Embed Size (px)
Citation preview

A 27-Amino-Acid Synthetic Peptide Corresponding to the
NH2-Terminal Zinc-Binding Domain of Endostatin Is
Responsible for Its Antitumor Activity
Robert M. Tjin Tham Sjin,1Ronit Satchi-Fainaro,
1Amy E. Birsner,
1V.M. Sadagopa Ramanujam,
2
Judah Folkman,1and Kashi Javaherian
1
1Vascular Biology Program, Department of Surgery, Children’s Hospital Boston, Harvard Medical School, Boston, Massachusetts and2Division of Human Nutrition, Department of Preventive Medicine and Community Health, University of Texas Medical Branch,Galveston, Texas
Abstract
The first recombinant endostatin that elicited strong anti-tumor activity was expressed in Escherichia coli and admin-istered as a suspension. Under these conditions, the proteinretained its full antiangiogenic activity. Lack of requirementfor a folded structure prompted us to investigate antitumorproperties of synthetic peptides corresponding to different re-gions of endostatin. Here, we show that the entire antitumor,antimigration, and antipermeability activities of endostatinaremimicked by a 27-amino-acid peptide corresponding to theNH2-terminal domain of endostatin. This peptide containsthree histidines that are responsible for zinc binding.Mutations of the zinc-binding histidines abolished its anti-tumor and antimigration activities, but not antipermeabilityproperties. (Cancer Res 2005; 65(9): 3656-63)
Introduction
Endostatin, a 183-amino-acid proteolytic cleavage fragmentcorresponding to the COOH terminus of collagen 18, has beenthe subject of investigation by a number of laboratories because ofits antitumor activity with no toxic side effects (1–5). Severalantiangiogenic activities have been reported for this protein, suchas inhibition of endothelial cell proliferation, migration, and tubeformation. Endostatin also suppresses vascular endothelial growthfactor (VEGF)–induced vascular permeability (6). However, themechanism of action of endostatin remains unknown. Endostatininhibits endothelial cell migration by inhibiting phosphorylation offocal adhesion kinase via binding to a5h1 integrin (7). It also hasbeen shown that cell surface glypicans are low-affinity endostatinreceptors (8). Endostatin has been implicated in several signalingpathways, such as down-regulation of c-myc (9), cyclin D1 (10) andRhoA activity (11), blockage of VEGF signaling (12, 13), andinhibition of the wnt-signaling pathway (14). Furthermore, endo-statin has been shown to bind and inactivate metalloproteinases(15–17) and to regulate a spectrum of genes that suppressangiogenesis (18).The crystal structures of both murine and human endostatin
have been elucidated (19, 20) and show a noncovalently held dimerat high concentration required for crystallization (20). Thepresence of two disulfide bonds results in a highly folded structure.
Endostatin binds one atom of zinc (Zn) per monomer via the threehistidines in the NH2 terminus of the molecule (histidines 1, 3,and 11) and aspartic acid 76. The heparin-binding property ofendostatin is mediated by noncontiguous arginines clustered overthe three-dimensional globular surface of the molecule (21).We have previously shown that oligomeric endostatin (NC1 and
dimer) is primarily associated with laminin in the basementmembrane (22). This association may be important for some of thebiological functions displayed by endostatin. On the other hand,the heparin-binding properties of endostatin manifest themselvesin its interaction with the cell surface. It is likely that endostatinhas several biological functions mediated by different regions of theprotein.Our goal in this report is to identify and characterize the
endostatin region responsible for its antitumor activity. Towardachieving this goal, a series of overlapping peptides correspondingto the complete sequence of endostatin were synthesized. We findthat the entire antiangiogenic activity of endostatin is located in a27-amino-acid peptide that binds Zn. We show that Zn binding isrequired for antitumor and antimigration activities of endostatinbut not its antipermeability property.
Materials and Methods
Cell culture and reagents. Human BxPC-3 pancreatic adenocarcinomaand murine Lewis lung carcinoma (LLC) cells (American Type Culture
Collection, Rockville, MD) were grown and maintained as described earlier
(2, 23). Human microvascular endothelial cells (HMVEC-d, Clonetics,
Walkersville, MD) were cultured in microvascular endothelial cell growthmedium (EGM-2 MV; Clonetics) and maintained at 5% CO2 in a 37jChumidified incubator.
Recombinant human endostatin was a generous gift from EntreMedCorporation (Rockville, MD) and recombinant human and murine FcEndos-
tatin were prepared as described earlier (24). Human and murine endostatin
peptides were synthesized by SynPep Corporation (Dublin, CA). Peptides
were resuspended in PBS or 50 mmol/L Tris, 150 mmol/L NaCl (pH 7.5).PECAM, purified rat anti-mouse CD31, was obtained from BD
PharMingen (San Diego, CA) and human recombinant VEGF was obtained
from the NIH (Bethesda, MD).
The thrombospondin-1 (TSP-1) type 1 repeats fragment 3TSR, containingamino acid 361 to 530 of TSP-1 (25), was kindly provided by Dr. Jack Lawler
(Beth Israel Deaconess Medical Center, Department of Pathology, Boston,
MA). TNP-470 was a generous gift from Takeda Chemical Industries Ltd.(Osaka, Japan).
Animal studies. All animal procedures were done in compliance with
Boston Children’s Hospital guidelines, and protocols were approved by the
Institutional Animal Care and Use Committee. Male (24-27 g) immuno-competent C57Bl/6J mice (The Jackson Laboratory, Bar Harbor, ME) and
immunocompromised severe combined immunodeficiency (SCID) mice
(Massachusetts General Hospital, Boston, MA) were used. Mice were 7 to 9
Requests for reprints: Kashi Javaherian, Vascular Biology Program, Department ofSurgery, Children’s Hospital Boston, Karp Family Research Laboratories, Room 11.213,One Blackfan Circle, Boston, MA 02115. Phone: 617-919-2392; Fax: 617-739-5891;E-mail: [email protected].
I2005 American Association for Cancer Research.
Cancer Res 2005; 65: (9). May 1, 2005 3656 www.aacrjournals.org
Research Article
Research. on April 21, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

weeks of age. They were acclimated, caged in groups of five in a barrier carefacility, and fed animal chow and water ad libitum . Animals were
anesthetized via inhalation of isoflurane (Baxter, Deerfield, IL) before all
surgical procedures and observed until fully recovered. Animals were
euthanized by a lethal dose of CO2 asphyxiation.Tumor models. BxPC-3 and LLC cells were grown in 900-cm2 roller
bottles. The cell concentration was adjusted to 50 � 106 cells/mL. Mice
were shaved and the dorsal skin was cleaned with ethanol before tumor cell
injection. A suspension of 5 � 106 tumor cells in 0.1 mL RPMI 1640 ( for
BxPC-3) or DMEM ( for LLC) was injected s.c. into the dorsa of mice at the
proximal midline. BxPC-3 cells were implanted in SCID mice and LLC in
C57Bl/6J mice as described earlier (2).
