Embed Size (px)
Citation preview

DOCENTE:
Dr. Víctor Chumpitaz Cerrate
ALUMNOS :
o Herrera Ccoicca, Marylin
o Huamaní Chipana, Karina
o Huapaya Pardavé, María
o Núñez Díaz, Daniella
o Parra Gálvez, Katherine
TURNO : 3 – 5pm
Ciudad universitaria, abril del 2011
UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS
FACULTAD DE ODONTOLOGÍA
FISIOLOGIA GENERAL

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
2
ÍNDICE
I. INTRODUCCIÓN -------------------------------------------------------------------------- 3
II. MARCO TEORICO ----------------------------------------------------------------------- 4
1. SISTEMA NERVIOSO AUTONOMO ----------------------------------------------- 4 1.1 El control autónomo de los músculos lisos ---------------------------------- 5 1.2 Los componentes centrales del sistema nervioso autónomo ----------- 5 1.3 Función Simpática: ---------------------------------------------------------------- 5 1.4 Función Parasimpática: ---------------------------------------------------------- 6
2. SISTEMA NERVIOSO SIMPÁTICO ------------------------------------------------ 6 2.1 Distribución segmentaria de las fibras nerviosas simpáticas ----------- 7 2.2 Síntesis de noradrenalina, su eliminación y duración de su acción --- 7 2.3 Efectos autónomos sobre los diversos órganos del cuerpo ------------- 9
3. RECEPTORES ADRENÉRGICOS ------------------------------------------------ 10 3.1 Bases moleculares de la función de los receptores adrenérgicos ---- 10 3.2 Estructura de los receptores adrenérgicos --------------------------------- 10 3.3 Clasificación ----------------------------------------------------------------------- 10 3.4 Receptores adrenérgicos alfa ------------------------------------------------- 10
3.4.1 RECEPTORES α1: ------------------------------------------------------------ 10 3.5 Receptores adrenérgicos beta ------------------------------------------------ 11
3.5.1 RECEPTORES ß1: ------------------------------------------------------------ 11 3.5.2 RECEPTORES ß2: ------------------------------------------------------------ 11 3.5.3 RECEPTORES ß3: ------------------------------------------------------------ 12
4. RECEPTORES DOPAMINÉRGICOS --------------------------------------------- 12 4.1 Receptores dopaminérgicos d1: ---------------------------------------------- 12 4.2 Receptores dopaminérgicos d2: ---------------------------------------------- 12
5. SISTEMA NERVIOSO PARASIMPÁTICO: -------------------------------------- 13 5.1 Distribución de las fibras del Sistema Nervioso Parasimpatico: ------ 14 5.2 Neurotransmisores del Sistema Nervioso Parasimpático: -------------- 16 5.3 Características de la transmisión colinérgica en diferentes sitios: --- 17
6. RECEPTORES DEL SISTEMA NERVIOSO PARASIMPÁTICO: ---------- 18 6.1 Receptores muscarínicos: ------------------------------------------------------ 18
6.1.1 TIPOS DE RECEPTORES MUSCARÍNICOS:-------------------------- 19 6.2 Receptores nicotínicos ---------------------------------------------------------- 20
6.2.1 TIPOS DE RECEPTORES NICOTÍNICOS ------------------------------ 20
III. CONCLUSIONES --------------------------------------------------------------------- 22
IV. COMENTARIOS DE LOS ARTICULOS ----------------------------------------- 23
V. BIBLIOGRAFIA --------------------------------------------------------------------------- 31

