Embed Size (px)
Citation preview

Cellular Immunology 237 (2005) 131–140
www.elsevier.com/locate/ycimm
A cytokine promoter/yellow Xuorescent protein reporter transgene serves as an early activation marker of lymphocyte subsets
Shantha Kumar a,1, Marianne J. Skeen a,1, YaVa Adiri a, Hyseuk Yoon a, Vaiva D. Vezys b, Aron E. Lukacher b, Brian D. Evavold a, H. Kirk Ziegler a, Jeremy M. Boss a,¤
a Department of Microbiology and Immunology, Emory University School of Medicine, 1510 Clifton Rd., Atlanta, GA 30322, USAb Department of Pathology and Laboratory Medicine, Emory University School of Medicine, 1510 Clifton Rd., Atlanta, GA 30322, USA
Received 12 July 2005; accepted 23 November 2005
Abstract
A mouse containing an IL-4 promoter linked to the yellow Xuorescent protein (YFP) reporter transgene was created to follow aspectsof lymphocyte development and function. Following stimulation with phorbol 12-myristate 13-acetate and ionomycin, anti-CD3/CD28,antigen-speciWc peptide, or allogeneic cells, both CD4 and CD8 T cells expressed the transgene within 24 h in a manner that was consis-tent with cellular activation markers. Transgene induction was inhibited by cyclosporine and FK506, suggesting that its activation occursin an NFAT-dependent manner. B lymphocytes were also able to express the transgene when stimulated with LPS. This induction wasinhibited in part by rapamycin. The results suggest that this transgene can function as an indicator of lymphocyte activation. BecauseYFP is not toxic and requires no preparation of the cells to view the reporter gene, this system provides a unique tool to follow lympho-cyte activation in a number of model systems, such as those involving transplantation, allergy, and vaccine development. 2005 Elsevier Inc. All rights reserved.
Keywords: T cell activation; Transgenic; CD4/CD8 cells
1. Introduction
During acquired immune responses both B and T cellpopulations are activated to proliferate. Antigen speciWccellular proliferation is the basis of clonal expansion andthe acquired immune response. Recent evidence has sug-gested that the initial exposure to antigen is suYcient tocontinue subsequent cell division in the absence of contin-ued stimulation [1–3]. Concomitant with B and T cell acti-vation is the acquisition of an eVector phenotype and achange in the gene expression proWles [4]. Some of thesechanges manifest in the expression of novel surface mark-ers, such as CD69 for T cells [5] and Syndecan-1 for B cells[6,7]. While changes in the appearance/disappearance ofsurface markers are easily monitored if an antibody is
* Corresponding author. Fax: +1 404 727 1719.E-mail address: [email protected] (J.M. Boss).
1 These authors contributed equally to this study.
0008-8749/$ - see front matter 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.cellimm.2005.11.003
available, other changes, such as those that include theinduction of cytokine synthesis, are less easily monitored.This is particularly true of T cells that have diVerentiatedfrom the Th0 state to Th1 or Th2, where cytokine synthesisof IFN-� and IL-4, respectively, distinguish between thesesubsets of cells [8].
The Th1 or Th2 bias of naive T cells occurs in a step-wisemanner with cytokine-dependent and independent phases.These phases include the activation of cell-type speciWc fac-tors, epigenetic chromatin modiWcation, and maintenance ofa stable phenotype. For example, in Th2 cells, STAT6 activa-tion by IL-4, together with GATA-3 and c-maf promote Th2diVerentiation [9–11]. This diVerentiation is facilitated by spe-ciWc remodeling and increased chromatin accessibility of theTh2-speciWc gene loci and enhanced secondary immuneresponses [9,12,13]. A common feature of the IFN-�, IL-2,and IL-4 gene promoters is the presence of cis-acting regula-tory elements that bind the nuclear factor of activated T cells(NFAT) and composite sites, which bind NFAT-AP1

132 S. Kumar et al. / Cellular Immunology 237 (2005) 131–140
complexes [14–18]. Phosphorylated NFAT is maintained inthe cytoplasm [18,19]. Upon TCR stimulation, NFAT isdephosphorylated by calcineurin, allowing its translocationto the nucleus. The competitive inhibition of calcineurin’sdephosphorylation of NFAT by the immunosuppressivedrugs cyclosporin A (CsA) and FK506 (reviewed in [17]),results in inhibition of IL-4, IL-2, IFN-�, and other NFAT-regulated genes [20,34].
To study the activation and subsequent fate of T cellsin vivo, a transgenic mouse strain was created that harborsin its genome a yellow Xuorescent protein (YFP) reportergene under the control of the immediate upstream IL-4promoter and regulatory sequences known to respond tocell activation signals [21–23]. The analysis of a transgenicfounder line from this vector, Line 19, was found to providea marker of T and B cell activation. T cells stimulated byphorbol 12-myristate 13-acetate (PMA) and ionomycin(Ion) showed robust expression of the transgene. Addition-ally, YFP expression was observed in T cells in response toconcanavalin A (ConA), as well as immobilized anti-CD3/CD28 antibodies. B cells activated by lipopolysaccharide(LPS) also expressed high levels of the transgene reporter.Moreover, induction of the transgene was detected follow-ing appropriate speciWc peptide stimulation in DO11.10 Tcell receptor transgenic mice crossed onto the Line 19 back-ground. The transgene was also expressed when spleen cellsfrom Line 19 mice were exposed to allogeneic APCs in anMLR. In an in vivo model for YFP transgene expression,MHC tetramer-positive T cells from mice infected withpolyoma virus were found to express the transgene. T cellinduction of the transgene was inhibited by cyclosporine A(CSA) and FK506, implicating a role for NFAT in thepathway leading to expression of the transgene. The Line19 “activation” mouse therefore provides a simple geneticbackground that can easily allow the identiWcation andsorting of activated lymphocyte populations duringimmune responses. Thus, this transgenic model would be ofpotential use in studies of NFAT-dependent activation, andas a marker of helper T cell activation.
