Embed Size (px)
Citation preview

Review of literature
5
Numerous definitions of viruses have been proposed from time to time, the definition by
Hull (2002) reads as: “A virus is a set of one or more nucleic acid template molecules,
normally encased in a protective coat or coats of protein or lipoprotein, that is able to
organize its own replication only within suitable host cells. It can usually be horizontally
transmitted between hosts. Within such cells, virus replication is (1) dependent on the
host’s protein-synthesizing machinery, (2) organized from pools of the required materials
rather than by binary fission, (3) located at sites that are not separated from the host cell
contents by a lipoprotein bilayer membrane, and (4) continually giving rise to variants
through various kind of change in the viral nucleic acid”.
The earliest known written record of a viral disease is available in a Japanese poem
written by Empress Koken in 752 AD, referring to leaf yellowing of Eupatorium
lindleyanum, a plant very susceptible to a virus disease (Hull, 2002). Later on these
symptoms were shown to be caused by Eupatorium yellow vein virus (a geminivirus) and a
betasatellite component (Saunders et al., 2003).
2.1 GEMINIVIRUSES
Plant viruses are divided into two groups: RNA and DNA viruses. Geminiviruses have
DNA genomes and cause significant losses to a wide variety of economically important
crops worldwide, including monocotyledonous and dicotyledonous plant species (Hull,
2002; Gutierrez, 2000). Geminiviruses come under the family Geminiviridae. In this
chapter, a brief summary of geminivirus taxonomy, genome organization, replication,
transcription and functions of various genes are presented. The chapter also summarizes the
emerging begomovirus problems of solanaceous crops in India, recent advancements in
geminivirus characterization and understanding of RNA silencing suppression mechanism.
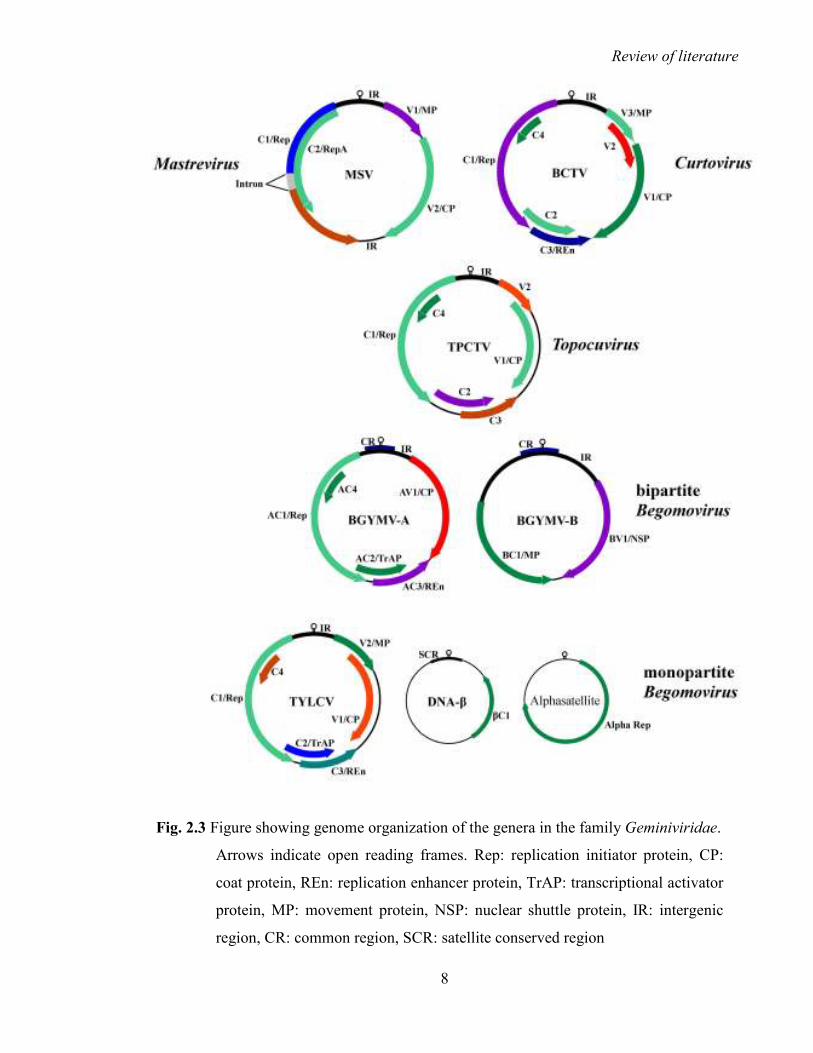
2.1.1 The family Geminiviridae: Taxonomy and genome organization
Members of the family Geminiviridae have small single-stranded circular DNA genomes
(2.5-3.0 kb in size) and have distinct twin-icosahedra shaped particles of approximately
18x30 nm in size (Fig. 2.1). The name geminivirus has been derived from the word

Review of literature
6
‘Gemini’, the sign of the zodiac symbolized by twins (Harrison et al., 1977), and represents
the unique particle shape of this family of viruses. The family is divided into four genera;
Mastrevirus, Curtovirus, Topocuvirus and Begomovirus (Fig. 2.2 and 2.3), based on
genome structure, insect vector and host range (Rybicki et al., 2000; Fauquet and Stanley,
2003; Stanley et al., 2005).
2.1.1.1 Mastrevirus
Maize streak virus (MSV) is the type member of the genus Mastrevirus. It contains a
monopartite genome encoding four open reading frames (ORFs; Fig 2.3). It contains two
intergenic regions (IRs) which are essential for genome replication (Fauquet and Mayo
2001). All the members are transmitted by leafhoppers, infect monocotyledonous plants
except Bean yellow dwarf virus (BeYDV) and Tobacco yellow dwarf virus (TYDV), and
are distributed in the Old World (Eastern Hemisphere, Europe, Africa, Asia and Australia)
(Fauquet and Mayo 2001; Stanley et al., 2005). They are transmitted in a non-propagative,
persistent and circulative manner, i.e. once the insect acquires the virus after feeding on the
infected plant, transmission can occur within hours and continue for the entire life of the
insect (Harrison, 1985). The replication initiator protein (Rep) contains an intron and
undergoes post-transcriptional splicing event, unlike all other geminiviruses. The
phenomenon is essential for Mastrevirus replication and gene expression in monocot plants
(Rojas et al., 2005a).
2.1.1.2 Curtovirus
Beet curly top virus (BCTV) is the type member of the genus Curtovirus. Curtoviruses are
transmitted by leafhoppers (Circulifer tenellus) in a persistent circulative manner to
dicotyledonous plants of both the New (The Americas) and Old World (Briddon and
Markham, 2001; Fauquet and Mayo 2001). The typical Curtovirus genome is monopartite
with seven ORFs and a single IR (Fig. 2.3) containing the origin of replication (ori;
Hormuzdi and Bisaro, 1993).

Review of literature
7
Fig. 2.2 Classification of the family Geminiviridae based on the transmitting vector, host
range and genome organization
Fig. 2.1 Cartoon showing the geminate particle morphology of geminiviruses
Adapted from: http://expasy.org/viralzone/all_by_species/111.html
Review of literature
8
Fig. 2.3 Figure showing genome organization of the genera in the family Geminiviridae.
Arrows indicate open reading frames. Rep: replication initiator protein, CP:
coat protein, REn: replication enhancer protein, TrAP: transcriptional activator
protein, MP: movement protein, NSP: nuclear shuttle protein, IR: intergenic
region, CR: common region, SCR: satellite conserved region

Review of literature
9
2.1.1.3 Topocuvirus
Tomato pseudo-curly top virus (TPCTV) is the type member of the genus Topocuvirus and
is the only member of this genus (Stanley et al., 2005). The sequence analysis of TPCTV
reveals that it has features common to both mastreviruses and begomoviruses, suggesting
that this virus is evolved from a natural recombination between the two genera (Fauquet
and Mayo, 2001). Its genome organization is similar to monopartite begomoviruses with
six ORFs and a single IR containing the ori (Fig. 2.3). It is transmitted by a treehopper to
the dicotyledonous plants in the New World (Briddon et al., 1996).
2.1.1.4 Begomovirus
Begomovirus is the largest and economically most important genus of the family
Geminiviridae. The type member of the genus is Bean golden mosaic virus (BGMV). Its
species are transmitted by the whitefly Bemisia tabaci (Gennadius) in a persistent,
circulative manner and infect dicotyledonous plants (Hanley-Bowdoin et al., 1999;
Harrison and Robinson, 2002). They cause several of the most devastating plant diseases
including tomato leaf curl, cassava mosaic, cotton leaf curl, bean golden mosaic with losses
estimated up to several billion dollars (Varma and Malathi, 2003). The genomes of
begomoviruses typically consist of one or two circular, single-stranded DNA components
of 2.5 to 3 kb in size, known as DNA-A and DNA-B (Fig. 2.3; Navot et al., 1991; Mayo
and Pringle, 1998). The DNA-A component is capable of autonomous replication and can
produce virus particles but requires DNA-B for efficient systemic infection. The two
components of bipartite begomoviruses share an approximately 200 nt long highly
conserved common region (CR), containing repetitive sequence-specific Rep binding
motifs called iterons and a nonanucleotide stem-loop structure (TAATATT↓AC),
essentially required for replication of the viral genome (Moffat, 1997; Fauquet et al., 2003).
Most of the monopartite begomoviruses are associated with approximately 1.4 kb
sized satellite molecules known as betasatellites (DNA-β; Fig. 2.3), which are essential for
symptom development but depend entirely upon the helper virus for their replication,
encapsidation, cell to cell movement and whitefly transmission (Briddon et al., 2001;
Briddon and Stanley, 2006; Mansoor et al., 2003; Saunders et al., 2004; Bull et al., 2004;
Stanley, 2004). Betasatellites are confined to the Old World and are specifically associated

