Embed Size (px)
Citation preview

A critical review of the literature on the pest Erionota spp.(Lepidoptera, Hesperiidae): taxonomy, distribution, food plants,early stages, natural enemies and biological control
Matthew J. W. Cock*
Address: CABI Europe – UK, Bakeham Lane, Egham TW20 9TY, UK
*Correspondence: Matthew J. W. Cock. Email: [email protected]
Received: 2 December 2014
Accepted: 15 February 2015
doi: 10.1079/PAVSNNR201510007
The electronic version of this article is the definitive one. It is located here: http://www.cabi.org/cabreviews
g CAB International 2015 (Online ISSN 1749-8848)
Abstract
The classification, taxonomy, distribution, spread, food plants, early stages, natural enemies
and biological control of South-East Asian Erionota spp. (Hesperiidae, Hesperiinae, incertae sedis)
are reviewed with specific attention to Erionota thrax (Linnaeus), Erionota torus Evans and Erionota
acroleuca (Wood-Mason and de Niceville) (=E. hiraca (Moore)). Erionota thrax and E. torus are
shown to be pests of Musa spp. but not palms, while E. acroleuca is a pest of various palms. Where
they overlap in distribution in mainland South-East Asia and the Southern Philippines, E. thrax
and E. torus have not been distinguished in the economic literature. The species which became
established in Mauritius around 1968 and subsequently the target of a successful biological control
programme is shown to be E. torus, and not E. thrax as hitherto reported. Research gaps are
identified, including the need to find diagnostic features or tools to separate the immature stages
of the three species in order to confirm these conclusions, and clarify the ecology and pest status
of the pest species.
Keywords: Erionota thrax, Erionota torus, Erionota acroleuca, Banana, Musa, Palm, Arecaceae.
Review Methodology: Because these pests are large butterflies, there is an extensive literature based on regional butterfly books and
lists as well as the applied entomological literature. The author attempted to track down and examine all available information on the
genus Erionota, particularly the applied literature dealing with the biology, pest status and control of the pest species. The taxonomic
work of Evans [1] and the CABI Crop Protection Compendium datasheet for E. thrax were important starting points. This was
supplemented by searching CAB Abstracts, internet searching, and consulting selected reference works dealing with butterflies and
pests in the region. The information sources were traced back to their original publication whenever possible. The author familiarised
himself with the biology of the two banana skippers (E. thrax and E. torus) by examination of field material in West Malaysia, which
helped considerably in interpreting the literature for this review. The Erionota spp. material in the insect collections of the Natural
History Museum, London, the Hope Entomological Collections (Oxford University Museum) and the Forest Research Institute of
Malaysia were examined to check for locality records, voucher specimens and biological information; the genitalia figures of Inoue and
Kawazoe [2] enabled several misidentifications in these collections to be recognised by examination of the externally visible genitalia.
The taxonomy of the natural enemies and plant names were checked against key online databases. As indicated in the
acknowledgements a variety of informed individuals were consulted on specific points.
Introduction
Erionota thrax (Linnaeus) is well known as ‘the banana
skipper’ or ‘banana leaf roller’ in South-East Asia, and
as an introduced pest in various Pacific territories. It
is amongst the largest members of the family Hesperiidae
(Figure 1). A second species, Erionota torus Evans
(Figure 2), has a very similar biology, also occurs in South-
East Asia and as an introduced in Mauritius, Taiwan and
Japan (Okinawa). Both have been the target of successful
biological control programmes where they have been
introduced. A third, Erionota acroleuca (Wood-Mason
and de Niceville) (Figure 3) is similar in appearance but
the larvae feed on palms. All three overlap in much
http://www.cabi.org/cabreviews
CAB Reviews 2015 10, No. 007

of their distribution in South-East Asia and there is an
extensive literature which indicates considerable confu-
sion as to the Erionota spp. concerned. In this review,
I attempt to untangle this literature, correct some errors,
and identify aspects needing further attention.
Classification
Erionota Mabille is a genus in the subfamily Hesperiinae,
family Hesperiidae. In his work on the Hesperiidae of
Europe, Asia and Australia, Evans [1] grouped seven
Figure 1 Pinned adults of E. thrax thrax, reared on Musa sp., Luzon Is, Philippines; male on the left, female on the right;dorsal surface above, ventral surface below; life size, the scale bar is 1 cm.
Figure 2 Pinned adults of E. torus; male on the left, female on the right; dorsal surface above, ventral surface below; lifesize, the scale bar is 1 cm. Male: SE China, Macao, 20 Aug 1904, J. C. Kershaw; female: west Malaysia, Negri Sembilan,Seremban, Feb 1914, V.G Bell and J. W. B. Bell (specimens in Hope Entomological Collections, Oxford UniversityMuseum). The specimens illustrated are less dark than those of the other two Erionota spp. illustrated because they havefaded with age. Erionota torus often cannot be reliably distinguished from E. thrax thrax (Figure 1) without examination of thegenitalia.
http://www.cabi.org/cabreviews
2 CAB Reviews

Oriental genera as the Erionota sub-group within the
Plastingia group of Hesperiinae. In their work on the
genitalia of the Hesperiidae of South Vietnam, Inoue and
Kawazoe [2] suggested affinities between Lotongus Distant
([1] Plastingia subgroup), Gangara Moore, Erionota, Matapa
Moore ([1] Erionota subgroup), Unkana Distant, Hidari
Distant and Pirdana Distant ([1] Unkana subgroup)
based primarily on characters of the male genitalia.
Although Warren [3] treated Erionotini as a provisional
tribe of Hesperiinae in his Ph.D. thesis on the classification
of the Hesperioidea, this was a much enlarged group of
genera, which in his subsequent co-authored publications
[4, 5] were placed incertae sedis.
Of the seven genera that Evans grouped in his Erionota
subgroup, no information has been located on the early
stages of species of Zela De Niceville, Ilma Swinhoe, Ge
De Niceville or Pudicitia De Niceville, but there is infor-
mation on Gangara, Erionota and Matapa. Gangara thyrsis
(Fabricius) feeds on various palms and the early
stages have been illustrated in Igarashi and Fukuda [6] and
Cock et al. [7] among others. Matapa aria (Moore) is
a bamboo-feeder [8], and the early stages are illustrated
by Bascombe et al. [9] and Saji et al. [10]. Erionota spp. feed
on palms (Arecaceae), Musa spp. (Musaceae, bananas and
plantains) and some related Zingiberales. Igarashi and
Fukuda [6] illustrate the early stages of E. torus, E. thrax
and E. grandis (Leech), while Bascombe et al. [9] also
illustrate the early stages of E. torus. The leaf shelters
of at least some members of all three genera are note-
worthy for being tubes made by rolling palm leaflets
or cut sections of Musa leaves into a tube, either in a
spiral (Gangara, Matapa) or in a roll (Erionota). This
behaviour separates them from all other Hesperiinae
familiar to the author, and supports their recognition as a
distinct group.
There is information available on the early stages of
the additional genera that Inoue and Kawazoe [2] sug-
gested could be grouped with Erionota. Lotongus calathus
(Hewitson) has an unusual biology as documented by
Igarashi and Fukuda [11]. The distinctive ova are flattened
with a grooved ring around the top. They were found laid
singly, and the newly hatched larvae make individual
leaf folds on their unidentified palm seedling food plant.
The second instar larvae then aggregate to make a
communal shelter, which is always co-occupied by a small
ant, Dolichoderus bituberculatus Mayr, which does not
attack the larvae. The mature larva has a four-lobed black
marking on the face, reminiscent of those of some African
Artitropa spp. [12, 13]. The larvae pupate in a stout
cocoon within the shelter, and the pupa is pale brown
with no striking features. Igarashi and Fukuda [11] docu-
ment the life history of Unkana ambasa (Moore) and
Unkana mytheca (Hewitson) on Pandanus odorifer (as
P. odoratissimus, Pandanaceae). The ova are smooth and
domed, similar to those of many other Hesperiinae;
the larvae are translucent grey-green with dark, oval
heads, and a dark anal plate in the case of U. ambasa;
the leaf shelters are cylindrical and made by rolling the
leaf along the axis of the mid-rib; the pupae are brown,
the proboscis reaching the cremaster. Rearing revealed
Figure 3 Pinned adults of E. acroleuca apicalis; male on the left, female on the right; dorsal surface above, ventral surfacebelow; life size, the scale bar is 1 cm. Both collected as larvae on Caryota sp. (Arecaceae), Peninsular Malaysia, Selangor,Kedong, Forest Research Institute of Malaysia, 2 Apr 1988, L. G. Kirton, rearing reference B:5 (specimens in ForestResearch Institute of Malaysia). The extent of the white apical forewing apex is variable, especially in the female, and whenit is reduced this species is difficult to distinguish from E. thrax (Figure 1) and E. torus (Figure 2).
http://www.cabi.org/cabreviews
Matthew J. W. Cock 3

that the all-female species Erionota harmachis (Hewitson)
is a synonym of the all-male species U. mytheca [14],
indicating affinities in adult structure characters. Hidari
irava (Moore) makes leaf shelters between leaflets of
its palm hosts where small numbers of larvae cohabit
[11, 15, 16]. Pirdana hyela (Hewitson) and Pirdana distanti
(Staudinger) feed on Dracaena spp. (Asparagaceae)
[11]. The ovum is strongly ribbed; the head and body of
the larva of P. hyela superficially resembles that of an
Artitropa sp. [12, 13], while that of P. distanti has a
similar head, but the body is dark with a yellow dorsal
line and yellow dorsolateral transverse bars; leaf shelters
are made by rolling the food plant leaf longitudinally; the
pupa has a short, blunt, frontal spike. The rather diverse
life histories of representatives of these genera do not
suggest any obvious affinities with Erionota, unlike that
pointed out for Gangara and Matapa above.
Taxonomy
The checklist in Table 1 presents the current position.
The species of immediate concern as pests are E. torus,
E. thrax and E. acroleuca. However, as some or all the
other species also feed on Musacaeae and Arecaceae as
larvae, they are all of concern as potential quarantine
pests.
Linnaeus [22] described Papilio thrax from Java, referring
to an un-named image in Clerck (36, plate 42.2). Honey
and Scoble [37] explain that there are two specimens
of E. thrax in the Linnean Collection, one of which has
been designated the lectotype, but as Mabille [20] also
pointed out, Clerck’s [36] image is of a different, unre-
lated species (a South American species of the genus
Thracides).
Evans [21] described E. torus from Sikkim and gave
characters to separate the adults from E. thrax based
on appearance and male genitalia. In his catalogue of the
Hesperiidae of Europe, Asia and Australia [1] he added his
diagrammatic figures of the male genitalia. Males of E. torus
(Figure 2) can usually but not always be distinguished
from those of E. thrax (Figure 1) by the more rounded
distal margin of the forewing in E. torus compared to
the straighter margin and apically more pointed forewing
of E. thrax. Separation of the two species, particularly
females, is only reliable if based on examination of the
genitalia ([38], H. Chiba, personal communication, 2014;
author’s observations). It is fortunate that the male
and female genitalia of both species have been accurately
documented and drawn by Inoue and Kawazoe [2] and
available on the internet. Corbet et al. [39] include
figures of the male genitalia of both species, and the male
genitalia and female venation of E. torus are illustrated
by Bascombe et al. [9] from Hong Kong. The diagnostic
characters of both male and female genitalia can usually be
seen by brushing off the scales of the abdomen tip using a
short, fine paint brush, obviating the need for traditional
dissection. In particular, the strongly upturned, more
heavily chitinized apical process of the valves of male
E. torus, compared with the simple, blunt apical process
of E. thrax can often be distinguished. A recent student
thesis yet to be published [40] concluded that ‘the banana
skippers, E. thrax and E. torus, are a monophyletic group
which represent a single host shift from Arecaceae or
Zingiberaceae to Musaceae’.
There is some confusion regarding the correct name
for E. acroleuca [19]. It was first described by Wood-
Mason and de Niceville [25] in August 1881 as Telegonus
acroleucus in the abstract of a paper which was not pub-
lished in full until December that year when the combi-
nation Hesperia acroleuca was used [26]. In the interim, in
September, Moore [27] independently described the same
species as Hesperia hiraca. The description in the abstract
[25] gives the new species name, indicates that it is new
species, gives a brief description and comparison with
E. thrax, and indicates many specimens from ‘S. Andaman’,
i.e. it meets the requirements for the publication of the
new name and so has precedence over Moore’s name.
Gender agreement has been applied under the ICZN [41],
i.e. E. acroleuca is correct rather than E. acroleucus fol-
lowing the treatments of De Jong and Treadaway [17] and
Xue and Lo [19]. The male of E. acroleuca has the apex
of the forewing upper side white, which distinguishes it
from the males of E. thrax and E. torus. The female is
generally smaller than those of E. thrax and E. torus, but
the white patch at the forewing apex is variable, and
some specimens are not reliably distinguishable except
by examination of the genitalia. Fortunately, Inoue and
Kawazoe [2] again provide good drawings of both male
and female genitalia of this species (ssp. apicalis).
The authorship of ssp. apicalis also needed clarification.
Xue and Lo [19] showed that when Evans [29] published
the name as var. apicalis, this made it an unavailable
infraspecific name under Article 45.5.1 of the International
Code of Zoological Nomenclature [41], and it should
take the authorship from the first publication to use it as
a valid name, i.e. De Jong and Treadaway [17], as listed in
Table 1.
Astictopterus harmachis Hewitson was described from
Sumatra [42], and was subsequently treated as a species
of Erionota [1, 21]. However, it is now known to be the
female of U. mytheca, which had previously only been
known from the male [11, 14].
Distribution and Spread
There are four subspecies of E. thrax recognized in
South-East Asia: E. thrax thrax widespread from north-east
India to the Philippines, Sulawesi and Java, E. thrax mindana
Evans in the southern Philippines but overlapping with
E. thrax thrax in the northern Philippines, E. thrax alexandra
Semper from upland northern Luzon and E. thrax has-
drubal Fruhstorfer from Kep. Sula (Mangole, Sanana),
http://www.cabi.org/cabreviews
4 CAB Reviews
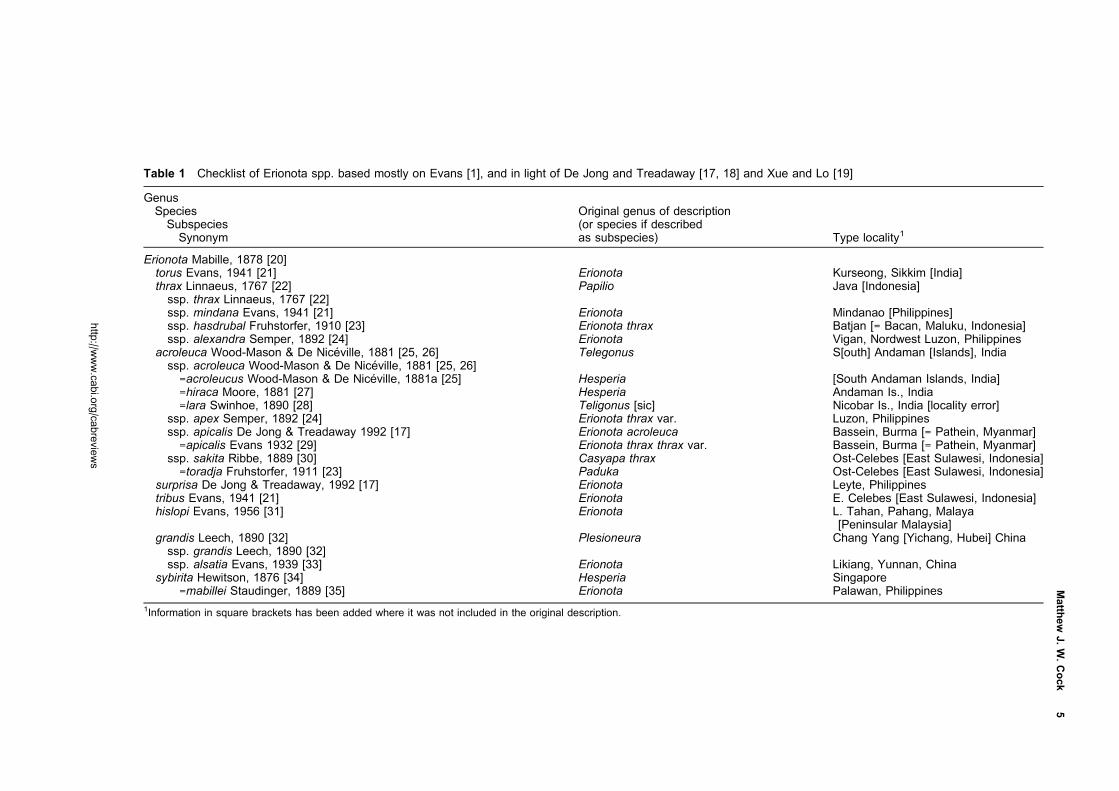
Table 1 Checklist of Erionota spp. based mostly on Evans [1], and in light of De Jong and Treadaway [17, 18] and Xue and Lo [19]
GenusSpeciesSubspeciesSynonym
Original genus of description(or species if describedas subspecies) Type locality1
Erionota Mabille, 1878 [20]torus Evans, 1941 [21] Erionota Kurseong, Sikkim [India]thrax Linnaeus, 1767 [22] Papilio Java [Indonesia]ssp. thrax Linnaeus, 1767 [22]ssp. mindana Evans, 1941 [21] Erionota Mindanao [Philippines]ssp. hasdrubal Fruhstorfer, 1910 [23] Erionota thrax Batjan [= Bacan, Maluku, Indonesia]ssp. alexandra Semper, 1892 [24] Erionota Vigan, Nordwest Luzon, Philippines
acroleuca Wood-Mason & De Niceville, 1881 [25, 26] Telegonus S[outh] Andaman [Islands], Indiassp. acroleuca Wood-Mason & De Niceville, 1881 [25, 26]=acroleucus Wood-Mason & De Niceville, 1881a [25] Hesperia [South Andaman Islands, India]=hiraca Moore, 1881 [27] Hesperia Andaman Is., India=lara Swinhoe, 1890 [28] Teligonus [sic] Nicobar Is., India [locality error]
ssp. apex Semper, 1892 [24] Erionota thrax var. Luzon, Philippinesssp. apicalis De Jong & Treadaway 1992 [17] Erionota acroleuca Bassein, Burma [= Pathein, Myanmar]=apicalis Evans 1932 [29] Erionota thrax thrax var. Bassein, Burma [= Pathein, Myanmar]
ssp. sakita Ribbe, 1889 [30] Casyapa thrax Ost-Celebes [East Sulawesi, Indonesia]=toradja Fruhstorfer, 1911 [23] Paduka Ost-Celebes [East Sulawesi, Indonesia]
surprisa De Jong & Treadaway, 1992 [17] Erionota Leyte, Philippinestribus Evans, 1941 [21] Erionota E. Celebes [East Sulawesi, Indonesia]hislopi Evans, 1956 [31] Erionota L. Tahan, Pahang, Malaya
[Peninsular Malaysia]grandis Leech, 1890 [32] Plesioneura Chang Yang [Yichang, Hubei] Chinassp. grandis Leech, 1890 [32]ssp. alsatia Evans, 1939 [33] Erionota Likiang, Yunnan, China
sybirita Hewitson, 1876 [34] Hesperia Singapore=mabillei Staudinger, 1889 [35] Erionota Palawan, Philippines
1Information in square brackets has been added where it was not included in the original description.
http
://www.cabi.o
rg/cabreviews
Matth
ew
J.W.Cock
5