2. Materials and methods
2.1. Transgene construction and generation of transgenic mice
The transgene construct pIL-4YFP consisted of themurine IL-4 promoter (¡742 to +63) ligated to the
enhanced YFP coding fragment and polyadenylation site(between the HindIII and BamHI sites) of the pEYFP-1vector purchased from Clontech (Fig. 1). The HindIII toAXII fragment was digested from the vector and used toinject mouse F2 embryos generated from a (C57Bl/6 £ SJL)cross. Creation of the transgenic mouse line was carried outby the Emory Transgenic Facility. Following identiWcationof founder animals by PCR and Southern blotting, thefounders were backcrossed to C57Bl/6 animals. Primersused for PCR analysis were 5�-ACCCTCGGACACCTGTGACC from the murine IL-4 promoter and 5�-GGTCAGCTTGCCGTAGGTGG from the EYFP vector. At thecurrent time, 10 generations of backcrossed animals havebeen performed. No diVerence in reactivity has beenobserved through the successive generations.
2.2. Cell preparation and culture
Single cell suspensions were prepared from spleens byhomogenization. Red blood cells were lysed with ammo-nium chloride buVer. Cells were washed and resuspendedfor culture in RPMI 1640 supplemented with 10% fetalbovine serum (FBS), 5£ 10¡5 M 2-� mercaptoethanol,0.5 mM sodium pyruvate, 10 mM Hepes buVer, 50 U/mlpenicillin, 50 �g/ml streptomycin, and 2 mM L-glutamine.For in vitro stimulation, cells were cultured with combina-tions of the following: PMA (10 ng/ml), ionomycin (1 �M),concanavilin A (2 �g/ml), LPS (2 �g/ml), immobilized anti-CD3 (10 �g/ml), or soluble anti-CD28 (2�g/ml). Immobi-lized anti-CD3 was coated in high pH buVer overnight at4 °C and plates were washed well before use. In some exper-iments, B cells were puriWed from splenic lymphocytes bynegative selection with CD43 (Ly-48) microbeads accord-ing to the manufacturer’s instructions (Miltenyi Biotec).This resulted in >95% B cell population as determined byXow cytometry for B220 and I-A.
2.3. Analysis of lymphocyte populations by Xow cytometry
Spleen cells (1£ 106/sample) were incubated for 30 min at4 °C with Xuorochrome-conjugated antibodies to surfacemarkers that characterize lymphocyte subsets. Cells werewashed twice with wash buVer (PBS with 3% FBS and 0.1%sodium azide) and examined by Xow cytometry. Data froma minimum of 10,000 cells per population were collectedusing a FACScalibur Xow cytometer (Becton–Dickinson)and analyzed using CellQuest software. The following
Fig. 1. Schematic of IL-4YFP transgene. The IL-4 promoter region from ¡742 to +63 was fused in frame to the EYFP reporter. The location of the probe
Probe
PCR Primers -111 +155
A
-742 +63pIL-4-YFP EYFP
+1070+831
IL-4 promoter
H
Poly A
N
used in Southern analysis and primers used for PCR analysis of the mice are shown.

S. Kumar et al. / Cellular Immunology 237 (2005) 131–140 133
reagents for Xow cytometry were purchased from BDPharmingen: GK1.5-FITC (anti CD4), 53-6.72-PerCP or -FITC (anti CD8), 23G2-FITC (CD45R/B220), MEL-14-PE(CD69), and TP1.55.3-APC (CD62L). Isotype controls forthese antibodies were as follows: rat IgG-FITC, rat IgG-PE, hamster IgG-PE, rat IgG-PerCP, and rat IgG-APCfrom BD Pharmingen. Antibody combinations used in eachexperiment are indicated in the Wgure legends.
2.4. Analysis of signal transduction inhibition
A representative group of signal transduction inhibitors(purchased from Sigma, MO and Calbiochem, CA) aimedat a variety of cellular targets (see Table 1) was added tospleen cell cultures 30–60 min prior to the addition of T orB cell stimuli. Cells were examined by Xow cytometry 24 hlater for expression of YFP.
2.5. Mixed lymphocyte reaction
Stimulator populations were enriched for dendriticcells (CD11c+) by incubating spleen cells on tissue culturedishes for 2 h at 37 °C to enrich for adherent cells. Thesecells were then incubated overnight at 37 °C to allow thedendritic population to loosen from the dish. Cells wereharvested by vigorous washing and were used as stimula-tors in MLR cultures. Responder populations wereenriched for CD4+ and CD8+ cells using mouse T cellenrichment columns (R&D Systems, Minneapolis, MN).These cells were also characterized by Xow cytometry.Responder and stimulator cells were incubated togetherat a ratio of 10:1 in 24-well culture plates. Cells were har-vested after 24, 48, or 72 h of culture and examined forYFP expression. YFP expression patterns at these timepoints were similar.
2.6. Antigen speciWc peptide responses
DO11.10 mice [24] were crossed with Line 19 homozy-gous mice and the F1 were used for peptide speciWcresponses. Spleen cells were incubated overnight with theindicated amount of OVA 323–339. Cells were stained withanti CD69 and CD4 and the CD4 population was analyzedby Xow cytometry. DO11.10 T cells were driven to Th1 orTh2 phenotype using appropriate cytokines and antibodies.Spleen cells (3£ 106) from DO11.10 mice were stimulatedwith 1�M OVA 323–339 peptide and either IL-12 (2 ng/ml)and anti-IL-4 antibody (10�g/ml) to produce Th1 cells orIL-4 (10 ng/ml) and anti-IFN-� antibody (10�g/ml) to gen-erate Th2 cells. After seven days, the T cells were puriWedand restimulated with the indicated doses of OVA peptidesand analyzed for IL-4YFP levels by Xow cytometry.