Review of literature
10
with their helper components, irrespective of hosts and geographical distribution (Briddon
et al., 2003). Betasatellites encode a single βC1 gene (13 kDa protein) in the
complementary strand, contain an adenine-rich region (~240 nts) and a satellite-conserved
region (SCR) of nearly 220 nts. The SCR is highly conserved among all betasatellites (Fig.
2.3). Betasatellite does not share any sequence homology with the helper virus but it has a
similar nona-nucleotide (TAATATT↓AC) stem-loop structure (Briddon et al., 2003).
Betasatellites generally have a loose specificity of trans-replication by different helper
components (Saunders et al., 2008).
Some nanovirus-like DNA components known as alphasatellites (DNA1) have also
been reported with many monopartite begomovirus disease complexes occurring in the Old
World. Alphasatellites are self-replicating molecules but depend entirely upon the helper
virus for movement, encapsidation and vector transmission (Briddon et al., 2004). The
genome organization of alphasatellites is highly conserved (Fig. 2.3), encoding a replication
associated protein (~36 kDa) and an adenine-rich region (~200 nts). They have a conserved
nona-nucleotide origin of replication (TAGTATT↓AC), similar to the one found in
nanoviruses (Briddon et al., 2004).
Nomenclature and classification of begomoviruses
Due to emergence of new strains and species of begomoviruses, their taxonomy and
nomenclature is becoming increasingly complex. A group of geminivirologists (ICTV
Geminiviridae Study Group) review the scenario of geminiviruses at regular intervals and
publish the updates related to their nomenclature and taxonomy (Fauquet et al., 2000, 2003,
2008; Fauquet and Stanley, 2003, 2005). The system and guidelines proposed by this group
are followed by a large number of geminivirologists in the world. In 2003, this group
proposed a threshold of 89% nucleotide identity between complete DNA-A nucleotide
sequences of begomoviruses for species demarcation (Fauquet et al., 2003).

Review of literature
11
Recent and most important criteria for the classification of begomovirus isolates in
variants, strains and species (Fauquet et al., 2008) are as follows:
1. After pairwise sequence comparison of a new begomovirus isolate with all known
sequences representative of species, using ClustalV algorithm;
� if its nucleotide identity is <88%, it belongs to a new species.
� if its nucleotide identity =88-89%, it belongs tentatively to the closest species.
� if its nucleotide identity >89%, it belongs definitively to that species.
2. After pairwise sequence comparison of a new begomovirus isolate with all known
sequences representative of strains and variants in the identified species, using ClustalV
algorithm;
� if its nucleotide identity <93% to all known members, it is a member of a new
strain in that species,
� if its nucleotide identity >94% to an existing isolate, it is a variant of that strain in
that species.
Virus names for descriptors under the species level should adopt the following
nomenclature structure (Fauquet et al., 2008):
Species name, strain descriptor (symptoms, location, host, if appropriate or a letter such
as A, B, C) [variant descriptor (country: location: [host]: year)]
Example: Mungbean yellow mosaic India virus-[India:Varanasi:Dolichos:2008]
2.1.1.5 Recently described divergent geminiviruses
Beet curly top Iran virus (BCTIV) has been reported as a distinct curtovirus in Iran. BCTIV
has an unusual TAAGATTCC virion strand replication origin nonanucleotide. The Rep
protein of this virus resembles to those of mastreviruses (having an intron) and the coat
protein (CP) is a typical curtovirus-like protein. It has been predicted that the genome of
BCTIV may have arisen by a recombination event involving curto- and mastrevirus
ancestors (Yazdi et al., 2008). Eragrostis curvula streak virus (ECSV) is another divergent
geminivirus reported from South Africa having an unusual TAAGATTCC virion strand
replication origin nonanucleotide similar to BCTIV. ECSV resembles a chimaera of
geminiviruses from different genera having transcription and replication origin structurally

Review of literature
12
more similar to those found in begomoviruses and curtoviruses than it is to those found in
mastreviruses. ECSV transcription activator protein (TrAP) gene is most similar to that
found in begomoviruses. Moreover ECSV has mastrevirus-like CP gene and two IRs.
ECSV genome is not recombinant, suggesting that its common features with other
geminiviruses might probably be present within the last common ancestor of these viruses
(Varsani et al., 2009). Turnip curly top virus (TCTV) is another recently described
divergent geminivirus from Iran, having a genome arrangement in the complementary-
sense similar to that of curtoviruses but in the virion-sense it encodes only two ORFs (the
curtoviruses encode three). The complementary-sense genes are homologous to those of
curtoviruses but show little sequence identity with them. TCTV features suggest that it is a
highly divergent geminivirus with similarities to viruses of the genus curtovirus (Briddon et
al., 2010). These divergent geminiviruses are important links to the understanding of their
evolution.
2.1.2 Evolution of geminiviruses
The greatest molecular diversity of geminiviruses and their associated satellites is found in
Southeast Asia suggesting that this area might be the center of origin of these viruses.
Probably, spread of geminivirus diseases has been promoted by domestication of crops
from reservoir plants and their extensive cultivation (Nawaz-ul-Rehman and Fauquet,
2009). It is believed that geminiviruses may have been evolved as episomal DNA replicons
of primeval prokaryotic organisms that later adapted to primitive eukaryotic progenitors of
modern plants (Rojas et al., 2005a). The hypothesis is supported by several evidences like
conserved features of the Rep protein of the contemporary eukaryotic and prokaryotic DNA
replicons (Ilyina and Koonin, 1992; Campos-Olivas et al., 2002), ability of the
geminiviruses to replicate in Agrobacterium tumefaciens and their polycistronic mRNAs
(Selth et al., 2002). During evolution of these replicons and their hosts, they might have
acquired new features due to recombination with host genome and/or other replicons (Rojas
et al., 2005a). According to one hypothesis, all geminiviruses were originally monopartite,
infected only monocots and were transmitted by leafhoppers (Rybicki, 1994; Padidam et
al., 1995a). Further evolution facilitated their transmission by whiteflies which could also
transmit them to dicot plants. The acquisition of DNA-B might have occurred later, perhaps

Review of literature
13
before the continental separation, as bipartite begomoviruses are present in Old as well as
New world (Rojas et al., 2005a). Two begomoviruses have been reported from Vietnam,
which are more closely related to New World than to Old World geminiviruses, inferring
the interesting hypothesis that the New World geminiviruses developed in the Old World
prior to continental separation and thereafter they spread and expanded their population in
New World (Ha et al., 2006, 2008). Recombination has been the major factor responsible
for geminivirus diversification, probably due to their recombination-dependent replication
along with their frequent natural mixed infections (Preiss and Jeske, 2003; Padidam et al.,
1999; Pita et al., 2001; Ribeiro et al., 2003). The fact that two begomoviruses can replicate
in a single nucleus, further strengthens this hypothesis (Morilla et al., 2004). Genomic
properties of curtoviruses suggest that they might have evolved from recombination
between mastreviruses and begomoviruses (Harrison and Robinson, 1999). Topocuvirus
also suggest to have originated due to recombination between curtoviruses and a genetically
very different virus, not related with present geminiviruses (Briddon et al., 1996).
Association of begomoviruses with alpha- and betasatellite molecules may be helping them
to adapt and infect new hosts (Briddon et al., 2003, 2004).
2.1.3 Geminivirus replication
Replication of geminiviruses takes place in the nucleus and is dependent upon the host
machinery. Replication is accomplished through double stranded DNA (dsDNA)
intermediates using rolling circle replication (RCR) and recombination-dependent
replication (RDR) mechanisms (Sunter and Bisaro, 1991; Gutierrez, 1999; Jeske et al.,
2001; Preiss and Jeske, 2003). The RCR cycle can be divided into three stages: in the first
stage, a covalently closed dsDNA intermediate also known as replicating form (RF) is
generated by synthesis of a complementary-sense (minus) strand using virion-sense (plus)
strand as a template. The process involves a Rep protein with a nicking-closing activity in a
host-directed, RNA primed synthesis of the complementary strand. The RF acts as a
template for viral transcription as well as further replication (Hanley-Bowdoin et al., 1999).
In the second stage, additional RF DNAs are generated, initiated by viral Rep protein by
nicking the plus strand at a specific sequence (TAATATT↓AC). After the cleavage, the
Rep protein covalently binds to the 5' terminus via a phosphotyrosine linkage. The 3' -OH

