Embed Size (px)
Citation preview

A bacterial process for seleniumnanosphere assemblyCharles M. Debieuxa, Elizabeth J. Dridgea,1, Claudia M. Muellera,1, Peter Splatta, Konrad Paszkiewicza, Iona Knighta,Hannah Florancea, John Lovea, Richard W. Titballa, Richard J. Lewisb, David J. Richardsonc, and Clive S. Butlera,2
aBiosciences, College of Life and Environmental Sciences, University of Exeter, Stocker Road, Exeter EX4 4QD, United Kingdom; bInstitute for Cell andMolecular Biosciences, University of Newcastle, Newcastle upon Tyne NE2 4HH, United Kingdom; and cSchool of Biological Sciences,University of East Anglia, Norwich NR4 7TJ, United Kingdom
Edited by Dianne K. Newman, California Institute of Technology/Howard Hughes Medical Institute, Pasadena, CA, and accepted by the Editorial Board July 1,2011 (received for review April 13, 2011)
During selenate respiration by Thauera selenatis, the reduction ofselenate results in the formation of intracellular selenium (Se)deposits that are ultimately secreted as Se nanospheres of approxi-mately 150 nm in diameter. We report that the Se nanospheresare associatedwith a protein of approximately 95 kDa. Subsequentexperiments to investigate the expression and secretion profile ofthis protein have demonstrated that it is up-regulated and secretedin response to increasing selenite concentrations. The protein waspurified from Se nanospheres, and peptide fragments from a tryp-tic digest were used to identify the gene in the draft T. selenatisgenome. A matched open reading frame was located, encoding aprotein with a calculated mass of 94.5 kDa. N-terminal sequenceanalysis of the mature protein revealed no cleavable signal pep-tide, suggesting that the protein is exported directly from thecytoplasm. The protein has been called Se factor A (SefA), andhomologues of known function have not been reported previously.The sefA gene was cloned and expressed in Escherichia coli, andthe recombinant His-tagged SefA purified. In vivo experiments de-monstrate that SefA forms larger (approximately 300 nm) Se nano-spheres in E. coliwhen treatedwith selenite, and these are retainedwithin the cell. In vitro assays demonstrate that the formation ofSe nanospheres upon the reduction of selenite by glutathione arestabilized by the presence of SefA. The role of SefA in seleniumnanosphere assembly has potential for exploitation in bionanoma-terial fabrication.
nanoparticles ∣ biomineralization ∣ anaerobic respiration
The utilization of oxygen or nitrogen oxyanions (nitrate andnitrite) as respiratory substrates presents a fortuitous advan-
tage to organisms, because their respiratory products are eitheraqueous or gaseous and simply diffuse away from the cell. How-ever, this is not always the case. Some microorganisms that livein niche environments have adapted to utilize more unusual sub-strates for energy conservation, such as metal ions or chalcogenoxides (1). Often, the reduction of these compounds can result inthe precipitation of insoluble products that ultimately accumulatewithin the cell (2). If such compounds are to be used as respira-tory substrates, mechanisms for the disposal of the insoluble pro-ducts are essential. A number of systems exist in Gram-negativebacteria for secretion out of the cell and are commonly referredto as the type 1–6 secretion systems (TxSS). A further mechanismfor secretion of both soluble and insoluble material is theprocess of outer membrane vesiculation. In response to stress,a section of the outer membrane forms a distinct spherical vesicle,composed of a lipid bilayer and encloses material exclusively fromthe periplasm (3, 4).
In the present work, the process of bacterial selenate (SeO42−)
respiration has been used to investigate the mechanism of Seprecipitation and secretion. The reduction of selenate followsa sequential series of reductive steps ultimately leading to thegeneration of elemental selenium (Se0). Eqs. 1 and 2 summarizethe overall reactions:
SeO42− þ 2e− þ 2Hþ ⇆ SeO3
2− þH2O [1]
andSeO3
2− þ 4e− þ 6Hþ ⇆ Se0 þ 3H2O: [2]
Thauera selenatis (a β-proteobacterium) is by far the best-studiedselenate respiring bacterium (5–8). The selenate reductase(SerABC) isolated from T. selenatis (6) is a soluble periplasmicenzyme. The enzyme is a type II molybdoenzyme that comprisesthree subunits, SerA (96 kDa), SerB (40 kDa), and SerC(23 kDa), and coordinates molybdenum, heme (b-type), andnumerous [Fe-S] centers as prosthetic constituents (9). SerABCcontributes to proton-motive force generation by accepting elec-trons from a diheme c-type cytochrome (cytc4), which mediateselectron flux from either a quinol-cytochrome c oxidoreductase(QCR) or quinol dehydrogenase. The use of QCR ensures thatselenate reduction is coupled to the Q-cycle mechanism provid-ing a minimum net gain of 2qþ∕2e− of proton electrochemicalgradient (10).
The resultant product from SerABC is selenite (SeO32−). The
reduction of selenite in T. selenatis does not support growth andis not a respiratory substrate. There has been much debate re-garding the mechanisms by which selenite is reduced to seleniumin bacterial cells. Early reports by Macy and coworkers (11)implicated a nitrite reductase in the process of selenite reduction,by virtue that a nonspecific mutant strain of T. selenatis, which wasdeficient in nitrite reductase activity, also failed to produce de-tectable Se0 upon growth in selenate-rich medium. The authorsspeculated that it was probably a periplasmic nitrite reductasethat was responsible for selenite reduction. Selenite reacts readilywith thiols following the reactions described by Painter (12). Glu-tathione (GSH) is the primary reduced thiol in Escherichia coli,and it is now widely believed that it is the prime candidate forbacterial intracellular selenite reduction. Bacteria belonging tothe α, β, and γ groups of the proteobacteria are all abundant inGSH (13), so the utilization of GSH for the reduction of seleniteduring selenate respiration would seem plausible. Selenite reactsreadily with GSH producing selenodiglutathione (GS–Se–SG).GS–Se–SG is a good substrate for GSH reductase and is subse-
Author contributions: R.J.L. D.J.R., and C.S.B. designed research; C.M.D., E.J.D., C.M.M., P.S.,I.K., and C.S.B. performed research; K.P. and H.F. contributed new reagents/analytic tools;J.L., R.W.T., D.J.R., and C.S.B. analyzed data; and C.S.B. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission. D.K.N. is a guest editor invited by the EditorialBoard.