2.7. Polyoma infection and tetramer staining
Line 19 homozygous mice were infected with 2£ 106 pfuof polyoma strain A2 subcutaneously. Spleens were har-vested at Wve days post infection and the polyoma speciWcCD8 T cells were identiWed by tetramer staining using Db-LT359 as previously described in Byers et al. [25] Thisexperiment was repeated two times with identical results.
3. Results
3.1. YFP is expressed in lymphocytes following stimulation in vitro
In an eVort to create reporter mice that would allow spe-ciWc markers of activation and diVerentiation to be deter-mined without sacriWce to the animal or to the cells, themurine IL-4 promoter from ¡742 to +63 was fused in
Table 1Inhibition of YFP expression
10–20% inhibition (+), 20–50% inhibition (++), or >50% inhibition of the response.Inhibition patterns were similar in CD8+ cells (data not shown).For comparison, inhibitors were also used at twice the concentration shown above. Protein and calmodulin inhibitors were used at four concentrationseach because diVerent kinases exhibit varying sensitivities to the same inhibitor. Results are presented for a single concentration for each inhibitor for sim-plicity. Use of other concentrations did not result in more inhibition.
Target of inhibition Inhibitor Concentration CD4+ cells B220+ cells
PMA+Ion CD3/CD28 PMA+Ion LPS
Immunophilins Cyclosporin A 1 �M +++ +++ +++ ¡FK506 10 nM +++ +++ ++ +
Rapamycin 10 nM ¡ + ++ +++
Protein and calmodulin kinases Staurosporin 20 nM ¡ + ¡ ¡H7 6 �M ¡ ¡ ¡ ¡K252a 25 nM ¡ ¡ ¡ ¡
Phosphatidylinositol kinase Wortmanin 5 nM ¡ ¡ ¡ ¡Tyrosine kinase Genistein 2.6 �M ¡ ¡ ¡ ¡MAP kinase and MEK SB 202190 350 nM ¡ ¡ ¡ +
5�-Iodotubericidin 530 nM + ¡ ¡ ¡U0126 75 nM ¡ ¡ ¡ ¡
I �B and NF-�B PDTC 50 �M ¡ ¡ ¡ ¡

134 S. Kumar et al. / Cellular Immunology 237 (2005) 131–140
frame to the enhanced, yellow Xuorescent protein (YFP)creating the pIL-4YFP construct (Fig. 1). Transient trans-fection of the pIL-4YFP reporter construct into EL-4 Tcells showed that the reporter could respond to general cel-lular stimulation by PMA/Ion (data not shown). The pIL-4YFP vector was then used to create a transgenic mouseline. Four founder mice carrying the IL-4YFP transgenewere identiWed by Southern blotting and PCR. Foundermice were backcrossed with C57Bl/6 mice and the N2 gen-eration was examined for copy number and tested for theirability to express the YFP reporter transgene followingin vitro stimulation. The copy number of the diVerent foun-der mice varied. For the “Line 19” mouse, approximately50 copies of the transgene were integrated (data notshown).
Splenic lymphocytes isolated from transgenic and non-transgenic mice were stimulated with PMA/Ion and ana-lyzed for the expression of YFP by Xow cytometry. Of thefour founder lines, only founder Line 19 lymphocytes dis-played a detectable response to the stimulation (Fig. 2Aand data not shown). To further characterize responsesgenerated from Line 19 lymphocytes, additional stimulantswere tested, including Con A, immobilized anti-CD3 andCD28 antibodies, and LPS. The lymphocytes were stainedwith antibodies to CD4, CD8, and B220, and each of thesecell populations was characterized for forward scatter andexpression of the YFP transgene (Fig. 2A). For compari-son, a non-transgenic littermate was examined for PMA/Ion and LPS induced auto-Xuorescence (Fig. 2B). With theexception of the LPS treatment, CD4+ and CD8+ T cellswere activated by each of the signals as evidenced byincreased forward scatter, a measurement indicatingincreased cell size. Moreover, the activated cell populationsdisplayed a robust YFP signal. While showing clear activa-tion in response to the appropriate signals (forward scatterFig. 2B), activated lymphocytes from the non-transgenic lit-termates displayed minimal yellow Xuorescence, indicatingthat the signal observed in the transgenic lymphocytes wasdue to the induction of the YFP transgene.
Representing the splenic B cell population, B220+ cellswere analyzed for activation by the above stimulants andexpression of the YFP reporter (Fig. 2A). As anticipatedthe B cells were highly activated by PMA/Ion, as well as byLPS. PuriWed B cells stimulated with LPS alone (Fig. 3C),LPS, IL-2, and IL-5, or LPS and anti-IgM also expressedthe transgene (data not shown). The activated cell popula-tions also showed a strong YFP signal. Unexpectedly, theimmobilized and anti-CD3/CD28 stimulation also resultedin activation of the B cell population with an accompaniedYFP signal. Because this eVect was likely due to the pro-duction of cytokines, crosstalk with T lymphocytes, or sig-naling through the B cell’s Fc receptors we puriWed theB cells using MACs beads and tested them separately fortheir ability to respond to CD3/CD28 stimulation. Asshown in Fig. 3B, a 5.5-fold reduction in YFP reporterexpression on the B cell population was observed in thisassay when puriWed B cells were used as compared to spleen
cells. This suggests that the major signal observed in thesplenic B cells was likely due to crosstalk and/or cytokinesproduced by the T cells in the initial culture.
Treatment with Con A resulted in a slight activation andYFP signal, which may have resulted through similarmechanisms. It should be noted that peritoneal myeloidcells from the Line 19 mice did not express the YFP trans-gene when cultured with several agents, including LPS, Lis-teria monocytogenes, or PMA/Ion (data not shown). Thissuggests that expression may be limited to the lymphoidcompartment.