North Moluku, Indonesia [1, 18, 43]. Of these, only
E. thrax thrax is known to have spread beyond its indi-
genous range.
In the Pacific E. thrax thrax spread to Guam by 1957
[44] and Saipan in the Northern Mariana Islands by
the late 1960s [45], to Oahu, Hawaii by 1973 [46], to
Irian Jaya, east Indonesia some years before 1983 [47],
to mainland Papua New Guinea (PNG) in 1983 [45],
to New Britain, PNG, in 1991 [48], to Duke of York
and New Ireland islands, PNG, by 1994, and possibly to
Bougainville, PNG, about this time [47]. Muniappan [49]
refers to the original introduction into Guam as being
from the Philippines, based on both localities having
American air force and naval bases, and the heavy civilian
traffic between the Philippines and Guam (R. Muniappan,
personal communication, 2014). Within the Hawaiian
Islands it spread from Oahu to Kauai and Maui in 1974 and
Hawaii, Molokoi and Lanai in 1975 [50, 51].
The records from Guam, Saipan and Hawaii have been
referred to incorrectly as E. torus, e.g. Igarashi and Fukuda
[11], Opler and Warren [52] and perhaps others. The
origin of this error and its correction are outlined in
Pelham [53]. Kawazoe and Wakabayashi [54] illustrated
E. thrax as E. torus and stated that it occurs in Hawaii
and Guam. Snyder and Kumon [55] and Fukuda and Nicho
[56] clarified that E. thrax is indeed the species occurring
in Hawaii, not E. torus.
Although Parsons [57] treats the population that
spread into PNG as E. thrax hasdrubal, the specimen that
he illustrates is E. thrax thrax, based on the characters in
Evans [1, 21], and is treated as such by Waterhouse and
Norris [45] and Sands et al. [58]. Apart from the records
from New Guinea, it is unclear to what extent E. thrax of
either subspecies thrax or subspecies hasdrubal has spread
through eastern Indonesia, either naturally, or with acci-
dental human assistance.
Erionota thrax was first reported in Mauritius in 1970
[59], although it was suspected to have been introduced in
1968 [45]. The author has examined the three Mauritius
specimens in the Natural History Museum, London
(NHM), collected by P. M. H. Davis at Black River in 1979,
and reported in Davis and Barnes [60], and finds that all
are E. torus, based on the visible external genitalia. The
original identification reported in Monty [59] was made
at the NHM, but only female specimens seem to have
been provided, and at that time the distinctive female
genitalia of E. thrax and E. torus had not been documented.
Although it is not impossible that both species were
accidentally introduced into Mauritius, in the absence
of any specimens of E. thrax, it seems safe to assume that
only E. torus is present.
Veenakumari and Mohanraj [61] documented the
first record of E. thrax thrax from the Andaman Islands,
although it would be desirable to confirm this identifica-
tion by examination of the genitalia (K. Veenakumari,
personal communication, 2014). The author was unable
to locate any specimens in the NHM to do so. Erionota
thrax was recently reported from Palpa District, central
southern Nepal [62], but this needs confirmation in
view of Smith’s [63] statement that only E. torus oc-
curs in Nepal (R. Muniappan, personal communication,
2014).
Erionota torus was more restricted in distribution, but is
now widely introduced. It is indigenous in northern India:
an early report of E. thrax as common around Dehradun
(=Dehra Dun) at the base of the Western Himalaya [64]
is considered to refer to E. torus since only specimens of
this species are present in the NHM from Dehra Dun [1].
It is also found in Nepal [63, 65] and Northeastern
India to Southern China and Peninsular Malaysia [1], but it
spread to Mauritius around 1968 (reported as E. thrax, but
see above), Southern Japan in 1971 [66], Taiwan in 1986
[67, 68], the southern Philippines probably in the early
1980s (Cebu, Dinagat, Leyte, Mindanao, Negros, Samar;
[18]), and has just been found in the Western Ghats
of India [69]. There is a single male specimen labelled
Sarawak in the NHM, but the absence of any other
records from Borneo suggests its presence there should
be considered unconfirmed. It does not seem to be pre-
sent in Java or the rest of Indonesia.
Erionota acroleuca occurs in the Andaman Islands
as the nominate subspecies. Although Swinhoe [28]
described a synonym from the Nicobar Islands, Evans [1]
treats this as an erroneous locality. Erionota acroleuca apex
is restricted to the Philippines [17, 18]. Erionota acroleuca
apicalis was formerly treated as a synonym of E. acroleuca
apex, but De Jong and Treadaway [17] established that it is
a separate subspecies found from Myanmar and Northern
Vietnam, through the Malaysian peninsula to Borneo,
Sumatra, Java and east to West Nusa Tenggara. Finally,
E. acroleuca sakita is found on Sulawesi, and the islands to
the east of Central Sulawesi [1, 43].
It likely seems that the major translocations of E. thrax
and E. torus were mediated by aircraft flights carrying
infested plants or planting materials, infested leaves or
adult butterflies. Ova occasionally occur on fruit in bun-
ches, and so may also be transported internationally [70],
but the challenge for the hatching larvae to reach Musa
spp. leaves after arrival is very considerable. Military flights
have been implicated or suggested as the route for
the spread to Mauritius and Hawaii: E. torus is ‘thought to
have been introduced in military aircraft based in Malaysia
in 1968’ [45], and E. thrax was discovered in Hawaii at
Hickam Air Force Base, Oahu [46]. The spread of E. thrax
thrax to Guam has been linked to American military bases
on Guam and the Philippines, as well as heavy civilian
traffic between the two (R. Muniappan, personal com-
munication). The fact that Guam and Saipan both had
strong military bases in the 1950s suggests how the sub-
sequent introduction to Saipan may have been facilitated,
rather than to other islands in the region. During its
outbreak phase in PNG, E. thrax spread throughout the
PNG mainland within 6 years, moving several hundred
km per year [58]. This would have been due to a mixture
http://www.cabi.org/cabreviews
6 CAB Reviews

of natural dispersal of gravid females, and accidental man-
assisted transport of planting materials, leaves and adult
butterflies.
What is less clear is the mechanism of spread between
relatively nearby islands, e.g. through the Hawaiian Islands
and from New Guinea to New Britain. Waterhouse
and Norris [45] speculate that ‘the water gaps between
Southern PNG and Northern Australia are probably well
within the flight range of adults. . . . Southwest Pacific
islands will be at risk if inter-island movement of fresh
banana leaves is not prohibited. In any case adults may be
able to fly to nearby islands and establish themselves.’ This
speculation by Waterhouse and Norris [45] has subse-
quently been presented in a pest risk assessment as
‘E. thrax has spread from Oahu to the other Hawaiian
Islands flying over water for distances of up to 150 km
[45]’ [71] – 150 km being approximately the distance
between PNG and the northern tip of Australia. No
published evidence to support the possibility of gravid
females flying these distances has been found. The fact
that E. thrax and E. torus have not spread more widely
already, e.g. within the parts of Indonesia where they
do not occur naturally, or within the Northern Mariana
Islands, suggests that such long distance flights over sea do
not occur. Similarly, the evolution of isolated subspecies
of E. thrax (above) indicates a species that is not highly
mobile.
Inter-island movement of boats seems a more likely
pathway of introduction, as Sands et al. [48] discuss:
‘Lights on board boats are considered to be a means for
attracting the crepuscular adult E. thrax and allowing them
to be carried from one island to another. A specimen in
the Natural History Museum, London, was collected
on board a ship in 1929, near Seram (less than 200 km
west of New Guinea), outside of its native range. From
the distinctive pattern of spots beneath the hind wings,
this specimen probably came from Java. A specimen from
Port Moresby, PNG in 1961 and referred to by Parsons
[57] probably travelled to PNG on board a ship since this
specimen pre-dated first establishment of E. thrax in
North-western PNG by 22 years.’
In view of the foregoing discussion and the confusion
that has arisen, it is worth recognizing the possibility that
in the last 65 years or so since Evans [1] completed his
review of the Hesperiidae of the region, E. torus could
have spread to areas where only E. thrax thrax was known
(or vica versa), and easily been overlooked. This possibi-
lity merits checking with recent material, for example
in the island of Borneo where there is already a single
record of E. torus, and Sumatra and Java which are in
relatively easy reach of Peninsular Malaysia.
Food Plants
I have reviewed much of the literature on food plants
of Erionota spp. and find considerable confusion and
indications of errors. Accordingly, I present my analysis
in some detail below. Erionota thrax thrax and E. torus are
similar in adult appearance [1, 21] and life history [6, 39],
and where their ranges overlap, have almost certainly
been confused – with each other and with other Erionota
spp., particularly E. acroleuca. Reports from localities
where only E. thrax or only E. torus occur are more likely
to be reliable, but where both occur there will be
confusion between the two. The situation is rendered
more difficult to interpret by the habit of many authors
of compiling food plant records from multiple sources
without attribution, so that it is not clear whether any
particular observation, such as a food plant record, is an
original observation or repeated from an earlier source,
which may be misidentified, represent a different sub-
species, or be from a different geographical area [72].
Food plants of E. thrax
I have found reports on the food plants and biology of
E. thrax thrax only, and none on the other subspecies of
E. thrax, although it seems likely that they will have a simi-
lar food plant range. In its indigenous range, many refer-
ences (e.g. [15, 39, 73–77]) indicate that E. thrax feeds
primarily on Musa spp. (Musaceae), especially banana
(M. � paradisiaca, = M. sapientum and M. paradisiaca) and
M. textilis (abaca or Manila hemp). Sands et al. [58] sur-
veyed E. thrax in PNG when it first spread through the
island of New Guinea. They report collecting immature
stages of E. thrax from Musa � paradisiaca, M. textilis, other
Musa spp. (including sections Australimusa and Eumusa)
and several varieties. Waterhouse and Norris [45] point
out that in its introduced range, E. thrax thrax is known
only from cultivated and wild species of banana (based on
personal communications of experienced entomologists
who worked on the introduced pest), although their
observation to this effect from Mauritius is actually based
on E. torus. However, records from other recorded food
plants, especially in the older literature, need critical
examination. It is possible that different species of Erionota
are responsible for the records on bamboo and palms.
Probably the oldest record of E. thrax as a pest
of banana (pisang, Musa paradisiaca) is that of Horsfield &
Moore [78] in A Catalogue of the Lepidopterous Insects in the
Museum of the Hon. East-India Company. As these obser-
vations were from Java, where E. torus is not reported,
they should be reliable. Soon afterwards, Distant [79] in
Rhopalocera Malayana, repeats Horsfield & Moore’s [78]
observation on the food plant, although his illustration of
the adult is E. torus according to Evans [1]. In Lepidoptera
Indica Vol. X, Swinhoe [80] gives the food plants as ‘wild or
cultivated Musa’. Although Evans [1] indicates that the
species illustrated is E. thrax, the observations could relate
to either E. thrax or E. torus or both. Duport [81] refers to
E. thrax as a banana pest in Indochina, and this could refer
to both E. thrax and E. torus. Williams [82] describes the
http://www.cabi.org/cabreviews
Matthew J. W. Cock 7

biology on banana on Luzon Island, Philippines, which
should be correctly associated with E. thrax. Beller and
Bhenchitr [83] include E. thrax thrax as a banana pest in
their list of insect pests of Thailand (Siam); this could be E.
thrax or E. torus.
There are many other food plant references compiled
in Robinson et al. [84] with sources, and in Vane-Wright
and De Jong [43] and CABI CPC [85] without sources.
These include various palms (Arecaceae), bamboos
and grasses (Poaceae), Heliconia spp. (Heliconiaceae),
Curcuma angustifolia (Zingiberaceae) and traveller’s palm
(Ravenala madagascariensis, Strelitziaceae), which are
assessed below.
Robinson et al. [84] cite Bhumannavar et al.’s [86]
Insects of Agricultural Importance in Andaman and Nicobar
Islands that E. thrax feeds in rolled leaves on the intro-
duced R. madagascariensis. Veenakumari et al. [87] made
new observations and reported the food plants of E. thrax
thrax in the Andaman Islands as only Musa�paradisiaca
and R. madagascariensis. As Veenakumari et al. [87] sepa-
rate their food plant records of E. thrax thrax from those
of E. acroleuca, and the leaf structure of R. madagascariensis
is suitable for leaf rolling as practiced by E. thrax, this
record is accepted here, even though Sands et al. [58] did
not find immature stages of E. thrax on this food plant in
PNG. Mau and Kessing [88] give Strelitzia (Strelitziaceae)
as a food plant genus in Hawaii, but this does not seem to
have been formally published. Nevertheless, it is a possible
food plant, although perhaps as a spill-over effect (below).
Similarly, Mau and Kessing [88] list the genus Heliconia
as a food plant in Hawaii. In PNG, the ornamental native
species Heliconia papuana is a widespread host for E. thrax
thrax (F. M. Dori and D. P. A. Sands, personal commu-
nication, in [47]), although the introduced Neotropical
Heliconia bihai is not, and is not in the laboratory [58].
Igarashi and Fukuda [6] list C. angustifolia (wild or Indian
arrowroot; Zingiberaceae) as a food plant. No other
source has been located for this record (which is repeated
in 43), but given that C. angustifolia has large leaves, similar
to those of Musa, Ravenala and Heliconia, it seems a pos-
sible food plant. A record of Canna (Cannaceae) as a food
plant genus in Hawaii [88] could be interpreted in this
way, but could also be due to a spill-over effect.
At this point it is worth noting that Musaceae
and Strelitziaceae are closely related families in the
order Zingiberales, while Heliconiaceae, Cannaceae and
Zingiberaceae although also placed in Zingiberales are
less closely related [89, 90]. Oligophagous species are
mostly, but not always, more likely to feed on species
closely related to their main food plant, than on those less
closely related (e.g. [91]). The remaining food plant
records are of species from other plant orders, and
correspondingly less likely.
The original records of E. thrax [thrax] feeding on
Arecaceae seem to be those of Piepers and Snellen [73]
from Java, who in addition to Musa spp., list Cocos nucifera
(coconut), Rhapis excelsa (as R. flabelliformis), Metroxylon
sagu (sago palm), Arenga pinnata (as A. saccharifera) (sugar
palm), and based on a verbal report Elaeis guineensis (West
African oil palm) as food plants. Keuchenius [92] and
Leefmans [93] also report E. thrax as a coconut pest in
Indonesia (Dutch East Indies). However, since Piepers
and Snellen [73] treat E. acroleuca as a synonym of E. thrax,
it is suggested here that the records from palms are
more likely to relate to E. acroleuca, which has since
been documented as a palm pest (below). Specimens from
P. C. T. Snellen’s collection (which includes material from
M. C. Piepers) are in the Naturalis Biodiversity Centre,
Leiden, Netherlands but they cannot be examined
at present during rearrangements of the collection
(R. De Jong, personal communication, 2014); perhaps in
the future examination of this collection will clarify some
of these records. It is furthermore suggested that Piepers
and Snellen’s records are the basis of the continuing
references to palms as food plants of E. thrax in the lit-
erature. Seitz [94] lists sugar-cane, coconut, Rhapis and
Metroxylon in addition to Musa spp.; the similarity to the
species listed in Piepers and Snellen [73] suggests that
their record of A. pinnata (as A. saccharifera) was mistaken
for sugar-cane.
P. B. Richards [95] and R. M. Richards [96] reported
H. irava Moore and E. thrax as hesperiid pests of coconut
in West Malaysia, but Corbett [74] in his Insects of Coco-
nuts in Malaya considers the two have been confused, that
H. irava is the coconut pest and E. thrax is only a banana
pest. Corbet et al. [39] in The Butterflies of the Malay
Peninsula highlight Musa spp. as the food plants of E. thrax
thrax and E. torus, point out that the two do not differ in
life history, and include coconut and sugar palm (i.e.
A. pinnata) as reported food plants.
In Les Insectes des Palmiers, Lepesme [75] considers
E. thrax to be very common on banana, but rarer on Elaeis,
Cocos, Metroxylon, Calamus, Cyrtostachys [Arecaceae],
sugar-cane and bamboo [Poaceae]’, citing De Joannis
(in [81]), Richards [95], Seitz [94], Williams [82],
Dammerman [97], Corbett [74] and Beller and Bhenchitr
[83]. As noted above, Duport [81], Williams [82] and
Beller and Bhenchitr [83] report E. thrax (and/or E. torus)
as a banana pest. Richards [95], Seitz [94] and Corbett
[74] are also discussed above. Dammerman [97] treated
E. thrax as a banana pest, also found on ‘Manila hemp,
bamboo, coconut, oil palm and other palms’ but does not
indicate his sources. None of these sources listed by
Lepesme [75] are the source of records on Calamus and
Cyrtostachys.
Hutacharern and Tubtim [98] listed E. thrax thrax from
Calamus sp. (rattan) in Thailand, and Johari and Che Aziz
[99] listed it as a pest of Calamus trachycoleus in Peninsular
Malaysia. Erionota thrax was listed as a pest of C. manan
in Peninsular Malaysia by Norani et al. [100], although
in a subsequent treatment of rattan pests in Peninsular
Malaysia, Maziah et al. [101] refer to and illustrate
the larva as E. torus, rather than E. thrax. In their survey
of hesperiids associated with C. manan, Steiner and
http://www.cabi.org/cabreviews
8 CAB Reviews

Aminuddin [102] also record E. torus, but later correct
this to E. acroleuca apicalis (as E. hiraca apicalis) [103]. This
correction was based on the advice of L. G. Kirton, who
indicates that E. acroleuca (as E. hiraca) is the Erionota sp.
that feeds on Calamus spp. and other palms. Hence, in his
recent Naturalist’s Guide, Kirton [77] refers to E. thrax
thrax feeding on bananas only.
In ‘Pests of Oil Palms in Malaysia and their Control’ Wood
[104] includes E. thrax as a minor pest, but the illustration
of the adult (plate XIVb above) is of a male E. acroleuca
apicalis with the distinctive white fore wing tips. There is a
relatively recent record of E. thrax hasdrubal as a pest of
oil palm in New Britain [105]. The adult illustrated in the
paper is not E. thrax hasdrubal, but it is not clear whether
it represents E. thrax thrax, E. torus, an individual of
E. acroleuca lacking the pale forewing apex, or some other
taxon of Erionota. Of these, only E. thrax thrax is known
from New Britain, but above we have discounted all
records of palms as food plants for this subspecies. It will
be necessary to examine and dissect voucher specimens
to confirm or clarify this record.
There are further records from Arecaceae catalogued
by Robinson et al. [84]. In their ‘List of economic pests,
host plants, parasites and predators in West Malaysia
(1920–1978)’ Yunus and Ho [106] include Caryota and
Nypa fruticans as food plants of E. thrax, but do not specify
the source for this record, and Pholboon [107] and
Hutacharern and Tubtim [98] in their compilations of
plant pests in Thailand, list Licuala grandis as a food plant of
E. thrax. These records from palms have not been tracked
back to their original source for this review, but below
we document that in West Malaysia E. acroleuca apicalis is
recorded to feed on Caryota spp. and E. sybirita on a Licuala
sp., indicating that these records are likely to be based on
misidentifications. Kitamura [108] reared a single Erionota
sp. from a leaf shelter on coconut on Panay Island,
Philippines, and illustrated the larva, pupa and shelter
(a longitudinally rolled leaflet). He identified the resultant
adult female as E. thrax. Could it have been another
species such as E. acroleuca or E. surprisa De Jong
and Treadaway? Kitamura [109] subsequently reared
E. acroleuca apex from a palm and compares the larva
with his coconut-reared specimen, so clearly considered
them different. Nevertheless, given the concerns raised
here, and the similarity of adult females, this record based
on a single female merits checking by examination of the
genitalia before accepting.
In their survey of E. thrax in New Guinea, Sands
et al. [58] specifically report that no immature stages
were found on Ensete glaucum (Musaceae), H. bihai
(Heliconiaceae), R. madagascariensis (Strelitziaceae),
N. fruticans, Metroxylon sp., Calamus spp., C. nucifera, Areca
catechu or E. guineensis (Arecaceae), even when infested
Musa spp. plants were growing close by. Furthermore,
in the laboratory, groups of ten newly-hatched of E. thrax
died without feeding when placed on fresh foliage of
H. bihai, C. nucifera and E. guineensis, whereas all larvae
placed on M. � paradisiaca (as M. sapientum) as controls,
began feeding. Furthermore, the leaf shelters that E. thrax
and E. torus make on banana (see below) involve beha-
viour patterns that would not be readily transferable to
a pinnate leaved palm. Hence, the evidence indicates
that E. thrax thrax does not use palms as food plants, and it
is anticipated that all the records from Arecaceae, should
be referred to E. acroleuca. It is unfortunate, therefore,
that the records from palms have led to E. thrax being
mentioned repeatedly as a pest of palms in the pest lit-
erature (e.g. [75, 110–115]). It is equally unfortunate that
the common name ‘palm redeye’ has been applied to this
species. The oldest use of this name appears to derive
from Evans [116, 117], and is assumed to be based on the
food plant records referred to above, ignoring the well-
documented damage to bananas. Although Brigadier
W. H. Evans was an excellent morphological taxonomist,
there is almost no reference to food plants and early
stages in any of his works, so it seems likely that he was
not that well-informed on this aspect, hence the use of
this inappropriate name. I recommend that the use of this
common name should be discontinued for all species of
Erionota to avoid future confusion.
There are also records from Poaceae. Robinson et al.
[84] attribute a record of Chrysopogon to Sevastopulo’s
[118] The Food Plants of Indian Rhopalocera, but this is
incorrect, as Sevastopulo does not list this grass.
Saccharum (sugar cane) is listed as food plant by
Sevastopulo [118] based on Seitz [94], which it was sug-
gested above is an error for sugar palm. Robinson et al.
[84] attribute a record of Saccharum to Corbet et al. [39],
but the food plant listed there is sugar palm, i.e. A. pinnata
as originally listed by Piepers and Snellen [73] and dis-
cussed above. Robinson et al. [84] also cite A Manuscript
List of Butterfly Hostplants in the Western Ghats and Southern
India by H. Gaonkar (2000), which does not seem to have
been subsequently published, that Saccharum officinarum
is a food plant of E. thrax thrax. Whether this is a repeat
of one of the earlier errors or a new record, it is anyway
unlikely to be an original record as E. thrax is not reported
from this part of India, and E. torus has only recently been
found there. It is concluded that none of the published
records of Poaceae as food plants for E. thrax should be
accepted.
Finally, Robinson et al. [84] cite Hutacharern and
Tubtim [98] that Butea monosperma (Fabaceae) is a food
plant of E. thrax thrax in Thailand, but this is an error for
the record of Calamus sp. on the same page ([98], p. 41),
and this record should not be perpetuated.
Interpreting these food plant records, it is worth
remembering two facts documented in weed biological
control. Firstly, oligophagous species are mostly, but not
always, more likely to feed on species closely related to
their main food plant, than on those less closely related
[91]. On this basis, Musaceae and Strelitziaceae are part of
one clade of Zingiberales, while Marantaceae, Cannaceae
and Heliconiaceae are in the other [89]. Families from
http://www.cabi.org/cabreviews
Matthew J. W. Cock 9

