Embed Size (px)
Citation preview

ORIGINAL PAPER
A comprehensive coastal seabird survey maps out the front linesof ecological change on the western Antarctic Peninsula
Paula Casanovas · Ron Naveen · Steve Forrest ·Jérôme Poncet · Heather J. Lynch
Received: 16 October 2014 / Revised: 14 January 2015 / Accepted: 25 January 2015
© Springer-Verlag Berlin Heidelberg 2015
Abstract Seabirds along the western Antarctic Peninsula
are known to be shifting in abundance and distribution in
response to changing sea ice and prey distributions, but the
spatial extent of these changes has remained an open
question because survey efforts have focused on the more
easily accessed northern coastline. We used a yacht-based
field expedition (January 5–21, 2013) to complete the first
comprehensive penguin (Pygoscelis spp.) and blue-eyed
shag (Phalacrocorax [atriceps] bransfieldensis) populationsurvey of the Graham and Loubet Coasts of the western
Antarctic Peninsula since the mid-1980s. Our surveys
document a sharp transition zone at the northern boundary
of Marguerite Bay; north of this boundary zone, we con-
firm widespread declines in Adelie penguins and increasing
populations of gentoo penguins, but south of this zone we
find Adelie populations that have remained stable or
increased in abundance since the previous surveys by
Poncet and Poncet (Br Antarct Surv Bull 77:109–129,
1987). Marguerite Bay has long been known as a predator
“hotspot,” but our findings suggest that Marguerite Bay has
actually been improving for marine predators for at least
several decades. Marguerite Bay, which has fundamentally
different ocean dynamics than in areas just outside Mar-
guerite Bay, has maintained persistent phytoplankton
blooms over the past decade even as summer sea ice extent,
which can inhibit access to breeding areas, has declined.
This provides further support for the hypothesis that ocean
productivity and sea ice dynamics are critical factors reg-
ulating Adelie penguin abundance in the region and that
Marguerite Bay is now at the front lines of ecological
change in this region.
Keywords Penguins · Blue-eyed shag · Marguerite
Bay · Sea ice · Phytoplankton blooms
Introduction
It is now well established that penguin populations on the
Antarctic Peninsula have been changing over the past three
decades (Trivelpiece et al. 2011; Lynch et al. 2012;
Ducklow et al. 2013). Particular attention has focused on
widespread declines in the abundance of Adelie penguin
(Pygoscelis adeliae) colonies situated along the western
coast of the Antarctic Peninsula (Lynch et al. 2012; Lynch
and LaRue 2014), as well as increasing abundance and
southward expansion of gentoo penguins (P. papua) (Lynchet al. 2012). Evidence suggests that environmental factors
associated with climate change, such as declining sea ice
and shifts in primary productivity, play a significant role in
these shifting patterns of abundance and distribution (Fra-
ser et al. 1992; Smith et al. 1999; Croxall et al. 2002;
Clarke et al. 2007; Murphy et al. 2007; Trivelpiece et al.
2011). Most studies to date, however, have focused on
colonies north of 65°S, and thus information on population
changes south of 65°S and in the vicinity of Marguerite
Bay has been limited. The survey work reported by Poncet
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00300-015-1651-x) contains supplementarymaterial, which is available to authorized users.
P. Casanovas · R. Naveen · S. Forrest
Oceanites, Inc., Chevy Chase, MD, USA
J. Poncet
Golden Fleece Expedition Cruises, Beaver Island,
Falkland Islands
H. J. Lynch (&)
Ecology and Evolution Department, Stony Brook University,
Stony Brook, NY 11794, USA
e-mail: [email protected]
123
Polar Biol
DOI 10.1007/s00300-015-1651-x

and Poncet (1987), who censused penguin colonies in this
region from 1983 to 1987, is the only comprehensive field
survey of the region prior to that reported here. Their data
provide the baseline against which we can assess changes
in the abundance and distribution of penguins breeding
south of 65°S on the western Antarctic Peninsula (WAP).
Monitoring seabird populations in the Antarctic is
important because changes in their abundance and distri-
bution may indicate regional climate change (Croxall et al.
2002) or direct or indirect human disturbance (e.g., tour-
ism, marine traffic, commercial harvesting of marine
resources). However, limited data south of 65°S have made
it impossible to know whether observed changes persist
south of the latitudinal range for which data have been
available. Lynch et al. (2012) found that while Adelie
penguins were declining over most of the western Antarctic
Peninsula (WAP), populations at Bongrain Point, Pourquoi
Pas Island (−67.72°, −67.73°) and Red Rock Ridge
(–68.29°, −67.18°) were actually increasing. At the same
time, gentoo penguins were found to be expanding their
range further south, prompting Lynch et al. (2012) to
predict further expansion south to the Berthelot Islands.
More recent evidence suggests that chinstrap penguins (P.antarctica), which are declining regionally (e.g., Lynch
et al. 2012), may also be establishing new colonies at their
southern range margin (~65°S), although only in areas
already occupied by other pygoscelid penguins (Lynch
et al. 2013). To address these open questions, we organized
a multipronged survey of the Graham and Loubet Coasts
(between 65° and 68° South, including the Marguerite Bay
area) using a comprehensive yacht-based field survey of
the region supported by a coastal survey of high-resolution
satellite imagery. While most of the WAP is being moni-
tored regularly (Lynch et al. 2013), the area south of 65°Sis not routinely visited by passenger vessels. Small yacht
operations allow for the surveying of areas that are extre-
mely difficult or impossible to reach by bigger vessels and
provide greater flexibility for scheduling travel among
sites.
Methods
Field survey
From January 5 to 21, 2013, we used the yacht GoldenFleece (with a 19.5 m steel hull, a beam of 6.19 m, and a
maximum draft of 2.48 m) to survey the WAP coastline
between 65°06′ and 68°18′S, including offshore islands in
the Grandidier Channel, Crystal Sound, the west and
southern coast of Adelaide Island, and the islands in
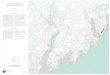
Marguerite Bay (Fig. 1). Over this period, we completed 43
site censuses, where some sites included multiple census
locations due to dispersal of individuals over small island
archipelagos or islands separated from the mainland. Ide-
ally, each “site” encompasses a single breeding population,
but we have defined their boundaries and naming schema
to remain consistent with historical precedent. Mild
weather conditions and the absence of pack ice facilitated
the work, allowing for surveying every day and access to
all targeted sites. We surveyed all of the known breeding
locations described in Poncet and Poncet (1987) south of
Moot Point except for Rhyolite Island (69°40′S, 68°47′W),
which could not be reached because of time/ice constraints
and distance. The survey occurred after the peak of pen-
guin egg-laying (for nest counts) and before the peak of
penguin chick-creching (for chick counts). We mapped,
through waypoints and tracklogs as appropriate, all sur-
veyed sites, as well as the Golden Fleece’s route.We systematically searched all known or discovered
islands, islets, and penguin/shag colonies thereon, relying
upon Capt. Jerome Poncet’s [30 years experience navi-
gating these waters and on his navigation notes from the
surveys described in Poncet and Poncet (1987). With the
exception of the shag survey at Green Island, all of our
surveys were accomplished by researchers (PC, RN, and
SF) on land, proceeding from Golden Fleece to shore by
zodiac. All encountered colonies of gentoo penguins,
chinstrap penguins, Adelie penguins, and blue-eyed shags
were mapped and censused by direct enumeration of
individual occupied nests or chicks [see Lynch et al. (2008)
for details on survey methodology]. We counted nests or
chicks individually three times and report the average of
those three counts; estimates of abundance are, except as
noted, accurate to ±5 %. We focused our survey on pen-
guin breeding sites reported by Poncet and Poncet (1987)
and the Wildlife Awareness Manual (Harris 2006), as well
as at locations that had been determined by remotely
sensed data to potentially contain rookeries.