3.2. YFP expression correlates with cellular activation markers
Induction of CD69 is concomitant with initial activationof T lymphocytes [1,5]. To correlate the increase in forwardscatter and YFP expression with T cell activation, Line 19T cells were analyzed for expression of the activationmarker CD69. Splenocytes were stimulated with immobi-lized anti-CD3/CD28 antibodies, stained with anti-CD4,CD8, and CD69 antibodies. CD4 and CD8 positive cellswere gated and analyzed for expression of CD69 and theexpression of the YFP transgene by Xow cytometry (Fig. 4).The results showed greater than 96 and 83% of the CD4+and CD8+ T cells, respectively, were induced under theseconditions to express CD69. Moreover, the bulk of thisCD69+ population also expressed YFP. Less than 7% ofthe unstimulated transgenic splenic T cells were positive forCD69 and even fewer for YFP. Additionally, a similarexperiment was performed on puriWed B cells stimulatedwith LPS. The results show a signiWcant shift in the popula-tion of B cells (compare Fig. 3B lower left with 3C). Cou-pled with the increases in forward scatter shown in Fig. 2,these results suggest that the YFP-reporter transgene isinduced in response to cellular activation.
3.3. The IL-4YFP transgene can be activated by antigenic peptides independent of IL-4 production
Line 19 mice were crossed with the DO11.10 T cell recep-tor transgenic mice to determine if the IL-4YFP transgenecould be activated by antigen-speciWc peptides. Treatmentof the splenic lymphocytes from the double transgenic micewith PMA/Ion showed that the »90% of the CD69+ lym-phocytes were also positive for the reporter, indicating thatthe transgene was functioning similarly to the single trans-genic animals (Fig. 5). In comparison to the no antigen con-trol in which 5.8% of the T cells were CD69+ and with 3.2%of that population expressing the YFP reporter, stimula-tion with OVA 323–339 peptide produced a dose-depen-dent increase in both CD69 and YFP expression (Fig. 5).The maximum level of peptide used resulted in 72% of theT cells expressing the transgene. These results demonstratedthat the IL-4YFP transgene expression is coordinate withspeciWc antigenic stimulation of T cells in a dose speciWcmanner.

S. Kumar et al. / Cellular Immunology 237 (2005) 131–140 135
To determine whether IL-4 gene expression aVected over-all levels of IL-4YFP, the DO11.10 T cells were polarized tothe Th1 (non-IL-4 production) or Th2 (IL-4 production)phenotype using antigen stimulation and appropriate cyto-kines [8]. To drive diVerentiation toward the Th1 phenotype,
DO11.10 T cells were stimulated with 1�M of OVA 323–339peptide in the presence of IL-12 and antibody speciWc forIL-4 for seven days. For generation of Th2 cells, the DO11.10T cells were stimulated with 1�M OVA peptide with IL-4and antibody to IFN-�. Polarization was conWrmed by
Fig. 2. YFP is expressed in lymphocyte subsets after stimulation in vitro. Spleen cells from Tg+ mice (A) or Tg¡ controls (B) were incubated in vitro for24 h with T or B cell stimuli as follows: 10 ng/ml PMA, 1 �M ionomycin, 2 �g/ml Con A, 10 �g/ml immobilized anti-CD3, 2 �g/ml anti-CD28, or 2 �g/mlLPS. Lymphocyte subsets were identiWed with Xuorochrome-conjugated antibodies and analyzed by Xow cytometry. Cells were evaluated for YFP expres-sion after gating on either CD4+, CD8+, or B220+ cells. Changes in the forward scatter properties reXect changes in cell size under diVerent incubationconditions. Numbers in individual dot plots indicate the frequencies of YFP-expressing cells under each condition.
YFP EXPRESSION
MEDIUM PMA + IONOMYCIN CON A Anti-CD3/CD28 LPS
YFP EXPRESSION
FO
RW
AR
D S
CA
TT
ER
A
0.9% 64.2% 32.6% 69.0% 8.0%
1.7% 49.8% 38.0% 72.7% 4.3%
4.0% 86.9% 13.8% 42.3% 45.3%
CD4+
CD8+
B220+
MEDIUM PMA + IONOMYCIN LPS
FO
RW
AR
D S
CA
TT
ER
0.2% 0.6% 0.2%
4.4% 12.1% 4.9%
0.1% 0.6% 0.1%
B
CD4+
CD8+
B220+

136 S. Kumar et al. / Cellular Immunology 237 (2005) 131–140
analysis of IL-4 and IFN-� production by ELISA (data notshown). There was no apparent diVerence in the Xuorescentlevels between Th1 and Th2 DO11.10 cells when stimulated
Fig. 4. CD69 is expressed on cells that express YFP after inactivationin vitro with anti-CD3 and anti-CD28. Spleen cells from Tg+ mice wereincubated for 24 h in medium alone or with immobilized anti-CD3 andsoluble anti-CD28 as described for Fig. 2. Cells expressing CD4, CD8, orCD69 were identiWed using Xuorochrome-conjugated antibodies. Co-expression of CD69 and YFP was evaluated after gating on either CD4+or CD8+ cells. Cells from Tg¡ mice expressed CD69 but not YFP afterstimulation with anti-CD3 and anti-CD28.
YFP EXPRESSION
CD
69
2.3% 5.1%
6.3%86.2%
14.3% 82.0%
3.6% 0.1%
4.1% 2.6% 19.8% 63.7%
88.1% 14.7%5.3% 1.8%
CD4+
CD8+
MEDIUM Anti-CD3/CD28
over a range of peptide antigen doses (Fig. 6). For instance,65% of the Th1 cells and 66% of the Th2 cells were YFP pos-itive when stimulated with 10�M of OVA peptide. Theseresults indicate that the IL-4YFP transgene reXects T cellactivation independent of IL-4 production.3.4. The IL-4YFP transgene is expressed in viral speciWc T cells
To determine if the YFP transgene could be activated ina natural setting, the ability to detect the reporter followinga viral infection was examined. T cells from mice infectedwith polyoma virus were harvested after Wve days of infec-tion and stained with a polyoma speciWc MHC tetramer todetect viral speciWc CD8 T cells. While the MHC tetramer-positive cells represent only a small fraction of the CD8 Tcells (1%), Xow cytometry demonstrated that the vastmajority of the tetramer positive cells were YFP positive(Fig. 7). This result indicates that the reporter can be acti-vated in response to an infection.