Data deposition: The sefA gene sequence and the Thauera selenatis AX draft genomesequence have been deposited in the NCBI GenBank database, www.ncbi.nlm.nih.gov/genbank (accession nos. HQ380173 and PRJNA53521, respectively).1E.J.D. and C.M.M. contributed equally to this work.2To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1105959108/-/DCSupplemental.
13480–13485 ∣ PNAS ∣ August 16, 2011 ∣ vol. 108 ∣ no. 33 www.pnas.org/cgi/doi/10.1073/pnas.1105959108
quently reduced to form a selenopersulfide of GSH (GS–Se−).GS–Se− is unstable and dismutates into elemental Se (Se0) andreduced GSH. The reaction has been studied for the phototrophicα-proteobacterium, Rhodospirillum rubrum, where the reductionof selenite by GSH results in the accumulation of Se particles inthe cytoplasm (13). Selenite can also be detoxified bymethylation,liberating volatile selenium compounds dimethylselenide anddimethyldiselenide (14, 15). Recently, the S-adenosyl-L-methio-nine (SAM)-dependent methyltransferase (TehB) from E. coli,which is involved in tellurite resistance, has been shown to beeffective in selenium methylation in vitro (16). It is thereforeof interest to investigate the mechanisms by which selenite is de-toxified in a true selenate respiring organism. If T. selenatis utilizesan intracellular reductant to detoxify selenite during selenaterespiration, then elemental Se would inevitably accumulate withinthe cell. Consequently, it is considered likely that an export systemis required for the secretion of the Se0 deposits in order to sustainthe use of selenate as the sole respiratory substrate. The aim of thepresent work was to resolve the mechanism by which T. selenatisdeposits Se during selenate respiration, which has led to the iden-tification of a Se-nanosphere assembly protein.
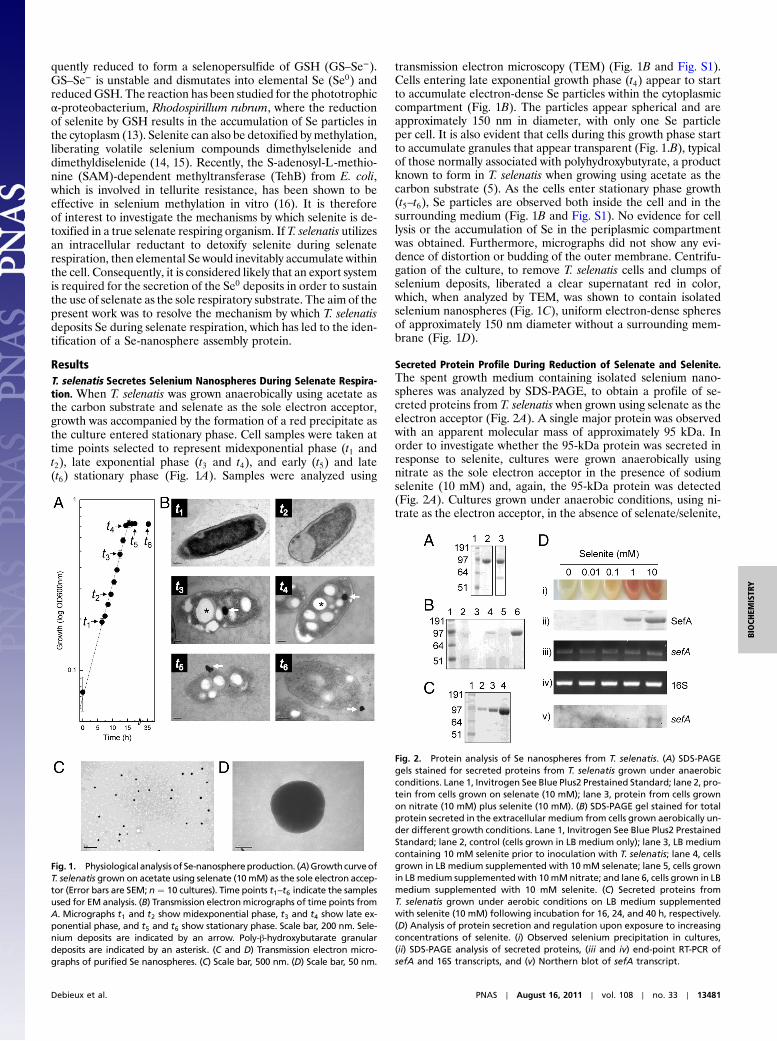
ResultsT. selenatis Secretes Selenium Nanospheres During Selenate Respira-tion. When T. selenatis was grown anaerobically using acetate asthe carbon substrate and selenate as the sole electron acceptor,growth was accompanied by the formation of a red precipitate asthe culture entered stationary phase. Cell samples were taken attime points selected to represent midexponential phase (t1 andt2), late exponential phase (t3 and t4), and early (t5) and late(t6) stationary phase (Fig. 1A). Samples were analyzed using
transmission electron microscopy (TEM) (Fig. 1B and Fig. S1).Cells entering late exponential growth phase (t4) appear to startto accumulate electron-dense Se particles within the cytoplasmiccompartment (Fig. 1B). The particles appear spherical and areapproximately 150 nm in diameter, with only one Se particleper cell. It is also evident that cells during this growth phase startto accumulate granules that appear transparent (Fig. 1.B), typicalof those normally associated with polyhydroxybutyrate, a productknown to form in T. selenatis when growing using acetate as thecarbon substrate (5). As the cells enter stationary phase growth(t5–t6), Se particles are observed both inside the cell and in thesurrounding medium (Fig. 1B and Fig. S1). No evidence for celllysis or the accumulation of Se in the periplasmic compartmentwas obtained. Furthermore, micrographs did not show any evi-dence of distortion or budding of the outer membrane. Centrifu-gation of the culture, to remove T. selenatis cells and clumps ofselenium deposits, liberated a clear supernatant red in color,which, when analyzed by TEM, was shown to contain isolatedselenium nanospheres (Fig. 1C), uniform electron-dense spheresof approximately 150 nm diameter without a surrounding mem-brane (Fig. 1D).
