates
that relationship between diet and dental caries is not as simplistic as it is often reported
and analysis of carious lesions needs to be performed with careful consideration of the
other factors that may affect dental health, such as food processing and other cultural
factors. In this study, cultural practice of betel-nut chewing, restricted to the Latte
period, had a beneficial effect on the dental health of the Chamorro and limited bacterial

159
infection and enamel destruction through its physical and chemical properties, despite
the highly cariogenic diet of the agricultural population.

160
Table 6-1. Individual occurrence carious lesion frequencies
Pre Latte Latte
n/Na
% n/Na %
Males 25/33 75.8 7/29 24.1
Females 29/38 76.3 11/37 29.7
Indeterminate 18/28 64.3 8/42 19.0
Total 72/99 72.7 26/108 24.1 a. n = number of individuals with carious lesions, N = number of
individuals analyzed Table 6-2. Tooth count of carious lesion frequencies
Pre Latte Latte
n/Na % n/Na %
Males 110/644 17.1 18/508 3.5
Females 94/858 11.0 46/701 6.6
Indeterminate 42/428 9.8 12/527 2.3
Total 246/1930 12.7 76/1736 4.4 a. n = number of individuals with carious lesions, N = number of
individuals analyzed
Table 6-3. Individual occurrence carious lesions by age
Pre Latte Latte
n/N % n/N %
Child 1/2 50.0 0/11 0.0
Juvenile 2/5 40.0 4/13 30.8
YA 50/64 78.1 10/40 25.0
MA 17/23 73.9 10/32 31.3
OA ---- ---- 0/4 0.0
n = number of individuals with carious lesions
N = number of individuals examined
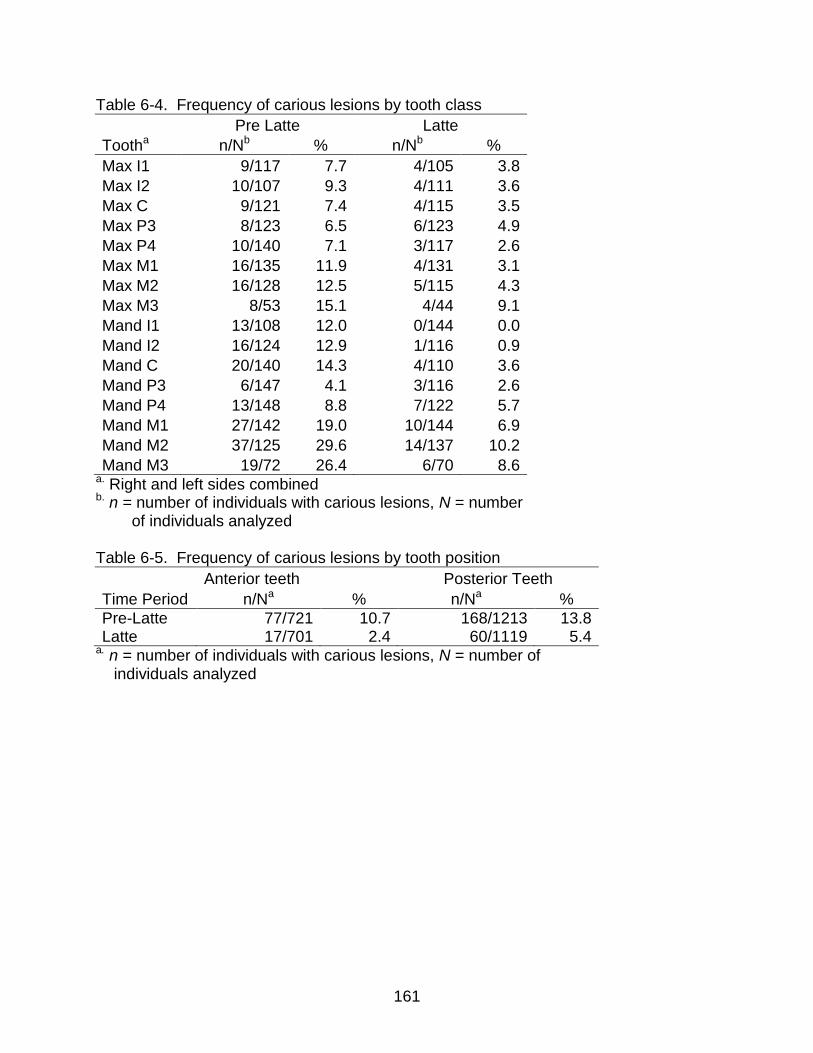
161
Table 6-4. Frequency of carious lesions by tooth class
Pre Latte Latte
Tootha n/Nb % n/Nb %
Max I1 9/117 7.7 4/105 3.8
Max I2 10/107 9.3 4/111 3.6
Max C 9/121 7.4 4/115 3.5
Max P3 8/123 6.5 6/123 4.9
Max P4 10/140 7.1 3/117 2.6
Max M1 16/135 11.9 4/131 3.1
Max M2 16/128 12.5 5/115 4.3
Max M3 8/53 15.1 4/44 9.1
Mand I1 13/108 12.0 0/144 0.0
Mand I2 16/124 12.9 1/116 0.9
Mand C 20/140 14.3 4/110 3.6
Mand P3 6/147 4.1 3/116 2.6
Mand P4 13/148 8.8 7/122 5.7
Mand M1 27/142 19.0 10/144 6.9
Mand M2 37/125 29.6 14/137 10.2
Mand M3 19/72 26.4 6/70 8.6 a. Right and left sides combined b. n = number of individuals with carious lesions, N = number
of individuals analyzed
Table 6-5. Frequency of carious lesions by tooth position
Anterior teeth Posterior Teeth
Time Period n/Na % n/Na %
Pre-Latte 77/721 10.7 168/1213 13.8 Latte 17/701 2.4 60/1119 5.4
a. n = number of individuals with carious lesions, N = number of individuals analyzed

162
Table 6-6. Pearson Chi-Square Test on carious lesion expression
Value Df Asymp. Sig
(2-sided) Exact Sig. (2-sided)
Exact Sig. (1-sided)
Time Periods 49.046 1 0.000* 0.000* 0.000*
Sex within Pre-Latte 0.003 1 0.956+ 1.000+ 0.587+
Sex within Latte 0.256 1 0.613+ 0.782+ 0.412+
Age within Pre-Latte 6.415 3 0.093+ - -
Age within Latte 1.971 4 0.741+ - -
* Indicates statistically significant differences at the 0.05 alpha level + Indicates no statistical relationship at the 0.05 alpha level
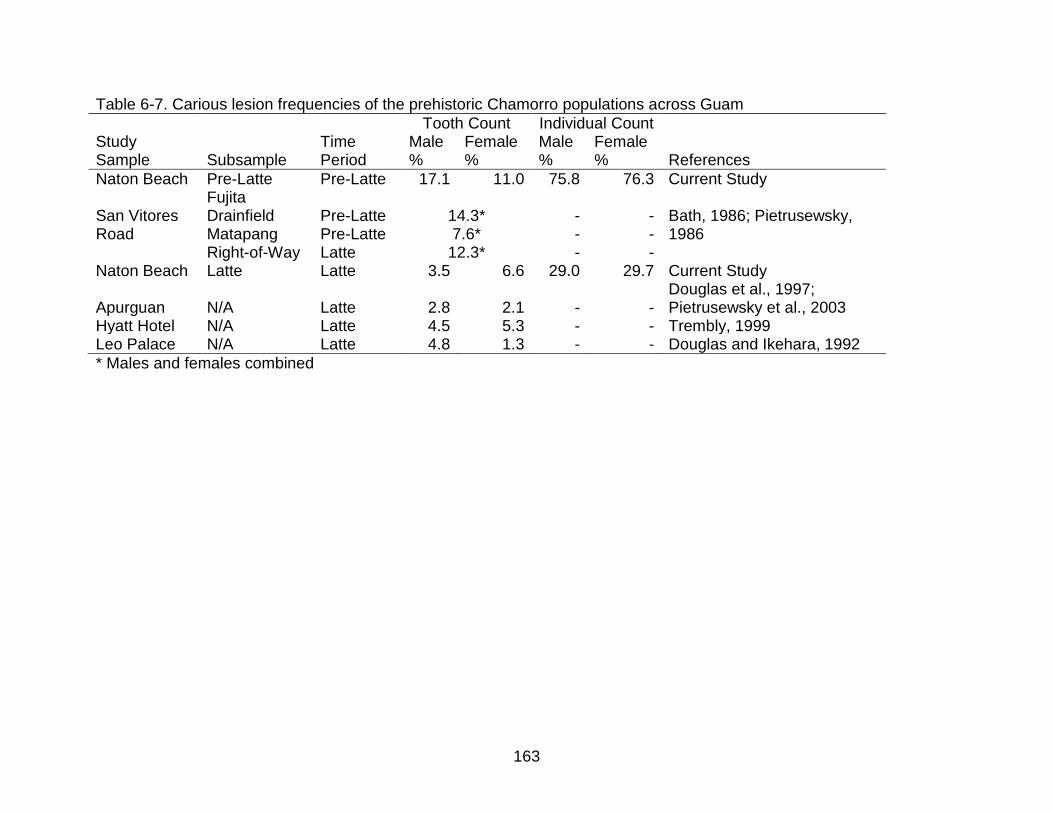
163
Table 6-7. Carious lesion frequencies of the prehistoric Chamorro populations across Guam
Tooth Count Individual Count Study Sample Subsample
Time Period
Male %
Female %
Male %
Female % References
Naton Beach Pre-Latte Pre-Latte 17.1 11.0 75.8 76.3 Current Study
San Vitores Road
Fujita Drainfield Pre-Latte 14.3* - - Bath, 1986; Pietrusewsky,
1986 Matapang Pre-Latte 7.6* - - Right-of-Way Latte 12.3* - -
Naton Beach Latte Latte 3.5 6.6 29.0 29.7 Current Study
Apurguan N/A Latte 2.8 2.1 - - Douglas et al., 1997; Pietrusewsky et al., 2003
Hyatt Hotel N/A Latte 4.5 5.3 - - Trembly, 1999 Leo Palace N/A Latte 4.8 1.3 - - Douglas and Ikehara, 1992
* Males and females combined

164
Figure 6-1. Pre-Latte carious lesions in the anterior dentition (Photo by author).
Figure 6-2. Pre-Latte dental crowding (Photo by author).
Figure 6-3. Betel-nut with piper leaf and slacked lime (Photo by author).

165
CHAPTER 7 SUMMARY
This research is a diachronic investigation of the evolutionary dynamics of the
prehistoric Chamorro population from the Naton Beach Burial Complex in Guam.
Patterns of health and disease are analyzed with a focus on biological processes to see
how they relate to biocultural and environmental change in the prehistoric society.
In an island wide survey of health and disease in the prehistoric Chamorro,
Pietrusewsky and colleagues (1997) hint at temporal changes in disease frequencies
between the time periods and suggest that elevated frequencies of disease are
indicative of higher stress levels in some of the earliest Chamorro populations.
However, their samples sizes are small and restricted to two sites, thus they hesitate on
making definitive conclusions until larger samples became available. The Naton Beach
skeletal population represents the largest and earliest mortuary assemblage excavated
in Guam (DeFant, 2008), and thus allows for a more detailed investigation into
diachronic changes occurring over time.
The transition between the Pre-Latte and Latte time periods are accompanied by
changes in population size, diet and subsistence strategies (Hunter-Anderson and
Butler, 1991; Moore, 2005; Amesbury, 2007). Agricultural intensification with an
increased reliance on staple crops of taro, yam, and rice supplemented by marine
resources replace the earlier marine-dependent, horticultural and forager subsistence
strategies (Hunter-Anderson, 1991; Ambrose et al., 1997; Hanson and Butler, 1997).
These transitions occur concomitantly with large-scale environmental and climatic
fluctuations such as sea-level decline and increased storminess, aridity, and drought
(Hunter-Anderson and Butler, 1991; Nunn, 2007, Hunter-Anderson, 2010). Thus, it was

166
predicted that cultural and environmental shifts are likely to be accompanied by
biological ones, due to increased stress levels associated with malnutrition, limited
access to resources, and increased prevalence of disease.
The dentition of the Pre-Latte sample is significantly larger than the Latte teeth,
with an 8% decrease in overall tooth size over time. Dental reduction did not occur
uniformly throughout the dentition and the greatest amount of reduction was observed in
the buccolingual dimensions and the posterior teeth. Odontometric trends were
analyzed in conjunction with data on craniometrics, stature, and pathological differences
between the periods (Walth, pers. comm.) to assess which of the proposed
mechanisms of dental reduction best fits the data.
The high rates of carious lesions and dental crowding in the Pre-Latte followed by
a significant decrease of caries and reduction in dental crowding in the Latte suggest
that there may have been selection for smaller, less complex teeth that are more
resistant to dental caries, particularly in populations with soft, cariogenic diets. This soft,
cariogenic diet would have occurred with a transition to agriculture and reliance on
staple crops of taro, yam, and rice accompanied by advanced food processing methods,
such as cooking and the use of mortar and pestles, which minimize the force necessary
to break down tough food. With decreased functional demands placed on the
masticatory apparatus, the maxilla and mandible reduce in size followed by a
subsequent decrease in tooth size, as is seen in the Latte population. These findings
best support a combination of Calcagno’s (1986; 1989) Selective Compromise Effect
and Carlson and Van Gerven’s (1977) Masticatory Functional Hypothesis.

167
There were no significant differences in the rates of juvenile and adult hypoplasias;
however, in terms of raw frequency data, juveniles displayed a higher percentage of
hypoplastic defects than adults. Thus, there may be a correlation between life
expectancy and physiological disruptions. Likewise, male and female hypoplasia
frequencies demonstrated no significant differences. Nonetheless, female susceptibility
to physiological stress differs between the Pre-Latte and Latte periods, where females
display fewer hypoplasias than men in the Pre-Latte and higher rates of hypoplastic
defects in the Latte. This transition indicates there may have been differential access to
resources based on gender-roles and division of labor.
Significant differences in hypoplasia frequencies are demonstrated between the
Pre-Latte and Latte populations. The Pre-Latte individuals are less prone to hypoplastic
defects and thus may not have been exposed to high degrees of physiological stressors
as the Latte. Climatic instability, such as typhoons, droughts, and increased aridity, was
more common in the Latte period, resulting in destruction of crops and reef systems,
and likely led to reduced access to nutritional resources and subsequent decrease in
health status.
Analysis of carious lesion frequencies indicates that the Pre-Latte display an
unusually elevated number of caries in the anterior dentition, which may be the result of
dental crowding. No significant differences in carious lesion frequencies were noted
between age groups or sex, but males have more infected teeth, overall, than females.
A significant decrease in caries rates occurs from the Pre-Latte to Latte periods. This
pattern is contrary to the expected results as intensification of agriculture is often
associated with a higher degree of caries, especially with reliance on highly cariogenic

168
staple crops of taro, yam, and rice. The practice of betel-nut chewing, which is
restricted almost entirely to the Latte period, could explain this disparity as it is known to
have cariostatic properties and may lead to better dental health in the Latte despite the
highly cariogenic diet brought on by an agricultural transition. Finally, this study is a
clear example of the multifactorial nature of carious lesions and demonstrates the
importance of evaluating factors other than diet and warns against making broad
generalizations about the relationship between subsistence strategies and carious
lesions.
Future Studies
The majority of archaeological, linguistic, and biological studies have focused on
settlement of Remote Oceania through the Lapita expansion that began around 3200
BP and led to the colonization of Polynesia (Kirch, 2010). However, settlement of the
Marianas Islands has been securely dated to 3600 to 3420 BP (Carson, 2008; Clarke et
al., 2010), approximately 400 years before the Lapita peoples ventured into the remote
Pacific.
Biodistance studies investigating settlement patterns of the Marianas Islands have
been restricted almost entirely to skeletal remains from the Latte period (Pietrusewsky,
1990a; 1990b; 1994; 2006; Ishida and Dodo, 1997). The current study demonstrates
that significant biological changes have occurred between the early and late prehistoric
Chamorro populations. Thus, comparisons of the Pre-Latte Naton Beach sample with
circum-Pacific skeletal populations may shed new light on settlement and migration
patterns of the Marianas Islands and Remote Oceania.

169
Archaeological, paleoenvironmental, and stable isotope data suggest
intensification of agriculture with an increased reliance on terrestrial crops and decrease
exploitation of marine resources (Hunter-Anderson and Butler, 1991; Ambrose et al.,
1997; Athens and Ward, 2004; Moore, 2005; Amesbury, 2007). However, only one
stable isotope study from Saipan incorporates samples from the Pre-Latte period (n = 5)
(McGovern and Wilson, 1996). Analysis of carbon and nitrogen stable isotope ratios
from the Pre-Latte Naton Beach sample in combination with indicators of dental health
will help clarify temporal variability in diet and health as it relates to subsistence change
in the prehistoric Chamorro.
This study utilizes a diachronic approach to evaluate changes in dental health
between the Pre-Latte and Latte periods and provides information on the health of some
of the earliest settlers of Guam, which was previously unknown. Archaeological data
was evaluated to see if biological disparities in the populations correlate to
environmental and cultural changes shifts. Large-scale environmental and ecosystem
fluctuations necessitated cultural shifts in subsistence strategies, resulting in dental
reduction and an increase in physiological disturbances. Carious lesion rates, however,
did not increase with the subsistence shifts, and instead improved in the Latte as the
result of betel-nut chewing. This temporal assessment of the dentition identifies cultural
and environmental processes that led to biological change in the prehistoric Chamorro.

170
APPENDIX A DENTAL METRICS
Table A-1. Descriptive statistics of dental measurements
Measurement N Minimum Maximum Mean Std. Dev.
LMax I1 MD 99 7.7 10.4 8.9 0.6
LMax I1 BL 100 6.9 8.9 7.7 0.4
LMax I2 MD 100 6.2 8.6 7.2 0.5
LMax I2 BL 98 6.1 8.7 7.1 0.4
LMax C MD 108 7.1 9.6 8.4 0.5
LMax C BL 111 7.2 10.1 8.8 0.5
LMax P3 MD 120 6.9 10.3 7.9 0.5
LMax P3 BL 121 7.5 11.6 10.1 0.6
LMax P4 MD 133 6.5 9.9 7.6 0.5
LMax P4 BL 130 7.1 12.0 10.1 0.6
LMax M1 MD 119 9.9 12.5 11.3 0.6
LMax M1 BL 127 9.6 13.6 11.9 0.6
LMax M2 MD 118 8.4 12.4 10.5 0.7
LMax M2 BL 124 9.6 14.3 11.9 0.8
LMax M3 MD 47 7.2 13.0 9.4 1.0
LMax M3 BL 50 9.1 13.0 11.4 0.8
RMax I1 MD 102 7.2 10.0 8.8 0.6
RMax I1 BL 107 6.9 8.6 7.7 0.4
RMax I2 MD 96 6.1 8.4 7.2 0.5
RMax I2 BL 97 6.0 8.1 7.1 0.4
RMax C MD 114 7.3 9.6 8.5 0.4
RMax C BL 111 6.7 10.0 8.7 0.5
RMax P3 MD 119 6.2 9.0 7.8 0.5
RMax P3 BL 121 8.5 11.5 10.2 0.6
RMax P4 MD 126 6.2 9.2 7.5 0.5
RMax P4 BL 126 8.5 12.0 10.1 0.6
RMax M1 MD 127 10.1 13.1 11.4 0.6
RMax M1 BL 137 10.4 13.7 11.8 0.6
RMax M2 MD 121 7.6 13.0 10.4 0.7
RMax M2 BL 128 10.0 13.6 11.7 0.8
RMax M3 MD 45 7.0 11.3 9.3 0.9
RMax M3 BL 45 9.6 13.5 11.3 0.8
LMand I1 MD 95 4.1 6.8 5.7 0.4
LMand I1 BL 88 5.4 7.6 6.3 0.4
LMand I2 MD 101 4.9 7.4 6.4 0.5
LMand I2 BL 104 5.6 7.9 6.7 0.4
LMand C MD 117 4.4 8.4 7.3 0.5
LMand C BL 120 6.8 9.6 8.1 0.6

171
Table A-1. Continued
Measurement N Minimum Maximum Mean Std. Dev.
LMand P3 MD 134 6.5 9.1 7.9 0.5
LMand P3 BL 133 7.4 10.4 8.9 0.6
LMand P4 MD 132 6.8 9.2 8.0 0.5
LMand P4 BL 128 7.5 10.6 9.0 0.6
LMand M1 MD 126 11.2 13.9 12.5 0.6
LMand M1 BL 140 9.4 12.3 10.8 0.5
LMand M2 MD 122 10.1 13.8 11.9 0.8
LMand M2 BL 130 6.7 12.7 10.6 0.8
LMand M3 MD 62 8.3 14.1 11.0 1.1
LMand M3 BL 67 8.8 12.3 10.3 0.8
RMand I1 MD 87 4.7 6.7 5.7 0.4
RMand I1 BL 83 5.6 7.3 6.3 0.4
RMand I2 MD 104 5.5 7.5 6.5 0.4
RMand I2 BL 105 5.9 7.7 6.8 0.4
RMand C MD 115 6.2 8.4 7.3 0.4
RMand C BL 114 7.0 9.5 8.1 0.5
RMand P3 MD 126 6.1 9.1 7.8 0.5
RMand P3 BL 125 6.1 10.3 8.9 0.6
RMand P4 MD 126 6.6 9.0 7.9 0.5
RMand P4 BL 133 7.3 11.8 9.0 0.6
RMand M1 MD 133 10.5 13.8 12.4 0.6
RMand M1 BL 145 9.6 12.2 10.9 0.5
RMand M2 MD 113 7.9 13.6 11.8 0.8
RMand M2 BL 127 9.2 12.2 10.6 0.6
RMand M3 MD 58 8.8 14.0 11.2 0.9
RMand M3 BL 64 8.4 12.8 10.4 0.8
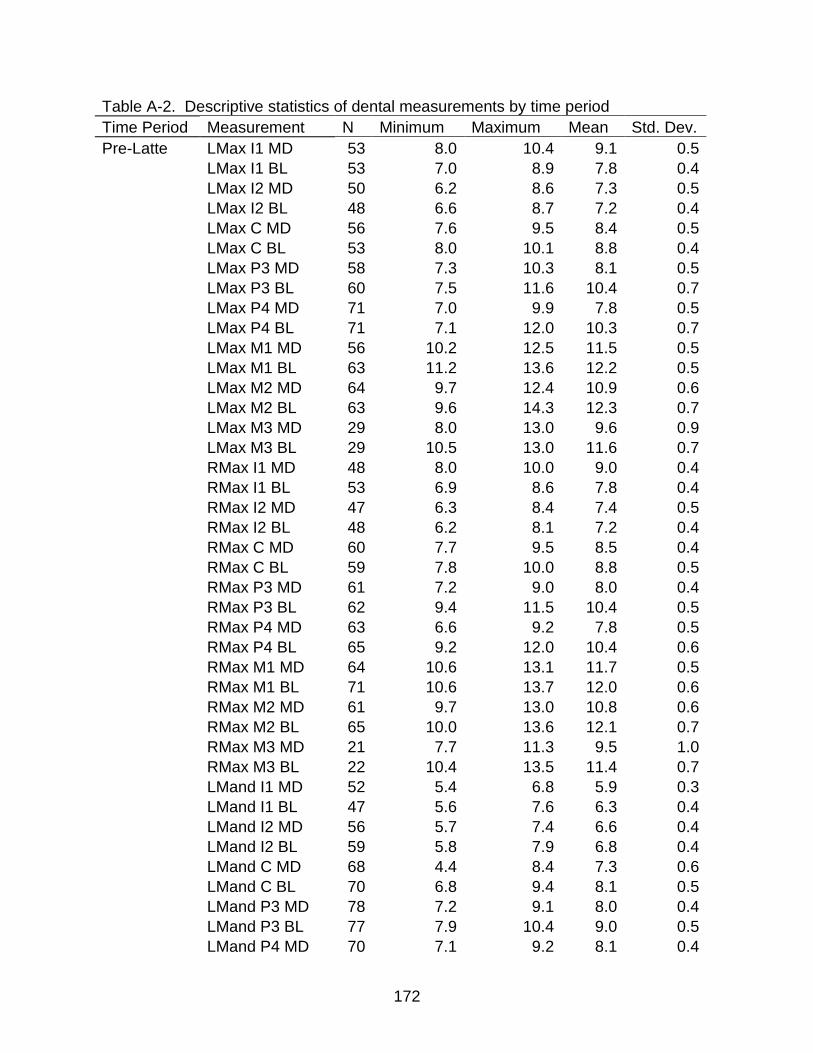
172
Table A-2. Descriptive statistics of dental measurements by time period
Time Period Measurement N Minimum Maximum Mean Std. Dev.
Pre-Latte LMax I1 MD 53 8.0 10.4 9.1 0.5
LMax I1 BL 53 7.0 8.9 7.8 0.4
LMax I2 MD 50 6.2 8.6 7.3 0.5
LMax I2 BL 48 6.6 8.7 7.2 0.4
LMax C MD 56 7.6 9.5 8.4 0.5
LMax C BL 53 8.0 10.1 8.8 0.4
LMax P3 MD 58 7.3 10.3 8.1 0.5
LMax P3 BL 60 7.5 11.6 10.4 0.7
LMax P4 MD 71 7.0 9.9 7.8 0.5
LMax P4 BL 71 7.1 12.0 10.3 0.7
LMax M1 MD 56 10.2 12.5 11.5 0.5
LMax M1 BL 63 11.2 13.6 12.2 0.5
LMax M2 MD 64 9.7 12.4 10.9 0.6
LMax M2 BL 63 9.6 14.3 12.3 0.7
LMax M3 MD 29 8.0 13.0 9.6 0.9
LMax M3 BL 29 10.5 13.0 11.6 0.7
RMax I1 MD 48 8.0 10.0 9.0 0.4
RMax I1 BL 53 6.9 8.6 7.8 0.4
RMax I2 MD 47 6.3 8.4 7.4 0.5
RMax I2 BL 48 6.2 8.1 7.2 0.4
RMax C MD 60 7.7 9.5 8.5 0.4
RMax C BL 59 7.8 10.0 8.8 0.5
RMax P3 MD 61 7.2 9.0 8.0 0.4
RMax P3 BL 62 9.4 11.5 10.4 0.5
RMax P4 MD 63 6.6 9.2 7.8 0.5
RMax P4 BL 65 9.2 12.0 10.4 0.6
RMax M1 MD 64 10.6 13.1 11.7 0.5
RMax M1 BL 71 10.6 13.7 12.0 0.6
RMax M2 MD 61 9.7 13.0 10.8 0.6
RMax M2 BL 65 10.0 13.6 12.1 0.7
RMax M3 MD 21 7.7 11.3 9.5 1.0
RMax M3 BL 22 10.4 13.5 11.4 0.7
LMand I1 MD 52 5.4 6.8 5.9 0.3
LMand I1 BL 47 5.6 7.6 6.3 0.4
LMand I2 MD 56 5.7 7.4 6.6 0.4
LMand I2 BL 59 5.8 7.9 6.8 0.4
LMand C MD 68 4.4 8.4 7.3 0.6
LMand C BL 70 6.8 9.4 8.1 0.5
LMand P3 MD 78 7.2 9.1 8.0 0.4
LMand P3 BL 77 7.9 10.4 9.0 0.5
LMand P4 MD 70 7.1 9.2 8.1 0.4
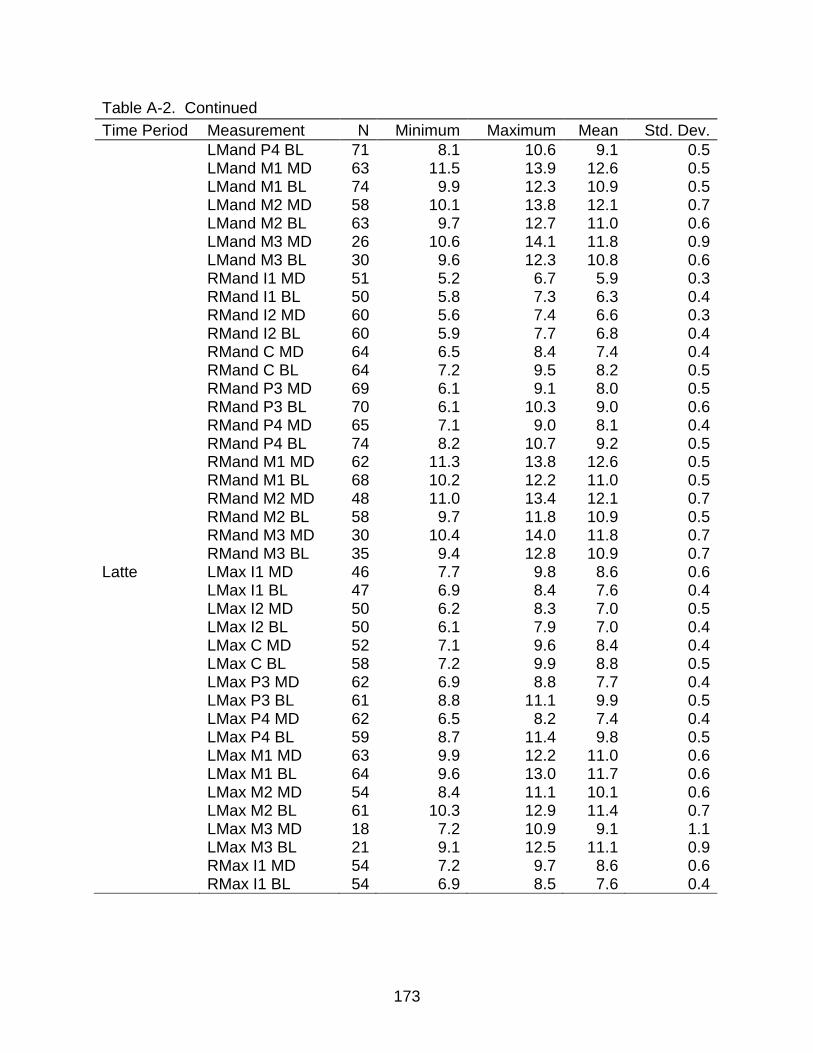
173
Table A-2. Continued
Time Period Measurement N Minimum Maximum Mean Std. Dev.
LMand P4 BL 71 8.1 10.6 9.1 0.5 LMand M1 MD 63 11.5 13.9 12.6 0.5 LMand M1 BL 74 9.9 12.3 10.9 0.5 LMand M2 MD 58 10.1 13.8 12.1 0.7 LMand M2 BL 63 9.7 12.7 11.0 0.6 LMand M3 MD 26 10.6 14.1 11.8 0.9 LMand M3 BL 30 9.6 12.3 10.8 0.6 RMand I1 MD 51 5.2 6.7 5.9 0.3 RMand I1 BL 50 5.8 7.3 6.3 0.4 RMand I2 MD 60 5.6 7.4 6.6 0.3 RMand I2 BL 60 5.9 7.7 6.8 0.4 RMand C MD 64 6.5 8.4 7.4 0.4 RMand C BL 64 7.2 9.5 8.2 0.5 RMand P3 MD 69 6.1 9.1 8.0 0.5 RMand P3 BL 70 6.1 10.3 9.0 0.6 RMand P4 MD 65 7.1 9.0 8.1 0.4 RMand P4 BL 74 8.2 10.7 9.2 0.5 RMand M1 MD 62 11.3 13.8 12.6 0.5 RMand M1 BL 68 10.2 12.2 11.0 0.5 RMand M2 MD 48 11.0 13.4 12.1 0.7 RMand M2 BL 58 9.7 11.8 10.9 0.5 RMand M3 MD 30 10.4 14.0 11.8 0.7 RMand M3 BL 35 9.4 12.8 10.9 0.7 Latte LMax I1 MD 46 7.7 9.8 8.6 0.6 LMax I1 BL 47 6.9 8.4 7.6 0.4 LMax I2 MD 50 6.2 8.3 7.0 0.5 LMax I2 BL 50 6.1 7.9 7.0 0.4 LMax C MD 52 7.1 9.6 8.4 0.4 LMax C BL 58 7.2 9.9 8.8 0.5 LMax P3 MD 62 6.9 8.8 7.7 0.4 LMax P3 BL 61 8.8 11.1 9.9 0.5 LMax P4 MD 62 6.5 8.2 7.4 0.4 LMax P4 BL 59 8.7 11.4 9.8 0.5 LMax M1 MD 63 9.9 12.2 11.0 0.6 LMax M1 BL 64 9.6 13.0 11.7 0.6 LMax M2 MD 54 8.4 11.1 10.1 0.6 LMax M2 BL 61 10.3 12.9 11.4 0.7 LMax M3 MD 18 7.2 10.9 9.1 1.1 LMax M3 BL 21 9.1 12.5 11.1 0.9 RMax I1 MD 54 7.2 9.7 8.6 0.6 RMax I1 BL 54 6.9 8.5 7.6 0.4