The mice were weighed and tumors were measured every 3 to 5 days in
two diameters with a dial caliper. Volumes were determined using the
formula a2 � b � 0.52 (where a is the shortest and b is the longest
diameter). Data is represented as volume of treated tumor over control
(T/C). At the completion of each experiment, the mice were euthanized
with CO2 asphyxiation. Tumors were fixed in 10% buffered formalin (Fisher
Scientific, Fair Lawn, NJ) and paraffin embedded.
For treatment of tumor-bearing mice, tumor volumes were allowed to
grow to f100 mm3, and mice were randomized. Treatment was done by
single bolus s.c. injections. Peptides were given twice a day (every 12 hours).
Doses indicated for peptides were corrected for the purity of peptides
(f70%). For example, mice given injections of 4 mg/kg/d peptide were
actually given 2.8 mg/kg/d after correction. BxPC-3 tumors were treated for
28 days and LLC tumors for 14 days. The unpaired Student t test was used
for statistical analysis.
Immunohistochemistry. Tumors were fixed in 10% buffered formalin
overnight at 4jC. The next day, tumors were washed thrice in PBS and
paraffin embedded. Sections (5 Am) were permeabilized with 40 Ag/mLproteinase K (Roche Diagnostics Corp., Indianapolis, IN) in 0.2 mol/L Tris-
HCl buffer (pH 7.6) for 25 minutes at 37jC and washed with PBS. PECAM
(1:250) was incubated at 4jC overnight. Stainings were amplified using
tyramide signal amplification direct and indirect kits (NEN Life ScienceProducts Inc., Boston, MA). Sections were photographed at 200� or 400�magnification using a NIKON TE300 microscope (Melville, NY). Vessel
density (average of 10 fields) was determined by IPLab software. Theunpaired Student t test was used for statistical analysis.
Endothelial cell migration assay. The motility response of HMVEC-d
cells was assayed using amodified Boyden chamber. Cells were plated in T75-
cm2 flasks at 0.5� 106 cells per flask and allowed to grow for 48 hours (f70%confluent) before the migration assay. To facilitate cell adhesion, the upper
membrane of a transwell (8-mm pore; Costar, Corning, NY) was coated with
fibronectin (10 Ag/mL; Becton Dickinson, Bedford, MA) for 1 hour at 37jC.Coatedmembranes were rinsed with PBS and allowed to air-dry immediatelybefore use. Cells were detached by trypsinization, treated with trypsinization
neutralization solution (Clonetics), and resuspended at a final concentration
of 1� 106 cells/mL in serum-free endothelial basal medium (EBM; Clonetics)
containing 0.1% bovine serum albumin (BSA). Cells (200,000 in 0.2 mL) werethen treated with 0.2 mL of EBM/BSA containing endostatin or peptides at
the indicated concentrations. Cells were incubated for 20 minutes at 37jCwith occasional shaking. Cells (50,000 in 100 AL) were added to the upperchamber of the transwell. EBM or EBM supplemented with VEGF (5 ng/mL)
was added to the lower chamber and cells were allowed to migrate toward
the bottom chamber for 4 hours in a humidified incubator containing 5%
CO2. Transwell filters were rinsed once with PBS and fixed and stained usinga Diff-Quik staining kit (Baxter) following the manufacturer’s protocol.
Nonmigrated cells were removed from the upper chamber with a cotton
swab. Stained filters were cut out of the chamber and mounted onto slides
using Permount (Fisher Scientific). The number of migrated cells wasmeasured using microscopy (three fields from each membrane were
captured using a 40� objective), and images were captured with a charge-
coupled device camera using SPOT software. Total migration per membranewas quantified from the captured images using Scion Image software (NIH).
All experiments were run in triplicate. Migration was normalized to percent
migration, with migration to VEGF alone representing 100% migration. The
data represents an average of five to six separate experiments. High error bars
were observed due to the variability inherent in endothelial cell migrationassays. A 3-fold increase in VEGF-induced migration over basal levels was
considered the minimum criterion for an experiment to be considered valid.
The unpaired Student t test was used for statistical analysis.
Miles vascular permeability assay. SCID mice (n = 12) were given s.c.injections of human endostatin (EntreMed; 100 mg/kg/d), murine
FcEndostatin (20 mg/kg/d), peptides (either 14 or 2.8 mg/kg/d), and saline
(200 AL) for 5 days before performing the Miles assay (26). Briefly, Evan’s blue
dye (100 AL of a 1% solution in PBS) was injected i.v. into mice. After 10minutes, 50 AL of human recombinant VEGF (1 ng/AL) or PBS were injectedintradermally into the preshaved back skin. After 20 minutes, the animals
were euthanized and an area of skin that included the blue spot resulting
from leakage of the dye was removed. Evan’s blue dye was extracted from theskin by incubation with formamide for 5 days at room temperature, and the
absorbance of extracted dye was measured at 620 nm with a spectropho-
tometer. The unpaired Student t test was used for statistical analysis.Statistical methods. Data are expressed as mean F SD. Statistical
significance was assessed using the Student t test. For all statistical
comparisons, treated groups were compared with PBS-treated controls or
VEGF alone (in migration assays). P < 0.05 was considered statisticallysignificant.
