Embed Size (px)
Citation preview

The American Journal of Clinicai Nutrition 42: AUGUST 1985, pp 214-219. Printed in USA© 1985 American Society for Clinical Nutrition
214
A comparative study of the effects of whitesugar, unrefined sugar and starch on theefficiency of food utilization and thermogenesis1’2AG Dulloo, OA Eisa, DS Miller, and J Yudkin
ABSTRACT Energy balance studies with weanling ratsand mice were carried out to investigate
the chronic effects of diets containing white sugar, unrefined sugar, and starch as sole carbohydrate
on body composition and energetic efficiency. The results show no significant differences inenergetic efficiency and in body composition of both rats and mice fed the different carbohydrate
diets. However, there is a tendency for the sugar-fed animals to have higher metabolizable energyintakes, but rarely was this sufficient to increase body energy since there were correspondingincreases in metabolic rate. These studies indicate that differences in energy balance between sugar
and starch are small and that any increase in thermogenesis that occurred in some of the groupsfed sugar-rich diets was in response to the higher energy intake rather than due to a lower efficiencyof utilization of sucrose per se. Am I Clin Nuir l985;42:2l4-219.
KEY WORDS Sugar, thermogenesis, obesity, energy balance
Introduction
The variable effects of different carbohy-drates in fattening animals have been appre-ciated for many years in animal feedingpractice. More than a century ago, Rubner(1) found that similar amounts of eitherstarch (232 g), cane sugar (234 g), or glucose(256 g) were required to replace 100 g of fatin the metabolism of a fasted dog. A fewdecades later, Keliner (2) from his studies ondomestic animals observed that while canesugar is equal to starch as a direct fuel, it isonly three-fourths as effective in laying downfat: he calculated that the formation of 100
g of fat required 403 g of starch comparedto 532 g of sucrose.
Since then, work on laboratory animalshas added to the conificting findings regardingthe effects of different carbohydrates on en-ergy metabolism and growth. While severalworkers have reported greater weight gainsin rats fed sugar in comparison with thosefed starch (3-6), others have failed to findsuch improved weight gain (7-12). Some ofthe discrepancies regarding relative growthrates of animals fed sucrose or other carbo-hydrates can, to some extent, be explained
by differences in food intake. But, even onsimilar food intakes, results are conificting.Feyder (3), pair-feeding weanling rats, foundthat the sucrose-fed animals gained moreweight than the glucose-fed rats, while othersfound no differences in body weight or bodyfat in experiments during which the animalsconsumed the same amount of calories(12, 13).
Several factors may be responsible for dif-
ferences seen in such studies. As pointed outby Bender and Damji (14), differences amongexperiments in the strain, sex, or age of theanimals and the length of time on the dietaryregime can all influence the metabolic fatesof sugar and lead to conflicting results. Fur-thermore, body weight alone may not be areliable indicator of the effects of sugar onenergy utilization since changes in body fatmay be masked by changes in body water.
‘From the Department of Nutrition, Queen ElizabethCollege, University of London, England.
2Add� reprint requests to: AG Dulloo, Department
of Nutrition, Queen Elizabeth College, University ofLondon, Campden Hill Road, London W8 7AH, En-gland.
Received July 16, 1984.Accepted for publication February 12, 1985.