174
Table A-2. Continued
Time Period Measurement N Minimum Maximum Mean Std. Dev.
RMax I2 MD 49 6.1 8.0 7.0 0.5 RMax I2 BL 49 6.0 8.0 7.0 0.4 RMax C MD 54 7.3 9.6 8.4 0.5 RMax C BL 52 6.7 9.8 8.7 0.6 RMax P3 MD 58 6.2 8.8 7.6 0.5 RMax P3 BL 59 8.5 11.3 9.9 0.5 RMax P4 MD 63 6.2 8.0 7.3 0.4 RMax P4 BL 61 8.5 11.0 9.7 0.5 RMax M1 MD 63 10.1 12.6 11.2 0.5 RMax M1 BL 66 10.4 12.9 11.5 0.5 RMax M2 MD 60 7.6 12.3 10.0 0.6 RMax M2 BL 63 10.4 12.8 11.3 0.6 RMax M3 MD 24 7.0 10.4 9.1 0.9 RMax M3 BL 23 9.6 12.8 11.1 0.8 LMand I1 MD 43 4.1 6.2 5.5 0.4 LMand I1 BL 41 5.4 6.8 6.2 0.4 LMand I2 MD 45 4.9 7.3 6.2 0.5 LMand I2 BL 45 5.6 7.4 6.6 0.4 LMand C MD 49 6.0 8.2 7.2 0.5 LMand C BL 50 6.9 9.6 7.9 0.6 LMand P3 MD 56 6.5 8.6 7.6 0.5 LMand P3 BL 56 7.4 10.4 8.7 0.6 LMand P4 MD 62 6.8 8.9 7.8 0.5 LMand P4 BL 57 7.5 10.2 8.8 0.6 LMand M1 MD 63 11.2 13.9 12.4 0.6 LMand M1 BL 66 9.4 12.1 10.6 0.6 LMand M2 MD 64 10.1 13.8 11.7 0.8 LMand M2 BL 67 6.7 11.8 10.3 0.8 LMand M3 MD 36 8.3 12.8 10.5 1.0 LMand M3 BL 37 8.8 11.2 9.9 0.7 RMand I1 MD 36 4.7 6.1 5.5 0.4 RMand I1 BL 33 5.6 6.8 6.2 0.3 RMand I2 MD 44 5.5 7.5 6.3 0.4 RMand I2 BL 45 5.9 7.4 6.6 0.4 RMand C MD 51 6.2 8.1 7.2 0.4 RMand C BL 50 7.0 9.1 8.1 0.5 RMand P3 MD 57 6.7 8.8 7.6 0.5 RMand P3 BL 55 7.6 10.3 8.8 0.6 RMand P4 MD 61 6.6 8.8 7.8 0.5 RMand P4 BL 59 7.3 11.8 8.8 0.7 RMand M1 MD 71 10.5 13.8 12.3 0.6 RMand M1 BL 77 9.6 12.1 10.7 0.6 RMand M2 MD 65 7.9 13.6 11.6 0.9
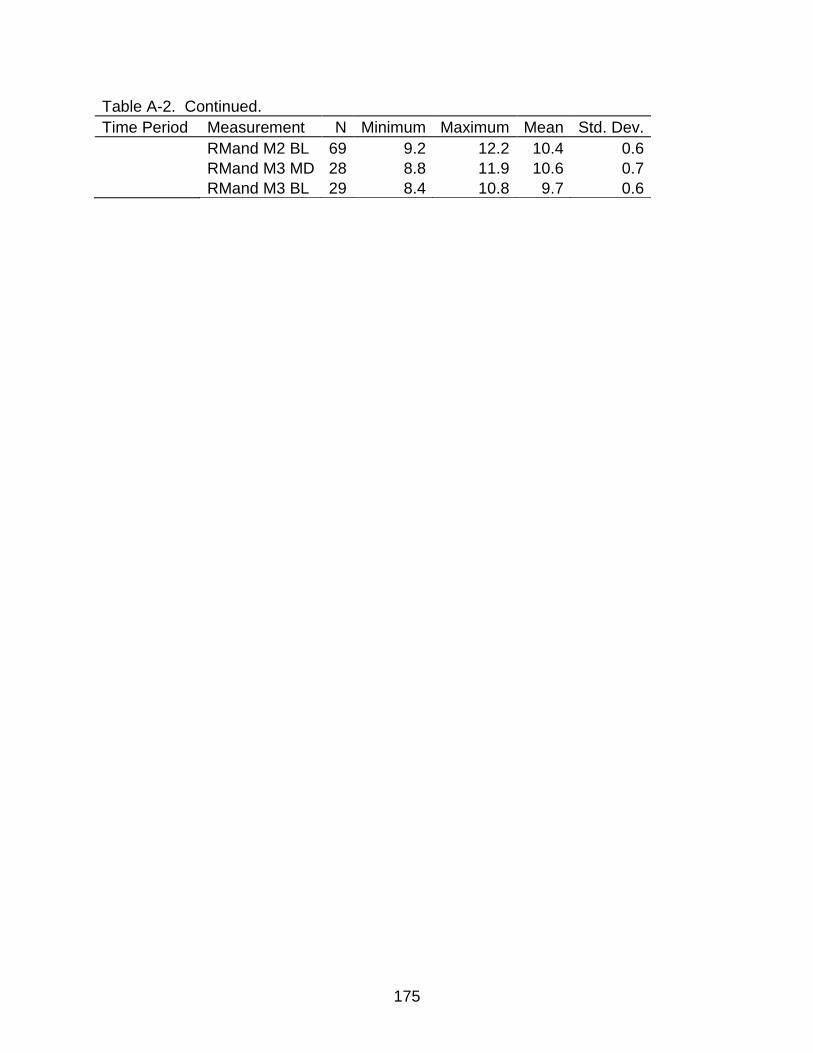
175
Table A-2. Continued.
Time Period Measurement N Minimum Maximum Mean Std. Dev.
RMand M2 BL 69 9.2 12.2 10.4 0.6
RMand M3 MD 28 8.8 11.9 10.6 0.7
RMand M3 BL 29 8.4 10.8 9.7 0.6

176
Table A-3. Descriptive statistics of dental measurements by time period and sex.
Time Period Sex Measurement N Minimum Maximum Mean Std. Dev.
Pre-Latte Male LMax I1 MD 17 8.2 10.1 9.2 0.5
LMax I1 BL 16 7.6 8.5 7.9 0.3
LMax I2 MD 13 6.2 8.4 7.3 0.7
LMax I2 BL 14 6.7 8.0 7.2 0.4
LMax C MD 17 7.8 9.5 8.7 0.5
LMax C BL 15 8.1 9.4 8.8 0.4
LMax P3 MD 21 7.4 10.3 8.2 0.6
LMax P3 BL 21 7.5 11.6 10.4 0.8
LMax P4 MD 22 7.0 9.9 7.9 0.7
LMax P4 BL 23 7.1 12.0 10.4 0.9
LMax M1 MD 15 11.0 12.5 11.8 0.4
LMax M1 BL 20 11.8 13.6 12.5 0.6
LMax M2 MD 20 9.7 11.9 10.9 0.5
LMax M2 BL 21 11.4 13.7 12.5 0.6
LMax M3 MD 10 8.9 13.0 9.9 1.2
LMax M3 BL 10 10.6 13.0 11.8 0.7
RMax I1 MD 14 8.0 9.8 9.1 0.5
RMax I1 BL 18 6.9 8.5 7.8 0.4
RMax I2 MD 14 6.8 8.4 7.5 0.5
RMax I2 BL 15 6.4 8.0 7.2 0.5
RMax C MD 20 8.0 9.5 8.7 0.4
RMax C BL 19 7.8 9.6 8.8 0.5
RMax P3 MD 19 7.4 8.9 8.0 0.4
RMax P3 BL 20 9.7 11.3 10.4 0.5
RMax P4 MD 17 7.3 9.2 7.9 0.6
RMax P4 BL 19 9.4 12.0 10.6 0.6
RMax M1 MD 20 10.6 12.9 11.8 0.5
RMax M1 BL 22 11.6 13.7 12.4 0.6
RMax M2 MD 24 9.9 11.9 10.9 0.6
RMax M2 BL 25 11.1 13.6 12.4 0.6
RMax M3 MD 6 8.4 11.3 9.9 1.2

177
Table A-3. Continued
Time Period Sex Measurement N Minimum Maximum Mean Std. Dev.
RMax M3 BL 7 10.8 13.5 12.0 0.8
LMand I1 MD 17 5.6 6.8 5.9 0.3
LMand I1 BL 16 5.6 7.6 6.4 0.5
LMand I2 MD 17 5.9 7.2 6.6 0.3
LMand I2 BL 20 6.3 7.6 6.8 0.3
LMand C MD 21 4.4 8.4 7.6 0.8
LMand C BL 24 7.2 9.2 8.4 0.5
LMand P3 MD 26 7.4 9.1 8.1 0.4
LMand P3 BL 26 8.1 9.9 9.0 0.4
LMand P4 MD 20 7.3 8.9 8.2 0.4
LMand P4 BL 22 8.5 10.6 9.3 0.6
LMand M1 MD 17 11.8 13.8 12.7 0.5
LMand M1 BL 22 10.4 12.0 11.1 0.4
LMand M2 MD 16 10.1 13.3 12.0 0.8
LMand M2 BL 18 10.1 12.0 11.2 0.6
LMand M3 MD 6 10.9 12.7 11.8 0.8
LMand M3 BL 9 9.8 11.6 11.0 0.6
RMand I1 MD 15 5.2 6.2 5.8 0.3
RMand I1 BL 15 5.9 6.9 6.3 0.3
RMand I2 MD 18 5.6 7.3 6.6 0.4
RMand I2 BL 21 6.1 7.6 6.8 0.4
RMand C MD 21 6.7 8.4 7.5 0.4
RMand C BL 21 7.2 9.0 8.2 0.5
RMand P3 MD 23 6.1 9.0 8.0 0.6
RMand P3 BL 24 6.1 10.1 8.9 0.8
RMand P4 MD 20 7.7 9.0 8.3 0.3
RMand P4 BL 24 8.5 10.7 9.4 0.6
RMand M1 MD 19 12.1 13.6 12.9 0.4
RMand M1 BL 22 10.6 12.2 11.3 0.4
RMand M2 MD 11 11.0 13.4 11.9 0.7
RMand M2 BL 15 10.3 11.6 11.1 0.4

178
Table A-3 Continued
Time Period Sex Measurement N Minimum Maximum Mean Std. Dev.
RMand M3 MD 8 11.4 13.1 12.1 0.6
RMand M3 BL 9 10.2 11.6 11.0 0.5
Female LMax I1 MD 21 8.0 9.8 8.9 0.4
LMax I1 BL 23 7.0 8.3 7.7 0.3
LMax I2 MD 22 6.5 8.6 7.3 0.5
LMax I2 BL 20 6.6 7.5 7.0 0.2
LMax C MD 24 7.7 8.8 8.2 0.4
LMax C BL 24 8.0 9.5 8.7 0.4
LMax P3 MD 23 7.3 9.0 8.0 0.4
LMax P3 BL 26 9.2 11.5 10.4 0.5
LMax P4 MD 31 7.1 8.3 7.6 0.4
LMax P4 BL 33 9.2 11.4 10.3 0.4
LMax M1 MD 27 10.4 12.5 11.4 0.5
LMax M1 BL 29 11.3 13.2 12.0 0.4
LMax M2 MD 30 9.7 11.9 10.7 0.5
LMax M2 BL 30 11.4 14.0 12.2 0.6
LMax M3 MD 15 8.0 10.2 9.3 0.7
LMax M3 BL 15 10.5 12.6 11.4 0.6
RMax I1 MD 20 8.3 9.7 9.0 0.4
RMax I1 BL 22 7.0 8.3 7.7 0.4
RMax I2 MD 20 6.3 8.0 7.3 0.5
RMax I2 BL 20 6.2 8.1 7.1 0.4
RMax C MD 27 7.7 9.1 8.3 0.3
RMax C BL 27 8.0 9.4 8.7 0.4
RMax P3 MD 25 7.2 9.0 7.9 0.4
RMax P3 BL 26 9.5 11.4 10.4 0.4
RMax P4 MD 29 6.6 8.5 7.6 0.4
RMax P4 BL 29 9.2 11.1 10.2 0.5
RMax M1 MD 23 10.9 13.1 11.7 0.5
RMax M1 BL 29 10.6 13.0 11.8 0.5
RMax M2 MD 25 9.7 11.7 10.6 0.5
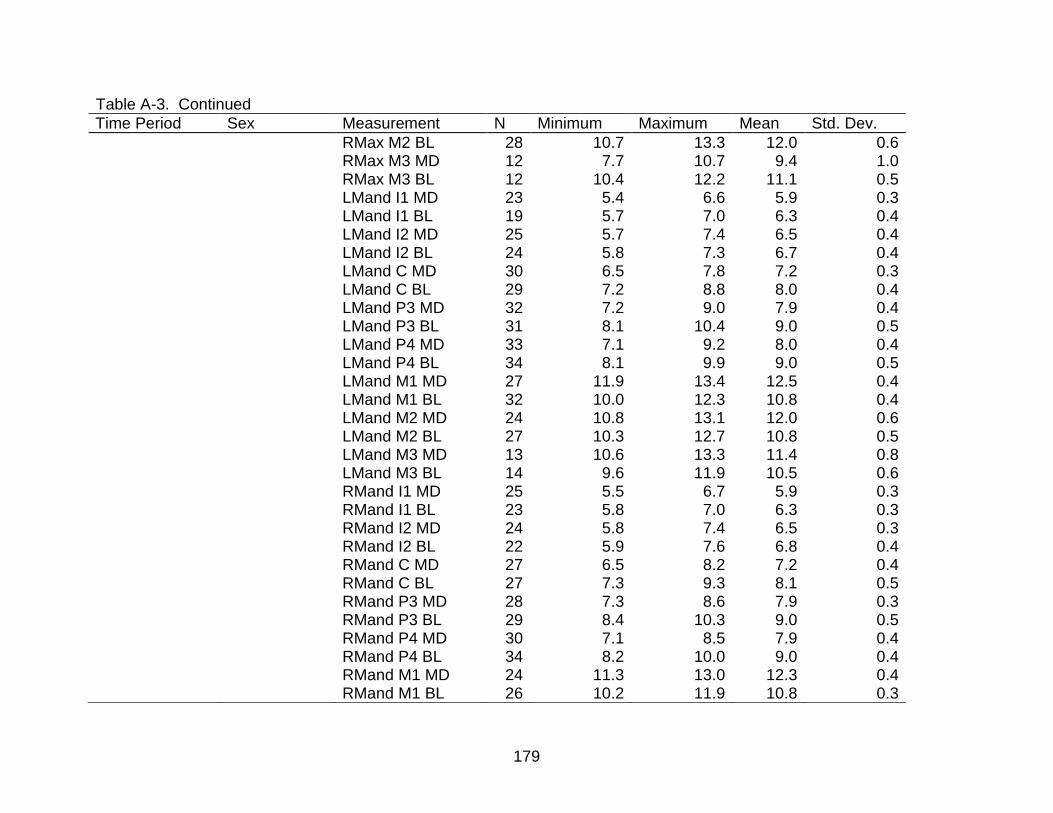
179
Table A-3. Continued
Time Period Sex Measurement N Minimum Maximum Mean Std. Dev.
RMax M2 BL 28 10.7 13.3 12.0 0.6
RMax M3 MD 12 7.7 10.7 9.4 1.0
RMax M3 BL 12 10.4 12.2 11.1 0.5
LMand I1 MD 23 5.4 6.6 5.9 0.3
LMand I1 BL 19 5.7 7.0 6.3 0.4
LMand I2 MD 25 5.7 7.4 6.5 0.4
LMand I2 BL 24 5.8 7.3 6.7 0.4
LMand C MD 30 6.5 7.8 7.2 0.3
LMand C BL 29 7.2 8.8 8.0 0.4
LMand P3 MD 32 7.2 9.0 7.9 0.4
LMand P3 BL 31 8.1 10.4 9.0 0.5
LMand P4 MD 33 7.1 9.2 8.0 0.4
LMand P4 BL 34 8.1 9.9 9.0 0.5
LMand M1 MD 27 11.9 13.4 12.5 0.4
LMand M1 BL 32 10.0 12.3 10.8 0.4
LMand M2 MD 24 10.8 13.1 12.0 0.6
LMand M2 BL 27 10.3 12.7 10.8 0.5
LMand M3 MD 13 10.6 13.3 11.4 0.8
LMand M3 BL 14 9.6 11.9 10.5 0.6
RMand I1 MD 25 5.5 6.7 5.9 0.3
RMand I1 BL 23 5.8 7.0 6.3 0.3
RMand I2 MD 24 5.8 7.4 6.5 0.3
RMand I2 BL 22 5.9 7.6 6.8 0.4
RMand C MD 27 6.5 8.2 7.2 0.4
RMand C BL 27 7.3 9.3 8.1 0.5
RMand P3 MD 28 7.3 8.6 7.9 0.3
RMand P3 BL 29 8.4 10.3 9.0 0.5
RMand P4 MD 30 7.1 8.5 7.9 0.4
RMand P4 BL 34 8.2 10.0 9.0 0.4
RMand M1 MD 24 11.3 13.0 12.3 0.4
RMand M1 BL 26 10.2 11.9 10.8 0.3

180
Table A-3. Continued
Time Period Sex Measurement N Minimum Maximum Mean Std. Dev.
RMand M2 MD 23 11.2 12.9 12.0 0.6
RMand M2 BL 27 10.2 11.7 10.8 0.4
RMand M3 MD 13 10.4 12.3 11.5 0.5
RMand M3 BL 16 9.4 12.8 10.7 0.8
Indet. LMax I1 MD 15 8.5 10.4 9.3 0.5
LMax I1 BL 14 7.1 8.9 7.9 0.6
LMax I2 MD 15 6.7 8.2 7.4 0.5
LMax I2 BL 14 6.7 8.7 7.4 0.5
LMax C MD 15 7.6 9.2 8.4 0.5
LMax C BL 14 8.0 10.1 8.8 0.6
LMax P3 MD 14 7.3 8.6 8.1 0.4
LMax P3 BL 13 9.0 11.4 10.2 0.7
LMax P4 MD 18 7.2 9.0 7.8 0.5
LMax P4 BL 15 9.2 11.3 10.4 0.6
LMax M1 MD 14 10.2 12.0 11.5 0.5
LMax M1 BL 14 11.2 12.9 12.1 0.4
LMax M2 MD 14 10.2 12.4 11.2 0.6
LMax M2 BL 12 9.6 14.3 12.2 1.1
LMax M3 MD 4 8.6 10.3 9.8 0.8
LMax M3 BL 4 11.3 12.5 11.8 0.6
RMax I1 MD 14 8.3 10.0 9.1 0.5
RMax I1 BL 13 7.2 8.6 7.9 0.5
RMax I2 MD 13 6.9 8.2 7.6 0.4
RMax I2 BL 13 6.5 8.0 7.2 0.4
RMax C MD 13 7.8 9.3 8.7 0.5
RMax C BL 13 7.9 10.0 8.8 0.7
RMax P3 MD 17 7.6 8.6 8.1 0.3
RMax P3 BL 16 9.4 11.5 10.4 0.6
RMax P4 MD 17 6.9 9.1 7.9 0.5
RMax P4 BL 17 9.2 11.1 10.3 0.6
RMax M1 MD 21 10.7 12.4 11.5 0.5

181
Table A-3. Continued
Time Period Sex Measurement N Minimum Maximum Mean Std. Dev.
RMax M1 BL 20 11.2 13.1 12.0 0.4
RMax M2 MD 12 9.9 13.0 11.0 0.8
RMax M2 BL 12 10.0 13.1 11.9 0.8
RMax M3 MD 3 8.8 9.7 9.3 0.5
RMax M3 BL 3 10.9 11.5 11.2 0.3
LMand I1 MD 12 5.5 6.6 5.9 0.4
LMand I1 BL 12 5.9 7.1 6.4 0.4
LMand I2 MD 14 6.1 7.2 6.7 0.3
LMand I2 BL 15 6.4 7.9 6.9 0.5
LMand C MD 17 6.6 8.2 7.3 0.5
LMand C BL 17 6.8 9.4 8.0 0.7
LMand P3 MD 20 7.3 9.0 8.1 0.5
LMand P3 BL 20 7.9 9.9 9.0 0.5
LMand P4 MD 17 7.7 9.0 8.2 0.4
LMand P4 BL 15 8.4 10.0 9.2 0.5
LMand M1 MD 19 11.5 13.9 12.7 0.6
LMand M1 BL 20 9.9 11.9 10.9 0.5
LMand M2 MD 18 11.0 13.8 12.4 0.8
LMand M2 BL 18 9.7 12.5 11.1 0.7
LMand M3 MD 7 11.5 14.1 12.4 0.8
LMand M3 BL 7 10.5 12.3 11.1 0.6
RMand I1 MD 11 5.5 6.6 6.0 0.4
RMand I1 BL 12 5.8 7.3 6.4 0.5
RMand I2 MD 18 6.0 7.1 6.5 0.3
RMand I2 BL 17 6.4 7.7 6.8 0.4
RMand C MD 16 6.8 8.3 7.4 0.5
RMand C BL 16 7.6 9.5 8.2 0.6
RMand P3 MD 18 7.3 9.1 8.1 0.5
RMand P3 BL 17 8.1 9.8 9.0 0.5
RMand P4 MD 15 7.4 8.7 8.1 0.4
RMand P4 BL 16 8.5 9.9 9.1 0.5

182
Table A-3. Continued
Time Period Sex Measurement N Minimum Maximum Mean Std. Dev.
RMand M1 MD 19 11.5 13.8 12.7 0.6
RMand M1 BL 20 10.4 12.0 10.9 0.4
RMand M2 MD 14 11.0 13.4 12.2 0.8
RMand M2 BL 16 9.7 11.8 11.0 0.6
RMand M3 MD 9 11.0 14.0 12.0 0.9
RMand M3 BL 10 10.2 12.2 11.0 0.6
Latte Male LMax I1 MD 13 7.8 9.8 8.7 0.6
LMax I1 BL 14 6.9 8.4 7.8 0.5
LMax I2 MD 15 6.4 8.1 7.0 0.5
LMax I2 BL 15 6.6 7.7 7.2 0.4
LMax C MD 15 8.1 9.4 8.6 0.4
LMax C BL 15 8.3 9.9 9.1 0.5
LMax P3 MD 17 7.1 8.8 7.8 0.4
LMax P3 BL 17 8.8 11.1 10.1 0.5
LMax P4 MD 16 7.0 8.1 7.6 0.4
LMax P4 BL 15 9.5 11.4 10.2 0.5
LMax M1 MD 15 10.5 12.0 11.3 0.5
LMax M1 BL 15 11.0 12.9 12.0 0.6
LMax M2 MD 10 9.7 11.1 10.4 0.5
LMax M2 BL 13 10.8 12.7 11.8 0.7
LMax M3 MD 8 8.1 10.9 9.7 0.9
LMax M3 BL 10 10.3 12.5 11.4 0.8
RMax I1 MD 15 7.6 9.7 8.7 0.6
RMax I1 BL 16 7.0 8.5 7.7 0.4
RMax I2 MD 13 6.3 7.9 6.9 0.4
RMax I2 BL 14 6.7 7.9 7.2 0.4
RMax C MD 16 8.0 9.5 8.7 0.4
RMax C BL 15 8.2 9.8 8.9 0.4
RMax P3 MD 20 7.2 8.8 7.8 0.4
RMax P3 BL 20 8.8 11.3 10.1 0.5
RMax P4 MD 17 6.8 8.0 7.5 0.3

183
Table A-3. Continued
Time Period Sex Measurement N Minimum Maximum Mean Std. Dev.
RMax P4 BL 16 9.4 11.0 10.1 0.5
RMax M1 MD 19 10.6 12.2 11.3 0.5
RMax M1 BL 19 11.0 12.9 11.7 0.5
RMax M2 MD 15 9.4 11.2 10.2 0.5
RMax M2 BL 16 10.6 12.8 11.6 0.7
RMax M3 MD 12 7.9 10.4 9.3 0.8
RMax M3 BL 11 10.1 12.8 11.3 0.9
LMand I1 MD 10 4.8 6.2 5.5 0.4
LMand I1 BL 10 5.5 6.7 6.2 0.4
LMand I2 MD 9 5.4 7.0 6.3 0.5
LMand I2 BL 10 6.4 7.4 6.9 0.3
LMand C MD 15 6.9 8.2 7.6 0.4
LMand C BL 16 7.5 9.6 8.2 0.6
LMand P3 MD 17 7.2 8.6 7.9 0.5
LMand P3 BL 18 8.0 9.8 8.9 0.5
LMand P4 MD 19 7.2 8.9 8.1 0.5
LMand P4 BL 18 8.4 10.2 9.1 0.5
LMand M1 MD 17 11.8 13.4 12.7 0.5
LMand M1 BL 17 10.1 12.1 11.0 0.5
LMand M2 MD 18 10.5 13.8 12.1 0.8
LMand M2 BL 19 10.0 11.8 10.7 0.6
LMand M3 MD 10 8.3 12.8 10.7 1.3
LMand M3 BL 10 9.1 10.8 10.2 0.6
RMand I1 MD 7 4.8 6.0 5.5 0.4
RMand I1 BL 8 5.6 6.8 6.2 0.4
RMand I2 MD 11 5.8 6.9 6.4 0.4
RMand I2 BL 13 5.9 7.4 6.7 0.4
RMand C MD 16 6.9 8.1 7.5 0.4
RMand C BL 16 7.6 9.1 8.4 0.5
RMand P3 MD 17 6.8 8.8 7.9 0.6
RMand P3 BL 16 7.8 10.3 9.0 0.7
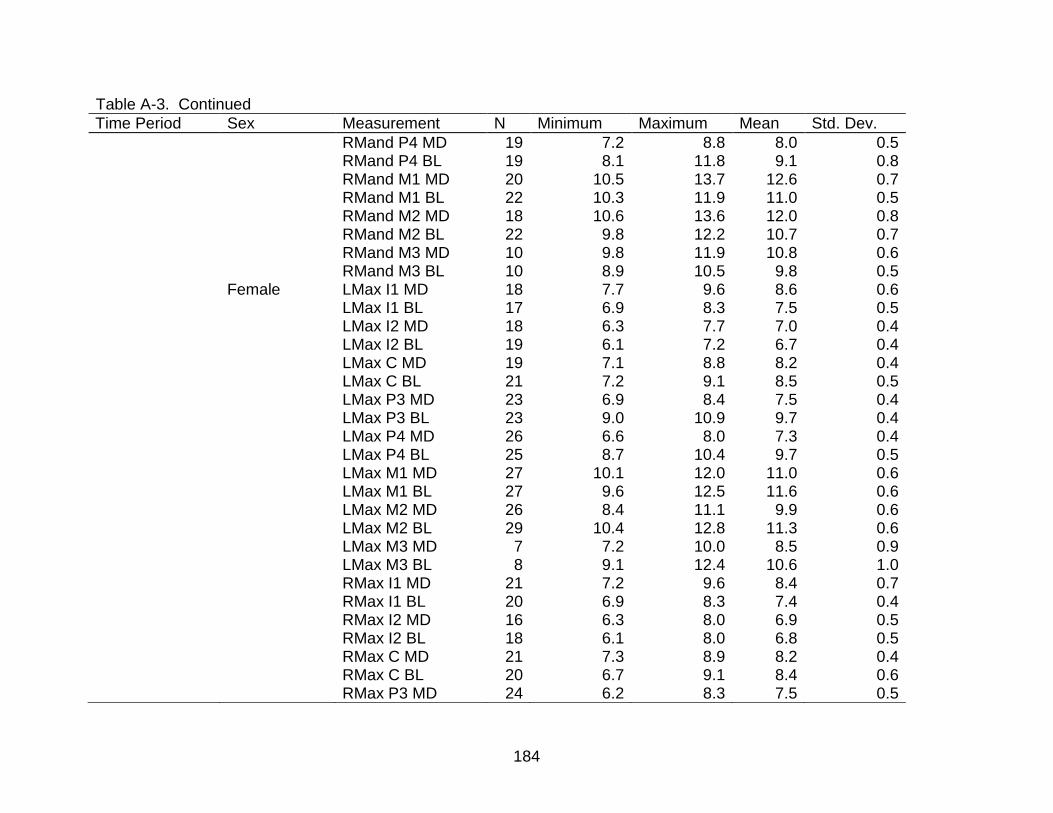
184
Table A-3. Continued
Time Period Sex Measurement N Minimum Maximum Mean Std. Dev.
RMand P4 MD 19 7.2 8.8 8.0 0.5
RMand P4 BL 19 8.1 11.8 9.1 0.8
RMand M1 MD 20 10.5 13.7 12.6 0.7
RMand M1 BL 22 10.3 11.9 11.0 0.5
RMand M2 MD 18 10.6 13.6 12.0 0.8
RMand M2 BL 22 9.8 12.2 10.7 0.7
RMand M3 MD 10 9.8 11.9 10.8 0.6
RMand M3 BL 10 8.9 10.5 9.8 0.5
Female LMax I1 MD 18 7.7 9.6 8.6 0.6
LMax I1 BL 17 6.9 8.3 7.5 0.5
LMax I2 MD 18 6.3 7.7 7.0 0.4
LMax I2 BL 19 6.1 7.2 6.7 0.4
LMax C MD 19 7.1 8.8 8.2 0.4
LMax C BL 21 7.2 9.1 8.5 0.5
LMax P3 MD 23 6.9 8.4 7.5 0.4
LMax P3 BL 23 9.0 10.9 9.7 0.4
LMax P4 MD 26 6.6 8.0 7.3 0.4
LMax P4 BL 25 8.7 10.4 9.7 0.5
LMax M1 MD 27 10.1 12.0 11.0 0.6
LMax M1 BL 27 9.6 12.5 11.6 0.6
LMax M2 MD 26 8.4 11.1 9.9 0.6
LMax M2 BL 29 10.4 12.8 11.3 0.6
LMax M3 MD 7 7.2 10.0 8.5 0.9
LMax M3 BL 8 9.1 12.4 10.6 1.0
RMax I1 MD 21 7.2 9.6 8.4 0.7
RMax I1 BL 20 6.9 8.3 7.4 0.4
RMax I2 MD 16 6.3 8.0 6.9 0.5
RMax I2 BL 18 6.1 8.0 6.8 0.5
RMax C MD 21 7.3 8.9 8.2 0.4
RMax C BL 20 6.7 9.1 8.4 0.6
RMax P3 MD 24 6.2 8.3 7.5 0.5

185
Table A-3. Continued
Time Period Sex Measurement N Minimum Maximum Mean Std. Dev.
RMax P3 BL 24 8.5 10.6 9.8 0.6
RMax P4 MD 29 6.2 8.0 7.2 0.4
RMax P4 BL 28 8.5 10.5 9.5 0.5
RMax M1 MD 23 10.1 12.0 11.1 0.5
RMax M1 BL 26 10.4 12.3 11.4 0.5
RMax M2 MD 27 7.6 10.7 9.8 0.6
RMax M2 BL 29 10.4 12.4 11.2 0.6
RMax M3 MD 9 7.0 10.1 9.0 1.0
RMax M3 BL 9 9.6 12.4 10.9 0.8
LMand I1 MD 14 4.1 6.0 5.4 0.5
LMand I1 BL 13 5.4 6.5 6.0 0.3
LMand I2 MD 19 4.9 6.9 6.1 0.5
LMand I2 BL 18 6.0 7.1 6.5 0.3
LMand C MD 19 6.0 7.3 6.9 0.3
LMand C BL 20 7.0 8.2 7.6 0.3
LMand P3 MD 21 6.5 7.9 7.4 0.4
LMand P3 BL 20 7.4 9.2 8.5 0.5
LMand P4 MD 26 6.9 8.3 7.6 0.4
LMand P4 BL 22 7.7 9.6 8.6 0.5
LMand M1 MD 18 11.2 13.1 12.1 0.5
LMand M1 BL 21 9.4 11.3 10.4 0.4
LMand M2 MD 25 10.1 12.8 11.5 0.7
LMand M2 BL 26 6.7 11.1 10.0 0.9
LMand M3 MD 16 8.7 12.0 10.3 0.9
LMand M3 BL 17 9.0 10.6 9.7 0.5
RMand I1 MD 11 5.0 5.9 5.5 0.3
RMand I1 BL 9 5.8 6.5 6.1 0.3
RMand I2 MD 15 5.5 6.8 6.2 0.4
RMand I2 BL 15 6.2 7.1 6.6 0.3
RMand C MD 17 6.2 7.4 6.9 0.3
RMand C BL 18 7.0 8.2 7.7 0.3