outside the order Zingiberales, i.e. Arecaceae in this case,
are much less likely to be suitable food plants. Secondly,
when a successful weed biological control agent is first re-
leased, there is often a population explosion, during which
there can be spill-over effects where feeding damage
occurs on other plants, usually because insects have
strayed or fallen from the normal food plant onto adjacent
plants. The same thing may happen with any newly
introduced species, such as E. thrax and E. torus in their
exotic range, so that food plant records during a popu-
lation explosion will include species that are not normally
suitable, on which eggs will not normally be laid, or which
will not sustain the full life cycle. Some of the observations
reported, e.g. from Hawaii [88], may be of this nature.
To summarize, it is concluded that the food plants of
E. thrax are restricted to Musa spp., perhaps some other
Musaceae and a few related, similarly broad-leaved species
in Heliconiaceae, Strelitziaceae and possibly Zingiberaceae
(all Zingiberales). No convincing food plant records have
been found of subspecies of E. thrax apart from the
nominate subspecies.
Food plants of Erionota torus
As has become evident during this analysis of E. thrax,
there has been confusion between E. thrax and E. torus
where the two co-occur in mainland South-East Asia.
Accordingly, food plant records without voucher speci-
mens can only be accepted for E. torus with full confidence
where E. thrax does not occur. It is clear that like E. thrax,
E. torus is primarily a pest of Musa spp. Only E. torus
occurs in Hong Kong and adjacent China, and here it has
been documented to feed on Musa spp. but not on
other plants since the late nineteenth century as E. thrax
[119–121], and more recently as E. torus (e.g. [9, 122]).
Similarly, only E. torus appears to be present in the
Western Himalayas [1], so Mackinnon & de Niceville’s
[64] report of E. thrax as common on cultivated plantains
around Dehradun is assumed to refer to E. torus.
Since the early stages of E. thrax and E. torus cannot
be distinguished, and they are both common pests of
Musa spp., it follows that where both species co-occur in
mainland South-East Asia and the Southern Philippines,
then any studies on the pest status, natural enemies,
control, etc. which do not address the presence of the
two species can be assumed de facto to treat either or
both. This would be the case for the detailed studies by
J. Okolle and colleagues in Peninsular Malaysia on the
population dynamics, within-field and within-plant dis-
tribution of the banana skipper and its parasitoids
[70, 123–129].
There are a few published records of palms as food
plants for E. torus. Robinson et al. [84] list Caryota
and Roystonea regia. As noted above, in their treatment of
pests of rattan palm in Peninsular Malaysia, Maziah et al.
[101] include E. torus, although Steiner [130] and Steiner
and Aminuddin [103] subsequently indicate that this
record should be treated as E. acroleuca (=E. hiraca).In a laboratory study in Taiwan, Tsai et al. [131] fed
cohorts of each of the five instars of larvae of E. torus with
cut portions of leaves of banana, Strelitzia reginae (bird
of paradise flower; Strelitziaceae), C. nucifera (coconut;
Arecaceae), areca nut (A. catechu; Arecaceae), Bambusa
oldhamii (bamboo; Poaceae) and sugar cane (Saccharum
officinarum; Poaceae). Feeding was most extensive on
banana, but almost as heavy on Strelitzia reginae; there was
very little feeding on coconut (none for the cohort of
instar 1), less on areca nut and none on bamboo or sugar
cane. Average survival for all instar cohorts was 50% on S.
reginae, 5% on coconut and 0% on bamboo and sugar
cane. This is a measure of the survival of larvae trans-
ferred to a plant other than banana, and does not reflect
what would happen following oviposition on that plant in
the field. If there was no feeding in instar 1 on coconut, all
larvae would die in the field, while the data does not allow
us to interpret what total mortality would be on S. regi-
nae, it would be more than 50%. In this experiment, the
larvae on banana are reported to have all completed
development, but in a parallel experiment, only 34 and
21% of individuals completed their metamorphosis on
banana at 20 and 30 �C, respectively, most mortality
occurring in instars 1 and 2. It is difficult to know how to
interpret these two apparently conflicting results. How-
ever, we can conclude that sugar cane, bamboo, areca nut
and coconut are unsuitable food plants, whereas S. reginae
is less suitable than banana.
There are internet records of E. torus using Canna sp(p).
(Cannaceae) as food plants (e.g. [132]), but these have not
been traced to any formal publication, and may well
be derived from the records for E. thrax, when the name
E. torus was misapplied to that species.
As noted above and discussed below, the early stages
and leaf shelters of E. thrax and E. torus are very similar.
The shelters can only be built using a leaf with a large flat
lamina, e.g. Musa spp. Heliconia spp., some Strelitziaceae,
Marantaceae, Zingiberaceae, etc. (all in the Zingiberales),
and unlikely to be used on pinnate palms. It is concluded
that all food plant records of E. torus from palms should
be disregarded, and although the possibility of other
Zingiberales being used as food plants cannot be discoun-
ted, none should be considered confirmed at this time.
Food plants of E. acroleuca
Erionota acroleuca acroleuca has been recorded using
palms as food plants in the Andaman Islands: C. nucifera,
Pinanga kuhlii, Calamus sp. [87]. Erionota acroleuca apicalis
from South-East Asia also feeds on palms. In Peninsular
Malaysia, it is recorded from ‘oil palm’ ([104] as E. thrax,
[76]), Caryota sp. and Orania sylvicola (=macrocladus) [106],
Calamus manan, C. ornatus, C. scipionum, Caryota mitis,
Areca triandra and Adonidia merrillii [130, 133]. Recently,
http://www.cabi.org/cabreviews
10 CAB Reviews

Xue and Lo [19] report rearing this subspecies from
Caryota maxima (as C. ochlandra) in Yunnan Province
and Arenga westerhoutii in Guangxi Province, China, and
an unidentified palm in Singapore; they conclude that
E. acroleuca is primarily a palm feeder. Kitamura [109]
reports Saribus rotundifolia (=Livistona rotundifolia) to be a
food plant of E. acroleuca apex on Samar Island, Philippines.
L. G. Kirton (in 103, 130) has suggested that all records
of Erionota spp. from rattan palms (Calamus spp.) should
be treated as E. acroleuca, and here this generalization is
extended to all records from palms (although some of
these could be errors for H. irava). In particular, the
records of E. thrax from C. nucifera, R. excelsa, M. sagu,
A. pinnata and E. guineensis in Java [73] and the various
repetitions of these in the literature (see above) are
considered to be in error for E. acroleuca. L. G. Kirton
(personal communication, 2014) transferred larvae of
E. acroleuca apicalis of various sizes from palms to Musa sp.
and they all died. The record of E. thrax from oil palm in
New Britain, PNG [105] needs clarification by examina-
tion of voucher specimens; it might prove to be E. acro-
leuca. In conclusion, E. acroleuca is the Erionota species that
is commonly found on a wide range of palms, and until
such time as there are authoritative records of E. thrax
or E. torus breeding on palms, all such existing records
should be considered misidentifications, most probably
for E. acroleuca.
Food plants of other Erionota spp.
The Chinese species, E. grandis feeds on palms [6], as does
E. sybirita (Hewitson). The former is a gregarious species
and so unlikely to be mistaken for other Erionota spp.
Steiner [130, 133] records the later feeding on Licuala
kunstleri in Peninsular Malaysia. It is a rare species found
from Southern Myanmar and Thailand to Borneo and
Palawan [1, 18], so is unlikely to account for many of the
records of other Erionota spp. from palms. The food plants
of the remaining Erionota spp. are unknown.
Immature Stages
Erionota thrax, E. torus and E. acroleuca are similar in adult
appearance [1, 21]. The life history and food plants of
E. thrax and E. torus are very similar [6, 39]. However,
while the early stages of E. acroleuca are also similar, the
food plants and shelters are not. Hence, at this time, the
early stages cannot be reliably separated by diagnostic
characters, and E. acroleuca can only be provisionally
separated by its food plants and shelters.
The larvae of E. thrax and E. torus make their leaf
shelters by cutting and rolling the lamina of their food
plant leaves. More detailed observations of shelter for-
mation are needed for both species, with information on
individual variation, feeding habits, etc. At present, the
shelter making behaviour cannot be used to separate
E. thrax and E. torus. In contrast, although hardly docu-
mented, the leaf shelter of E. acroleuca on palm leaves may
be constructed similarly to those of E. thrax and E. torus,
but is also made by longitudinally rolling a leaflet or pulling
together two leaflets or parts of a larger leaf. This latter
type of shelter seems typical for palm-feeding Hesperiinae
[7], and contrasts with the shelters of E. thrax and E. torus
where the leaf is rolled transversely.
Life history of E. thrax
Horsfield and Moore [78] provide the first illustrations of
the larva and pupa. As these observations were from Java,
where E. torus is not reported, they should be reliably
associated with E. thrax. The figure portrays the larva as
white, with long erect setae on the body, and a dark
brown-black head. The pupa is incorrectly shown, in that
it has a short, strong, pointed, slightly upturned frontal
spike. Horsfield’s emerged pupa is preserved in the NHM,
and is accurately drawn, so is here assumed to belong to
some other species. Distant [79] reproduces these
figures of the larva and pupa in Rhopalocera Malayana,
perpetuating this error regarding the pupa. Semper [24]
illustrates the larva, correct pupa and a large leaf shelter
implicitly from Manila, Luzon Is, Philippines, in which case
they are safe to associate with E. thrax.
Piepers and Snellen [73] document the biology: the
larva ‘lives in a part of a leaf that is rolled up into a case
and fastened with threads, it is chalk-white with a black
head overgrown with very short hairs and wholly covered
with a white waxy powder, that also entirely fills its
abode’. Although Piepers and Snellen treat E. acroleuca as
a synonym of E. thrax, it seems likely given the description
of the shelter on Musa in Java that they are dealing with
the immature stages of E. thrax. Their painting of the pupa
also appears to represent this species, having no frontal
spike, and a long proboscis extending beyond the cre-
master.
In Lepidoptera Indica Vol. X, Swinhoe [80] gives the food
plants as ‘wild or cultivated Musa’ and states that the larva
‘lives in a shelter made of a portion of rolled-up leaf. To
make this shelter it has to cut into the edge of the
enormous leaves of the wild or cultivated Musa, or
plantains, to obtain a suitable segment to be rolled up.’
The larva is figured from ‘Grote’s original drawing’.
Arthur Grote prepared the first catalogue of the butterfly
collection of the Asiatic Museum in 1857–1859; he also
took notes on the life histories, making use of a Munshi
Zynulabdin, a local artist [134], so the material for these
figures was likely to have been collected in the vicinity
of Kolkata (Calcutta), and could represent either E. thrax
or E. torus. Although not acknowledged, the figure of a
pupa is copied from [78] and so is incorrect. Evans [1]
indicates that the adult illustrated by Swinhoe [80] is
E. thrax; however, the observations on early stages and
http://www.cabi.org/cabreviews
Matthew J. W. Cock 11

the figure of the larva could refer to either or both
E. thrax or E. torus.
Williams [82] describes the biology on banana on Luzon
Island, Philippines, which should be correctly associated
with E. thrax. ‘The eggs are hemispherical and well ribbed
from the apex and are not quite 2 mm in diameter. The
little larvae cut obliquely inward from the edge of the
banana leaf and form a tube by fastening this rolled strip
with silk; a large larva makes a large roll as illustrated.
Here it also pupates. A full-fed larva is fusiform-cylindrical
with a large head set on a small neck, as is usual among the
Hesperiidae, and is covered with a whitish flocculent
substance.’ Williams [82] includes drawings of a banana
leaf with many small shelters, a leaf with a single large
shelter, and a parasitized pupa. The figure of the large
shelter is not entirely clear, nor is it explained in the text.
It can be interpreted as indicating that the larva makes
two parallel cuts along the leaf and rolls the resultant flap,
leaving the leaf edge intact. An alternative explanation is
that the area of leaf adjacent to the edge of the lamina
reflects the portion of the leaf that is eaten or not eaten
by the larva.
The following account is synthesized from several
sources. Ashari and Eveleens [135] provide drawings of
the life cycle and provide some additional information.
More recent observations are similar, but some have
been made from areas where E. thrax and E. torus occur
together and so cannot be allocated to species with
confidence (e.g. [16, 70]). Igarashi and Fukuda [6] include
the life history of E. thrax from India (in their text they
only mention Sikkim). As noted above, Igarashi and
Fukuda [6] refer to the species that has spread to the
Pacific and Asia as E. torus, rather than E. thrax. As they do
not refer to the diagnostic differences in the genitalia, one
cannot be sure that their identifications were thus con-
firmed. However, although both E. torus and E. thrax occur
in Sikkim, India, and the possibility of confusion cannot
be ignored, the adults that they illustrate are correctly
associated (the difference in male forewing shape is very
clear), so it is assumed that the early stages are also
correctly associated. Although there are photographs and
brief descriptions of the biology of E. thrax in its intro-
duced range [45, 136] no detailed accounts have been
located in this review.
Eggs are laid singly or in groups, mostly on the under-
side of the leaves, but also on the upper side. Eggs are
2.3 mm wide�1.1 mm high, with about 22 fine ribs stop-
ping short of the micropyle.
Detailed descriptions of the leaf shelter building beha-
viour are not available. The newly hatched larvae form a
little leaf roll from the edge of the leaf, inside which they
feed [135]; it is not clear whether the first shelters
are simple leaf folds from the leaf edge, or the transverse
leaf roll observed later – or both. Igarashi and Fukuda
[6] note that ‘the larva rolls a part of a food plant leaf
inwards . . . to make a nest in which to rest with its head
directed upwards. . . . The larva eats the wall of the nest
from below . . . Feeding occurs only during the night
�time. The bottom of the nest is usually closed, and often
filled with a large amount of frass pellets. . . . the larva
moves and nests are renewed several times. Moving takes
place in the direction towards the basal part of the leaf. It
takes a few days to make a new nest. Some larvae,
however, may show behaviour patterns exceptional to
the habits described above.’ Leaf rolls made by the mature
larvae extend all the way from the edge to the leaf midrib,
which leads to considerable loss of leaf area (Figure 4.2).
The leaf rolls in Figure 4.1 appear to have five rolls of leaf,
and dangling from the bottom is a narrow strip of leaf
margin which has not been eaten. This can be interpreted
as the larva feeding on the leaf edge within its shelter, but
not the section furthest from the midrib, which would
form the distal end of the leaf roll, where a feeding larva
might be vulnerable to predators or disturbance. This may
be the reason for the two lines in Williams’ [82] drawing
mentioned above.
There are five larval instars and the final instar grows to
50 mm or more. All instars have dark brown to black
heads and pale bodies. After the second instar there is an
increasingly dense cover of white powder on the body.
The body has fairly inconspicuous pale, erect setae; the
head is dark brown, rounded, indent at vertex, widest
near base, with setae similar to those on the body; the
pronotum, spiracles, true legs and prolegs are all pale,
Figure 4 Shelters of E. thrax thrax and E. torus on Musasp. 1, E. thrax, Maui, Hawaii, 13 Aug 2002 (
gForest and
Kim Starr); 2, E. thrax outbreak, Fly River, Papua NewGuinea (
gC. Dewhurst); 3, E. torus, Serdang, Malaysia, 6
Feb 2003, M. J. W. Cock. Differences observed betweenindividual shelters are not considered to be diagnostic, butto reflect individual differences.
http://www.cabi.org/cabreviews
12 CAB Reviews

more or less concolorous with the body. It can be seen
that there is minimal white powder on the head, thorax
and ventral part of the abdomen of the individual shown in
Figure 5.1, but the covering becomes more extensive as
the larva becomes fully grown. The pupa (Figure 6.1) is
pale white-brown, nearly cylindrical but deeper and wider
at the thorax; no frontal projection and the proboscis
sheath projects beyond the cremaster. The pupa and the
inside of the pupal shelter are lined with white waxy
powder.
Life history of E. torus
Walker [119] wrote about the butterflies of Hong Kong,
and was the first to comment on the leaf-roll shelters,
stating that E. thrax (i.e. E. torus) has been reared ‘in 1892
from larvae found in rolled-up leaves of banana’. Mack-
innon and de Niceville [64] include a brief description of
the larva in northern India, and mention that pupation is in
a portion of a rolled-up leaf. In the Palaearctic butterflies
section of the Macrolepidoptera of the World, Seitz [137]
wrote ‘The larvae of Erionota . . . feed on the gigantic
leaves of Musa, which are yards long. In forming their
shelter they commence by making two incisions from the
edge of the leaf towards the midrib at a distance of about
8–10 cm one from the other, and then proceed to roll this
piece of the leaf up like a cigar, the larva living in the
cylindrical hollow within this roll.’ When dealing with the
Indo-Australian butterflies, Seitz [94] expands on this ‘The
larva is snow-white, covered with a very short wax-like
pubescence, and with a black head. It is easily discovered,
as it gnaws out of the gigantic banana leaves, by two
parallel-cuts a longitudinal piece which it rolls up like a
cigar, in the centre of which it lives. These characteristic
cuts you can see in the Musa-plantations from a distance
of 20 or more paces, so that the larvae, and still more
easily the pupae, may be collected in numbers ad lib. The
pupa is of a dingy pale yellow with a very long case of the
proboscis projecting beyond the end of the body like a
spear . . .’. These seem to be Seitz’ own observations as
he refers to collecting many specimens in Hong Kong, i.e.
they are E. torus not E. thrax. The reference to two cuts to
make the shelter are not clear, but may parallel those
shown by Williams [82] for E. thrax, and perhaps explained
by the feeding behaviour of the larvae inside the shelters
as extrapolated above for E. thrax based on Figure 4.1, i.e.
the cut nearer the midrib is used to make the shelter, but
the second ‘cut’ is actually where the larva stops feeding
within the shelter in order not to expose itself at the
bottom.
Based on his observations at Hong Kong and Canton
(Guangdong, China), Hoffmann [121] provided a quite
detailed description of the early stages and their biology,
together with drawings of the ovum, larva and pupa, and
photographs of adults and shelters. Eggs are laid singly or
in irregular groups on the upper and lower surfaces of
leaves (more frequently the latter). They measured 2 mm
in diameter and 1.13 mm in height, with the flat area on
top 0.45 mm wide; there are 24–29 fine ribs. They are
usually orange in colour, but there is considerable varia-
tion over time and within batches. There are five larval
Figure 5 Final instar larva of E. thrax thrax and E. torus..1, E. thrax, Maui, Hawaii (g Forest and Kim Starr); 2, E. torus,Mauritius (g T. C. E. Congdon). The apparent difference in head colour is not considered diagnostic.
Figure 6 Pupa of E. thrax thrax and E. torus on banana, lateral view. 1, E. thrax, Papua New Guinea (g C. Dewhurst);2, E. torus, Mauritius (g T. C. E. Congdon).
http://www.cabi.org/cabreviews
Matthew J. W. Cock 13