Gentoo penguins were counted at eight census sites. All
gentoo penguins were past the peak of egg-laying, many
brooding 1–2 week old chicks, and no creching was
observed. In this circumstance, the focus throughout was
on obtaining nest counts. Chinstrap penguins were cens-
used at seven locations and found breeding in small
numbers. Only nests with eggs or 1–2 week old chicks
were observed, and thus, the focus was nest counts. Adelie
penguins were censused at 31 census sites. Adelies were
past the peak of egg-laying, with many chicks already
hatched (mostly 2–4 weeks of age) but not yet starting to
creche, so we focused on nest counts. However, at five sites
in Marguerite Bay (Red Rock Ridge, Lagotellerie Island,
Avian Island, Chatos Island, and Cone Island), a significant
proportion of the colony had creched and we counted
chicks instead of occupied nests. Where it was possible to
do so, both occupied nests and chicks were simultaneously
Polar Biol
123

3
8
9
5
4
7
6
12
1113
10
16
23
19
1514
21
20
27
2824
1817
22
25
31 32
26
33
3529 30
3637
39
4342
4140
38
21
8 km0 4
A B
C
ED
7.50 15 km
0 25 50 km
34
0 30 60 km0 20 40 km
AB
C
D
E
120 Wo
30 Wo 60 So
70 So
80 So
60 Wo
90 Wo
150 Wo
180 Wo
150 Eo
120 Eo
90 Eo
60 Wo
30 Wo
180 Wo
0 70 140 km
65 So
66 So
Fig. 1 Track of the GoldenFleece route representing the
area searched and surveyed for
this study. Areas along this track
not reported in this paper
represent true absence within
the detection limits of the
survey (up to 5 km). a Gerlache
Strait area, b Lemaire Channel
to Green Island area, c Renaud
Island area, d Crystal Sound
area, e Marguerite Bay area.
The numbers on the maps
correspond to the Map ID
numbers in Table 1. All imagery
Copyright (2014) by Digital
Globe, Inc.
Polar Biol
123

censused. We also surveyed all blue-eyed shag (Phala-crocorax [atriceps] bransfieldensis) nests in the region.
Data collected by Poncet and Poncet (1987) were similar
in methodology, though the earlier survey included some
estimates obtained from offshore aboard a yacht as
reflected in their lower precision. Our assessment of pop-
ulation change is based on all the available data for each
site surveyed, including data from Poncet and Poncet
(1987), the Antarctic Site Inventory database (Naveen and
Lynch 2011), and other published sources.
Environmental drivers of Adelie population change
We analyzed changes in penguin abundance in the context of
the oceanographic characteristics of the study area, in par-
ticular the patterns of phytoplankton blooms and summer sea
ice extent. Phytoplankton blooms were quantified using the
merged satellite measurements of ocean chlorophyll-a data
derived from the moderate resolution imaging spectroradi-
ometer (MODIS), in orbit on theAqua platform, and the Sea-
viewing Wide Field-of-View Sensor (SeaWIFS), in orbit on
the OrbView-2. We used the level 3 monthly merged chlo-
rophyll product (concentration of chlorophyll-a in mg/m3),
with a spatial resolution of 9 km2. The data were obtained
from the Ocean Biology Processing Group (OBPG), Global
Change Data Center, Earth Sciences Division, Science and
Exploration Directorate, Goddard Space Flight Center,
NASA (http://oceancolor.gsfc.nasa.gov) in hierarchical data
format and converted to ASCII format using ESRI Arc-
GIS10.0. We traced a circular buffer around each colony,
with radius (50 km) equal to the maximum Adelie foraging
distance (Trivelpiece et al. 1987). We calculated the mean
values for chlorophyll-a concentration over a period of
10 years for the month of January, in the over-water portion
of the buffer. Due to small sample size and non-normality
in the within-group distribution of chlorophyll-a concen-
tration, differences between Adelie penguin colonies
increasing in abundance and decreasing in abundance were
assessed using a permutation test (n = 999 random permu-
tations of the values between the “increasing” and
“decreasing” categories).
Sea ice spatiotemporal distribution was estimated from
NIMBUS-7 Scanning Multichannel Microwave Radiome-
ter (SMMR) and the Defense Meteorological Satellite
Programs (DMSP) Special Sensor Microwave/Imager
(SSM/I) and the Special Sensor Microwave Imager/Soun-
der (SSMIS) passive microwave data (25 km2 spatial
resolution). The data were obtained from the National
Snow and Ice Data Center in raw binary format and con-
verted to ASCII format using ESRI ArcGIS 10.0. We
calculated the minimum sea ice concentration for the
month of December for the same buffers described above
from 1978 to 2011.
Results
Field survey
The list of sites visited along with the presence–absence
summary for each site (describing the status of the colony
if possible) is presented in Table 1. Sites listed in Table 1
were surveyed extensively on the ground; under these
conditions, the probability of detection for penguins and
shags approaches 100 % and thus absence in Table 1
should be interpreted as true absence at the time of the
survey. Maps showing the track of the yacht expedition and
all sites are shown in Fig. 1, and maps of the survey sites
requiring further detail are presented in Online Resource 1.
The numbers on the maps correspond to the “Map ID”
numbers on the tables. The distribution of colonies by
species is shown in Online Resource 2. Photodocumenta-
tion of all sites surveyed is available on request; we hope
that our efforts to document these infrequently surveyed
seabird colonies will be useful for future survey work in
this area.