Results
Identification of the endostatin peptide that is responsiblefor antitumor activity. The first recombinant endostatin wasexpressed in Escherichia coli and purified using 8 mol/L urea andreducing conditions (1). After dialysis against PBS, the finalproduct, in the form of an insoluble suspension, was injected intoimmunocompetent C57Bl/6J mice bearing murine LLC, resulting intumor inhibition (1). Although the above protocol has beenrepeated by several laboratories, some laboratories failed toreproduce these results, leading to a controversy on theeffectiveness of endostatin (27). One possible explanation for theantitumor activity of the E. coli–derived insoluble endostatin is thata folded endostatin structure is not required. We hypothesized thatendostatin antitumor activity might be mediated by a shortsynthetic peptide. Toward this goal, we synthesized eight over-lapping peptides with 24 to 27 amino acids derived from humanendostatin (Table 1). Peptides were approximately one seventh toone eighth the size of full-length endostatin. Three cys-teines33,165,173 were replaced by alanines, and cysteine135 wasomitted, to prevent the formation of disulfide bonds. Twoadditional lysines were added at the COOH-terminal of hP8 toincrease its solubility. Most peptides were water soluble, except forhP2 at high concentrations (z2.5 mg/mL). In addition, all peptides
Table 1. Overlapping human endostatin peptides
hP1 1HSHRDFQPVLHLVALNSPLSGGMRG25
hP2 23MRGIRGADFQAFQQARAVGLAGTFR47
hP3 45TFRAFLSSRLQDLYSIVRRADRAAV69
hP4 67AAVPIVNLKDELLFPSWEALFSGSE91
hP5 89GSEGPLKPGARIFSFDGKDVLRHPT113
hP6 111HPTWPQKSVWHGSDPNGRRLTESY134
hP7 136ETWRTEAPSATGQASSLLGGRLLGQ160
hP8 158LGQSAASAHHAYIVLAIENSFMTASKKK183
NOTE: Cysteines (33, 165, 173) were substituted by alanines (under-
lined) or omitted (Cys135). The bold letters represent the overlapping
amino acids. Two additional lysines were added at the COOHterminus of hP8 to increase its solubility (double underlined).
Antitumor Activity of Endostatin Peptide
www.aacrjournals.org 3657 Cancer Res 2005; 65: (9). May 1, 2005
Research. on April 21, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

were f70% pure. However, no difference in tumor inhibition wasobserved when peptides of more than 95% purity were used (datanot shown).These peptides were initially tested for antitumor activity using
the human pancreatic tumor cells BxPC-3 in SCID mice. Peptides(7 mg/kg/d) were given s.c. twice a day due to the high clearancerate from mouse circulation, whereas full-length endostatin(human FcEndostatin, hFcES; 20 mg/kg/d) was given only onceper day. We have previously shown that FcES could successfullyinhibit tumor growth (24). The NH2-terminal hP1 peptide ofendostatin inhibited BxPC-3 by 39% (P = 0.077) and full-lengthendostatin by 44% (P = 0.0057; Fig. 1). Two other peptides, hP2 andhP5, also showed some small antitumor activity, wherein hP2inhibited BxPC-3 by 19% (P = 0.48) and hP5 by 29% (P = 0.15). Theremaining peptides had no effect on inhibiting tumor growth (Fig. 1).Thus, most of the antitumor activity was associated with the NH2-terminal hP1 peptide compared with full-length endostatin. Tumorinhibition by hP1, hP2, and hP5 was statistically not significant,probably due to the small number of mice per group (n = 3) used.Although peptides were not used at equimolar ratios to full-lengthendostatin, this data still suggests that the antitumor property ofendostatin may be located within its NH2-terminal domain.The NH2-terminal 27 amino acid peptide of endostatin is
responsible for its antitumor property. We next examined theeffect of all 8 endostatin peptides on murine Lewis lung carcinoma(LLC) tumor growth. For these experiments, murine analogues of theendostatin peptides were synthesized. The only difference betweenthe human and murine peptides was that murine P1 peptidecontained 27 amino acids instead of 25 amino acids for human P1.Unlike the treatment of BxPC-3 tumors, LLC tumors were treatedat equimolar concentrations of murine full-length endostatin(20 mg/kg/d) and murine peptides (2.8 mg/kg/d). Eight mice pergroup were used and control mice were treated with PBS. Peptideswere injected (s.c.) twice a day, whereas full-length endostatin andPBS were given once a day. The NH2-terminal mP1 endostatinpeptide inhibited LLC by 45% (P = 0.0021), which is similar to theinhibition by full-length endostatin (48%, P = 0.0068; Fig. 2A).No significant tumor inhibition was detected using the remaining
peptides (mP2 to mP8) at the same concentration used as the mP1peptide (Fig. 2A). Thus, this result suggests that the 27-amino-acidmP1 peptide contains all of the antitumor activity associated withendostatin.We also wanted to determine the effect on angiogenesis after
mP1 treatment. Therefore, LLC tumors treated with the endostatinpeptides and full-length endostatin at equimolar concentrationwere analyzed for vessel density (CD31; Fig. 2B and C). Treatmentof LLC with mP1 and full-length endostatin reduced vessel densitysignificantly (f50%), whereas the remaining peptides and PBS hadno effect. These results suggest that mP1 can inhibit LLC tumorgrowth by reducing vessel density in similar manner to full-lengthendostatin.Histidine residues at position 1 and 3 of endostatin are
critical for zinc binding. The crystal structure of endostatinreveals a highly folded molecule (ref. 20; Fig. 2D). However, theNH2-terminal region resembles a random coil structure consistentwith our analysis that a synthetic peptide corresponding to thisdomain can mimic the native molecule (Fig. 2D). There is an atomof Zn associated with each molecule of endostatin (20). Based onour crystal structure analysis, three histidines at positions 1, 3, and11, plus aspartic acid at position 76, form the four coordinates forthis Zn atom (20). Initially, it was found that trypsin digestion ofhuman endostatin resulted in cleavage of four amino acids (HSHR)from the NH2 terminus, which lacked Zn binding (20). BecausemP1 contains the three histidine residues mentioned above, itraised the possibility that this peptide was able to bind Zn byhaving a molecule of water occupying the fourth coordinate(Fig. 3 A , left). Previously, our group has shown that site-directedmutagenesis of histidine residues 1 and 3 of endostatin to alaninesresulted in the loss of its antitumor property (28). Furthermore, itwas shown that deleting the first three amino acids HTH of murineendostatin disrupted Zn binding (28). Therefore, a mutant ofpeptide mP1 was synthesized and the histidines at positions 1 and3 were mutated to alanines. This mutant peptide was called mP1-H1/3A. To determine the Zn-binding capacity of mP1 and mP1-H1/3A, flame atomic absorption was done. Each peptide was dissolvedin 20 mmol/L Tris (pH 8.0) mixed with excess Zn chloride andextensively dialyzed against the above buffer. These data yielded Znratios of 0.1 per molecule of mP1-H1/3A and 0.9 for mP1 (Fig. 3B).Therefore, mutating the histidines at positions 1 and 3 to alaninesabolished Zn binding (Fig. 3A , right).The zinc-binding domain of endostatin is important for its
antitumor activity. To determine if Zn binding is also importantfor the antitumor property of endostatin, mP1 and mP1-H1/3Awere tested using the LLC tumor model. Peptides were adminis-tered twice a day (s.c.) at a dose of 2.8 mg/kg/d. Peptide mP1inhibited LLC by 42% (P = 0.031), whereas mP1-H1/3A had noeffect (Fig. 3C). To determine if there was a difference inangiogenesis, vessel density (CD31) of LLC tumors was analyzedafter mP1 and mP1-H1/3A treatment. There was a considerabledecrease in vessel density after mP1 treatment (67% reduction,P < 0.01), whereas mP1-H1/3A was similar to the PBS-treatedcontrol (Fig. 3D and E). These data suggest that Zn binding isimportant for the antitumor property of endostatin.The NH2-terminal fragment of endostatin inhibits endo-
thelial cell migration. Endostatin has previously been shown toinhibit endothelial cell migration (2, 29). Therefore, we tested thehP1 peptide and the mutant hP1 peptide (hP1-H1/3A) for anti–endothelial cell migration activity. Inhibition of VEGF-inducedmigration of HMVECs was determined using several doses of
Figure 1. Treatment of human pancreatic carcinoma with human endostatinpeptides. BxPC-3 cells were implanted in the s.c. dorsa of SCID mice andsystemically treated with human endostatin peptides (7 mg/kg/d s.c.),full-length human FcEndostatin (hFcES ; 20 mg/kg/d s.c.), and PBS (as control).Peptides were administered twice a day. Tumors were measured every 3 daysand the final measurements at day 28 are shown. T/C is indicated within thecolumns . Group sizes: n = 3. P values (against PBS control): hFcES,P = 0.0057; hP1, P = 0.077; hP2, P = 0.48; hP5; P = 0.15.