186
Table A-3. Continued
Time Period Sex Measurement N Minimum Maximum Mean Std. Dev.
RMand P3 MD 22 6.7 7.8 7.3 0.3
RMand P3 BL 22 7.6 9.1 8.5 0.4
RMand P4 MD 23 6.6 8.2 7.5 0.4
RMand P4 BL 21 7.3 9.4 8.5 0.6
RMand M1 MD 21 11.2 13.0 12.1 0.5
RMand M1 BL 23 9.6 11.5 10.5 0.5
RMand M2 MD 26 7.9 12.7 11.3 1.0
RMand M2 BL 26 9.3 11.1 10.2 0.5
RMand M3 MD 11 8.8 11.5 10.3 0.7
RMand M3 BL 11 8.4 10.8 9.6 0.6
Indet. LMax I1 MD 15 7.7 9.4 8.5 0.5
LMax I1 BL 16 7.0 8.3 7.7 0.3
LMax I2 MD 17 6.2 8.3 7.1 0.5
LMax I2 BL 16 6.4 7.9 7.2 0.4
LMax C MD 18 7.6 9.6 8.6 0.5
LMax C BL 22 7.9 9.9 8.9 0.5
LMax P3 MD 22 7.2 8.7 7.7 0.4
LMax P3 BL 21 9.1 10.7 10.0 0.4
LMax P4 MD 20 6.5 8.2 7.3 0.4
LMax P4 BL 19 8.8 10.4 9.7 0.4
LMax M1 MD 21 9.9 12.2 11.0 0.6
LMax M1 BL 22 10.6 13.0 11.6 0.6
LMax M2 MD 18 9.3 10.9 10.1 0.4
LMax M2 BL 19 10.3 12.9 11.4 0.7
LMax M3 MD 3 7.9 10.0 9.1 1.1
LMax M3 BL 3 10.9 12.5 11.5 0.8
RMax I1 MD 18 7.7 9.7 8.7 0.6
RMax I1 BL 18 7.0 8.5 7.6 0.4
RMax I2 MD 20 6.1 8.0 7.0 0.5
RMax I2 BL 17 6.0 7.7 7.0 0.4
RMax C MD 17 7.6 9.6 8.5 0.5
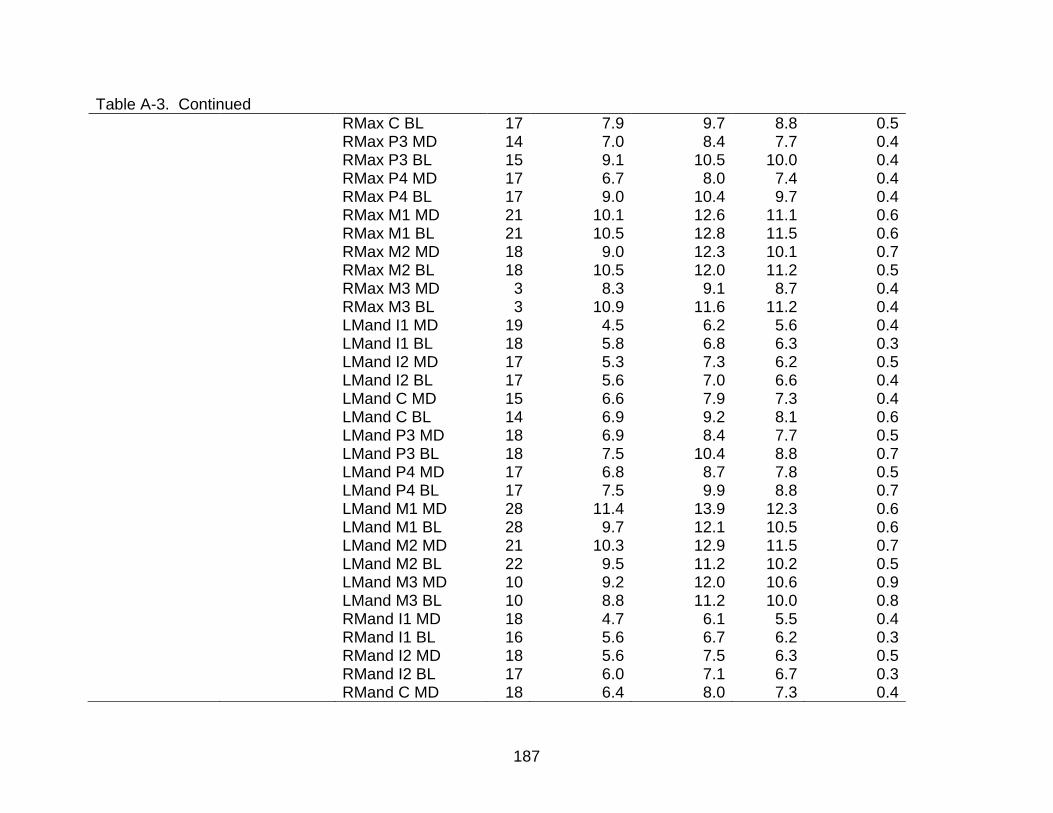
187
Table A-3. Continued
RMax C BL 17 7.9 9.7 8.8 0.5
RMax P3 MD 14 7.0 8.4 7.7 0.4
RMax P3 BL 15 9.1 10.5 10.0 0.4
RMax P4 MD 17 6.7 8.0 7.4 0.4
RMax P4 BL 17 9.0 10.4 9.7 0.4
RMax M1 MD 21 10.1 12.6 11.1 0.6
RMax M1 BL 21 10.5 12.8 11.5 0.6
RMax M2 MD 18 9.0 12.3 10.1 0.7
RMax M2 BL 18 10.5 12.0 11.2 0.5
RMax M3 MD 3 8.3 9.1 8.7 0.4
RMax M3 BL 3 10.9 11.6 11.2 0.4
LMand I1 MD 19 4.5 6.2 5.6 0.4
LMand I1 BL 18 5.8 6.8 6.3 0.3
LMand I2 MD 17 5.3 7.3 6.2 0.5
LMand I2 BL 17 5.6 7.0 6.6 0.4
LMand C MD 15 6.6 7.9 7.3 0.4
LMand C BL 14 6.9 9.2 8.1 0.6
LMand P3 MD 18 6.9 8.4 7.7 0.5
LMand P3 BL 18 7.5 10.4 8.8 0.7
LMand P4 MD 17 6.8 8.7 7.8 0.5
LMand P4 BL 17 7.5 9.9 8.8 0.7
LMand M1 MD 28 11.4 13.9 12.3 0.6
LMand M1 BL 28 9.7 12.1 10.5 0.6
LMand M2 MD 21 10.3 12.9 11.5 0.7
LMand M2 BL 22 9.5 11.2 10.2 0.5
LMand M3 MD 10 9.2 12.0 10.6 0.9
LMand M3 BL 10 8.8 11.2 10.0 0.8
RMand I1 MD 18 4.7 6.1 5.5 0.4
RMand I1 BL 16 5.6 6.7 6.2 0.3
RMand I2 MD 18 5.6 7.5 6.3 0.5
RMand I2 BL 17 6.0 7.1 6.7 0.3
RMand C MD 18 6.4 8.0 7.3 0.4

188
Table A-3. Continued
Time Period Sex Measurement N Minimum Maximum Mean Std. Dev.
RMand C BL 16 7.7 9.0 8.3 0.4
RMand P3 MD 18 6.9 8.5 7.8 0.5
RMand P3 BL 17 7.8 10.3 8.8 0.7
RMand P4 MD 19 7.1 8.7 7.8 0.4
RMand P4 BL 19 7.4 10.2 8.9 0.6
RMand M1 MD 30 11.3 13.8 12.3 0.6
RMand M1 BL 32 9.6 12.1 10.7 0.6
RMand M2 MD 21 10.3 13.3 11.7 0.7
RMand M2 BL 21 9.2 11.5 10.3 0.6
RMand M3 MD 7 9.6 11.5 10.8 0.6
RMand M3 BL 8 8.5 10.7 9.8 0.8

189
Table A-4. Descriptive statistics of cross-sectional area by time period
Time Period Measurement N Minimum Maximum Mean Std. Dev.
Pre-Latte AVG Max I1 CX 38 61.6 81.7 71.0 5.9
AVG Max I2 CX 34 42.9 66.9 52.6 5.4
AVG Max C CX 37 61.6 93.0 74.1 6.4
AVG Max P3 CX 45 70.3 103.2 84.2 7.4
AVG Max P4 CX 50 64.5 102.7 80.3 8.4
AVG Max M1 CX 43 124.6 162.3 141.1 9.8
AVG Max M2 CX 47 111.4 157.2 131.3 10.4
AVG Max M3 CX 13 86.4 120.5 105.1 10.4
AVG Mand I1 CX 36 31.7 45.2 37.6 3.5
AVG Mand I2 CX 39 36.0 55.2 44.6 4.0
AVG Mand C CX 49 43.8 77.4 59.9 7.1
AVG Mand P3 CX 58 54.0 88.2 72.1 7.0
AVG Mand P4 CX 52 60.2 95.6 74.3 6.7
AVG Mand M1 CX 45 120.6 165.2 137.3 10.0
AVG Mand M2 CX 32 106.6 155.6 131.8 11.2
AVG Mand M3 CX 15 101.8 171.9 125.9 17.7
Latte AVG Max I1 CX 35 54.5 80.6 66.3 7.4
AVG Max I2 CX 32 39.2 63.3 49.4 5.5
AVG Max C CX 36 56.8 92.4 74.8 7.6
AVG Max P3 CX 47 61.1 98.9 76.3 7.6
AVG Max P4 CX 49 55.3 88.6 72.5 7.0
AVG Max M1 CX 52 106.8 157.6 129.3 11.9
AVG Max M2 CX 48 87.5 139.2 114.0 11.5
AVG Max M3 CX 11 73.2 127.1 101.2 17.1
AVG Mand I1 CX 26 26.6 40.9 33.6 3.8
AVG Mand I2 CX 31 34.0 51.6 42.0 4.0
AVG Mand C CX 33 44.7 72.3 58.2 7.5
AVG Mand P3 CX 45 53.8 87.5 66.6 8.1
AVG Mand P4 CX 44 52.8 87.5 68.4 7.8
AVG Mand M1 CX 53 111.6 164.3 131.4 12.1
AVG Mand M2 CX 50 94.6 162.8 119.7 14.7
AVG Mand M3 CX 17 85.4 123.3 103.3 10.9
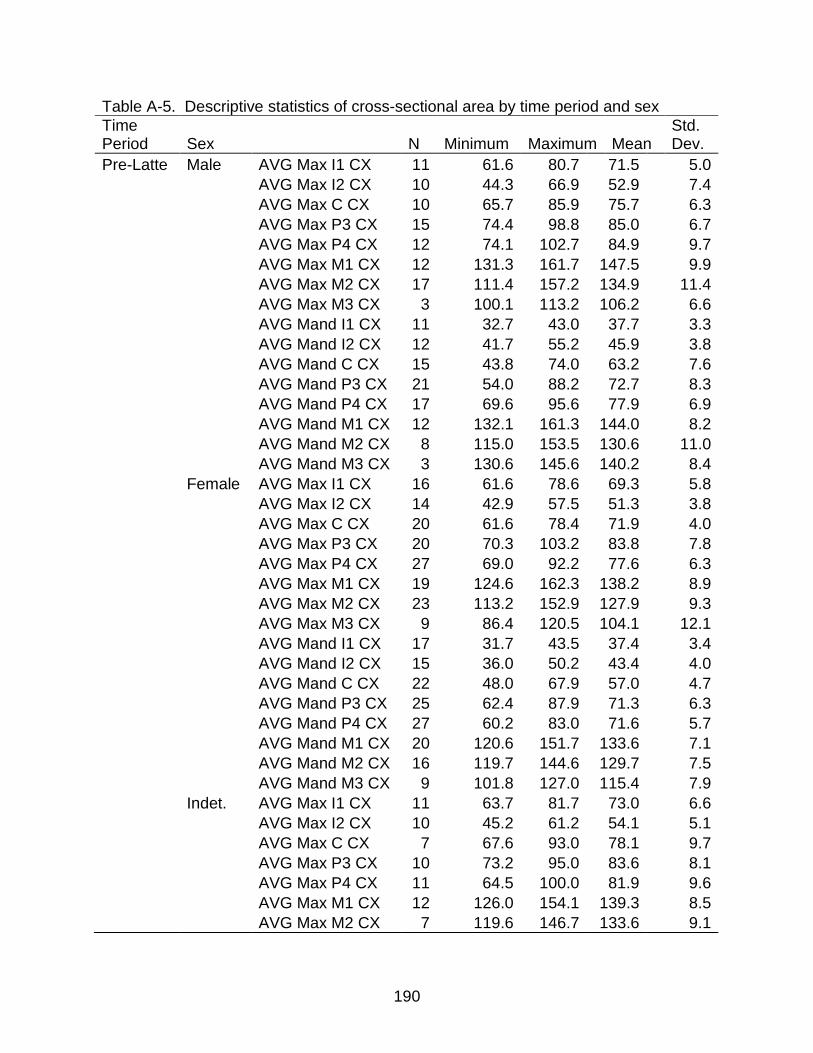
190
Table A-5. Descriptive statistics of cross-sectional area by time period and sex
Time Period Sex N Minimum Maximum Mean
Std. Dev.
Pre-Latte Male AVG Max I1 CX 11 61.6 80.7 71.5 5.0
AVG Max I2 CX 10 44.3 66.9 52.9 7.4
AVG Max C CX 10 65.7 85.9 75.7 6.3
AVG Max P3 CX 15 74.4 98.8 85.0 6.7
AVG Max P4 CX 12 74.1 102.7 84.9 9.7
AVG Max M1 CX 12 131.3 161.7 147.5 9.9
AVG Max M2 CX 17 111.4 157.2 134.9 11.4
AVG Max M3 CX 3 100.1 113.2 106.2 6.6
AVG Mand I1 CX 11 32.7 43.0 37.7 3.3
AVG Mand I2 CX 12 41.7 55.2 45.9 3.8
AVG Mand C CX 15 43.8 74.0 63.2 7.6
AVG Mand P3 CX 21 54.0 88.2 72.7 8.3
AVG Mand P4 CX 17 69.6 95.6 77.9 6.9
AVG Mand M1 CX 12 132.1 161.3 144.0 8.2
AVG Mand M2 CX 8 115.0 153.5 130.6 11.0
AVG Mand M3 CX 3 130.6 145.6 140.2 8.4
Female AVG Max I1 CX 16 61.6 78.6 69.3 5.8
AVG Max I2 CX 14 42.9 57.5 51.3 3.8
AVG Max C CX 20 61.6 78.4 71.9 4.0
AVG Max P3 CX 20 70.3 103.2 83.8 7.8
AVG Max P4 CX 27 69.0 92.2 77.6 6.3
AVG Max M1 CX 19 124.6 162.3 138.2 8.9
AVG Max M2 CX 23 113.2 152.9 127.9 9.3
AVG Max M3 CX 9 86.4 120.5 104.1 12.1
AVG Mand I1 CX 17 31.7 43.5 37.4 3.4
AVG Mand I2 CX 15 36.0 50.2 43.4 4.0
AVG Mand C CX 22 48.0 67.9 57.0 4.7
AVG Mand P3 CX 25 62.4 87.9 71.3 6.3
AVG Mand P4 CX 27 60.2 83.0 71.6 5.7
AVG Mand M1 CX 20 120.6 151.7 133.6 7.1
AVG Mand M2 CX 16 119.7 144.6 129.7 7.5
AVG Mand M3 CX 9 101.8 127.0 115.4 7.9
Indet. AVG Max I1 CX 11 63.7 81.7 73.0 6.6
AVG Max I2 CX 10 45.2 61.2 54.1 5.1
AVG Max C CX 7 67.6 93.0 78.1 9.7
AVG Max P3 CX 10 73.2 95.0 83.6 8.1
AVG Max P4 CX 11 64.5 100.0 81.9 9.6
AVG Max M1 CX 12 126.0 154.1 139.3 8.5
AVG Max M2 CX 7 119.6 146.7 133.6 9.1
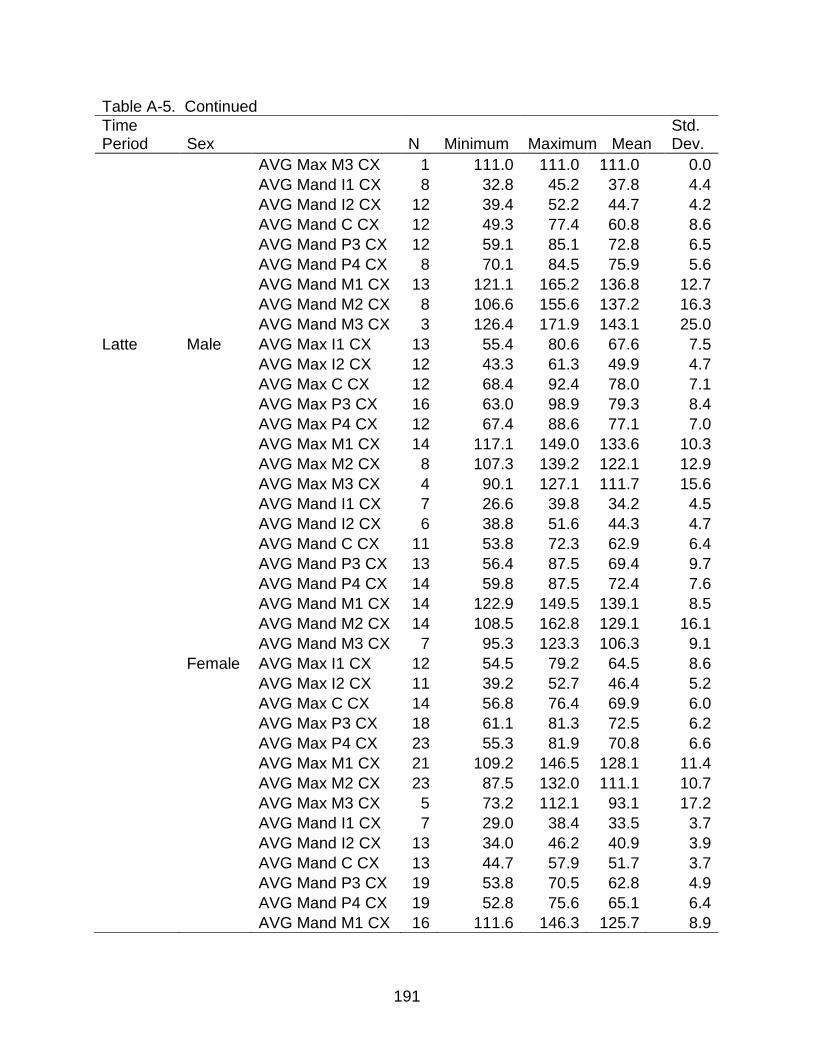
191
Table A-5. Continued
Time Period Sex N Minimum Maximum Mean
Std. Dev.
AVG Max M3 CX 1 111.0 111.0 111.0 0.0
AVG Mand I1 CX 8 32.8 45.2 37.8 4.4
AVG Mand I2 CX 12 39.4 52.2 44.7 4.2
AVG Mand C CX 12 49.3 77.4 60.8 8.6
AVG Mand P3 CX 12 59.1 85.1 72.8 6.5
AVG Mand P4 CX 8 70.1 84.5 75.9 5.6
AVG Mand M1 CX 13 121.1 165.2 136.8 12.7
AVG Mand M2 CX 8 106.6 155.6 137.2 16.3
AVG Mand M3 CX 3 126.4 171.9 143.1 25.0
Latte Male AVG Max I1 CX 13 55.4 80.6 67.6 7.5
AVG Max I2 CX 12 43.3 61.3 49.9 4.7
AVG Max C CX 12 68.4 92.4 78.0 7.1
AVG Max P3 CX 16 63.0 98.9 79.3 8.4
AVG Max P4 CX 12 67.4 88.6 77.1 7.0
AVG Max M1 CX 14 117.1 149.0 133.6 10.3
AVG Max M2 CX 8 107.3 139.2 122.1 12.9
AVG Max M3 CX 4 90.1 127.1 111.7 15.6
AVG Mand I1 CX 7 26.6 39.8 34.2 4.5
AVG Mand I2 CX 6 38.8 51.6 44.3 4.7
AVG Mand C CX 11 53.8 72.3 62.9 6.4
AVG Mand P3 CX 13 56.4 87.5 69.4 9.7
AVG Mand P4 CX 14 59.8 87.5 72.4 7.6
AVG Mand M1 CX 14 122.9 149.5 139.1 8.5
AVG Mand M2 CX 14 108.5 162.8 129.1 16.1
AVG Mand M3 CX 7 95.3 123.3 106.3 9.1
Female AVG Max I1 CX 12 54.5 79.2 64.5 8.6
AVG Max I2 CX 11 39.2 52.7 46.4 5.2
AVG Max C CX 14 56.8 76.4 69.9 6.0
AVG Max P3 CX 18 61.1 81.3 72.5 6.2
AVG Max P4 CX 23 55.3 81.9 70.8 6.6
AVG Max M1 CX 21 109.2 146.5 128.1 11.4
AVG Max M2 CX 23 87.5 132.0 111.1 10.7
AVG Max M3 CX 5 73.2 112.1 93.1 17.2
AVG Mand I1 CX 7 29.0 38.4 33.5 3.7
AVG Mand I2 CX 13 34.0 46.2 40.9 3.9
AVG Mand C CX 13 44.7 57.9 51.7 3.7
AVG Mand P3 CX 19 53.8 70.5 62.8 4.9
AVG Mand P4 CX 19 52.8 75.6 65.1 6.4
AVG Mand M1 CX 16 111.6 146.3 125.7 8.9
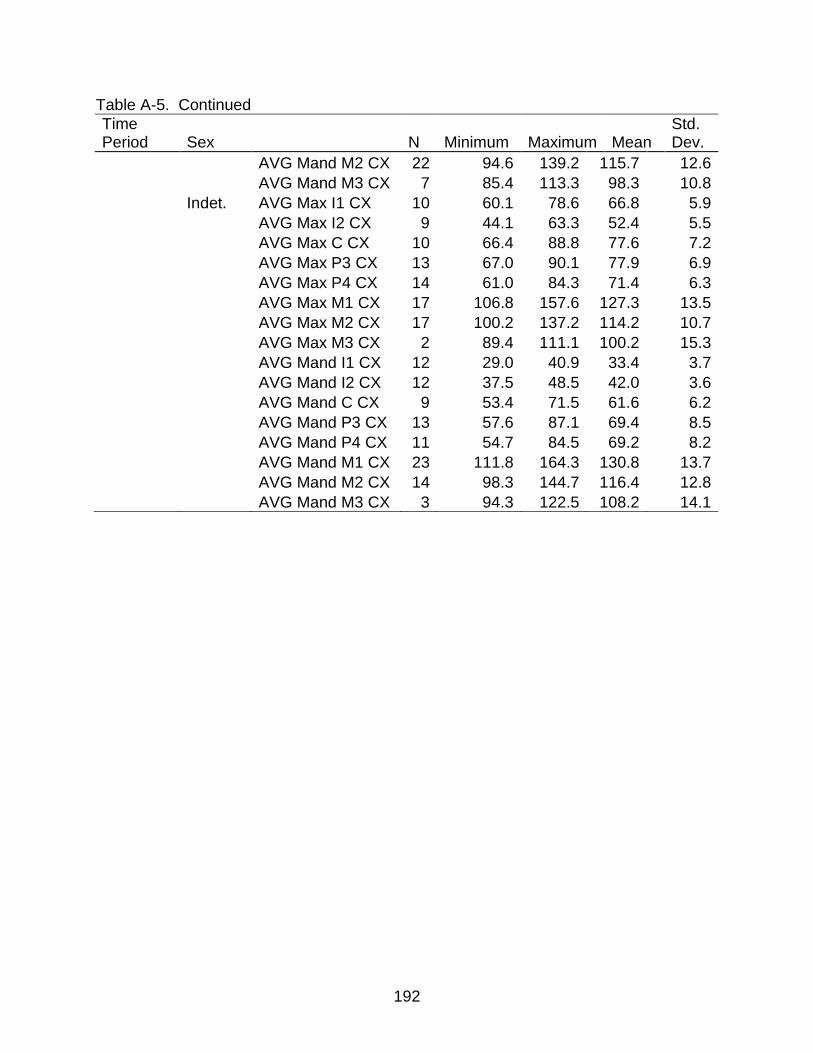
192
Table A-5. Continued
Time Period Sex N Minimum Maximum Mean
Std. Dev.
AVG Mand M2 CX 22 94.6 139.2 115.7 12.6
AVG Mand M3 CX 7 85.4 113.3 98.3 10.8
Indet. AVG Max I1 CX 10 60.1 78.6 66.8 5.9
AVG Max I2 CX 9 44.1 63.3 52.4 5.5
AVG Max C CX 10 66.4 88.8 77.6 7.2
AVG Max P3 CX 13 67.0 90.1 77.9 6.9
AVG Max P4 CX 14 61.0 84.3 71.4 6.3
AVG Max M1 CX 17 106.8 157.6 127.3 13.5
AVG Max M2 CX 17 100.2 137.2 114.2 10.7
AVG Max M3 CX 2 89.4 111.1 100.2 15.3
AVG Mand I1 CX 12 29.0 40.9 33.4 3.7
AVG Mand I2 CX 12 37.5 48.5 42.0 3.6
AVG Mand C CX 9 53.4 71.5 61.6 6.2
AVG Mand P3 CX 13 57.6 87.1 69.4 8.5
AVG Mand P4 CX 11 54.7 84.5 69.2 8.2
AVG Mand M1 CX 23 111.8 164.3 130.8 13.7
AVG Mand M2 CX 14 98.3 144.7 116.4 12.8
AVG Mand M3 CX 3 94.3 122.5 108.2 14.1
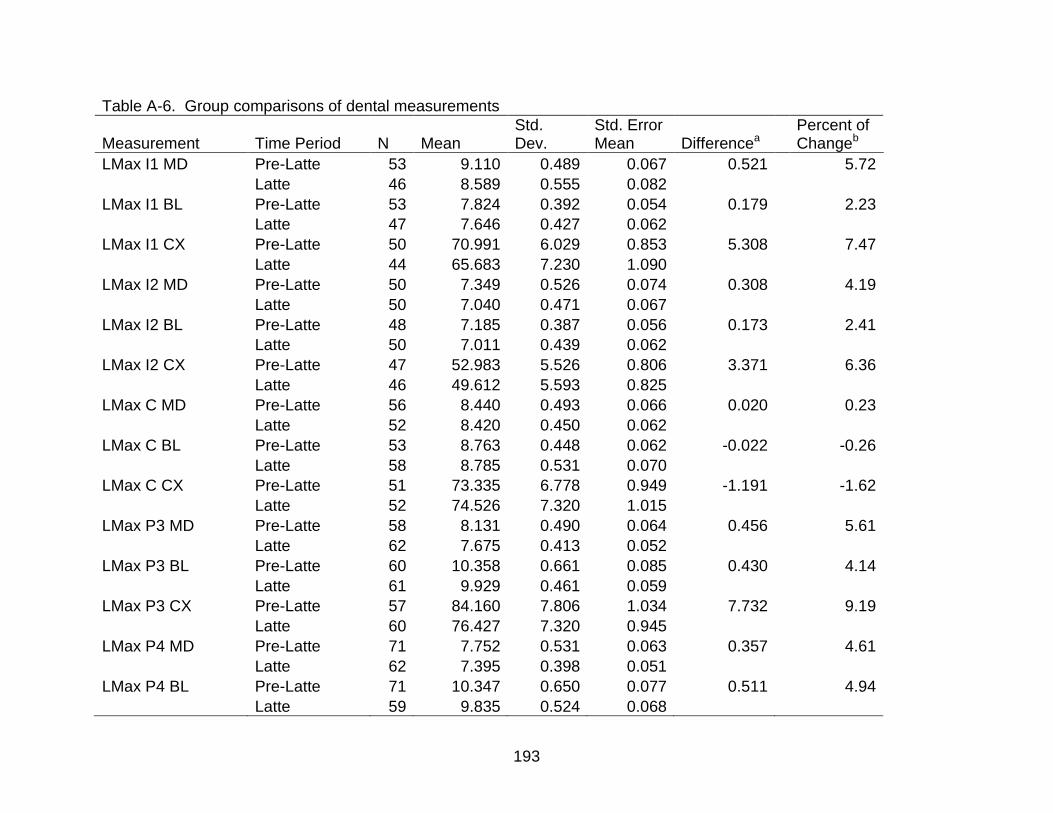
193
Table A-6. Group comparisons of dental measurements
Measurement Time Period N Mean Std. Dev.
Std. Error Mean Differencea
Percent of Changeb
LMax I1 MD Pre-Latte 53 9.110 0.489 0.067 0.521 5.72
Latte 46 8.589 0.555 0.082
LMax I1 BL Pre-Latte 53 7.824 0.392 0.054 0.179 2.23
Latte 47 7.646 0.427 0.062
LMax I1 CX Pre-Latte 50 70.991 6.029 0.853 5.308 7.47
Latte 44 65.683 7.230 1.090
LMax I2 MD Pre-Latte 50 7.349 0.526 0.074 0.308 4.19
Latte 50 7.040 0.471 0.067
LMax I2 BL Pre-Latte 48 7.185 0.387 0.056 0.173 2.41
Latte 50 7.011 0.439 0.062
LMax I2 CX Pre-Latte 47 52.983 5.526 0.806 3.371 6.36
Latte 46 49.612 5.593 0.825
LMax C MD Pre-Latte 56 8.440 0.493 0.066 0.020 0.23
Latte 52 8.420 0.450 0.062
LMax C BL Pre-Latte 53 8.763 0.448 0.062 -0.022 -0.26
Latte 58 8.785 0.531 0.070
LMax C CX Pre-Latte 51 73.335 6.778 0.949 -1.191 -1.62
Latte 52 74.526 7.320 1.015
LMax P3 MD Pre-Latte 58 8.131 0.490 0.064 0.456 5.61
Latte 62 7.675 0.413 0.052
LMax P3 BL Pre-Latte 60 10.358 0.661 0.085 0.430 4.14
Latte 61 9.929 0.461 0.059
LMax P3 CX Pre-Latte 57 84.160 7.806 1.034 7.732 9.19
Latte 60 76.427 7.320 0.945
LMax P4 MD Pre-Latte 71 7.752 0.531 0.063 0.357 4.61
Latte 62 7.395 0.398 0.051
LMax P4 BL Pre-Latte 71 10.347 0.650 0.077 0.511 4.94
Latte 59 9.835 0.524 0.068