No breeding gentoos were observed south of Cape
Tuxen (site 11 in Table 1 and Online Resource 2). This
represents the most southerly breeding colony for this
species. Comparing our data with the 2010/2011 survey
data reported in Pilipenko (2013), we find that the Cape
Tuxen colony increased in the 2 years separating the sur-
veys. We report on the first complete site censuses of the
three gentoo penguin breeding colonies in the Lemaire
Channel (Glandaz Point, Loubat Point, and Humphries
Heights). Compared with historic data from these sites
(Poncet and Poncet 1987; Woehler 1993), we find that
gentoo populations in the Lemaire Channel are not
declining (cf. Lynch et al. 2012) but rather, like other
gentoo colonies in the Peninsula, are either stable or
increasing in abundance. Table 2 lists the counts of gentoo
penguin colonies by this survey, as well as previous data
that allowed for the estimation of population changes.
Breeding chinstrap penguins were found as far south as
Emperor Island (67°51′S). The breeding pairs found at
Emperor Island, Cone Island and the Islet north east of
Weller Island are recorded here for first time. Chinstrap
penguins were always found in very small groups (mostly
in groups of two breeding pairs, with a maximum of 21
breeding pairs at Point Charcot, Booth Island) or as single
pairs. Table 3 lists the counts of chinstrap penguins for the
sites where they were present.
All colonies of Adelie penguins north of Marguerite
Bay, with the exception of the colonies next to Winkle
Island, declined since the 1980s (Table 4). However, col-
onies in Marguerite Bay did not match this pattern, and
abundances were found to be similar in size or larger than
reported by Poncet and Poncet (1987).
Polar Biol
123

Table 1 Sites surveyed, describing the status of the colonies if available
Map ID Site Latitude Longitude Gentoo Chinstrap Adelie Blue-eyed shags
1 Pursuit Point (Wiencke I.) −64.905 −63.434 I A A D
2 Truant I. −64.915 −63.411 I A A Pb
3 Loubat Point −65.061 −63.9061 I A A A
4 Humphries Heights −65.065 −64.033 I A A Pb
5 Port Charcot (Booth I.) −65.065 −64.033 I I D Pb
6 Glandaz Point −65.091 −63.957 I A A A
7 Petermann I. −65.167 −64.143 I Pa D Pb
8 Moot Point −65.204 −64.075 I A A A
9 Penguin Point (Galindez I.) −65.248 −64.240 I A A A
10 Yalour Is. −65.254 −64.133 I A D A
11 Cape Tuxen −65.268 −64.124 I A A A
12 Green Island −65.322 −64.153 A A A P
13 Berthelot Is. −65.332 −64.116 A A D A
14 Rocks west of Trundle I. −65.399 −65.300 A Pa P D
15 Jingle Is. −65.438 −65.344 A S D P
16 Weller I. −65.442 −65.364 A A D A
17 Islet southeast of Weller I. −65.451 −65.346 A A D A
18 Smiggers I. −65.459 −65.347 A S D A
19 Islet north east of Fitzkin I. −65.513 −65.479 A A D P
20 Two islets west of Winkle I. −65.519 −65.686 A Pa S P
21 Vieugue I. −65.650 −65.225 A A D A
22 Islets south of Martin I. −65.687 −65.335 A A D P
23 Islet next to Lacktinov I. −65.758 −65.791 A A P P
24 Islets near Armstrong I. −65.889 −66.196 A S D P
25 Fish Is. −66.029 −65.381 A A D I
26 Islet 1 km west of Lavosier I. −66.120 −66.832 A A D P
27 Cape Evensen −66.152 −65.709 A A D A
28 Darbel Is. −66.373 −65.870 A A D P
29 Kuno Point −66.402 −67.168 A A P P
30 Barcroft Is. −66.461 −67.139 A A D P
31 Mist Rocks −66.800 −66.624 A A A P
32 Holdfast Point −66.809 −66.563 A A D P
33 Detaille I. −66.870 −66.786 A A D A
34 Andressen Island (west) −66.874 −66.709 A A D P
35 Andressen Island (east) −66.878 −66.648 A A D P
36 Chatos I. −67.660 −69.152 A A S P
37 Cone I. −67.674 −69.153 A P S P
38 Bongrain Point (Pourquoi Pas I.) −67.722 −67.732 A A I A
39 Ginger I. −67.749 −68.685 A A I I
40 Avian I. −67.769 −68.888 A A S A
41 Emperor I. −67.863 −68.708 A P I P
42 Lagotellerie I. −67.888 −67.379 A A I P
43 Red Rock Ridge −68.291 −67.186 A A I P
I increasing, D decreasing, S stable, A absent, P present—no data available for estimating statusa Intermittently occupied by a single nestb From Lynch et al. (2013)
Polar Biol
123

The distribution of blue-eyed shag colonies follows
closely the distribution of the penguin colonies, sharing
most sites with one or more species of penguins in this
survey (Table 5). Only at Mist Rocks and Green Island
were blue-eyed shags found to be breeding by themselves.
Environmental drivers of Adelie penguin population
change
We find a relationship between the persistence (as fre-
quency across years) of phytoplankton blooms and the
population trajectories of Adelie penguin populations; in
the vicinity of Marguerite Bay, phytoplankton blooms are
more persistent, and Adelie penguin populations have
either maintained their size or grown over the last
30 years, whereas chlorophyll-a poor waters north of
Marguerite Bay with smaller (in spatial extent), less fre-
quent phytoplankton blooms host Adelie penguin
populations that are in decline (Figs. 2, 3). There is a
statistically significant (p \ 0.001) difference in mean
chlorophyll-a between those colonies increasing in abun-
dance and those decreasing in abundance, though it is
important to recognize that the strong spatial autocorre-
lation in both abundance trends and chlorophyll-a make it
difficult to interpret the true statistical significance of this
comparison.