Cancer Research
Cancer Res 2005; 65: (9). May 1, 2005 3658 www.aacrjournals.org
Research. on April 21, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

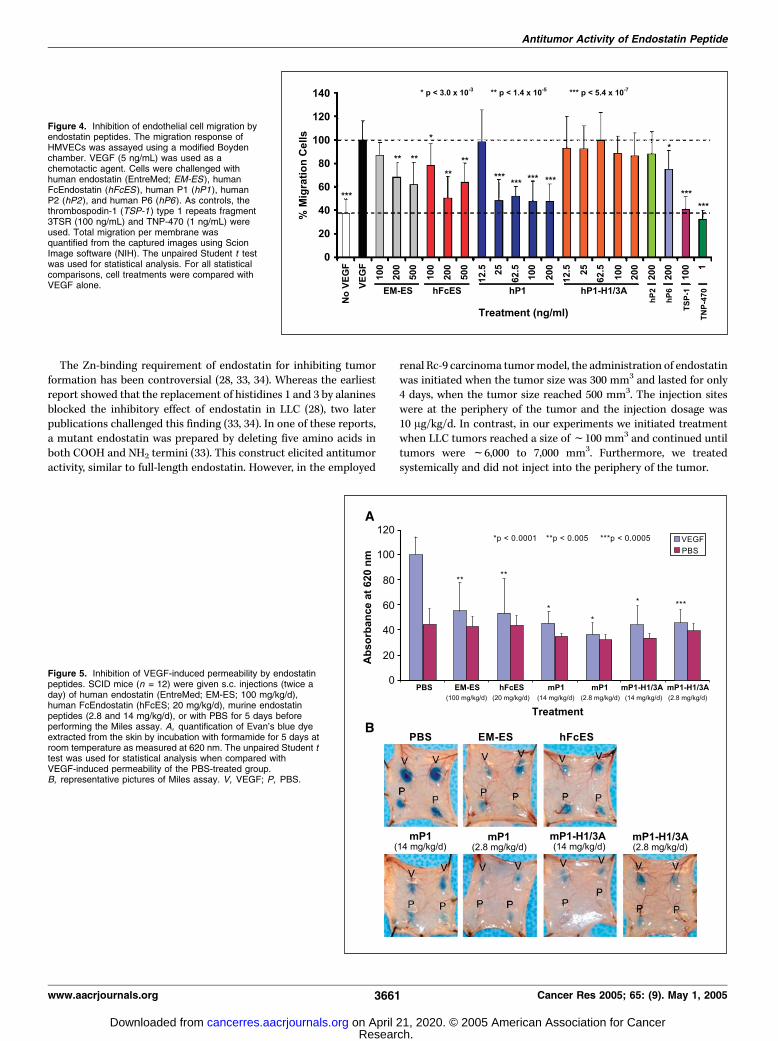
endostatin and endostatin peptides (Fig. 4). Human peptideswere used because the cells were of human origin. Two sourcesof full-length endostatin [EntreMed endostatin (EM-ES) andhuman FcEndostatin (hFcES)] were used in the migration assay,both of which inhibited migration. The inhibition of full-lengthendostatin was dose responsive between 100 and 200 ng/mL.Interestingly, there was no additional inhibition with 500 ng/mL.Furthermore, we found that hP1 inhibited endothelial cellmigration at lower concentrations than full-length endostatin.Maximal inhibition seems to occur with doses between 25 and200 ng/mL (Fig. 4).To determine if the Zn-binding site is important for anti–
endothelial cell migration activity, hP1-H1/3A was also tested. Noinhibition of endothelial cell migration was observed for hP1-H1/3A at any dose (Fig. 4). As positive controls, two other angiogenesisinhibitors previously used in VEGF-induced endothelial cell
migration were also included. As expected, the TSP-1 fragment3TSR (30) and TNP-470 (31) significantly inhibited endothelial cellmigration (Fig. 4).We also examined two other peptides, hP2 and hP6. As expected,
hP2 had no effect on migration at 200 ng/mL (Fig. 4). There wasalso no inhibition observed at lower concentrations (data notshown). hP6 between 100 and 200 ng/mL had a slight effect oninhibition (only 200 ng/mL is shown). However, lower doses had noeffect on inhibiting migration (data not shown).The endostatin hP1 peptide could inhibit VEGF-induced
endothelial cell migration at equimolar concentrations (25 ng/mL)to full-length endostatin (200 ng/mL), whereas hP6 only inhibitedat doses of 100 and 200 ng/mL. These results show that the NH2-terminal P1 peptide of endostatin maintains the ability to inhibitVEGF-induced endothelial cell migration and that the Zn-bindingsite is critical for this activity.