instars. The first instar is 3 mm long when recently hat-
ched, pale green with the head and pronotum black, and
no wax powder. The width of the head capsules of the five
instars were 1.08, 1.45, 2.08, 2.9 and 4.26 mm. After the
first moult, the larva is covered with an increasingly
dense layer of white powder. The fifth instar measures
about 44 mm long; the surface of the blackish-brown head
is ‘more or less granulose’, with a moderate covering of
weak, silky setae, thicker on the front. The pupa measures
40–46 mm, and is about 7 mm wide and deep at the
thorax and is covered with white powder. It is pale dirty
yellow apart from the dark brown cremaster; the pro-
boscis extends 5 mm beyond the cremaster; spiracles
brown; the eyes become reddish about halfway through
the development period.
After the early reports considered above, there
seems to be little in the way of original observations, as
opposed to repeating information from the literature,
until late last century, when accounts illustrated with
photographs started to appear. There is a brief summary
from Mauritius in Monty [59] as E. thrax. Bascombe et al.
[9] describe and illustrate the early stages of from Hong
Kong, and Igarashi and Fukuda [6] include the life history
from Japan. Additional information is provided from the
observations of T. C. E. Congdon (personal communica-
tion, 2014) and the author’s unpublished observations.
The following is largely based on Bascombe et al.’s [9]
account, except as indicated. The egg ([9], Plate 103.1–2)
is 1.8–2.2 mm in diameter, dome-shaped with 22–24 fine
ribs stopping short of the micropyle; they are pink or
variegated pink and yellowish-white when laid, becoming
paler as they develop. They are usually laid on the leaf
underside, and may be laid singly, but more often in
clusters of up to 30, rarely 50. In contrast, Igarashi and
Fukuda [6] state that the ova are 2.2 mm in diameter, and
normally laid singly on the leaf upper side, although bat-
ches of 3–36 are recorded; this may reflect a gen-
eralization on a relatively small number of observations.
On hatching the larva eats the egg shell, and then moves
to the edge of the leaf to make its first shelter, which is
either a simple fold parallel and close to the edge of the
leaf ([9], Plate 103.3), or a cone shaped roll formed by
making a curved cut from the edge of the leaf and rolling
the resultant flap under. This first cut may be curved
towards the leaf base or away from it, but all subsequent
shelters are made with the cut curved towards the base of
the leaf. Larvae feed on the inner parts of the roll, which is
closed at the inner end (nearest midrib) and open at
the distal end, where frass may accumulate. As the larva
grows, it extends the cut and rolls more of the leaf into
the shelter. Additional shelters are constructed if the
shelter is damaged, unable to roll further or encounters
another shelter.
Igarashi and Fukuda [6] indicate that the leaf shelters of
E. torus and E. thrax are rolled in opposite directions,
although it is not clear from the text and diagrams whe-
ther this means clockwise and anticlockwise when seen
from above, or one with the leaf under surface on the
inside of the shelter and the other with the leaf under
surface on the outside of the shelter. In any case, they
seem to be incorrect, as both species roll the leaf flap
clockwise or anticlockwise depending on which side of
the leaf the shelter is formed, and the shelters normally
have the leaf underside inside the shelter for both species.
Igarashi and Fukuda [6] also indicate that the top end of
the larval leaf shelter is open and that the larva comes out
of this opening to feed on the leaf by night; this also seems
to be incorrect as photographs of the leaf shelters indicate
no feeding outside the shelters [9, 66, 121, 138], and the
indications are that the larvae feed inside the tube, as
discussed below. Working in Japan, Makibayashi [139]
provides a detailed description with clear diagrams to
show how a final instar caterpillar makes a new leaf
shelter. His diagrams show that the leaf roll is pulled into
position by silk strands between the leaf roll and the
adjacent unaffected part of the leaf, which is in due course,
is pulled into the shelter in turn.
The following notes on E. torus in Mauritius were pre-
pared by T. C. E. Congdon (personal communication,
2014). The larva makes a cut in the leaf, and rolls the cut
portion, extending the cut across the leaf as it grows. No
evidence of eating can be seen outside the shelter, and
when the tube is unrolled, the unrolled leaf does not refill
the space it originally occupied. It appears that the larva
feeds by eating the inner layers of the tube, thus avoiding
the dangers inherent in emerging from its shelter. One
effect of this is that the inside diameter of the tube
becomes much greater than is usual in Hesperiinae tube
shelters. The bottom end of the shelter is loosely sealed
with silk, retaining the frass, thus hindering access by
predators. The top of the shelter is crudely closed with
silk, but must be opened when the larva extends the cut,
and rolls more of the leaf. Old strands of silk are cut and
remain visible along the cut on the section not incorpo-
rated into the shelter. This detail is at variance with the
description and diagrams of Makibayashi [139] above, and
the author’s unpublished observations from western
Malaysia that the silk strands used to form the shelter
are attached to parts of the leaf that are subsequently
incorporated into the shelter. Further observations are
needed to clarify whether these are local or individual
variation in behaviour. The larva pupates within the
shelter, head up and without a girdle.
From an early age, the larva is covered with an
increasingly thick layer of white powder, so that the body
appears white with erect pale setae partially covered with
the powder, and the uniformly dark head is partially
obscured with the white powder (Figure 5.2). The larva
grows to 55 mm in length [6] and is covered with white
flocculence as it prepares for pupation.
Pupation takes place in the final leaf roll shelter which is
15–20 mm long, lined with a thin layer of silk and closed
at the distal end by a thin mesh, which is easily broken by
the emerging adult. Ito and Nakamori [140] refer to the
http://www.cabi.org/cabreviews
14 CAB Reviews

pupal shelters in Japan as ‘trumpet shaped’, 1.5–2.0 cm
diameter at the smaller [basal] opening, and 3.0–3.5 cm at
the terminal opening. This does not seem to be typical.
Igarashi and Fukuda [6] note that the pupal shelter is
simpler than the larval shelter, having 3–4 layers rather
than 5–6, and the pupa is formed with the head upwards,
although the adult butterfly emerges from the bottom of
the shelter.
The pupa (Figure 6.2) is 45 mm long, pale brown with a
variable and patchy covering of white powder. The pro-
boscis extends almost to the cremaster. It is attached at
the cremaster, and unlike many Hesperiinae does not
construct a silk girdle.
Life history of E. acroleuca
Wood [104] treats E. acroleuca apicalis as E. thrax
and includes a photograph of a hesperiid larva and
shelter (plate XIVa) without explicitly stating whether it
represents E. thrax or Cephrenes chrysozona (Plotz), the
two named species of Hesperiidae treated. The angle of
view does not permit a definitive identification to be
made, but it is more likely to be E. acroleuca than
C. chrysozona. The colouring of the larva suggests an
Erionota sp., rather than a Cephrenes sp., which has a green
body and white head with dark lines [9, 39]. The leaf
shelter appears to have been formed by making a diagonal
fold across the oil palm leaflet, but since this may have
been constructed in captivity, it is not necessarily indica-
tive of what happens in the field. Mature C. chrysozona
larvae roll palm leaflets parallel to the mid rib (author’s
unpublished observations).
Steiner [133] records observations on the ova of
E. acroleuca apicalis (as E. hiraca). The ova are white, with
a smooth surface and hemispherical in shape. They can
be found on all leaves of C. manan, but the majority are
found on the youngest leaf, and although found on both
surfaces of the leaf, they are mostly found on the upper
surface. Xue and Lo [19] indicate that the ova of
E. acroleuca apicalis are laid in a cluster on the frond
underside.
Maziah et al. [101] treat E. acroleuca apicalis as E. torus
in their illustrated account of the pests of rattan, and
illustrate the larva with a photograph. They note that the
larva ‘rolls up the leaf from the tip parallel to the veins and
lives and feeds from one end of the leaf in this shelter’.
This wording is not completely clear, but in view of sub-
sequent observations, it is clear that the leaflet is rolled
transversely (more or less at right angles to the mid rib),
unlike the longitudinally rolled shelter (parallel to the
midrib), made by most palm-feeding Hesperiinae [7]. They
describe the larva as ‘pale green covered with short silky
hairs and a whitish powdery substance and has a dark
brown head capsule’. Their photograph shows a larva
similar to those of E. thrax and E. torus. Steiner’s [133]
description and photograph of a 34 mm larva match
earlier observations. To make the larval leaf shelter, larvae
cut the leaflets near the base about two-thirds of the
distance to the midrib, and wrap the distal part in several
turns to form a roll (Steiner refers to this as a longitudinal
roll, but I would interpret this term as parallel to the mid-
rib, which is not the case here). The larva rests and feeds
inside the leaf roll. Prior to pupation, the larva creates a
similar kind of leaf roll, which is heavily lined with wax.
L. G. Kirton (personal communication, 2014) confirms
these observations, and has also observed that on Caryota
sp(p)., which have laminate fishtail pinnae, the larva makes
a shelter in a similar manner to those of E. thrax and
E. torus on Musa spp., i.e. by cutting an arc from the edge
of the lamina and rolling the resultant flap into a tube, in
which the larva hides and feeds. Xue and Lo [19] provide
some additional information regarding E. acroleuca apicalis.
Larvae, hatching from egg clusters develop in separate
pinnae on the same rachis, rolling the pinnae into a ‘cone-
shape shelter’. They illustrate in colour a larva similar to
those above, with a dark brown head with short setae,
conspicuous long setae on the body, and the head and
body covered with white waxy powder.
Maziah et al. [101] record that pupation is in the final
larval shelter and ‘the pupa is long, slender and also
covered with the same whitish powdery substance pre-
sent on the larva. When disturbed the pupa wriggles
violently in the shelter’. Xue and Lo [19] indicate that the
pupa is 33 mm, pale cream in colour with brown spiracles,
the proboscis extending beyond the cremaster. It is
‘sealed in a cocoon’ in a shelter formed by ‘rolling the
undersurface of the apex half of the rachis [pinna?] lat-
erally’. This description of the shelter is not entirely clear,
and although their illustrations of the shelter and pupa
show a silk cocoon within a leaf shelter, the structure of
the shelter cannot be interpreted, although it appears to
be aligned longitidinally with the pinna veins.
Finally, Kitamura ([109], Figure 18, upper photo)
includes a photograph of the final instar of E. acroleuca
apex in his report on this butterfly feeding on L. rotundifolia
on Samar Island, Philippines. The larva is similar to those
reported above (and to other Erionota spp.), but the leaf
shelters are not documented.
The shelter building behaviour of the larvae seems
to vary with the leaf structure of the food plant palm.
Further observations from different food plants and from
different parts of the range of this species and its sub-
species, supported with voucher specimens, are needed
to clarify the biology of this species.
Life history of E. sybirita
There are also some details on the life history of E. sybirita
on L. kunstleri in Steiner’s [133] thesis. The eggs are white
to grey, hemispherical with a smooth surface; almost
all were found on the underside of the leaflets. The
larvae have a whitish green body, but unlike the other
http://www.cabi.org/cabreviews
Matthew J. W. Cock 15

Erionota spp. treated above, no wax layer. The head
capsule is dark brown to black. The pupae are brownish,
covered with wax, and about 36 mm long. The very long
proboscis sheath extends beyond the abdomen by up to
40 mm. The pupae are attached only by the cremaster and
do not have a silk girdle. Like the larvae of E. hiraca, the
larvae of E. sybirita cut the leaflets near the base and wrap
the distal part into a roll with several turns (again referred
to as longitudinal, but this term should be avoided for the
leaf rolls of Erionota spp.). The larva stays in the roll and
eats it from the inside. Prior to pupation the lower end of
the tube is sealed with silk. It appears that the absence of a
covering of white waxy powder on the larva (and perhaps
also on the pupa) as well as the specific host plant should
facilitate separation of the early stages from E. acroleuca.
Natural Enemies of E. thrax and E. torus
Waterhouse and Norris [45] summarized the available
information on the natural enemies of E. thrax, many only
identified to genus or family, but as has become apparent
above this summary would not have separated natural
enemies of E. thrax from those of E. torus where they
occur together. This table is updated as three separate
tables below, broken up into records that can be un-
equivocally associated with E. thrax (Table 2), E. torus
(Table 3), or because of their location could be associated
with either species (Table 4). Summaries of parasitoid
records in Gold et al. [171], Tinzaara and Gold [172],
and Muniappan et al. [115] are not repeated here. Key
information on what appears to be the most common
and important parasitoids (all Hymenoptera) is sum-
marized in the following paragraphs.
Brachymeria spp. (Chalcididae)
Several Brachymeria spp. that attack pupae of Erionota spp.
appear to be polyphagous species, including B. lasus
(Walker), B. albotibialis (Ashmead), B. euploeae (West-
wood) and B. thracis (Crawford). Okolle et al. [127] found
that B. albotibialis also attacks prepupae. Where recorded,
they are gregarious; Hasyim et al. [147, 148] record an
average of nine parasitoids per host, and Okolle et al.
[127] report seven per host for B. albotibialis. The incon-
sistency of names used in different publications, even from
the same area, suggests that at least some material has
been misidentified in the past.
Ooencyrtus pallidipes (Ashmead) (Encyrtidae)
This species was better known as O. erionotae Ferriere,
which was described from Peninsular Malaysia from ‘eggs
of E. thrax L. and of an unknown Lepidopteron on wild
plantain leaf’ [169]. Given the locality, this could have
been from either E. thrax or E. torus. Huang and Noyes
[150] point out that O. pallidipes is the senior synonym.
Ashmead [173] described O. pallidipes from the Philippines
from ‘two specimens bred by Father W. A. Stanton from
an undetermined aphid’. This now seems to be a host
error, perhaps because a mummified aphid may be similar
in size, shape and colour to a parasitized ovum of E. thrax.
Ooencyrtus pallidipes is a common and widespread egg
parasitoid of E. thrax (Table 2) that also parasitizes ova of
E. torus (Table 4). It is reported parasitizing E. thrax (or
E. torus) from Nepal, Thailand, Malaysia, Sumatra, Java,
PNG and Guam, although the last two may represent
accidental introductions. It has been introduced as a
biological control agent in Mauritius and Hawaii for the
control of E. thrax (although the pest in Mauritius is now
known to be E. torus). Females are reported to prefer
younger ova for oviposition [45] and once the larvae start
to form within the ovum, they are not parasitized [127].
Females avoid parasitized ova for oviposition [127].
In Sumatra, it is reported to cause 10–27% mortality
[148, 151] and Hasyim et al. [147] reared an average of
3.8 adults per parasitized egg. It has been recorded from
several Lepidoptera, including Eupterotidae, Gracillariidae,
Nymphalidae, Papilionidae and Pieridae [141], but the ova
of Gracillariidae are most probably too small to support
this species. A record from ova of a buprestid represents
a misidentification [150].
Pediobius erionotae Kerrich (Eulophidae)
This species was described from specimens reared from
ova of E. thrax (or E. torus) in peninsular Malaysia, and the
type series includes material from Java, and R. A. Syed’s
material from Sabah, Malaysia reared from ova, larvae
and cocoons of Cotesia erionotae (Wilkinson) [149, 155].
Kerrich [155] does not seem entirely convinced by the
records from E. thrax larvae and even less so by the
record from a C. erionotae cocoon. This is another com-
mon egg parasitoid of E. thrax (or E. torus), which has
also been recorded Sumatra, where it causes 16–39% egg
mortality [148, 151].
Cotesia erionotae (Wilkinson) (Braconidae)
This gregarious larval parasitoid was described from
E. thrax (or E. torus) in West Malaysia [157], and is known
to attack one or both Erionota spp. in Thailand, West
and East Malaysia, Sumatra and Java (Table 3); it attacks
E. thrax in Sabah, Malaysia (Table 2) and E. torus in
Mauritius (Table 4). It was originally described in the
genus Apanteles, but Austin and Dangerfield [174] trans-
ferred it to Cotesia. Larvae of instars 2–4 are parasitized
and the mature parasitoid larvae emerge mainly from
instar 5 [127]. Hasyim et al. [147, 148] record an average
of 58 adults per parasitization, although cocoons are often
hyperparasitized, while Okolle et al. [127] record an
http://www.cabi.org/cabreviews
16 CAB Reviews