Table 2 Gentoo penguin (Pygoscelis papua) colonies surveyed and references to data collected in previous years by other authors
Map ID Locality Count Accuracy Year Reference
1 Pursuit Point 200 N3/4 1986 Poncet and Poncet (1987)
567 N1 2012 This paper
2 Truant I. 717 C1+ 1986 Poncet and Poncet (1987)
921 N1 2012 This paper
3 and 4 Northern Lemaire (Loubat Point and
Humphries Heights combined)
238 N1 1982 Poncet and Poncet (1987)
237 N1 2010 Pilipenko (2013)
626 N1 2012 This paper
5 Port Charcot (Booth I.) 318 N1 1982 Poncet and Poncet (1987)
1,074 C1 2010 Central Lemaire in Pilipenko (2013)
1,484 N1 2012 This paper
6 Glandaz Point 75 N1 1982 Poncet and Poncet (1987)
115 N1 2010 “Central Lemaire” in Pilipenko (2013)
145 N1 2012 This paper
7 Petermann I. 75 N3 1909 Gain (1914)
480 N3 1971 Muller-Schwarze and Muller-Schwarze (1975)
2,148 N1 2010 Pilipenko (2013)
3,300 N1 2012 This paper
8 Moot Point 74a N1 2005 Lynch et al. (2013)
389 N1 2010 Pilipenko (2013)
479 N1 2012 This paper
9 Penguin Point (Galindez I.) 93a N1 2008 Lynch et al. (2013)
238 N1 2010 Pilipenko (2013)
273 N1 2012 This paper
10 Yalour Is. 0 N1 1982 Poncet and Poncet (1987)
24a C1 2010 Pilipenko (2013)
28 N1 2012 This paper
11 Cape Tuxen 197a N1 2010 Foothill of Mount Demaria in Pilipenko (2013)
282 N1 2012 This paper
“Year” refers to the year at the beginning of the austral summer (e.g., the survey here reported occurred during the 2012/2013 field season and is
listed as 2012). In most cases, we have restricted Table 2 to counts available from Poncet and Poncet (1987) (or earlier references where
available) and the current survey. Counts from Pilipenko (2013) are also included, where available, as this reference may not be widely available
we would like to clarify how the site names contained therein match up with our current naming scheme. Additional survey counts are available
in Lynch et al. (2013). We have referred to all counts as either nest counts (N), chick counts (C), or adult counts (A), interpreting the intent of the
original survey reports as appropriate, with precision reported as follows: (1) error less than ±5 %, (2) ±5–10 %, (3) ±10–25 %, (4) ±25–50 %a These colonies had never been recorded as existing prior to the survey indicated
Polar Biol
123

Satellite data on sea ice extent show that sea ice con-
centration in the Marguerite Bay area has been declining in
the last 30 years. More importantly, perhaps, the frequency
of years in which the areas surrounding the Marguerite Bay
colonies is free of sea ice in December has increased in the
last 20 years (Fig. 4).
Discussion
Despite a concerted search effort for new gentoo penguin
colonies, we did not find any colonies south of Cape
Tuxen. Our results coincide with the data collected by
Pilipenko (2013), which also confirmed the breeding pop-
ulation at Cape Tuxen. All gentoo colonies for which it was
possible to estimate population change showed a signifi-
cant increase in their breeding abundance, consistent with
the general trend of gentoo penguin population growth
along the WAP (Lynch et al. 2012).
Despite widespread population declines along most of
the WAP, we found that small numbers of chinstrap pen-
guins south of 65°S have persisted (Table 3). We report
here for the first time chinstrap penguins breeding in small
numbers as far south as Cone Island and Emperor Island,
the latter of which may now be the southern-most breeding
population [given the apparent disappearance of the colony
at Peter I Island; see Woehler (1993)]. There are significant
environmental and sea ice phenology differences between
these southern-most chinstrap occupied islands and the
South Sandwich Islands where chinstrap penguins are most
abundant. It remains to be seen whether these southern
WAP chinstrap penguins differ from their more northern
conspecifics with respect to breeding phenology, migration
strategy, or foraging strategies. While chinstrap penguins
are apparently able to successfully breed along the southern
WAP, the biotic and abiotic constraints on greater expan-
sion of chinstrap colonization and persistence remain an
open question. We speculate that the rates of extinction for
these small, isolated groups [as well as on other extremely
isolated islands such as Peter I Island, Heard Island, and the
Balleny Islands; see Woehler (1993)], would be high
absent at least occasional migration from more abundant
populations at the core of their breeding range, but the rates
of these processes are unknown. Given the lack of physi-
ological (or obvious ecological) barriers for breeding this
far south, it is not clear why chinstraps are not more
abundant in this region.
Relatively little is known about the abundance and
distribution of blue-eyed shags in the area covered by this
survey, and we have only sparse historical data for these
populations to assess how the population might have
changed in the last few decades. At two of the more
Table 3 Chinstrap penguin (Pygoscelis antarctica) colonies surveyed and references to data collected in previous years by other authors
Map ID Locality Count Accuracy Year Reference
5 Port Charcot (Booth I.) 3 C1 1982 Poncet and Poncet (1987)
21 N1 2010 Pilipenko (2013)
22 N1 2012 This paper
7 Petermann I. 1 N1 2010 Pilipenko (2013)
0 N1 2012 This paper
14 Rocks west of Trundle I. 1 N1 2012 This paper
15 and 18 Pitt Is. North (Jingle I. and
Smiggers I. combined)
0 N1 1983 Poncet and Poncet (1987)
4 N1 2012 This paper
20 Two islets next to Winkle I. 0 N1 1982 Pitt Islands South in Poncet and Poncet (1987)
1 N1 2012 This paper
24 Islets near Armstrong I. 1 N1 1983 Poncet and Poncet (1987)
6 N1 1990 Woehler (1993)
1 N1 2012 This paper
37 Cone I. 2 N1 2012 This paper
41 Emperor I. 1 N1 2012 This paper
“Year” refers to the year at the beginning of the austral summer (e.g., the survey here reported occurred during the 2012/2013 field season and is
listed as 2012). In most cases, we have restricted Table 2 to counts available from Poncet and Poncet (1987) (or earlier references where
available) and the current survey. Counts from Pilipenko (2013) are also included, where available, as this reference may not be widely available
we would like to clarify how the site names contained therein match up with our current naming scheme. Additional survey counts are available