Figure 2. The NH2-terminal domain ofendostatin is responsible for its antitumorproperties. A, treatment of LLC with murineFcEndostatin and murine endostatinpeptides. LLC cells were implanted in thes.c. dorsa of C57Bl/6J mice, andsystemically treated. Murine FcEndostatin(mES ) was given at 20 mg/kg/d s.c., oneinjection per day. Peptides were given at2.8 mg/kg/d s.c., two injections per day.Control mice were treated with PBS. T/C isindicated. Group sizes: n = 8. The unpairedStudent t test was used for statisticalanalysis. B, CD31 staining. LLC sectionswere from PBS-, FcEndostatin- (20 mg/kg/d),and murine endostatin peptide(2.8 mg/kg/d)–treated mice. Peptides wereadministered s.c. twice a day. LLC tumorsections from day 14 were formalin fixed,paraffin embedded, and then stained withCD31 (PECAM). Sections werephotographed at 200� magnification usinga NIKON TE300 microscope. CD31staining is in brown . C, quantification ofvessel density based on CD31 staining.Y axis, % CD31/high power field (at 200�magnification). D, crystal structure ofendostatin. The orange-shaded areacorresponds to P1, which is randomlycoiled. Two a helixes are designated a1and a2. Letters A to P, h sheets.Yellow-red strings, disulfide bonds. Blackcircle, Zn binding to the NH2 terminus ofendostatin.
Antitumor Activity of Endostatin Peptide
www.aacrjournals.org 3659 Cancer Res 2005; 65: (9). May 1, 2005
Research. on April 21, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

Antipermeability activities of endostatin peptides. Severalpathologic diseases, including tumors, have been associated withincreased vascular permeability. Endostatin has been shown tosuppress VEGF-induced permeability (6). Therefore, the ability ofendostatin peptides to inhibit VEGF-induced permeability was alsotested using the Miles assay (26). Previously, endostatin has beenshown to inhibit VEGF-induced permeability using the Milesassay.3 Immunocompromised SCID mice were treated for 5 daysbefore performing the Miles assay. Interestingly, both mP1 andmP1-H1/3A inhibited VEGF-induced permeability when tested atboth 2.8 mg/kg/d (which is at equimolar ratio to full-lengthendostatin) and 14 mg/kg/d (Fig. 5). This effect on permeabilitywas similar to that with full-length endostatin. These data indicatethat two separate mechanisms or separate regions of the P1peptide may regulate antitumor and antipermeability activities.
Discussion
We have shown that a synthetic peptide, corresponding to theNH2 terminus of endostatin, is responsible for its antitumor,
antimigration, and antipermeability activities. Zinc binding isrequired for the antitumor and antimigration activities becausesubstitution of the two histidines at amino acid positions 1 and 3 inthe peptide completely blocks these properties. According to ourcrystallography data, histidines 1, 3, 11, and aspartic acid 76 ofendostatin are important for Zn binding (20). Mutating theseresidues to alanine showed reduced antitumor activity of full-length endostatin and the double H1/3A mutant abolishedendostatin antitumor activity completely (28). The removal of fouramino acids, including the above two histidines from the NH2
terminus of endostatin, resulted in the loss of Zn binding byendostatin (20). In another study, it was shown that deletion of thefirst three amino acids HTH from the NH2 terminus of murineendostatin prevented Zn incorporation by endostatin (28).However, Zn binding was not required for the antipermeabilityproperty of endostatin because the mutant mP1-H1/3A retained itsfull antipermeability property. It is known that hyperpermeability isregulated by RhoA activity, calcium, tyrosine kinase, and celljunctions, resulting in the disruption of endothelial cell barrierfunction (32). Endostatin regulates many signaling pathways and itis likely that different mechanisms or separate regions of the P1peptide regulate endostatin antitumor and antipermeabilityactivities.
Figure 3. The Zn-binding site ofendostatin is important for antitumoractivity. A, schematic diagram of mP1 andmP1-H1/3A. Zinc binding to mP1 ismediated by three histidines at positions1, 3, and 11 (left). Histidines at positions1 and 3 are mutated to alanines in mP1-H1/3A. These mutations prevent Zn binding(right ). Sequences of mP1 and mP1-H1/3Aare indicated. B, Zn binding to mP1 andmP1-H1/3A. Each peptide was dissolved in20 mmol/L Tris (pH 8) at a concentration of0.5 mg/mL and 1 mmol/L Zn chloride wasadded. Each sample was dialyzed againstTris buffer for 72 hours with three changesin dialysis solution [molecular weight cutoff(MWCO) = 1,000 Da]. Atomic absorptionreadings of the final Zn concentrations(Ag/mL) were determined to be 9.63 and1.05 for mP1 and mP1-H1/3A,respectively. These data yielded Zn ratiosof 0.9 per molecule of mP1 and 0.1 formP1-H1/3A. Molecular weight of thepeptides was taken to be 3,000 Da. C,treatment of LLC with mP1 and mP1-H1/3A. Peptides were given s.c. twice a day.Both mP1 and mP1-H1/3A were given at2.8 mg/kg/d. D, LLC tumor sectionsstained with CD31. Sections werephotographed at 400� magnification.E, determination of vessel density. Theunpaired Student t test was used forstatistical analysis.
3 Shay Soker, personal communication.
Cancer Research
Cancer Res 2005; 65: (9). May 1, 2005 3660 www.aacrjournals.org
Research. on April 21, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from
The Zn-binding requirement of endostatin for inhibiting tumorformation has been controversial (28, 33, 34). Whereas the earliestreport showed that the replacement of histidines 1 and 3 by alaninesblocked the inhibitory effect of endostatin in LLC (28), two laterpublications challenged this finding (33, 34). In one of these reports,a mutant endostatin was prepared by deleting five amino acids inboth COOH and NH2 termini (33). This construct elicited antitumoractivity, similar to full-length endostatin. However, in the employed
renal Rc-9 carcinoma tumormodel, the administration of endostatinwas initiated when the tumor size was 300 mm3 and lasted for only4 days, when the tumor size reached 500 mm3. The injection siteswere at the periphery of the tumor and the injection dosage was10 Ag/kg/d. In contrast, in our experiments we initiated treatmentwhen LLC tumors reached a size off100 mm3 and continued untiltumors were f6,000 to 7,000 mm3. Furthermore, we treatedsystemically and did not inject into the periphery of the tumor.