Review of literature
14
terminus is used as a primer for the synthesis of nascent plus strand. This nascent plus
strand displaces the parental plus strand from the intact minus-strand template. The process
utilizes host replication proteins. The origin of replication is regenerated on completion of
the nascent plus strand, which again is nicked by Rep to release the displaced unit-length
plus strand, which is simultaneously ligated to circular form by the closing activity. Rep is
again transferred to the newly created 5' terminus (Gutierrez, 1999). The third stage of RCR
occurs late in the replication cycle. In this stage, accumulation of the viral genomes takes
place for encapsidation. Priming is prevented and ssDNA is the predominant product
(Bisaro, 1996).
Some defective DNA molecules may also accumulate during improper replication,
which may interfere with the replication of the parent virus and attenuate symptom
development (Patil and Dasgupta, 2006). For this reason, they are also known as defective
interfering (DI) DNAs. They are usually half the size of viral DNA (~1,300 bp) and retain
mainly the left side of the DNA component, including the viral ori (Jeske, 2009). A number
of evidences indicate that viral DNA replication might be coupled to the cell cycle
regulatory network of the infected cell. Therefore, geminiviruses are excellent model
systems to understand the regulation of DNA replication and cell cycle in plant cells
(Gutierrez, 1999).
2.2 BEGOMOVIRUS TRANSCRIPTION AND PROTEIN FUNCTIONS
Transcription takes place in the nucleus and begomovirus genes are transcribed from
dsDNA intermediates. Transcription is bidirectional i.e. proteins are encoded either in
virion sense or the complementary sense from the common region that contains the
bidirectional promoter sequences. Transcription frequently gives rise to multiple
overlapping polycistronic mRNAs. The virion sense and the complementary sense
transcripts overlap by 4 bp at their 3’ ends (Hanley-Bowdoin et al., 1999; Chellappan et al.,
2004; Shivaprasad et al., 2005).
2.2.1 Functions of AV1 or V1 gene (coat protein)
AV1 encodes the only structural protein of the begomoviruses i.e. the coat protein (CP). CP
is ~30 kDa sized multifunctional protein, which forms multimers resulting in the geminate

Review of literature
15
particle and encapsidates the viral ssDNA (Harrison et al., 2002; Hallan and Gafni, 2001;
Unseld et al., 2004). The CP acts as determinant of vector specificity and plays an
important role in transmission of the virus within and between the plant hosts (Briddon et
al., 1990; Azzam et al., 1994; Qin and Petty, 2001) and in case of monopartite
begomoviruses, CP is essentially required for the systemic movement of the virus and
accumulation of viral ssDNA (Azzam et al., 1994; Wartig et al., 1997; Noris et al., 1998;
Qin et al., 1998; Gafni and Epel, 2002). CP of tomato leaf curl Sardinia virus interacts with
a member of the small heat-shock protein family (termed BtHSP16). CP is also reported to
interact with AC1 gene and down regulate its expression, indicating its probable role in
controlling geminiviral DNA replication (Malik et al., 2005). CP interaction is also
reported with host karyopherin (importin) α which suggests that it might be utilizing this
pathway for docking the viral particles to the nucleus in infected cells (Gafni and Epel,
2002; Guerra-Peraza et al., 2005). Nuclear shuttle protein (NSP) plays a role in cell-to-cell
and long distance movement of the virus but if it is mutated, CP is shown to complement
the respective defects. Probably, intimate interaction of both these proteins is required for
efficient trafficking (Qin et al., 1998; Carvalho and Lazarowitz 2004; Carvalho et al. 2006).
For all monopartite begomoviruses, CP is essential for the viral long distance movement
(Gafni and Epel, 2002).
2.2.2 Functions of AV2 or V2 gene (pre-coat protein)
The AV2 or V2 gene (pre-coat protein; ~13.5 kDa) is only present in Old World
begomoviruses. It is called pre-coat protein (PCP) because of its genomic position as it is
located in front of CP (Padidam et al., 1996; Harrison and Robinson, 1999). In case of
monopartite begomoviruses, the PCP has been implicated in symptom development and
ssDNA accumulation (Rigden et al., 1993). In Tomato leaf curl New Delhi virus
(ToLCNDV), PCP is involved in systemic movement of the virus (Padidam et al., 1996). In
TYLCV, PCP is involved in viral DNA export from the nucleus to the plasmodesmata
(Rojas et al., 2001). PCPs of monopartite TYLCV and bipartite Indian cassava mosaic
virus (ICMV) were targeted to the cell periphery and indicated plasmodesmal localization
particularly in plasmolysed cells (Selth et al., 2006; Rothenstein et al., 2007). TYLCV-
Israel and Tomato leaf curl Java virus PCPs are also reported to be suppressors of RNA

Review of literature
16
silencing (Zrachya et al., 2007; Sharma and Ikegami, 2010) and its interaction with host
SGS3 is required for the suppression activity (Glick et al., 2008).
2.2.3 Functions of AC1 or C1 gene (Rep protein)
The AC1 or C1 protein (Rep) is implicated in replication of both DNA-A and DNA-B
components and is responsible for the initiation of RCR. It is ~40 kDa sized highly
conserved protein, localized in the nuclei of the infected plant cells (Gutierrez, 1999; Elmer
et al., 1988 Hanley-Bowdoin et al., 1999). The protein has a nucleoside triphospahte
binding site and binds to highly specific viral DNA sequences (iterons), located in the
conserved CR, repressing its own promoter (Sunter et al., 1993; Fontes et al., 1994; Eagle
et al., 1994; Desbiez et al., 1995; Chatterji et al., 2000). The protein has a DNA
nicking/ligating property and it can also act as a DNA helicase (Laufs et al., 1995a; Pant et
al., 2001; Choudhury et al., 2006). Interaction of the Rep protein with itself and with a
number of host proteins has been reported (Orozco et al., 1997; Ach et al., 1997; Luque et
al., 2002; Kong and Hanley-Bowdoin, 2002; Castillo et al., 2003; Castillo et al., 2004).
Rep is shown to interact with the host retinoblastoma homologue protein, required in cell
cycle and differentiation (Arguello-Astorga et al., 2004). This interaction is helpful in
regulating the cell cycle of mature plant cells for supporting viral DNA replication and
infection (Kong et al., 2000). During infection, AC1 down-regulates transcription of
complementary sense genes AC2 and AC3 (Sunter et al., 1993). Rep can form homo- and
hetero-oligomers and interact with the viral replication enhancer protein (REn) which
enhances viral DNA accumulation. It also interacts with proliferating cell nuclear antigen
(PCNA), a conserved plant protein and an important replication factor. Hence, efficient
begomovirus replication requires the formation of Rep, REn and PCNA complexes (Sunter
et al., 1990; Settlage et al., 1996; Castillo et al., 2003; Bagewadi et al., 2004; Settlage et
al., 2005).
2.2.4 Functions of AC2 or C2 gene (Transcriptional activator protein; TrAP)
The TrAP is ~15 kDa protein, functions as a regulator of viral gene expression and is
essential for efficient transcription of the late expressing viral genes (AV1 and BV1)
located in positive sense orientation. TrAP activates CP promoter in mesophyll cells but

Review of literature
17
derepresses it in phloem tissue, suggesting that its regulation of transcription is specific to
cell types (Sunter and Bisaro 1991, 1992, 1997; Hartitz et al., 1999). It has been reported to
be a suppressor of RNA silencing in a number of begomoviruses (Voinnet et al., 1999; van
Wezel et al., 2001; Dong et al., 2003; Selth et al., 2004; Vanitharani et al., 2004; Trinks et
al., 2005; Wang et al., 2005a; Kon et al., 2007; Gopal et al., 2007). Plants over-expressing
TrAP are more susceptible to viral infection (Sunter et al., 2001). TrAP localizes in the
nucleus, containing a putative nuclear localization signal consisting of five residues
(PRRRR), which are essential for suppression of RNA silencing (Chowda-Reddy et al.,
2009). In case of Tomato leaf curl virus-India, AC2 is reported to be essential for
infectivity of the virus (Padidam et al., 1995b). TrAP of tomato golden mosaic virus
(TGMV) binds non-specifically to ssDNA and weakly interacts with dsDNA, which
suggests that this protein might be interacting with host proteins for transcriptional
activation. It consists of three functional domains viz. nuclear localization signal domain at
the N-terminus, DNA-binding Zinc-finger domain in the centre and an acidic activator
domain at the C-terminal (Hartitz et al., 1999). TrAP has an ability to interact with, and
inactivate, serine/threonine-related kinases such as sucrose non-fermenting 1 protein kinase
(SNF1) and adenosine kinases (ADK). SNF1 and ADK are involved in host defense
response and mediate various plant anti-stress pathways (Hao et al., 2003; Wang et al.,
2003). Arabidopsis PEAPOD2 (PPD2) transcription factor interacts with TrAP and its CP
promoter, suggesting that TrAP is targeted to the CP promoter through interaction with
PPD2, leading to activation of CP gene expression (Lacatus and Sunter, 2009). It has been
studied that TrAP of mungbean yellow mosaic virus transactivates the viral promoter-
driven transgene and causes toxicity in transgenic tobacco plants. All three domains of
TrAP are essential for transactivation, suppression of gene silencing and the toxic effect
(Rajeswaran et al., 2007).
2.2.5 Functions of AC3 or C3 gene (Replication enhancer protein; REn)
REn is ~16 kDa nuclear targeted protein that enhances the virus replication (Sunter et al.,
1994). REn forms homo-oligomers and interacts with Rep to enhance its ATPase activity,
probably resulting in replication enhancement. It also interacts with at least two host-
encoded proteins i.e. PCNA and retinoblastoma-related protein (pRBR). Begomovirus