in Lynch et al. (2013). We have referred to all counts as either nest counts (N), chick counts (C), or adult counts (A), interpreting the intent of the
original survey reports as appropriate, with precision reported as follows: (1) error less than ±5 %, (2) ±5–10 %, (3) ±10–25 %, (4) ±25–50 %
Polar Biol
123

Table 4 Adelie penguin (Pygoscelis adeliae) colonies surveyed and references to data collected in previous years by other authors
Map ID Locality Count Accuracy Year Reference
5 Port Charcot (Booth I.) 500 A5 1909 Croxall and Kirkwood (1979)
83 C1 1982 Poncet and Poncet (1987)
9 C1 2010 Pilipenko (2013)
15 N1 2012 This paper
7 Petermann I. 925 A3 1909 Croxall and Kirkwood (1979)
1,540 N3 1971 Muller-Schwarze and Muller-Schwarze (1975)
987 N1 1982 Airey (1982), in Poncet and Poncet (1987)
251 N1 2010 Pilipenko (2013)
283 N1 2012 This paper
10 Yalour Is. 10,400 N1, N3 1958 Croxall and Kirkwood (1979)
5,865 N1 1982 Airey (1982), in Poncet and Poncet (1987)
2,555 N1 2010 Pilipenko (2013)
2,411 N1 2012 This paper
13 Berthelot Is. 904 N1 1982 Airey (1982), in Poncet and Poncet (1987)
346 N1 2010 Pilipenko (2013)
313 N1 2012 This paper
14 Rocks west of Trundle I. 347 N1 2012 This paper
15, 16, 17 and 18 Pitt Is. North (Jingle I., Weller I.,
Islet southeast of Weller I., and
Smiggers I. combined)
3,424 + 350 C1 + N3/4 1983 Poncet and Poncet (1987)
1,059 N1 2012 This paper
19 Islet north east of Fitzkin I. 250 N3/4 1983 Poncet and Poncet (1987)
113 N1 2012 This paper
20 Two islets next to Winkle I. 52 C1 1982 Pitt Is. south in Poncet and Poncet (1987)
32 N1 2012 This paper
21 Vieugue I. 600 N4 1986 Poncet and Poncet (1987)
359 N1 2012 This paper
22 Islets south of Martin I. 1,277 C1 1982 “Kim Is.” in Poncet and Poncet (1987)
477 N1 2012 This paper
23 Islet next to Lacktinov I. 38 N1 2012 This paper
24 Islets near Armstrong I. 9,626 + 700 C1 + N 3/4 1983 Poncet and Poncet (1987)
2,813 C1 2012 This paper
25 Fish Is. 4,000 N3/4 1983 Poncet and Poncet (1987)
912 N1 2012 This paper
26 Islet 1 km west of Lavosier I. 150 N4/5 1986 Poncet and Poncet (1987)
17 N1 2012 This paper
27 Cape Evensen (including
penguins on offshore rocks)
1,200 N3/4 1983 Poncet and Poncet (1987)
1,100 N3/4 1990 Woehler (1993)
374 N1 2012 This paper
28 Darbel Islands 650 N3/4 1983 Poncet and Poncet (1987)
324 N1 2012 This paper
29 Kuno Point 235 N1 2012 This paper
30 Barcroft Is. 1,600 N4/5 1982 Poncet and Poncet (1987)
211 N1 2012 This paper
32 Holdfast Point 625 N3/4 1983 Poncet and Poncet (1987)
435 N1 2012 This paper
Polar Biol
123

northern sites in our survey area (Pursuit Point and Rocks
west of Trundle Island), shag populations appear to be
decreasing, while at two of the sites further south (Fish
Islands and Ginger Island), they appear to be stable or even
increasing. This pattern is consistent with the general
north–south trend seen for Adelie penguins, though con-
tinued observation of blue-eyed shag populations will be
required to understand their dynamics in the context of
underlying environmental conditions. Using the estimated
global population of 10,900 breeding pairs by Orta (1992)
as reported by Casaux and Barrera-Oro (2006), our survey
covered approximately 25 % of the global population of
Phalacrocorax (atriceps) bransfieldensis.
Primary productivity strongly correlated with trends
in Adelie abundance
Persistent phytoplankton blooms in Marguerite Bay are
congruent with population numbers for Adelie populations
that are stable or increasing, suggesting a relationship
between persistent primary productivity and maintenance
of Adelie populations. Referencing previous surveys in this
region (Poncet and Poncet 1987; Woehler 1993) and the
analyses reported in Lynch et al. (2012), it is clear that all
Adelie populations from the Yalour Islands south to Mar-
guerite Bay have declined, some by as much as 75 %. By
contrast, the Adelie populations at eight open-ocean-facing
Table 4 continued
Map ID Locality Count Accuracy Year Reference
33 Detaille I. 10,000 A5 1908 Croxall and Kirkwood (1979)
350 N3 1957 Croxall and Kirkwood (1979)
858 C1 1985 Poncet and Poncet (1987)
561 C1 2012 This paper
34 Andresen I. (northwest coast) 2,200 B3/4 1983 Poncet and Poncet (1987)
1,104 C1 2012 This paper
35 Andresen I. (northeast coast) 281 N1 2012 This paper
36 Chatos I. 100 N4/5 1982 Poncet and Poncet (1987)
135 N1 2012 This paper
37 Cone I. 3,000 N4/5 1982 Poncet and Poncet (1987)
3,051 N1 2012 This paper
38 Bongrain Point (Pourquoi Pas I.) 700 N3/4 1983 Poncet and Poncet (1987)
2,284 N1 2012 This paper
39 Ginger I. 2,790 C1, C3 1982 Poncet and Poncet (1987)
3,334 C1 2012 This paper
40 Avian I. 25,000 A4 1968 Croxall and Kirkwood (1979)
35,600 N2 1978 Poncet and Poncet (1987)
77,515a N (unknown
precision)
2012 Sailley et al. (2013)
47,146 C1 2012 This paper
41 Emperor I. 700 N4/5 1985 Poncet and Poncet (1987)
1,420 C1 2012 This paper
42 Lagotellerie I. 400 N3 1936 Croxall and Kirkwood (1979)
2,402 N1 1955 Croxall and Kirkwood (1979)
1,000 N3 1960 Croxall and Kirkwood (1979)
1,720 C1 1982 Poncet and Poncet (1987)
7,482 C1 2012 This paper
43 Red Rock Ridge 540 N3 1936 Croxall and Kirkwood (1979)
900 N3 1960 Croxall and Kirkwood (1979)
1,820 C1 1983 Poncet and Poncet (1987)
3,760 C1 2012 This paper
A complete list of abundance estimates for many of these sites may be found in Lynch et al. (2013). We have referred to all counts as either nest
counts (N), chick counts (C), or adult counts (A), interpreting the intent of the original survey reports as appropriate, with precision reported as
follows: (1) error less than ±5 %, (2) ±5–10 %, (3) ±10–25 %, (4) ±25–50 %a There is a significant disparity between two surveys of Avian Island in 2013; it is not clear whether the difference stems from differing survey
targets (nests vs. chicks), a massive loss in the number of chicks between the two surveys, or another unidentified difference in survey protocols
Polar Biol
123

sites in Marguerite Bay (Chatos Island, Cono Island, Gin-
ger Islands, Bongrain Point at Pourquois-Pas Island, Red
Rock Ridge, Lagotellerie Island, Emperor Island, and
Avian Island) appear to be maintaining their size or
increasing, consistent with other research suggesting that
northern Marguerite Bay is a biological “hotspot” with
high occurrence rates of krill predators (Friedlaender et al.
2011).