3.5. Expression of the IL-4YFP transgene is inhibited by cyclosporine A
To decipher the signaling pathway that led to the induc-tion of the line 19 transgene, a panel of signaling inhibitors(Table 1), which block immunophilin, protein and calmod-ulin kinases, phosphatidylinositol kinases, tyrosinekinases, MAP and MEK kinases, and I�B/NF-�B
Fig. 3. PuriWed B cells do not respond to CD3/CD28 stimulation. (A) Splenic B cells were puriWed by negative selection using CD43 microbeads and wereanalyzed by staining with I-A antibodies. (B) Flow cytometry of splenic and puriWed B cells gated on B220+ cells co-stained with CD69 for populations ofcontrol cells or cells stimulated with CD3/CD28 antibodies for 48 h. (C) PuriWed B cells from the above experiment were treated with LPS for 24 h andanalyzed for CD69 and YFP expression.
10 0 10 1 10 2 10 3 10 40
20
40
60
Cel
l Nu
mb
er
I-A
0.9
4.5
YFP
CD
69
100
101
102
103
104
0.6 41.7
5.5
27.5
15.9 5.1
3.7
10 0 10 1 10 2 10 3 10 4100
101
102
103
104
1.1 1.0
5.1
10 0 10 1 10 2 10 3 10 4
Splenic B
cellsP
urified B cells
Control CD3/ CD28 Stimulated
C
A B
100 101 102 103 104100
101
102
103
104
37.9 20.1
9.3
CD
69
YFP

S. Kumar et al. / Cellular Immunology 237 (2005) 131–140 137
activation, were examined for their ability to inhibitexpression of the transgene. Line 19 spleen cells were stim-ulated with PMA/Ion, immobilized anti-CD3/CD28 anti-bodies, or LPS, treated with the inhibitor, and examined
Fig. 5. DO11.10 £ IL-4YFP splenocytes respond to OVA 323–339 in adose-dependent manner. Splenocytes were harvested from the doubletransgenic DO11.10 £ YFP mice and restimulated for 24 h in the presenceor absence of OVA 323–329 peptide or PMA/Ion. The cells were washed,stained with anti-CD4, anti-CD69, and KJ126 (the DO11.10 clonotypicantibody), and analyzed by Xow cytometry. The data shown are gated onKJ126+ and CD4+ lymphocytes. The percentage of CD69+ and YFP+cells with each subset is indicated.
0.1 1.0 10 1000
10
20
30
40
50
60
70Th1Th2
OVA peptide dose µM
% Y
EP
po
siti
ve T
cel
ls
by Xow cytometry for the expression of the transgene inCD4+ T and B cells. At the dosages tested, only those thatblocked the action of the immunophilins cyclosporine A,FK506, and rapamycin were able to substantially reducethe induction of the transgene, suggesting that the NFATpathway, which is known to regulate IL-4, is likely to beinvolved in the expression of the transgene. Cyclosporin Ainhibited PMA/Ion induction of the transgene by >50% inboth CD4+ T and B220+ B cells, but did not inhibit LPS-induced B cell expression of the transgene. FK506 had amore pronounced eVect on the induction of the transgenein T cells than in B cells. In contrast, rapamycin displayedlittle eVect on T cells, but was best at inhibiting transgeneexpression in LPS-stimulated B220+ B cells. The ability ofthese agents to diVerentially inhibit transgene activation ina manner that was dependent on the activating signal sug-gests that the IL-4YFP reporter can be induced throughseveral distinct signals. Mild eVects on the activation of theLine 19 transgene were also observed in CD4+ T cellstreated with the general serine/threonine protein kinaseinhibitor staurosporin and the MAP kinase inhibitor5�-iodotubericidin. These mild eVects may implicate addi-tional or convergent pathways that inXuence the expres-sion of the IL-4YFP transgene.
Fig. 6. DO11.10 Th1 and Th2 cells express equivalent levels of YFP. DO11.10 T cells were polarized toward the Th1 or Th2 phenotype for seven days. TheT cells were puriWed and rechallenged with the indicated dose of OVA 323–339 peptide presented by spleen cells as APC. After 24 h, YFP expression of the
89.7 13.9 54.8 72.1
No antigen PMA/ionomycin 0.1µM 1µM 10µM
CD
69
104
3.16104
103
102
101
1 102 1031011
OVA 323-339
DO11.10 T cells (KJ126+) was analyzed by Xow cytometry.
Fig. 7. Polyoma speciWc T cells express the YFP transgene. IL-4YFP transgenic and non-transgenic animals were infected sc with polyoma virus. Five dayslater spleens were analyzed for the presence of polyoma virus Db-LT359 speciWc CD8 T cells using MHC tetramers. The plot is gated on CD8 T cells andthe number indicates the percent tetramer+ cells within the CD8 population. Note that the tetramer positive population of cells is directly over the peak ofnegative cells in the non-transgenic control group but shifted to the right in the YFP-transgene group. These experiments were performed two times with
1.51.1
YFP
Db L
T35
9 te
tram
er 0.1
infected Tginfected B6naive B6
similar results.