194
Table A-6. Continued
Measurement Time Period N Mean Std. Dev.
Std. Error Mean Differencea
Percent of Changeb
LMax P4 CX Pre-Latte 67 80.442 8.136 0.994 7.553 9.39
Latte 59 72.889 7.251 0.944
LMax M1 MD Pre-Latte 56 11.513 0.502 0.067 0.464 4.03
Latte 63 11.049 0.575 0.072
LMax M1 BL Pre-Latte 63 12.191 0.519 0.065 0.504 4.13
Latte 64 11.688 0.597 0.075
LMax M1 CX Pre-Latte 55 140.416 10.928 1.474 11.066 7.88
Latte 62 129.350 12.511 1.589
LMax M2 MD Pre-Latte 64 10.875 0.568 0.071 0.819 7.53
Latte 54 10.055 0.559 0.076
LMax M2 BL Pre-Latte 63 12.316 0.719 0.091 0.900 7.31
Latte 61 11.416 0.661 0.085
LMax M2 CX Pre-Latte 62 133.699 12.378 1.572 18.631 13.94
Latte 54 115.068 11.628 1.582
LMax M3 MD Pre-Latte 29 9.609 0.933 0.173 0.471 4.60
Latte 18 9.138 1.083 0.255
LMax M3 BL Pre-Latte 29 11.629 0.678 0.126 0.506 4.35
Latte 21 11.122 0.919 0.200
LMax M3 CX Pre-Latte 29 111.867 13.421 2.492 8.727 7.80
Latte 18 103.139 17.202 4.054
RMax I1 MD Pre-Latte 48 9.036 0.433 0.063 0.446 4.94
Latte 54 8.589 0.614 0.084
RMax I1 BL Pre-Latte 53 7.753 0.430 0.059 0.172 2.21
Latte 54 7.581 0.407 0.055
RMax I1 CX Pre-Latte 44 70.210 5.723 0.863 4.908 6.99
Latte 51 65.302 7.722 1.081
RMax I2 MD Pre-Latte 47 7.404 0.458 0.067 0.427 5.76
Latte 49 6.977 0.481
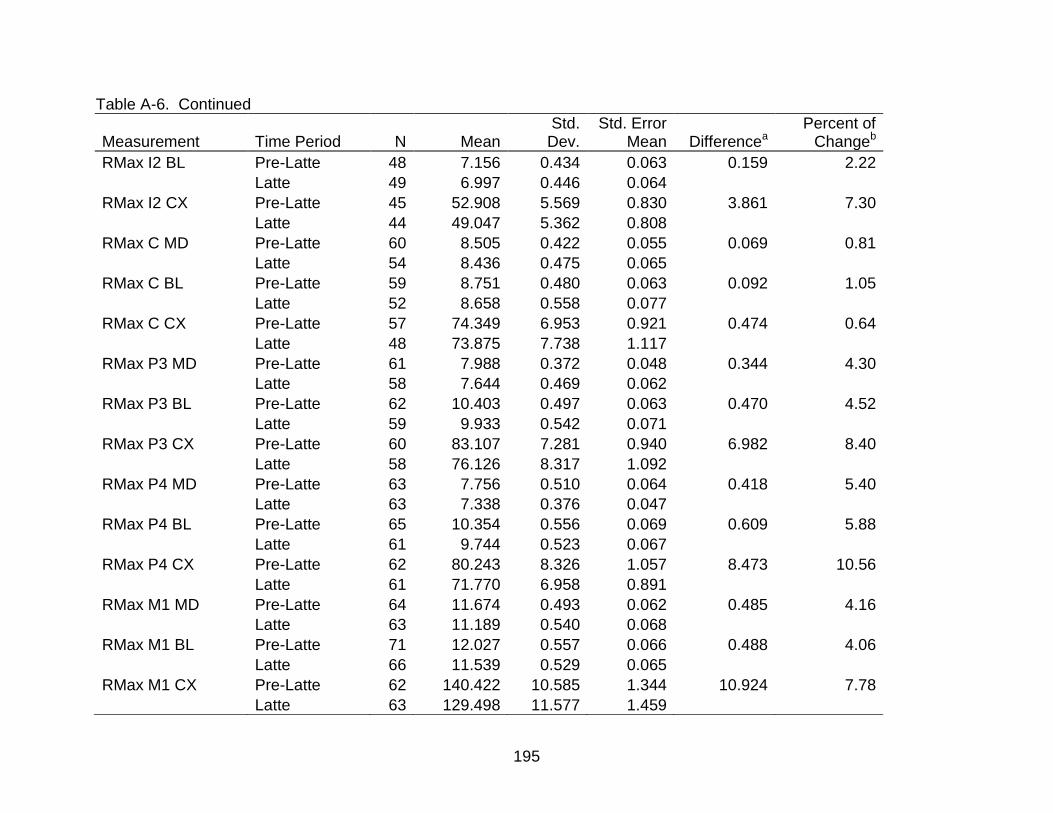
195
Table A-6. Continued
Measurement Time Period N Mean Std. Dev.
Std. Error Mean Differencea
Percent of Changeb
RMax I2 BL Pre-Latte 48 7.156 0.434 0.063 0.159 2.22
Latte 49 6.997 0.446 0.064
RMax I2 CX Pre-Latte 45 52.908 5.569 0.830 3.861 7.30
Latte 44 49.047 5.362 0.808
RMax C MD Pre-Latte 60 8.505 0.422 0.055 0.069 0.81
Latte 54 8.436 0.475 0.065
RMax C BL Pre-Latte 59 8.751 0.480 0.063 0.092 1.05
Latte 52 8.658 0.558 0.077
RMax C CX Pre-Latte 57 74.349 6.953 0.921 0.474 0.64
Latte 48 73.875 7.738 1.117
RMax P3 MD Pre-Latte 61 7.988 0.372 0.048 0.344 4.30
Latte 58 7.644 0.469 0.062
RMax P3 BL Pre-Latte 62 10.403 0.497 0.063 0.470 4.52
Latte 59 9.933 0.542 0.071
RMax P3 CX Pre-Latte 60 83.107 7.281 0.940 6.982 8.40
Latte 58 76.126 8.317 1.092
RMax P4 MD Pre-Latte 63 7.756 0.510 0.064 0.418 5.40
Latte 63 7.338 0.376 0.047
RMax P4 BL Pre-Latte 65 10.354 0.556 0.069 0.609 5.88
Latte 61 9.744 0.523 0.067
RMax P4 CX Pre-Latte 62 80.243 8.326 1.057 8.473 10.56
Latte 61 71.770 6.958 0.891
RMax M1 MD Pre-Latte 64 11.674 0.493 0.062 0.485 4.16
Latte 63 11.189 0.540 0.068
RMax M1 BL Pre-Latte 71 12.027 0.557 0.066 0.488 4.06
Latte 66 11.539 0.529 0.065
RMax M1 CX Pre-Latte 62 140.422 10.585 1.344 10.924 7.78
Latte 63 129.498 11.577 1.459
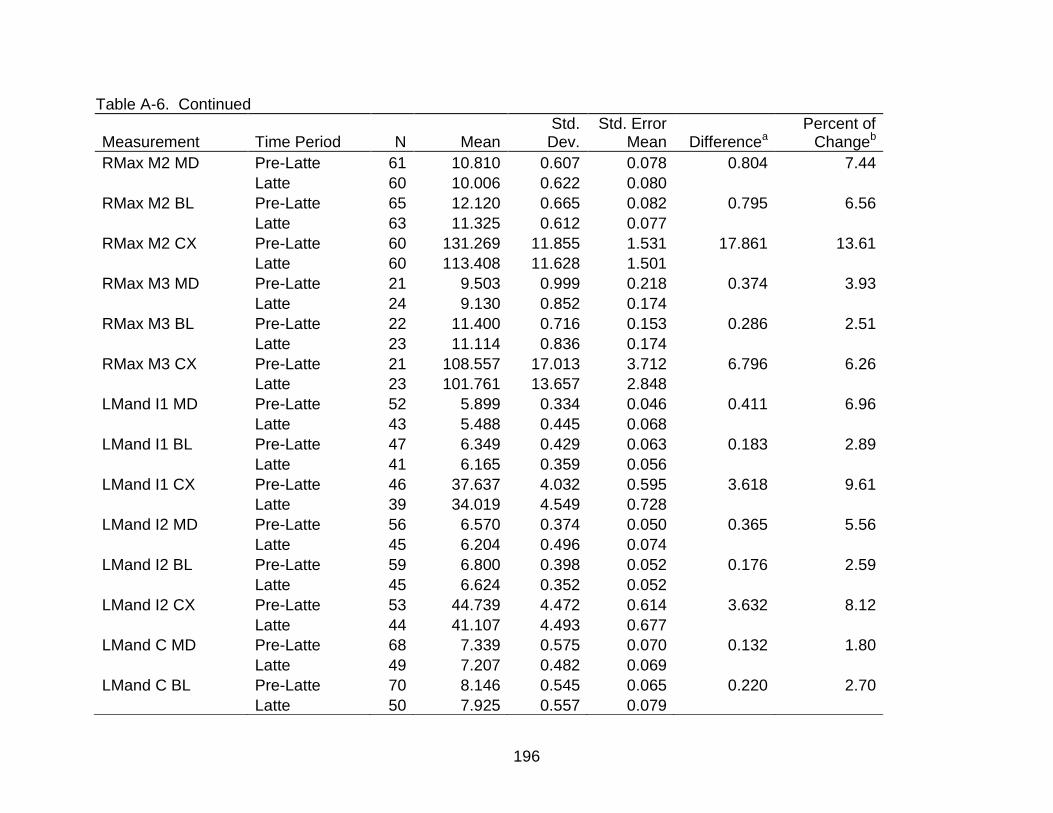
196
Table A-6. Continued
Measurement Time Period N Mean Std. Dev.
Std. Error Mean Differencea
Percent of Changeb
RMax M2 MD Pre-Latte 61 10.810 0.607 0.078 0.804 7.44
Latte 60 10.006 0.622 0.080
RMax M2 BL Pre-Latte 65 12.120 0.665 0.082 0.795 6.56
Latte 63 11.325 0.612 0.077
RMax M2 CX Pre-Latte 60 131.269 11.855 1.531 17.861 13.61
Latte 60 113.408 11.628 1.501
RMax M3 MD Pre-Latte 21 9.503 0.999 0.218 0.374 3.93
Latte 24 9.130 0.852 0.174
RMax M3 BL Pre-Latte 22 11.400 0.716 0.153 0.286 2.51
Latte 23 11.114 0.836 0.174
RMax M3 CX Pre-Latte 21 108.557 17.013 3.712 6.796 6.26
Latte 23 101.761 13.657 2.848
LMand I1 MD Pre-Latte 52 5.899 0.334 0.046 0.411 6.96
Latte 43 5.488 0.445 0.068
LMand I1 BL Pre-Latte 47 6.349 0.429 0.063 0.183 2.89
Latte 41 6.165 0.359 0.056
LMand I1 CX Pre-Latte 46 37.637 4.032 0.595 3.618 9.61
Latte 39 34.019 4.549 0.728
LMand I2 MD Pre-Latte 56 6.570 0.374 0.050 0.365 5.56
Latte 45 6.204 0.496 0.074
LMand I2 BL Pre-Latte 59 6.800 0.398 0.052 0.176 2.59
Latte 45 6.624 0.352 0.052
LMand I2 CX Pre-Latte 53 44.739 4.472 0.614 3.632 8.12
Latte 44 41.107 4.493 0.677
LMand C MD Pre-Latte 68 7.339 0.575 0.070 0.132 1.80
Latte 49 7.207 0.482 0.069
LMand C BL Pre-Latte 70 8.146 0.545 0.065 0.220 2.70
Latte 50 7.925 0.557 0.079
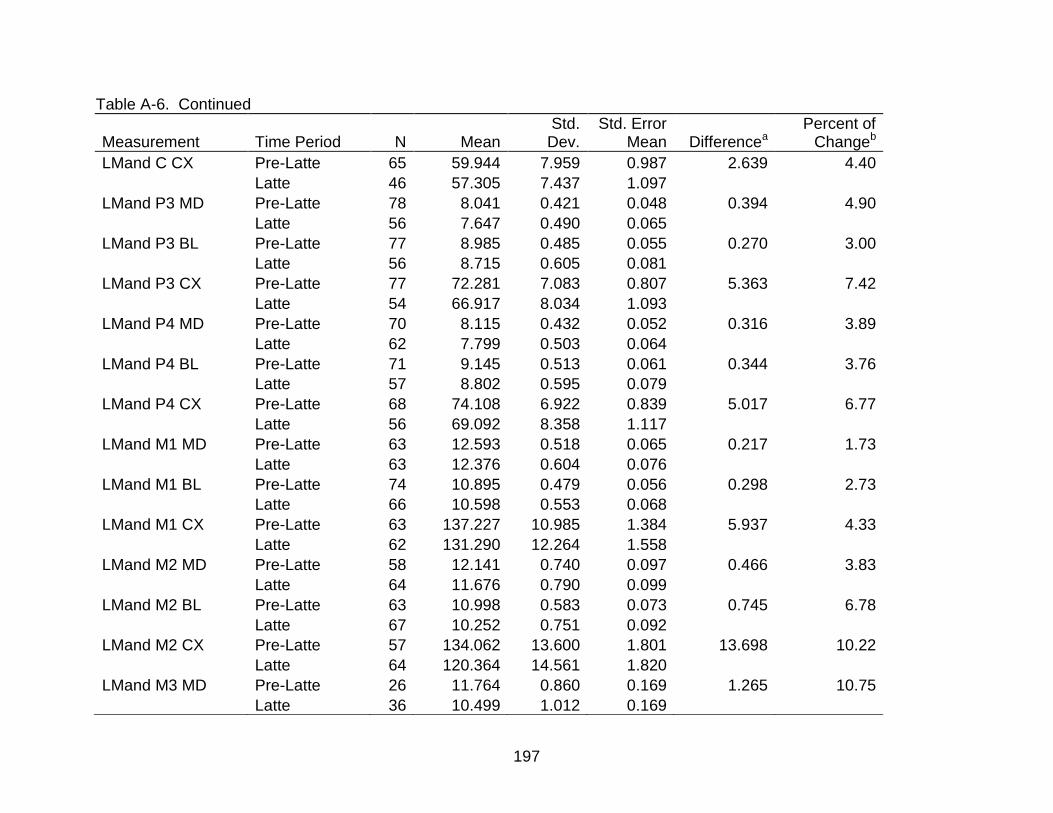
197
Table A-6. Continued
Measurement Time Period N Mean Std. Dev.
Std. Error Mean Differencea
Percent of Changeb
LMand C CX Pre-Latte 65 59.944 7.959 0.987 2.639 4.40
Latte 46 57.305 7.437 1.097
LMand P3 MD Pre-Latte 78 8.041 0.421 0.048 0.394 4.90
Latte 56 7.647 0.490 0.065
LMand P3 BL Pre-Latte 77 8.985 0.485 0.055 0.270 3.00
Latte 56 8.715 0.605 0.081
LMand P3 CX Pre-Latte 77 72.281 7.083 0.807 5.363 7.42
Latte 54 66.917 8.034 1.093
LMand P4 MD Pre-Latte 70 8.115 0.432 0.052 0.316 3.89
Latte 62 7.799 0.503 0.064
LMand P4 BL Pre-Latte 71 9.145 0.513 0.061 0.344 3.76
Latte 57 8.802 0.595 0.079
LMand P4 CX Pre-Latte 68 74.108 6.922 0.839 5.017 6.77
Latte 56 69.092 8.358 1.117
LMand M1 MD Pre-Latte 63 12.593 0.518 0.065 0.217 1.73
Latte 63 12.376 0.604 0.076
LMand M1 BL Pre-Latte 74 10.895 0.479 0.056 0.298 2.73
Latte 66 10.598 0.553 0.068
LMand M1 CX Pre-Latte 63 137.227 10.985 1.384 5.937 4.33
Latte 62 131.290 12.264 1.558
LMand M2 MD Pre-Latte 58 12.141 0.740 0.097 0.466 3.83
Latte 64 11.676 0.790 0.099
LMand M2 BL Pre-Latte 63 10.998 0.583 0.073 0.745 6.78
Latte 67 10.252 0.751 0.092
LMand M2 CX Pre-Latte 57 134.062 13.600 1.801 13.698 10.22
Latte 64 120.364 14.561 1.820
LMand M3 MD Pre-Latte 26 11.764 0.860 0.169 1.265 10.75
Latte 36 10.499 1.012 0.169
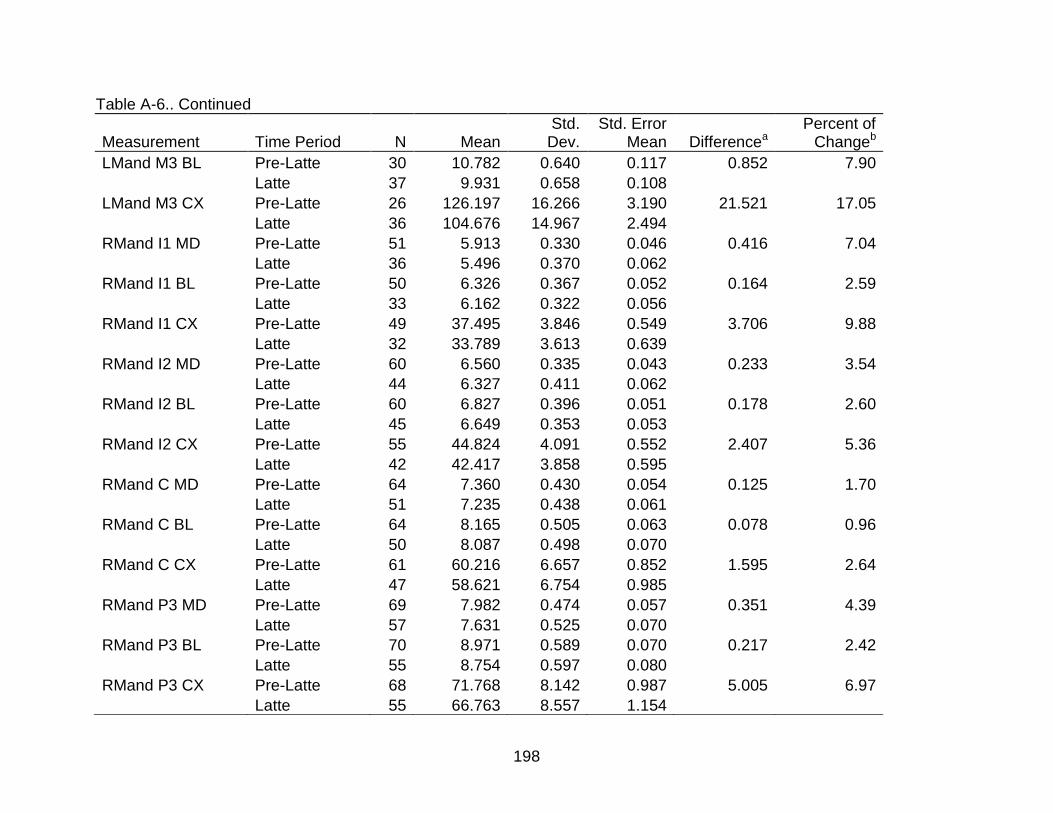
198
Table A-6.. Continued
Measurement Time Period N Mean Std. Dev.
Std. Error Mean Differencea
Percent of Changeb
LMand M3 BL Pre-Latte 30 10.782 0.640 0.117 0.852 7.90
Latte 37 9.931 0.658 0.108
LMand M3 CX Pre-Latte 26 126.197 16.266 3.190 21.521 17.05
Latte 36 104.676 14.967 2.494
RMand I1 MD Pre-Latte 51 5.913 0.330 0.046 0.416 7.04
Latte 36 5.496 0.370 0.062
RMand I1 BL Pre-Latte 50 6.326 0.367 0.052 0.164 2.59
Latte 33 6.162 0.322 0.056
RMand I1 CX Pre-Latte 49 37.495 3.846 0.549 3.706 9.88
Latte 32 33.789 3.613 0.639
RMand I2 MD Pre-Latte 60 6.560 0.335 0.043 0.233 3.54
Latte 44 6.327 0.411 0.062
RMand I2 BL Pre-Latte 60 6.827 0.396 0.051 0.178 2.60
Latte 45 6.649 0.353 0.053
RMand I2 CX Pre-Latte 55 44.824 4.091 0.552 2.407 5.36
Latte 42 42.417 3.858 0.595
RMand C MD Pre-Latte 64 7.360 0.430 0.054 0.125 1.70
Latte 51 7.235 0.438 0.061
RMand C BL Pre-Latte 64 8.165 0.505 0.063 0.078 0.96
Latte 50 8.087 0.498 0.070
RMand C CX Pre-Latte 61 60.216 6.657 0.852 1.595 2.64
Latte 47 58.621 6.754 0.985
RMand P3 MD Pre-Latte 69 7.982 0.474 0.057 0.351 4.39
Latte 57 7.631 0.525 0.070
RMand P3 BL Pre-Latte 70 8.971 0.589 0.070 0.217 2.42
Latte 55 8.754 0.597 0.080
RMand P3 CX Pre-Latte 68 71.768 8.142 0.987 5.005 6.97
Latte 55 66.763 8.557 1.154

199
Table A-6. Continued
Measurement Time Period N Mean Std. Dev.
Std. Error Mean Differencea
Percent of Changeb
RMand P4 MD Pre-Latte 65 8.074 0.411 0.051 0.308 3.81
Latte 61 7.766 0.477 0.061
RMand P4 BL Pre-Latte 74 9.154 0.496 0.058 0.318 3.48
Latte 59 8.836 0.705 0.092
RMand P4 CX Pre-Latte 64 74.145 7.185 0.898 5.331 7.19
Latte 56 68.814 7.829 1.046
RMand M1 MD Pre-Latte 62 12.609 0.508 0.064 0.306 2.43
Latte 71 12.303 0.639 0.076
RMand M1 BL Pre-Latte 68 10.993 0.453 0.055 0.261 2.37
Latte 77 10.732 0.555 0.063
RMand M1 CX Pre-Latte 62 138.419 10.024 1.273 6.794 4.91
Latte 71 131.624 12.390 1.470
RMand M2 MD Pre-Latte 48 12.054 0.675 0.097 0.463 3.84
Latte 65 11.591 0.876 0.109
RMand M2 BL Pre-Latte 58 10.935 0.472 0.062 0.581 5.31
Latte 69 10.355 0.619 0.075
RMand M2 CX Pre-Latte 48 131.708 12.162 1.755 11.882 9.02
Latte 64 119.826 14.788 1.849
RMand M3 MD Pre-Latte 30 11.806 0.700 0.128 1.218 10.32
Latte 28 10.588 0.679 0.128
RMand M3 BL Pre-Latte 35 10.859 0.654 0.111 1.119 10.30
Latte 29 9.740 0.615 0.114
RMand M3 CX Pre-Latte 30 128.416 13.708 2.503 25.477 19.84
Latte 28 102.939 10.877 2.056 a. Calculated as Latte mean subtracted from Pre-Latte mean b. Calculated as mean of Pre-Latte subtracted from the mean of the Latte divided by 100 multiplied by 100

200
Table A-7. Kolmogorov-Smirnova Test for normality
Measurement Time Period n Statistic df Sig.
LMax I1 MD Pre-Latte 53 0.094 53 0.200*
Latte 46 0.086 46 0.200*
LMax I1 BL Pre-Latte 53 0.080 53 0.200*
Latte 47 0.075 47 0.200*
LMax I2 MD Pre-Latte 50 0.066 50 0.200*
Latte 50 0.109 50 0.193
LMax I2 BL Pre-Latte 48 0.109 48 0.200*
Latte 50 0.055 50 0.200*
LMax C MD Pre-Latte 56 0.094 56 0.200*
Latte 52 0.071 52 0.200*
LMax C BL Pre-Latte 53 0.072 53 0.200*
Latte 58 0.060 58 0.200*
LMax P3 MD Pre-Latte 58 0.103 58 0.196
Latte 62 0.079 62 0.200*
LMax P3 BL Pre-Latte 60 0.130 60 0.013
Latte 61 0.113 61 0.050
LMax P4 MD Pre-Latte 71 0.106 71 0.046
Latte 62 0.101 62 0.182
LMax P4 BL Pre-Latte 71 0.106 71 0.046
Latte 59 0.101 62 0.182
LMax M1 MD Pre-Latte 56 0.085 56 0.200*
Latte 63 0.100 63 0.195
LMax M1 BL Pre-Latte 63 0.120 63 0.024
Latte 64 0.074 64 0.200*
LMax M2 MD Pre-Latte 64 0.117 64 0.030
Latte 54 0.068 54 0.200*
LMax M2 BL Pre-Latte 63 0.091 63 0.200*
Latte 61 0.126 61 0.018
LMax M3 MD Pre-Latte 29 0.164 29 0.045
Latte 18 0.170 18 0.179
LMax M3 BL Pre-Latte 29 0.091 29 0.200*
Latte 21 0.100 21 0.200*
RMax I1 MD Pre-Latte 48 0.108 48 0.200*
Latte 54 0.078 54 0.200*
RMax I1 BL Pre-Latte 53 0.089 53 0.200*
Latte 54 0.101 54 0.200*
RMax I2 MD Pre-Latte 47 0.060 47 0.200*
Latte 49 0.075 49 0.200*
RMax I2 BL Pre-Latte 48 0.081 48 0.200*
Latte 54 0.076 49 0.200*
RMax C MD Pre-Latte 60 0.074 60 0.200*
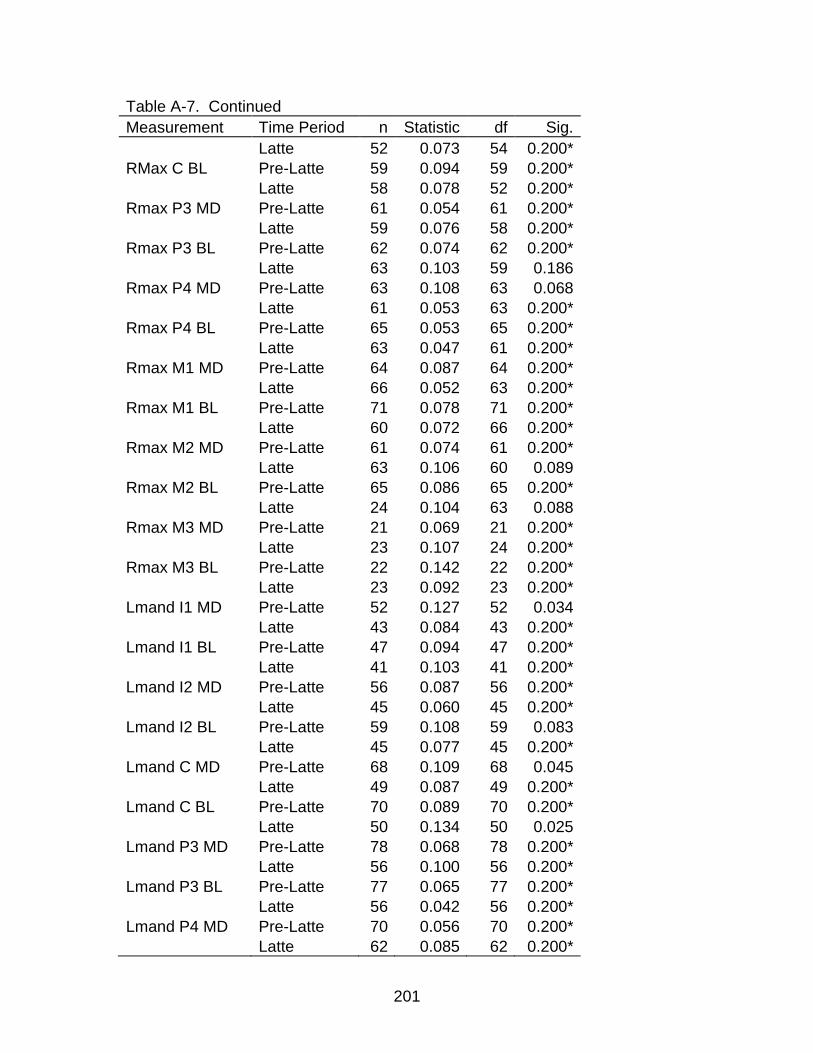
201
Table A-7. Continued
Measurement Time Period n Statistic df Sig.
Latte 52 0.073 54 0.200*
RMax C BL Pre-Latte 59 0.094 59 0.200*
Latte 58 0.078 52 0.200*
Rmax P3 MD Pre-Latte 61 0.054 61 0.200*
Latte 59 0.076 58 0.200*
Rmax P3 BL Pre-Latte 62 0.074 62 0.200*
Latte 63 0.103 59 0.186
Rmax P4 MD Pre-Latte 63 0.108 63 0.068
Latte 61 0.053 63 0.200*
Rmax P4 BL Pre-Latte 65 0.053 65 0.200*
Latte 63 0.047 61 0.200*
Rmax M1 MD Pre-Latte 64 0.087 64 0.200*
Latte 66 0.052 63 0.200*
Rmax M1 BL Pre-Latte 71 0.078 71 0.200*
Latte 60 0.072 66 0.200*
Rmax M2 MD Pre-Latte 61 0.074 61 0.200*
Latte 63 0.106 60 0.089
Rmax M2 BL Pre-Latte 65 0.086 65 0.200*
Latte 24 0.104 63 0.088
Rmax M3 MD Pre-Latte 21 0.069 21 0.200*
Latte 23 0.107 24 0.200*
Rmax M3 BL Pre-Latte 22 0.142 22 0.200*
Latte 23 0.092 23 0.200*
Lmand I1 MD Pre-Latte 52 0.127 52 0.034
Latte 43 0.084 43 0.200*
Lmand I1 BL Pre-Latte 47 0.094 47 0.200*
Latte 41 0.103 41 0.200*
Lmand I2 MD Pre-Latte 56 0.087 56 0.200*
Latte 45 0.060 45 0.200*
Lmand I2 BL Pre-Latte 59 0.108 59 0.083
Latte 45 0.077 45 0.200*
Lmand C MD Pre-Latte 68 0.109 68 0.045
Latte 49 0.087 49 0.200*
Lmand C BL Pre-Latte 70 0.089 70 0.200*
Latte 50 0.134 50 0.025
Lmand P3 MD Pre-Latte 78 0.068 78 0.200*
Latte 56 0.100 56 0.200*
Lmand P3 BL Pre-Latte 77 0.065 77 0.200*
Latte 56 0.042 56 0.200*
Lmand P4 MD Pre-Latte 70 0.056 70 0.200*
Latte 62 0.085 62 0.200*

202
Table A-7. Continued
Measurement Time Period n Statistic df Sig.
LMand P4 BL Pre-Latte 71 0.078 71 0.200*
Latte 57 0.082 57 0.200*
LMand M1 MD Pre-Latte 63 0.056 63 0.200*
Latte 63 0.067 63 0.200*
LMand M1 BL Pre-Latte 74 0.068 74 0.200*
Latte 66 0.107 66 0.061
LMand M2 MD Pre-Latte 58 0.096 58 0.200*
Latte 64 0.112 64 0.044
LMand M2 BL Pre-Latte 63 0.097 63 0.200*
Latte 67 0.099 67 0.176
LMand M3 MD Pre-Latte 26 0.133 26 0.200*
Latte 36 0.092 36 0.200*
LMand M3 BL Pre-Latte 30 0.133 30 0.185
Latte 37 0.104 37 0.200*
RMand I1 MD Pre-Latte 51 0.127 51 0.040
Latte 36 0.103 36 0.200*
RMand I1 BL Pre-Latte 50 0.097 50 0.200*
Latte 33 0.109 33 0.200*
RMand I2 MD Pre-Latte 60 0.099 60 0.200*
Latte 44 0.070 44 0.200*
RMand I2 BL Pre-Latte 60 0.073 60 0.200*
Latte 45 0.092 45 0.200*
RMand C MD Pre-Latte 64 0.069 64 0.200*
Latte 51 0.052 51 0.200*
RMand C BL Pre-Latte 64 0.059 64 0.200*
Latte 50 0.057 50 0.200*
RMand P3 MD Pre-Latte 69 0.094 69 0.200*
Latte 57 0.116 57 0.055
RMand P3 BL Pre-Latte 70 0.085 70 0.200*
Latte 55 0.083 55 0.200*
RMand P4 MD Pre-Latte 65 0.062 65 0.200*
Latte 61 0.071 61 0.200*
RMand P4 BL Pre-Latte 74 0.095 74 0.097
Latte 59 0.135 59 0.009
RMand M1 MD Pre-Latte 62 0.055 62 0.200*
Latte 71 0.073 71 0.200*
RMand M1 BL Pre-Latte 68 0.103 68 0.071
Latte 77 0.081 77 0.200*
RMand M2 MD Pre-Latte 48 0.143 48 0.015
Latte 65 0.092 65 0.200*
RMand M2 BL Pre-Latte 58 0.087 58 0.200*
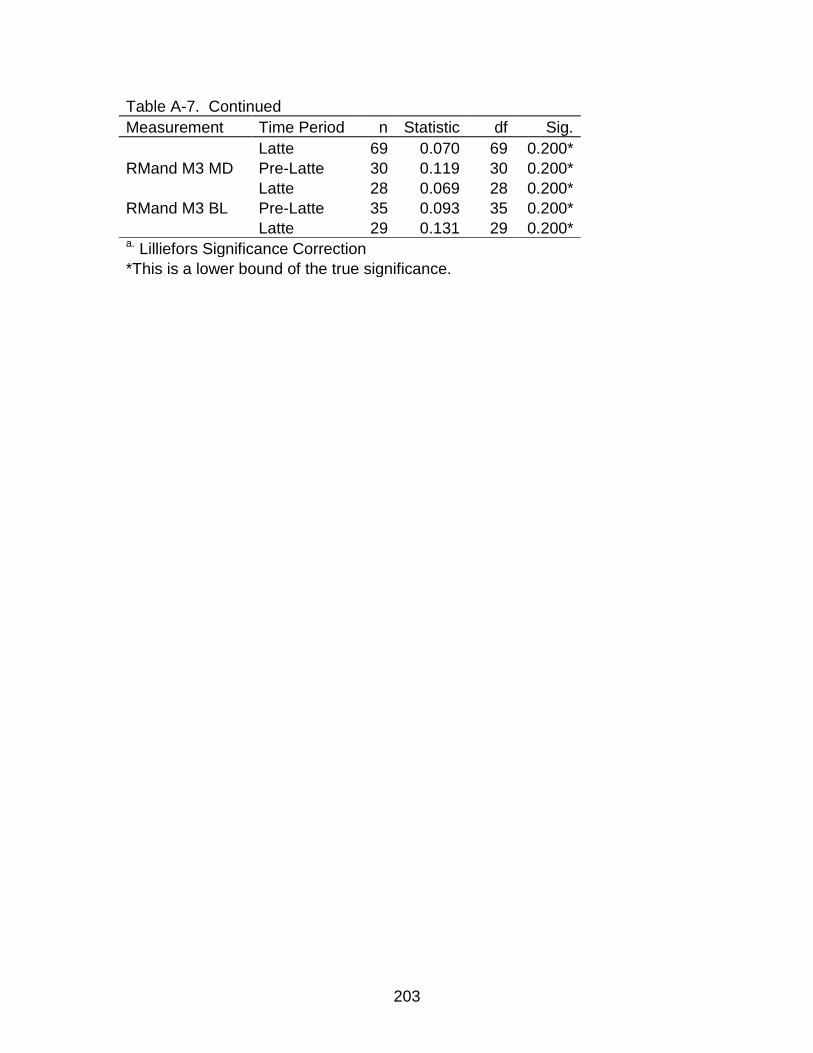
203
Table A-7. Continued
Measurement Time Period n Statistic df Sig.
Latte 69 0.070 69 0.200*
RMand M3 MD Pre-Latte 30 0.119 30 0.200*
Latte 28 0.069 28 0.200*
RMand M3 BL Pre-Latte 35 0.093 35 0.200*
Latte 29 0.131 29 0.200* a. Lilliefors Significance Correction
*This is a lower bound of the true significance.