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
3
I. INTRODUCCIÓN
El sistema nervioso autónomo (SNA) o vegetativo es un componente importante del
sistema nervioso constituido por un complejo conjunto de neuronas y vías nerviosas
que controlan la función de los diferentes sistemas viscerales del organismo. Su
función global consiste en mantener la situación de homeostasis del organismo y
efectuar las respuestas de adaptación ante cambios del medioambiente externo e
interno.
El SNA a través de los tres componentes eferentes que lo integran: simpático,
parasimpático y entérico, inerva el músculo cardíaco, el músculo liso de todos los
órganos y las glándulas exocrinas y endocrinas. Así, regula la respiración, la
circulación, la digestión, el metabolismo, la secreción glandular, la temperatura
corporal, la reproducción y, además, coordina todas estas funciones vitales para
mantener la homeostasis.
Este sistema, como su propio nombre indica (autónomo o vegetativo) no se encuentra
sujeto al control voluntario o consciente. Las disfunciones del SNA comportan
manifestaciones clínicas múltiples y variables, que, a menudo, son infravaloradas en la
práctica clínica. Sin embargo, los síntomas o alteraciones autonómicos deben tenerse
siempre en cuenta por su implicación diagnóstica, por la repercusión sobre la calidad
de vida del paciente y por la influencia pronóstica sobre la expectativa de vida.
La estructura y funcionamiento básicos del SNA fueron definidos en los inicios de este
siglo primordialmente por Gaskell y Langley, quienes reconocieron sus dos divisiones
principales: el sistema simpático y el parasimpático. Además, Langley designó como
una tercera división el conjunto de los plexos submucoso de Meissner y mientérico de
Auerbach, inmersos en la pared del tracto gastrointestinal, que recientemente ha
ganado aceptación como el sistema nervioso entérico, aunque controlado por las
divisiones simpática y parasimpática. Superando el concepto clásico de un sistema
puramente eferente, en la actualidad se acepta que el SNA está integrado por vías
aferentes viscerales, centros de integración, especialmente en el hipotálamo y el
córtex cerebral, y vías eferentes viscerales simpáticas y parasimpáticas, de forma que
el SNA se extiende tanto en el sistema nervioso central como en el periférico.

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
4
II. MARCO TEÓRICO
RECEPTORES ADRENÉRGICOS Y COLINERGICOS
1. SISTEMA NERVIOSO AUTONOMO
El SNA es la parte del sistema nervioso que controla gran diversidad de funciones viscerales del organismo y su función es la de mantener la compleja homeostasia del organismo en respuesta tanto a las alteraciones del medio interno como a los estímulos exteriores; llega virtualmente a todas las partes del organismo, afectando directa o indirectamente a todos los órganos y sistemas. El sistema nervioso autónomo controla la actividad de los componentes viscerales del cuerpo humano. Se denomina “autónomo” porque realiza sus funciones de forma automática, lo que significa que su funcionamiento no puede controlarse fácilmente a voluntad. El SNA forma el soporte visceral para el comportamiento somático ajustando el organismo anticipadamente para responder al estrés y su actividad ocurre de forma independiente de la voluntad. Tiene un control parcial sobre la tensión arterial, la motilidad y secreciones gastrointestinales, el vaciamiento de la vejiga urinaria, la sudoración, la temperatura corporal, la regulación del músculo cardíaco, del músculo liso y muchas otras funciones viscerales del organismo. Una de las características más llamativas es la rapidez y la intensidad con la que puede cambiar las funciones viscerales. Por ejemplo, en 3 a 5 segundos, puede duplicar la frecuencia cardiaca, y en 10 a 15 segundos la tensión arterial.
Desde un punto de vista anatómico, el sistema autónomo tiene dos componentes principales: el sistema nervioso simpático y el parasimpático. Los nervios simpáticos que controlan la actividad visceral proceden de las porciones torácica y lumbar de la médula espinal; los nervios parasimpáticos proceden del tronco encefálico y de la porción sacra de la médula espinal.
Desde un punto de vista fisiológico, no puede hacerse una generalización única que se aplique a la forma en que los sistemas nerviosos simpático y parasimpático controlan diferentes órganos corporales.
En la mayor parte de los casos, los órganos viscerales están inervados por ambos sistemas, y cada uno de ellos tiene un efecto opuesto en un sistema de controles y equilibrios. El corazón, por ejemplo, está inervado por nervios simpáticos cuya excitación produce una aceleración del ritmo cardíaco, y también por nervios parasimpáticos cuya excitación produce un retardo del mismo. Cada sistema puede estimular o inhibir los órganos que inerva. En otros casos, los órganos son controlados de forma predominante o exclusiva por uno de los dos sistemas. Una función vital del sistema nervioso autónomo es el mantenimiento de la
homeostasia (estado estable de equilibrio) y la adaptación del organismo animal a su medio ambiente externo.
La homeostasia es el estado de equilibrio de las funciones corporales conseguido por un proceso activo; el control de la temperatura corporal, del agua y de los electrólitos son ejemplos de procesos homeostáticos.
Desde el punto de vista farmacológico, no hay un neurotransmisor único asociado a funciones simpáticas o parasimpáticas, como se creyó en tiempos.
La antigua creencia de que la acetilcolina era el transmisor predominante del sistema autónomo tuvo que abandonarse cuando se encontraron nuevas clases de