138 S. Kumar et al. / Cellular Immunology 237 (2005) 131–140
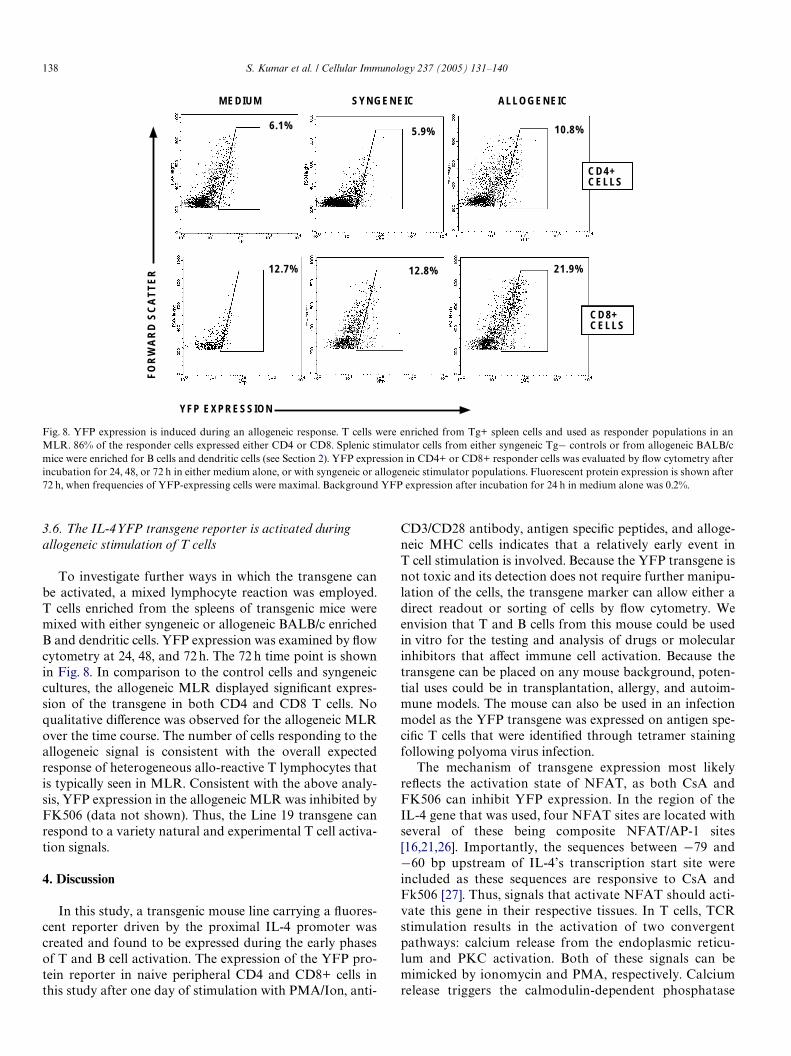
3.6. The IL-4YFP transgene reporter is activated during allogeneic stimulation of T cells
To investigate further ways in which the transgene canbe activated, a mixed lymphocyte reaction was employed.T cells enriched from the spleens of transgenic mice weremixed with either syngeneic or allogeneic BALB/c enrichedB and dendritic cells. YFP expression was examined by Xowcytometry at 24, 48, and 72 h. The 72 h time point is shownin Fig. 8. In comparison to the control cells and syngeneiccultures, the allogeneic MLR displayed signiWcant expres-sion of the transgene in both CD4 and CD8 T cells. Noqualitative diVerence was observed for the allogeneic MLRover the time course. The number of cells responding to theallogeneic signal is consistent with the overall expectedresponse of heterogeneous allo-reactive T lymphocytes thatis typically seen in MLR. Consistent with the above analy-sis, YFP expression in the allogeneic MLR was inhibited byFK506 (data not shown). Thus, the Line 19 transgene canrespond to a variety natural and experimental T cell activa-tion signals.
4. Discussion
In this study, a transgenic mouse line carrying a Xuores-cent reporter driven by the proximal IL-4 promoter wascreated and found to be expressed during the early phasesof T and B cell activation. The expression of the YFP pro-tein reporter in naive peripheral CD4 and CD8+ cells inthis study after one day of stimulation with PMA/Ion, anti-
CD3/CD28 antibody, antigen speciWc peptides, and alloge-neic MHC cells indicates that a relatively early event inT cell stimulation is involved. Because the YFP transgene isnot toxic and its detection does not require further manipu-lation of the cells, the transgene marker can allow either adirect readout or sorting of cells by Xow cytometry. Weenvision that T and B cells from this mouse could be usedin vitro for the testing and analysis of drugs or molecularinhibitors that aVect immune cell activation. Because thetransgene can be placed on any mouse background, poten-tial uses could be in transplantation, allergy, and autoim-mune models. The mouse can also be used in an infectionmodel as the YFP transgene was expressed on antigen spe-ciWc T cells that were identiWed through tetramer stainingfollowing polyoma virus infection.
The mechanism of transgene expression most likelyreXects the activation state of NFAT, as both CsA andFK506 can inhibit YFP expression. In the region of theIL-4 gene that was used, four NFAT sites are located withseveral of these being composite NFAT/AP-1 sites[16,21,26]. Importantly, the sequences between ¡79 and¡60 bp upstream of IL-4’s transcription start site wereincluded as these sequences are responsive to CsA andFk506 [27]. Thus, signals that activate NFAT should acti-vate this gene in their respective tissues. In T cells, TCRstimulation results in the activation of two convergentpathways: calcium release from the endoplasmic reticu-lum and PKC activation. Both of these signals can bemimicked by ionomycin and PMA, respectively. Calciumrelease triggers the calmodulin-dependent phosphatase
Fig. 8. YFP expression is induced during an allogeneic response. T cells were enriched from Tg+ spleen cells and used as responder populations in anMLR. 86% of the responder cells expressed either CD4 or CD8. Splenic stimulator cells from either syngeneic Tg¡ controls or from allogeneic BALB/cmice were enriched for B cells and dendritic cells (see Section 2). YFP expression in CD4+ or CD8+ responder cells was evaluated by Xow cytometry afterincubation for 24, 48, or 72 h in either medium alone, or with syngeneic or allogeneic stimulator populations. Fluorescent protein expression is shown after72 h, when frequencies of YFP-expressing cells were maximal. Background YFP expression after incubation for 24 h in medium alone was 0.2%.