Review of literature
18
replication is dependent upon the host-encoded polymerases and replications factors, and it
is believed that an interaction between Rep, REn, pRBR and PCNA reprograms the plant
cells into S phase where these enzymes are abundantly available (Settlage et al., 2001;
Nagar et al., 2002; Castillo et al., 2003). This interaction also enhances the recognition of
ori and probably PCNA recruits various host cell factors required for the replication. REn is
essential for efficient viral replication in differentiated cells and is also required for optimal
replication in undifferentiated cells (Settlage et al., 2005). The enhancement of replication
is also thought to be achieved by an interaction between REn and a transcription factor
involved in the induction of SINAC1 expression, a class of proteins that are often induced
upon pathogen infection (Selth et al., 2005).
2.2.6 Functions of AC4 or C4 gene
AC4 or C4 is ~12 kDa protein and its ORF is completely embedded in AC1/C1 coding
region but in a different frame (Hanley-Bowdoin et al., 1999). The amino acid sequence of
this protein is the least conserved among all begomovirus proteins. Early studies with
TGMV, African cassava mosaic virus (ACMV), Bean golden mosaic virus (BGMV) and
Potato yellow mosaic virus (PYMV) showed that C4 protein is not essential for viral
replication and symptom development (Elmer et al., 1988; Etessami et al., 1991; Sung and
Coutts, 1995; Hoogstraten et al., 1996; Pooma and Petty, 1996). But, mutagenesis studies
with Tomato yellow leaf curl Sardinia virus (TYLCSV) and Tomato leaf curl virus
(ToLCV) revealed that C4 protein is an essential pathogenicity factor (Jupin et al., 1994;
Rigden et al., 1994; Krake et al., 1998; Selth et al., 2004). In case of TYLCV with a mutant
C4 gene, the virus was unable to move systemically in tomato plants suggesting its role in
virus movement (Jupin et al., 1994). In case of TGMV, C4 protein plays a role in
regulation of Rep expression (Gröning et al., 1994). In case of ACMV, Sri Lankan cassava
mosaic virus (SLCMV), Bhendi yellow vein mosaic virus (BYVMV) and East African
cassava mosaic Cameroon virus (EACMCV), C4 protein has been reported as suppressor
of RNA silencing (Vanitharani et al., 2004, 2005; Fondong et al., 2007; Gopal et al., 2007).
But in case of East African cassava mosaic virus (EACMV) and Indian cassava mosaic
virus (ICMV), the respective AC2 proteins rather than the AC4 proteins are the RSSs,

Review of literature
19
suggesting that AC4 and AC2 proteins probably target different steps in the RNA silencing
pathways (Vanitharani et al., 2004; Chellappan et al., 2005).
2.2.7 Functions of BV1 and BC1 genes
ORF BV1 encodes for ~29 kDa nuclear shuttle prtein (NSP) while BC1 encodes for a ~34
kDa movement protein (MP). Both NSP and MP coordinate the movement of the viral
DNA from nucleus to other plant cells (Noueiry et al., 1994; Sanderfoot and Lazarowitz,
1995, 1996; Gafni and Epel, 2002). NSP transfers viral DNA between the nucleus and
cytoplasm, whereas MP facilitates transport between cells through plasmodesmata and
long-distance via phloem. NSP serves its function by localizing itself in the nucleus,
nucleolus and cell periphery of the infected cells (Rojas et al., 2001; Zhang et al., 2001;
Kleinow et al., 2009). It is reported that a NSP-interacting GTPase (NIG) interacts with
NSP in vitro and in vivo to redirect the viral protein from the nucleus to the cytoplasm
(Carvalho et al., 2008a). When co-expressed with MP, NSP is translocated to the nucleus
of the adjacent cells. The co-operation of both NSP and MP is direct and does not need any
plant-specific protein (Frischmuth et al., 2007). NSP and MP can bind to the ssDNA and
dsDNA based upon its size, regardless of its sequence (Rojas et al., 1998; Lazarowitz,
1999; Radhakrishnan et al., 2008). NSP packages the viral DNA and interacts with BC1 in
the cytoplasm to transport the DNA into the neighboring cells through the plasmodesmata
(Lazarowitz and Beachy, 1999; Hehnle et al., 2004). A tomato kinase protein and its
soybean homolog are reported to interact with NSP of TGMV and Tomato crinkle leaf
yellows virus (Mariano et al., 2004). Both NSP and MP of different bipartite
begomoviruses are reported as virulence or avirulence determinants in various host plants
(von Arnim and Stanley, 1992; Pascal et al., 1993; Ingham et al., 1995; Duan et al., 1997;
Hou et al., 2000; Saunders et al., 2001; Carvalho and Lazarowitz, 2004; Fontes et al., 2004;
Hussain et al., 2005; Zhou et al., 2007). MP localizes at the cell periphery and increases the
size selection limit of plasmodesmata to facilitate the movement of viral DNA across the
cell (von Arnim et al., 1993; Noueiry et al., 1994; Rojas et al., 2001; Sanderfoot and
Lazarowitz 1995; Zhang et al., 2002). MP has also been reported to localize in specialized
endoplasmic reticulum-derived tubules in developing phloem cells (Ward et al., 1997), and
in the cytoplasmic leaflet of endoplasmic reticulum vesicles and plasma membranes in

Review of literature
20
yeast cells (Aberle et al. 2002). A NSP-interacting kinase (NIK) involved in plant
development and defense, is believed to transduce a defense signal against virus infection
and regulate NSP function (Florentino et al., 2006; Carvalho et al., 2008b; Santos et al.,
2009, 2010). Synaptotagmins (SYTA), calcium sensors that regulates endocytosis, are also
suggested to help MP to transport its cargos to plasmodesmata for cell-to-cell spread via an
endocytic recycling pathway (Lewis et al., 2010).
2.2.8 Functions of betasatellite
Betasatellite is reported to be involved in host range determination and induction of typical
disease symptoms in a number of monopartite begomoviruses (Saunders et al., 2000;
Briddon et al., 2001; Zhou et al., 2003; Jose and Usha 2003; Mansoor et al., 2003; Cui et
al., 2004; Saeed et al., 2005; Briddon and Stanley, 2006; Qazi et al., 2007). Betasatellite
encodes a single protein (βC1) of ~13.7 kDa size, which localizes in the nucleus and cell
periphery, and acts as a suppressor of RNA silencing (Briddon et al., 2003; Mansoor et al.,
2003; Stanley, 2004; Cui et al., 2005; Kumar et al., 2006; Kon et al., 2007; Gopal et al.,
2007). The βC1 protein has an ability to bind to ssDNA or dsDNA without size or sequence
specificity (Cui et al., 2005). Betasatellite molecules can also affect the accumulation of the
helper virus and may help in providing an appropriate cellular environment for efficient
DNA replication (Saunders et al., 2000). Betasatellite can take over the movement function
of DNA-B in case of bipartite begomoviruses (Saeed et al., 2007). Interaction of CP and
βC1 protein has been suggested to play a collaborative role in inter- and intracellular
dynamics of bhendi yellow vein mosaic disease (Kumar et al., 2006). In a study, it has been
shown that betasatellite of tomato leaf curl virus can override the pathogenicity phenotype
of the C4 gene but cannot compensate for loss of function CP and PCP (Saeed et al., 2008).
It is found that efficient gene expression and replication of betasatellite requires an
interaction of the G-box motif with host nuclear factors (Eini et al., 2009).
2.2.9 Functions of alphasatellite (DNA1)
Almost all begomoviruses carrying betasatellite are also associated with the alphasatellite
(DNA1) subviral components (Briddon and Stanley, 2006). These molecules are not
important in the disease process and are not essential for infectivity and symptom induction

Review of literature
21
(Briddon et al., 2004; Wu and Zhou, 2005). However in a recent study, some DNA-2 type
alphasatellites have been reported to encode a pathogenicity determinant that may attenuate
symptoms and reduce betasatellite accumulation (Idris et al., 2011). Replication-associated
protein encoded by two alphasatellites associated with Cotton leaf curl Rajasthan virus
(CLCuRaV) have been reported to interact with the Rep and C4 proteins encoded by their
helper begomovirus, in a yeast two-hybrid assay. Rep proteins encoded by these two
alphasatellites were found to have strong gene silencing suppressor activity, suggesting that
these alphasatellites provide a selective advantage to their helper virus/betasatellite
complex (Nawaz-Ul-Rehman et al., 2010). Alphasatellite associated with tobacco curly
shoot virus (TbCSV) has been used to design a VIGS vector by inserting a multiple cloning
site (MCS) between its Rep protein ORF and the A-rich region. DNA fragments of genes
targeted for silencing can be cloned into this MCS (Huang et al., 2009).
2.3 DETECTION OF GEMINIVIRUSES
Detection of the virus is first and foremost requirement in geminivirus research. Efficient
diagnostic methods are the primary tools, required to combat the increasing incidences and
epidemics of geminiviruses (Haible et al., 2006). Although geminivirus diseases were
known much earlier but viral etiology was recognized only in the 1970s, and a virus with
geminate structure was detected even later. Geminate particles were observed in electron
microscopic (EM) studies of thin sections from TYLCV-infected tomato leaves, indicating
localization in the nuclei of phloem parenchyma cells (Russo et al., 1980; Goodman, 1981;
Cherif and Russo, 1983). EM was the only possible way to detect geminiviruses in those
times. Since then, several diagnostic tools have been developed for mass screening and for
more specific characterization of the virus.
2.3.1 Serological methods
Serological tests in the form of enzyme linked immunosorbent assay (ELISA) have been
widely used for diagnosis of geminiviruses but they have limitations because of the
requirement of sufficient purified coat protein for the production of antisera (Czosnek et al.,
1988a; Credi et al., 1989; Chiemsombat et al., 1991). Moreover, geminiviruses are found in
low concentrations in infected tissue and their particles are extremely fragile, due to which