Secreted Protein Profile During Reduction of Selenate and Selenite.The spent growth medium containing isolated selenium nano-spheres was analyzed by SDS-PAGE, to obtain a profile of se-creted proteins from T. selenatis when grown using selenate as theelectron acceptor (Fig. 2A). A single major protein was observedwith an apparent molecular mass of approximately 95 kDa. Inorder to investigate whether the 95-kDa protein was secreted inresponse to selenite, cultures were grown anaerobically usingnitrate as the sole electron acceptor in the presence of sodiumselenite (10 mM) and, again, the 95-kDa protein was detected(Fig. 2A). Cultures grown under anaerobic conditions, using ni-trate as the electron acceptor, in the absence of selenate/selenite,
Fig. 1. Physiologicalanalysisof Se-nanosphereproduction. (A)Growth curveofT. selenatis grown on acetate using selenate (10mM) as the sole electron accep-tor (Error bars are SEM; n ¼ 10 cultures). Time points t1–t6 indicate the samplesused for EM analysis. (B) Transmission electronmicrographs of time points fromA. Micrographs t1 and t2 show midexponential phase, t3 and t4 show late ex-ponential phase, and t5 and t6 show stationary phase. Scale bar, 200 nm. Sele-nium deposits are indicated by an arrow. Poly-β-hydroxybutarate granulardeposits are indicated by an asterisk. (C and D) Transmission electron micro-graphs of purified Se nanospheres. (C) Scale bar, 500 nm. (D) Scale bar, 50 nm.
Fig. 2. Protein analysis of Se nanospheres from T. selenatis. (A) SDS-PAGEgels stained for secreted proteins from T. selenatis grown under anaerobicconditions. Lane 1, Invitrogen See Blue Plus2 Prestained Standard; lane 2, pro-tein from cells grown on selenate (10 mM); lane 3, protein from cells grownon nitrate (10 mM) plus selenite (10 mM). (B) SDS-PAGE gel stained for totalprotein secreted in the extracellular medium from cells grown aerobically un-der different growth conditions. Lane 1, Invitrogen See Blue Plus2 PrestainedStandard; lane 2, control (cells grown in LB medium only); lane 3, LB mediumcontaining 10 mM selenite prior to inoculation with T. selenatis; lane 4, cellsgrown in LB medium supplemented with 10 mM selenate; lane 5, cells grownin LBmedium supplementedwith 10mMnitrate; and lane 6, cells grown in LBmedium supplemented with 10 mM selenite. (C) Secreted proteins fromT. selenatis grown under aerobic conditions on LB medium supplementedwith selenite (10 mM) following incubation for 16, 24, and 40 h, respectively.(D) Analysis of protein secretion and regulation upon exposure to increasingconcentrations of selenite. (i) Observed selenium precipitation in cultures,(ii) SDS-PAGE analysis of secreted proteins, (iii and iv) end-point RT-PCR ofsefA and 16S transcripts, and (v) Northern blot of sefA transcript.
Debieux et al. PNAS ∣ August 16, 2011 ∣ vol. 108 ∣ no. 33 ∣ 13481
BIOCH
EMISTR
Y

produced either very little or failed to secrete detectable levels ofthe 95-kDa protein. The secretion profile of the 95-kDa proteinwas further investigated from cells grown under aerobic condi-tions in the presence/absence of various oxyanions (Fig. 2B).When T. selenatis was grown aerobically on LB medium alone, orin the presence of nitrate, the 95-kDa protein was not detected.Upon the addition of selenate (10 mM), a faint band was resolvedat 95 kDa, the amount of which increased when the medium wassupplemented with 10 mM selenite. The quantity of protein se-creted as a function of time following exposure to selenite wasalso investigated (Fig. 2C). The amount of the 95-kDa proteinsecreted increased for 16, 24, and 40 h postincubation with10 mM selenite. To investigate whether or not a threshold levelof selenite concentration was needed to induce secretion of the95-kDa protein, aerobic cultures of T. selenatis were supplemen-ted with selenite at increasing concentration (0, 0.01, 0.1, 1, and10 mM) and incubated for 24 h. Culture growth was monitored atOD600 nm to ensure that the presence of selenite did not have adeleterious effect on cell growth. Red elemental selenium wasdetected in cultures exposed to >1 mM selenite (Fig. 2D). Ana-lysis of the secreted protein profile shows that the 95-kDa proteinis only detectable from cultures where the selenite concentrationwas >1 mM (Fig. 2D). Analysis using inductively coupled plasmaatomic emission spectroscopy detected selenium in purified pro-tein samples, giving calculated molar ratios of approximately320∶1 selenium to protein.