by guest on June 19, 2013ajcn.nutrition.org
Dow
nloaded from

SUGAR AND ENERGY BALANCE 215
In the present investigation, we have com- I �. �
pared the effects of diets rich in refined I � � �
000
(white) sugar, unrefined sugar, and starch on � 1� e�
energy balance and body composition of I � �weanling rats and mice. In these studies, Imetabolizable energy intake was monitored Icontinuously over the whole experimental Iperiod, and energy expenditure was estimated � � ��o ‘r, ‘i,
by calculation from energy balance data and � � � 4�
also by indirect calorimetry. � � � �..,rb 00 �(‘4r’4r9
Materials and methods o
� IMale and female Sprague-Dawley weanling rats (Queen I � � �
Elizabeth College, London) and male weanling mice of I � .,� � �the CFLP strain (Anglia Laboratory Animals, Hunting- I - �
don, UK) were maintained on a stock diet (CRM, I � �Christopher Hill Group, London) for 10 days. At the Istart of each energy balance experiment, the animals � .� Iwere divided into four groups with similar mean body � � Iweight. One group was killed at the beginning to provide � I
I.. I �‘0Nthe initial carcass energy. The remaining groups were ,�g I � � �fed ad libitum isocaloric diets containing either refined � I � � ciwhite sugar, unrefined sugar (Light Muscovado, Billing- � I � 4� �tons, Ltd. UK), or corn starch as the sole carbohydrate .a I o� � u� v
C.) � �o�#{243}i..:(66%), casein protein (25%), corn oil (0.9%), mineral �mixture (4%), vitamin mixture (2%), and solka flocfiber (2%). ri
“
The mice were put three to a cage while the rats were ‘-0 Uhoused in pairs in metabolism rooms maintained at 25 #{149}
.c I oqoqo�� .�± 1#{176}Cwith a 12:12 h light/dark cycle. At the end ofevery week, the animals were transferred to clean cages; � I +� +� +� �000
all spilled food, feces, and urine were carefully scraped ‘ I � � � cioffthe bottom trays, dried to constant weight in an oven � I � ± �kept at 105#{176}C,homogenized and the energy content U Idetermined on triplicate samples using a ballistic bomb � Icalorimeter (15). Metabolizable energy (ME) intake was � Icalculated over the entire course ofeach 4-wk experiment I E
II � -�
from the gross energy content of the diet presented � �‘ ‘� .�c Iminus the energy content of spilled food, feces, and � I
urine over that period. .� I +1 +1 +1 �
At the end ofthe experiments, the animals were killed E I 0� 0� 0 U-U I ,e:,r,,#{243}
with ether, the skull and thoracic and abdominal cavitieswere incised after which the carcasses were dried in an Eoven at 105#{176}Cfor 48 h. Each dried carcass was then 0
homogenized in a grinder, and triplicate samples wereanalyzed for energy content by bomb calorimetry (15).Carcass fat was measured by the soxhlet fat extraction ci ci ci 0
+I+IHcimethod (16) and carcass protein was calculated from a E � v �general formula relating the energy derived from fat to 8 �o oo �
the total energetic value of the carcass and the energy m C’� L)derived from protein (17). Body energy gain was calcu-lated from the final body energy content minus theenergy content of the corresponding initial group killedat the start of the experiment. Energy expenditure (total i�.