Review of literature
22
they are very difficult to isolate and purify (Sharma et al., 2004). Using ELISA, it is almost
impossible to resolve mixed infections, for which molecular techniques are necessary.
Initially, polyclonal antibodies were mainly used until the production of monoclonal
antibodies (Harrison et al., 1991 and 2002; Macintosh et al., 1992). Another approach has
also been used for trapping virus particles, in which the ELISA plates are coated with
GroEL protein rather than the virus-specific antibodies (Akad et al., 2004). Immunoblotting
methods in the form of tissue-printing and dot-blotting have also been reported (Hajimorad
et al., 1996; Pico et al., 1999; Dalmon et al., 2000). In comparison with ELISA, blotting
methods have limitations because of relatively high backgrounds and their inability to
distinguish among similar species.
2.3.2 DNA hybridization
DNA hybridization is another widely used method for geminivirus detection (Czosnek et
al., 1988b; Navot et al., 1989; Nakhla et al., 1993; Sharma et al., 2004). In this method,
radiolabelled probes prepared from cloned viral cDNAs are used in Southern blots to detect
the viral DNA in infected plants. A modification in this technique called tissue printing
employs blotting of leaf squashes rather than the viral DNA on to nylon membranes in the
form. These methods have been used for large-scale epidemiological studies and for use in
breeding programmes for virus resistance (Lapidot et al., 1997, 2001; Sanchez-Campos et
al., 1999; Davino et al., 2006). Use of radioactive material for preparing the probes has
been the major limitation of this technique. To avoid this hazardous material, non-
radioactive labelling techniques have been developed (Crespi et al., 1991; Abou Jawdah et
al., 1995; Quiñónez et al., 2004). Usually chemiluminescent substrates are used in non-
radioactive hybridization and probes can be easily removed for reprobing the blot for other
viruses (Caciagli and Bosco, 1996, 1997; Czosnek and Laterrot, 1997). Hybridization
technique employing radioactive or non-radioactive probes has been the best method for
simultaneous analysis of a large number of samples.
2.3.3 Polymerase chain reaction (PCR)
Polymerase chain reaction using specific or degenerate primers, along with restriction
fragment length polymorphism (RFLP), or tissue printing, or immunoblotting or molecular

Review of literature
23
hybridization have been used as common tools for geminivirus diagnostics (Navot et al.,
1992; Rojas et al., 1993; Deng et al., 1994; Briddon and Markham, 1995; Wyatt and
Brown, 1996). Print-capture PCR protocols have also been reported, in which squashes of
plant or whitefly tissue are directly transferred on small pieces of nylon membranes or
Whatman paper and directly used in amplification reactions (Atzmon et al., 1998; Navas-
Castillo et al., 1998). Immunocapture-PCR has also been tried to study transmission of
begomoviruses by whiteflies (Ghanim et al., 2001). Another technique named loop-
mediated isothermal amplification (LAMP), which amplifies DNA with high efficiency
under isothermal conditions without being much influenced by co-presence of non-specific
DNA has also been used for TYLCV detection (Fukuta et al., 2003). Multiplex-PCR has
also been successfully used to identify more than one geminiviruses from a single source
(Potter et al., 2003; Gorsane et al., 2005). These techniques have been constrained by their
higher costs and requirement of prior sequence information of target DNA.
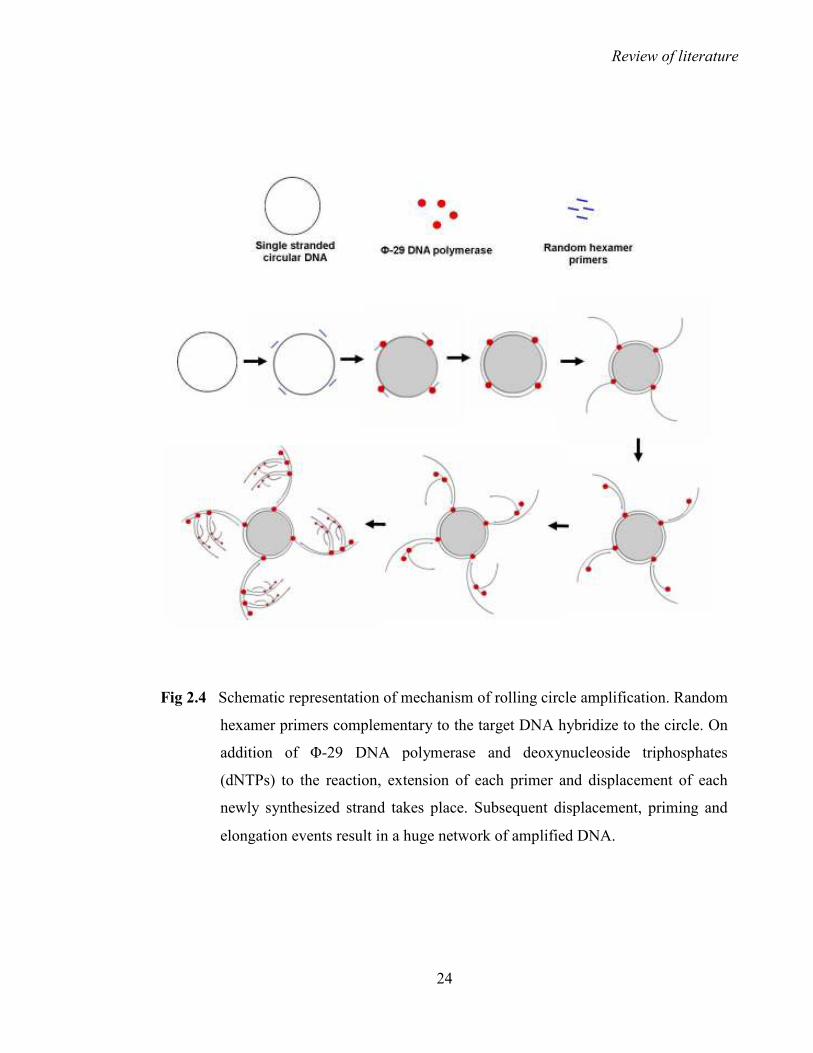
2.3.4 Rolling circle amplification (RCA)
A highly efficient solution to above mentioned problems has been provided by the use of
high fidelity Φ-29 DNA polymerase, which is extracted from the bacteriophage Bacillus
subtillis (Blanco et al., 1989; Dean et al., 2001; Inoue-Nagata et al., 2004; Haible et al.,
2006). This enzyme selectively amplifies only circular DNA and has polymerase, strand
displacement (helicase) and 3’ to 5’ exonuclease (proof reading) activities. With these
properties, it is ideal to amplify geminiviral, nanoviral, circoviral and plasmid DNAs of
small or moderate sizes, through rolling circle amplification (RCA) mechanism. To start
RCA, the enzyme requires a free 3’-OH, which is either provided by a nick in dsDNA or by
primers that are annealed to complementary sequences. Usually random hexamer primers
are used in RCA for an increased efficiency of multiplication. The polymerase extends the
primer continuously around the circular DNA template generating a long DNA product
(Fig. 2.4) that consists of many repeated copies of the circle (Dean et al., 2001).
Review of literature
24
Fig 2.4 Schematic representation of mechanism of rolling circle amplification. Random
hexamer primers complementary to the target DNA hybridize to the circle. On
addition of Φ-29 DNA polymerase and deoxynucleoside triphosphates
(dNTPs) to the reaction, extension of each primer and displacement of each
newly synthesized strand takes place. Subsequent displacement, priming and
elongation events result in a huge network of amplified DNA.

Review of literature
25
2.4 EMERGING BEGOMOVIRUS DISEASES OF SOLANACEOUS CROPS IN
INDIA
Begomoviruses infect a wide range of crops viz. vegetables, cash crops, ornamentals,
tropical fruits and even weeds which sometimes can act as latent hosts of the virus (Varma
and Malathi, 2003). Among these, solanaceous plants are important as food crops and are
grown in a large part of the world and also cultivated throughout India. Hot and humid
climate in some parts of India is suitable for spread of begomoviruses, especially affecting
communities dependent on the produce from these crops as sole source of very valuable
protein component of diet. However, there had been some scattered reports of begomovirus
infections in sub-temperate areas as well (Sharma et al., 2001; Sambyal et al., 2006;
Sharma et al., 2007). Bemisia tabaci, the begomovirus vector is very prevalent in hot and
humid climate; in such conditions dual effect of virus infection and vector
feeding/multiplication is highly damaging to the crops. This section summarizes the
emerging begomoviral disease of solanaceous crops in India.
2.4.1 Tropical and sub-tropical areas
This section describes incidences of emerging begomoviruses infecting solanaceous crops
in last ten years in tropical and sub-tropical parts of India.
2.4.1.1 Tomato
Tomatoes rank fourth among cultivated vegetable crops worldwide with a production of
more than 100 million metric tons a year (Peralta et al., 2005). It is one of the most
important vegetables in India and its cultivation is constrained by tomato leaf curl disease
(ToLCD), which was first reported in Northern India in 1948 (Vasudeva and Sam Raj,
1948). The symptoms induced by begomovirus infection include leaf curling, mosaic,
bright yellow spots on leaves, reduction in leaf size, vein clearing, crinkling, chlorosis,
stunting, reduced or no fruiting and occasional formation of enations (Varma and Malathi,
2003). The disease causes considerable crop losses. In a study, tomato cv. Pusa Ruby, Pusa
Early Dwarf and Maitri showed 66.40, 67.00 and 88.00% reduction in yield respectively
due to ToLCD. Maitri showed the highest disease incidence of 62.16%, while Pusa Ruby
showed the lowest, 58.27% (Shahnaz and Kumar, 2004).