The Marguerite Bay area is of particular interest because
it has unique oceanographic characteristics (e.g., Schofield
et al. 2013) and frequently hosts the largest and most
persistent phytoplankton blooms on the WAP (Marrari
et al. 2008). Spring and summer phytoplankton blooms are
an important factor influencing Antarctic krill (Euphausiasuperba) recruitment in the Antarctic Peninsula region
(Marrari et al. 2008), which is widely recognized as a
major link between primary producers and many popula-
tions of krill-feeding vertebrates, including penguins
(Ballerini et al. 2014). It has been shown that there is an
alongshore gradient of phytoplankton biomass in the
Table 5 Blue-eyed shag (Phalacrocorax [atriceps] bransfieldensis) colonies surveyed
Map ID Locality Count Accuracy Year Reference
1 Pursuit Point 140 Unknown 1986 Harris et al. (2011)
21 N1 2012 This paper
12 Green I. 89 N1 2012 This paper
14 Rocks west of Trundle I. 140 Unknown 1989 Harris et al. (2011)
71 N1 2012 This paper
15 Jingle Is. 7 N1 2012 This paper
19 Islet north east of Fitzkin I. 40 N1 2012 This paper
20 Two islets west of Winkle I. 38 N1 2012 This paper
22 Islets south of Martin I. 24 N1 2007 Lynch et al. (2013)
32 N1 2012 This paper
23 Islet next to Lacktinov I. 60 N1 2012 This paper
24 Islets near Armstrong I. 114 N1 2012 This paper
25 Fish Is. 78 C1 1994 Lynch et al. (2013)
31 N1 2002 Lynch et al. (2013)
55 N1 2007 Lynch et al. (2013)
137 C1 2007 Lynch et al. (2013)
65 C1 2008 Lynch et al. (2013)
78 N1 2012 This paper
26 Islet 1 km west of Lavosier I. 39 N1 2012 This paper
28 Darbel Is. 39 N1 2012 This paper
29 Kuno Point 15 N1 2012 This paper
30 Barcroft Is. 277 N1 2012 This paper
31 Mist Rocks 17 N1 2012 This paper
32 Holdfast Point 45 N1 2012 This paper
34 and 35 Andressen I. 30 N1 2012 This paper
36 Chatos I. 10 N1 2012 This paper
37 Cone I. 130 N1 2012 This paper
39 Ginger I. 275 Unknown 1980s Harris et al. (2011)
504 N1 2012 This paper
41 Emperor I. 810 N1 2012 This paper
42 Lagotellerie I. 270 N1 2012 This paper
43 Red Rock Ridge 3 N1 2007 Lynch et al. (2013)
19 C1 2009 Lynch et al. (2013)
9 C1 2012 This paper
A complete list of abundance estimates for many of these sites may be found in Lynch et al. (2013). Green Island was surveyed from an offshore
zodiac. We have referred to all counts as either nest counts (N), chick counts (C), or adult counts (A), interpreting the intent of the original survey
reports as appropriate, with precision reported as follows: (1) error less than ±5 %, (2) ±5–10 %, (3) ±10–25 %, (4) ±25–50 %
Polar Biol
123

northern part of the Antarctic Peninsula, with higher bio-
mass in the northern sectors earlier in the productive
season, which later progresses to the southeast as the sea
ice retreats (Smith et al. 1998; Garibotti et al. 2003). In this
area, intermediate values of chlorophyll-a were observed
(0.1–2 mg/m3) with small spatial extent, short-lived blooms
(chlorophyll values greater than 2 mg/m3 in some near
shore regions, Marrari et al. 2008). However, the Mar-
guerite Bay area showed persistently high chlorophyll
concentrations (0.1–47 mg/m3) during spring and summer
in comparison with any other sector west of the Antarctic
Peninsula (Marrari et al. 2008). Reports on chlorophyll-aconcentrations in the Marguerite Bay area vary from
2.30 mg/m3 (Arrigo and van Dijken 2003) to up to 17.86
DecreasingIncreasing
No changeNo data
201120102009
200820072006
200520042003
Low High
Chlorophyll-a concentration
A
B
N
Fig. 2 Adelie penguin
population trends along with the
distribution of chlorophyll-aconcentration (available from:
http://oceancolor.gsfc.nasa.
gov). The dotted line indicates
the approximate location of the
transition zone described in the
text. Note that the color scale is
a stretched scale based on stan-
dard deviation and is therefore
not a linear representation of
relative chlorophyll-a values.
a Mean January chlorophyll-aconcentration (2003–2014),
b mean January chlorophyll-aconcentration for each year
from 2003 to 2011. Note the
persistence of the phytoplankton
blooms in the Marguerite Bay
area
Polar Biol
123

and 25 mg/m3 (Garibotti et al. 2003; Meyer et al. 2003,
respectively).
It is interesting to compare our findings with Montes-
Hugo et al. (2009), who find an unambiguous demarcation
between areas of decreasing chlorophyll-a (northern WAP)
and increasing chlorophyll-a (southern WAP). At the
broadest scale, our findings are consistent with this picture.
However, Montes-Hugo et al. (2009) found that chloro-
phyll-a has increased 66 % on average from 1978–1986 to
1998–2006 between Marguerite Bay and south of Anvers
Island (63.8°–67.8°S, 64.4°–73.0°W)—an area where
Adelie penguin populations have been declining in the last
decades. We propose two possible explanations for this
paradox. One is suggested by the fact that the increase in
chlorophyll-a in this region is driven by high chlorophyll
values during only 2 years, and it may be that a change in
the frequency of years with persistently high concentration
in the phytoplankton blooms over time may be more rel-
evant to penguin population trends than an average trend
on phytoplankton concentration skewed by infrequent high
blooms (cf. Fraser and Hofmann 2003). It may also be the
case that this ecological boundary continues to move
southward, so the line between increasing and decreasing
chlorophyll-a concentrations may have shifted further
south since 2006 [the last year included in the analysis by
Montes-Hugo et al. (2009)].
Summer sea ice declines improves colony access
In the last 30 years, summer sea ice extent has been
declining in Marguerite Bay (Fig. 4). Our analysis of
summer sea ice in the proximities of penguin colonies
showed that summer sea ice has declined at all sites,
especially since the late 1980s. The Marguerite Bay area
has essentially shifted from a regime of extensive summer
sea ice to less extensive summer sea ice, including years
with no summer sea ice (Fig. 4). Several studies have
highlighted that in breeding seasons with reduced summer
sea ice adjacent to rookeries Adelie penguins are able to
use fat reserves and food for breeding purposes (egg pro-
duction, courtship, incubation and chick feeding) instead of
using those resources for traveling to reach open water (at
Cape Royds, Stonehouse 1963; at Cape Crozier, Ainley and
Leresche 1973; in the Ross Sea area, Taylor et al. 1990).
Highlighting the potentially negative consequences of
heavy summer sea ice, Croxall and Kirkwood (1979) noted
that during the 1958–1959 breeding season in Detaille
Island, penguins had to walk nearly 50 km to reach open
water, which led to almost complete nest failure in that
year.
Our survey data suggest that Adelie penguin populations
flourish where persistently high primary productivity
occurs in areas without extensive summer sea ice that
might block access to the colony during the austral spring.