204
Table A-8. Levene's Test of Homogeneity of Variance based on the mean
Measurement Levene Statistic df1 df2 Sig.
LMax I1 MD 0.674 1 97 0.414 LMax I1 BL 0.884 1 98 0.349 LMax I2 MD 0.427 1 98 0.515 LMax I2 BL 1.638 1 96 0.204 LMax C MD 1.952 1 106 0.165 LMax C BL 1.145 1 109 0.287 LMax P3 MD 0.448 1 118 0.504 LMax P3 BL 2.538 1 119 0.114 LMax P4 MD 1.869 1 131 0.174 LMax P4 BL 0.113 1 128 0.737 LMax M1 MD 1.520 1 117 0.220 LMax M1 BL 0.871 1 125 0.352 LMax M2 MD 0.405 1 116 0.526 LMax M2 BL 0.072 1 122 0.790 LMax M3 MD 2.212 1 45 0.144 LMax M3 BL 1.829 1 48 0.183 RMax I1 MD 6.234 1 100 0.014 RMax I1 BL 0.512 1 105 0.476 RMax I2 MD 0.302 1 94 0.584 RMax I2 BL 0.003 1 95 0.960 RMax C MD 0.368 1 112 0.545 RMax C BL 0.896 1 109 0.346 RMax P3 MD 1.952 1 117 0.165 RMax P3 BL 0.169 1 119 0.682 RMax P4 MD 4.465 1 124 0.037 RMax P4 BL 0.045 1 124 0.833 RMax M1 MD 0.704 1 125 0.403 RMax M1 BL 0.015 1 135 0.902 RMax M2 MD 0.286 1 119 0.594 RMax M2 BL 0.068 1 126 0.795 RMax M3 MD 0.859 1 43 0.359 RMax M3 BL 0.882 1 43 0.353 LMand I1 MD 3.333 1 93 0.071 LMand I1 BL 0.256 1 86 0.614 LMand I2 MD 4.023 1 99 0.048 LMand I2 BL 0.187 1 102 0.666 LMand C MD 0.008 1 115 0.929 LMand C BL 0.034 1 118 0.855 LMand P3 MD 2.809 1 132 0.096 LMand P3 BL 2.807 1 131 0.096 LMand P4 MD 2.733 1 130 0.101 LMand P4 BL 0.857 1 126 0.356
Table A-8. Continued
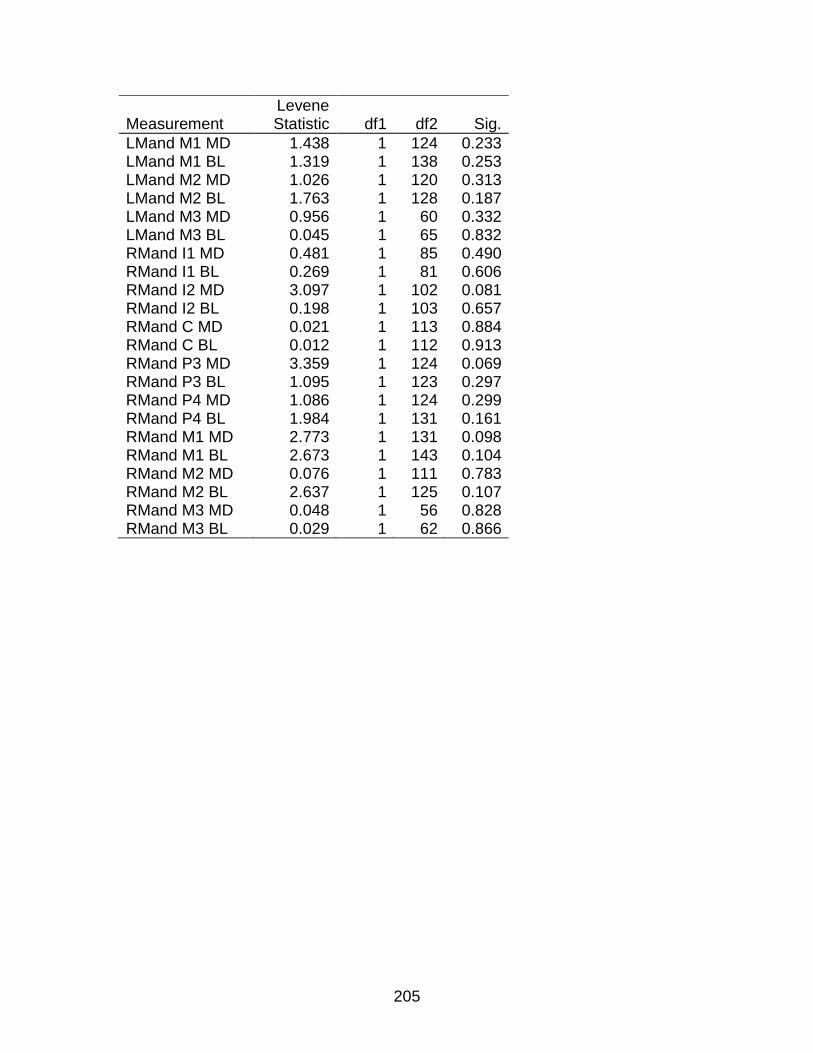
205
Measurement Levene Statistic df1 df2 Sig.
LMand M1 MD 1.438 1 124 0.233 LMand M1 BL 1.319 1 138 0.253 LMand M2 MD 1.026 1 120 0.313 LMand M2 BL 1.763 1 128 0.187 LMand M3 MD 0.956 1 60 0.332 LMand M3 BL 0.045 1 65 0.832 RMand I1 MD 0.481 1 85 0.490 RMand I1 BL 0.269 1 81 0.606 RMand I2 MD 3.097 1 102 0.081 RMand I2 BL 0.198 1 103 0.657 RMand C MD 0.021 1 113 0.884 RMand C BL 0.012 1 112 0.913 RMand P3 MD 3.359 1 124 0.069 RMand P3 BL 1.095 1 123 0.297 RMand P4 MD 1.086 1 124 0.299 RMand P4 BL 1.984 1 131 0.161 RMand M1 MD 2.773 1 131 0.098 RMand M1 BL 2.673 1 143 0.104 RMand M2 MD 0.076 1 111 0.783 RMand M2 BL 2.637 1 125 0.107 RMand M3 MD 0.048 1 56 0.828 RMand M3 BL 0.029 1 62 0.866

206
APPENDIX B ANALYSES OF VARIANCE TESTS FOR DENTAL MEASUREMENTS

207
Table B-1. Two-Way Factorial ANOVA for LMax I1 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 3.596a 3 1.199 A.53 0.00604
Intercept 5239.096 1 5239.096 19797.321 0.00
TimePeriod 2.852 1 2.852 10.777 0.00
SexCombined 0.622 1 0.622 2.351 0.13 TimePeriod * SexCombined 0.008 1 0.008 0.031 0.86
Error 17.201 65 0.265 Total 5429.299 69
Corrected Total 20.798 68
R Squared = 0.173 (Adjusted R Squared = 0.135) Table B-2. Two-Way Factorial ANOVA for LMax I1 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 1.209a 3 0.403 2.664 0.05503
Intercept 4046.654 1 4046.654 26742.88 0.00
TimePeriod 0.418 1 0.418 2.763 0.10
SexCombined 0.727 1 0.727 4.807 0.03 TimePeriod * SexCombined 0.092 1 0.092 0.606 0.44
Error 9.987 66 0.151 Total 4189.417 70
Corrected Total 11.196 69
R Squared = 0.108 (Adjusted R Squared = 0.067)
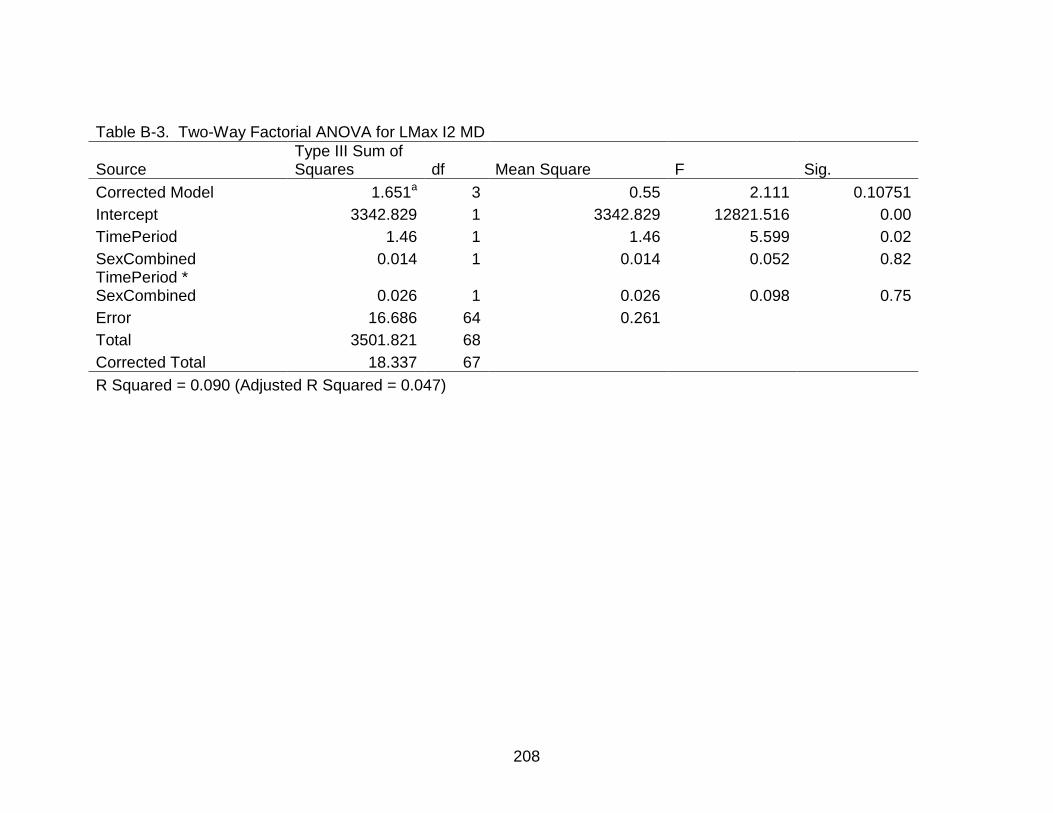
208
Table B-3. Two-Way Factorial ANOVA for LMax I2 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 1.651a 3 0.55 2.111 0.10751
Intercept 3342.829 1 3342.829 12821.516 0.00
TimePeriod 1.46 1 1.46 5.599 0.02
SexCombined 0.014 1 0.014 0.052 0.82 TimePeriod * SexCombined 0.026 1 0.026 0.098 0.75
Error 16.686 64 0.261 Total 3501.821 68
Corrected Total 18.337 67
R Squared = 0.090 (Adjusted R Squared = 0.047)

209
Table B-4. Two-Way Factorial ANOVA for LMax I2 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 3.054 a 3 1.018 8.652 0.00007a
Intercept 3285.352 1 3285.352 27927.215 0.00
TimePeriod 0.494 1 0.494 4.202 0.04b
SexCombined 1.848 1 1.848 15.709 0.00b
TimePeriod * SexCombined 0.573 1 0.573 4.875 0.03b
Error 7.529 64 0.118 Total 3350.988 68
Corrected Total 10.583 67
R Squared = 0.289 (Adjusted R Squared = 0.255) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

210
Table B-5. Two-Way Factorial ANOVA for LMax C MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 3.989a 3 1.33 8.015 0.00011a
Intercept 5170.027 1 5170.027 31161.041 0.00
TimePeriod 0.193 1 0.193 1.161 0.28
SexCombined 3.781 1 3.781 22.787 0.00b TimePeriod * SexCombined 0.008 1 0.008 0.048 0.83
Error 11.78 71 0.166 Total 5305.585 75
Corrected Total 15.769 74
R Squared = 0.253 (Adjusted R Squared = 0.221) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

211
Table B-6. Two-Way Factorial ANOVA for LMax C BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 3.367a 3 1.122 5.939 0.00113027
Intercept 5511.714 1 5511.714 29166.158 0.00
TimePeriod 0.006 1 0.006 0.034 0.85
SexCombined 2.056 1 2.056 10.881 0.00 TimePeriod * SexCombined 1.387 1 1.387 7.337 0.01
Error 13.417 71 0.189 Total 5730.832 75
Corrected Total 16.785 74
R Squared = 0.201 (Adjusted R Squared = 0.167)

212
Table B-7. Two-Way Factorial ANOVA for LMax P3 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 6.572a 3 2.191 9.997 0.00001a
Intercept 5168.768 1 5168.768 23586.009 0.00
TimePeriod 4.613 1 4.613 21.049 0.00b
SexCombined 1.462 1 1.462 6.673 0.01b
TimePeriod * SexCombined 0.072 1 0.072 0.329 0.57
Error 17.532 80 0.219 Total 5268.44 84
Corrected Total 24.104 83
R Squared = 0.273 (Adjusted R Squared = 0.245) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level
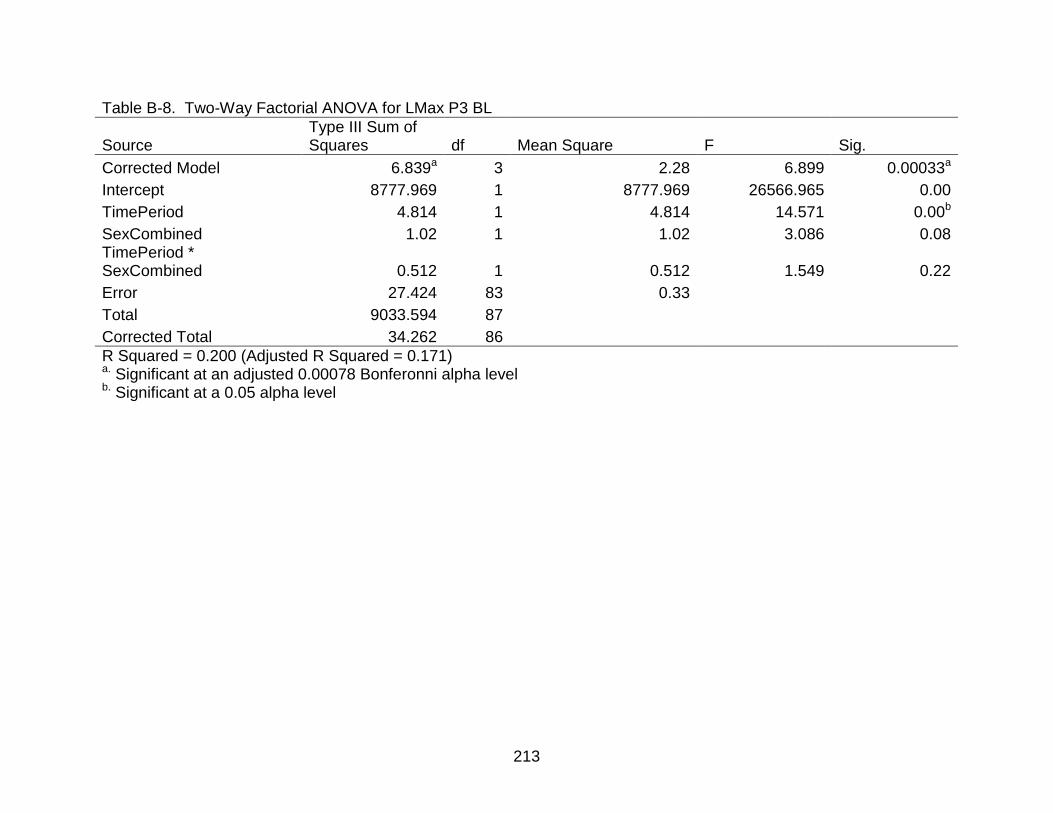
213
Table B-8. Two-Way Factorial ANOVA for LMax P3 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 6.839a 3 2.28 6.899 0.00033a
Intercept 8777.969 1 8777.969 26566.965 0.00
TimePeriod 4.814 1 4.814 14.571 0.00b
SexCombined 1.02 1 1.02 3.086 0.08 TimePeriod * SexCombined 0.512 1 0.512 1.549 0.22
Error 27.424 83 0.33 Total 9033.594 87
Corrected Total 34.262 86
R Squared = 0.200 (Adjusted R Squared = 0.171) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

214
Table B-9. Two-Way Factorial ANOVA for LMax P4 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 3.817a 3 1.272 5.717 0.00125
Intercept 5169.992 1 5169.992 23228.853 0.00
TimePeriod 2.061 1 2.061 9.262 0.00
SexCombined 1.526 1 1.526 6.857 0.01 TimePeriod * SexCombined 0.018 1 0.018 0.079 0.78
Error 20.254 91 0.223 Total 5496.688 95
Corrected Total 24.071 94 R Squared = 0.159 (Adjusted R Squared = 0.131)
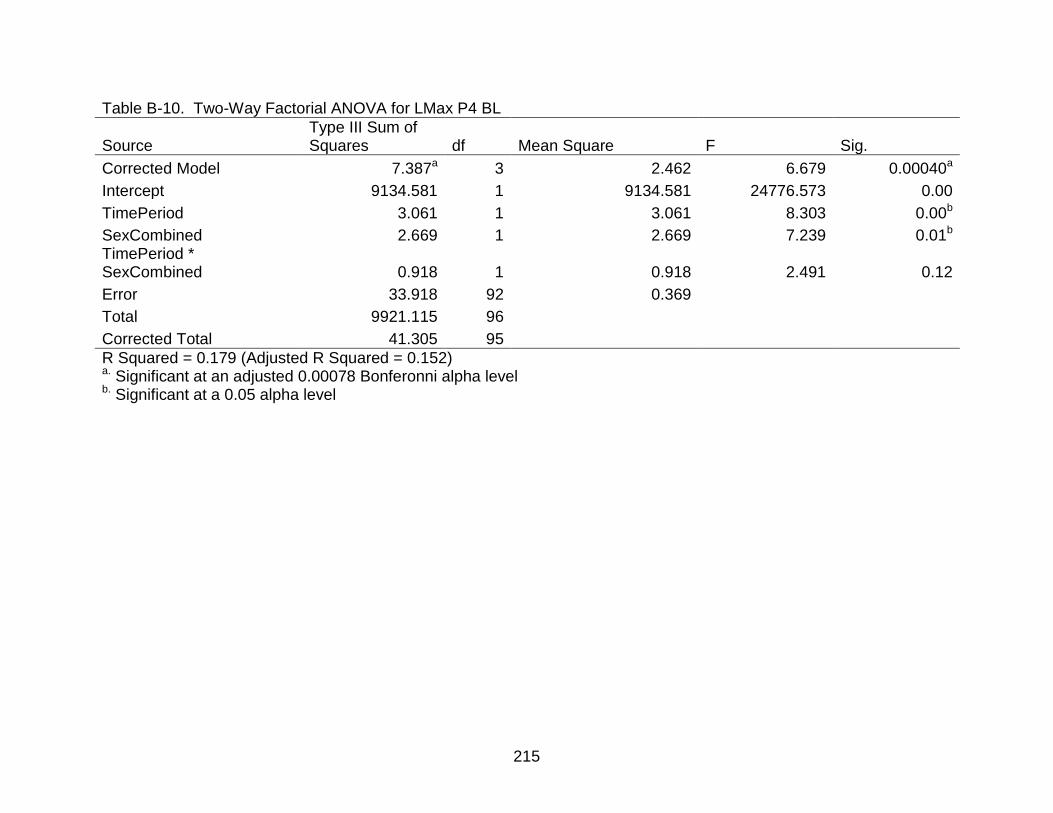
215
Table B-10. Two-Way Factorial ANOVA for LMax P4 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 7.387a 3 2.462 6.679 0.00040a
Intercept 9134.581 1 9134.581 24776.573 0.00
TimePeriod 3.061 1 3.061 8.303 0.00b
SexCombined 2.669 1 2.669 7.239 0.01b
TimePeriod * SexCombined 0.918 1 0.918 2.491 0.12
Error 33.918 92 0.369 Total 9921.115 96
Corrected Total 41.305 95
R Squared = 0.179 (Adjusted R Squared = 0.152) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

216
Table B-11. Two-Way Factorial ANOVA for LMax M1 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 6.835a 3 2.278 8.657 0.00005a
Intercept 9960.838 1 9960.838 37847.924 0.00
TimePeriod 3.734 1 3.734 14.187 0.00b
SexCombined 2.77 1 2.77 10.524 0.00b
TimePeriod * SexCombined 0 1 0 0 1.00
Error 21.054 80 0.263 Total 10771.033 84
Corrected Total 27.89 83
R Squared = .245 (Adjusted R Squared = .217) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

217
Table B-12. Two-Way Factorial ANOVA for LMax M1 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 9.023a 3 3.008 10.362 0.00001a
Intercept 12272.892 1 12272.892 42282.653 0.00
TimePeriod 4.685 1 4.685 16.141 0.00b
SexCombined 3.665 1 3.665 12.626 0.00b
TimePeriod * SexCombined 0.099 1 0.099 0.34 0.56
Error 25.252 87 0.29 Total 13107.333 91
Corrected Total 34.275 90
R Squared = 0.263 (Adjusted R Squared = 0.238) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level
Table B-13. Two-Way Factorial ANOVA for LMax M2 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 13.957a 3 4.652 15.346 0.00000a
Intercept 7942.252 1 7942.252 26198.798 0.00
TimePeriod 7.42 1 7.42 24.475 0.00b
SexCombined 2.185 1 2.185 7.208 0.01b
TimePeriod * SexCombined 0.623 1 0.623 2.056 0.16
Error 24.859 82 0.303 Total 9494.712 86
Corrected Total 38.815 85
R Squared = 0.360 (Adjusted R Squared = 0.336) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

218
Table B-14. Two-Way Factorial ANOVA for LMax M2 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 22.942a 3 7.647 21.584 0.0000a
Intercept 11879.491 1 11879.491 33527.741 0.00
TimePeriod 14.303 1 14.303 40.368 0.00b
SexCombined 4.26 1 4.26 12.023 0.00b
TimePeriod * SexCombined 0.165 1 0.165 0.466 0.50
Error 31.534 89 0.354 Total 13283.297 93
Corrected Total 54.477 92
R Squared = 0.421 (Adjusted R Squared = 0.402) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level
Table B-15. Two-Way Factorial ANOVA for LMax M3 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 9.775a 3 3.258 3.705 0.02019
Intercept 3234.598 1 3234.598 3677.787 0.00
TimePeriod 2.543 1 2.543 2.892 0.10
SexCombined 7.939 1 7.939 9.027 0.00
TimePeriod * SexCombined 1.06 1 1.06 1.205 0.28
Error 31.662 36 0.879 Total 3588.821 40
Corrected Total 41.437 39
R Squared = 0.236 (Adjusted R Squared = 0.172)

219
Table B-16. Two-Way Factorial ANOVA for LMax M3 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 6.851a 3 2.284 3.934 0.01518
Intercept 5238.222 1 5238.222 9024.631 0.00
TimePeriod 3.945 1 3.945 6.796 0.01
SexCombined 3.695 1 3.695 6.366 0.02
TimePeriod * SexCombined 0.427 1 0.427 0.736 0.40
Error 22.637 39 0.58 Total 5588.395 43
Corrected Total 29.488 42
R Squared = 0.232 (Adjusted R Squared = 0.173)
Table B-17. Two-Way Factorial ANOVA for RMax I1 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 4.297a 3 1.432 4.882 0.00398
Intercept 5248.785 1 5248.785 17890.567 0.00
TimePeriod 3.281 1 3.281 11.182 0.00
SexCombined 0.496 1 0.496 1.692 0.20
TimePeriod * SexCombined 0.157 1 0.157 0.537 0.47
Error 19.363 66 0.293 Total 5410.721 70
Corrected Total 23.661 69
R Squared = 0.182 (Adjusted R Squared = 0.144)

220
Table B-18. Two-Way Factorial ANOVA for RMax I1 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 1.381a 3 0.46 2.781 0.04712
Intercept 4379.103 1 4379.103 26459.067 0.00
TimePeriod 0.436 1 0.436 2.637 0.11
SexCombined 0.726 1 0.726 4.388 0.04
TimePeriod * SexCombined 0.181 1 0.181 1.093 0.30
Error 11.916 72 0.166 Total 4447.095 76
Corrected Total 13.297 75
R Squared = 0.104 (Adjusted R Squared = 0.066)
Table B-19. Two-Way Factorial ANOVA for RMax I2 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 3.021a 3 1.007 4.563 0.00609
Intercept 3129.101 1 3129.101 14179.744 0.00
TimePeriod 2.903 1 2.903 13.156 0.00
SexCombined 0.087 1 0.087 0.396 0.53 TimePeriod * SexCombined 0.201 1 0.201 0.911 0.34
Error 13.02 59 0.221 Total 3238.189 63
Corrected Total 16.041 62
R Squared = 0.188 (Adjusted R Squared = 0.147)

221
Table B-20. Two-Way Factorial ANOVA for RMax I2 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 2.116a 3 0.705 3.709 0.01598
Intercept 3287.914 1 3287.914 17291.212 0.00
TimePeriod 0.347 1 0.347 1.827 0.18
SexCombined 1.476 1 1.476 7.764 0.01 TimePeriod * SexCombined 0.263 1 0.263 1.383 0.24
Error 11.979 63 0.19 Total 3353.617 67
Corrected Total 14.095 66
R Squared = 0.150 (Adjusted R Squared = 0.110)
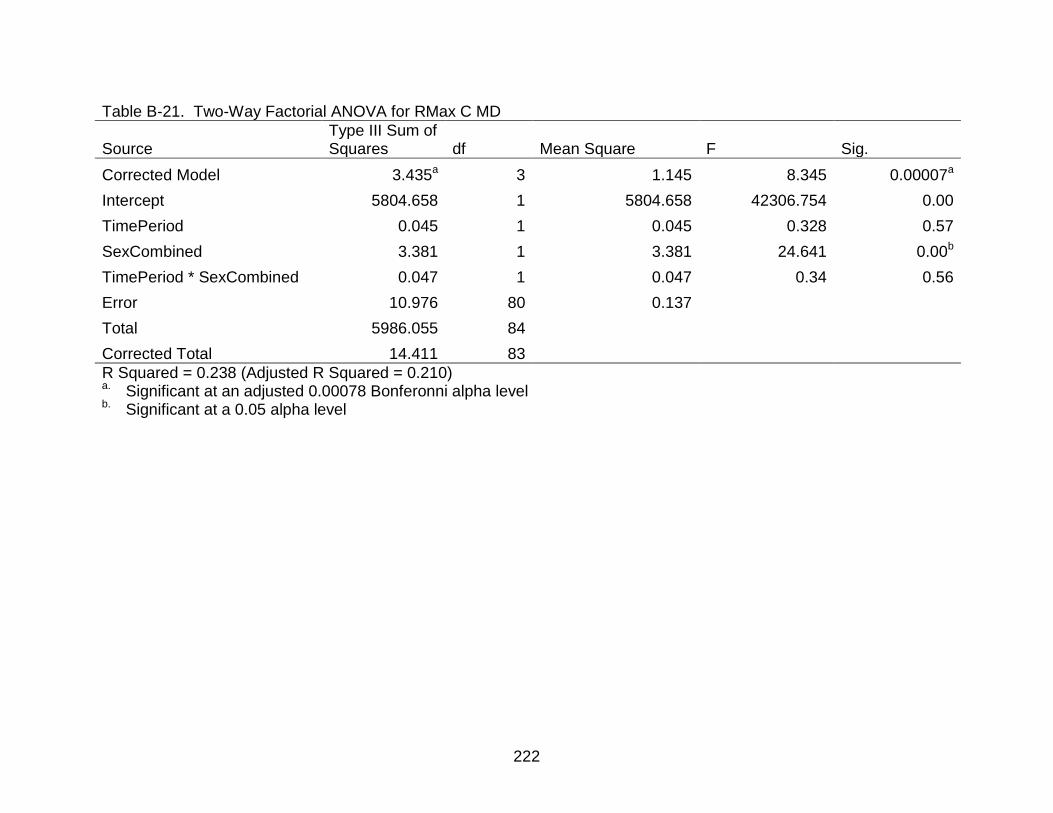
222
Table B-21. Two-Way Factorial ANOVA for RMax C MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 3.435a 3 1.145 8.345 0.00007a
Intercept 5804.658 1 5804.658 42306.754 0.00
TimePeriod 0.045 1 0.045 0.328 0.57
SexCombined 3.381 1 3.381 24.641 0.00b
TimePeriod * SexCombined 0.047 1 0.047 0.34 0.56
Error 10.976 80 0.137 Total 5986.055 84
Corrected Total 14.411 83
R Squared = 0.238 (Adjusted R Squared = 0.210) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

223
Table B-22. Two-Way Factorial ANOVA for RMax C BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 3.010a 3 1.003 4.726 0.00443
Intercept 5859.032 1 5859.032 27594.638 0.00
TimePeriod 0.114 1 0.114 0.539 0.47
SexCombined 1.852 1 1.852 8.723 0.00
TimePeriod * SexCombined 1.26 1 1.26 5.933 0.02
Error 16.349 77 0.212 Total 6118.622 81
Corrected Total 19.359 80
R Squared = 0.155 (Adjusted R Squared = 0.123)
Table B-23. Two-Way Factorial ANOVA for RMax P3 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 3.284a 3 1.095 5.804 0.00118
Intercept 5279.585 1 5279.585 27987.8 0.00
TimePeriod 2.313 1 2.313 12.262 0.00
SexCombined 0.637 1 0.637 3.378 0.07
TimePeriod * SexCombined 0.198 1 0.198 1.049 0.31
Error 15.846 84 0.189 Total 5358.863 88
Corrected Total 19.13 87
R Squared = 0.172 (Adjusted R Squared = 0.142)
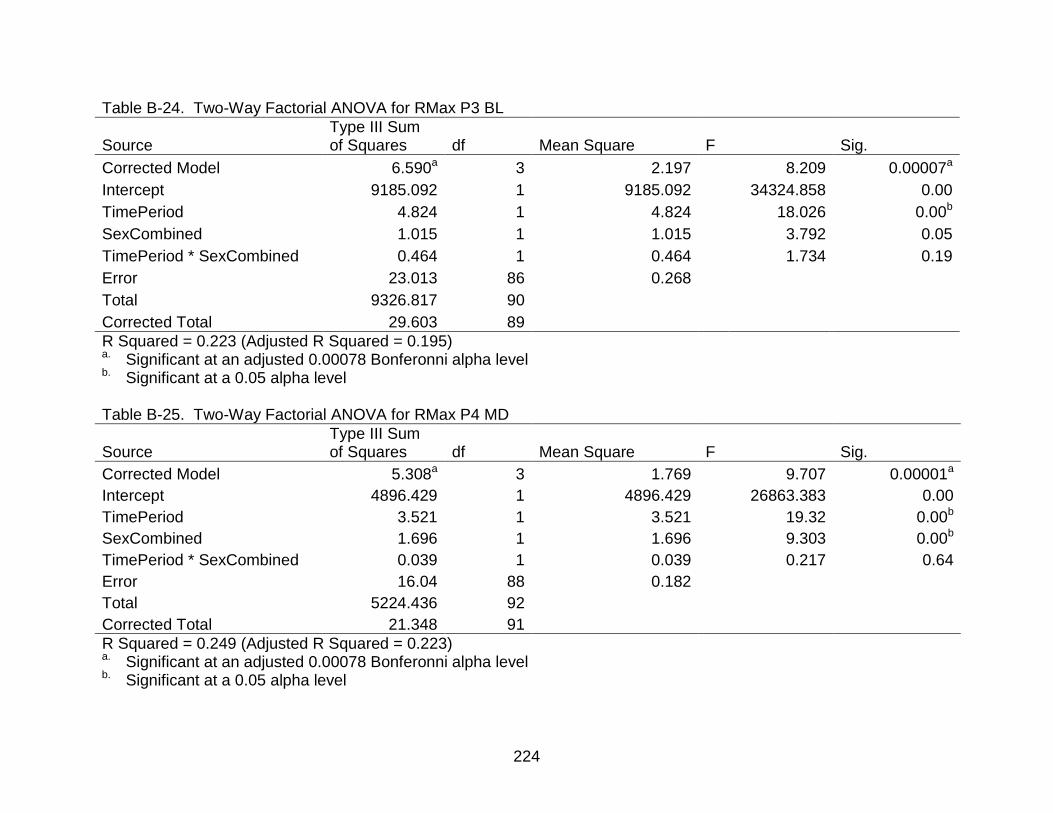
224
Table B-24. Two-Way Factorial ANOVA for RMax P3 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 6.590a 3 2.197 8.209 0.00007a
Intercept 9185.092 1 9185.092 34324.858 0.00
TimePeriod 4.824 1 4.824 18.026 0.00b
SexCombined 1.015 1 1.015 3.792 0.05
TimePeriod * SexCombined 0.464 1 0.464 1.734 0.19
Error 23.013 86 0.268 Total 9326.817 90
Corrected Total 29.603 89
R Squared = 0.223 (Adjusted R Squared = 0.195) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level
Table B-25. Two-Way Factorial ANOVA for RMax P4 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 5.308a 3 1.769 9.707 0.00001a
Intercept 4896.429 1 4896.429 26863.383 0.00
TimePeriod 3.521 1 3.521 19.32 0.00b
SexCombined 1.696 1 1.696 9.303 0.00b
TimePeriod * SexCombined 0.039 1 0.039 0.217 0.64
Error 16.04 88 0.182 Total 5224.436 92
Corrected Total 21.348 91
R Squared = 0.249 (Adjusted R Squared = 0.223) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

225
Table B-26. Two-Way Factorial ANOVA for RMax P4 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 13.591a 3 4.53 17.023 0.00000a
Intercept 8849.841 1 8849.841 33254.095 0.00
TimePeriod 7.204 1 7.204 27.07 0.00b
SexCombined 4.575 1 4.575 17.19 0.00b
TimePeriod * SexCombined 0.295 1 0.295 1.107 0.30
Error 23.419 88 0.266 Total 9386.412 92
Corrected Total 37.01 91
R Squared = 0.367 (Adjusted R Squared = 0.346) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level
Table B-27. Two-Way Factorial ANOVA for RMax M1 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 6.817a 3 2.272 9.041 0.00003a
Intercept 11153.872 1 11153.872 44379.428 0.00
TimePeriod 6.286 1 6.286 25.011 0.00b
SexCombined 0.293 1 0.293 1.166 0.28
TimePeriod * SexCombined 0.06 1 0.06 0.238 0.63
Error 20.358 81 0.251 Total 11257.388 85
Corrected Total 27.175 84
R Squared = 0.251 (Adjusted R Squared = 0.223) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

226
Table B-28. Two-Way Factorial ANOVA for RMax M1 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 10.285a 3 3.428 13.18 0.00000a
Intercept 13092.688 1 13092.688 50334.755 0.00
TimePeriod 5.236 1 5.236 20.132 0.00b
SexCombined 4.575 1 4.575 17.59 0.00b
TimePeriod * SexCombined 0.53 1 0.53 2.037 0.16
Error 23.93 92 0.26 Total 13430.062 96
Corrected Total 34.215 95
R Squared = 0.301 (Adjusted R Squared = 0.278) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level
Table B-29. Two-Way Factorial ANOVA for RMax M2 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 16.801a 3 5.6 18.605 0.0000a
Intercept 9320.397 1 9320.397 30962.461 0.00
TimePeriod 11.614 1 11.614 38.581 0.00b
SexCombined 2.76 1 2.76 9.168 0.00c
TimePeriod * SexCombined 0.044 1 0.044 0.145 0.70
Error 26.189 87 0.301 Total 9869.333 91
Corrected Total 42.99 90
R Squared = 0.391 (Adjusted R Squared = 0.370) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

227
Table B-30. Two-Way Factorial ANOVA for RMax M2 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 19.825a 3 6.608 17.436 0.00000a
Intercept 12923.433 1 12923.433 34099.631 0.00
TimePeriod 13.13 1 13.13 34.645 0.00b
SexCombined 4.096 1 4.096 10.807 0.00c
TimePeriod * SexCombined 0.001 1 0.001 0.002 0.97
Error 35.625 94 0.379 Total 13716.55 98
Corrected Total 55.45 97
R Squared = 0.358 (Adjusted R Squared = 0.337) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level
Table B-31. Two-Way Factorial ANOVA for RMax M3 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 3.030a 3 1.01 1.057 0.37980
Intercept 3176.591 1 3176.591 3323.831 0.00
TimePeriod 1.985 1 1.985 2.077 0.16
SexCombined 1.799 1 1.799 1.882 0.18
TimePeriod * SexCombined 0.052 1 0.052 0.055 0.82
Error 33.45 35 0.956 Total 3445.583 39
Corrected Total 36.48 38
R Squared = 0.083 (Adjusted R Squared = 0.004)