MEDIUM SYNGENEIC ALLOGENEIC
YFP EXPRESSION
FO
RW
AR
D S
CA
TT
ER
6.1% 5.9% 10.8%
21.9%12.8%12.7%
CD4+CELLS
CD8+CELLS

S. Kumar et al. / Cellular Immunology 237 (2005) 131–140 139
activity of calcineurin and the subsequent activation ofNFAT, whereas PKC induces the activity of AP-1 familymembers, including the binding of AP-1 to the NFATcomposite sequences [16]. Thus, the T cell activationsignals used in this study likely induce NFAT and AP-1factors, which bind the transgenic promoter and transacti-vate the YFP gene.
In addition to NFAT, several other factors have beenfound to be associated with IL-4 expression and includeGATA3 [10], c-maf [9], and STAT6 [11]. Each of these fac-tors can interact with the promoter region included in thetransgene used in this report and are likely candidates forits expression. While the above factors are likely to play adirect role in the activation of the transgene in the T cellpopulation, we can only speculate that NFAT family mem-bers along with unknown factors that regulate the trans-gene in other cell types, such as B cells [28–30].
One of our initial objectives was to determine if theYFP transgene’s expression would follow the IL-4 tran-scription program associated with Th2 cells. Severalexperimental conditions were examined to polarize Th1and Th2 cells (Fig. 6). While high levels of IFN-� and IL-4were detected by ELISA in their respective polarized pop-ulations (data not shown), expression of the transgene inthese long-term cultures could not be directly correlatedwith IL-4 expression in the polarized cells, as both Th1and Th2 DO11.10 T cells were equally Xuorescent. In con-trast to our system, a EGFP/IL-4 bicistronic reporterknock-in mouse model [31] found that T cells primedunder Th2 conditions showed sensitive EGFP expressionand maintained endogenous IL-4 expression. Similarly,the system described by Paul and co-workers using anIL-4-GFP knock-in also behaves faithfully with respect toIL-4 regulated expression [32]. Additionally, when a simi-lar IL-4 upstream sequence was used to drive a �-galacto-sidase transgene, Th2 speciWc transgene expression wasobserved in some but not all of the founders [22]. The lat-ter system indicates that the location of the transgeneintegrating event is critical to the ability of the transgeneto be appropriately regulated. IL-4 transcription regula-tion is complex and its locus requires the reorganizationof the local chromatin structure through the aid of multi-ple regulatory elements located at various sites within thelocus [12,33]. Thus, these results may suggest that theadditional control regions in the IL-4 gene not only allowinitiation of IL-4 gene expression but also provide for itsmaintenance during Th1/Th2 diVerentiation. Thus, wesuggest that the IL-4YFP transgene described here hasintegrated into a chromosomal location that provides aunique regulation and response to immune signals.
Despite the lack of Wdelity with IL-4 gene activity, theexpression of this transgene in CD4, CD8, and B cells, andits ease of detection make this system ideal to track and iso-late activated cell populations. Additionally, the use of thistransgene as a reporter for immunosuppressive drugswould be extremely valuable as it marks cells at an earlystep in cellular activation.
Acknowledgments
We thank the Emory University Transgenic facility fortheir eVorts in the generation and initial breeding of theanimals in this study. We also thank Tracey Walden fortechnical assistance. This work was supported by an NIHgrant R21RR15183 and an Emory University FacultyResearch Award.
References
[1] S.M. Kaech, R. Ahmed, Memory CD8+ T cell diVerentiation: initialantigen encounter triggers a developmental program in naive cells,Nat. Immunol. 2 (2001) 415–422.
[2] K.H. Lee, A.D. Holdorf, M.L. Dustin, A.C. Chan, P.M. Allen, A.S.Shaw, T cell receptor signaling precedes immunological synapse for-mation, Science 295 (2002) 1539–1542.
[3] M.J. van Stipdonk, E.E. Lemmens, S.P. Schoenberger, Naive CTLsrequire a single brief period of antigenic stimulation for clonal expan-sion and diVerentiation, Nat. Immunol. 2 (2001) 423–429.
[4] S.M. Kaech, S. Hemby, E. Kersh, R. Ahmed, Molecular and func-tional proWling of memory CD8 T cell diVerentiation, Cell 111(2002) 837–851.
[5] S. Nakamura, S.S. Sung, J.M. Bjorndahl, S.M. Fu, Human T cell acti-vation. IV.T cell activation and proliferation via the early activationantigen EA 1, J. Exp. Med. 169 (1989) 677–689.
[6] R.D. Sanderson, P. Lalor, M. BernWeld, B lymphocytes express andlose syndecan at speciWc stages of diVerentiation, Cell Regul. 1(1989) 27–35.
[7] C. Angelin-Duclos, G. Cattoretti, K.I. Lin, K. Calame, Commitmentof B lymphocytes to a plasma cell fate is associated with Blimp-1expression in vivo, J. Immunol. 165 (2000) 5462–5471.
[8] T.R. Mosmann, R.L. CoVman, TH1 and TH2 cells: diVerent patternsof lymphokine secretion lead to diVerent functional properties, Annu.Rev. Immunol. 7 (1989) 145–173.
[9] L.-C. Ho, M.R. Hodge, J.W. Rooney, L.H. Glimcher, The proto-onco-gene c-maf is responsible for tissue-speciWc expression of interleukin-4, Cell 85 (1996) 973–983.
[10] W. Zheng, R.A. Flavell, The transcription factor GATA-3 is necessaryand suYcient for Th2 cytokine gene expression in CD4 T cells, Cell 89(1997) 587–596.
[11] M. Kubo, J. Ransom, D. Webb, Y. Hashimoto, T. Tada, T. Nakay-ama, T-cell subset-speciWc expression of the IL-4 gene is regulated bya silencer element and STAT6, EMBO J. 16 (1997) 4007–4120.