Review of literature
26
In November 2001, tomato plants showing yellowing of leaf lamina, upward leaf
curling, leaf distortion, shrinking of leaf surface, and stunted growth were observed in
tomato-growing areas in Varanasi and Mirzapur districts of eastern Uttar Pradesh, India.
Based on DNA sequences obtained, the causal pathogen was identified as a begomovirus
(Chakraborty et al., 2003a).
In 2002, a new monopartite begomovirus, tomato leaf curl Karnataka virus
(U38239) was characterized from Bangalore, South India, infecting tomatoes. The virus
was transferred to tomato by whitefly biotype B, sap inoculation and particle bombardment
and it produced typical yellowing and leaf curl symptoms. The isolate appeared to be a
recombinant virus (Chatchawankanphanich and Maxwell, 2002).
In 2003, a new bipartite species was found causing a severe leaf curl disease of
tomato in Varanasi (Chakraborty et al., 2003b), Uttar Pradesh, India. The virus was named
as Varanasi isolate of Tomato leaf curl Gujarat virus (ToLCGV-[Var]; AY190290,
AY190291). The virus could be transmitted by grafting and through whitefly transmission
in a persistent manner. ToLCGV-[Var] could infect and cause symptoms in tomato, pepper
(Capsicum annuum cv. California Wonder), Nicotiana benthamiana and N. tabacum (cv.
Xanthi) when partial tandem dimeric constructs of DNA-A and DNA-B were co-inoculated
by particle bombardment. DNA-A alone was also infectious, but symptoms were milder
and took longer to develop (Chakraborty et al., 2003b).
In order to assess variability of tomato leaf curl begomovirus (TLCB) in India,
Reddy et al. tested 69 tomato plants and 34 weeds or neighbouring crops for
begomoviruses, collected from all the major tomato producing areas of India. Eighty-one of
the 103 samples were found positive in PCR using begomovirus genus-specific primers. CP
genes from 29 samples were PCR amplified and characterized. Phylogenetic analyses of the
CP sequences revealed five different TLCB clusters each having <88% identity to the
others. Four clusters represented known Indian begomoviruses, whereas one cluster
contained sequences originating from Haryana State with highest identity (89%) with
Croton yellow vein mosaic virus. Sixty-five begomovirus positive samples were
characterized further by PCR for betasatellite and DNA-B for four Indian TLCB species,
using universal begomovirus primers. The majority of samples represented monopartite
TLCBs associated with betasatellite components. All four known TLCBs appeared to be

Review of literature
27
present throughout India. TLCBs were also found to be present in chilli, cowpea, okra and
tobacco crops and in some common weeds. Papaya leaf curl virus and Pepper leaf curl
Bangladesh virus were also detected in tomato. Mixed begomovirus infections, were
evident in 13 samples (Reddy et al., 2005).
In 2008, isolates of two distinct begomovirus species, the bipartite severe strain of
the species Tomato leaf curl New Delhi virus (ToLCNDV-[IN:ND:Svr:92]) and the
Varanasi strain of the species Tomato leaf curl Gujarat virus (ToLCGV-[IN:Var:01])
infecting tomato, causing severe yield losses in northern India were characterized. This
study also investigated the infectivity properties of genomic components of these two
species. Both were infectious in N. benthamiana, N. tabacum and L. esculentum.
Pathogenicity was enhanced when DNA-A of ToLCNDV-[IN:ND:Svr:92] was trans-
complemented with ToLCGV-[IN:Var:01] DNA-B, and increase in accumulation of
ToLCGV-[IN:Var:01] DNA-B was consistently observed with it. A synergistic interaction
between these two viruses was also evident in the study. This was the first report
demonstrating a virulent pseudorecombinant between two distinct species of
begomoviruses that infect tomato, and was the second report on synergism between
begomoviruses (Chakraborty et al., 2008). In 2010, diversity of tomato infecting
betasatellites was analyzed and nucleotide identity of various sequences taken in the study
varied from 45 to 93%. In phylogenetic analysis, these sequences clustered according to
their geographic locations rather than the host species. Two new betasatellite sequences,
tomato leaf curl Bangalore betasatellite and tomato leaf curl Maharashtra betasatellite, were
also identified (Sivalingam et al., 2010).
Three distinct tomato infecting begomoviruses have been characterized in year 2010
itself. One of these distinct species was characterized from the gangetic plain of Northern
India, associated with a betasatellite. The viral sequence shared maximum (85.8%) identity
with Tomato leaf curl Laos virus-[Laos] reported from Laos; whereas sequence of the
betasatellite shared maximum (75.8%) identity with tomato leaf curl Joydebpur betasatellite
reported from Bangladesh. This virus was proposed a name tomato leaf curl Patna virus
(ToLCPaV) and DNA-β was named tomato leaf curl Patna betasatellite (ToLCPaB)
(Kumari et al., 2010). The other two distinct monopartite begomoviruses have been
reported from New Delhi (ToLCV-CTM) and Kerala (ToLCVK3/K5). These isolates were

Review of literature
28
considered to be novel begomoviruses based on <89 % nucleotide identity of their complete
genomes. Sequence and phylogenetic analysis study revealed that these isolates have
evolved by recombination between viruses related to two or more viral ancestors (Pandey et
al., 2010).
2.4.1.2 Potato
ToLCNDV has emerged as a serious virus disease in India. Delayed setting in winter in
subtropical countries due to global warming and changed cropping pattern has helped this
virus to establish on potato (Garg, 2005). Potato leaf curl disease (PoLCD) was observed
for the first time in northern India in 1999 (Usharani et al., 2004a). A distinct strain of
tomato leaf curl New Delhi virus (ToLCNDV-Pot) was found to be associated the disease.
The virus was sap-transmissible and bipartite in nature. A defective betasatellite was also
detected, associated with an old world bipartite begomovirus for the first time (Usharani et
al., 2004a).
2.4.1.3 Chilli
A leaf curl disease in the chilli (Capsicum annuum) cultivars Surajmukhi, Anhra Jyoti, X
235, Chandramukhi, Pusa Jawala, Longi-Jpani, Chanchal, Chaman and Selection 54 was
observed in Uttar Pradesh, India. The causal virus could be transmitted from infected to
healthy chilli plants through whiteflies. The virus was detected by dot-blot hybridization,
using the total DNA extracted from infected chilli leaf samples and an Indian tomato leaf
curl virus-specific probe. This study was the first evidence of association of a begomovirus
with leaf curl disease of chilli in India (Raj et al., 2005).
C. annuum plants were observed to exhibit symptoms consisting of leaf curling,
shortening of internodes and petioles, crowding of leaves and stunting of whole plants in
fields around Lucknow, Uttar Pradesh, India. The disease was experimentally transmitted
from infected to healthy chilli and tomato seedlings by whiteflies. Inoculated chilli plants
developed typical symptoms of the disease. Tomato plants developed severe leaf curl
symptoms similar to those of leaf curl disease of tomato caused by ToLCNDV. PCR
amplification and sequence analyses confirmed the identity of the virus as ToLCNDV. This
was the first report of this begomovirus affecting chilli crops in India (Khan et al., 2006).

Review of literature
29
In December 2004, up to 100% disease incidence was observed in C. annuum plants
in Jodhpur district, Rajasthan, India. Infected plants exhibited symptoms consisting of
upward curling, puckering and reduced size of leaves. Severely affected plants were stunted
and produced no fruit. The virus was transmitted to healthy chilli plants by whiteflies. PCR
amplification and sequence analysis (DQ445255) showed close similarity of the isolate
with the intergenic region and part of the replication initiator protein, AV1 and AV2 genes
of Chilli leaf curl virus-[Pakistan: Multan] (ChiLCuV-[Pk:Mul]; AF336806), reported from
Pakistan. This was the first report of ChiLCuV affecting chilli in India (Senanayake et al.,
2007).
Symptoms of mild yellowing, severe leaf curling, and leaf distortion, stunting and
blistering were observed in June 2004 in chilli fields in Ludhiana, Punjab, India. The
pathogen was identified as Tomato leaf curl Joydebpur virus (ToLCJV), based on the
sequence data. This was the first report of ToLCJV infecting chilli naturally in India (Shih
et al., 2007).
Another incidence of chilli leaf curl disease was observed in 2008 in Varanasi,
India. Sequence analysis revealed that the monopartite genome (EF190217) of causal virus
had the highest sequence identity (95%) with that of ChiLCuV-PK[PK:Mul:98]. The
betasatellite (EF190215) shared maximum sequence identity (88%) with a betasatellite
associated with tomato leaf curl disease from Rajasthan (ToLCBDB-[IN:Raj:03]).
Infectious clones of the viral genome and the betasatellite produced typical leaf curl
symptoms in chilli and N. benthamiana (Chattopadhyay et al., 2008).
2.4.1.4 Weeds
Tomato leaf curl virus was found to naturally infect Parthenium hysterophorus and
Sonchus asper plants in Uttar Pradesh, India, during 2002-03. B. tabaci could transmit the
virus from these symptomless infected weeds to tomato plants, which exhibited leaf curling
and twisting symptoms at 15-25 days after transmission (Ansari et al., 2005).
In 2007, disease incidence of yellow vein mosaic (YVM) disease of kenaf (Hibiscus
cannabinus) was observed in northern India. The disease was found in both commercial
and experimental farms in the Bahraich district, Uttar Pradesh, India and caused significant
reduction in plant height and crop yield. Southern hybridization with begomovirus-specific