228
Table B-32. Two-Way Factorial ANOVA for RMax M3 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 4.869a 3 1.623 2.617 0.06636
Intercept 4784.046 1 4784.046 7712.077 0.00
TimePeriod 2.036 1 2.036 3.282 0.08
SexCombined 3.336 1 3.336 5.377 0.03
TimePeriod * SexCombined 0.56 1 0.56 0.903 0.35
Error 21.712 35 0.62 Total 4975.126 39
Corrected Total 26.581 38
R Squared = 0.183 (Adjusted R Squared = 0.113)
Table B-33. Two-Way Factorial ANOVA for LMand I1 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 3.498a 3 1.166 8.023 0.00014a
Intercept 1881.394 1 1881.394 12944.752 0.00
TimePeriod 3.145 1 3.145 21.636 0.00b
SexCombined 0.153 1 0.153 1.054 0.31 TimePeriod * SexCombined 0.029 1 0.029 0.202 0.66
Error 8.72 60 0.145 Total 2109.172 64
Corrected Total 12.219 63
R Squared = 0.286 (Adjusted R Squared = 0.251) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

229
Table B-34. Two-Way Factorial ANOVA for LMand I1 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 1.021a 3 0.34 1.996 0.12549
Intercept 2117.819 1 2117.819 12423.168 0.00
TimePeriod 0.805 1 0.805 4.724 0.03
SexCombined 0.147 1 0.147 0.86 0.36 TimePeriod * SexCombined 0.029 1 0.029 0.17 0.68
Error 9.206 54 0.17 Total 2266.611 58
Corrected Total 10.226 57
R Squared = 0.100 (Adjusted R Squared = 0.050)
Table B-35. Two-Way Factorial ANOVA for LMand I2 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 2.137a 3 0.712 3.621 0.01749
Intercept 2482.62 1 2482.62 12616.504 0.00
TimePeriod 1.588 1 1.588 8.072 0.01
SexCombined 0.147 1 0.147 0.747 0.39
TimePeriod * SexCombined 0.032 1 0.032 0.161 0.69
Error 12.987 66 0.197 Total 2883.989 70
Corrected Total 15.125 69
R Squared = 0.141 (Adjusted R Squared = 0.102)

230
Table B-36. Two-Way Factorial ANOVA for LMand I2 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 1.000a 3 0.333 2.778 0.04775
Intercept 2933.052 1 2933.052 24446.84 0.00
TimePeriod 0.035 1 0.035 0.295 0.59
SexCombined 0.823 1 0.823 6.858 0.01
TimePeriod * SexCombined 0.186 1 0.186 1.552 0.22
Error 8.158 68 0.12 Total 3256.129 72
Corrected Total 9.158 71
R Squared = 0.109 (Adjusted R Squared = 0.070)
Table B-37. Two-Way Factorial ANOVA for LMand C MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 7.250a 3 2.417 9.571 0.00002a
Intercept 4254.588 1 4254.588 16849.096 0.00
TimePeriod 0.472 1 0.472 1.869 0.18
SexCombined 6.667 1 6.667 26.402 0.00b
TimePeriod * SexCombined 0.53 1 0.53 2.101 0.15
Error 20.453 81 0.253 Total 4524.127 85
Corrected Total 27.704 84
R Squared = 0.262 (Adjusted R Squared = 0.234) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level
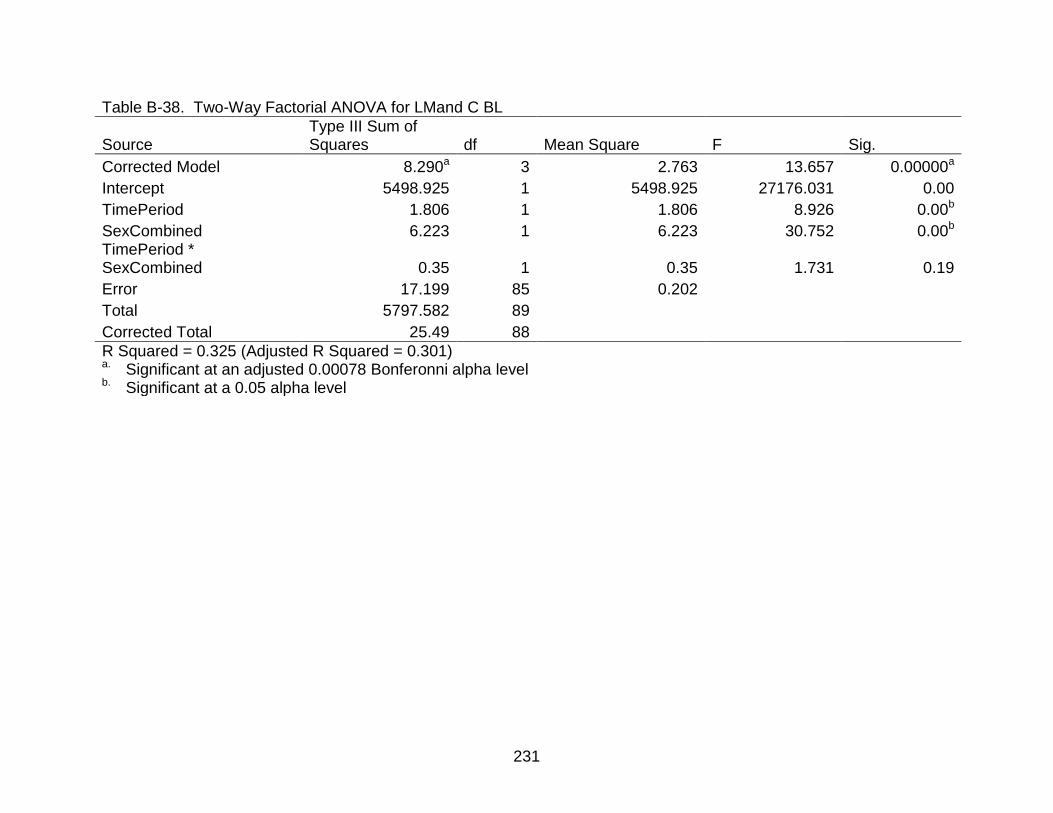
231
Table B-38. Two-Way Factorial ANOVA for LMand C BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 8.290a 3 2.763 13.657 0.00000a
Intercept 5498.925 1 5498.925 27176.031 0.00
TimePeriod 1.806 1 1.806 8.926 0.00b
SexCombined 6.223 1 6.223 30.752 0.00b
TimePeriod * SexCombined 0.35 1 0.35 1.731 0.19
Error 17.199 85 0.202 Total 5797.582 89
Corrected Total 25.49 88
R Squared = 0.325 (Adjusted R Squared = 0.301) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level
232
Table B-39. Two-Way Factorial ANOVA for LMand P3 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 7.152a 3 2.384 14.168 0.00000a
Intercept 5576.195 1 5576.195 33138.75 0.00
TimePeriod 3.141 1 3.141 18.665 0.00b
SexCombined 3.193 1 3.193 18.977 0.00c
TimePeriod * SexCombined 0.978 1 0.978 5.81 0.02b
Error 15.481 92 0.168 Total 5948.445 96
Corrected Total 22.633 95 R Squared = 0.316 (Adjusted R Squared = 0.294)
a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level
Table B-40. Two-Way Factorial ANOVA for LMand P3 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 3.386a 3 1.129 4.522 0.00530
Intercept 7101.785 1 7101.785 28448.67 0.00
TimePeriod 1.873 1 1.873 7.501 0.01
SexCombined 1.085 1 1.085 4.348 0.04 TimePeriod * SexCombined 0.58 1 0.58 2.325 0.13
Error 22.717 91 0.25 Total 7498.103 95
Corrected Total 26.103 94 R Squared = 0.130 (Adjusted R Squared = 0.101)

233
Table B-41. Two-Way Factorial ANOVA for LMand P4 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 5.820a 3 1.94 10.709 0.00000a
Intercept 5933.765 1 5933.765 32751.291 0.00
TimePeriod 1.424 1 1.424 7.861 0.01b
SexCombined 3.369 1 3.369 18.594 0.00b
TimePeriod * SexCombined 0.891 1 0.891 4.917 0.03c
Error 17.031 94 0.181 Total 6216.696 98
Corrected Total 22.851 97 R Squared = 0.255 (Adjusted R Squared = 0.231)
a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

234
Table B-42. Two-Way Factorial ANOVA for LMand P4 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 6.441a 3 2.147 8.575 0.00004a
Intercept 7381.609 1 7381.609 29484.726 0.00
TimePeriod 2.376 1 2.376 9.489 0.00b
SexCombined 3.874 1 3.874 15.473 0.00b
TimePeriod * SexCombined 0.456 1 0.456 1.82 0.18
Error 23.033 92 0.25 Total 7816.277 96
Corrected Total 29.473 95 R Squared = 0.219 (Adjusted R Squared = 0.193)
a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

235
Table B-43. Two-Way Factorial ANOVA for LMand M1 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 4.344a 3 1.448 6.442 0.00061a
Intercept 11906.606 1 11906.606 52971.467 0.00
TimePeriod 0.538 1 0.538 2.392 0.13
SexCombined 3.361 1 3.361 14.951 0.00b
TimePeriod * SexCombined 0.787 1 0.787 3.501 0.07
Error 16.858 75 0.225 Total 12364.952 79
Corrected Total 21.202 78 R Squared = 0.205 (Adjusted R Squared = 0.173)
a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level
Table B-44. Two-Way Factorial ANOVA for LMand M1 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 6.261a 3 2.087 11.262 0.00000a
Intercept 10230.494 1 10230.494 55205.338 0.00 TimePeriod 1.255 1 1.255 6.774 0.01b
SexCombined 4.932 1 4.932 26.615 0.00b
TimePeriod * SexCombined 0.348 1 0.348 1.876 0.17 Error 16.308 88 0.185
Total 10773.979 92 Corrected Total 22.569 91 R Squared = 0.277 (Adjusted R Squared = 0.253)
a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

236
Table B-45. Two-Way Factorial ANOVA for LMand M2 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 5.804a 3 1.935 3.727 0.01461
Intercept 11366.565 1 11366.565 21895.661 0.00
TimePeriod 0.875 1 0.875 1.685 0.20
SexCombined 2.399 1 2.399 4.621 0.03
TimePeriod * SexCombined 1.841 1 1.841 3.546 0.06
Error 41.011 79 0.519 Total 11757.644 83
Corrected Total 46.815 82 R Squared = 0.124 (Adjusted R Squared = 0.091)
Table B-46. Two-Way Factorial ANOVA for LMand M2 BL.
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 16.986a 3 5.662 13.04 0.00000a
Intercept 9889.247 1 9889.247 22776.433 0.00
TimePeriod 9.171 1 9.171 21.123 0.00b
SexCombined 6.205 1 6.205 14.291 0.00b
TimePeriod * SexCombined 0.686 1 0.686 1.58 0.21
Error 37.34 86 0.434 Total 10185.178 90
Corrected Total 54.326 89
R Squared = 0.313 (Adjusted R Squared = 0.289) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

237
Table B-47. Two-Way Factorial ANOVA for LMand M3 MD
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 14.361a 3 4.787 5.102 0.00430
Intercept 4826.761 1 4826.761 5144.283 0.00
TimePeriod 11.881 1 11.881 12.662 0.00
SexCombined 1.593 1 1.593 1.698 0.20
TimePeriod * SexCombined 0.002 1 0.002 0.002 0.97
Error 38.469 41 0.938 Total 5423.724 45
Corrected Total 52.831 44 R Squared = 0.272 (Adjusted R Squared = 0.219)
238
Table B-48. Two-Way Factorial ANOVA for LMand M3 BL
Source Type III Sum of Squares df Mean Square F Sig.
Corrected Model 10.903a 3 3.634 10.974 0.00001a
Intercept 5020.924 1 5020.924 15161.674 0.00
TimePeriod 7.384 1 7.384 22.297 0.00b
SexCombined 2.949 1 2.949 8.905 0.00b
TimePeriod * SexCombined 0.004 1 0.004 0.011 0.92
Error 15.233 46 0.331 Total 5287.874 50
Corrected Total 26.136 49
R Squared = 0.417 (Adjusted R Squared = 0.379) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level Table B-49. Two-Way Factorial ANOVA for RMand I1 MD
Source Type III Sum of Squares
df Mean Square F Sig.
Corrected Model 2.074a 3 0.691 6.384 0.00088 Intercept 1525.705 1 1525.705 14086.23 0.00 TimePeriod 1.681 1 1.681 15.524 0.00 SexCombined 0.011 1 0.011 0.105 0.75 TimePeriod * SexCombined
0.07 1 0.07 0.644 0.43
Error 5.849 54 0.108 Total 1943.646 58 Corrected Total 7.923 57
.R Squared = 0.262 (Adjusted R Squared = 0.221)

239
Table B-50. Two-Way Factorial ANOVA for RMand I1 BL
Source Type III Sum of Squares
df Mean Square F Sig.
Corrected Model 0.313a 3 0.104 1.015 0.39384 Intercept 1800.943 1 1800.943 17491.938 0.00 TimePeriod 0.215 1 0.215 2.088 0.15 SexCombined 0.064 1 0.064 0.622 0.43 TimePeriod * SexCombined
0.05 1 0.05 0.49 0.49
Error 5.251 51 0.103 Total 2165.768 55 Corrected Total 5.564 54
R Squared = 0.056 (Adjusted R Squared = 0.001) Table B-51. Two-Way Factorial ANOVA for RMand I2 MD
Source Type III Sum of Squares
df Mean Square F Sig.
Corrected Model 1.458a 3 0.486 3.851 0.01345 Intercept 2626.457 1 2626.457 20807.176 0.00 TimePeriod 1.09 1 1.09 8.632 0.00 SexCombined 0.291 1 0.291 2.304 0.13 TimePeriod * SexCombined
0.021 1 0.021 0.165 0.69
Error 8.079 64 0.126 Total 2872.665 68 Corrected Total 9.537 67
R Squared = 0.153 (Adjusted R Squared = 0.113)
240
Table B-52. Two-Way Factorial ANOVA for RMand I2 BL
Source Type III Sum of Squares
df Mean Square F Sig.
Corrected Model 0.696a 3 0.232 1.416 0.24585 Intercept 3056.193 1 3056.193 18653.853 0.00 TimePeriod 0.617 1 0.617 3.764 0.06 SexCombined 0.015 1 0.015 0.091 0.76 TimePeriod * SexCombined
0.05 1 0.05 0.305 0.58
Error 10.977 67 0.164 Total 3239.594 71 Corrected Total 11.673 70
R Squared = 0.060 (Adjusted R Squared = 0.018) Table B-53. Two-Way Factorial ANOVA for RMand C MD
Source Type III Sum of Squares
df Mean Square F Sig.
Corrected Model 4.763a 3 1.588 11.801 0.00000a Intercept 4121.565 1 4121.565 30632.515 0.00 TimePeriod 0.424 1 0.424 3.151 0.08 SexCombined 4.405 1 4.405 32.741 0.00b TimePeriod * SexCombined
0.221 1 0.221 1.645 0.20
Error 10.36 77 0.135 Total 4308.722 81 Corrected Total 15.124 80
R Squared = 0.315 (Adjusted R Squared = 0.288) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

241
Table B-54. Two-Way Factorial ANOVA for RMand C BL
Source Type III Sum of Squares
df Mean Square F Sig.
Corrected Model 5.060a 3 1.687 8.182 0.00008a Intercept 5160.1 1 5160.1 25033.04 0.00 TimePeriod 0.379 1 0.379 1.84 0.18 SexCombined 3.81 1 3.81 18.481 0.00b TimePeriod * SexCombined
1.459 1 1.459 7.079 0.01b
Error 16.078 78 0.206 Total 5378.017 82 Corrected Total 21.138 81
R Squared = 0.239 (Adjusted R Squared = 0.210) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

242
Table B-55. Two-Way Factorial ANOVA for RMand P3 MD
Source Type III Sum of Squares
df Mean Square F Sig.
Corrected Model 6.317a 3 2.106 10.207 0.00001a Intercept 5266.893 1 5266.893 25532.716 0.00 TimePeriod 2.49 1 2.49 12.073 0.00b SexCombined 2.508 1 2.508 12.159 0.00b TimePeriod * SexCombined
1.202 1 1.202 5.826 0.02c
Error 17.74 86 0.206 Total 5464.768 90 Corrected Total 24.057 89
R Squared = 0.263 (Adjusted R Squared = 0.237) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level Table B-56. Two-Way Factorial ANOVA for RMand P3 BL
Source Type III Sum of Squares
df Mean Square F Sig.
Corrected Model 3.900a 3 1.3 3.774 0.01344 Intercept 6819.722 1 6819.722 19797.856 0.00 TimePeriod 0.935 1 0.935 2.715 0.10 SexCombined 0.609 1 0.609 1.767 0.19 TimePeriod * SexCombined
2.201 1 2.201 6.39 0.01
Error 29.969 87 0.344 Total 7183.181 91 Corrected Total 33.869 90
R Squared = 0.115 (Adjusted R Squared = 0.085)

243
Table B-57. Two-Way Factorial ANOVA for RMand P4 MD
Source Type III Sum of Squares
df Mean Square F Sig.
Corrected Model 7.144a 3 2.381 15.137 0.00000a Intercept 5637.778 1 5637.778 35835.681 0.00 TimePeriod 2.59 1 2.59 16.462 0.00b SexCombined 4.774 1 4.774 30.344 0.00b TimePeriod * SexCombined
0.032 1 0.032 0.204 0.65
Error 13.844 88 0.157 Total 5810.048 92 Corrected Total 20.989 91
R Squared = 0.340 (Adjusted R Squared = 0.318) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

244
Table B-58. Two-Way Factorial ANOVA for RMand P4 BL
Source Type III Sum of Squares
df Mean Square F Sig.
Corrected Model 8.684a 5 1.736 5.319 0.00018a Intercept 10197.624 1 10197.624 31229.701 0.00 TimePeriod 3.274 1 3.274 10.028 0.00b SexCombined 5.262 2 2.631 8.058 0.00b TimePeriod * SexCombined
0.467 2 0.233 0.715 0.49
Error 41.470 127 0.326 Total 10854.678 133 Corrected Total 50.155 132
R Squared = 0.173 (Adjusted R Squared = 0.141) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level
245
Table B-59. Two-Way Factorial ANOVA for RMand M1 MD
Source Type III Sum of Squares
df Mean Square F Sig.
Corrected Model 8.627a 5 1.725 5.646 0.00000a Intercept 20146.669 1 20146.669 65924.701 0.00 TimePeriod 3.391 1 3.391 11.095 0.00b SexCombined 5.407 2 2.704 8.847 0.00b TimePeriod * SexCombined
0.107 2 0.054 0.176 0.84
Error 38.811 127 0.306 Total 20649.328 133 Corrected Total 47.439 132
R Squared = 0.182 (Adjusted R Squared = 0.150) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level

246
Table B-60. Two-Way Factorial ANOVA for RMand M1 BL
Source Type III Sum of Squares
df Mean Square F Sig.
Corrected Model 8.435a 3 2.812 15.045 0.00000a Intercept 11000.381 1 11000.381 58862.344 0.00 TimePeriod 2.322 1 2.322 12.427 0.00b SexCombined 6.328 1 6.328 33.859 0.00b TimePeriod * SexCombined
0 1 0 0.002 0.96
Error 16.633 89 0.187 Total 11058.707 93 Corrected Total 25.068 92
R Squared = 0.336 (Adjusted R Squared = 0.314) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level Table B-61. Two-Way Factorial ANOVA for RMand M2 MD
Source Type III Sum of Squares
df Mean Square F Sig.
Corrected Model 8.581a 3 2.86 4.426 0.00645 Intercept 9745.587 1 9745.587 15078.329 0.00 TimePeriod 2.103 1 2.103 3.254 0.08 SexCombined 1.378 1 1.378 2.132 0.15 TimePeriod * SexCombined
2.81 1 2.81 4.348 0.04
Error 47.828 74 0.646 Total 10819.881 78 Corrected Total 56.41 77
R Squared = 0.152 (Adjusted R Squared = 0.118)
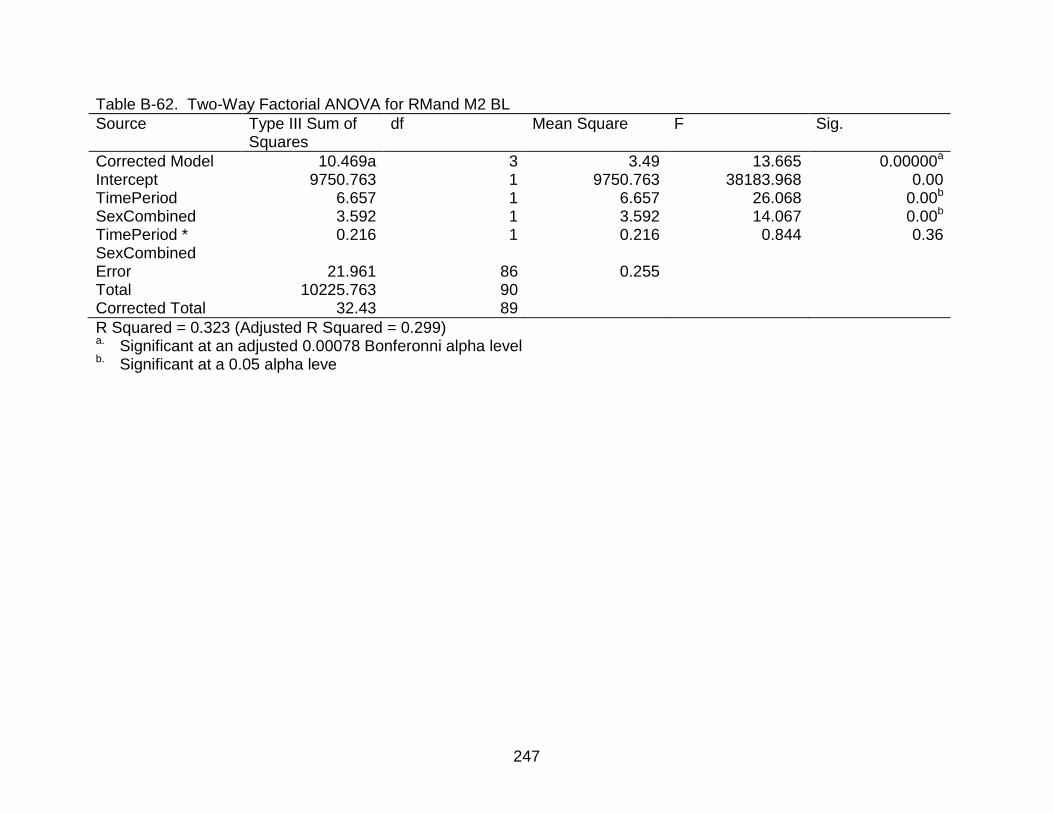
247
Table B-62. Two-Way Factorial ANOVA for RMand M2 BL
Source Type III Sum of Squares
df Mean Square F Sig.
Corrected Model 10.469a 3 3.49 13.665 0.00000a Intercept 9750.763 1 9750.763 38183.968 0.00 TimePeriod 6.657 1 6.657 26.068 0.00b SexCombined 3.592 1 3.592 14.067 0.00b TimePeriod * SexCombined
0.216 1 0.216 0.844 0.36
Error 21.961 86 0.255 Total 10225.763 90 Corrected Total 32.43 89
R Squared = 0.323 (Adjusted R Squared = 0.299) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha leve
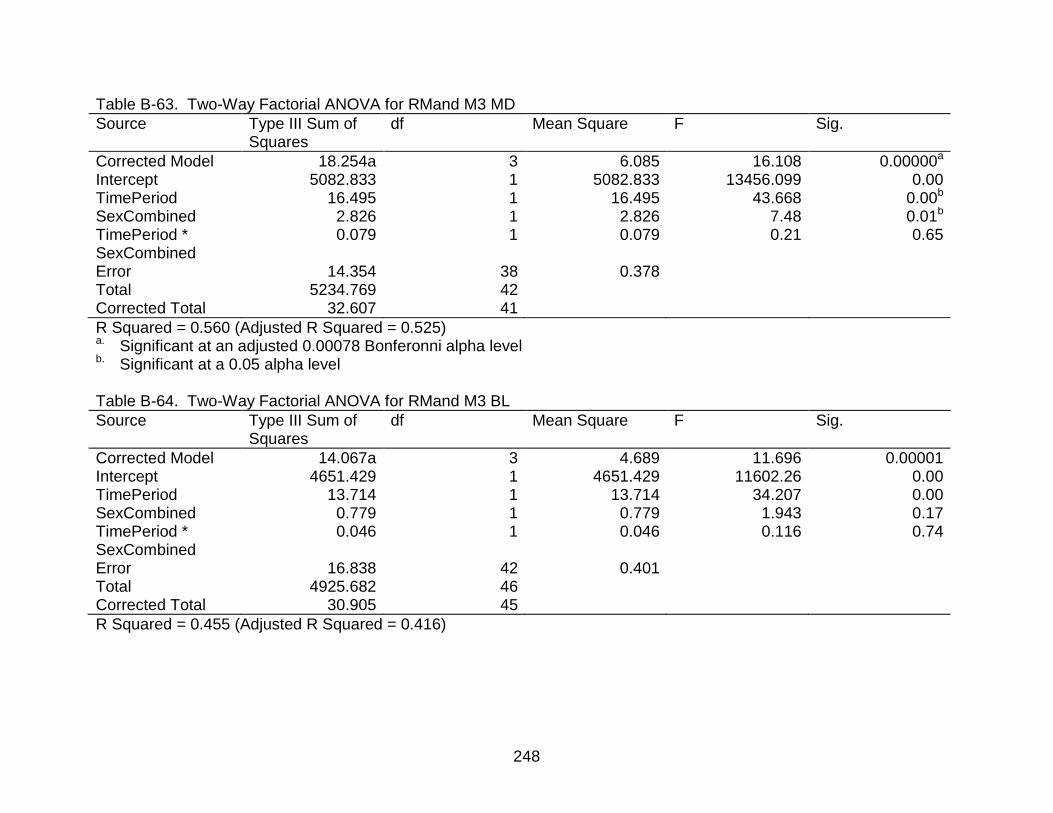
248
Table B-63. Two-Way Factorial ANOVA for RMand M3 MD
Source Type III Sum of Squares
df Mean Square F Sig.
Corrected Model 18.254a 3 6.085 16.108 0.00000a Intercept 5082.833 1 5082.833 13456.099 0.00 TimePeriod 16.495 1 16.495 43.668 0.00b SexCombined 2.826 1 2.826 7.48 0.01b TimePeriod * SexCombined
0.079 1 0.079 0.21 0.65
Error 14.354 38 0.378 Total 5234.769 42 Corrected Total 32.607 41
R Squared = 0.560 (Adjusted R Squared = 0.525) a. Significant at an adjusted 0.00078 Bonferonni alpha level b. Significant at a 0.05 alpha level Table B-64. Two-Way Factorial ANOVA for RMand M3 BL
Source Type III Sum of Squares
df Mean Square F Sig.
Corrected Model 14.067a 3 4.689 11.696 0.00001 Intercept 4651.429 1 4651.429 11602.26 0.00 TimePeriod 13.714 1 13.714 34.207 0.00 SexCombined 0.779 1 0.779 1.943 0.17 TimePeriod * SexCombined
0.046 1 0.046 0.116 0.74
Error 16.838 42 0.401 Total 4925.682 46 Corrected Total 30.905 45
R Squared = 0.455 (Adjusted R Squared = 0.416)

249
LIST OF REFERENCES
Abdi H. 2007. The Bonferonni and Šidák corrections for multiple comparisons. In N Salkind, ed., Encyclopedia of Measurment and Statistics. Thousand Oaks: Sage. p 1-9.
Ambrose SH, Butler BM, Hanson DB, Hunter-Anderson RL, Krueger HW. 1997. Stable isotopic analysis of human diet in the Marianas Archipelago, Western Pacific. Am J of Phys Anthropol 104:343-361.
Amesbury JR. 1999. Changes in species composition of archaeological marine shell assemblages in Guam. Micronesica 31:347-366.
Amesbury JR. 2007. Mollusk collecting and environmental change during the prehistoric period in the Mariana Islands. Coral Reefs 26:947-958.
Amesbury JR, Hunter-Anderson RL, Moore DR. 1991. An archaeological study of the San Antonio Burial Trench and a report on the archaeological monitoring of road construction along Marine Drive between Rts. 8 and 4, Agana, Guam. A Report Prepared for Government of Guam Public Works, Division of Highway Engineering, and Black Construction Corporation, Agana, Guam.
Amesbury JR, Hunter-Anderson RL. 2003. Review of archaeological and historical data concerning reef fishing in the U.S. Flag Islands of Micronesia: Guam and the Northern Mariana Islands. Prepared for Western Pacific Regional Fishery Management Council, Honolulu. Micronesian Archaeological Research Services, Guam.
Amesbury JR, Hunter-Anderson RL. 2008. An analysis of archaeological and historical data on fisheries for the pelagic species in Guam and the Northern Marianas Islands. A report prepared for Pelagic Fisheries Research Program, Joint Institute for Marine and Atmospheric Research, School of Ocean and Earth Science and Technology, University of Hawai’i at Manoa. Micronesian Archaeological Research Services, Guam.
Amesbury JR, Moore DR, Hunter-Anderson RL. 1996. Cultural adaptations and late Holocene sea level change in the Marianas: recent excavations at Chalan Piao, Saipan, Micronesia. Indo-Pacific Prehistory Association Bulletin 15:53-69.
Anderson AJ. 1991. The chronology of colonization in New Zealand. Antiquity. 65:767-795.
Anderson AJ. 1996. Adaptive voyaging and subsistence strategies in the early settlement of East Polynesia. In T Akazawa and EJE Szathmary, eds.: Prehistoric Mongoloid dispersals. Oxford: Oxford University Press. p 359-373.

250
Anderson BE. 1992. Preliminary report on the human skeletal remains from the Gognga-Gun Beach Project, Tumon Bay, Guam. In K Hanihara, ed.: Japanese as a Member of the Asian and Pacific Populations. International Symposium 4. Kyoto, International Research Center for Japanese Studies. p 238-243.
Anderson A. 2005. Crossing the Luzon Strait: archaeological chronology of the Batanes Islands, Philippines and the regional sequence of Neolithic dispersal. J Austrones Stud 1:25–45.
Armelagos GJ, Van Gerven DP. 2003. A century of skeletal biology and paleopathology: contrasts, contradictions, and conflicts. Am Anthropol 105:53-64.
Athens JS, Ward JV. 1993. Paleoenvironment of the Orote Peninsula. In J Carucci, ed.: The archaeology of Orote Peninsula: Phase I and II, archaeological inventory survey of areas proposed for projects to accommodate relocation of Navy activities from the Philippines to Guam, Mariana Islands pp. 153–197. Report prepared for Dept. of the Navy, Pacific Division, Naval Facilities Engineering Command, Pearl Harbor, Hawai’i, under contract to Belt Collins and Associates, Honolulu. Honolulu: International Archaeological Research Institute, Inc.
Athens SJ, Ward JV. 1995. Paleoenvironment of the Orote Peninsula, Guam. Micronesica 28:51–76.
Athens JS, Ward JV. 1999. Paleoclimate, vegetation, and landscape change on Guam: The Laguas Core. In B Dixon, JS Athens, JV Ward, T Mangieri, T Rieth: Archaeological inventory survey of the Sasa Valley and Tenjo Vista Fuel Tank Farms, Piti District, Territory of Guam, Mariana Islands. pp. 121–151. Department of the Navy, Pacific Division, Naval Facilities Engineering Command, Pearl Harbor, Hawaii. International Archaeological Research Institute, Inc., Honolulu.
Athens JS, Ward JV. 2004. Holocene vegetation, savanna origins and human settlement of Guam. Re Aust Mus Supp 29:5–30.
Bath J. 1986. The San Vitores Road Project. Part 1: Final Report. Prepared for Maeda Pacific Corporation and Maeda Road Construction Co., Ltd. Unpublished manuscript.
Beaglehole JC. 1955. The Journals of Captain James Cook on His Voyages of Discovery, Vol. I: The Voyage of the Endeavor 1768-1771. Hakluyt Soc. Extra Ser. NLo. XXXIV. Cambridge: Cambridge University Press.
Bellwood P. 1975. The prehistory of Oceania. Curr Anthropol 16:9-28.
Bellwood P. 1979. The Oceanic context. In JD Jennings, ed.: The prehistory of Polynesia. Cambridge: Harvard University Press. p 6-26.
Bellwood P. 1991. The Austronesian dispersal and the origin of languages. Sci Am 265:88-93.

251
Bellwood P. 1989. The colonization of the Pacific: Some current hypotheses. In AVS Hill, SW Serjeantson, eds.: The Colonization of the Pacific: a genetic trail. Oxford: Clarendon Press. p 1-59.
Bellwood P. 2000. Some thoughts on understanding the human colonization of the Pacific. People Cult Oceania 16:5-17.
Berbesque JC, Doran GH. 2008. Brief Communications: Physiological stress in the Florida Archaic: enamel hypoplasia and patterns of developmental insult in the early North American hunter-gatherers. Am J Phys Anthropol 136:351-356.
Bernal V, Novellino P, Gonzalez PN, Perez SI. 2007. Role fo wild plant foods among Late Holocene Hunter-Gatherers from Central and North Patagonia (South America): an approach from dental evidence. Am J Phys Anthropol 133:1047-1059.
Binford LR. 1962. Archaeology as anthropology. Am Antiq 28:217–225.
Birkby WH. 1977 Report on the teeth recovered from pit 6-1-1 Burial No. 1 (Your No.7-2), Guam, Mariana Islands. In FM Reinman: An Archaeolgical Survey and Preliminary Test Excavations on the Island of Guam, Mariana Islands, 1965-1966. Miscellaneous Publication No. 1.
Blakely RL. 1977a. Biocultural adaptation in prehistoric America, Southern Anthropological Society Proceedings, No. 11. Athens: The University of Georgia Press.
Blakely RL. 1977b. Introduction: changing strategies for the biological anthropologist. In RL Blakey ed.: Biocultural Adaptation in Prehistoric America, 1–9, Southern Anthropological Society Proceedings, No. 11. Athens: The University of Georgia Press.
Blust R. 2000. Chamorro historical phonology. Oceanic Linguist 39:83-122.
Blust R. 2009. The Austronesian languages (Pacific Linguistics 602). Canberra: Department of Linguistics, Research School of Pacific and Asian Studies, The Australian National University.
Boldsen JL. 2007. Early childhood stress and adult age mortality: a study of dental enamel hypoplasia in the medieval Danish village of Tirup. Am J Phys Anthropol 132:59–66.
Bonhomme T, Craib JL. 1987. Radiocarbon dates from Unai Bapot, Saipan: implications for the prehistory of the Mariana Islands. J Polynes Soc 96:95-105.
Brace CL. 1963. Structural reduction in evolution. Am Nat 97:39-49.
Brace CL. 1964. The probable mutation effect. Ame Natural 98:453-455.