average of 87 per larva. This species has been released
and established for biological control of E. torus in Maur-
itius and Taiwan and E. thrax in Hawaii, Guam, Saipan and
PNG. It was initially thought to be already present in
Taiwan, attacking Udaspes folus (Cramer) (Hesperiidae,
Hesperiinae, incertae sedis) which feeds on gingers
(Zingiberaceae), but closer examination showed differ-
ences in microsculpture and colouring [175], suggesting
this is a different species. In light of this, a recent record
of C. erionotae parasitizing U. folus in India [176, 177] may
need verification.
Casinaria sp. (Ichneumonidae)
This is a common larval parasitoid of E. thrax in Sumatra
and Java. In Java, 10–80% larval parasitism is reported, and
it is commonly hyperparasitized by Brachymeria lasus, and
rarely by Eurytoma sp. B and Theronia zebra [147]. It is
possible that the records of a Scenocharops sp. a common
larval parasitoid of E. thrax in Sabah [149] represent the
same species as the two genera are similar and closely
related in the tribe Campoplegini.
Xanthopimpla gampsura Krieger (Ichneumonidae)
This is a common pupal parasitoid of E. thrax in Sumatra
and Java. It was described from Borneo and Sumatra
[178], and has also been recorded as a parasitoid of the
palm-feeding hesperiids H. irava and Cephrenes chrysozona.
Earlier records from Java of Xanthopimpla sp. are referable
to this species, but it is not clear whether the record
of X. regina from Sabah represents a different species or a
misidentification.
Biological Control of E. thrax and E. torus
There have been several programmes of biological control
against E. thrax (Guam, Saipan, Hawaii, PNG) and E. torus
(Mauritius, Taiwan). Waterhouse and Norris [45] provide
a review of the earlier programmes against E. torus
and E. thrax (summarized below and in Table 5). There
are numerous natural enemies in the indigenous range of
E. thrax (previous section, Tables 2–4), and several have
been used as biological control agents (Table 5).
Erionota torus was first reported in Mauritius in 1970 as
E. thrax [59], although it was suspected to have been
introduced with military air traffic from Malaysia in 1968
[45]. Once established in Mauritius, it soon became a
serious pest on tall banana varieties in urban areas, but
not in rural commercial plantations of dwarf Cavendish-
type bananas [45, 59]. R. A. Syed of the Commonwealth
Institute of Biological Control (part of CABI) studied the
parasitoids of E. thrax in Sabah [188] and sent shipments
of larval parasitoids Scenocharops sp. and Cotesia erionotae,
and egg parasitoids Ooencyrtus pallidipes and Agiommatus
sp. nr. sumatraensis to Mauritius in 1971–1973. All four
species were released in Mauritius. Cotesia erionotae
and O. pallidipes became established, and although there
were recoveries of Agiommatus sp. it did not persist. The
parasitoids quickly provided effective biological control.
In 1975 a cyclone severely damaged banana plants and
drastically reduced banana skipper and parasitoid popu-
lations which, nevertheless, built up again in 1976. None
of the parasites established earlier was recovered in the
two years following the cyclone, but damage to banana by
the skipper was recorded as being very low in 1978 and it
remained uncommon. Davis and Barnes [60] report that
specimens of E. torus (as E. thrax) were taken on the west
coast of Mauritius in 1979, but it was not seen elsewhere
between the years 1976 and 1980, so that its status
was considered unclear. Williams [189] considered it (as
E. thrax) ‘uncommon for reasons that are not clear’.
Collectors from the African Butterfly Research Institute
found early stages in 2014 (T. C. E. Congdon, personal
communication, 2014), so E. torus continues as an
uncommon species of no significant pest status in
Mauritius, almost certainly kept under effective biological
control by its introduced parasitoids. None of the other
species of Hesperiidae found on Mauritius are known
to be parasitized by any of the parasitoid species intro-
duced against E. torus (J. Monty, personal communication,
1987 in 45).
Based on the programme already carried out
for Mauritius (purportedly on E. thrax but actually on
E. torus) when E. thrax appeared in Hawaii, a biological
control programme was immediately started [45]. In
1973, O. pallidipes was introduced from Guam where
it seemed to be an accidental introduction itself, and in
1974 C. erionotae was introduced and released, initially
from Thailand (early 1974) and then from Sabah (late
1974). Based on the information presented above (e.g.
Tables 3 and 4), the population of C. erionotae from Sabah
would have been collected from E. thrax only, since
E. torus does not occur there, whereas that from Thailand
is likely to have been reared from both E. thrax and
E. torus. Control in Hawaii was rapid, and the strain of
C. erionotae from Thailand was probably already estab-
lished and bringing the pest under control before that
from Sabah was released. In 1974, C. erionotae cultures of
the Thailand strain were sent from Hawaii to Guam, and
from Guam to Saipan in 1974–75, in both cases giving
good control [45, 49]. Both parasitoids were also sent
from Hawaii to Taiwan for the control of E. torus; they
were released in 1987 and both became established
[182]. The impact has not been fully reported, although
Teruya [138] found E. torus to have ‘a low pest status’,
infestation ranging from 0.1 to 7% with an average of 4%,
but he did not find the introduced parasitoids.
Erionota thrax may have spread into Irian Jaya before
1983 as it appeared by the border in north-western
mainland PNG in 1983, and then spread rapidly through
http://www.cabi.org/cabreviews
Matthew J. W. Cock 17
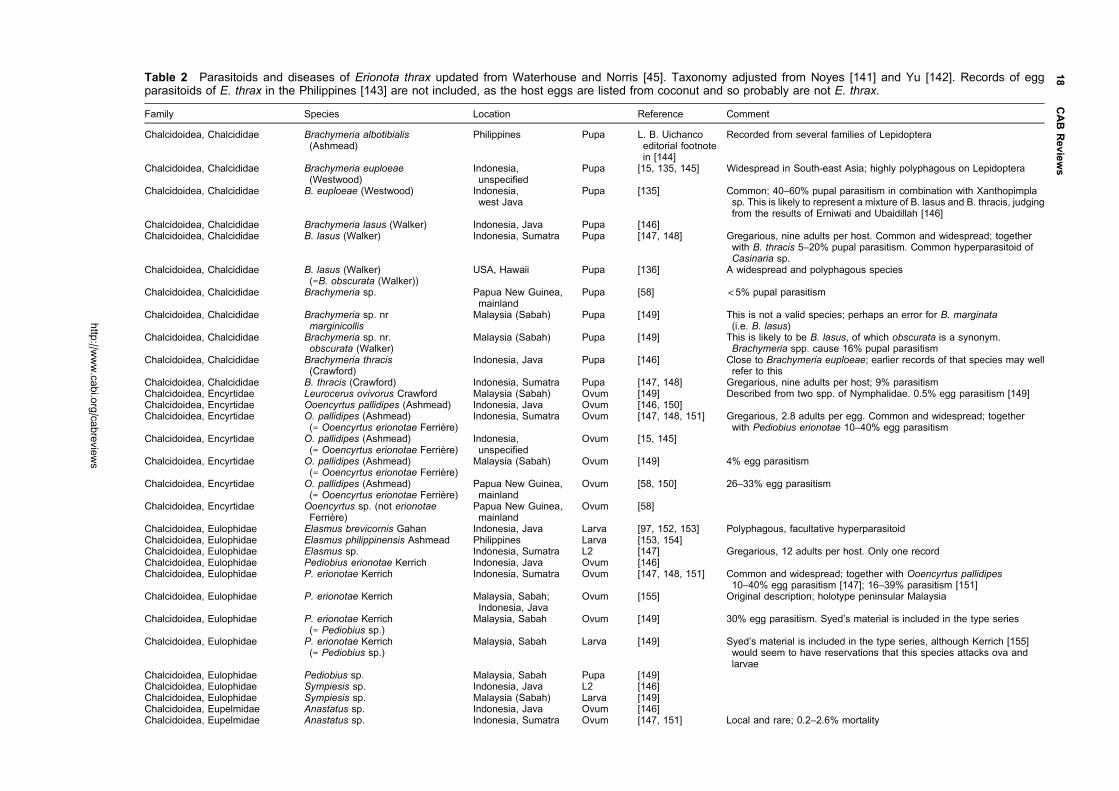
Table 2 Parasitoids and diseases of Erionota thrax updated from Waterhouse and Norris [45]. Taxonomy adjusted from Noyes [141] and Yu [142]. Records of eggparasitoids of E. thrax in the Philippines [143] are not included, as the host eggs are listed from coconut and so probably are not E. thrax.
Family Species Location Reference Comment
Chalcidoidea, Chalcididae Brachymeria albotibialis(Ashmead)
Philippines Pupa L. B. Uichancoeditorial footnotein [144]
Recorded from several families of Lepidoptera
Chalcidoidea, Chalcididae Brachymeria euploeae(Westwood)
Indonesia,unspecified
Pupa [15, 135, 145] Widespread in South-east Asia; highly polyphagous on Lepidoptera
Chalcidoidea, Chalcididae B. euploeae (Westwood) Indonesia,west Java
Pupa [135] Common; 40–60% pupal parasitism in combination with Xanthopimplasp. This is likely to represent a mixture of B. lasus and B. thracis, judgingfrom the results of Erniwati and Ubaidillah [146]
Chalcidoidea, Chalcididae Brachymeria lasus (Walker) Indonesia, Java Pupa [146]Chalcidoidea, Chalcididae B. lasus (Walker) Indonesia, Sumatra Pupa [147, 148] Gregarious, nine adults per host. Common and widespread; together
with B. thracis 5–20% pupal parasitism. Common hyperparasitoid ofCasinaria sp.
Chalcidoidea, Chalcididae B. lasus (Walker)(=B. obscurata (Walker))
USA, Hawaii Pupa [136] A widespread and polyphagous species
Chalcidoidea, Chalcididae Brachymeria sp. Papua New Guinea,mainland
Pupa [58] < 5% pupal parasitism
Chalcidoidea, Chalcididae Brachymeria sp. nrmarginicollis
Malaysia (Sabah) Pupa [149] This is not a valid species; perhaps an error for B. marginata(i.e. B. lasus)
Chalcidoidea, Chalcididae Brachymeria sp. nr.obscurata (Walker)
Malaysia (Sabah) Pupa [149] This is likely to be B. lasus, of which obscurata is a synonym.Brachymeria spp. cause 16% pupal parasitism
Chalcidoidea, Chalcididae Brachymeria thracis(Crawford)
Indonesia, Java Pupa [146] Close to Brachymeria euploeae; earlier records of that species may wellrefer to this
Chalcidoidea, Chalcididae B. thracis (Crawford) Indonesia, Sumatra Pupa [147, 148] Gregarious, nine adults per host; 9% parasitismChalcidoidea, Encyrtidae Leurocerus ovivorus Crawford Malaysia (Sabah) Ovum [149] Described from two spp. of Nymphalidae. 0.5% egg parasitism [149]Chalcidoidea, Encyrtidae Ooencyrtus pallidipes (Ashmead) Indonesia, Java Ovum [146, 150]Chalcidoidea, Encyrtidae O. pallidipes (Ashmead)
(= Ooencyrtus erionotae Ferriere)Indonesia, Sumatra Ovum [147, 148, 151] Gregarious, 2.8 adults per egg. Common and widespread; together
with Pediobius erionotae 10–40% egg parasitismChalcidoidea, Encyrtidae O. pallidipes (Ashmead)
(= Ooencyrtus erionotae Ferriere)Indonesia,unspecified
Ovum [15, 145]
Chalcidoidea, Encyrtidae O. pallidipes (Ashmead)(= Ooencyrtus erionotae Ferriere)
Malaysia (Sabah) Ovum [149] 4% egg parasitism
Chalcidoidea, Encyrtidae O. pallidipes (Ashmead)(= Ooencyrtus erionotae Ferriere)
Papua New Guinea,mainland
Ovum [58, 150] 26–33% egg parasitism
Chalcidoidea, Encyrtidae Ooencyrtus sp. (not erionotaeFerriere)
Papua New Guinea,mainland
Ovum [58]
Chalcidoidea, Eulophidae Elasmus brevicornis Gahan Indonesia, Java Larva [97, 152, 153] Polyphagous, facultative hyperparasitoidChalcidoidea, Eulophidae Elasmus philippinensis Ashmead Philippines Larva [153, 154]Chalcidoidea, Eulophidae Elasmus sp. Indonesia, Sumatra L2 [147] Gregarious, 12 adults per host. Only one recordChalcidoidea, Eulophidae Pediobius erionotae Kerrich Indonesia, Java Ovum [146]Chalcidoidea, Eulophidae P. erionotae Kerrich Indonesia, Sumatra Ovum [147, 148, 151] Common and widespread; together with Ooencyrtus pallidipes
10–40% egg parasitism [147]; 16–39% parasitism [151]Chalcidoidea, Eulophidae P. erionotae Kerrich Malaysia, Sabah;
Indonesia, JavaOvum [155] Original description; holotype peninsular Malaysia
Chalcidoidea, Eulophidae P. erionotae Kerrich(= Pediobius sp.)
Malaysia, Sabah Ovum [149] 30% egg parasitism. Syed’s material is included in the type series
Chalcidoidea, Eulophidae P. erionotae Kerrich(= Pediobius sp.)
Malaysia, Sabah Larva [149] Syed’s material is included in the type series, although Kerrich [155]would seem to have reservations that this species attacks ova andlarvae
Chalcidoidea, Eulophidae Pediobius sp. Malaysia, Sabah Pupa [149]Chalcidoidea, Eulophidae Sympiesis sp. Indonesia, Java L2 [146]Chalcidoidea, Eulophidae Sympiesis sp. Malaysia (Sabah) Larva [149]Chalcidoidea, Eupelmidae Anastatus sp. Indonesia, Java Ovum [146]Chalcidoidea, Eupelmidae Anastatus sp. Indonesia, Sumatra Ovum [147, 151] Local and rare; 0.2–2.6% mortality
http
://www.cabi.o
rg/cabreviews
18
CAB
Reviews
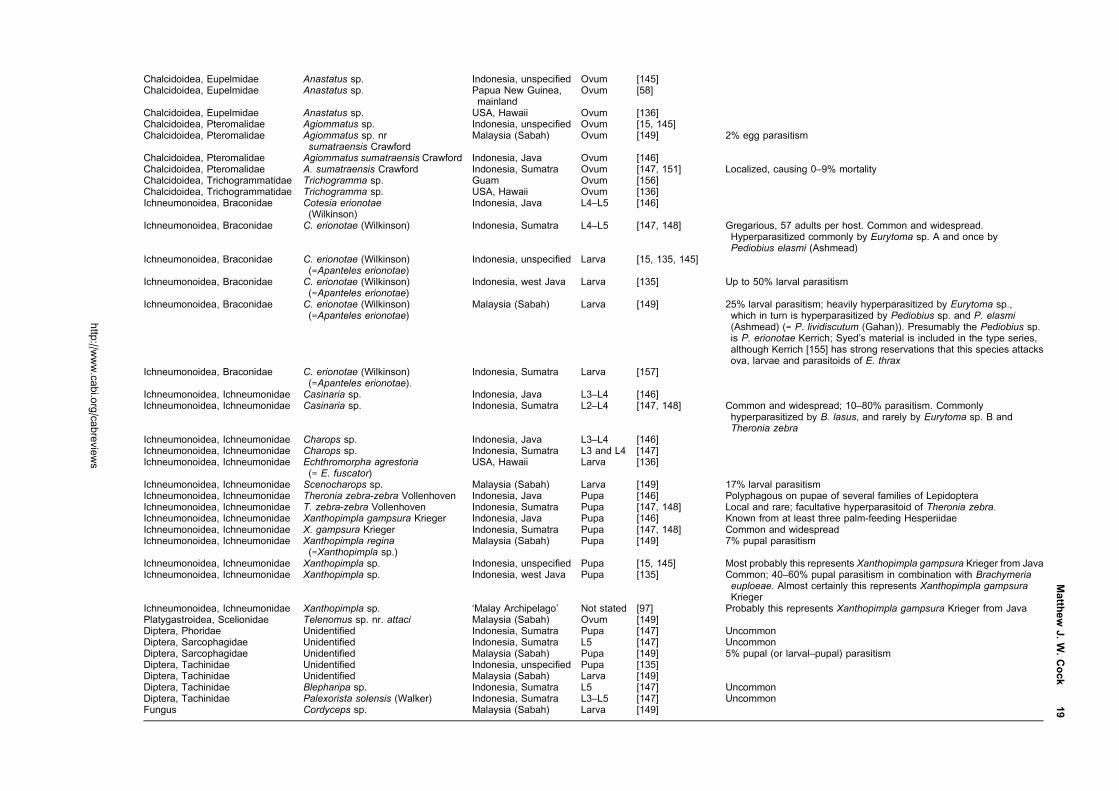
Chalcidoidea, Eupelmidae Anastatus sp. Indonesia, unspecified Ovum [145]Chalcidoidea, Eupelmidae Anastatus sp. Papua New Guinea,
mainlandOvum [58]
Chalcidoidea, Eupelmidae Anastatus sp. USA, Hawaii Ovum [136]Chalcidoidea, Pteromalidae Agiommatus sp. Indonesia, unspecified Ovum [15, 145]Chalcidoidea, Pteromalidae Agiommatus sp. nr
sumatraensis CrawfordMalaysia (Sabah) Ovum [149] 2% egg parasitism
Chalcidoidea, Pteromalidae Agiommatus sumatraensis Crawford Indonesia, Java Ovum [146]Chalcidoidea, Pteromalidae A. sumatraensis Crawford Indonesia, Sumatra Ovum [147, 151] Localized, causing 0–9% mortalityChalcidoidea, Trichogrammatidae Trichogramma sp. Guam Ovum [156]Chalcidoidea, Trichogrammatidae Trichogramma sp. USA, Hawaii Ovum [136]Ichneumonoidea, Braconidae Cotesia erionotae
(Wilkinson)Indonesia, Java L4–L5 [146]
Ichneumonoidea, Braconidae C. erionotae (Wilkinson) Indonesia, Sumatra L4–L5 [147, 148] Gregarious, 57 adults per host. Common and widespread.Hyperparasitized commonly by Eurytoma sp. A and once byPediobius elasmi (Ashmead)
Ichneumonoidea, Braconidae C. erionotae (Wilkinson)(=Apanteles erionotae)
Indonesia, unspecified Larva [15, 135, 145]
Ichneumonoidea, Braconidae C. erionotae (Wilkinson)(=Apanteles erionotae)
Indonesia, west Java Larva [135] Up to 50% larval parasitism
Ichneumonoidea, Braconidae C. erionotae (Wilkinson)(=Apanteles erionotae)
Malaysia (Sabah) Larva [149] 25% larval parasitism; heavily hyperparasitized by Eurytoma sp.,which in turn is hyperparasitized by Pediobius sp. and P. elasmi(Ashmead) (= P. lividiscutum (Gahan)). Presumably the Pediobius sp.is P. erionotae Kerrich; Syed’s material is included in the type series,although Kerrich [155] has strong reservations that this species attacksova, larvae and parasitoids of E. thrax
Ichneumonoidea, Braconidae C. erionotae (Wilkinson)(=Apanteles erionotae).
Indonesia, Sumatra Larva [157]
Ichneumonoidea, Ichneumonidae Casinaria sp. Indonesia, Java L3–L4 [146]Ichneumonoidea, Ichneumonidae Casinaria sp. Indonesia, Sumatra L2–L4 [147, 148] Common and widespread; 10–80% parasitism. Commonly
hyperparasitized by B. lasus, and rarely by Eurytoma sp. B andTheronia zebra
Ichneumonoidea, Ichneumonidae Charops sp. Indonesia, Java L3–L4 [146]Ichneumonoidea, Ichneumonidae Charops sp. Indonesia, Sumatra L3 and L4 [147]Ichneumonoidea, Ichneumonidae Echthromorpha agrestoria
(= E. fuscator)USA, Hawaii Larva [136]
Ichneumonoidea, Ichneumonidae Scenocharops sp. Malaysia (Sabah) Larva [149] 17% larval parasitismIchneumonoidea, Ichneumonidae Theronia zebra-zebra Vollenhoven Indonesia, Java Pupa [146] Polyphagous on pupae of several families of LepidopteraIchneumonoidea, Ichneumonidae T. zebra-zebra Vollenhoven Indonesia, Sumatra Pupa [147, 148] Local and rare; facultative hyperparasitoid of Theronia zebra.Ichneumonoidea, Ichneumonidae Xanthopimpla gampsura Krieger Indonesia, Java Pupa [146] Known from at least three palm-feeding HesperiidaeIchneumonoidea, Ichneumonidae X. gampsura Krieger Indonesia, Sumatra Pupa [147, 148] Common and widespreadIchneumonoidea, Ichneumonidae Xanthopimpla regina
(=Xanthopimpla sp.)Malaysia (Sabah) Pupa [149] 7% pupal parasitism
Ichneumonoidea, Ichneumonidae Xanthopimpla sp. Indonesia, unspecified Pupa [15, 145] Most probably this represents Xanthopimpla gampsura Krieger from JavaIchneumonoidea, Ichneumonidae Xanthopimpla sp. Indonesia, west Java Pupa [135] Common; 40–60% pupal parasitism in combination with Brachymeria
euploeae. Almost certainly this represents Xanthopimpla gampsuraKrieger
Ichneumonoidea, Ichneumonidae Xanthopimpla sp. ‘Malay Archipelago’ Not stated [97] Probably this represents Xanthopimpla gampsura Krieger from JavaPlatygastroidea, Scelionidae Telenomus sp. nr. attaci Malaysia (Sabah) Ovum [149]Diptera, Phoridae Unidentified Indonesia, Sumatra Pupa [147] UncommonDiptera, Sarcophagidae Unidentified Indonesia, Sumatra L5 [147] UncommonDiptera, Sarcophagidae Unidentified Malaysia (Sabah) Pupa [149] 5% pupal (or larval–pupal) parasitismDiptera, Tachinidae Unidentified Indonesia, unspecified Pupa [135]Diptera, Tachinidae Unidentified Malaysia (Sabah) Larva [149]Diptera, Tachinidae Blepharipa sp. Indonesia, Sumatra L5 [147] UncommonDiptera, Tachinidae Palexorista solensis (Walker) Indonesia, Sumatra L3–L5 [147] UncommonFungus Cordyceps sp. Malaysia (Sabah) Larva [149]
http
://www.cabi.o
rg/cabreviews
Matth
ew
J.W.Cock
19