Characterization of the 95-kDa Secreted Protein.N-terminal sequen-cing (Pinnacle Proteomic Facility, Newcastle University) andliquid chromatography/tandem mass spectroscopy of tryptic-digest fragments (obtained from both University of York Proteo-mics Department and Biosciences, University of Exeter) resultedin 124 amino acids of sequence data that was used to identify thegene in the draft assembly of the T. selenatis genome [NationalCenter for Biotechnology Information (NCBI) accession no.PRJNA53521]. Open reading frame (ORF) prediction was car-ried out using the EMBOSS getorf package (17), and candidateregions were searched via BLAST (BlastX and BlastP) againstthe nonredundant database of protein sequences. A matchedORF was located on contig 179, and its translation predicted aprotein with a calculated mass of 94532.73 Da (Fig. S2). Indepen-dent N-terminal sequence analysis of the mature protein pro-duced a peptide sequence (AITATQRT), which aligns adjacentto the start methionine of the target protein identified. The pro-tein does not possess a leader peptide that is cleaved duringexport from the cell (18–20). Consequently, translocation to theperiplasmic compartment directly via a Sec (18) or twin-argininetranslocation (TAT) (19, 20) pathway seems unlikely. Interest-ingly, 64.3% of the primary sequence is derived from only fiveamino acids (16.4% Ala; 15.4% Thr; 12% Gly; 10.4% Val; 10.1%Asp). BLASTsearches against the NCBI nonredundant databaseand UniProt database revealed that there are few statisticallysignificant matches to putative and hypothetical proteins in othermicrobial organisms. Alignment of SefA with a hypotheticalprotein [NAL212_3002 (839aa)] identified in the genome ofNitrosomonas sp. AL212 [isolated from laboratory activatedsludge (21)] reveals another family member (Fig. S2). Other re-sults indicate that similarity to other proteins is limited only to aregion covering amino acids 100–200 from the start methionine.One of these was identified in the genome of Desulfurispirillumindicum strain S5 (Selin_0231) (22), a known selenate respiringbacterium. Other hits reveal similarity to S-layer proteins, such asSapA from Campylobacter fetus. Additional Interpro (23) andPfam (24) scans of the gene revealed no significant profile hits.Attempts to elucidate gross structure by hidden-Markov-modelcomparison using HHPred (http://toolkit.lmb.uni-muenchen.de/hhpred) yielded no additional clues as to the structure and func-tion of the protein, which we have named Se factor A (SefA). To
determine whether the expression of SefA correlated with the se-cretion profile observed in the presence of increasing selenite(Fig. 2D), the expression of SefA upon exposure of cells to sele-nite was evaluated at the mRNA transcription level by RT-PCRand Northern blotting (Fig. 2D). Cells grown in the presence ofincreasing concentrations of selenite were sampled for mRNA inthe stationary phase, just prior to the observation of SefA andSe nanospheres in the extracellular medium. End-point RT-PCRdetected sefA transcripts in all samples; however, quantitativeNorthern blot analysis revealed that sefA was up-regulated incultures supplemented with 10 mM selenite.
Analysis of the sefA Locus. Annotation of the ORFs adjacent toSefA (Fig. 3A) revealed that upstream are a putative CHASE2extracellular sensory domain/guanylate cyclase and a proteincontaining a tetratricopeptide (TPR) repeat. Downstream is aputative SAM-dependent methyltransferase, a further putativepeptide, and a peptide with a domain of unknown function(DUF). A search for putative transcriptional regulator bindingsites upstream of the sefA gene was performed using the Prokar-yotic Promoter Prediction system (http://bioinformatics.biol.rug.nl/websoftware/ppp/ppp_start.php). This revealed the presenceof a putative FNR promoter binding site between positions −87and −103 relative to the sefA gene start site. FNR is a transcrip-tional regulator active at low oxygen levels. The motif TGTGA-TTCCCATCACA falls within the FnrBac∕PrfA Group motifs(TGTGA-N6-TCACA) classified as Crp/FNR factors by Körneret al. (25). Analysis of the intergenic region between sefA andthe putative SAM-dependent methyltransferase cannot indentifya known transcriptional regulator binding site. It is thus consid-ered likely that both sefA and the gene encoding the putativemethyltransferase are in the same operon. Consequently, we havecalled the methyltransferase SefB. SefB is a 513 aa protein with acalculated molecular weight of 56.9 kDa. Pfam analysis identifiesboth putative methyltransferase and regulatory domains. A SefBhomologue has also been located in D. indicum strain S5(Selin_0233).
Fig. 3. Analysis of the sef operon. (A) Schematic representation of the sefgene locus. Putative annotations are as follows: CHASE2 extracellular sensorydomain and guanylate cyclase (with sec leader peptide in red); SefA, seleniumnanosphere assembly protein; SefB, SAM-methyltransferase;?, putative pep-tide. The (B) nucleotide sequence of the promoter region of sefA. (C) Nucleo-tide sequence upstream from sefB. A putative Shine–Dalgarno (SD) sequenceand putative FnrBac∕PrfA binding motif (TGTGA-N6-TCACA) are located up-stream of sefA. No obvious promoter binding sequences are identified be-tween sefA and sefB. A putative SD sequence upstream of sefB is also located.
13482 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1105959108 Debieux et al.

Expression of SefA in E. coli, in Vivo Localization, and Resistance toSelenite. The gene encoding SefA was cloned and expressed inE. coli (see SI Materials and Methods and Fig. S3). Western blotanalysis of soluble cell fractions and extracellular medium areshown in Fig. 4A. After 4 h induction, SefA is detected predomi-nantly in the soluble cell fraction but is also detectable in theextracellular medium. Analysis of the medium by SDS-PAGEindicated that this was not due to cell lysis. At 19 h after induc-tion, the amount of SefA detected in both fractions increased sub-stantially. In the presence of selenite (10 mM), SefA was againdetected in both the soluble cell fractions and the extracellularmedium at both time points. Samples that were not treated withIPTG failed to produce detectable SefA in either fraction.Further analysis of the selenite-treated samples by TEM revealedthat cells not expressing SefA were shown to accumulate Separticles ranging in size from 30–50 nm (Fig. 4B). Cells treated
with selenite/IPTG accumulated larger particles (approximately300 nm) in the cytoplasm (Fig. 4C). No evidence was found forextracellular Se nanospheres. The results show that SefA can beexpressed in E. coli and indicate it might be a substrate for proteinexport. The addition of selenite to the culture had no obviouseffect on the level of exported SefA, suggesting that secretionfrom E. coli was not conditional upon Se binding. These data sug-gest that although apo-SefA can be secreted from E. coli, a spe-cific export system is required for Se-nanosphere secretion.Analysis of E. coli (pET33b-sefA) to selenite resistance shows thatcells in which SefA is expressed are more resistant to selenite con-centrations up to 25 mM than those cells without SefA (Fig. 4D).