252
Brace CL. 1967. Environment, tooth form and size in the Pleistocene. J Dent Res 46:809-816.
Brace CL. 1978. Tooth reduction in the Orient. Asian Persp 19:203-219.
Brace CL. 1979. Krapina, “Classic” Neanderthals, and the evolution of the European face. J Hum Evol 8:527-550.
Brace CL. 1980. Australian tooth-size clines and the death of a stereotype. Curr Anthropol 21:141-164.
Brace CL, Hinton, RJ. 1981. Oceanic tooth-size variations a reflection of biological and cultural mixing. Curr Anthropol 22:549-569.
Brace CL, Mahler PE. 1971. Post-Pleistocene changes in the human dentition. Am J Phys Anthropol. 34:191-204.
Brace CL, Rosenberg KR, Hunt KD. 1987. Gradual change in human tooth size in the late Pleistocene and early Holocene. Evolution. 41:705–720.
Brace CL, Brace ML, Dodo Y, Leonard WR, Yongyi L, Sangvichien S, Xiangqing S, Zhenbiao Z. 1990. Micronesians, Asians, Thais and relations: a craniofacial and odontometric perspective. Micronesica Supp 2:323-348.
Braidwood RJ. 1960. The agricultural revolution. Sci Am 203:130-148.
Bridgman HA 1983. Could climatic change have had an influence on the Polynesian migrations? Palaeogeogr Palaeoclimatol Palaeoecol 41: 193-206.
Buikstra JE. 1977. Biocultural dimensions of archaeological study: a regional perspective. In R Blakely ed.: Biocultural adaptation in prehistoric America. Southern Anthropological Society Proceedings No. 11. Athens: University of Georgia Press. p 67–84.
Buikstra JE, Beck LA. 2006. Bioarchaeology: The contextual analysis of human remains. Academic Press: New York.
Buikstra JE, Ubelaker DH. 1994. Standards for data collection from human skeletal remains: proceedings of a seminar at the Field Museum of Natural History. Arkansas Archeological Survey Research Series; no. 44. Arkansas Archeological Survey, Fayetteville, Ark.
Butler BM. 1994. Early prehistoric settlement in the Mariana Islands: new evidence from Saipan. Man and Cult in Ocean 10:15-38.

253
Butler BM.1995. Archaeological investigations in the Achugao and Matansa areas of Saipan, Mariana Islands. Micronesian Archaeological Survey Report, No. 30. Division of Historic Preservation, Department of Community and Cultural Affairs, CNMI, Saipan.
Buzon MR. 2005. Health of the non-elites at Tombos: nutritional and disease stress in New Kingdom Nubia. Am J Phys Anthropol 130:26-37.
Callaghan R, Fitzpatrick SM. 2008. Examining prehistoric migration patterns in the Palauan archipelago: a computer simulated analysis of drift voyaging. Asian Persp 47:28-44.
Campos FZ. 2009. The ichthyoarchaeology of Batanes Islands, Northern Philippines. M.A. Thesis, Department of Archaeology, University of Philippines.
Carson MT. 2008. Refining earliest settlement in Remote Oceania: renewed archaeological investigation at Unai Bapot, Saipan. J of Island and Coast Arch 3:115–139.
Carson MT. 2010. Radiocarbon chronology with marine reservoir correction for the Ritidian archaeological site, northern Guam. Radiocarbon 52:1627–1638.
Carson MT. 2011. Palaeohabitat of first settlement sites, 1500–1000 B.C. in Guam, Mariana Islands, western Pacific. J Archeol Sci 38: 2207–2221.
Carson MT, Welch D. 2005. Archaeological survey, mapping, and testing of Bapot Latte Site (SP-1-0013) in Laulau, Saipan, Commonwealth of the Northern Mariana Islands. Report prepared for the Commonwealth of the Northern Mariana Islands Division of Historic Preservation, Saipan, CNMI.
Chandra S, Desai VM. 1970. Relationship of betel chewing and dental caries. J Ind Dent Assoc 42:269– 276.
Chatrchaiwiwatana S. 2006. Dental caries and periodontics associated with betel quid chewing: analysis of two data sets. J Med Assoc Thai 89:1004-1010.
Chu NS. 2001. Effects of betel chewing of the central and autonomic nervous system. J Biomed Sci 8:229-236.
Chu NS. 2002. Neurological aspects of areca and betel chewing. Addic Biol 7:111-114.
Clark G. 2005. A 3000-year culture sequence from Palau, Western Micronesia. Asian Persp 44:349–380.
Clark G, Petchey F, Winter O, Carson MT, O’Day P. 2010. New radiocarbon dates from the Bapot-1 site in Saipan an dNeolithic dispersal by stratified diffusion. J Pac Archaeol 1:21-35.

254
Clark JGD. 1972. Starr Carr: A case study in bioarchaeology, Modular Publications 10. London: Addison-Wesley.
Cloud P, Schmidt RG, Burke HW. 1956. Geology of Saipan, Marianas Islands. Part I. General Geology. Geological Survey Professional Paper 280-A. Washington, DC.
Cohen MN, Armelagos GJ. 1984. Paleopathology and the Origins of Agriculture. New York: Academic Press.
Collins SK & DM Pearsall. 2001a. Phytolith analysis. In RL Hunter-Anderson et al., Cultural resources survey of five Navy surplus Guam land use plan parcels, Territory of Guam, Mariana Islands. Department of the Navy, Pacific Division, Naval Facilities Engineering Command, Pearl Harbor, Hawaii. International Archaeological Research Institute, Inc., Honolulu. p E-1-E-14.
Collins SK, Pearsall DM. 2001b. Report of phytolith analysis. In RL Hunter-Anderson: Archaeological investigations at Ngachang, Ordot-Chalan Pago Municipality, Guam. Micronesian Archaeological Research Services, Guam. p 60–68.
Conrad CD, Galea LA, Kuroda Y, McEwen BS. 1996. Chronic stress impairs rat spatial memory on the Y maze, and this effect is blocked by tianeptine pretreatment. Behav Neurosci 110:1321–1334.
Cook DC. 1984. Subsistence and health in the Lower Illinois Valley: osteological evidence. In MN Cohn and GJ Armelagos, eds.: Paleopathology at the Origins of Agriculture. Orlando: Academic Press. p 235-269.
Cook DC, Buikstra JE. 1979. Health and differential survival in prehistoric populations: prenatal dental defects. Am J Phys Anthropol 51:649–664.
Craib JL. 1986. Casas de los antiguos: social differentiation in Protohistoric Chamorro society, Mariana Islands. Doctoral dissertation. University of Sydney, Australia.
Craib JL. 1993. Early occupation at Unai Chulu, Tinian, Commonwealth of the Northern Mariana Islands. Bulletin of the Indo-Pacific Prehistory Association 13:116-134.
Craib JL. 1998. Archaeological surveys of military lease lands on Tinian for the partial mitigation of adverse effects in conjunction with the FY93 Tandem Thrust Training Exercise. Prepared for the Department of the Navy, Pacific Division, Naval Facilities Engineering Command, Pearl Harbor, Hawai‘i. Ogden Environmental and Energy Services Co., Inc., Honolulu.
Craib JL. 1999. Colonisation of the Mariana Islands: new evidence and implications for human movements in the Western Pacific. In JC Galipaud, I Lilley: The Pacific from 5000 to 2000 BP, Colonisation and Transformations. IRD Editions:Paris. p 477-487.

255
Crowther AM, Halsam TH, Loy TH. 2003. Residue analysis of ten potsherds from Guam. Report Number: MARS 390. University of Queensland, St. Lucia, Australia.
Cucina A. 2002. Diachronic investigation of linear enamel hypoplasia in prehistoric skeletal samples from Trentino, Italy. Am J Phys Anthropol 119:283–287.
Cummings LS, Puseman K. 1998. Pollen, phytolith, and macrofloral analysis at sites in the NAVACTS Ordnance Annex. In JD Henry, AE Haun, MA Kirkendall, DG DeFant: Vol II: Appendices, Final report Phase I archaeological survey and subsurface testing, U.S. Naval Activities Ordnance Annex, Guam. Department of the Navy, Pacific Division, Naval Facilities Engineering Command, Pearl Harbor, Hawaii. Paul H. Rosendahl, Ph.D., Inc., Hilo, Hawaii. p L-1-L-37.
Cummings LS. 2002. Pollen analysis, Guam Ordnance Annex. In J Allen, B Dixon, K Aronson, D Gosser, L Gilda, T Moorman, C O’Hare, M Riford, J Toenjes: Final report, archaeological survey and excavations at Naval Ordnance Annex, Territory of Guam. Department of the Navy, Pacific Division, Naval Facilities Engineering Command, Pearl Harbor, Hawaii. Ogden Environmental and Energy Services Co., Inc., Honolulu. p E-1-E-9.
Cunha E, Rozzi FR, Bermudez de Castro JM, Martinon-Torres M, Wasterlain SN, Sarmiento S. 2004. Enamel hypoplasias and physiological stress in the Sima de los Huesos Middle Pleistocene Hominins. Am J Phys Anthrop. 125:220-231.
Cybulski JS. 1974. Tooth wear and material culture: Precontact patterns in the Tsimshian area, British Columbia. 7: 31-35.
Davidson JM. 1984. The Prehistory of New Zealand. London: Longman.
Davis BD, Tomonari-Tuggle MJ, Wickler S, Favreau CK. 1992 Archaeological investigations at the Leo Palace Hotel Site, Naton Beach, Tumon Bay. Vol. I: Archaeological Data Recovery, Burial Recovery, and Monitoring. Prepared for Sumitomo Construction Company, Tamuning, Guam. International Archaeological Research Institute, Inc., Honolulu.
DeFant DG. 2008. Early human burials from the Naton Beach Site, Tumon Bay, Island of Guam, Mariana Islands. J of Island and Coastal Archaeology. 3: 149-153.
Delgado-Darias T, Velasco-Vazques J, Arnay de la Rosa M, Martin-Rodriguez E, Gonzalez-Reimer E. 2005. Dental caries among the Prehispanic populations from Gran Canaria. Am J Phys Anthropol 128:560-568.
Dickinson WR. 2000. Hydro-isostatic and tectonic influences on emergent Holocene paleoshorelines in the Mariana Islands, western Pacific Ocean. J Coast Res 16:735-746.
Dickinson WR. 2001. Paleoshoreline record of relative Holocene sea levels on Pacific islands. Earth Sci Rev 55:191-234.

256
Dickinson 2003. Impact of mid-Holocene hydro-isostatic highstand in regional sea level on habitability of islands in Pacific Oceania. J Coast Res 19:489-502.
Dilli BJ, Haun AE, Goodfellow ST, DeRoo B. 1998. Volume II: Data analyses, archaeological mitigation program, Mangilao Golf Course Project Area, Mangilao Municipality, Territory of Guam. Prepared for Mr. Jetan Sahni, Guam. Prepared by Paul H. Rosendahl, PhD, Inc, Hilo, Hawaii.
DePaola DP. 1982. The influence of food carbohydrates on dental caries. In DR Lineback, GE Inglett, eds.: Food carbohydrates. Westport: AVI.
DeRoo B, Goodfellow ST. 1998. Section IV-II, Ceramic Analysis. In Dilli et al.: Volume II: Data analyses, archaeological mitigation program, Mangilao Golf Course project area, Mangilao Municipality, Territory of Guam. Paul H. Rosendahl, Ph.D., Inc., Hilo.
Dixon BJ, Athens S, Ward JV, Mangieri T, Rieth T. 1999. Archaeological inventory survey of the Sasa Valley and Tenjo Vista Fuel Tank Farms, Piti District, Territory of Guam, Mariana Islands. Prepared for Department of the Navy, Pacific Division, Naval Facilities Engineering Command, Pearl Harbor, Hawaii. Prepared by International Archaeological Research Institute, Inc, Honolulu.
Domett K, Tayles N. 2007. Population health from the Bronze to the Iron Age in the Mun river valley, Northeastern Thailand. In MN Cohen and G Crane-Kramer, eds.: Ancient Health. Gainesville: University Press of Florida. p 286-299.
Doods ML. 1960. The cariogenic property of cereal foods. J Nutrition 71:317-321.
Douglas MT and Ikehara RM. 1992. Archaeological Investigations at the Leo Palace Hotel Site, Naton Beach, Tumon Bay. Volume II: Osteological study of prehistoric Chamorro skeletal remains. Honolulu: International Archaeological Research Institute, Inc. Unpublished manuscript.
Douglas MT, Pietrusewsky M, Ikehara-Quebral RM. 1997. Skeletal biology of Apurguan: A precontact Chamorro site on Guam. Am J Phys Anthrop 104:291-313.
Douglas MT, Pietrusewsky M. 2007. Biological consequences of sedentism: agricultural intensification in Northeast Thailand. In: Cohen MN, Crane-Kramer GMM, editors. Ancient health: skeletal indicators of agricultural and economic intensification. Gainesville: University Press of Florida. p 300–319.
Doyle WJ, Kelley C, Siegel MI. 1977. The effects of audiogenic stress on the growth of long bones in the laboratory rat. Growth 41:183-189.
Driver MC. 1988. Cross, sword, and silver: the nascent Spanish colony in the Mariana Islands. Pacific Studies 11:21-313.

257
Driver MG. 1993. Fray Juan Pobre de Zamora and his account of the Mariana Islands. The J Pac Hist 18:198–216.
Driver MG. 1991. An account of the Islands of the Ladrones. The J Pac Hist 26:103-106.
d’Urville DJSC. 1832. Sur les îles du grand ocean. Bulletin de la Societé de Géographie 17:1-21.
Duray SM. 1996. Dental indicators of stress and reduced age at death in prehistoric Native Americans. Am J Phys Anthropol 99:275–286.
Dyen I. 1965. A lexicostatistical classification of the Austronesian languages. Int J Am Linguist. Memoir 19.
Easton WH, Ku TL, Randall RH. 1978. Recent reefs and shore lines of Guam. Micronesica 14:1–11.
FDI. 1982. An epidemiological index of developmental defects of dental enamel (DDE Index). Int Dent J 32:159–167.
Fitzpatrick SM. 2003. Early human burials in the western Pacific: evidence for a c.3000 year old occupation on Palau. Antiquity 77:719–731.
Fitzpatrick SM, Nelson GC. 2008. From limestone caves to concrete graves: 3000 years of mortuary practice in the Palauan archipelago, Micronesia. Int J Osteoarchaeol 18:439–457.
Fitzpatrick SM, Nelson GC. 2011. Purposeful commingling of adult and child cranial elements from the Chelechol ra Orrak Cemetery, Palau. Int J Osteoarchaeol 21:360-366.
Foss LB, Davidson JM. 2006a. Analysis of faunal material from an archaeological site complex at Mangilao, Guam. Technical Report 38, Museum of New Zealand Te Papa Tongarewa.
Foss LB, Davidson JM. 2006b. Analysis of faunal material from an archaeological site at Ylig, Guam. Technical Report 39, Museum of New Zealand Te Papa Tongarewa.
Frencken JE, Rugarabamu P, Mulder J. 1989. The effect of sugar cane chewing on the development of caries. J Dent Res. 68:1102-1104.
Fulmer J, Kirkendall MA, DeFant DB, Haun AE. 1999. Archaeological subsurface testing, Sumay Village Site, Fort Santiago Site, and the Orote Point Cave Site, US Naval Activities (NAVACTS) Waterfront Annex, Santa Rita Municipality, Territory of Guam. Prepared for Commander, Pacific Division, Naval Facilities Engineering Command. Paul H. Rosendahl, PhD, Inc, Hilo.

258
Gingerich SB. 2003. Hydrologic Resources of Guam. Water-Resources Investigation Report 03-4126. US Geological Survey, Honolulu.
Goodenough WH. 1957. Oceania and the problem of controls in the study of cultural and human evolution. The J Polynesian Soc. 66:146-155.
Goodman AH. 1991. Health, adaptation, and maladaptation in past societies. In H Bush and M Zvelebil: Health in Past Societies: Biocultural Interpretations of Human Skeletal Remains in Archaeological Contexts. British Archaeological Reports, International Series 567:31-38.
Goodman AH, Armelagos GJ. 1985. Factors affecting the distribution of enamel hypoplasias within the human permanent dentition. Am J Phys Anthropol 68:479–493.
Goodman AH, Armelagos GJ. 1989. Infant and childhood morbidity and mortality risks in archaeological populations. World Arch 21:225-243.
Goodman AH, Martin DL. 2002. Reconstructing health profiles from skeletal remains. In RH Steckel and JC Rose, eds.: The Backbone of History: Health and Nutrition in the Western Hemisphere. Cambridge University Press: Cambridge. p 11-60.
Goodman AH, Rose JC. 1990. Assessment of systemic physiological perturbations from dental enamel hypoplasias and associated histological structures. Yearb Phys Anthropol 33:59–110.
Goodman AH, Rose JC. 1991. Dental enamel hypoplasias as indicators of nutritional status. In MA Kelley and CS Larsen, eds.: Advances in Dental Anthropology. Wiley-Liss, New York. p 279-293.
Goodman AH, Armelagos GJ, Rose JC. 1980. Enamel hypoplasias as indicators of stress in three prehistoric populations from Illinois. Hum Biol 52:512–528.
Goodman AH, Thomas RB, Swedlund AC, Armelagos GJ. 1988. Biocultural perspectives on sress in prehistoric, historical, and contemporary population research. Yearb Phys Anthropol. 31:169-202.
Grant MP, Knutsson I, Stodder ALW. 1992. Archaeological Monitoring and Data Recovery AT&T Tumon Bay Cable Easement, Manhole, and Conduit System. Report prepared for AT&T Communications. PHRI Inc. Unpublished manuscript.
Graves MW. 1986. Organization and Differentiation Within Late Prehistoric Ranked Social Units, Mariana Islands, Western Pacific. Journal of Field Archaeol 13:139-154.

259
Graves MW, Moore DR. 1985. Tumon Bay Area Overview: Cultural and Historical Researches. A report prepared for the Historic Preservation Office, Department of Parks and Recreation, Government of Guam. Micronesian Area Research Center and Department of Anthropology, University of Guam, Mangilao.
Green RC. 1979. Lapita. In J Jennings, ed.: The prehistory of Polynesia. Harvard University Press, Cambridge. p 27-60.
Green RC. 1991a. Near and remote Oceania – disestablishing “Melanesia” in culture history. In A Pawley ed.: Man and a Half. Essays in Pacific Anthropology and Ethnobiology in Honour of Ralph Bulmer. Auckland: The Polynesian Society. p 591-502.
Green RC. 1991b. A reappraisal of the dating for some Lapita sites in the Reef/Santa Cruz group of the Southeast Solomon Islands. J Polynesian Soc. 100:197-207.
Green RC. 1999. Integrating historical linguistics with archaeology: insights from research in Remote Oceania. Indo-Pac Prehist Assoc Bull 18:3-16.
Grenby TH. 1997. Summary of the dental effects of starch. Int J Food Sci Nut 48:411-416.
Griffin MC, Lambert PM, Monahan-Driscoll E. 2001. Biological relationships and population history of native peoples in Spanish Florida and the American Southeast. In: Larsen CS, editor. Bioarchaeology of Spanish Florida: The impact of colonialism. Gainesville: University Press of Florida. p 226– 273.
Grivet L, Glaszmann JC, D’Hont A. 2004. Molecular genetics evidence for sugarcane evolution and domestication. Ethnobot Res App 2:9-17.
Guatelli-Steinberg D. 2003. Macroscopic and microscopic analysis of linear enamel hypoplasia in Plio-Pleistocene South African hominins with respect to aspects of enamel development and morphology. Am J Phy Anthropol 12:309-322.
Guatelli-Steinberg D, Benderlioglu Z. 2006. Brief communication: linear enamel hypoplasia and the shift from irregular to regular provisioning in Cayo Santiago rhesus monkeys (Macaca mulatta). Am J Phy Anthropol 131:416-119.
Guatelli-Steinberg D, Lukacs JR. 1999. Interpreting sex differences in enamel hypoplasia in human and non-human primates: developmental, environmental, and cultural considerations. Yearb Phys Anthropol 42:73–126.
Guatelli-Steinberg D, Larsen CS, Hutchinson DL. 2004. Prevalence and the duration of linear enamel hypoplasia: a comparative study of Neandertals and Inuit foragers. J Hum Evol 47:65–84.
Hanihara T. 1992. Dental and cranial affinities among populations of East Asia and the Pacific: The basic populations in East Asia, IV. Am J Phys Anthropol. 88:163-182.

260
Hanihara T, Ishida H. 2005. Metric dental variation of major human populations. Am J Phys Anthropol 128:287–298.
Hanson DB. 1988. Prehistoric mortuary practices and human biology. In BM Butler, ed.: Archaeological investigations on the North Coast of Rota, Mariana Islands. Micronesian Archaeological Survey Report No. 23, Center for Archaeological Investigations Occasional Paper No. 8. Carbondale: Southern Illinois University at Carbondale. p 375–435.
Hanson DB, Butler BM. 1997. A biocultural perspective on Marianas prehistory: Recent trends in bioarchaeological research. Am J Phys Anthropol. 104:271-290.
Hanson DB, Gordon CC. 1989. Mortuary practices and social complexity in Micronesia: problems and prospects of an emerging archaeological database. Man and Cult in Ocean 5:37–66.
Hartnady P, Rose JC. 1991. Abnormal tooth-loss patterns among Archaic-period inhabitants of the lower Pecos region, Texas. In MA Kelley, CS Larsen, eds.: Advances in dental anthropology. New York: Wiley-Liss. p 267-278.
Han SS, Baek KW, Shin MH, Kim J, Oh CS, Lee SJ. 2010. Dental caries prevalence of medieval Korean people. Arch Oral Biol 55:535–40.
Haun AE, Jimenez JA, Kirkendall MA, Goodfellow ST. 1999. Archaeological investigations at Unai Chulu, Island of Tinian, Commonwealth of the Northern Mariana Islands. Prepared for Department of the Navy, Commander, Pacific Division, Naval Facilities Engineering Command, Pearl Harbor, Hawai`i. Paul H. Rosendahl, Inc., Hilo, Hawai’i.
Hezel FX.1982 From Conquest to colonization: the early Spanish mission in the Marianas. J Pac Hist 17:115-137.
Heyerdall T. 1952. American Indians of the Pacific. London: Allen and Unwin.
Hill K, Hurtado AM. 1989. Hunter-gatherers of the New World. Am Sci 77:436-443.
Hillier RJ, Craig GT. 1992. Human dental enamel in the determination of health patterns in children. J Paleopathol 2:381-390.
Hillson S. 1979. Diet and dental disease. World Archaeol 2:147– 162.
Hillson S. 1996. Dental anthropology. Cambridge: Cambridge University Press.
Hodges DC. 1987. Health and the agricultural intensification in the prehistoric valley of Oaxaca, Mexico. Am J Phys Anthrop 73:323-332.
Houghton P. 1991a. Selective influences and morphological variation amongst Pacific Homo sapiens. J Human Evol 21:49-59.

261
Houghton P. 1991b. The early human biology of the Pacific; some considerations. J Polynesian Soc. 100: \167-196.
Houghton P. 1996. People of the great ocean. Cambridge: University Press.
Howden GF. 1984. The cariostatic effect of betel nut chewing. PNG Med J 27:123– 131.
Howells WW. 1970. Anthropometric grouping analysis of Pacific peoples. Archeol Phys Anthropol Ocean 5:192-217.
Howells WW.1973. The Pacific Islanders. Wellington: AH & AW Reed.
Howells WW. 1977. The sources of human variation in Melanesia and Australia. In J Allen, R Golson, R Jones, eds.: Sunda and Sahul: prehistoric studies in Southeast Asia, Melanesia, and Australia. London: Academic Press. p 169-186.
Howells WW. 1979. Physical anthropology. In JD Jennings ed.: The Prehistory of Polynesia. Canberra: The Australian National University. p 271-285.
Hung H, Carson MT, Bellwood P, Campos FZ, Piper PJ, Dizon E, Bolunia MJLA, Oxenham M, Chi Z. 2011. The first settlement of Remote Oceania: the Philippines to the Marianas. Antiquity. 85:909-926.
Hunter-Anderson RL, BM Butler. 1991. An Overview of Northern Marianas Prehistory. Micronesian Archaeological Research Services, Guam.
Hunter-Anderson RL, Moore DR. 2002. Phase I and phase II archaeological survey at Waterfront Annex and Ordnance Annex, Territory of Guam. Report prepared for International Archaeological Research Institute, Inc., Honolulu, under contract to Department of the Navy, Pacific Division, Pearl Harbor. Micronesian Archaeological Research Services, Inc., Mangilao.
Hunter-Anderson RL, Moore DR, Amesbury JR. 1994. The Manenggon Hills archaeological project. In RL Hunter-Anderson, ed.: Volume I, Project history, research design, methods of site discovery, data recovery, and analysis. MDI Guam Corporation, Yona, Guam. Micronesian Archaeological Research Services, Mangilao, Guam. p 1.1–1.100.
Hunter-Anderson RL, Thompson GB, Moore DR. 1995. Rice as a prehistoric variable in the Mariana Islands, Micronesia. Asian Persp 34:69-89.
Hunter-Anderson, RL, Amesbury JR, Moore DR. 1998. Results of monitoring and data recovery at Naton Beach, Tumon Bay, Guam. Prepared for Architectural-Engineering Services, Inc. and Tanota Partners, Inc., Tumon, Guam. Prepared by Micronesian Archaeological Research Services, Guam.

262
Hunter-Anderson RL, Dixon, B, Mangieri. 2001. Cultural resources survey of five Navy surplus Guam land use parcels, Territory of Guam. Department of the Navy, Pacific Division, Naval Facilities Engineering Command, Pearl Harbor, Hawaii. International Archaeological Research Institute, Inc., Honolulu.
Hurles ME, Nicholson J, Bosch E, Renfrew C, Sykes BC, Jobling MA. 2002. Y chromosomal evidence for the origins of Oceanic-speaking peoples. Genetics. 160:289-303.
Ikehara RM and Douglas MT. 1997. Cultural alteration of human teeth in the Mariana Islands. Am J Phys Anthrop. 104:381-391.
Irwin G. 1992. The Prehistoric Exploration and Colonization of the Pacific. Cambridge: Cambridge University Press.
Ishida H, Dodo Y. 1997. Cranial variation in prehistoric human skeletal remains from the Marianas. Am J Phys Anthropol 104: 399-410.
Kayanne H, Ishii T, Matsumoto E, Yonekura N. 1993. Late Holocene sea-level change on Rota and Guam, Mariana Islands, and its constraint on geophysical predictions. Quat Res 40:189-200.
Kedjarune U, Migasena P, Changbumrung S, Pongapaew P, Tungtronchitr R. 1997. Flow rate and composition of whole saliva in children from rural and urban Thailand with different caries prevalence and dietery intake. Caries Res 31:148-157.
Keegan WF, Diamond, JM. 1987. Colonization of islands by humans: a biogeographical perpective. Adv Arch Method Theory. 10:49-92.
Keita SOY. 2003. A study of vault porosities in early Upper Egypt from the Badarian through Dynasty I. World Archaeol 35:210–222.
Keita SOY, Boyce AJ. 2001. Diachronic patterns of dental hypoplasias and vault porosities during the predynastic in the Naqada region, Upper Egypt. Am J Hum Biol 13:733–743.
Kelley MA, Levesque DR, Weidl E. 1991. Contrasting patterns of dental disease in five early northern Chilean groups. In: Kelley MA, Larsen CS, eds.: Advances in dental anthropology. New York: Wiley-Liss. p 179–202.
King T, Humphrey LT, Hillson SW. 2005. Linear enamel hypoplasias as indicators of systemic physiological stress: evidence from two known age-at-death and sex populations from post-medieval London. Am J Phys Anthropol 128:547–559.
Kirch PV. 1997. The Lapita Peoples: Ancestors of the Oceanic World. Oxford: Blackwell Scientific.

263
Kirch PV. 2000. On the Road of the Winds. Berkeley: University of California Press.
Kirch PV. 2010. Peopling of the Pacific: A holistic anthropological perspective. Annu Rev Anthropol. 39:131-148.
Kirch PV, Hunt TL. 1988. The spatial and temporal boundaries of Lapita. In PV Kirch and TL Hunt, eds.: Archaeology of the Lapita cultural complex: a critical review. Seattle: Thomas Burke Memorial Washington State Museum Research Report 5. p 9-31.
Krasse B. 1985. The cariogenic potential of foods—a critical review of current methods. Int Dent J 35:36-42.
Kurashina H, Clayshulte RN. 1983. Site formation processes and cultural sequence at Tarague, Guam. IPPA Bulletin No. 4. Canberra: Australian National University Press.
Lanfranco LP, Eggers S. 2010. The usefulness of caries frequency, depth, and location in determining cariogenicity and past subsistence: a test on early and later agriculturalists from the Peruvian coast. Am J Phys Anthrop 143:75-91.
Lanphear KM. 1990. Enamel hypoplasias in a historic skeletal sample. Am J Phys Anthropol 81:37-43.
Larsen CS. 1981. Skeletal and dental adaptations to the shift to agriculture on the Georgia Coast population. Curr Anthropol 22:422–423.
Larsen CS. 1983. Behavioural implications of temporal change in cariogenesis. J Archaeol Sci 10:1–8.
Larsen CS. 1984. Health and disease in prehistoric Georgia: the transition to agriculture. In MN Cohn and GJ Armelagos, eds.: Paleopathology at the Origins of Agriculture. Orlando: Academic Press. p 367-392.
Larsen CS. 1995. Biologcial changes in human populations with agriculture. Annu Rev Anthropol 24:185-213.
Larsen CS. 1997. Bioarchaeology: Interpreting Behavior from the Human Skeleton. Cambridge University Press: Cambridge.
Larsen CS, Shavit R, Griffin MC. 1991. Dental caries as evidence for dietary change: an archaeological context. In: Kelley MA, Larsen CS, eds.: Advances in dental anthropology. New York: Wiley-Liss. p 179–192.
Larsen CS, Teaford MF, Sandford MK. 1998. Teeth as tools at Tutu: Extramasticatory behavior in prehistoric St. Thomas, U.S. Virgin Islands. In JR Lukacs, ed., Human Dental Development, Morphology, and Pathology: A Tribute to Albert A. Dahlberg. Eugene: University of Oregon Anthropological Papers: Number 54).

264
Larsen CS, Crosby AW, Griffin MC, Hutchinson DL, Ruff, CB, Russell KR, Schoeninger MJ, Sering LE, Simpson SW, Takacs JL, Teaford MF. 2002. A biohistory of health and behavior in the Georgia Bight: the agricultural transition and the impact of European contact. In: Steckel RH, Rose JC, editors. The backbone of history: health and nutrition in the western hemisphere. Cambridge: Cambridge University Press. p 406-439.
Leigh RW. 1929. Dental morphology and pathology of prehistoric Guam. Memoirs of the Bernice P. Bishop Museum 11:1–18.
Li, K. 2002. Prehistoric marine fishing adaptation in southern Taiwan. J East Asian Archaeol 3:47-74.
Li, K. 1997. Change and stability in the dietary system of a prehistoric coastal population in Southern Taiwan. Doctoral dissertation, Arizona State University.
Liston J, Amesbury JR, Ikehara-Quebral R, Moore D, Olmo RK, Wells EF. 1996 The legacy of Tarague embayment and its inhabitants, Andersen AFB, Guam. Vol. I: Archaeology. International Archaeology, Inc., Honolulu, HI.
Loy TH. 2001a. Cooking residues on four potsherds, Nimitz Hill, Guam. Appendix B in Phase II Archaeological Survey: Detailed Recording of Sites on the Lands to be Retained by the Navy, Nimitz Hill, Territory of Guam, Mariana Islands, by Moore DR, Hunter-Anderson RL, p B-1-B-30. Prepared by Micronesian Archaeological Research Services, Guam, for International Archaeological Research Institute, Inc., Honolulu, under contract with the Department of the Navy, Pacific Division, Naval Facilities Engineering Command, Pearl Harbor, Hawaii.
Loy TH. 2001b. Analysis of six pottery sherds from the west coast of Guam. Appendix F in Hunter-Anderson RL et al.: Cultural resources survey of five navy surplus Guam land use plan parcels, Territory of Guam, Mariana Islands., p F-1-F-21. Prepared by Micronesian Archaeological Research Services, Guam, and International Archaeological Research Institute, Inc., Honolulu, under contract with the Department of the Navy, Pacific Division, Naval Facilities Engineering Command, Pearl Harbor, Hawaii.
Loy TH, Crowther A. 2002. Analysis of pottery sherds from two archaeological sites: Guam. In DR Moore: Guam’s prehistoric pottery and its chronological sequence. Prepared for the US Department of the Navy, Pacific Division by Micronesian Archaeological Research Services, Guam p 125-158.
Luine V, Villegas M, Martinez C, McEwen BS. 1994. Repeated stress causes reversible impairments of spatial memory performance. Brain Res 639:167–170.
Lukacs JR 1989. Dental paleopathology: methods for reconstructing health status and dietary patterns in prehistory. In MY İşcan and KAR Kennedy, eds: Reconstructing Life from the Skeleton. New York: Alan R. Liss. p 261-286.