the island over five years, and started to colonize
the other islands of PNG [58]. It spread together with its
egg-parasitoid, O. pallidipes. However, control was not
adequate with the egg parasitoid alone, and so C. erionotae
from Guam was introduced after quarantine safety tests
[48]. This rapidly resulted in good biological control of
the pest. The programme in PNG was supported by
ACIAR (Australian Centre for International Agricultural
Research), who subsequently funded assessments of the
benefits to PNG and Australia [47] and the impact on
poverty in PNG [190].
Waterhouse et al. [47] extrapolated from several
studies on defoliation and fruit loss indicating that more
than 16% defoliation will cause loss of yield, and 50%
defoliation will cause 28% yield loss. As the banana skipper
spread, it destroyed an average of some 60% of banana
leaves, leading to both a serious delay in fruit maturation
and reduced weight of banana bunches. After C. erionotae
was introduced and established, there was a reduction in
the estimated average leaf damage from 60 to 5%. The
losses due to E. thrax each year were calculated at 30% of
crop, of which 95% was saved by the biological control
introduction. Using a discount rate of 5% per year over
the 30 year period, the net present value of lost pro-
duction due to E. thrax amounts to approximately
A$301.8 million. The value of damage prevented by bio-
logical control is estimated at A$201.6 million.
The reduction of banana skipper abundance by 90%
in Southern PNG has correspondingly reduced the
chance of adults invading not only Australian islands in
the Torres Strait but also the Australian mainland.
The successful biological control programme in PNG
has therefore provided significant benefits to banana
production in Australia. Assuming similar levels of damage
in Australia as in PNG, these benefits were estimated to
be A$223 million to the year 2020. The assessment
of benefits did not consider the social and cultural uses
of banana leaves, nor the possible benefits in Irian Jaya, to
which the parasitoid almost certainly spread.
Bauer et al. [190] considered the impact of the
PNG biological control programme on poverty. Poverty
through income deprivation is a significant problem in
PNG. Estimates from a 1996 household income survey
suggested that around 30% of the PNG population have
household incomes below the poverty line (i.e. income is
not sufficient to sustain 2200 calories per day of food
consumption and cover the cost of essential non-food
items). The biological control of E. thrax has had a sig-
nificant beneficial impact on the effective incomes of most
PNG households by improving the supply of bananas
and by lowering the price of bananas to purchasers.
The improvement in income due to biological control
measures against the banana skipper would put between
6000 and 15 000 rural farmers (depending on assump-
tions) above the poverty line who would otherwise have
been below it. The successful implementation of the pro-
ject has also contributed to lower urban banana prices,Table
3ParasitoidsanddiseasesofErionota
torus.Taxonomyadjustedfrom
Noyes[141]andYu[142]
Family
Species
Location
Reference
Comment
Chalcidoidea,Encyrtidae
Leurocerushongkongensis
SubbaRao
China,HongKong
Ovum
[9,158]
Solitary
species
Chalcidoidea,Encyrtidae
Ooencyrtuspallidipes(Ashmead)
(=OoencyrtuserionotaeFerriere)
Mauritius
Ovum
[45,159]
Introducedto
anareawhere
itis
anticipatedonly
E.torusoccurs
Chalcidoidea,Eulophidae
Elasmusphilippinensis
Ashmead
Taiwan
Larva
[131,160]
Ectoparasitoid
Chalcidoidea,Eupelm
idae
Anastatussp.
Japan,Okinawa
Larva
[161]
Chalcidoidea,Pteromalidae
Agiommatuserionotae
China,Fujian
Ovum
[150,162]
Chalcidoidea,Trichogrammatidae
Unidentified
Japan,Okinawa
Ovum
[161]
Chalcidoidea,unspecified
Unidentified
China,HongKong
Ovum
[121]
Ichneumonoidea,Braconidae
Cotesia
erionotae(W
ilkinson)
(=Apanteleserionotae).
Mauritius
Larva
[45,159]
Introducedto
anareawhere
itis
anticipatedonly
E.torusoccurs
Ichneumonoidea,Ichneumonidae
Xanthopim
pla
sp.reginagroup
China,HongKong
Pupa
[9]
Diptera,Tachinidae
Unidentified
Taiwan
Pupa
[138]
Diptera,Tachinidae
Exorista
japonica(Townsend)
Japan,Okinawa
Pupa
[161]
Unspecifieddisease
Japan,Okinawa
Larva,pupa
[161]
http://www.cabi.org/cabreviews
20 CAB Reviews
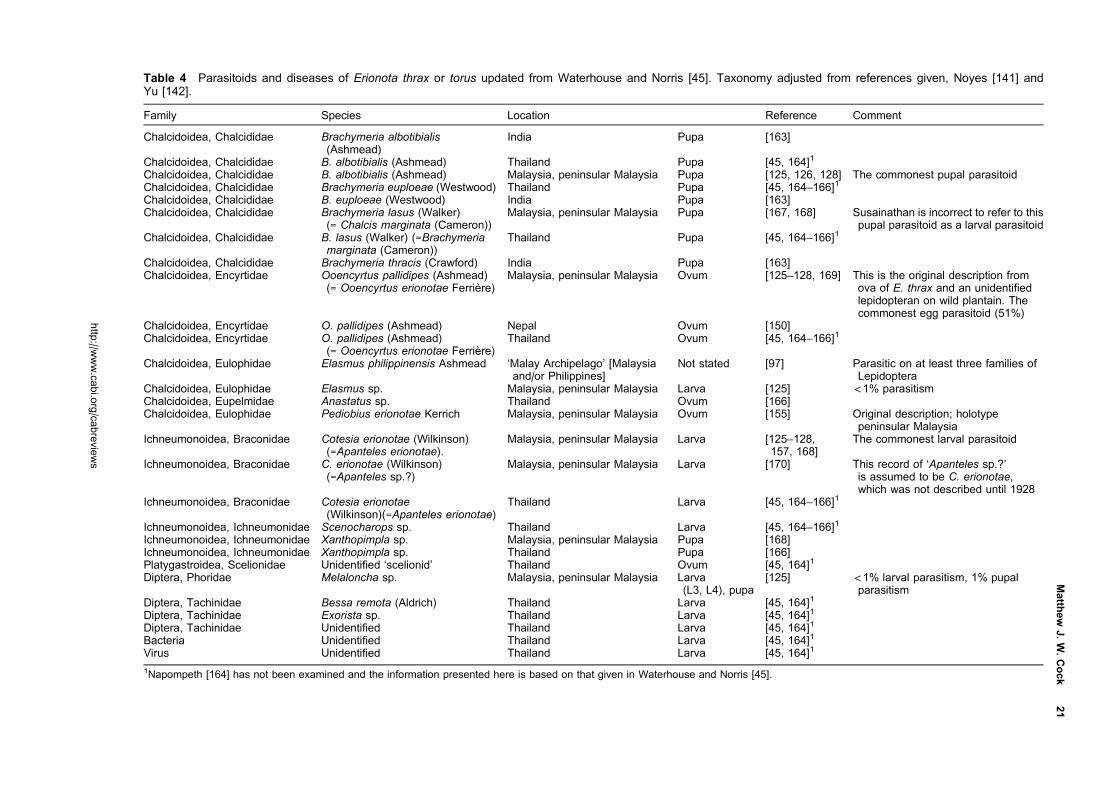
Table 4 Parasitoids and diseases of Erionota thrax or torus updated from Waterhouse and Norris [45]. Taxonomy adjusted from references given, Noyes [141] andYu [142].
Family Species Location Reference Comment
Chalcidoidea, Chalcididae Brachymeria albotibialis(Ashmead)
India Pupa [163]
Chalcidoidea, Chalcididae B. albotibialis (Ashmead) Thailand Pupa [45, 164]1
Chalcidoidea, Chalcididae B. albotibialis (Ashmead) Malaysia, peninsular Malaysia Pupa [125, 126, 128] The commonest pupal parasitoidChalcidoidea, Chalcididae Brachymeria euploeae (Westwood) Thailand Pupa [45, 164–166]1
Chalcidoidea, Chalcididae B. euploeae (Westwood) India Pupa [163]Chalcidoidea, Chalcididae Brachymeria lasus (Walker)
(= Chalcis marginata (Cameron))Malaysia, peninsular Malaysia Pupa [167, 168] Susainathan is incorrect to refer to this
pupal parasitoid as a larval parasitoidChalcidoidea, Chalcididae B. lasus (Walker) (=Brachymeria
marginata (Cameron))Thailand Pupa [45, 164–166]1
Chalcidoidea, Chalcididae Brachymeria thracis (Crawford) India Pupa [163]Chalcidoidea, Encyrtidae Ooencyrtus pallidipes (Ashmead)
(= Ooencyrtus erionotae Ferriere)Malaysia, peninsular Malaysia Ovum [125–128, 169] This is the original description from
ova of E. thrax and an unidentifiedlepidopteran on wild plantain. Thecommonest egg parasitoid (51%)
Chalcidoidea, Encyrtidae O. pallidipes (Ashmead) Nepal Ovum [150]Chalcidoidea, Encyrtidae O. pallidipes (Ashmead)
(= Ooencyrtus erionotae Ferriere)Thailand Ovum [45, 164–166]1
Chalcidoidea, Eulophidae Elasmus philippinensis Ashmead ‘Malay Archipelago’ [Malaysiaand/or Philippines]
Not stated [97] Parasitic on at least three families ofLepidoptera
Chalcidoidea, Eulophidae Elasmus sp. Malaysia, peninsular Malaysia Larva [125] < 1% parasitismChalcidoidea, Eupelmidae Anastatus sp. Thailand Ovum [166]Chalcidoidea, Eulophidae Pediobius erionotae Kerrich Malaysia, peninsular Malaysia Ovum [155] Original description; holotype
peninsular MalaysiaIchneumonoidea, Braconidae Cotesia erionotae (Wilkinson)
(=Apanteles erionotae).Malaysia, peninsular Malaysia Larva [125–128,
157, 168]The commonest larval parasitoid
Ichneumonoidea, Braconidae C. erionotae (Wilkinson)(=Apanteles sp.?)
Malaysia, peninsular Malaysia Larva [170] This record of ‘Apanteles sp.?’is assumed to be C. erionotae,which was not described until 1928
Ichneumonoidea, Braconidae Cotesia erionotae(Wilkinson)(=Apanteles erionotae)
Thailand Larva [45, 164–166]1
Ichneumonoidea, Ichneumonidae Scenocharops sp. Thailand Larva [45, 164–166]1
Ichneumonoidea, Ichneumonidae Xanthopimpla sp. Malaysia, peninsular Malaysia Pupa [168]Ichneumonoidea, Ichneumonidae Xanthopimpla sp. Thailand Pupa [166]Platygastroidea, Scelionidae Unidentified ‘scelionid’ Thailand Ovum [45, 164]1
Diptera, Phoridae Melaloncha sp. Malaysia, peninsular Malaysia Larva(L3, L4), pupa
[125] < 1% larval parasitism, 1% pupalparasitism
Diptera, Tachinidae Bessa remota (Aldrich) Thailand Larva [45, 164]1
Diptera, Tachinidae Exorista sp. Thailand Larva [45, 164]1
Diptera, Tachinidae Unidentified Thailand Larva [45, 164]1
Bacteria Unidentified Thailand Larva [45, 164]1
Virus Unidentified Thailand Larva [45, 164]1
1Napompeth [164] has not been examined and the information presented here is based on that given in Waterhouse and Norris [45].
http
://www.cabi.o
rg/cabreviews
Matth
ew
J.W.Cock
21
Table 5 Introductions for the biological control of E. torus and E. thrax, updated from Waterhouse and Norris [45]
Target Country Species Released Source Result References
E. torus Mauritius Agiommatus sp.nr. sumatraensis
1971–1972 Malaysia (Sabah) Recoveries, but notestablished
[149, 179], J. Monty, personalcommunication, 1987 (in [45])
Cotesia erionotae 1971–1974 Malaysia (Sabah) Established [149, 159, 179, 180]Ooencyrtus pallidipes 1971–1972 Malaysia (Sabah) Established [149, 159, 179]Scenocharops sp. 1971–1974 Malaysia (Sabah) Not established [149, 179, 180, 181], J. Monty,
personal communication (in [45])E. torus Taiwan C. erionotae 1987 Thailand via Hawaii Established [182]
O. pallidipes 1987 Guam via Hawaii Established [182]E. thrax Hawaii C. erionotae 1973–1974 Thailand Established [136, 183, 184]
1974 Malaysia (Sabah) Unknown (no way todistinguish from Thailandstrain already becomingestablished)1
[136, 180, 184]
O. pallidipes 1973 Guam2 Established [183, 184]Scenocharops sp. 1974 Malaysia (Sabah) Not established [136, 180, 184]
E. thrax Guam3 C. erionotae 1974 Thailand via Hawaii Established [49, 156, 185, 186]4
E. thrax Saipan C. erionotae 1974–1975 Thailand via Guam Established [49, 156]E. thrax Papua New
Guinea (mainland)C. erionotae 1990 Thailand via Hawaii
and GuamEstablished [48]
1This could be done today using molecular methods.2Ooencyrtus pallidipes seems to have spread accidentally to Guam without deliberate human assistance.3A Brachymeria sp. was introduced from Papua New Guinea for the biological control of Pericyma cruegeri (Butler) (Erebidae), became established and has been reared from pupae of E. thrax[49]. It has been referred to as Brachymeria sp. [49], B. albotibialis [156, 186], B. euploeae [45] and B. lasus [187]; all three Brachymeria names are currently valid species [141], and all have beenreported to parasitize E. thrax in South-East Asia (Table 4).4Reference [186] not examined, but based on citation in Waterhouse and Norris [45].
http
://www.cabi.o
rg/cabreviews
22
CAB
Reviews

in that the fall in supply of bananas caused by the pest
would have forced banana prices up. Banana prices
are likely to have risen by about 15% had the biological
control of the skipper not been achieved. Preventing this
increase in market prices for bananas of about 15% by the
introduction of the biological control measures resulted
in about 28 000 purchasers of bananas living just above
the poverty line when they otherwise would have been
below it. The biological control program increased
effective incomes across all income levels of banana pro-
ducers and banana consumers.
Discussion
The available information on the classification, taxonomy,
food plants, natural enemies and biological control of two
species of Erionota that feed on Musa spp. and one that
feeds on palms have been presented. It is concluded that
E. thrax and E. torus feed as larvae on Musa spp., and to a
limited extent some related species of Zingiberales, but
records of these species from palms and other plant
groups are in error. The records from palms are now
attributed to E. acroleuca. This distinction is important in
order to understand the ecology of these crop-feeding
butterflies so that management strategies are appropriate.
Where E. thrax and E. torus occur together, economic
entomologists have hitherto treated them as a single
species. There is no published information to show
whether the occasional outbreaks of banana skipper in
this region where both species are indigenous are due
to one or both of these species. Similarly, it is not
known whether the associated parasitoids attack both
species equally or show preference for one or the other.
Nevertheless, biological control has been successful
without knowing about the preferences of parasitoids for
the two species, or whether the material used was col-
lected from one or both Erionota spp. in Thailand. As
E. torus in Mauritius was misidentified as E. thrax, it
was controlled using parasitoids of E. thrax from Sabah.
There are plenty of examples where a lack of under-
standing of the taxonomy of the target pest of a biological
control programme has led to a mis-match of parasitoids
and target (see examples in [191]), and the author firmly
supports the view that good taxonomy is critical to suc-
cess in many biological control programmes, but in the
case of Erionota spp. this does not seem to have been a
critical factor.
For the foreseeable future, it would be a sensible pre-
caution to ensure that voucher specimens are preserved
and documented for all observations on Erionota spp. All
three species should be considered quarantine risks for
tropical areas where they do not naturally occur or have
not already spread, and an improved understanding of
their ecology should improve pest risk assessments
relating to their spread in future.
Research Gaps
In light of the foregoing, the following research questions
and gaps are identified:
� Molecular methods, particularly barcoding, are needed
to confirm identifications of early stages and develop
identification tools.
� Further collections and reliable identification of
adults (by examination of the genitalia and/or barcod-
ing) are needed to test the conclusion that E. thrax
and E. torus feed only on Musa spp. and a few similar
species, whereas E. acroleuca feeds only on various palm
species.
� Material from the Snellen collection should be ex-
amined to see if some of their food plant records can
be aligned with modern taxonomy of Erionota spp.; in
particular should their records of E. thrax from palms
be associated with E. acroleuca, which they considered a
synonym of E. thrax.
� The life history and food plants of the other three
subspecies of E. thrax should be established, which may
help to clarify their taxonomic status.
� As yet, there is no reliable way to separate the early
stages of E. thrax, E. torus and E. acroleuca morpholo-
gically. Careful study and documentation of reared
material, supported by reared vouchers and barcoding
of individuals, is needed to see if diagnostic criteria can
be found.
� Careful observation and behavioural studies of how
the larvae construct their shelters on different food
plants may provide additional diagnostic characters.
� Once the larvae of E. thrax and E. torus can be dis-
tinguished, the host-parasitoid relations for the genus
can be clarified; do parasitoids accept both equally or is
there specialization for one or the other?
� The identity of E. thrax rather than E. torus in the
Andaman Islands may need confirmation. It does not
seem to be a significant pest in the Andaman Islands. If
this is correct, is it because of indigenous or ac-
cidentally introduced parasitoids, perhaps also attacking
E. acroleuca?
� Recognizing the possibility that E. torus may have spread
into areas where only E. thrax thrax was known, it
would be worth making fresh collections to assess the
situation, particularly in Borneo, Sumatra and Java.
� It is considered that E. torus is most probably the
only Erionota sp. in Mauritius, but further collection
should be made to confirm this. Erionota torus is present
in Mauritius at low densities, and seems to cause
no economic damage. It seems most likely that this
is due to the continuing action of the introduced
parasitoids, but this should be investigated. The
reasons for the low population density of E. torus in
Mauritius have implications for the quarantine risk
that it presents to La Reunion and other Afrotropical
areas.
http://www.cabi.org/cabreviews
Matthew J. W. Cock 23

� The new report of E. torus devastating bananas in the
Western Ghats of India suggests that biological control
should be considered in this situation. However, a
careful risk analysis with regard to indigenous species of
Hesperiinae would be needed first. To support this, a
phylogenetic classification of the Hesperiinae would be
necessary, to identify the most closely related genera
which would be most at risk.
� Further collections are required in New Britain, PNG,
to clarify the record of an Erionota sp. feeding on oil
palm.
Acknowledgements
Dr Hideyuki Chiba (Research Associate, Bishop Museum)
and Dr Laurence G. Kirton (Forest Research Institute of
Malaysia, retired) generously shared information, views
and documentation. Dr Chiba and Dr Rienk De Jong
(National Museum of Natural History, The Netherlands)
kindly reviewed parts of the manuscript paper prior to
submission. I thank Dr Feng Zhang (CABI China Regional
Centre), Dr Hiroko Kamei (University of Dundee), and
Dr Tim Haye (CABI Europe-Switzerland) for translation
and interpretation of Chinese, Japanese and German texts
respectively. In addition to the library resources of the
African Butterfly Research Institute (ABRI), the
Biodiversity Heritage Library, the British Library,
CABI, and the Royal Entomological Society, I have been
helped by a variety of people with reprints and copies
of literature, including Dr Rienk De Jong, Dr Ahsol
Hasyim (Indonesia), Suli Khing, Dr Lum Keng Yeang and
Dr Soetikno S Sastroutomo (CABI SE Asia), Dr Banpot
Napompeth (Thailand), Dr Justin Okolle (IRAD/CARBAP/
UB, Cameroon), Dr Don Sands (CSIRO Australia,
retired), Dr K. Veenakumari (India), Dr Janny Vos
(CABI Netherlands), as well as all those scientists and
organizations who have generously made their
publications and information available via the internet.
I thank Dr Charles Dewhurst (PNG Oil Palm Research
Association, retired), Colin Congdon and Forest and
Kim Starr (http://www.starrenvironmental.com/) for
allowing me to use their images. The images of adult
E. torus (Figure 2) are of specimens in Hope Entomological
Collections, Oxford University Museum, and I thank
Dr James Logan and Ms Molly Carter who facilitated
my visit. The images of adults of E. acroleuca (Figure 3)
are of specimens in the collection of the Forest Research
Institute of Malaysia and I thank Phon Chooi-Khim for
facilitating my visit there. I also thank Dr Blanca Huertas
and Mr John Chainey who facilitated a visit to the Natural
History Museum, London. Preparation of this review was
partially funded by the CABI Development Fund, which
is supported by contributions from the Australian Centre
for International Agricultural Research, the UK’s
Department for International Development, the Swiss
Agency for Development and Cooperation and others.
References
1. Evans WH. A Catalogue of the Hesperiidae from Europe,
Asia and Australia in the British Museum (Natural History).
British Museum (Natural History), London; 1949, 502 pp,
53 plates.
2. Inoue S, Kawazoe A. Hesperiid butterflies from South
Vietnam (5). Tyo ta Ga 1970;21(1–2):1–14. Available from:
URL: http://ci.nii.ac.jp/naid/110007706966.
3. Warren AD. The Higher Classification of the Hesperiidae
(Lepidoptera: Hesperioidea) [Ph.D. thesis]. Oregon State
University, Corvalis, OR, USA; 2006, 458 pp.
4. Warren AD, Ogawa JR, Brower AVZ. Phylogenetic
relationships of subfamilies and circumscription of tribes in
the family Hesperiidae (Lepidoptera: Hesperioidea).
Cladistics 2008;24:1–35.
5. Warren AD, Ogawa JR, Brower AVZ. Revised classification
of the family Hesperiidae (Lepidoptera: Hesperioidea) based
on combined molecular and morphological data. Systematic
Entomology 2009;34:467–523.
6. Igarashi S, Fukuda H. The Life Histories of Asian Butterflies,
Volume 2. Tokai University Press, Tokyo, Japan; 2000, 711 pp.
7. Cock MJW, Congdon TCE, Collins SC. Observations on
the biology of Afrotropical Hesperiidae (Lepidoptera). Part 6.
Hesperiinae incertae sedis: palm feeders. Zootaxa
2014;3831(1):1–61.
8. De Jong R. Revision of the Oriental genus Matapa Moore
(Lepidoptera, Hesperiidae) with discussion of its phylogeny
and geographic history. Zoologische Mededelingen
1983;57:243–70.
9. Bascombe MJ, Johnston G, Bascombe FS. The Butterflies
of Hong Kong. Academic Press, London; 1999, xiii + 422 pp,
222 plates.
10. Saji K, Kale P, Kane A. Matapa aria Moore, 1865 – Common
Branded Redeye. In: Kunte K, Kalesh S, Kodandaramaiah U,
editors. Butterflies of India, v. 2.10. Indian Foundation for
Butterflies; 2014. Available from: URL: http://
www.ifoundbutterflies.org/sp/599/Matapa-aria [Last
accessed 1 December 2014].
11. Igarashi S, Fukuda H. The Life Histories of Asian butterflies,
Volume1. Tokai University Press, Tokyo, Japan; 1997, 547 pp.
12. Riley ND. A revision of the continental African species of
the genus Artitropa (Lep. Hesperiidae). Transactions of the
Entomological Society of London 1925;1925:281–8, plate 35.
13. Dickson CGC, Kroon DM (editors). Pennington’s Butterflies of
Southern Africa. A.D. Donker, Johannesburg, South Africa;
1978, 670 pp.
14. Igarashi S. The Hesperiidae loses one species Erionota
harmachis proves conspecific with Unkana mytheca
[In Japanese]. Butterflies, The Butterfly Society of Japan
1996;14:54–5.
15. Kalshoven LGE, van den Laan PA. The Pests of Crops in
Indonesia. P.T. Ichtiar Baru-Van Hoeve, Jakarta, Indonesia;
1981, 701 pp.
http://www.cabi.org/cabreviews
24 CAB Reviews