Figure 4. Inhibition of endothelial cell migration byendostatin peptides. The migration response ofHMVECs was assayed using a modified Boydenchamber. VEGF (5 ng/mL) was used as achemotactic agent. Cells were challenged withhuman endostatin (EntreMed; EM-ES ), humanFcEndostatin (hFcES ), human P1 (hP1), humanP2 (hP2), and human P6 (hP6). As controls, thethrombospodin-1 (TSP-1 ) type 1 repeats fragment3TSR (100 ng/mL) and TNP-470 (1 ng/mL) wereused. Total migration per membrane wasquantified from the captured images using ScionImage software (NIH). The unpaired Student t testwas used for statistical analysis. For all statisticalcomparisons, cell treatments were compared withVEGF alone.
Figure 5. Inhibition of VEGF-induced permeability by endostatinpeptides. SCID mice (n = 12) were given s.c. injections (twice aday) of human endostatin (EntreMed; EM-ES; 100 mg/kg/d),human FcEndostatin (hFcES; 20 mg/kg/d), murine endostatinpeptides (2.8 and 14 mg/kg/d), or with PBS for 5 days beforeperforming the Miles assay. A, quantification of Evan’s blue dyeextracted from the skin by incubation with formamide for 5 days atroom temperature as measured at 620 nm. The unpaired Student ttest was used for statistical analysis when compared withVEGF-induced permeability of the PBS-treated group.B, representative pictures of Miles assay. V, VEGF; P, PBS.
Antitumor Activity of Endostatin Peptide
www.aacrjournals.org 3661 Cancer Res 2005; 65: (9). May 1, 2005
Research. on April 21, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

Another publication that dealt with the relevance of Zn bindingto antitumor activity of endostatin showed that the removal of fouramino acids ‘‘HSHR’’ from the NH2 terminus of human endostatindid not affect its antitumor activity (34). Measurements of Znbinding revealed that this mutant bound 2 atoms of Zn per mol-ecule of endostatin, whereas the wild-type bound 10 atoms of Znper endostatin molecule. However, in our crystal structure studiesof endostatin, we have shown that endostatin contains 1 atom ofZn/endostatin molecule and the removal of the four amino acidsHSHR from the NH2 terminus results in loss of Zn binding (20).Endostatin is generated by proteolytic cleavage of collagen 18
(1, 35, 36). The first amino acid at the NH2 terminus of endostatin isa histidine. The presence of histidine is important for conferring Znbinding to endostatin. Consequently, we are led to conclude thatthe processing of collagen 18 to endostatin may be highly regulated.Several groups have shown that peptides derived from endostatin
have antiangiogenic effects (29, 37–40). An NH2-terminal peptidecomposed of amino acids 6 to 49 (lacking the Zn-binding histidines)has inhibited endothelial cell proliferation and migration (37, 38).A Matrigel assay using this peptide has resulted in the inhibition ofangiogenesis in vivo . However, no antitumor data was presented.In another study, a COOH-terminal peptide (amino acids 135-184)retaining the Cys135–Cys165 disulfide bond, has shown antitumoractivity (39). However, the peptide was administered at the tumorperiphery and not systemically. Cho et al. have shown that the NH2
terminus, which includes the Zn-binding site, and the COOHterminus of endostatin are not required for antitumor activity (40).However, this peptide and full-length endostatin were not tested atequimolar concentrations. Our results differ from these groups inthat the P1 peptide systemically inhibited tumor formation,endothelial cell migration, and permeability at equimolar concen-trations to full-length endostatin. Furthermore, at higher concen-trations (14 mg/kg/d), mP2 inhibited LLC tumor formation as wellas mP1 at 2.8 mg/kg/d (data not shown). However, mP1 at 14 mg/kg/d inhibited LLC tumor formation less than at 2.8 mg/kg/d (datanot shown). Thus, a U-shaped curve seems to be associated withantitumor activity of endostatin as a function of the proteinconcentration. Similar results were observed for full-length endo-statin using the pancreatic BxPC-3 and ASPC-1 tumor models.4
Therefore, determination of optimum endostatin concentrationmay be an important factor. In vitro assays have shown a similarbiphasic characteristic by endostatin with the use of matrixmetalloproteinases (MMP; ref. 16).Similar to the mP1 endostatin peptide, antiangiogenic peptides
from TSP-1 (ABT-510; ref. 41) and tumstatin (42), which is afragment of collagen IV a3 chain, have been shown to haveantitumor activities. Employing the LLC tumor model, wecompared the antitumor activities of the mP1 endostatin peptidewith the tumstatin T8 peptide and the ABT-510/TSP-1 peptideusing equimolar concentrations. The antitumor activity of mP1 andthe tumstatin T8 peptide was similar, whereas mP1 was moreeffective in inhibiting tumor growth than the ABT-510/TSP-1peptide (data not shown).The fact that full-length endostatin is not required for its
antitumor activity explains the initial inconsistencies of endostatinactivity (27). Endostatin has two disulfide bonds. Aggregation ofendostatin in E. coli preparations is caused by random intermo-lecular disulfides after PBS dialysis. Whereas endostatin shows a
single protein molecule under reducing conditions, most of theprotein in an identical sample does not enter the polyacrylamidegel under nonreducing condition (data not shown). It is probablythe degree of nonspecific aggregation that is responsible for thelack of activity in some of the preparations. Endostatin is mostlikely released from the aggregate in animals over a period of time,resulting in a denatured protein or partial fragments, which arecapable of demonstrating antitumor properties due to their NH2-terminal peptide. Presumably, some of the preparations yield largeraggregates, which make such a release inefficient and give rise toa product that is incapable of eliciting an antiangiogenic responsein mice.What is the basis of endostatin’s antitumor activity? Several
mechanisms have been proposed. Binding of endostatin to integrina5h1 has been studied in more detail (7). Based on the findings ofthese authors, an assembly of several cell surface proteins andcomponents, including a5h1, is responsible for interactionsbetween endostatin and this integrin (11). However, no antitumordata were presented to confirm the above mechanism. Morerecently, the same authors have shown that an 11-amino-acidpeptide derived from endostatin containing arginines and showingheparin binding is responsible for antiangiogenic activity ofendostatin (29). We speculate that the phenomena observed bythese investigators reflects some of the properties associated withthe heparin-binding characteristic of endostatin and not itsantitumor activity. Previously, we reported that disruption ofheparin binding of endostatin (accomplished by the mutation oftwo discontinuous arginines on the protein surface) blocked cellmotility (43). Furthermore, our endostatin hP3 peptide (see Table 1),which contains the peptide reported by the authors, failed toinhibit tumor growth.The peptide P1 resembles the catalytic Zn-binding regions of
MMPs. Three separate groups have investigated endostatin bindingto MMPs. One group has reported that endostatin forms a complexwith MMP-2, inhibiting its enzymatic activity (15). A second grouphas reported the absence of association between MMP-2 andendostatin (44). Finally, a third group obtained data indicatingpossible interactions between MMP-2 and endostatin (16).Therefore, we investigated the binding of endostatin to MMP-2and to determine the effect of enzymatic activity using gelzymography. Both endostatin and the peptides could not inhibitthe MMP-2 activity when using recombinant MMP-2 and MMP-2derived from human umbilical vascular endothelial cells (data notshown). However, there still is a possibility of coparticipation ofMMPs and endostatin in biological reactions. It is possiblethat endostatin may bind to the cellular target of MMPs(i.e., extracellular matrix and cell surface proteins) and block theirenzymatic activities. Such specific cellular recipients have beenpoorly characterized at the present time.The mechanism of endostatin action is unknown. Here, we have
shown that a 27-amino-acid peptide corresponding to the NH2
terminal of endostatin, which contains a Zn atom, is responsiblefor the entire antitumor activity of endostatin. The binding of Zn iscrucial for this phenomenon. We believe that the work presented inthis publication is probably the most detailed investigation of allthe peptides reported for endostatin thus far. This finding mayenable us to substitute endostatin with mP1 peptide in clinicaltrials, hence providing a major saving in time and resources.Moreover, because this peptide retains its antipermeability activity,it can be used to treat other hyperpermeability diseases (i.e., retinaldiseases, rheumatoid arthritis, and endometriosis). In addition,4 Ilhan Celik and Oliver Kisker, personal communication.