Review of literature
30
probe and PCR amplification with betasatellite and coat protein primers confirmed the
association of begomovirus with the disease (Ghosh et al., 2007). Later in 2007, full-length
DNA-A of a begomovirus infecting mesta (H. cannabinus L.) was cloned and sequenced.
The complete nucleotide sequence of the DNA-A molecule shared highest sequence
identity (83.5%) with an Indian begomovirus causing cotton leaf curl disease. Thus it was
considered as a novel begomovirus species, and the name mesta yellow vein mosaic virus
was given (Chatterjee and Ghosh, 2007a). A betasatellite component distinct from other
known geminiviruses or nanovirus components was also identified associated with mesta
yellow vein mosaic disease (Chatterjee and Ghosh, 2007b).
In 2008, six betasatellite isolates associated with YVM disease in mesta crops from
three different geographical locations of India were characterized. These six isolates and
the one previously reported from eastern India could be divided into two distinct types
based on the sequence identities. Another incidence of YVM disease of mesta in northern
India was recorded and the causal begomovirus was identified as a distinct monopartite
species associated with a betasatellite. This begomovirus species shared low sequence
identity with the mesta yellow vein mosaic virus (Das et al., 2008a, 2008b). A begomovirus
with DNA-A and a betasatellite molecule was detected for the first time in the eastern part
of India in Urena lobata showing yellow vein mosaic disease symptoms (Chatterjee et al.,
2007b). In 2008, nine isolates of betasatellite infecting kenaf were characterized from three
geographical locations in India. Betasatellites from northern and eastern region of India
showed 84.3% nucleotide identity and clustered with various isolates of Cotton leaf curl
Multan betasatellite (CLCuMB) reported from Indian subcontinent. Betasatellite sequences
from the Southern India were closely related to Tomato leaf curl Joydebpur betasatellite
and showed only 45.2 and 44.9% identity with betasatellite sequences from Eastern and
Northern India, respectively (Paul et al., 2008).
In 2010, yellow vein disease of weed Vernonia cinerea was observed around crop
fields in Madurai. Associated pathogen was identified as a new species of begomovirus
which showed highest identity (82.4%) with Emilia yellow vein virus reported from China.
A betasatellite was found to be associated with this disease and it shared highest identity
(56.8%) with DNA-β of Sigesbeckia yellow vein Guangxi betasatellite. This happened to be

Review of literature
31
the first report of any begomovirus infecting V. cinerea in India (Packialakshmi et al.,
2010).
2.4.2 Sub-temperate areas
Sambyal et al. (2006) collected tomato samples showing leaf curl symptoms from different
areas of Solan district of Himachal Pradesh at different altitudes ranging from 600-1650
mts. The samples showed positive reaction with antisera of tomato leaf curl virus. PCR also
confirmed the presence of tomato leaf curl virus in the samples. Begomovirus infection
could be detected from samples collected from locations with 1650 m altitude. This was the
first molecular evidence of a natural begomovirus infection in a sub-temperate area in India
(Sambyal et al., 2006).
Ageratum is an obnoxious weed, commonly called as “Neela Phulnu” is widespread
in different parts of Himachal Pradesh and two species viz., Ageratum conyzoides Linn. and
A. houstonianum Mill are found widely. A. houstonianum grows best in moist and shady
conditions. It is spread in abundance along the road sides, field bunds, water channels, tea
gardens, orchards, pastures and grasslands. The occurrence of a virus like disease showing
yellowing of veins, crinkling of young leaves, non-flowering and marginal necrosis in
severely infected plants has been reported on A. houstonianum (Sharma et al., 2001). In a
study, the virus did not show seed transmission but it could be transmitted through whitefly
and grafting only to A. houstonianum, A. conyzoides and Abelmoschus esculentus. Immuno-
diagnostic studies (ISEM and ELISA) revealed the association of geminate particles
measuring 17-28 nm in diameter with the disease. The infected samples reacted positively
with the antiserum of ICMV, suggesting association of begomovirus. Based on the particle
morphology, serological relationship, ultrastructural changes and transmission studies it
was evident that a begomovirus is associated with yellow mosaic disease of A.
houstonianum in Himachal Pradesh. The virus seemed to be similar to okra yellow vein
mosaic virus and thus, these weed species (A. houstonianum and A. conyzoides) could be an
alternative host of okra yellow vein mosaic virus in the region (Sharma et al., 2007).

Review of literature
32
2.5 RNA SILENCING AND VIRAL SUPPRESSION
As the name suggests, ‘RNA silencing’ refers to a novel mechanism of gene regulation in
eukaryotic cells and takes place at RNA level. RNA silencing was first discovered in
transgenic petunia plants, when an attempt to over-express chimeric chalcone synthase gene
for enhanced purple colour of the flowers, resulted in reversible suppression of the
introduced as well as homologous genes (Napoli et al., 1990). This phenomenon was called
co-suppression, while it is known as quelling in fungi and RNA interference (RNAi) in
animals (Cogoni and Macino, 1997; Fire et al., 1998). There are at least three pathways of
RNA silencing, involved in: 1) cellular defense against viruses and transgenes via short
interfering RNAs (siRNAs), also known as post transcriptional gene silencing (PTGS) in
plants, 2) regulation of gene expression via microRNAs (miRNAs), and 3) protection of
genome from transposons by DNA methylation and suppression of transcription via
siRNAs, also known as transcriptional gene silencing (TGS) (Baulcombe, 2004; Voinnet,
2005).
2.5.1 Pathways of RNA silencing
PTGS pathway is triggered by production of double stranded RNAs (dsRNAs) as a
replication intermediate or a secondary-structure feature of single-stranded viral RNA. In
plant DNA viruses the dsRNA may be formed by annealing of overlapping complementary
transcripts (Baulcombe, 2004). This dsRNA is then cleaved into characteristic 20-25 nt
long siRNAs with 2 nt overhangs at 3’ ends by RNaseIII-like proteins known as
Dicer/Dicer-like (DCL) proteins. Four different DCL proteins have been described in
Arabidopsis which probably cleave dsRNAs from different origins (Xie et al., 2005;
Deleris et al., 2006). DCL1 is involved in the biogenesis of miRNAs (Bartel, 2004),
whereas DCL2, DCL3, and DCL4 produce siRNAs typically of 22, 24, and 21 nt in length,
respectively (Dunoyer et al., 2005; Xie et al., 2005).
The 3’-terminal nucleotide of miRNAs and siRNAs are methylated by the
methyltransferase HEN1 for their protection from degradation and polyuridylation (Boutet
et al., 2003; Chen, 2005). Host RNA dependent RNA polymerase (RdRP) and several other
cellular proteins are involved in siRNA amplification process which helps in systemic
spread of RNA silencing, even in the absence of the original dsRNA trigger (Baulcombe,

Review of literature
33
2004). The siRNAs are denatured and recruited into a multi-protein RNA induced silencing
complex (RISC), via a specific Dicer associated dsRNA-binding (DRB) protein in an ATP-
dependent step. This complex carrying single-stranded siRNA is called activated RISC, of
which Argonaute protein (AGO) is a core component. RISC is then guided by the siRNA to
contact complementary mRNAs, causing their degradation (Mlotshwa et al., 2002; Roth et
al., 2004). An RNA slicer AGO1 protein binds to the target mRNA and cleaves it at 10-11th
nucleotide position of the siRNA (Elbashir et al., 2001; Bartel, 2004). Arabidopsis genome
encodes at least four DCL, ten AGO, five DRB, and six RdRP proteins that control the
RNA silencing pathways (Csorba et al., 2007; Hutvagner and Simard, 2008; Díaz-Pendón
and Ding, 2008).
The miRNA and trans-acting siRNA (ta-siRNA) pathways play crucial roles in gene
regulation and development of plants (Bartel, 2004; Voinnet, 2005; Meins et al., 2005).
These pathways are related to PTGS, in which the respective siRNAs, miRNAs and ta-
siRNAs are derived from separate genetic loci and act in trans to silence their target
mRNAs (Bisaro, 2006). After denaturation, one of the siRNA strands is recruited into the
RISC or an RNA-induced initiation of TGS complex to guide these complexes to their
respective targets (Baulcombe, 2004). Distinct AGO proteins act as active components in
each pathway (Zilberman et al., 2003; Baumberger and Baulcombe, 2005). TGS is usually
triggered by transcription of inverted repeats or tandemly repeated sequences (Jones et al.,
2001; Mette et al., 2000). In TGS, slightly larger siRNAs (24-26 nt) are involved, which
are generated by DCL3 in conjunction with AGO4 and RdDRP2 (Hamilton et al., 2002;
Xie et al., 2004; Qi et al., 2005). In addition to silencing elements, some other components
like methyltransferases are required in TGS (Bender, 2004; Gehring and Henikoff, 2007).
2.5.2 RNA silencing signal
Most of the plant viruses systemically infect their hosts (Hull, 2002). Probably in a co-
evolution process, the host plant triggers RNA silencing against the viral nucleic acid and
generates a mobile silencing signal that can spread along with/ahead of the virus (Roth et
al., 2004; Kalantidis et al., 2008). The systemic silencing patterns suggest that the signal
moves from cell-to-cell (short-distance) and through the phloem (long-distance),
resembling viral movement through the plant (Mlotshwa et al. 2002; Voinnet, 2005). It has