In areas north of Marguerite Bay, loss of sea ice has
increased access to the colony for breeding but at the
expense of reduced food availability for krill specialist
predators (Trivelpiece et al. 2011). We believe that Mar-
guerite Bay benefits from consistently high primary
productivity, in part due to its unique hydrogeography
(Ashjian et al. 2004), and this mitigates regional losses of
krill habitat due to declining annual sea ice cover. At the
No data Decreasing No change Increasing
0.8
1.0
1.2
1.4
1.6
1.8
Change in Adelie penguin colony abundance
chlo
roph
yll -
a (m
g/m
)3
Difference in group means
Freq
uenc
y
−0.2 0.0 0.2 0.4
5010
015
0data
permuted0
Fig. 3 Box plot showing January chlorophyll-a concentration (avail-
able from: http://oceancolor.gsfc.nasa.gov) around the colonies
grouped by the population status of the colonies. Boxes extend to the
first and third quartiles and whiskers extend to the extrema
(points[1.5 times the inter-quartile range from the box represented
by circles). Inset mean difference between the “increasing” and
“decreasing” group means under random permutation of the group
labels (n = 999). Arrow indicates the empirical difference in group
means, which is larger than any of the 999 random permutations
(hence, p\ 0.001)
Min
imum
sea
−ice
con
cent
ratio
n (%
)
1980 1985 1990 1995 2000 2005 2010
C
Year
0
40
80
1980 1985 1990 1995 2000 2005 2010
B
0
40
80
1980 1985 1990 1995 2000 2005 2010
A
0
40
80 p = 0.02R = 0.14
p = 0.003R = 0.23
p < 0.001R = 0.28
2
2
2
Fig. 4 Minimum sea ice concentration for December around the
Marguerite Bay colonies. (Sea ice data available from the National
Snow and Ice Data Center: http://nsidc.org/). a Cone Island and
Chatos Island; b Avian Island, Ginger Island and Emperor Island; and
c Bongrain Point (Pourquoi Pas Island), Lagotellerie Island and Red
Rock Ridge. The dotted line shows the general trend of sea ice con-
centration for the sites
Polar Biol
123

same time, decreasing sea ice is likely allowing Adelie
penguins to recoup any reduction in prey availability by
reductions in net reproductive energy, with reduced for-
aging effort and greater efficiencies due to access to open
water. It is possible that continued reduction in sea ice will
exceed the threshold where the net benefit of access to the
colony is outweighed by concomitant loss of prey.
While the biggest gains in Adelie penguin abundance in
the Marguerite Bay area have occurred between the mid-
1980s and the 2012/2013 season, it is interesting to note
that these populations have been increasing at least since
the 1940s. For example, Croxall and Kirkwood (1979)
noted that “a real increase in numbers [at Lagotellerie
Island] had taken place by the late 1940s–early 1950s”.
Therefore, while recent gains in abundance may be
explained by improving sea ice conditions (from the per-
spective of access to breeding colonies), increases earlier in
the twentieth century prior to increasing air temperatures
and declining sea ice (King and Turner 1997; Smith and
Stammerjohn 2001) are not easily explained by abiotic
drivers. These increases may reflect a response to har-
vesting of other krill predators (e.g., seals and whales), as
reviewed by Ainley et al. (2009), though the data on pen-
guin abundance we have for this region are insufficient to
make any firm conclusions in this regard.
While logistical difficulties in accessing the southern
portion of the WAP make it difficult to census these pop-
ulations regularly, it is clear that Marguerite Bay and areas
just north are not just static “predator hotspots” but rep-
resent the leading edge of climatic changes that can bring
transient benefits to species such as penguins. Whether
Marguerite Bay will remain a refugium for Adelie pen-
guins on the WAP in the long term remains to be seen, but
clearly this is a region deserving of focused attention over
the next several decades.
Acknowledgments We gratefully acknowledge the support of The
Tinker Foundation, whose support enabled Oceanites, Inc. and the
Antarctic Site Inventory team to charter Golden Fleece for this
directed research, and in addition, thank the U.S. National Science
Foundation Office of Polar Programs and Geography and Spatial
Sciences (Awards Nos. 07-39515 and 12-55058). We also acknowl-
edge the support of NASA headquarters under the NASA Earth and
Space Fellowship Program—Grant NNX10AN55H to P. C. We are
grateful to the crew of the yacht Golden Fleece, Yoann Gourdet and
Cathy Colle, for ensuring the safety and well-being of the Antarctic
Site Inventory research team during their work period in the Antarctic
Peninsula. Cheesemans’ Ecology Safaris and One Ocean Expeditions,
which graciously provided transportation to and from the Peninsula.
References
Ainley DG, Leresche RE (1973) The effects of weather and ice
conditions on breeding in Adelie Penguins. Condor 75:235–239
Ainley D, Ballard G, Blight LK, Ackley S, Emslie SD, Lescroel A,
Olmastroni S, Townsend SE, Tynan CT, Wilson P, Woehler E
(2009) Impacts of cetaceans on the structure of Southern Ocean
food webs. Mar Mamm Sci 26:482–498
Airey LR (1982) General report, 11 March 1982–28 February 1983.
AD6/2F/1982/A. British Antarctic Survey unpublished report
Arrigo KR, van Dijken GL (2003) Phytoplankton dynamics within 37
Antarctic coastal polynya systems. J Geophys Res 108:3271
Ashjian CJ, Rosenwaks GA, Wiebe PH, Davis CS, Gallager SM,
Copley NJ, Lawson GL, Alatalo P (2004) Distribution of
zooplankton on the continental shelf off Marguerite Bay,
Antarctic Peninsula, during austral fall and winter, 2001. Deep
Sea Res II 51:2073–2098
Ballerini T, Hofmann EE, Ainley DG, Daly K, Marrari M, Ribic CA,
Smith WO, Steele JH (2014) Productivity and linkages of the
food web of the southern region of the western Antarctic
Peninsula continental shelf. Prog Oceanogr 122:10–29
Casaux R, Barrera-Oro E (2006) Shags in Antarctica: their feeding
behaviour and ecological role in the marine food web. Antarct
Sci 18:3–14
Clarke A, Murphy EJ, Meredith MP, King JC, Peck LS, Barnes DKA,
Smith RC (2007) Climate change and the marine ecosystem of
the western Antarctic Peninsula. Philos Trans R Soc Lond Ser B
362:149–166
Croxall JP, Kirkwood ED (1979) The distribution of penguins on the
Antarctic Peninsula and islands of the Scotia Sea. British
Antarctic Survey, Cambridge, UK
Croxall JP, Trathan PN, Murphy EJ (2002) Environmental change and
Antarctic seabird populations. Science 297:1510–1514
Ducklow HW, Fraser WR, Meredith MP, Stammerjohn SE, Doney
SC, Martinson DG, Sailley SF, Schofield OM, Steinberg DK,
Venables HJ, Amsler CD (2013) West Antarctic Peninsula: an
ice-dependent coastal marine ecosystem in transition. Oceanog-
raphy 26:190–203
Fraser WR, Hofmann EE (2003) A predator’s perspective on causal
links between climate change, physical forcing and ecosystem
response. Mar Ecol Prog Ser 265:1–15
Fraser WR, Trivelpiece WZ, Ainley DG, Trivelpiece SG (1992)
Increases in Antarctic penguin populations: reduced competition
with whales or a loss of sea ice due to environmental warming?