Cancer Research
Cancer Res 2005; 65: (9). May 1, 2005 3662 www.aacrjournals.org
Research. on April 21, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

it may be useful in clinical applications by reducing edema orrepairing the vascular leak syndrome, as seen in pulmonary edema,ascites, and inflammation. Moreover, these results shed light on ourunderstanding of the molecular mechanism of endostatin.
Acknowledgments
Received 5/25/2004; revised 2/14/2005; accepted 2/16/2005.Grant support: NIH grant R01 CA064481 and the Breast Cancer Research
Foundation.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
We thank Dr. Carmen Barnes for helpful insights and discussions; Dr. Sarah Shortfor helpful discussions, critical reading of the manuscript, and providing assistancewith the migration assay; Drs. Arja Kaipainen and Dipak Panigraphy for assisting usto implant LLC tumors and for their assistance with immunohistochemistry; Drs.Ilhan Celik and Oliver Kisker for assisting us with the BxPC-3 tumor model; Dr.William Fogler from EntreMed for kindly providing human recombinant endostatin;Drs. Stephen Gillies and Kin-Ming Lo from EMD Lexington Research Center forproviding us with human and murine FcEndostatin constructs; Rickey Sanchez fortechnical assistance with preparing immunohistochemistry slides; and KristinGullage for photography.
References1. O’Reilly MS, Boehm T, Shing Y, et al. Endostatin: anendogenous inhibitor of angiogenesis and tumorgrowth. Cell 1997;88:277–85.
2. Kisker O, Becker CM, Prox D, et al. Continuousadministration of endostatin by intraperitoneallyimplanted osmotic pump improves the efficacy andpotency of therapy in a mouse xenograft tumor model.Cancer Res 2001;61:7669–74.
3. Dhanabal M, Ramchandran R, Volk R, et al. Endo-statin: yeast production, mutants, and antitumor effectin renal cell carcinoma. Cancer Res 1999;59:189–97.
4. Yoon SS, Eto H, Lin CM, et al. Mouse endostatininhibits the formation of lung and liver metastases.Cancer Res 1999;59:6251–6.
5. Folkman J, Kalluri R. Tumor angiogenesis. In: Kufe DW,Pollack RE, Weichselbaum RR, Bast RC, Gansler TS,Holland JF, Frei E, editors. Cancer medicine. Hamilton:B.C. Decker Inc.; 2003. p. 161–94.
6. Takahashi K, Saishin Y, Silva RL, et al. Intraocularexpression of endostatin reduces VEGF-induced retinalvascular permeability, neovascularization, and retinaldetachment. FASEB J 2003;17:896–8.
7. Wickstrom SA, Alitalo K, Keski-Oja J. Endostatinassociates with integrin a5h1 and caveolin-1, andactivates Src via a tyrosyl phosphatase-dependentpathway in human endothelial cells. Cancer Res 2002;62:5580–9.
8. Karumanchi SA, Jha V, Ramchandran R, et al. Cellsurface glypicans are low-affinity endostatin receptors.Mol Cell 2001;7:811–22.
9. Shichiri M, Hirata Y. Antiangiogenesis signals byendostatin. FASEB J 2001;15:1044–53.
10. Hanai J, Dhanabal M, Karumanchi SA, et al. Endo-statin causes G1 arrest of endothelial cells throughinhibition of cyclin D1. J Biol Chem 2002;277:16464–9.
11. Wickstrom SA, Alitalo K, Keski-Oja J. Endostatinassociates with lipid rafts and induces reorganization ofthe actin cytoskeleton via down-regulation of RhoAactivity. J Biol Chem 2003;278:37895–901.
12. Hajitou A, Grignet C, Devy L, et al. The antitumoraleffect of endostatin and angiostatin is associated witha down-regulation of vascular endothelial growthfactor expression in tumor cells. FASEB J 2002;16:1802–4.
13. Kim YM, Hwang S, Pyun BJ, et al. Endostatin blocksvascular endothelial growth factor-mediated signalingvia direct interaction with KDR/Flk-1. J Biol Chem2002;277:27872–9.
14. Hanai J, Gloy J, Karumanchi SA, et al. Endostatin is apotential inhibitor of Wnt signaling. J Cell Biol 2002;158:529–39.
15. Kim YM, Jang JW, Lee OH, et al. Endostatin inhibitsendothelial and tumor cellular invasion by blocking the
activation and catalytic activity of matrix metallopro-teinase. Cancer Res 2000;60:5410–3.
16. Nyberg P, Heikkila P, Sorsa T, et al. Endostatin inhibitshuman tongue carcinoma cell invasion and intravasationand blocks the activation of matrix metalloprotease-2, -9,and -13. J Biol Chem 2003;278:22404–11.
17. Lee SJ, Jang JW, Kim YM, et al. Endostatin binds tothe catalytic domain of matrix metalloproteinase-2.FEBS Lett 2002;519:147–52.