[12] P.E. Fields, G.R. Lee, S.T. Kim, V.V. Bartsevich, R.A. Flavell, Th2-speciWc chromatin remodeling and enhancer activity in the Th2 cyto-kine locus control region, Immunity 21 (2004) 865–876.
[13] L. Guo, J. Hu-Li, J. Zhu, C.J. Watson, M.J. DiWlippantonio, C. Panne-tier, W.E. Paul, In TH2 cells the Il4 gene has a series of accessibilitystates associated with distinctive probabilities of IL-4 production,Proc. Natl. Acad. Sci. USA 99 (2002) 10623–10628.
[14] H. Yoshida, H. Nishina, H. Takimoto, L.E. Marengere, A.C. Wake-ham, D. Bouchard, Y.Y. Kong, T. Ohteki, A. Shahinian, M. Bach-mann, P.S. Ohashi, J.M. Penninger, G.R. Crabtree, T.W. Mak, Thetranscription factor NF-ATc1 regulates lymphocyte proliferation andTh2 cytokine production, Immunity 8 (1998) 115–124.
[15] T.M. Aune, R.A. Flavell, DiVerential expression of transcriptiondirected by a discrete NF-AT binding element from the IL-4 pro-moter in naive and eVector T cells, J. Immunol. 159 (1997) 36–43.
[16] J.W. Rooney, Y.L. Sun, L.H. Glimcher, T. Hoey, Novel NFATsites that mediate activation of the interleukin-2 promoter inresponse to T-cell receptor stimulation, Mol. Cell. Biol. 15 (1995)6299–6310.
[17] A. Rao, NF-ATp: a transcription factor required for the co-ordinateinduction of several cytokine genes, Immunol. Today 15(1994) 274–281.

140 S. Kumar et al. / Cellular Immunology 237 (2005) 131–140
[18] A. Rao, C. Luo, P.G. Hogan, Transcription factors of the NFAT fam-ily: regulation and function, Annu. Rev. Immunol. 15 (1997) 707–747.
[19] G.R. Crabtree, E.N. Olson, NFAT signaling: choreographing thesocial lives of cells, Cell 109 (Suppl.) (2002) S67–S79.
[20] S.L. Schreiber, G.R. Crabtree, The mechanism of action of cyclo-sporin A and FK506, Immunol. Today 13 (1992) 136–142.
[21] M.A. Brown, J. Hural, Functions of IL-4 and control of its expression,CRC Crit. Rev. Immunol. 17 (1997) 1–32.
[22] C. Wenner, S.J. Szabo, K. Murphy, IdentiWcation of IL-4 promoterelements conferring Th2-restricted expression during T helper cellsubset development, J. Immunol. 158 (1997) 765–773.
[23] N. Takemoto, N. Koyano-Nakagawa, N. Arai, K.-I. Arai, T. Yokota,Four P-like elements are required for optimal transcription of themouse IL-4 gene: involvement of a distinct set of nuclear factor ofactivated T cells and activator protein-1 family proteins, Int. Immu-nol. 9 (1997) 1329–1338.
[24] K.M. Murphy, A.B. Heimberger, D.Y. Loh, Induction by antigen ofintrathymic apoptosis of CD4+CD8+TCRlo thymocytes in vivo, Sci-ence 250 (1990) 1720–1723.
[25] A.M. Byers, C.C. Kemball, J.M. Moser, A.E. Lukacher, Cutting edge:rapid in vivo CTL activity by polyoma virus-speciWc eVector andmemory CD8+ T cells, J. Immunol. 171 (2003) 17–21.
[26] S.J. Szabo, J.S. Gold, T.L. Murphy, K.M. Murphy, IdentiWcation ofcis-acting regulatory elements controlling interleukin-4 gene expres-sion in T cells: roles for NF-Y and NF-ATc, Mol. Cell. Biol. 13 (1993)4793–4805.
[27] M. Kubo, R.L. Kincaid, J.T. Ransom, Activation of the interleukin-4gene is controlled by the unique calcineurin-dependent transcriptionalfactor NF(P), J. Biol. Chem. 269 (1994) 19441–19446.
[28] K.B. Meyer, J. Ireland, PMA/ionomycin induces Ig kappa 3� enhanceractivity which is in part mediated by a unique NFAT transcriptioncomplex, Eur. J. Immunol. 28 (1998) 1467–1480.
[29] R. Berland, H.H. Wortis, An NFAT-dependent enhancer is necessaryfor anti-IgM-mediated induction of murine CD5 expression in pri-mary splenic B cells, J. Immunol. 161 (1998) 277–285.
[30] S.L. Peng, A.J. Gerth, A.M. Ranger, L.H. Glimcher, NFATc1 andNFATc2 together control both T and B cell activation and diVerenti-ation, Immunity 14 (2001) 13–20.
[31] M. Mohrs, K. Shinkai, K. Mohrs, R.M. Locksley, Analysis of type 2immunity in vivo with a bicistronic IL-4 reporter, Immunity 15 (2001)303–311.
[32] J. Hu-Li, C. Pannetier, L. Guo, M. Lohning, H. Gu, C. Watson, M.Assenmacher, A. Radbruch, W.E. Paul, Regulation of expression ofIL-4 alleles: analysis using a chimeric GFP/IL-4 gene, Immunity 14(2001) 1–11.
[33] C.G. Spilianakis, R.A. Flavell, Long-range intrachromosomal interac-tions in the T helper type 2 cytokine locus, Nat. Immunol. 5 (2004)1017–1027.
[34] J. Jain, P.G. McCaVrey, Z. Miner, T.K. Kerppola, J.N. Lambert, G.L.Verdine, T. Curran, A. Rao, The T-cell transcription factor NFATp isa substrate for calcineurin and interacts with Fos and Jun, Nature 365(1993) 352–355.





