heat production) over the entire experimental period was aderived from the difference between the ME intake andthe body energy gain for each individual animal. Energetic Iefficiency (gross) was calculated from the body energy ..� .n �n �ngain per unit ME intake, and net efficiency from thebody energy gain per unit ME intake above maintenance I-. iZ
by guest on June 19, 2013ajcn.nutrition.org
Dow
nloaded from
216 DULLOO ET AL
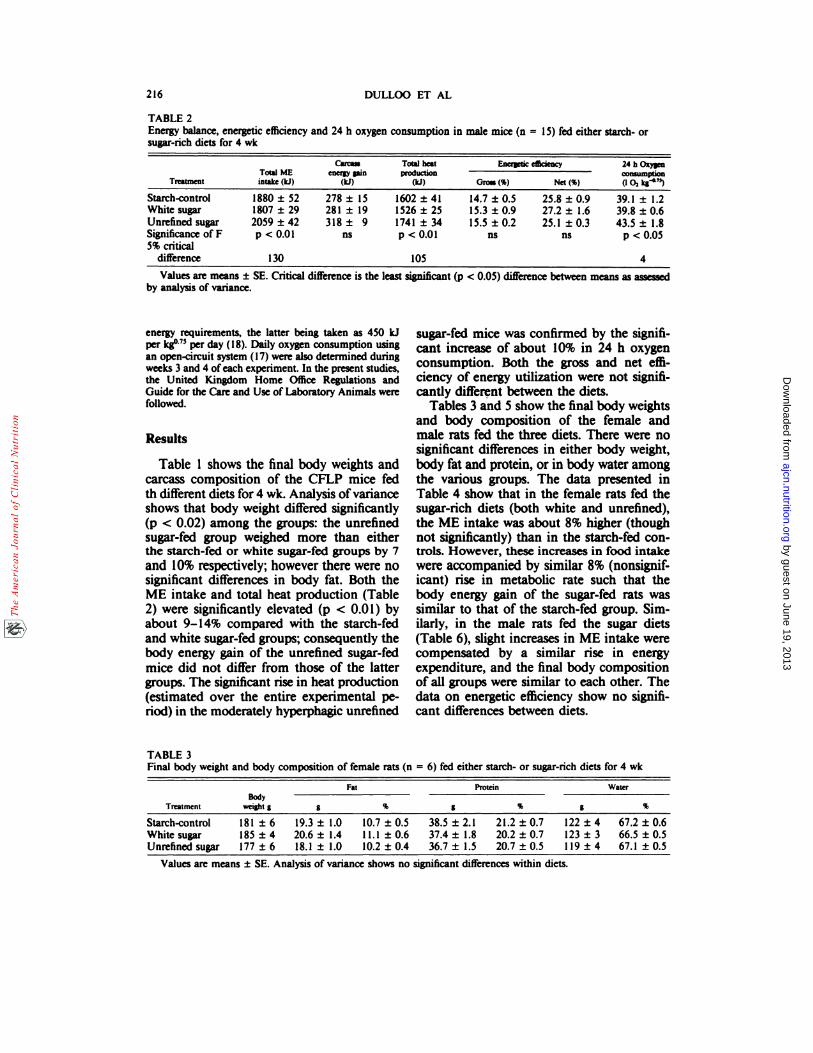
TABLE 2Energy balance, energetic efficiency and 24 h oxygen consumption in male mice (n = 15) fed either starch- orsugar-rich diets for 4 wk
TrcatmentTotal MEintake (ki)
Carcassener� gain
(ki)
Total heatproduction
(ki)
Energetic efficiency 24 h Oxygenconauznption(102 kg�”)Gross(%) Net (%)
Starch-control 1880 ± 52 278 ± 15 1602 ± 41 14.7 ± 0.5 25.8 ± 0.9 39.1 ± 1.2White sugar 1807 ± 29 281 ± 19 1526 ± 25 15.3 ± 0.9 27.2 ± 1.6 39.8 ± 0.6Unrefined sugar 2059 ± 42 318 ± 9 1741 ± 34 15.5 ± 0.2 25.1 ± 0.3 43.5 ± 1.8Significance of F5% critical
p <0.01 ns p <0.01 ns ns p <0.05
difference 130 105 4
Values are means ± SE. Critical difference is the least significant (p <0.05) difference between means as assessedby analysis of variance.
energy requirements, the latter being taken as 450 Uper kg#{176}”per day (18). Daily oxygen consumption usingan open-circuit system (17) were also determined duringweeks 3 and 4 ofeach experiment. In the present studies,the United Kingdom Home Office Regulations andGuide for the Care and Use of Laboratory Animals werefollowed.
Results
Table 1 shows the final body weights andcarcass composition of the CFLP mice fedth different diets for 4 wk. Analysis of varianceshows that body weight differed significantly(p <0.02) among the groups: the unrefinedsugar-fed group weighed more than eitherthe starch-fed or white sugar-fed groups by 7
and 10% respectively; however there were nosignificant differences in body fat. Both theME intake and total heat production (Table2) were significantly elevated (p <0.01) byabout 9-14% compared with the starch-fedand white sugar-fed groups; consequently thebody energy gain of the unrefined sugar-fedmice did not differ from those of the lattergroups. The significant rise in heat production(estimated over the entire experimental pe-nod) in the moderately hyperphagic unrefined
TABLE 3
sugar-fed mice was confirmed by the signifi-cant increase of about 10% in 24 h oxygenconsumption. Both the gross and net effi-ciency of energy utilization were not signifi-cantly different between the diets�
Tables 3 and 5 show the final body weightsand body composition of the female andmale rats fed the three diets. There were nosignificant differences in either body weight,body fat and protein, or in body water amongthe various groups. The data presented inTable 4 show that in the female rats fed thesugar-rich diets (both white and unrefined),the ME intake was about 8% higher (thoughnot significantly)than in the starch-fed con-trols.However, these increases in food intake
were accompanied by similar 8% (nonsignif-icant) rise in metabolic rate such that thebody energy gain of the sugar-fed rats wassimilar to that of the starch-fed group. Sim-ilarly, in the male rats fed the sugar diets(Table 6), slight increases in ME intake werecompensated by a similar rise in energyexpenditure, and the final body compositionof allgroups were similar to each other. Thedata on energetic efficiency show no signifi-cant differences between diets.