In Vitro Assembly of Selenium Nanospheres Is Facilitated by SefA.rSefA was purified using immobilized nickel affinity chromato-graphy and gel filtration (Fig. S3). Purified rSefA was analyzedby SDS-PAGE and demonstrated a migration position similar tothat of the native protein from T. selenatis (Fig. S3). The Se par-ticles, as secreted by T. selenatis during selenate respiration andonce separated by filtration through a 0.2 μM filter, displayed anabsorbance peak at 400 nm when analyzed by electron absorptionspectroscopy. Consequently, absorbance at 400 nm was used tomonitor the formation of Se particles in vitro from the reactionof reduced GSH (4 mM) with selenite (0.5 mM) in the presenceor absence of rSefA. Using molar ratios of GSH:selenite >4∶1ensured that GS–Se–SG was reduced to GSSeH, which readilydecomposed to Se0 (13). Fig. 4E shows formation of seleniumparticles as a function of time. Under these conditions, the reac-tion in the absence of SefA proceeded to give a maximum absor-bance of approximately 1 unit and produced a distinct blackprecipitate in the reaction cuvette. In the presence of SefA, themaximum absorbance was stabilized at 0.5 units, and the particlesremained in solution giving the typical red/orange color. TEManalysis of the particle formation under the reaction conditionsare shown in Fig. 4 F and G. The black precipitate observed inthe absence of SefAwas visible under TEMas vitreous Se deposits(Fig. 4F). In the presence of SefA, distinct Se nanospheres wereobserved with sizes ranging from 20–300 nm (Fig. 4G).
DiscussionBacteria that are capable of the respiration of selenium oxyanionshave been isolated from a number of selenium-rich environments(26, 27). Using selenium oxides as the sole electron acceptorpresents the organism with the challenge of what to do withthe elemental Se product following the subsequent reductions.For some bacteria that catalyze the reduction of selenate to Se0,Se deposits have been observed to accumulate within the cyto-plasmic compartment, whereas in others, they are secreted fromthe cell (28). For example; Enterobacter cloacae SLD1a-1 (29),R. rubrum (30), and Desulfovibrio desulfuricans (31) have all beenfound to bioreduce selenite to selenium both inside and outsidethe cell. E. coli is also able to reduce selenite-forming depositsboth in the periplasmic and cytoplasmic compartments (32).However, for a selenate respirer such as T. selenatis, a build-up ofSe in the cytoplasm or periplasm might not be sustainable andcould lead to necrosis. Consequently, mechanisms have evolvedfor the secretion of solid Se0 precipitates. In the present work, theprocess by which T. selenatis deposits Se during selenate respira-tion has been investigated. A secreted protein, SefA, has beenidentified, and homologues of known function have not beenreported previously. Evidence is presented that SefA facilitatesSe-nanosphere assembly prior to export from the cell.
The observation that SefA has no detectable selenite reductaseactivity would suggest that SefA functions only to bind to, or tostabilize, Se0. Because selenate reduction in T. selenatis is peri-plasmic, the location of the selenite reductive pathway remainsunclear. A previous study has suggested that a periplasmic nitritereductase could be responsible for selenite reduction (11); how-
Fig. 4. Expression of SefA in E. coli and in vitro formation of Se nanospheres.(A) Western blot analysis of the localization of SefA expression in E. colifollowing exposure to selenite. Samples (10 μg) were prepared from cellsþ∕− IPTG and þ∕− selenite (10 mM). Lanes 2, 4, and 6 represent extracellularprotein; lanes 3, 5, and 7 represent soluble cell extracts. Samples were ana-lyzed at both 4 and 19 h after the addition of IPTG and selenite. Lanes 1 and 8show an 80-kDa marker. (B) Transmission electron micrograph of an E. colisingle cell harboring plasmid pET33b-sefA grown in the presence of selenite(10 mM), but not induced with IPTG. (C) Transmission electron micrograph ofan E. coli single cell harboring plasmid pET33b-sefA grown in the presence ofboth selenite (10 mM) and IPTG. Three replicate cells are shown in Fig. S3.Selenium deposits are indicated by an arrow. (D) Growth yield at stationaryphase of E. coli cells exposed to increasing selenite concentrations. E. coli har-boring plasmid pET33b-sefA in the absence (hashed bars) or presence (solidbars) of IPTG, and E. coli cells harboring plasmid pET33b in the presence ofIPTG (open bars). Error bars are SEM (n ¼ 3). ** indicates t test, p < 0.01 com-pared to the nonrecombinant (pET33b) control. (E) The reaction of GSH(4 mM) with selenite (0.5 mM), either in the presence (○) or absence (▪)of purified rSefA (0.5 μg), monitored at 400 nm. (F and G) Transmission elec-tron micrographs of the reaction product from E, in the absence (F) and in thepresence (G) of SefA. Scale bars, 0.5 μm and 200 nm for F and G, respectively.