Review of literature
34
been shown that viral suppressors of RNA silencing are typically required for long-distance
spread of the virus in the infected plant, suggesting that RNA silencing signal is a crucial
component of the antiviral silencing pathway (Voinnet et al., 2000; Li and Ding, 2001;
Kasschau and Carrington, 2001; Ding et al., 2004). Highly specific nature of silencing
signals suggests the association of a nucleic acid component. Short-distance cell to cell
spread of the silencing signal takes place through plasmodesmata, mediated by secondary
21 nt siRNAs which are dependent upon RDR6 and are generated by DCL4 in a process
that requires RdRP2 and Pol-IVa (Himber et al., 2003; Dunoyer et al., 2005, 2007; Smith et
al., 2007). In contrast, long-distance movement is mediated by primary siRNAs (24 nt)
derived from viral dsRNA, requiring RdRP6, SDE3 (RNA helicase) and probably SDE5 to
produce dsRNA by using short-distance signals (Voinnet, 2005; Schwach et al., 2005;
Brosnan et al., 2007).
2.5.3 Viral strategies to overcome host antiviral defense
Viruses have evolved various counter-defensive strategies to overcome host antiviral
defense system. Some plant viruses evade RNA silencing by localizing and replicating
themselves in sub-cellular sites that are not exposed to RNA silencing machinery. Brome
mosaic virus, a Bromovirus is an example that replicate in membrane bound vesicles and
keeps its RNA away from host ribonucleases (Schwartz et al., 2002). Viroids escape RNA
silencing by forming strong secondary structures. In case of Hop stunt viroid, it is
experimentally demonstrated that although it is able to induce RNA silencing, mature forms
of the viroid RNA are resistant to RNA silencing (Gómez and Pallás, 2007). It is believed
that some viruses may multiply and spread very fast so as to avoid RNA silencing against
them.
Most of the viruses encode RNA silencing suppressors (RSSs) as an adaptive
response against RNA silencing. The discovery of RSSs strongly suggested that RNA
silencing is an antiviral mechanism and uncovered the viral counter-defensive strategy by
which they suppress the host defense. More than 35 RSSs have been identified from plant
and animal viruses, and evidences indicate that at least one RSS is encoded by each plant
virus (Li and Ding, 2006; Ding and Voinnet, 2007). The majority of RSSs do not share any
obvious structural or sequence identities across families and groups except that most of

Review of literature
35
them have been identified as pathogenicity determinants (Li and Ding, 2006). A number of
RSSs are reported to be viral pathogenicity determinants and produce phenotypic
abnormalities resembling with those of ago1, hyl1, hen1 and dcl1 mutants (Dunoyer et al.,
2005; Voinnet, 2005; Zhang et al., 2006).
RSSs adopt different strategies to overcome RNA silencing. Cucumber mosaic virus
(CMV) 2b and Beet western yellows virus (BWYV) P0 proteins have been reported to
physically interact with AGO1 (Zhang et al., 2006; Bortolamiol et al., 2007). Tombusviral
P19 protein suppresses RNA silencing by sequestering duplex siRNAs, which may prevent
siRNA incorporation into the RISC (Lakatos et al., 2004). HcPro of Tobacco etch virus
(TEV) and p21 protein of Beet yellows virus can bind and inactivate siRNAs (Lakotas et
al., 2004). TrAP of ACMV, Bhendi yellow vein virus (BYMV) and Mungbean yellow
mosaic virus, and HcPro of potyviruses may transactivate host suppressor genes (Voinnet,
2005; Trinks et al., 2005), while AC4 protein of ACMV captures single stranded siRNAs
rendering RISC inactive (Chellapan et al., 2005). TGMV TrAP and curtoviruses L2 protein
inactivate ADK proteins involved in host defense response (Wang et al., 2005a). BYMV
associated βC1 protein is reported to interact with tomato karyopherin α for suppression of
RNA silencing (Gopal et al., 2007). These studies suggest that the RSSs act at different
biochemical steps of RNA silencing; therefore they can serve as important biological tools
to dissect the detailed RNA silencing pathways (Karjee et al., 2008).
2.5.4 Begomoviruses and RNA silencing
Begomoviruses can both induce and be the targets of RNA silencing (Vanitharani et al.,
2005). The siRNAs of begomoviruses can be of 21, 22 and 24 nt long, implying that all the
four DCL protein are involved in their generation. However, DCL3 and DCL4 were found
to be more important for siRNA production in the plants infected with DNA viruses
(Blevins et al., 2006). In response to begomovirus infection, many segments of the viral
DNAs are methylated in a siRNA-dependent manner (Seemanpillai et al., 2003).
Begomoviruses can trigger RNA silencing, though they do not have a dsRNA phase in their
replication cycle. Possibly their bi-directional transcription results in overlapping of the
transcripts at their 3’ ends forming dsRNA (Townsend et al., 1985; Chellappan et al.,
2004). Alternatively, host RdRP can also generate the dsRNA from the replicase transcript

Review of literature
36
of begomoviruses, which is present abundantly in the early stages of infection (Dalmay et
al., 2000). The strong secondary structures of begomovirus transcripts can also be used as a
template by DCL proteins for the production siRNAs (Chellappan et al., 2004). However,
most of the begomoviruses encode suppressor proteins against the host RNA silencing
machinery. Various RNA silencing suppressor proteins (RSSs) encoded by begomoviruses
have been identified till date (Sharma and Ikegami, 2008), which are listed in Table 2.1.
2.5.5 Assays for suppressor identification
Screening of viral genes for suppressor activity is of utmost significance and a number of
assays have been developed to identify plant viral RSSs (Roth et al., 2004; Karjee et al.,
2008; Yadava et al., 2010). However, two approaches have been most commonly used for
this purpose:
In a transient expression or Agrobacterium co-infiltration assay (Fig. 2.5a), a mixture of
two Agrobacterium strains (one carrying a reporter gene for inducing silencing, the other is
viral ORF being tested for suppressor activity) are co-infiltrated into a plant leaf and the
infiltrated patch is examined over time for silencing of the reporter gene (Voinnet et al.,
2000; Llave et al., 2000). Transgenic Nicotiana spp. expressing the reporter gene (usually
GFP) as well as non-transgenic plants can be used (Zrachya et al., 2007). In a typical
experiment with Agrobacterium, green fluorescent protein (GFP) gene is used as the
inducer. Initially, the infiltrated patch expresses high levels of GFP and fluoresce bright
green under ultraviolet (UV) light. However, local RNA silencing gets triggered in three to
five days post infiltration (dpi), and the patch appears dirty red in colour under UV light
due to presence of chlorophyll in the leaves (Fig. 2.5a). If the co-infiltrated viral ORF
possesses a suppressor activity, it interferes with RNA silencing and the patch remains
bright green even after 20 dpi (Roth et al., 2004).

Review of literature
37
Suppressor
protein
Begomoviral species Mechanism
suppressed
Sub-cellular
localization
References
AC2/C2
African cassava mosaic virus-[Kenya] Local NK Vanitharani et al. 2004
Tomato golden mosaic virus Local Nucleus Wang et al. 2003
Mungbean yellow mosaic virus NK Nucleus Trinks et al. 2005
Indian cassava mosaic virus NK NK Vanitharani et al. 2004
East African cassava mosaic Cameroon virus NK NK Vanitharani et al. 2004
Tomato yellow leaf curl China virus NK Nucleus Dong et al. 2003
Tomato leaf curl Java virus Systemic Nucleus Kon et al. 2007
Bhendi yellow vein mosaic virus Local NK Gopal et al. 2007
AC4/C4
African cassava mosaic virus-[Cameroon] Systemic NK Vanitharani et al. 2004
Sri Lankan cassava mosaic virus Systemic NK Vanitharani et al. 2004
East African cassava mosaic Cameroon virus Systemic Membrane Fondong et al. 2007
Bhendi yellow vein mosaic virus Systemic NK Gopal et al. 2007
AV2/V2 Tomato yellow leaf curl virus-[Israel] NK Cytoplasm Zrachya et al. 2007
Tomato leaf curl Java virus NK NK Sharma & Ikegami 2010
βC1
Bhendi yellow vein mosaic virus Systemic Cytoplasm Gopal et al. 2007
Tomato leaf curl Java virus Systemic Cytoplasm Kon et al. 2007
Tomato yellow leaf curl China virus-[Y10] Systemic Nucleus Cui et al. 2005
Alpha Rep Cotton leaf curl Rajasthan virus NK NK Nawaz-Ul-Rehman et al. 2010
Table 2.1 List of begomovirus encoded RSSs identified till date. NK: Not known
37
Review
of litera
ture

Review of literature
38
In another approach called reversal of silencing assay, a plant silenced for a reporter
gene is infiltrated with the candidate virus or its individual ORFs to determine whether the
silenced phenotype is reversed (Fig. 2.5b). Most commonly, GFP-expressing transgenic N.
benthamiana line 16C is used in this assay (Ruiz et al., 1998). Soon after germination of
the plant, it is infiltrated with Agrobacterium expressing GFP to trigger local and systemic
silencing. After that the plant gets silenced for GFP and appears red in UV light. The plant
is then inoculated with the virus/ORF being tested for suppressor activity and if it can
reverse the silencing and allows the plant to express GFP, the infiltrated patch will glow
green under UV (Voinnet et al., 1999). Transgenic Nicotiana tabacum (cv. Xanthi) plants
expressing GFP as well as siRNAs against GFP (GFP silent line) has also been developed
by which individual genes can be assayed for suppressor activity without requiring initial
triggering of RNA silencing (Karjee et al., 2008). Agrobacterium as well as potato virus x
(PVX) vector can be used for expressing various ORFs being tested (Roth et al., 2004).
Fig. 2.5 Cartoon demonstrating a) co-infiltration and b) reversal of silencing assays








![2007 ANNUAL REPORT [E] - MSV Life · 2020. 6. 12. · Group (“MSV”), notwithstanding some ... we seek to exercise in our investment operations. During 2007, MSV continued to experience](https://img.pdfslide.us/doc/110x75/601191cba6c5644c480c8a3a/2007-annual-report-e-msv-life-2020-6-12-group-aoemsva-notwithstanding.jpg)




