Final body weight and body composition of female rats (n = 6) fed either starch- or sugar-rich diets for 4 wk
Fat ProteinBody
Treatment weight g $ g I
Water
%
Starch-control 181 ± 6 19.3 ± 1.0 10.7 ± 0.5 38.5 ± 2.1 21.2 ± 0.7 122 ± 4 67.2 ± 0.6Whitesugar 185±4 20.6± 1.4 11.1±0.6 37.4± 1.8 20.2±0.7 123±3 66.5±0.5Unrefinedsugar 177±6 18.1± 1.0 10.2±0.4 36.7± 1.5 20.7±0.5 119±4 67.1±0.5
Values are means ± SE. Analysis of variance shows no significant differences within diets.
by guest on June 19, 2013ajcn.nutrition.org
Dow
nloaded from

SUGAR AND ENERGY BALANCE 217
TABLE 4Energy balance, energeticefficiency and 24 h oxygen consumption in female rats(n = 6) fed either starch- orsugar-rich diets for 4 wk
TreatmentTotal MEintake (U)
Carcassenergy gain
(U)Total heat
production (U)
Energetic efficiency 24 h Oxygenconsumption
(102 kg�”)Gross (%) Net (%)
Starch-control 5082 ± 132 1180 ± 98 3902 ± 96 23.2 ± 1.6 49.5 ± 3.1 30.5 ± 0.9White sugar 5470 ± 213 1240 ± 67 4266 ± 196 22.0 ± 1.2 44.0 ± 3.3 31.3 ± 1.2Unrefined sugar 5451 ± 128 1090 ± 70 4361 ± 137 20.0 ± 1.3 39.7 ± 2.9 29.9 ± 0.9
Values are mea ns ± SE. Analysis of variance sho ws no significant differences within diets.
Discussion
The present studies indicate that in wean-ling rats and mice, sucrose does not alterbody composition nor the efficiency of foodutilization when compared with starch.Clearly, these experiments also demonstratethat, at least in energetic terms, there doesnot seem to be any difference between theconsumption of refined (white) and unrefinedsugars, except that the latter seemed to bemore palatable to the mice studied.
Diets containing sucrose have often beenreported to produce a lower growth rate inweanling rats or a loss of body weight inadult rats.These effects occurred despite thefact that food intake was not reduced andwould therefore suggest the existence of alower efficiency of food utilization in thesesucrose-fed animals (7, 19). However, anapparent growth depressant effect of sucroseappears to occur only in animals fed sucrosediets which were inadequate in protein, andthe difference in efficiency of weight gainbetween sucrose and starch tended to disap-pear as the level of protein was increased(19). In fact, in the present studies in whichthe protein level of all the diets was relativelyhigh (25% casein), no differences in the effi-ciency of energy utilization were observed.
TABLE 5
It could be argued that the failure to findsignificant difference between the effect ofsucrose and that of starch on metabolic ratewas due partly because we used weanlinganimals that were growing at a high foodefficiency which sucrose cannot further in-crease. However other comparative studieson the effect of sucrose and starch in adultrats, although not involving measurementsof energy expenditure, nevertheless indicatedthat sucrose-rich diet does not increase foodefficiency (19, 20). There is therefore noevidence to support the suggestion that su-crose per se is utilized more efficiently thanstarch in older animals.
It is noticeable in the present studies thatin those sugar-fed groups with a tendency toconsume more calories than the starch-fedcontrols, the accompanying rise in metabolicrate prevented the otherwise expected greatergain in body energy. Similar increases inmetabolic rate (ie diet-induced thermogenesis)in hyperphagic sucrose-fed rats have alsobeen reported by Rothwell et al (11). Thesefindings therefore indicate that the increasesin voluntary food intake in sucrose-fed ani-mals does not necessarily increase the rate ofbody energy gain as reported by some otherworkers (5, 6). Such discrepancies wouldseem to be mainly due to differences in the
Final body weight and body composition of male rats (n = 4) fed either starch- or sugar-rich diets fo r 4 wk
Fat ProteinBody weight
Treatment $ $ $ $
Water
%
Starch-control 223 ± 14 22.7 ± 1.7 10.2 ± 0.2 45.0 ± 1.1 20.3 ± 0.9 148 ± 9White sugar 241 ± 10 24.2 ± 2.1 10.0 ± 0.6 46.2 ± 1.0 19.3 ± 0.8 162 ± 7Unrefined sugar 236 ± 6 24.6 ± 1.3 10.4 ± 0.6 45.3 ± 1.5 19.2 ± 0.8 158 ± 4
66.6 ± 0.f67.1 ± 1.067.0 ± 0.7
Values are means ± SE. Analysis of variance shows no significant differences within diets.