Debieux et al. PNAS ∣ August 16, 2011 ∣ vol. 108 ∣ no. 33 ∣ 13483
BIOCH
EMISTR
Y

ever, selenite reductase activity in periplasmic fractions, usingreduced methyl viologen as the electron donor, has not been de-tected. Based upon the present data, it is suggested that selenitereduction occurs in the cytoplasm. The reduction to Se0 could bedue to interaction with thiols, possibly GSH, because GSH isabundant in both the β-proteobacteria (T. selenatis) and γ-proteo-bacteria (E. coli) (12, 13). GSH is synthesized by sequentialactions of γ-glutamylcysteine synthetase (GshA) and GSHsynthetase (GshB), and homologues for both GshA and GshBhave been identified in the draft T. selenatis genome. The reactionof selenite with GSH can result in the generation of O2
−, whichis detoxified in part by cytochrome c (13). Interestingly, we havepreviously identified a c-type cytochrome (cytc-Ts7) that is up-regulated during selenate respiration but is not involved in elec-tron transfer to SerABC (10). It is possible that the function ofcytc-Ts7 is to help detoxify any O2
− generated.The diagram presented in Fig. 5 shows the model for the
selenate respiration pathway in T. selenatis. The reduction ofselenate draws electrons from the membrane-bound QCR, whichconcomitantly provides a net gain of 2qþ∕2e− of proton electro-chemical gradient (10). Although the utilization of a QCR isunusual for a periplasmic molybdoenzyme, the additional 2Hþtranslocated via the link to the Q-cycle could provide the drivingforce for the translocation of selenite across the cytoplasmicmembrane. In this case, energy would also need to be conservedby the utilization of an electrogenic primary dehydrogenase. Theexact nature of the selenite transporter in T. selenatis is unknown,but in E. coli it has been suggested that selenite crosses the cyto-plasmic membrane via the sulphate transporter (33). Once in thecytoplasm, selenite could be reduced by GSH (or other reduc-tants), and the resultant Se0 binds to SefA, forming a nanosphereinside the cell. In addition, it is likely that the putative SAM-dependent methyltransferase (SefB), downstream from SefA,might function to generate methyl selenite or other volatile sele-nium compounds. The involvement of the tellurite resistanceSAM-dependent methyltransferase (TehB) has been shownrecently to be effective in the methylation of both tellurite andselenite (16). Interestingly, although E. coli TehB has been iden-
tified as responsible for the methylation of tellurite in vitro (16),in vivo assays have not established the production of volatilemethylated tellurium, yet precipitation of tellurium within cellsis clearly seen as black deposits (34). The identification of SefAand SefB in the same operon, suggests that in T. selenatis therecould be a link between both reductive and methylation-depen-dent selenite detoxification.
How does the Se nanosphere get out of the cell? SefA does nothave an N-terminal signal sequence for targeting to the periplam-sic compartment. Furthermore, analysis of neighboring geneson the chromosome does not identify any likely TAT substratecandidates that might “carry” SefA to the periplasm. The size ofthe particles is consistent with those seen during outer membranevesiculation (20–250 nm) (4), but by TEM we see no evidence ofouter membrane distortion or bulging. Furthermore, outer-mem-brane vesicles contain exclusively material from the periplasm,whereas the Se particles are not associated with other periplasmicproteins (such as SerABC) and from the EM images SefA–Senanospheres would appear to be translocated directly from thecytoplasm. Analysis of the T. selenatis draft genome assemblyusing an Interpro scan of all ORFs >50 amino acids revealedthe presence of all but Type V secretion systems. SefA could thusbe exported via the T6SS because other proteins secreted by thissystem also lack an N-terminal signal sequence and are not firsttranslocated to the periplasmic compartment. S-layer proteins(which show some sequence similarity to SefA) are typicallysubstrates for the T1SS. Most notably, T1SS substrates share acommon distinctive glycine-rich repeat (GGXGXDXXX) andcontain very few or no cysteine residues (35, 36), both features ofSefA. These characteristics might explain why SefA was detectedin the extracellular medium when expressed in E. coli. However,it would appear that in T. selenatis the SefA–Se complex is as-sembled in the cell and only exported once at an optimum size.T. selenatis is somehow capable of sensing the size of the internalparticles and selectively only secretes Se nanospheres of approxi-mately 150 nm diameter. Evidently, E. coli lacks this capabilityand accumulated much larger Se–SefA particles, suggesting thatthe Se-nanosphere export system is specific to T. selenatis. Themechanism of how the Se nanospheres as a whole are assembledand secreted across the inner and outer membranes remainsunknown.
The identification of another SefA family member in Nitroso-monas sp. (AL212 NAL212_3002) is interesting and deservescomment. The strain AL212 was isolated from cultures that couldgrow in up to 10.7 mM ðNH4ÞSO4 and along with strain JL21(grown in 3.57 mM) were called the “sensitive” strains (21). Ithad been noted in previous studies that some of the other strainsisolated from treatment plants and laboratory sludges, onceenriched in culture with ðNH4ÞSO4, showed the formation of“particles” in the cytoplasm (37), but not in strain JL21. Cur-iously, the EM images of strain JL21 (and to a lesser extentAL212) reported in Suwa et al. (21) show evidence of extracel-lular spherical particles reminiscent of those created by SefA. Itwould seem likely that in this case, these particles could be basedupon the chalcogen sulfur rather than selenium.
Various proteins that bind Se have been reported. Rhodanese(a sulfur transferase) (38), cysteine/selenocysteine lyases (39),SeBP (40) from Methanococcus vannielii, and the glycolyticenzyme glyceraldehyde-3-phosphate dehydrogenase (41) can allbind selenium at a reactive cysteine residue. Analysis of the ami-no acid sequence of SefA reveals no cysteine residues are present,indicating that the Se–SefA interaction is not via a direct thiolligand. BSA has also been shown to stabilize nano-Se followingreduction by GSH. Because BSA is (moderately) soluble in saltsolutions, it serves well as a carrier for molecules of low watersolubility. The nonspecific interaction with BSA and Se is thoughtto stabilize the Se nanosphere by allowing interactions of its func-tional groups with water and by sterically avoiding Se aggregation
Fig. 5. Schematic diagram showing the proposed pathway of selenium oxy-anion reduction and Se-nanosphere assembly in T. selenatis. The reduction ofselenate draws electrons from the membrane-bound QCR, generating a netgain of 2qþ∕2e− of proton electrochemical gradient, which could provide thedriving force for the translocation of selenite across the cytoplasmic mem-brane. Once in the cytoplasm, selenite reduction occurs, and the resultantSe0 binds to SefA, forming a Se nanosphere prior to export from the cell.The process by which SefA–Se is exported remains unknown. The identifica-tion of a gene (sefB) encoding a putative SAM-dependent methyltransferasemight also provide a mechanism for selenite detoxification via volatilizationto methylated selenides (R–Se–R). OM, outer membrane; IM, cytoplasmicinner membrane.
13484 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1105959108 Debieux et al.