265
Lukacs JR. 1996. Sex differences in dental caries with the origin of agriculture in South Asia. Curr Anthropol 37:147– 153
Lukacs JR. 1992. Dental paleopathology and agricultural intensification in South Asia: new evidence. Am J Phys Anthropol 87:133–150.
Lukacs JR. 1999. Enamel hypoplasia in deciduous teeth of great apes: Do differences in defect prevalence imply differential levels of physiological stress? Am J Phys Anthropol 110:351–363.
Lukacs JR. 2001. Enamel hypoplasia in the deciduous teeth of great apes: variation in prevalence and timing of defects. Am J Phys Anthropol 116:199–208.
Lukacs JR, Walimbe. 1998. Physiological stress in prehistoric India: new data on localized hypoplasia of primary canines linked to climate and subsistence change. J Arch Sci 25:571-583.
Lum JK, Cann RL. 1998. mtDNA and language support a common origin of Micronesians and Polynesians in Island Southeast Asia. Am J Phys Anthropol 105:109-119.
Lum JK, Cann RL. 2000. mtDNA lineage analyses: origins and migrations of Micronesians and Polynesians. Am J Phys Anthropol 113:151-168.
Lum JK, Jorde LB, Schiefenhovel W. 2002. Affinities among Melanesians, Micronesians, and Polynesians: A neutral, biparental, genetic perspective. Hum Biol 74:413-430.
Maat GJR, Van der Velde. 1987. The caries-attrition competition. Int J Anthropol 2:282-292.
Magnuson CM, Olmo R, Welch DJ. 2000. archaeological survey and data recovery investigations, Duty Free Shoppers expansion site,Tumon, Guam. International Archaeological Research Institute, Inc., Honolulu.
Malville NJ. 1997. Enamel hypoplasia in ancestral Puebloan populations from southwestern Colorado. I. Permanent dentition. Am J Phys Anthropol 102:351–367.
Martin DL, Armelagos GJ, Goodman AH, Van Gerven DP. 1984. The effects of socioeconomic change in prehistoric Africa: Sudanese Nubia as a case study. In MN Cohn and GJ Armelagos, eds.: Paleopathology at the Origins of Agriculture. Orlando: Academic Press. p 193-214.
Massler M, Schour I, Poncher HG. 1941. Developmental pattern of the child as reflected in the calcification pattern of the teeth. Am J Dis Child 62:33–67.
McElroy A. 1990. Biocultural models in studies of human health and adaptation. Medical Anthropology Quarterly, New Series. 4: 243-265

266
McGovern-Wilson R, Quinn C. 1996. Stable isotope analysis of ten individuals from Afetna, Saipan, Northern Marianas Islands. J Arch Sci. 23:59-65.
Meiklejoh C, Wyman JM, Schentag CT. Caries and attrition: dependent or independent variables? In J Anthropol 7:17-22.
Meng Y, Zhang H, Pan F, He Z, Shao J, Ding Y. 2011. Prevalence of dental caries and tooth wear in a Neolithic population (6700-5600 years BP) from northern China. Arch Oral Biol 56:1424-1435.
Moller IJ, Pindborg JJ, Effendi I. 1977. The relation between betel chewing and dental caries. Scand J Dent Res 85:64–70.
Moore. DR. 1983. Measuring change in Marianas pottery: the sequence of pottery production at Tarague, Guam. Unpublished M.A. thesis, University of Guam, Mangilao, Guam.
Moore DR. 1994. Prehistoric ceramics. In RL Hunter-Anderson, ed.: Archaeology in Manenggon Hills, Yona, Guam, Volume III: 3.1-3.71.
Moore DR. 2002. Ceramic analysis. In J Allen et al.: Final Report, Archaeological survey and excavations at the Naval Ordnance Annex, Territory of Guam, p B.1.1-B.1-18. Prepared for Department of the Navy, Pacific Division, Naval Facilities Engineering Command, Pearl Harbor, Hawaii. Prepared by Ogden Environmental and Energy Services Co., Inc., Honolulu.
Moore. DR. 2002 . Guam’s prehistoric pottery and its chronological sequence. prepared for the US Department of the Navy, Pacific Division by Micronesian Archaeological Research Services, Guam.
Moore. DR. 2001. 2001. Results of subsurface testing and monitoring at the Kallingal property, Tumon Bay, Guam. Prepared for Mr. Tom Kallingal, Tamuning, Guam. Micronesian Archaeological Research Services, Guam.
Moore, DR, Hunter-Anderson RL. 1999 Pots and pans in the Intermediate Pre-Latte (2500-1600 BP), Mariana Islands, Micronesia. In JC Galipaud, I Lilley, eds.: The Pacific from 5000 to 2000 BP, Colonisation and Transformations. p 487-503. IRD Editions, Paris.
Moore, DR, Hunter-Anderson RL. 2001. Phase II archaeological survey: detailed recording of sites on the lands to be retained by the Navy, Nimitz Hill, Territory of Guam, Mariana Islands. Prepared by Micronesian Archaeological Research Services, Guam, for International Archaeological Research Institute, Inc., Honolulu, under contract with the Department of the Navy, Pacific Division, Naval Facilities Engineering Command, Pearl Harbor, Hawaii.

267
Moore, DR, Hunter-Anderson RL, Wells E. 1993. Results of archaeological data recovery during monitoring at the Pacific Islands Club Property, Tumon, Guam. Report prepared for Pacific Islands Club, Tamuning, Guam. Guam: Micronesian Archaeological Research Services.
Morgan WN. 1988. Prehistoric Architecture in Micronesia. University of Texas Press: Austin.
Murakami G. 2000. Charcoal taxa identification, Navy Guam land use plan parcels project, Guam. In RL Hunter-Anderson, B Dixon, T Mangieri: Cultural resources survey of five Navy surplus guam land use plan parcels, Territory of Guam, Mariana Islands, pp. C-1-C-8. Department of the Navy, Pacific Division, Naval Facilities Engineering Command, Pearl Harbor, Hawaii. International Archaeological Research Institute, Inc., Honolulu.
Nelson GC, Fitzpatrick SM. 2006. Preliminary investigations of the Chelechol ra Orrak cemetery, Republic of Palau: I, skeletal biology and paleopathology. Anthropol Sci 114:1-12.
Neves WA, Wesolowski V. 2002. Economy, nutrition, and disease in prehistoric coastal Brazil. In: Steckel RH, Rose JC, editors. The backbone of history: health and nutrition in the western hemisphere. Cambridge: Cambridge University Press. p 376-405.
Newbrun E. 1982. Sugar and dental caries: a review of human studies. Science 217:418-423.
Nunn PD. 1999. Environmental change in the Pacific Basin: chronologies, causes, consequences. London: Wiley.
Nunn PD. 2000 illuminating sea-level fall around AD 1220-1510 (730-440 Cal Yr BP) in the Pacific Islands: implications for environmental change and cultural transformation. New Zealand Geograph 56: 46-54.
Nunn PD. 2007. The AD 1300 event in the Pacific Basin. Geograph Rev 97:1-23.
Nunn PD. Britton JMR. 2001. Human-environment relationships in the Pacific Islands around AD 1300. Environ Hist 7:3-22.
Nunn PD, Hunter-Anderson RL, Carson MT, Thomas F, Ulm, S, Rowland MJ. 2007. Times of plenty, times of less: last-millennium societal disruption in the Pacific Basin. Human Ecol 35:385-401.
Ogilvie MD, Curran BK, Trinkaus, E. 1989. Incidence and patterning of dental enamel hypoplasia among the Neandertals. Am J Phys Anthrop. 79:25-41.

268
Olmo RK, Goodman WL. 1994. archaeological investigations for Ypao Beach Park ground penetrating radar survey, Guam. Prepared for Department of Parks and Recreation, Government of Guam. International Archaeological Research Institute, Inc., Honolulu.
Olmo RK. 1997. archaeological subsurface testing of Baba Commercial Center, Tumon Bay. Report prepared for MCL/Martin Christobal Laguana, Dededo, Guam. International Archaeological Research Institute, Inc., Honolulu.
Ortner DJ. 2003. Identification of pathological conditions in human skeletal remains. San Diego: Academic Press.
Osborne D. 1958. The Palau islands: an intensive survey, Bulletin 230. Honolulu: Bernice P Bishop Museum.
Oxenham M, Nguyen L, Nguyen K. 2006. The oral health consequences of the adoption and intensification of agriculture in Southeast Asia. In M Oxenham, N Tayles, eds.: Bioarchaeology of Southeast Asia. p 263-289.
Oxenham MF, Matsumura H. 2008. Oral and physiological paleohealth in cold adapted peoples: Northeast Asia, Hokkaido. Am J Phys Anthropol 135:64-74.
Parr NM, Walth CK. 2011. An Analysis of Non-Masticatory Wear in the Prehistoric Chamorro from Tumon Bay, Guam in the Marianas Islands. Am J Phys Anthropol Supp 52:235.
Parr NM. 2012. Dental Modification in a prehistoric Chamorro population from Tumon Bay, Guam. Poster presented at the 80th Annual Meeting of the American Association of Physical Anthropologists: 11-14 April 2011, Portland, OR. Am J Phys Anthropol Supp 147:233.
Pate FD, Craib JL, Heathcote GM. 2001. Stable isotopic analysis of prehistoric human diet in the Mariana Islands, western Pacific. Aust Archaeol 52:1-4.
Pawley A. 1972. On the internal relationships of Eastern Oceanic languages. Pac Anthropol Rec 13:1-142.
Pawley A. 1999. Chasing rainbows: implications of the rapid dispersal of Austronesian languages. In E Zeitoun and P Li, eds.: Selected papers from the eights International Conference on Austronesian Linguistics. Taipai: Institute of Linguistics, Academia Sinica. p 95-138.
Pawley A. 1999. Language. In M Rappaport, ed.: The Pacific Islands environment and society. Honolulu: The Bess Press. p 181-194.

269
Pawley A. 1999. Chasing rainbows: implications of the rapid dispersal of Austronesian languages. In E Zeitoun, PJK Li, eds.: Selected papers from the eighth International Conference on Austronesian Linguistics. Taipei: Academia Sinica. p 95-138.
Pawley A. 2002. The Austronesian dispersal: languages, technologies and people. In P Bellwood, C Renfrew, eds.: Examining the farming/language dispersal hypothesis. McDonald Institute monographs: 251–73. Cambridge: McDonald Institute for Archaeological Research.
Pawley A, Greene RC. 1984. The Proto-Oceanic Language Community. J Pac Hist 19: 123-146.
Pearsall DM, Collins SK. 2000. Report of phytolith analysis of samples from Nimitz Hill, Guam. In RL Hunter-Anderson, DR Moore, Phase I Archaeological survey and specialized studies in undeveloped portions of Nimitz Hill, Territory of Guam, Mariana Islands, p F-1-F-18. Department of the Navy, Pacific Division, Naval Facilities Engineering Command, Pearl Harbor, Hawaii. International Archaeological Research Institute, Inc., Honolulu.
Pechenkina E, Benfer RA, Zhijun W. 2002. Diet and health changes at the end of the Chinese Neolithic: The Yangshao/ Longshan transition in Shaanxi Province. Am J Phys Anthro- pol 117:15–36.
Pechenkina EA, Delgado M. 2006. Dimensions of health and social structure in the early intermediate period cemetery at Villa El Salvador, Peru. Am J Phys Anthropol 131: 218–235.
Pedersen PO. 1952. Some dental aspects of anthropology. Dental Record. 72: 170-178.
Pietrusewsky M. 1986. Report on the Human Skeletal and Dental Remains Excavated During the San Vitores Road Project, Tumon Bay, Guam, Mariana Islands. Prepared for Maeda Pacific Corporation and Maeda Road Construction Company, Ltd. A Joint Venture. Unpublished manuscript.
Pietrusewsky M. 1990a. Craniofacial variation in Australasian and Pacific populations. American Journal of Physical Anthropology 82:319–40.
Pietrusewsky M. 1990b. Craniofacial variation in Micronesia and the Pacific: a multivariate study. Micronesia Suppl 2:373–402.
Pietrusewsky M. 1994. Pacific-Asian relationships: a physical anthropological perspective. Ocean Linguist 33:407–30.
Pietrusewsky M. 1995. Taiwan Aboriginals, Asians, and Pacific Islanders: a multivariate investigation of skulls. In PJ Li, C Tsang, Y Huang, D Ho, C Tseng, eds.: Austronesian studies relating to Taiwan, Taipei: symposium series of the Institute of History and Philology.

270
Pietrusewsky M. 1996. Multivariate craniometric investigations of Japanese, Asians, and Pacific Islanders. In K Omoto, ed.: interdisciplinary perspectives on the origins of the Japanese: international symposium No. 11-B, Kyoto: International Research Center for Japanese Studies.
Pietrusewsky M. 2000. Metric analysis of skeletal remains: Methods and applications. In Katzenberg MA, Saunders SR, editors. Biological anthropology of the human skeleton. New York: Wiley-Liss. p 375-415.
Pietrusewsky M. 2005. The physical anthropology of the Pacific, East Asia, and Southeast Asia. In Blench R, Sagart L, Sanchez-Mazas A, editors. The peopling of East Asia. New York: Routledge Curzon. p 201-229.
Pietrusewsky M. 2006. The initial settlement of remote Oceania: The evidence from physical anthropology. In T Simanjuntak, IHE Pojoh, M Hisyam, eds.: Austronesian diaspora and the ethnogenesis of people in Indonesian archipelago. Proceedings of the International Symposium. Jakarta: Indonesian Institute of Sciences, LIPI Press. p 320-347.
Pietrusewsky M. 2008. Craniometric variation in Southeast Asia and neighboring regions: A multivariate analysis of cranial measurements. Human Evol 23:49-86.
Pietrusewsky M and Batista C. 1980. Human Skeletal and Dental Remains from Four Sites on Tinian and Saipan, Commonwealth of the Northern Mariana Islands. Final report to the Northern Marianas Archaeological Society, Inc. Unpublished manuscript.
Pietrusewsky M, Douglas MT. 2002. Intensification of agriculture at Ban Chiang: is there evidence from the skeletons? Asian Persp 40:157-178.
Pietrusewsky M, Tsang C. A preliminary assessment of health and disease in human skeletal remains from Shi San Hang: a prehistoric aboriginal site on Taiwan. Anthorpol Sci 111:203-223.
Pietrusewsky M, Douglas MT, and Ikehara RM. 1992. Prehistoric Chamorro Remains from the Apurguan Site, Tamuning District, Guam: An Osteological Investigation and Comparisons with Other Micronesian Series. Honolulu: International Archaeological Research Institute, Inc. Honolulu, Hawaii Unpublished manuscript.
Pindborg JJ. 1970. Pathology of the dental hard tissues. Philidelphia: W.B. Saunders Company.
Powell ML. 1985. Dental Wear and Caries in Dietary Reconstruction. In RI Gilbert, JH Mielke, eds.: The Analysis of Prehistoric Diets. Academic Press, Inc.: New York.
Pregill GK Steadman DW. 2009. The prehistory and biogeography of terrestrial
vertebrates on Guam, Mariana Islands. Divers Distrib 15:983–996.

271
Prosser CL. 1964. Perspectives of adaptation: Theoretical aspects. In DB Dill, EF Adolph, eds.: Adaptation to the environment: section 4, handbook of physiology. Washington: American Physiology Society p 11-25.
Rainbird P. 2003. The archaeology of Micronesia. Cambridge: Cambridge University Press.
Ray ER. 1981. the material culture of prehistoric Tarague Beach, Guam. Unpublished M.S. thesis, Arizona State University.
Reid LA. Sources of Proto-Oceanic initial prenasalization: The view from outside Oceanic. In VP DeGuzman, Bender B, eds.: Grammatical analysis: Morphology, syntax and semantics: Studies in honor of Stanley Starosta.. Ocean Linguist Special Publication No. 29. Honolulu: University of Hawai‘i Press. p 30-45
Reid LA. 2002. Morphosyntactic evidence for the position of Chamorro in the Austronesian language family. In RS Bauer, ed.: Collected papers on Southeast Asian and Pacific languages, Pacific Linguistics 530. Canberra: Department of Linguistics, Research School of Pacific and Asian Studies, The Australian National University. p 63–94.
Reinman FR. 1977. An archaeological survey and preliminary test excavations on the Island of Guam, Mariana Islands, 1965-1966. Micronesian Area Research Center, University of Guam, Mangilao, Guam.
Roberts C, Manchester K. 2005. The Archaeology of Disease, Third Ed. Sutton Publishing: Gloucestershire.
Rodríguez N, Mayer N, Gauna HF. 2007. Effects of prenatal stress on male offspring sexual maturity. Biocell 311:67–74.
Rose JC, Condon KW, Goodman AH. 1985. Diet and dentition: developmental disturbances. In: Gilbert RI, Mielke JH, editors. The analysis of prehistoric diets. Orlando, FL: Academic Press. p 281–305.
Ross M, Pawley A, Osmond M. 1998. The lexicon of proto-Oceanic: the culture and environment of ancestral Oceanic society. Volume 1: Material culture, Pacific Linguistics series C 152. Canberra: Pacific Linguistics, Research School of Pacific and Asian Studies, The Australian National University.
Rowe N. 1982. Dental caries. In PF Steele, ed.: Dimensions of dental hygiene, 3rd ed. Philadelphia: Lea and Febiger. p 209-237.
Roy KJ. 1989. A Study of Prehistoric Marianas Islanders: The Skeletons Under Their Skins. M.A. Thesis. University of Otago, Dunedin.

272
Rudyk O, Makra P, Jansen E, Shattock MJ, Poston L, Taylor PD. 2011. Increased cardiovascular reactivity to acute stress and salt-loading in male offspring of fat fed non-obese rats. PLoS One 6: e25250.
Russell S. Tiempon I Manmopo’na: Ancient Chamorro Culture and History of the Northern Mariana Islands. 1998. Micronesian Archaeological Survey Report, No. 32. Division of Historic Preservation.
Ryan EM. 2010. Chamorro health and illness at a prehistoric coastal habitation site in Tumon Bay, Guam. PhD Thesis. Australian National University, Canberra.
Safford WM. 1905. The Useful Plants of Guam. Government Printing Office, Washington, D.C.
Safford WM. 1909. Chamorro language of Guam: a grammar of the idiom spoken by the inhabitants of the Marianne, or Ladrones, Islands. Washington DC: WH Lowdermilk and Co.
Sava VJ. 2001. Bioarchaeological Study of Prehistoric Human Skeletal Remains. Archaeological Mitigation Program Phase I Tumon Bay Infrastructure and Beautification Project. Report prepared for Guam Department of Public Works. PHRI Inc. Unpublished manuscript.
Sciulli PW, Doyle WJ, Kelly S, Siegel P, Siegel MI. 1979. The interaction of stressors in the induction of increased levels of fluctuating asymmetry in the laboratory rat. American Journal of Physical Anthropology. 50: 279-284.
Scott GR, Turner CG. 1997. The anthropology of modern human teeth: dental morphology and its variation in recent human populations. Cambridge: Cambridge University Press.
Selye H. 1936. A syndrome produced by noxious agents. Nature 138:32.
Selye H. 1956.The Stress of Life. New York: Mc- Graw-Hill.
Selye H. 1973. The evolution of the stress concept. Am Sci 61:692-699.
Seow WK, Amaratunge A, Bennett R, Bronsch D, Lai PY. 1996. Dental health of Aboriginal pre-school children in Brisbane, Austalia. Community Dent Oral Epidemiol 24: 187–190.
Serjeantson SW. 1985. Migration and admixture in the Pacific: insights provided by human leukocyte antigens. In RL Kirk, E Szathmary, eds.: Out of Asia: peopling of the Americas and the Pacific. Canberra: Journal of Pacific History, Australian National University. p 133-146.
Serjeantson SW. 1989. HLA antigens. In AVS Hill, SW Serjeantson, eds.: The colonization of the Pacific: A genetic trail. Oxford: Clarendon Press. p 286-294.

273
Shapiro HL, Buck PH. 1936. The physical characters of Cook Islands. Memoirs of the BP Bishop Museum, 12. No. 1.
Siegel MI, Doyle W. 1975a. The differential effects of prenatal and postnatal audiogenic stress on fluctuating dental asymmetry. Journal of Experimental Zoology. 191: 211-214.
Siegel MI, Doyle W. 1975b. The effects of cold stress on fluctuating asymmetry in the dentition of the mouse. Journal of Experimental Zoology. 193: 385-389.
Siegel MI, Doyle W, Kelly C. 1977. Heat stress, fluctuating asymmetry and prenatal selection in the laboratory rat. American Journal of Physical Anthropology. 46: 121-126.
Siegel MI, Smookler HH. 1973. Fluctuating dental asymmetry and audiogenic stress. Growth. 37: 35-39.
Skinner M. 1996. Developmental stress in immature hominines from Late Pleistocene Eurasia: evidence from enamel hypoplasia. J Archaeol Sci 23:833–852.
Skinner MF, Hopwood D. 2004. Hypothesis of the causes and periodicity of repetitive linear enamel hypoplasia in large, wild African (Pan troglodytes and Gorilla gorilla) and Asian (Pongo pygmaeus) apes. Am J Phys Anthropol 123:216–235.
Šlaus M. 2000. Biocultural analysis of sex differences in mortality profiles and stress levels in the Late Medieval population from Nova Rača. Croatia. Am J Phys Anthropol 111:193–209.
Slonim NB. 1974. Introduction. In NB Slonim, ed.: Environmental physiology. St. Louis: C.V. Mosby. p 1-9.
Smith PO, Bar-Yosef, Sillen A. 1984. Archaeological and skeletal evidence for dietary change during the LatePleistocene/Early Holocene in the Levant. In MN Cohn and GJ Armelagos, eds.: Paleopathology at the Origins of Agriculture. Orlando: Academic Press. p 101-136.
Spoehr A. 1954. Saipan, the ethnology of a war-devastated island. Fieldiana: Anthropology 41:1-383. Chicago Natural History Museum.
Spoehr A. 1957. Marianas Prehistory, Archaeological Survey and Excavations on Saipan, Tinian and Rota. Fieldiana: Anthropology 48. Chicago Natural History Museum.
Spriggs M. 1999. Archaeological dates and linguistic subgroups in the settlement of the Island Southeast Asian-Pacific Region. Bull Indo Pac Pre Hist 8:17-24.
Steadman DW. 1999. The prehistory of vertebrates, especially birds, on Tinian, Aguiguan, and Rota, Northern Mariana Islands. Micronesica, 31:319–345.

274
Steckel RH, Rose JC. 2002. The backbone of history: health and nutrition in the western hemisphere. Cambridge: Cambridge University Press.
Stefan VH, Chapman PM. 2003. Cranial variation in the Marquesas Islands. . Am J Phys Anthropol 121: 319–331.
Stodder ALW. 1993. Human Skeletal Remains from the Mangilao Golf Course Project, Mangilao Municipality, Territory of Guam. Paul H. Rosendahl, Ph.D., Inc. Hilo, HI. PHRI Project No. 89-744.
Stodder ALW. 1997. Subadult stress, morbidity, and longevity in Latte Period Populations on Guam, Mariana Islands. Am J Phys Anthropol 104:363–380.
Stodder ALW, Martin DL, Goodman AH, Reff DT. 2002. Cultural longevity in the face of biological stress. In: Steckel RH, Rose JC, editors. The backbone of history: health and nutrition in the Western hemisphere. Cambridge: Cambridge University Press. p 481–505.
Stojanowski CM. 2003. Differential phenotypic variability among the Apalachee populations of La Florida: A diachronic perspective. Am J Phys Anthropol 120:352–363.
Stojanowski CM. 2005. The bioarchaeology of identity in Spanish colonial Florida: Social and evolutionary transformation before, during, and after demographic collapse. Am Anthropol 107:417–431.
Stojanowski CM, Knudson KJ. 2011. Biogeochemical inferences of mobility of Early Holocene fisher-foragers from the southern Sahara desert. Am J Phys Anthrop 146:49-61.
Stoppelaar JD, van Houte J, Dirks OB. 1970. The effect of carbohydrate restriction on the presence of Streptococcus mutans, Streptococcus sanguis and iodophilic polysaccharide-producing bacteria in human dental plaque. Caries Res 4:114-123.
Strickland SS. 2002. Anthropological perspectives on use of the areca nut. Addic Biol. 7:85-97.
Sutton DG. 1980. A culture history of the Chatham Islands. J Polynesian Soc. 89:67-93.
Sutton PRN. 1978. Unusual forms of abrasion of teeth: Unintentional ‘filing’ in Micronesians and Polynesians. Archaeology and Physical Anthropology in Oceania 8: 30–39.
Taboroši D, Jenson JW, Myroie JE. 2005. Karst features of Guam, Mariana Islands. Micronesica 38: 17-46.
Tayles N, Domett K, Nelson K. 2000. Agriculture and dental caries? The case of rice in prehistoric Southeast Asia. World Archaeol 32:68-83.

275
Temple DH. 2007. Dietary variation and stress among prehistoric Jomon foragers from Japan. Am J Phys Anthropl 133:1035-1046.
Temple DH, Larsen CS. 2007. Dental caries prevalence as evidence for agriculture and subsistence variation during the Yayoi Period in prehistoric Japan: biocultural interpretations of an economy in transition. Am J Phys Anthropl 134:501-512.
Temple DH. 2010. Patterns of systemic stress during the agricultural transition in prehistoric Japan. Am J Phys Anthropol 142:112-124.
Terrell J. 1986. Prehistory in the Pacific Islands: a study of variation in language, customs, and human biology. Cambridge: Cambridge University Press.
Terrell J, Hunt TL, Gosden C. 1997. The dimensions of social life in the Pacific: human diversity and the myth of the primitive isolate. Curr Anthropol 38:155-196.
Thomas N. 1989. The force of ethnology: Origins and significance of the Melanesia/Polynesia division. Curr Anthropol 30: 27-41.
Thompson LM. 1932. Archaeology of the Marianas Islands. Honolulu: Bernice P. Bishop Museum Bulletin 100.
Thompson LM. 1940. The function of Latte in the Marianas. J Polynesian Soc 49:447-465.
Thompson LM. 1945. The Native Culture of the Marianas Islands. Honolulu: Bernice P. Bishop Museum Bulletin, No. 85.
Tomonari-Tuggle MJ, Tuggle HD, Welch DJ. 2005. Final Report: Regional integrated cultural resources management plan for COMNAVREG Marianas lands. Volume I: Guam. Prepared for Commander, Navy Region, Marianas by International Archaeological Research Institute, Inc., Honolulu.
Trembly D. 1999. Human skeletal and dental remains from the Tumon Bay Hyatt Hotel site. Prepared for EIE Guam Corporation. Paul H. Rosendahl, Ph.D., Inc., Hilo, Hawaii.
Topping D, Ogo P, Dungca B.. 1975 Chamorro-English Dictionary. Honolulu : University of Hawai`i Press.
Turner CG. 1972. Comment. Current Anthropology. 13:520-521.
Turner CG. 1979. Dental anthropological indication of agriculture among the Jomon people of central Japan. Am J Phys Anthrop 51:610-636.
Turner CG. 1987. Late Pleistocene and Holocene population history of East Asia based on dental variation. Am J Phys Anthrop 73:305-321.

276
Turner CG. 1990a. Major features of sundadonty and sinodonty, including suggestions about East Asia microevolution, population history, and Late Pleistocene relationships with Australian Aboriginals. Am J Phys Anthrop 82:295-317.
Turner CG. 1990b. Origin and affinity of the people of Guam: a dental anthropological assessment. Micronesica Supplement. 2:403-416.
Ubelaker DH. 1980. Human skeletal remains from site OGSE- SO, a preceramic site on the Santa Elena Peninsula, coastal Ecuador. J Wash Acad Sci 70:3–24.
Ubelaker DH. 1984. Prehistoric human biology of Ecuador: possible temporal trends and cultural considerations. In MN Cohn and GJ Armelagos, eds.: Paleopathology at the Origins of Agriculture. Orlando: Academic Press. p 495-513.
Ubelaker DH, Newson LA. 2002. Patterns of health and nutrition in prehistoric Ecuador. In: Steckel RH, Rose JC, editors. The backbone of history: health and nutrition in the western hemisphere. Cambridge: Cambridge University Press. p 343-375.
Underwood J. 1973. Population history of Guam: context of microevolution. Micronesica 9:11-44.
Vodanovic M, Brkic H, Slaus M, Demo Z. 2005. The frequency and distribution of caries in the mediaeval population of Bijelo Brdo in Croatia (10th–11th century). Arch Oral Biol 50:669–80.
Walker PL, Erlandson JM. 1986. Dental evidence for prehistoric dietary change on the northern Channel Islands, California. Am Antiq 51:375-383.
Walker PL, Hewlett BS. 1990. Dental health, diet, and social status among central Afrian foragers and farmers. Am Anthropol 92:383-389.
Ward JV. 1994. A Holocene pollen record from the Pago River Valley, Guam. In RL Hunter-Anderson Archaeology, ed.: Manenggon Hills, Yona, Guam, vol. III, Results of Analyses. Report prepared for MDI Guam Corporation, Yona, Guam. Guam: Micronesian Archaeological Research Services. Athens & Ward: Holocene Vegetation, Guam 2930 p 9.34–9.52.
Ward JV. 1995. A Holocene pollen record from Laguas River, Guam. In EF Wells, UK Prasad, RL Hunter-Anderson, eds.: Archaeology in the Marine Drive corridor from Agana to Agat, Guam, Appendix B. Draft final report prepared for Guam Department of Public Works, Agana, Guam. Guam: Micronesian Archaeological Research Services. p 205–224.

277
Ward JV. 2000. Holocene pollen record from Nimitz Hill, Guam, Mariana Islands. In RL Hunter-Anderson, DR Moore, eds.: Phase I archaeological survey and specialized studies in undeveloped portions of Nimitz Hill, Territory of Guam, Mariana Islands, Department of the Navy, Pacific Division, Naval Facilities Engineering Command, Pearl Harbor, Hawaii. International Archaeological Research Institute, Inc., Honolulu. p C-1-C-11.
Watt ME, Lunt DA, Gilmour WH. 1997. Caries prevalence in the permanent dentition of a mediaeval population from the south-west of Scotland. Arch Oral Biol 42:601–20.
Webb S. 1984. Intensification, population and social change in Southeastern Australia: the skeletal evidence. Aboriginal Hist 8:154-172.
Wellin E. 1978. Theoretical orientations in medical anthropology: change and continuity over the past half-century. In MH Logan, EE Hunt, eds.: Health and the Human Condition. Belmont: Wadsworth Publishing. P 23-39.
Wickler S. 2004. Modelling colonization and migration in Micronesia from a zooarchaeological perspective. In M Mondini, S Munoz, S Wickler, eds.: Proceedings of the 9th Conference of the International Council of Archaeozoology, Durham, August 2002. Oxford: Oxbow Books. p 28–40.
Wickler SK. 1993. Analysis of prehistoric pottery. In James Carucci, ed.: The archaeology of Orote Peninsula: phase i and ii archaeological inventory survey of areas proposed for projects to accommodate relocation of Navy activities from the Philippines to Guam, Mariana Islands. pp. 131-151. Prepared for Belt, Collins & Associates, Honolulu. International Archaeological Research Institute, Inc., Honolulu.
Wiles GJ, Amerson AB, Beck RE. 1989. Notes on the herpetofauna of Tinian, Mariana Islands. Micronesica 22:107–118.
Wright LE, Schwarcz HP. 1999. Correspondence between stable carbon, oxygen and nitrogen istotopes from human tooth enamel and dentine: infant diets at Kaminaljuyu. J Archaeol Sci 26:1159-1170.
Yamamoto M. 1988. Enamel hypoplasia of the permanent teeth in Japanese from the Jomon to the Modern Period. J Anthropol Soc Nippon 96:417-433.
Yawata I. 1961. Burial Systems of Ancient Mariana Islanders. Asian Persp 5–6:164–165.
Zhou L, Corruccini RS. 1998. Enamel hypoplasia related to famine stress in living Chinese. Am J Hum Biol 10:723-733.
Zobel E. 2002. The position of Chamorro and Palauan in the Austronesian family tree: evidence from verb morphology and morphosyntax. In F Wouk, M Ross, eds.: The history and typology of western Austronesian voice systems. Canberra: Pacific Linguistics. p 405-434.

278
Zumbroich TJ. 2007. The origin and diffusion of betel chewing: a synthesis of evidence from South Asia, Southeast Asia and beyond. J Indian Med 1:87-140.

279
BIOGRAPHICAL SKETCH
Nicolette Maria Luney Parr graduated with highest honors from the University of
Florida in 2002 with a bachelor’s degree in anthropology and minor in religion. Ms.
Parr’s graduate studies began at the University of Indianapolis, were she completed a
Master of Science degree in human biology. While at the University of Indianapolis, Ms.
Parr served as the Laboratory Coordinator for the Archaeology and Forensics
Laboratory, performed recovery of human remains from crime scenes, and assisted with
forensic casework, and served as a teaching assistant for anatomy, skeletal biology,
and dental anthropology courses. After an internship at the University of Pretoria in
South Africa, Ms. Parr completed her master’s thesis on determining ancestry from the
mandible.
Ms. Parr entered the anthropology Ph.D. program at the University of Florida in
2005, where she served as a graduate analyst at the C.A. Pound Human Identification
Laboratory, was a teaching assistant for an introductory biological anthropology course,
and served as in instructor for an introductory course on forensic anthropology, and a
laboratory based course in human osteology. She co-authored a book entitled Bare
Bones: An Introduction to Forensic Anthropology, which is currently in its second
edition. After completing coursework, Ms. Parr worked part-time as the physical
anthropologist for Garcia and Associates in Guam. Her work as an osteologist in Guam
led to the culmination of this project. Ms. Parr received her Ph.D. from the University of
Florida in August 2012. Upon graduation, Ms. Parr was hired by the Joint Pow/MIA
Accounting Command-Central Identification Laboratory in Honolulu, Hawaii, where she
works as a forensic anthropologist.