by guest on June 19, 2013ajcn.nutrition.org
Dow
nloaded from

218 DULLOO ET AL
TABLE 6Energy balance, energetic efficiency and 24 h oxygen consumption in male rats (n = 4) fed either starch- or sugar-rich diets for 4 wk
Treatment
Total MEintake (U)
Carcass energygain (U)
Total heat
production (U)
Energetic efficiency 24 h Oxygen
consumption
(102 kg�”)Gross (%) Net (%)
Starch-controlWhite sugarUnrefined sugar
5638 ± 4576162 ± 361
5811 ± 355
1400 ± 1551482 ± 63
1478 ± 136
4238 ± 3034680 ± 298
4333 ± 220
24.8 ± 0.824.1 ± 0.425.4 ± 0.8
50.9 ± 1.1
47.3 ± 0.951.2 ± 1.6
27.5 ± 2.2
29.4 ± 0.329.0 ± 1.0
Values are means ± SE. Analysis of variance shows no significant differences within diets.
genetic background of the animals used inthe various studies. In fact, strain differencesin energy deposition have been shown toresult primarily from variations in the levelofenergy expenditure in hyperphagic rats feda palatable cafeteria diet (21), and in ratsand mice fed a high-protein high-fat diet (22,23). Thus, while some strains are resistant toan extra intake of calories by exhibiting diet-induced thermogenesis others are less capableof burning off the extra calories and as aresult, develop obesity. It should be pointedout that in those studies using strains with apropensity to obesity, the increased fat de-position on a sucrose diet (5, 6, 20) probablyresulted from the increased caloric load ratherthan from a more efficient utilization ofsucrose per se. Similarly in strains resistantto obesity as in the mice fed high sucrosediet in the form of unrefined sugar, theelevated metabolic rate would seem to be inresponse to the increased energy intake ratherthan to a poorer utilization of the sucrosediet, because when the energy intake onsucrose-rich diet was similar to starch-fedcontrols, as in the case of the white sugar-fed mice, no differences in metabolic rate orenergetic efficiency were observed.
Further support for the claim that sucroseis not utilized more or less efficiently thanother carbohydrates comes from acute exper-iments in rats showing that the thermic re-sponses to sucrose were no different from theresponse to glucose and other carbohydrates(11). This contradicts previous findings of agreater thermic response to sucrose than toglucose in rats (24), but the results of thelatter studies could be criticized on the basisthat the experiments were performed onfasted rats given very small quantities ofcarbohydrates and under anesthesia. Further-
more, the metabolic rate after intubation ofsucrose tended to return to the fasting levelssooner than that occurring after intubationof other carbohydrates, thereby suggestingthat, over longer periods of investigations,the overall thermic response to sucrose mightnot have been different from that to othercarbohydrates.
Metabolic differences between starch andsugar are not universally accepted, but severalstudies have indicated that prolonged con-sumption of high sugar diets leads to increasesin blood levels of cholesterol, triglycerides,fasting insulin, and alters platelet behavior.These observations suggest that a high intakeof sugar might be a cause of coronary heartdisease and diabetes (25-27), although sucroseas an independent factor in these diseaseshas not been shown (28). In addition therehas been discussion of the possibility thatsucrose (compared to other carbohydrates) isinvolved in the current high prevalence ofobesity in affluent countries, although thepresent studies demonstrate that in weanlingrats and mice, sucrose is not utilized moreefficiently than starch. Of course, this doesnot necessarily imply that the level of sugarintake is unimportant in energy balance andobesity; further investigations involving hu-man subjects are required before the signifi-cance of these findings can be extended to asystematic evaluation of the role of a highintake of sugar in man’s dietary regime andin the etiology of human obesity. II
References
1. Rubner M. Cited by Lusk 0. In: The science ofnutrition. Philadelphia: WB Saunders Co. 1928.
2. Kellner D. Die Verfutterung der Zuckerfuttermittel.Dtsch Landw Bes Berlin, 1909.
by guest on June 19, 2013ajcn.nutrition.org
Dow
nloaded from

SUGAR AND ENERGY BALANCE 219
3. Feyder S. Fat formation from sucrose and glucose.J Nutr 1935;9:457-68.
4. Maodonald I, Grenby TH. Some differences betweencarbohydrates in their effects on weight loss andbody fat in rats. Proc Nutr Soc l979;38:30A.