in aqueous solution (42), thus capping the particle surface. Itwould seem likely that, in the absence of any thiol ligands, SefAwould also function as a capping agent, providing reaction sitesfor the creation of Se nanospheres and providing a shell to pre-vent aggregation of the secreted particles.
The involvement of SefA in Se-nanoshpere assembly mighthave applications pertinent to nanotechnology. In particular,selenium nanoparticles have excellent bioavailability, high biolo-gical activity, and relatively low toxicity. A number of methodshave been developed to generate Se nanoparticles, nanorods, andnanowires (43), but most require the use of hydrazine, glycol, sur-factant, or high temperature. A number of studies have also takenadvantage of biomolecules to generate Se nanoparticles in vitro.For example, Abdelouas et al. (44) have used reduced cyto-chrome c to synthesize Se nanowires at room temperature. Morerecently, Zhang et al. (45) synthesized t-Se nanowires using β-car-otene as an in situ soft template. Having identified a protein thatcan stabilize Se nanospheres secreted from T. selenatis, this could,through molecular engineering, enable particles to be producedwith structural arrangements that are not only unique, but alsohave yet to be reproduced by conventional chemical synthesis.The extracellular secretion of such nanoparticles from T. selenatis
also has distinct benefits when compared to those bacterial sys-tems that display intracellular accumulation, and as such thecurrent work could present an opportunity for the synthesis ofsecreted Se nanomaterials.
Materials and MethodsSee SI Materials andMethods for details of growth of T. selenatis, isolation ofSefA, identification of the sefA gene, generation of recombinant SefA inE. coli, assays for in vitro selenium nanosphere formation, imaging T. selenatiscells and Se nanospheres by TEM, RT-PCR, and Northern blotting reactionsand protocols.
ACKNOWLEDGMENTS.We thank Gavin Wakely, Edgar Dawkins, and Oluwake-mi Obasa for help with preliminary experiments. Thauera selenatis waskindly provided by Dr. Joanne Santini (University College London). We thankDr. David Studholme (University of Exeter), Dr. Ian Henderson (University ofBirmingham), Dr. Jon Marles-Wright, and Dr. Elisabeth Lowe (University ofNewcastle) for useful discussions and Kevin Brigden (University of Exeter)for preliminary ICP analysis. We also thank Carmen Denman and Dr. AlanBrown (University of Exeter) for help with the Northern blotting. C.M.D.,E.J.D., and I.K. were funded by a research grant from the Biotechnologyand Biological Sciences Research Council and the Engineering and PhysicalSciences Research Council (Life Science Interface) (BB/D00781X/1 to C.S.B.,R.J.L. and D.J.R.).
1. Richardson DJ (2000) Bacterial respiration: A flexible process for a changing environ-ment. Microbiology 146:551–571.
2. Lloyd JR (2003) Microbial reduction of metals and radionuclides. FEMS Microbiol Rev27:411–425.
3. McBroom AJ, Kuehn MJ (2007) Release of outer membrane vesicles by Gram-negativebacteria is a novel envelope stress response. Mol Microbiol 63:545–558.
4. Kulp A, Kuehn MJ (2010) Biological functions and biogenesis of secreted bacterialouter membrane vesicles. Annu Rev Microbiol 64:163–184.
5. Macy JM, et al. (1993) Thauera selenatis gen-nov, sp-nov, a member of the beta-sub-class of proteobacteria with a novel type of anaerobic respiration. Int J Syst Bacteriol43:135–142.
6. Schröder I, Rech S, Krafft T, Macy JM (1997) Purification and characterization of theselenate reductase from Thauera selenatis. J Biol Chem 272:23765–23768.
7. Krafft T, Bowen A, Theis F, Macy JM (2000) Cloning and sequencing of the genesencoding the periplasmic-cytochrome B-containing selenate reductase of Thaueraselenatis. DNA Seq 10:365–377.
8. Maher MJ, et al. (2004) X-ray absorption spectroscopy of selenate reductase. InorgChem 43:402–404.
9. Dridge EJ, et al. (2007) Investigation of the redox centres of periplasmic selenatereductase from Thauera selenatis by EPR spectroscopy. Biochem J 408:19–28.
10. Lowe EC, et al. (2010) Quinol-cytochrome c oxidoreductase and cytochrome c4 med-iate electron transfer during selenate respiration in Thauera selenatis. J Biol Chem285:18433–18442.
11. DeMoll-Decker H, Macy JM (1993) The periplasmic nitrite reductase of Thauera sele-natis may catalyze the reduction of selenite to elemental selenium. Arch Microbiol160:241–247.
12. Painter EP (1941) The chemistry and toxicity of selenium compounds with specialreference to the selenium problem. Chem Rev 28:179–213.
13. Kessi J, Hanselmann KW (2004) Similarities between the abiotic reduction of selenitewith glutathione and the dissimilatory reaction mediated by Rhodospirillum rubrumand Escherichia coli. J Biol Chem 279:50662–50669.
14. Ranjard L, Nazaret S, Cournoyer B (2003) Freshwater bacteria can methylateselenium through thiopurine methyltransferase pathway. Appl Environ Microbiol69:3784–3790.
15. Ranjard L, et al. (2004) Characterization of a novel selenium methyltransferase fromfreshwater bacteria showing strong similarities with the calicheamicin methyltransfer-ase. Biochim Biophys Acta 1679:80–85.
16. Choudhury HG, Cameron AD, Iwata S, Beis K (2011) Structure and mechanism ofthe chalcogen detoxifying protein TehB from Escherichia coli. Biochem J 435:85–91.
17. Rice P, Longden I, Bleasby A (2000) EMBOSS: The European Molecular Biology OpenSoftware Suite. Trends Genet 16:276–277.
18. Pugsley AP (1993) The complete general secretory pathway in gram-negative bacteria.Microbiol Rev 57:50–108.
19. Berks BC, Sargent F, Palmer T (2000) The Tat protein export pathway. Mol Microbiol35:260–274.
20. Sargent F (2007) Constructing the wonders of the bacterial world: Biosynthesis ofcomplex enzymes. Microbiology 153:633–651.