Polar Biol 11:525–531
Friedlaender AS, Johnston DW, Fraser WR, Burns J, Halpin PN,
Costa DP (2011) Ecological niche modeling of sympatric krill
predators around Marguerite Bay, Western Antarctic Peninsula.
Deep Sea Res II 58:1729–1740
Gain L (1914) Oiseaux antarctiques. Deuxieme Expedition Antarc-
tique Francaise 1908–1910
Garibotti IA, Vernet M, Ferrario ME, Smith RC, Ross RM, Quetin LB
(2003) Phytoplankton spatial distribution patterns along the
western Antarctic Peninsula (Southern Ocean). Mar Ecol Prog
Ser 261:21–39
Harris CM (ed) (2006) Wildlife awareness manual: Antarctic
Peninsula, South Shetland Islands, South Orkney Islands.
Wildlife Publication No. 1. Prepared for the UK foreign and
commonwealth office and HMS endurance. Environmental
Research and Assessment, Cambridge
Harris CM, Carr R, Lorenz K, Jones S (2011) Important bird areas in
Antarctica: Antarctic Peninsula, South Shetland Islands, South
Orkney Islands—final report. Prepared for Birdlife International
and the Polar Regions Unit of the UK foreign and common-
wealth office. Environmental Research and Assessment Ltd.,
Cambridge
King JC, Turner J (1997) Antarctic meteorology and climatology.
Cambridge University Press, Cambridge
Lynch HJ, LaRue MA (2014) First global survey of Adelie penguin
populations. Auk 131:457–466
Polar Biol
123

Lynch HJ, Naveen R, Fagan WF (2008) Censuses of penguins, blue-
eyed shags, and southern giant petrel populations in the Antarctic
Peninsula. Mar Ornithol 36:83–97
Lynch HJ, Naveen R, Trathan PN, Fagan WF (2012) Spatially
integrated assessment reveals widespread changes in penguin
populations on the Antarctic Peninsula. Ecology 93:1367–1377
Lynch HJ, Naveen R, Casanovas PV (2013) Antarctic site inventory
breeding bird survey data 1994/95–2012/13. Ecology 94:2653
Marrari M, Daly KL, Hu C (2008) Spatial and temporal variability of
SeaWiFS chlorophyll a distributions west of the Antarctic
Peninsula: implications for krill production. Deep Sea Res II
55:377–392
Meyer B, Atkinson A, Blume B, Bathmann U (2003) Feeding and
energy budgets of larval Antarctic krill, Euphausia superba, insummer. Mar Ecol Prog Ser 257:167–177
Montes-Hugo M, Doney SC, Ducklow HW, Fraser W, Martinson D,
Stammerjohn SE, Schofield O (2009) Recent changes in
phytoplankton communities associated with rapid regional
climate change along the western Antarctic Peninsula. Science
323:1470–1473
Muller-Schwarze C, Muller-Schwarze D (1975) A survey of twenty-
four rookeries of pygoscelid penguins in the Antarctic Peninsula
region. In: Stonehouse B (ed) The biology of penguins.
University Park Press, Baltimore, pp 309–320
Murphy EJ, Watkins JL, Trathan PN, Reid K, Meredith MP, Thorpe
SE, Johnston NM, Clarke A, Tarling GA, Collins MA, Forcada J,
Shreeve RS, Atkinson A, Korb R, Whitehouse MJ, Ward P,
Rodhouse PG, Enderlein P, Hirst AJ, Martin AR, Hill SL,
Staniland IJ, Pond DW, Briggs DR, Cunningham NJ, Fleming
AH (2007) Spatial and temporal operation of the Scotia Sea
ecosystem: a review of large-scale links in a krill centred food
web. Philos Trans R Soc Lond Ser B 362:113–148
Naveen R, Lynch HJ (2011) Antarctic Peninsula compendium, 3rd
edn. Environmental Protection Agency, Washington, DC
Orta J (1992) Family Phalacrocoracidae (Cormorants). In: del Hoyo J,
Elliot A, Sargatal J (eds) Handbook of the birds of the world, vol
1. Lynx editions, Barcelona, pp 326–353
PilipenkoDV (2013)Colonial bird species breeding in theUAS,Akademik
Vernadsky2010–2011season.УКPAЇHCЬКИЙAHTAPКTИЧHИЙЖУPHAЛ УAЖ (Ukr Antarct J UAZH) 12:206–216
Poncet S, Poncet J (1987) Censuses of penguin populations of the
Antarctic Peninsula, 1983–87. Br Antarct Surv Bull 77:109–129
Sailley SF, Ducklow HW, Moeller HV, Fraser WR, Schofield OM,
Steinberg DK, Garzio LM, Doney SC (2013) Carbon fluxes and
pelagic ecosystem dynamics near two western Antarctic Penin-
sula Adelie penguin colonies: an inverse model approach. Mar
Ecol Prog Ser 492:253–272
Schofield O, Ducklow H, Bernard K, Doney S, Patterson-Fraser D,
Gorman K, Martinson D, Meredith M, Saba G, Stammerjohn S,
Steinberg D, Fraser W (2013) Penguin biogeography along the
West Antarctic Peninsula. Oceanography 26:204–206
Smith RC, Stammerjohn SE (2001) Variations of surface air
temperature and sea-ice extent in the western Antarctic Penin-
sula region. Ann Glaciol 33:493–500
Smith RC, Baker KS, Vernet M (1998) Seasonal and interannual
variability of phytoplankton biomass west of the Antarctic
Peninsula. J Mar Syst 17:229–243
Smith RC, Ainley D, Baker K, Domack E, Emslie S, Fraser B,
Kennett J, Leventer A, Mosley-Thompson E, Stammerjohn S
(1999) Marine ecosystem sensitivity to climate change. Biosci-
ence 49:393–404
Taylor RH, Wilson RP, Thomas BW (1990) Status and trends of
Adelie penguin populations in the Ross Sea region. Polar Record
26:293–304
Trivelpiece WZ, Trivelpiece SG, Volkman NJ (1987) Ecological
segregation of Adelie, gentoo, and chinstrap penguins at King
George Island, Antarctica. Ecology 68:351–361
Trivelpiece WZ, Hinke JT, Miller AK, Reiss CS, Trivelpiece SG,
Watters GM (2011) Variability in krill biomass links harvesting
and climate warming to penguin population changes in Antarc-
tica. Proc Natl Acad Sci USA 108:7625–7628
Woehler EJ (1993) The distribution and abundance of Antarctic and
sub-Antarctic penguins. Scientific Committee on Antarctic
Research, Cambridge
Polar Biol
123