5. Kanarek RB, Hirsch E. Dietary-induced overeatingin experimental animals. Fed Proc 1977:36:154-8.
6. Kanarek RB, Orthen-Gambill N. Differential effectsof sucrose, fructose and glucose on carbohydrate-induced obesity in rats. J Nutr l982;l 12:1546-54.
7. A1-Nagdy 5, Miller DS, Qureshi RU, Yudkin J.Metabolic differences between starch and sucrose.Nature 1966;209:8 1-2.
8. Bruckdorfer KR, Kari-Kari BPB, Kahn IH, YudkinJ. Activity of lipogenic enzymes and plasma triglyc-eride levels in the rat and chicken as determined bythe nature of the dietary fat and dietary carbohydrate.Nutr Metab 1972;l4:228-37.
9. Reiser S, Michaeis 0, Putney J, Hallfrisch J. Effectof sucrose feeding on the intestinal transport ofsugars in two strains of rats. J Nutr l975;l05:894-905.
10. Kang SS, Bruckdorfer KR, Yudkin J. Influence ofdifferent dietary carbohydrates on liver and plasmaconstituents in rats adapted to meal feeding. NutrMetab l979;23:30l-l5.
11. Rothwell NJ, Saville ME, Stock MJ. Acute andchronic effects of sucrose and glucose on thermogenicresponses and weight gain in the rat. Proc Nutr Socl982;41:8A.
12. Rothwell NJ, Stock Mi. Chronic effects of alcoholand sucrose consumption on energy balance andbrown adipose tissue in the rat. Proc Nutr Soc (inpress).
13. Harper AE, Katyama MC. The influence of variouscarbohydrates on the utilisation of low protein rationsby the white rat. J Nutr l953;49:261-75.
14. Bender AE, Damji KB. Some effects of dietarysucrose. World Rev Nutr Diet l972;15:l04-55.
15. Miller DS, Payne PR. A ballistic bomb calorimeter.Br i Nutr l959;13:501-8.
16. Entenman C. General procedures for separatingcomponents of tissue. In: Colowick SP, Kaplan 0,eds. Methods of enzymology, vol III, New York:Academic Press, 1957:299-317.
17. Dulloo AG, Miller DS. Energy balance followingsympathetic denervation of brown adipose tissue.Can J Physiol Pharmacol l984;62:235-40.
18. Miller DS, Payne PR. A theory of protein metabo-lism. J Theoret Biol l963;5:398-4l 1.
19. Al-Nagdy 5, Miller DS, Yudkin J. Changes in bodycomposition and metabolism induced by sucrose inthe rat.Nutr Metab 1970;l2: 193-219.
20. Williams Vi, Szepesi B. Effect of dietary carbohy-drates on food efficiency and body composition inadult male rats during normal growth and duringrecovery from food restriction. Nutr Res 1983;5:457-68.
21. Rothwell NJ, Saville ME, Stock Mi. Effects offeeding a “cafeteria” diet on energy balance anddiet-induced thermogenesis in four strains of rat. JNutr l982;112:l515�-24.
22. Miller DS. Non-genetic models of obesity. In: FestingMFW, ed. Animal models of obesity, London: Mac-millan, 1979:131-40.
23. Dulloo AG. Presence and absence of dietary-inducedthermogenesis in two strains of mice fed a high-protein high-fat diet. J Physiol l984;348:35P.
24. Sharief N, Macdonald I. The effects of respiratoryquotient and metabolic rate of glucose, fructose andsucrose in rats.Proc Nutr Soc l979;38:83A.
25. Yudkin J. Patterns and trends in carbohydrate con-sumption and their relation to disease. Proc NutrSoc 1964,23:149-62.
26. Elwood PC, Waters WE, Moore 5, Sweetnam P.Sucrose consumption and ischaemic heart disease inthe community. Lancet l970;l:10l4-6.
27. Cleave TL, Campbell GD, Painter NS. Diabetes,coronary thrombosis, and the saccharine disease.Bristol, England: John Wright and Sons, 1969.
28. Bierman EL. Carbohydrates, sucrose and humandisease. Am J Clin Nutr l979;32:27l2-22.
by guest on June 19, 2013ajcn.nutrition.org
Dow
nloaded from