16. Khoo KC, Ooi PAC, Ho CT. Crop Pests and their
Management in Malaysia. Tropical Press Sdn. Bhd.,
Kuala Lumpur, Malaysia; 1991, ix + 242 pp.
17. De Jong R, Treadaway CG. Notizen uber einige
Erionota-Arten nebst Beschreibung einer neuen Art
(Lepidoptera: Hesperiidae). Entomologische Zeitschrift
1992;102(8):133–52.
18. De Jong R, Treadaway CG. The Hesperiidae (Lepidoptera)
of the Philippines. Zoologische Verhandelingen
1993;288:1–139.
19. Xue G-X, Lo YFP. A taxonomic note on Erionota acroleuca
(Wood-Mason & de Niceville, 1881) stat. rest. (Lepidoptera:
Hesperiidae). Zootaxa 2015;3926(3):445–7.
20. Mabille P. Catalogue des hesperides du Musee royal
d’Histoire naturelle de Bruxelles. Annales de la Societe
entomologique de Belgique 1878;21:12–36.
21. Evans WH. A revision of the genus Erionota Mabille
(Lep.: Hesp.). The Entomologist 1941;74:158–60.
22. Linnaeus C. Systema Naturae. Editio duodecima reformata.
Holmiae, Laurentius Salvius 1767;1(2):[ii] + 533–1328 +
[36] pp.
23. Fruhstorfer H. Neue Hesperiden des Indo-Malayischen
Faunengebietes und Besprechung verwandter Formen.
Deutsche entomologische Zeitscrift [Iris] 1910–1911;
24(3–5):58–104; 25(2–6), 9–78, plate 1.
24. Semper G. Reisen im Archipel der Philippinen. Zweiter Theil.
Wissenschaftliche Resultate. Funfter Band. Die
SchmetterlingederPhilippinischen Inseln.ErsteAbtheilungdie
Tagfalter – Rhopalocera. C.W.KKreidel’s Verlag,Wiesbaden,
Germany; 1886–1892, xiv + 380 pp, Plates A–B, 1–49.
25. Wood-Mason J, de Niceville L. Second list of Rhopalocerous
Lepidoptera from the Andaman Islands with descriptions
of new or little-known species and varieties (Abstract).
Proceedings of the Asiatic Society of Bengal
1881a;1881:142–3.
26. Wood-Mason J, de Niceville L. Second list of Rhopalocerous
Lepidoptera from the Andaman Islands with descriptions of
new or little-known species and varieties. Journal of the
Asiatic Society of Bengal 1881b;1881:243–62, plate XIV.
27. Moore F. Descriptions if new Asiatic diurnal Lepidoptera.
Transactions of the Entomological Society of London
1881;1881:305–13.
28. Swinhoe C. New species of Indian butterflies. Annals and
Magazine of Natural History, Series 6 1890;5:354–65.
29. Evans WH. The Identification of Indian Butterflies. 2nd ed.
Bombay Natural History Society, Madras; 1932, x + 454 pp.
30. von Ribbe C. Beitrage zur Lepidopteren-Fauna von Gross-
Ceram. Deutsche entomologische Zeitscrift. herausgegeben
von der Gesellschaft Iris zu Dresden in Verbindung mit der
Deutschen Entomologischen Gesellschaft zu Berlin.
Lepidopterologisches Heft 1889;2(2):187–265, plate 5.
31. Evans WH. Revisional Notes on the Hesperiidae of Europe,
Asia and Australia. Annals and Magazine of Natural History,
Series 1956;12(9):749–52.
32. Leech JH. New species of Lepidoptera from China.
The Entomologist 1890;23:26–50.
33. Evans WH. New species and subspecies of Hesperiidae
(Lepidoptera) obtained by Herr H. Hone in China in
1930–1936. Proceedings of the Royal Entomological Society
of London. Series B, Taxonomy 1939;8(8):163–6.
34. Hewitson WC. Descriptions of twenty-five new species
of Hesperidae from his own collection. Annals and
Magazine of Natural History, Series 4 1876;18:449–57.
35. Staudinger P. Lepidoptera der Insel Palawan.
Deutsche entomologische Zeitscrift. Herausgegeben
von der Gesellschaft Iris zu Dresden in Verbindung mit der
Deutschen Entomologischen Gesellschaft zu Berlin.
Lepidopterologisches Heft 1889;2(1):3–180, plates 1–2.
36. Clerck C. Icones insectorum rariorum. Sect. secunda cum
nominibus trivialibus logisque e nob. equ. Linni systemate
naturali adjectis. Holmiae; 1764, pp. 1–5, [Register 1–3],
Tab. 17–55.
37. Honey MR, Scoble MJ. Linnaeus’s butterflies (Lepidoptera:
Papilionoidea and Hesperioidea.). Zoological Journal of the
Linnean Society 2001;132:277–399.
38. Kunte K. [ButterflyIndia] Three new species pages, and
notes on Erionota torus from Kerala; 2014. Available from:
URL: https://groups.google.com/forum/#!topic/butterflyindia/
nU4CthWX_2Y
39. Corbet AS, Pendlebery HM, Eliot JN. The Butterflies of the
Malay Peninsula. 3rd ed. Malayan Nature Society, Kuala
Lumpur, Malaysia; 1978, xiv + 578 pp, 35 plates.
40. Yen CY. Molecular phylogeny of the genus Erionota
(Lepidoptera: Hesperiidae), with inferring the origins of two
notorious banana pest species [MSc thesis]. Taiwan National
University, Taipei, Taiwan; 2013, 73 pp.
41. ICZN (International Commission on Zoological
Nomenclature). International Code of Zoological
Nomenclature. 4th ed. The International Trust for Zoological
Nomenclature, London; 1999, xxix + 306 pp.
42. Hewitson WC. Descriptions of twenty new species of
Hesperidae from his own collection. Annals and Magazine
of Natural History, Series 5 1878;1:340–8.
43. Vane-Wright RI, de Jong R. The butterflies of Sulawesi:
annotated checklist for a critical island fauna. Zoologische
Verhandelingen 2003;343:3–267.
44. Gressitt JL. New insects and mites on Guam and Bonin
Islands. In: Notes and Exhibitions, July 15th 1957.
Proceedings of the Hawaiian Entomological Society
1958;16(3):331.
45. Waterhouse DF, Norris KR. 8. Erionota thrax (Linnaeus).
In: Waterhouse DF & Norris KR, editors. Biological Control
Pacific Prospects – Supplement 1. Australian Centre for
International Agricultural Research, Canberra; 1989,
frontispiece + pp. 89–99.
46. Davis CJ, Kawamura K. Notes and exhibitions. Proceedings
of the Hawaiian Entomological Society 1975;22(1):21.
47. Waterhouse D, Dillon B, Vincent D. Economic Benefits to
Papua New Guinea and Australia from the Biological Control
of Banana Skipper (Erionota thrax) ACIAR Project CS2/1988/
002-C. Impact Assessment Series 12. Australian Centre for
International Agricultural Research, ACIAR, Canberra,
Australia; 1998, 36 pp.
48. Sands DPA, Bakker P, Dori FM. Cotesia erionotae
(Wilkinson) (Hymenoptera: Braconidae), for biological control
of banana skipper, Erionota thrax (L.) (Lepidoptera:
Hesperiidae) in Papua New Guinea. Micronesica 1993;Suppl.
4:99–105.
http://www.cabi.org/cabreviews
Matthew J. W. Cock 25

49. Muniappan R. The influence of pests on traditional crops. In:
Proceedings of the Sub-Regional Training Course on
Methods of Controlling Diseases, Insects and Other Pests of
Plants in the South Pacific, October 4–20, 1982, Vaini,
Kingdom of Tonga. MAFF, Tonga/GTZ/USAID/CICP,
Nuku’alofa, Tonga; 1984, pp. 110–9.
50. Nakao HK, Funasaki GY. Introductions for biological control
in Hawaii, 1975 & 1976. Proceedings of the Hawaiian
Entomological Society 1979;23(1):125–8.
51. Riotte JCE, Uchida G. Butterflies of the Hawaiian Islands
according to the stand of late 1976. Proceedings of the
Hawaiian Entomological Society 1979;23(1):33–9.
52. Opler PA, Warren AD. Butterflies of North America. 2.
Scientific Names List for Butterfly Species of North America,
North of Mexico. Contributions of the C. P. Gillette Museum of
Arthropod Diversity, Colorado State University, Fort Collins,
CO, USA; 2002, 79pp.
53. Pelham JP. A catalogue of the butterflies of the United States
and Canada with a complete bibliography of the descriptive
and systematic literature. Journal of Research on the
Lepidoptera 2008;40:i–xiii, 1–652.
54. Kawazoe W, Wakabayashi M. Colored Illustrations of the
Butterflies of Japan. Hoikushu Publishing Co. Ltd., Osaka,
Japan; 1976, viii + 423 pp, 72 plates.
55. Snyder JF, Kumon T. Butterflies of Hawaii; 1999 onwards.
Available from: URL: http://www.asahi-net.or.jp/~ak5t-kmn/
hawaii.htm [Last accessed 1 December 2014]
56. Fukuda H, Nicho K. Butterflies recorded in 2000 and the
succession in butterfly fauna in Guam Is., and Mariana Isles
[In Japanese]. Butterflies, The Butterfly Society of Japan
2001;30:25–35.
57. Parsons M. The Butterflies of Papua New Guinea: their
Systematics and Biology. Academic Press, London; 1999, xvi
+ 736 pp.
58. Sands DPA, Sands MC, Arura M. Banana skipper, Erionota
thrax (L.) (Lepidoptera: Hesperiidae) in Papua New Guinea:
a new pest in the South Pacific region. Micronesica
1991;Suppl. 3:93–8.
59. Monty J. Notes on a new insect pest in Mauritius: the banana
leaf-roller Erionota thrax L. (Lepidoptera, Hesperiidae).
Revue Agricole et Sucriere de l’ile Maurice 1970;49:107–9.
60. Davis PMH, Barnes MJC. The butterflies of Mauritius.
Journal of Research on the Lepidoptera 1991;30:145–61.
61. Veenakumari K, Mohanraj P. Erionota thrax thrax L.
(Lepidoptera: Hesperiidae), a new record to Andaman
Islands. Journal of the Andaman Science Association
1991;7(2):91–2.
62. Muniappan R, Bhattarai SS, Paudel S. Banana leaf roller,
a new pest to Nepal. Newsletter, International Association
of Plant Protection Sciences 2014;9:1.
63. Smith C. Butterflies of Nepal. Revised edition.
Tecpress Service L.P., Bangkok, Thailand; 1994, 368 pp.
64. Mackinnon PW, de Niceville L. A list of the butterflies of
Mussoorie in the Western Himalayas and Neighbouring
Regions. Journal of the Bombay Natural History Society
1897–1898;11:205–21, (1897), 368–406 (1898), 585–604
(1898), plates U (1897), V (1898), W (1898).
65. Smith C. Illustrated Checklist of Nepal’s Butterflies.
New revised and updated edition. Ram Hari Thapur,
Kathmandu, Nepal; 2006, ii+ 129 pp.
66. Teruya T, Araki Y, Osada M. Erionota torus, a new insect
pest of banana plant [In Japanese]. Shokubutu Boeki
1973;27:17–9.
67. Chu YI. The arrival of the banana skipper and its
subsequent propagation in Taiwan [In Chinese].
Yadoriga 1998;135:22–5.
68. Chiang H-S, Hwang M-T. The banana skipper, Erionota torus
Evans (Hesperidae: Lepidoptera): Establishment, distribution
and extent of damage in Taiwan. Tropical Pest Management
1991;37(3):207–10.
69. Raju D, Kunte K, Kalesh S, Manoj P. Erionota torus Evans,
1941 – rounded palm-redeye. In: Kunte K, Kalesh S,
Kodandaramaiah U, editors. Butterflies of India, v. 2.10.
Indian Foundation for Butterflies; 2014. Available from: URL:
http://www.ifoundbutterflies.org/sp/2756/Erionota-torus
[Last accessed 1 December 2014].
70. Okolle JN, Abu Hassan A, Mashhor M, Tripathi L. Bioecology
and management of the banana skipper (Erionota thrax).
Tree and Forestry Science and Biotechnology 2010;4(Special
Issue 1):22–31.
71. EFSA (European Food Safety Authority). Scientific opinion of
the Panel on Plant Health on a request from the European
Commission on pest risk assessment made by France on
Erionota thrax L. considered by France as harmful in French
overseas departments of French Guiana, Guadeloupe,
Martinique and Reunion. The EFSA Journal 2008;672:1–23.
72. Cock MJW. Observations on the biology of Afrotropical
Hesperiidae (Lepidoptera) principally from Kenya. Part 1.
Introduction and Coeliadinae. Zootaxa 2010;2547:1–63.
73. Piepers MC, Snellen PCT. The Rhopalocera of Java
Hesperidae. Martinus Nijhoff & London, William Wesley,
The Hague; 1910, xxvi + 60 + plates V–X.
74. Corbett GH. Insects of coconuts in Malaya. Agricultural
Bulletin of the Straits and Federated Malay States,
General Series 1932;10:106 pp.
75. Lepesme P. Les Insectes des Palmiers. Paul Lechevalier,
Paris; 1947, 909 pp.
76. Fleming WA. Butterflies of West Malaysia and Singapore,
Volume 2. Classey Publications, Farringdon, UK and
Longmans Malaysia; 1975, xi + 93 pp. plates 55–90.
77. Kirton LG. A Naturalist’s Guide to the Butterflies of Peninsular
Malaysia, Singapore and Thailand. John Beaufoy, Oxford,
UK and Forest Research Institute Malaysia, Kepong,
Malaysia; 2014, 176 pp.
78. Horsfield T, Moore F. A Catalogue of the Lepidopterous
Insects in the Museum of the Hon. East-India Company,
Volume I. W.M.H. Allen & Co, London; 1857, v + 278 pp.
+ IV pp. appendix + 11 pp. index + IX plates larvae + VIa
plates adults.
79. Distant WL. Rhopalocera Malayana: a Description of the
Butterflies of the Malay Peninsula. West, Newman & Co.,
London; 1886, xvi + 481 pp. + [5 pp.], 46 plates.
80. Swinhoe C. Lepidoptera Indica. Vol. X. Rhopalocera.
Family Hesperiidae (concluded). Sub-families
Celaenorrhinae, Hesperiinae, Pamphilinae, Astictopterinae,
Suastinae, Erionotinae, Matapinae, Notocryptinae,
Plastingiinae, Erynninae. L. Reeve & Co., London; 1912–13,
vx + 361 pp., plates 757–835.
81. Duport L. Notes sur quelques maladies et ennemis des
plantes cultivees en Extreme Orient. Bulletin Economique
http://www.cabi.org/cabreviews
26 CAB Reviews

de l’Indochine, Hanoi-Haiphong, Nov.–Dec. 1912,
1913;99:147 pp.
82. Williams FX. The natural history of a Philippine nipa house
with descriptions of new wasps. Philippine Journal of Science
1928;35:53–115, plates 1–8.
83. Beller S, Bhenchitr P. A Preliminary List of Insect Pests
and their Host Plants in Siam. (With Notes on their Injury,
Miscellaneous Foods and Utilization of the Host Plants).
Department of Agriculture and Fisheries of Siam, Technical
Bulletin 1, Bangkok, Siam [Thailand]; 1936, 68 pp.
84. Robinson GS, Ackery PR, Kitching IJ, Beccaloni GW,
Hernandez LM. Hostplants of the Moth and Butterfly
Caterpillars of the Oriental Region. The Natural History
Museum, London; 2001, 744 pp.
85. CABI CPC. Datasheet. Erionota thrax; 2014.
Available from: URL: http://www.cabi.org/cpc/datasheet/
21833 [Last accessed 1 December 2014].
86. Bhumannavar BS, Mohanraj P, Ranganath HR, Jacob TK,
Bandyopadhyay AK. Insects of Agricultural Importance in
Andaman and Nicobar Islands. Research Bulletin of the
Central Agricultural Research Institute, Port Blair; 1991, p. 6,
i–xi, 1–49.
87. Veenakumari K, Mohanraj P, Sreekumar PV. Host plant
utilization by butterfly larvae in the Andaman and Nicobar
Islands (Indian Ocean). Journal of Insect Conservation
1997;1:235–46.
88. Mau RFL, Kessing JLM. Pelopidas [sic] thrax (Linnaeus).
Crop Knowledge Master; 1993. Available from: URL: http://
www.extento.hawaii.edu/Kbase/Crop/Type/pelopida.htm
[Last accessed 1 December 2014].
89. Kress WJ, Prince LM, Hahn WJ, Zimmer EA. Unraveling
the evolutionary radiation of the families of the Zingiberales
using morphological and molecular evidence. Systematic
Biology 2001;50(6):926–44.
90. Kress WJ, Specht CD. Between cancer and capricorn:
phylogeny, evolution and ecology of the primarily tropical
Zingiberales. Biologiske Skrifter 2005;55:459–78.
91. Pemberton RW. Predictable risk to native plants in weed
biological control. Oecologia 2000;125:489–94.
92. Keuchenius PE. Ziekten en plagen van de klapperkultuur in
nederlands�indie en hun bestrijding. Teysmannia
1916;27:608–42, plates 1–8.
93. Leefmans S. De gestreepte Dikkoprups van den Klapper
(Hidari irava, Moore). [The striped skipper caterpillar of the
coconut (Hidari irava, Moore)]. [In Dutch with extended
English summary and plate legends]. Mededeelingen van
het Laboratorium voor Plantenziekten 1919;35:15–31 pp,
4 plates.
94. Seitz A. Macrolepidoptera of the World: a Systematic
Description of the Hitherto Known Macrolepidoptera. Division
II: Fauna Exotica. Vol. 9: the Indo-Australian Butterflies.
Lehmann, Stuttgart, Germany; 1927, 1197 pp, 177 plates.
95. Richards PB. Note on the occurrence of three caterpillar
pests on coconuts. Agricultural Bulletin of the Federated
Malay States 1914;3(2):43–5.
96. Richards RM. The diseases and pests of the coconut
palm. Agricultural Bulletin of the Federated Malay States
1917;5(8–9):327–87.
97. Dammerman KW. The Agricultural Zoology of the
Malay Archipelago. J.H. de Bussy Ltd., Amsterdam; 1929,
xi + 473 pp.
98. Hutacharern C, Tubtim N. Checklist of Forest Insects in
Thailand. OEPP Biodiversity Series Vol. 1. Office of
Environmental Policy and Planning, Bangkok, Thailand;
1995, 392 pp.
99. Johari B, Che Aziz A. The growth of one-year old Calamus
trachycoleus at the Forest Research Institute, Kepong.
Malaysian Forester 1981;44(1):23–7.
100. Norani A, Tho YP, Hong LT. Pests and diseases of rattans
and rattan products in Peninsular Malaysia. In: Wong KM,
Manokaran N, editors. Proceedings of the Rattan Seminar,
2–4 October 1984, Kuala Lumpur, Malaysia. Rattan
Information Centre, Forest Research Institute of Malaysia,
Kepong, Malaysia; 1985, pp. 131–5.
101. Maziah Z, Azmi M, Kirton LG. Pests and diseases of rattans.
In: Wan Razali WM, Dransfield J, Manokaran N, editors.
A Guide to the Cultivation of Rattans. Malayan Forest
Records, Forest Research Institute Malaysia, No. 35; 1992,
pp. 127–41.
102. Steiner H, Aminuddin M. Survey of hesperiids found on
Calamus manan in a natural forest and three types of
plantation. Journal of Tropical Forest Science
1997;10(2):256–61.
103. Steiner H, Aminuddin M. Survey of hesperids found in three
age groups of Calamus manan planted under rubber trees.
Journal of Tropical Forest Science 2001;13(2):270–282.
104. Wood BJ. Pests of Oil Palms in Malaysia and their Control.
Incorporated Society of Planters, Kuala Lumpur, Malaysia;
1968, 204 pp.
105. Dewhurst CF, Tennent WJ. New records of butterflies
(Lepidoptera: Hesperiidae, Nymphalidae) feeding on oil
palm (Arecaceae) in West New Britain, Papua New Guinea.
Australian Entomologist 2011;38(4):155–60.
106. Yunus A, Ho TH. List of economic pests, host plants,
parasites and predators in West Malaysia (1920–1978).
Bulletin Ministry of Agriculture, Malaysia 1980;153:538 pp.
107. Pholboon P. A Host List of the Insects of Thailand.
Department of Agriculture, Royal Thai Government and U.S.
Operations Mission to Thailand, Bangkok; 1965, vi + 149 pp.
108. Kitamura M. Notes on the larvae and their foodplants of some
butterflies from the Philippines No. 44 [In Japanese]. Tsu I So
2001;1021/1022:1–27.
109. Kitamura M. A letter from Samar and Leyte (7): on
three hesperiid larvae [In Japanese]. Tsu I So 2003;1112/
1113:1–19.
110. Lever RJAW. Pests of the Coconut Palm. Food and
Agriculture Organization of the United Nations, Rome, Italy;
1969, 190 pp.
111. Hill DS. Agricultural Insect Pests of the Tropics and their
Control. 2nd ed. Cambridge University Press, Cambridge,
UK; 1983, x + 749 pp.
112. Mariau D. The Fauna of Oil Palm and Coconut: Insect
and Mite Pests and their Natural Enemies. Editions Quae,
Paris; 2001, 249 pp, 16 plates.
113. Howard FW, Abad RG. Lepidoptera. In: Howard FW, Moore
D, Giblin-Davis RM, Abad RG, editors. Insects on Palms.
CABI Publishing, Wallingford, Oxon, UK; 2001, pp. 34–81.
http://www.cabi.org/cabreviews
Matthew J. W. Cock 27