18. Abdollahi A, Hahnfeldt P, Maercker C, et al. Endo-statin’s antiangiogenic signaling network. Mol Cell 2004;13:649–63.
19. Hohenester E, Sasaki T, Olsen BR, Timpl R. Crystalstructure of the angiogenesis inhibitor endostatin at1.5 A resolution. EMBO J 1998;17:1656–64.
20. Ding YH, Javaherian K, Lo KM, et al. Zinc-dependentdimers observed in crystals of human endostatin. ProcNatl Acad Sci U S A 1998;95:10443–8.
21. Sasaki T, Larsson H, Kreuger J, et al. Structural basisand potential role of heparin/heparan sulfate binding tothe angiogenesis inhibitor endostatin. EMBO J 1999;18:6240–8.
22. Javaherian K, Park SY, Pickl WF, et al. Lamininmodulates morphogenic properties of the collagen XVIIIendostatin domain. J Biol Chem 2002;277:45211–8.
23. O’Reilly MS, Holmgren L, Shing Y, et al. Angiostatin: anovel angiogenesis inhibitor that mediates the suppres-sion of metastases by a Lewis lung carcinoma. Cell1994;79:315–28.
24. Bergers G, Javaherian K, Lo KM, Folkman J,Hanahan D. Effects of angiogenesis inhibitors onmultistage carcinogenesis in mice. Science 1999;284:808–12.
25. Miao WM, Seng WL, Duquette M, et al. Thrombo-spondin-1 type 1 repeat recombinant proteins inhibittumor growth through transforming growth factor-h-dependent and -independent mechanisms. Cancer Res2001;61:7830–9.
26. Miles AA, Miles EM. Vascular reactions to histamine,histamine-liberator and leukotaxine in the skin ofguinea-pigs. J Physiol 1952;118:228–57.
27. Marshall E. Cancer therapy. Setbacks for endostatin.Science 2002;295:2198–9.
28. Boehm T, O’Reilly MS, Keough K, et al. Zinc-binding of endostatin is essential for its antiangio-genic activity. Biochem Biophys Res Commun 1998;252:190–4.
29. Wickstrom SA, Alitalo K, Keski-Oja J. An endostatin-derived peptide interacts with integrins and regulatesactin cytoskeleton and migration of endothelial cells.J Biol Chem 2004;279:20178–85.
30. Short SM, Derrien A, Narsimhan RP, et al. Inhibitionof endothelial cell migration by thrombospondin-1type-1 repeats is mediated by h1 integrins. J Cell Biol2005;168:643–53.
31. Satchi-Fainaro R, Mamluk R, Wang L, et al. Inhibitionof vessel permeability by TNP-470 and its polymerconjugate, caplostatin. Cancer Cell 2005;7:251–61.
32. van Nieuw Amerongen GP, van Delft S, Vermeer MA,Collard JG, van Hinsbergh VW. Activation of RhoA bythrombin in endothelial hyperpermeability: role of Rhokinase and protein tyrosine kinases. Circ Res 2000;87:335–40.
33. Yamaguchi N, Anand-Apte B, Lee M, et al. Endostatininhibits VEGF-induced endothelial cell migration andtumor growth independently of zinc binding. EMBO J1999;18:4414–23.
34. Sim BK, Fogler WE, Zhou XH, et al. Zinc ligand-disrupted recombinant human Endostatin: potentinhibition of tumor growth, safety and pharmacokineticprofile. Angiogenesis 1999;3:41–51.
35. Wen W, Moses MA, Wiederschain D, Arbiser JL,Folkman J. The generation of endostatin is mediated byelastase. Cancer Res 1999;59:6052–6.
36. Felbor U, Dreier L, Bryant RA, et al. Secretedcathepsin L generates endostatin from collagen XVIII.EMBO J 2000;19:1187–94.
37. Cattaneo MG, Pola S, Francescato P, Chillemi F,Vicentini LM. Human endostatin-derived syntheticpeptides possess potent antiangiogenic propertiesin vitro and in vivo . Exp Cell Res 2003;283:230–6.
38. Chillemi F, Francescato P, Ragg E, et al. Studies onthe structure-activity relationship of endostatin:synthesis of human endostatin peptides exhibitingpotent antiangiogenic activities. J Med Chem 2003;46:4165–72.
39. Morbidelli L, Donnini S, Chillemi F, Giachetti A,Ziche M. Angiosuppressive and angiostimulatoryeffects exerted by synthetic partial sequences ofendostatin. Clin Cancer Res 2003;9:5358–69.
40. Cho H, Kim WJ, Lee YM, et al. N-/C-terminal deletedmutant of human endostatin efficiently acts as an anti-angiogenic and anti-tumorigenic agent. Oncol Rep 2004;11:191–5.
41. Reiher FK, Volpert OV, Jimenez B, et al. Inhibition oftumor growth by systemic treatment with thrombo-spondin-1 peptide mimetics. Int J Cancer 2002;98:682–9.
42. Maeshima Y, Yerramalla UL, Dhanabal M, et al.Extracellular matrix-derived peptide binds to a(v)h(3)integrin and inhibits angiogenesis. J Biol Chem 2001;276:31959–68.
43. Kuo CJ, LaMontagne KR Jr, Garcia-Cardena G, et al.Oligomerization-dependent regulation of motility andmorphogenesis by the collagen XVIII NC1/endostatindomain. J Cell Biol 2001;152:1233–46.
44. Wickstrom SA, Veikkola T, Rehn M, et al. Endostatin-induced modulation of plasminogen activation withconcomitant loss of focal adhesions and actin stressfibers in cultured human endothelial cells. Cancer Res2001;61:6511–6.
Antitumor Activity of Endostatin Peptide
www.aacrjournals.org 3663 Cancer Res 2005; 65: (9). May 1, 2005
Research. on April 21, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

2005;65:3656-3663. Cancer Res Robert M. Tjin Tham Sjin, Ronit Satchi-Fainaro, Amy E. Birsner, et al. Responsible for Its Antitumor Activity-Terminal Zinc-Binding Domain of Endostatin Is2
A 27-Amino-Acid Synthetic Peptide Corresponding to the NH
Updated version
http://cancerres.aacrjournals.org/content/65/9/3656
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/65/9/3656.full#ref-list-1
This article cites 42 articles, 27 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/65/9/3656.full#related-urls
This article has been cited by 13 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://cancerres.aacrjournals.org/content/65/9/3656To request permission to re-use all or part of this article, use this link
Research. on April 21, 2020. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from