21. Suwa Y, Sumino T, Noto K (1997) Phylogenetic relationships of activated sludge iso-lates of ammonia oxidizers with different sensitivities to ammonia sulphate. J GenAppl Microbiol 43:373–379.
22. Rauschenbach I, Narasingarao P, Haggblom M (2011) Desulfurispirillum indicumsp.nov., a selenate and selenite respiring bacterium isolated from an estuarine canalin southern India. Int J Syst Evol Microbiol 61:654–658.
23. Quevillon E, et al. (2005) InterProScan: Protein domains identifier. Nucleic Acids Res 33(Web Server issue):W116–W120.
24. Finn RD, et al. (2010) The Pfam protein families database. Nucleic Acids Res 38(Data-base issue):D211–D222.
25. Körner H, Sofia HJ, ZumftWG (2003) Phylogeny of the bacterial superfamily of Crp-Fnrtranscription regulators: Exploiting the metabolic spectrum by controlling alternativegene programs. FEMS Microbiol Rev 27:559–592.
26. Stolz JF, Oremland RS (1999) Bacterial respiration of arsenic and selenium. FEMS Mi-crobiol Rev 23:615–627.
27. Stolz JF, Basu P, Santini JM, Oremland RS (2006) Arsenic and selenium in microbialmetabolism. Annu Rev Microbiol 60:107–30.
28. Oremland RS, et al. (2004) Structural and spectral features of selenium nanospheresproduced by Se-respiring bacteria. Appl Environ Microbiol 70:52–60.
29. Losi ME, Frankenberger WT (1997) Reduction of selenium oxyanions by Enterobactercloacae SLD1a-1: Isolation and growth of the bacterium and its expulsion of seleniumparticles. Appl Environ Microbiol 63:3079–3084.
30. Kessi J, Ramuz M, Wehrli E, Spycher M, Bachofen R (1999) Reduction of selenite anddetoxification of elemental selenium by the phototrophic bacterium Rhodospirillumrubrum. Appl Environ Microbiol 65:4734–4740.
31. Tomei FA, et al. (1995) Transformation of selenate and selenite to elemental seleniumby Desulfovibrio desulfuricans. J Ind Microbiol Biotechnol 14:329–336.
32. Gerrard TL, Telford JN, Williams HH (1974) Detection of selenium deposits in Escher-ichia coli by electron microscopy. J Bacteriol 119:1057–1060.
33. Turner RJ, Weiner JH, Taylor DE (1998) Selenium metabolism in Escherichia coli.Biometals 11:223–7.
34. Liu M, et al. (2000) Escherichia coli requires S-adenosylmethionine as a cofactor tomediate tellurite resistance. J Bacteriol 182:6509–6513.
35. Baumann U, Wu S, Flaherty KM, McKay DB (1993) Three-dimensional structure of thealkaline protease of Pseudomonas aeruginosa: A two-domain protein with a calciumbinding parallel beta roll motif. EMBO J 12:3357–64.
36. Delepelaire P (2004) Type I secretion in gram-negative bacteria. Biochim Biophys Acta1694:149–161.
37. Suwa Y, Imamura Y, Suzuki T, Tashiro T, Urushigawa Y (1994) Ammonia-oxidizingbacteria with different sensitivies to ðNH4ÞSO4 in activated sludges. Water Res28:1523–1532.
38. Ogasawara Y, Lacourciere GM, Stadtman TC (2001) Formation of a selenium-substi-tuted rhodanese by reaction with selenite and glutathione: Possible role of a proteinperselenide in a selenium delivery system. Proc Natl Acad Sci USA 98:9494–9498.
39. Mihara H, et al. (1999) A nifS-like Gene, csdB, Encodes an Escherichia coli counterpartof mammalian selenocysteine lyase:gene cloning, purification, characterisation andpreliminary X-ray crystallographic studies. J Biol Chem 274:14768–14772.
40. Patteson KG, Trivedi N, Stadtman TC (2005) Methanococcus vannielii selenium-bindingprotein (SeBP): Chemical reactivity of recombinant SeBP produced in Escherichia coli.Proc Natl Acad Sci USA 102:12029–12034.
41. Lacourciere GM, Levine RL, Stadtman TC (2002) Direct detection of potential seleniumdelivery proteins by using an Escherichia coli strain unable to incorporate seleniumfrom selenite into proteins. Proc Natl Acad Sci USA 99:9150–9153.
42. BückingW, Massadeh S, Merkulov A, Xu S, Nann T (2010) Electrophoretic properties ofBSA-coated quantum dots. Anal Bioanal Chem 396:1087–1094.
43. Gates B, Mayers B, Cattle B, Xia Y (2002) Synthesis and characterization of uniformnanowires of trigonal selenium. Adv Funct Mater 12:219–227.
44. Abdelouas A, et al. (2000) Using cytochrome c, to make selenium nanowires. ChemMater 12:1510–1512.
45. Zhang B, et al. (2006) Biomolecule-assisted synthesis of single-crystalline selenium na-nowires and nanoribbons via a novel flake-cracking mechanism. Nanotechnology17:385–390.
Debieux et al. PNAS ∣ August 16, 2011 ∣ vol. 108 ∣ no. 33 ∣ 13485
BIOCH
EMISTR
Y









![A bacterial process for selenium nanosphere assembly › content › pnas › 108 › 33 › 13480.full.pdf · SeO4 2− þ2e− þ2Hþ ⇆ SeO 3 2− þH 2O [1] and SeO3 2− þ4e−](https://img.pdfslide.us/doc/110x75/5f20c89cfd9fad22b515f978/a-bacterial-process-for-selenium-nanosphere-assembly-a-content-a-pnas-a-108.jpg)






![Nanosphere [Ag(SR)]n: coordination polymers of Ag+ with a](https://img.pdfslide.us/doc/110x75/61dae58b31fddd7393715b24/nanosphere-agsrn-coordination-polymers-of-ag-with-a-.jpg)