114. Hill DS. Pests of Crops in Warmer Climates and their Control.
Springer, Dordrecht, Netherlands; 2008, 704 pp.
115. Muniappan R, Shepard BM, Carner GR, Ooi PAC. Arthropod
Pests of Horticultural Crops in Tropical Asia. CAB
International, Wallingford, UK; 2012, xi + 168 pp.
116. Evans WH. The identification of Indian Butterflies. Part X.
Journal of the Bombay Natural History Society 1926;31:
427–46, 1 plate.
117. Evans WH. The Identification of Indian Butterflies. Bombay
Natural History Society, Madras; 1927, x + 302 pp. + 32
plates.
118. Sevastopulo DG. The food plants of Indian Rhopalocera.
Journal of the Bombay Natural History Society
1973;72(3):156–83.
119. Walker JJ. A preliminary list of the butterflies of Hong-Kong;
based on observations and captures made during the winter
and spring months of 1892 and 1893. Transactions of the
Royal Entomological Society of London 1895;43(4):433–77.
120. Kershaw JC. Butterflies of HongKong. Kelly & Walsh Ltd,
HongKong; 1907, v + 184 pp.
121. Hoffmann WE. Observations on a hesperid leaf-roller and a
lace-bug, two pests of banana in Kwangtung. Lingnan
Science Journal 1935;14:634–49.
122. Marsh JCS. Hong Kong Butterflies. Updated reprint. Shell
Company of Hong Kong Ltd, Hong Kong; 1968, viii + 115 pp,
34 plates.
123. Okolle JN. Population dynamics, within-field and within-plant
distribution of the banana skipper (Erionota thrax L.)
(Lepidoptera: Hesperiidae) and its parasitoids in Penang,
Malaysia [PhD thesis]. School of Biological Sciences,
Science University of Malaysia, Pulau Pinang, Malaysia;
2006, 199 pp.
124. Okolle JN, Mashhor M, Abu Hassan A. Folivorous insect
fauna on two banana cultivars and their association with
non-banana plants. Jurnal Biosains 2006;17(1):89–101.
125. Okolle JN, Mashhor M, Abu Hassan A. Seasonal abundance
of the banana skipper, Erionota thrax (Lepidoptera:
Hesperiidae) and its parasitoids in a commercial plantation
and a subsistence farm in Penang, Malaysia. International.
Journal of Tropical Insect Science 2006;26(3):197–206.
126. Okolle JN, Mashhor M, Abu Hassan A. Spatial distribution
of banana skipper (Erionota thrax L.) (Lepidoptera:
Hesperiidae) and its parasitoids in a Cavendish banana
plantation, Penang, Malaysia. Insect Science
2006;13(5):381–9.
127. Okolle JN, Abu Hassan A, Mashhor M. Host stage
preferences of three major parasitoids of the banana
skipper (Erionota thrax) (Lepidoptera: Hesperiidae).
Indian Journal of Biological Control 2008;22(2):271–6.
128. Okolle JN, Abu Hassan A, Mashhor M. Infestation and
parasitism of banana skipper (Erionota thrax) (Lepidoptera:
Hesperiidae) in relation to banana leaf age, and surface
and distance from field edge. Asian and Australian Journal of
Plant Science and Biotechnology 2009;3(1):61–5.
129. Okolle JN, Abu Hassan A, Mashhor M. Evaluation of selected
insecticides for managing larvae of Erionota thrax and effects
on its parasitoid (Brachymeria albotibialis). Pest Technology
2011;5(1):39–43.
130. Steiner H. The Insect Fauna of Rattan. A Study of Potential
Pest Species on Calamus Manan and Other Rattan Palms in
Peninsula Malaysia. Deutsche Gesellschaft fur Technische
Zusammenarbeit (GTZ) GmbH, Eschborn, Germany; 2001b,
v + 57 pp + [30 pp.] annexes. Available from: URL: http://
www2.gtz.de/dokumente/bib/02-0306.pdf [Last accessed
14 July 2014].
131. Tsai YP, Hwang MT, Tsai JM, Chang HS. Bionomics and
biological control of banana skipper, Erionota torus Evans, in
Taiwan [In Chinese with English abstract, figure legends and
tables]. Chinese Journal of Entomology 1990;10(4):419–26.
132. Taiwan. Digital Taiwan – Culture & Nature. Butterflies
of Formosa – details; 2014. Available from: URL:
http://culture.teldap.tw/culture/index.php?option=
com_content&view= article&catid=163%3Abiosphere-and-
nature&id=1942%3Abutterflies-of-formosa-
details&Itemid=209 [Last accessed 1 December 2014].
133. Steiner H. Rattan und Insekten – eine vergleichende Studie
der Herbivorenfauna von Calamus manan und anderen
Palmenarten auf der Halbinsel Malaysia. Dissertation zur
Erlangung des Doktorgrades der Naturwissenschaften,
Fachbereich Biologie der Johann Wolfgang Goethe –
Universitat, Frankfurt am Main; 2001a, Z1–6, S1–6, I1–10,
D1–2, 298 pp, A1–39, Lebenslauf, Veroffentlichungen.
Available from: URL: http://core.kmi.open.ac.uk/download/
pdf/14505403.pdf
134. Wikipedia. Lepidoptera Indica; 2014. Available from:
URL: http://en.wikipedia.org/wiki/Lepidoptera_Indica
[Last accessed 1 December 2014].
135. Ashari, Eveleens KG. The banana leaf roller (Erionota thrax):
population dynamics, natural biological control by parasites,
and timing of chemical control. In Agricultural Cooperation
Indonesia-The Netherlands. Research Reports 1968–1974.
Section II. Technical contributions; 1974, pp. 364–9.
136. Mau RFL, Murai K, Kumashiro B, Teramoto K. Biological
control of the banana skipper, Pelopidas [sic] thrax
(Linnaeus), (Lepidoptera: Hesperiidae) in Hawaii.
Proceedings of the Hawaiian Entomological Society
1980;23:231–8.
137. Seitz A. Macrolepidoptera of the World: a Systematic
Description of the Hitherto Known Macrolepidoptera.
Section I: The Macrolepidoptera of the Palearctic Region.
Volume 1: The Palearctic Butterflies. Alfred Kernan, Stuttgart,
Germany; 1909, 378 pp, 89 plates.
138. Teruya T. A brief report about the distribution of the banana
skipper, Erionota torus Evans in Taiwan [In Chinese with
English tables and summary]. Okinawa Agriculture
1997;32(1):62–7.
139. Makibayashi I. An observation of the nest making of Erionota
torus final instar larva [In Japanese]. Cho Cho, The
Rhopalocerists’ Magazine, Fukuoka, Japan 1981;4(9):14–18.
140. Ito Y, Nakamori H. The banana skipper’s leaf scroll: a new
overwintering site for the Okinawan paper wasp, Polistes
japonicus. Kontyu, Tokyo 1986;54(3):519–20.
141. Noyes JS. Universal Chalcidoidea Database; 2014. Available
from: URL: http://www.nhm.ac.uk/chalcidoids [Last accessed
1 December 2014].
142. Yu DSK. Home of Ichneumonoidea; 2014. Available from:
URL: http://www.taxapad.com/ [Last accessed 1 December
2014].
143. Alba MC. Egg Parasitoids of Lepidopterous Pests of
Economic Importance in the Philippines. Biotrop Special
Publication No. 36, Bogor, Indonesia; 1989, pp. 123–38.
http://www.cabi.org/cabreviews
28 CAB Reviews

144. Roxas HA. Pericyma cruegeri (Butler): its life history and
economic importance (Noctuidae, Lepidoptera). Philippine
Agriculturist 1927;16(4):229–33, plates 1–2.
145. Kalshoven LGE, Van der Vecht J. De plagen van
de cultuurgewassen in Indonesie, Volume 2. W. Van Hoeve,
‘s-Gravenhage; 1951, pp. 515–1063.
146. Erniwati, Ubaidillah R. Hymenopteran parasitoids associated
with the banana-skipper Erionota thrax L. (Insecta:
Lepidoptera, Hesperiidae) in Java, Indonesia. Biodiversitas
2011;12:76–85.
147. Hasyim A, Hasan N, Syafril H, Nakamura K. Parasitoids of
the banana skipper Erionota thrax (L.) in Sumatera Barat,
Indonesia, with notes on their life history, distribution and
abundance. Tropics 1994;3(2):131–42.
148. Hasyim A, Kamisar, Nakamura K. Mortalitas stadia
pradewasa hama penggulung daun pisang Erionota thrax (L)
yang disebabkan oleh parasitoid. [Mortality of immature
stage banana leaf roller Erionota thrax (L.) attacked by
parasitoids.] [In Indonesian, with English abstract,
figure legend and tables.]. Jurnal Hortikultura
2003;13(2):114–9.
149. Syed RA. Biological Control of Erionota thrax (for Mauritius).
Commonwealth Institute of Biological Control, Annual Report;
1971, pp. 110–1.
150. Huang D-W, Noyes JS. A revision of the Indo-Pacific species
of Ooencyrtus (Hymenoptera: Encyrtidae), parasitoids of the
immature stages of economically important insect species
(mainly Hemiptera and Lepidoptera). Bulletin of the Natural
History Museum, London (Entomology) 1994;63(1):1–136.
151. Hasyim A, Hasan N, Syafril H, Nakamura K. Deteksi dan
tingkat parasitasi parasitoid telur hama penggulung daun
pisang, Erionota thrax (L.) di Sumatera Barat. [Detection of
egg parasitism and its mortality on the banana skipper,
Erionota thrax (L.) eggs in the province of West Sumatera
(Indonesia).] [In Indonesian, with English abstract,
figure legend and tables.]. Jurnal Hortikultura
1999;8(4):1278–83.
152. Gahan AB. Report on a small collection of parasitic
Hymenoptera from Java and Sumatra. Treubia 1922;3(1):
47–52.
153. Ferriere C. The Asiatic and African Species of the Genus
Elasmus, Westw. (Hym., Chalcid.). Bulletin of Entomological
Research 1929;20(4):411–23.
154. Gahan AB. A second lot of parasitic Hymenoptera
from the Philippines. Philippine Journal of Science
1925;27(1):83–110, plate 1.
155. Kerrich GJ. A revision of the tropical and subtropical species
of the eulophid genus Pediobius Walker (Hymenoptera:
Chalcidoidea). Bulletin of the British Museum (Natural
History) (Entomology) 1973;29:113–99.
156. Nafus D, Schreiner I. Biological control activities in the
Mariana Islands from 1911 to 1988. Micronesica
1989;22(1):65–106.
157. Wilkinson DS. A revision of the Indo-Australian species of the
genus Apanteles (Hym. Bracon.). Part 1. Bulletin of
Entomological Research 1928;19:79–105.
158. Subba Rao BR. New genera and species of encyrtids
(Hymenoptera: Encyrtidae). Journal of Natural History
1971;5(2):209–24.
159. Monty J. Entomological news from the Agricultural Services
of the Ministry of Agriculture, Natural Resources and the
Environment. Revue Agricole et Sucriere de l’Ile Maurice
1977;56(3):107–9.
160. Chiang HS. Bionomics and control of banana skipper,
Erionota torus Evans, in Taiwan [In Chinese]. Chinese
Journal of Entomology 1988;Special Publication 2:167–74.
161. Higa T, Ginoza T, Zakimi M. Biological studies on the banana
skipper, Erionota torus Evans in Okinawa. [In Japanese, no
English summary, some tables and figures in English].
Okinawa Agriculture 1979;15:19–37.
162. Huang D-W. Agiommatus erionotus a new species of
Miscogasterinae (Hymenoptera: Chalcidoidea, Pteromalidae)
[In Chinese with English abstract]. Wuji Science Journal
1986;6:103–5.
163. Narendran TC. Family Chalcididae. In: Subba Rao BR and
Hayat M (eds) The Chalcidoidea (Insecta: Hymenoptera) of
India and the adjacent countries. Oriental Insects
1986;20:11–41, 307–310.
164. Napompeth B. Biological Control of Banana Skipper
(Erionota thrax L.) by Using the Larval Parasite (Apanteles
erionotae Wilkinson) in Thailand and Hawaii. National
Biological Control Research Center Annual Report 1975–76.
National Research Council of Thailand, Bangkok, Thailand;
1976, pp. 52–8.
165. Napompeth B. Natural Enemies of the Banana Leaf Roller,
Erionota thrax: L. (Lepidoptera: Hesperiidae) in Thailand.
Technical Paper 2. National Biological Control Research
Center, Kasetsart University, Bangkok; 1978, 11 pp.
166. Napompeth B. Biological Control of Insect Pests andWeeds in
Thailand. Biotrop Special Publication No. 36, Bogor,
Indonesia; 1989, pp. 51–67.
167. Susainathan P. Some important pests of the Malay
Peninsula. In: Fletcher TB, editor. Report of the Proceedings
of the Fifth Entomological Meeting Held at Pusa on the 5th to
10th February 1923. Superintendent Government Printing,
India; 1924, pp. 28–33, xiii + 422 pp.
168. Corbett GH, Miller NCE. A list of insects with their parasites
and predators in Malaya. Malayan Agricultural Journal
1928;16(12):404–24.
169. Ferriere C. New chalcid egg-parasites from South Asia.
Bulletin of Entomological Research 1931;22(2):279–95.
170. Gater BAR. Notes on miscellaneous insects in 1924.
Malayan Agricultural Journal 1925;12:160–7.
171. Gold CS, Pinese B, Pena JE. Pests of banana. In: Pena JE,
Sharp JL, Wysoki M, editors. Tropical Fruit Pests and
Pollinators: Biology, Economic Importance, Natural Enemies
and Control. CAB International, Wallingford, UK; 2002,
pp. 13–56.
172. Tinzaara W, Gold CS. Banana pests and their management.
In: Capinera JL, editor. Encyclopaedia of Entomology,
Volume 2. Springer, Dordrecht, Netherlands; 2008,
pp. 359–78.
173. Ashmead WH. A list of Hymenoptera of the Philippine Islands
with descriptions of new species. Journal of the New York
Entomological Society 1904;12(1):1–22.
174. Austin AD, Dangerfield PC. Synopsis of Australian
Microgastrinae (Hymenoptera: Braconidae), with a key to
genera and description of new taxa. Invertebrate Taxonomy
1992;6:1–76.
http://www.cabi.org/cabreviews
Matthew J. W. Cock 29

175. Chou LY, Chiang HS, Hwang MT. The discovery of
Cotesia erionotae in Taiwan (Hymenoptera: Braconidae)
[In Chinese]. [Journal of Agricultural Research
of China] 1989;38(4):468–70.
176. Gupta A, Kalesh S. Reared parasitic wasps attacking
hesperiids from Western Ghats (Kerala, India) with
description of a new species of Dolichogenidea
(Hymenoptera: Braconidae) as a larval parasitoid of
Thoressa evershedi (Evans) (Lepidoptera: Hesperiidae).
Zootaxa 2012;3413:29–43.
177. Gupta A, Lokhande SA, Soman A. Parasitoids of Hesperiidae
from peninsular India with description of a new species of
Dolichogenidea (Hymenoptera: Braconidae) parasitic on
caterpillar of Borbo cinnara (Wallace) (Lepidoptera:
Hesperiidae). Zootaxa 2013;3701(2):277–90.
178. Krieger R. Ueber die Ichneumonidengattung Xanthopimpla
Sauss. Archiv fur Naturgeschichte 1914;80(6):1–148.
179. Syed RA. Biological Control of Erionota thrax (for Mauritius).
Commonwealth Institute of Biological Control, Annual Report;
1972, p. 81.
180. Syed RA. Supply of parasites of Erionota thrax (for
Mauritius and Hawaii). Commonwealth Institute of
Biological Control, Annual Report; 1974, p. 84.
181. Syed RA. Biological control of Erionota thrax (for Mauritius).
Commonwealth Institute of Biological Control, Annual Report;
1973, p. 81.
182. Lo KC. Biological control of insect and mite pests on crops
in Taiwan – a review and prospection. [In Chinese with
English abstract, tables and references]. Symposium on
the Biological Control of Agricultural Insects and Mites.
Formosan Entomologist 2002;Special Publication, 3:1–25.
183. Nakao HK, Funasaki GY, Davis CJ. Introductions for
biological control in Hawaii, 1973. Proceedings of the
Hawaiian Entomological Society 1975;22(1):109–12.
184. Nakao HK, Funasaki GY. Introduction for biological control
in Hawaii, 1974. Proceedings of the Hawaiian Entomological
Society 1976;22(2):329–31.
185. Stevens LM, Kikuchi R. The control of Erionota thrax (Linn.)
(Lepidoptera: Hesperiidae) by three parasites; Ooencyrtus
erionotae Ferriere, Apanteles erionotae Wilkinson and
Brachymeria obscurataWilkinson with special observation for
B. albotibialis Ashm. University of Guam, unpublished report;
1978.
186. Schreiner I, Nafus D. Accidental introductions of insect
pests to Guam, 1945–1985. Proceedings of the Hawaiian
Entomological Society 1986;27:45–52.
187. Nafus DM, Nechols JR. 55/Poinciana looper. In: Nechols JR,
editor. Biological Control in the Western United States.
Accomplishments and Benefits of Regional Research Project
W-84, 1964–1989. University of California Division of
Agriculture and Natural Resources Publication 3361, Davis,
CA, USA; 1995, pp. 203–4, 357 pp.
188. Syed RA. Biological Control of Erionota thrax (for Mauritius).
Commonwealth Institute of Biological Control, Annual Report;
1970, p. 96.
189. Williams JR. Butterflies of Mauritius. The Author, Mauritius;
1989, 41 pp.
190. Bauer M, Pearce D, Vincent D. Saving a Staple Crop: . . .
Impact of Biological Control of the Banana Skipper on
Poverty Reduction in Papua New Guinea. ACIAR project
CS2/1988/002-C. ACIAR Impact Assessment Series, No 22.
Australian Centre for International Agricultural Research,
Canberra, Australia; 2003, 23 pp.
191. Smith R, Rassmann K, Davies H, King N (editors).
Why Taxonomy Matters. BioNET-INTERNATIONAL,
Egham, UK; 2011. Available from: URL: http://
www.bionet-intl.org/opencms/opencms/caseStudies/
default.jsp [Last accessed 1 December 2014].
http://www.cabi.org/cabreviews
30 CAB Reviews











![Pyroneura liburnia (H [1868]) (Lepidoptera: Hesperiidae) · Rienk DE JONG and Colin G. TREAD AWAY ... [1868]) (Lepidoptera: Hesperiidae) ... collections of the National Museum of](https://img.pdfslide.us/doc/110x75/5ad3c5857f8b9a48398b7d4e/pyroneura-liburnia-h-1868-lepidoptera-hesperiidae-de-jong-and-colin-g-tread.jpg)
