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
5
neurotransmisores y neuromoduladores (p. ej., dopamina, serotonina, purinas y diversos neuropéptidos). Recientemente, los científicos han resucitado el enfoque conductual del sistema nervioso autónomo. El sistema nervioso autónomo participa en la reacción instintiva de lucha o huida, todavía presente en el ser humano, que constituye en esencia la base de las reacciones fisiológicas causadas por el estrés. Las interacciones entre el sistema nervioso y las funciones inmunológicas son posibles a través del sistema nervioso autónomo. Las emociones que tienen su origen en el sistema nervioso autónomo pueden expresarse a través de los músculos esqueléticos.1
1.1 El control autónomo de los músculos lisos
Los músculos de las vísceras, exceptuando los del corazón, son músculos lisos. El músculo cardíaco tiene características de músculo esquelético y liso al mismo tiempo. Al igual que los músculos esqueléticos, los músculos lisos contienen también las dos proteínas actina y, en menor proporción, miosina. A diferencia de los músculos esqueléticos, no presentan la organización regular en sarcolemas, la unidad contráctil de la fibra muscular. El corazón tiene la singularidad de que puede generar actividad miógena, es decir, incluso después de haberse seccionado sus inervaciones neurales, puede contraerse y relajarse durante varias horas por sí mismo. El acoplamiento neuromuscular en los músculos lisos es diferente al de los músculos esqueléticos. En los músculos esqueléticos, la unión neuromuscular es la conexión entre el nervio y las fibras musculares. En el músculo liso no hay unión neuromuscular; las terminaciones nerviosas entran en el músculo, y se extienden en todas direcciones. Por tanto, los acontecimientos eléctricos en el interior del músculo liso son mucho más lentos que en los músculos esqueléticos. Finalmente, el músculo liso tiene la característica exclusiva de presentar contracciones espontáneas, como las mostradas por el intestino. En gran parte, el sistema nervioso autónomo regula la actividad espontánea de los músculos lisos.
1.2 Los componentes centrales del sistema nervioso autónomo
El principal papel del sistema nervioso autónomo es regular la actividad de los músculos lisos, del corazón, de las glándulas del aparato digestivo, de las glándulas sudoríparas y de las suprarrenales y otras glándulas endocrinas. El sistema nervioso autónomo tiene un componente central, el hipotálamo, situado en la base del cerebro, donde se integran muchas funciones autónomas. Y lo que es más importante, los componentes centrales del sistema nervioso autónomo participan directamente en la regulación de los impulsos biológicos (regulación de la temperatura, del hambre, de la sed, del impulso sexual, de la micción, de la defecación y otros), la motivación, la emoción y, en gran medida, de funciones “psicológicas” como el estado de ánimo, la afectividad y los sentimientos.
1.3 Función Simpática:
Los efectos más importantes del SNS están relacionados con la circulación y la respiración. La estimulación adrenérgica produce un aumento del gasto cardíaco, así como una broncodilatación. Se inhiben las secreciones gastrointestinales y se estimula el metabolismo en general. El SNS juega un papel fundamental en la preservación del organismo, ya que ocasiona de forma rápida y muy efectiva una respuesta a estímulos exteriores que puedan amenazar la integridad del individuo.

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
6
1.4 Función Parasimpática:
La activación del sistema parasimpático está orientada, al contrario de la del simpático, a la conservación de la energía. La estimulación del SNP produce una disminución de la frecuencia cardíaca y de la velocidad de conducción auriculo-ventricular. Origina constricción del músculo liso con afectación bronquial, miosis, etc. Los signos de descarga parasimpática son: náusea, vómito, movimientos intestinales, enuresis, defecación. También origina un aumento de las secreciones.2
2. SISTEMA NERVIOSO SIMPÁTICO
El sistema nervioso simpático, ortosimpático o torácolumbar, llamado así porque se extiende desde el primer segmento torácico (T1), hasta el segundo o tercer segmento lumbar (L2 o L3). El sistema nervioso simpático se caracteriza por presentar una fibra nerviosa preganglionar corta y una fibra nerviosa postganglionar larga, por lo tanto el ganglio donde se realiza la última sinapsis está lejos del órgano efector Las neuronas que originan las fibras preganglionares se ubican en las columnas intermedio- laterales de la médula espinal. Los axones de las neuronas preganglionares simpáticas abandonan la médula espinal a través de la raíz anterior de un nervio raquídeo junto con las fibras motoras somáticas del mismo nivel segmentario. A continuación, los axones preganglionares mielínicos entran en una vía corta denominada ramo comunicante blanco y terminan en los ganglios paravertebrales. Los ganglios paravertebrales están conectados entre sí por fibras nerviosas formando los ganglios de la cadena simpática, que se encuentran a ambos lados de la médula espinal y se extienden desde la base del cráneo hasta el cóccix. Cada cadena dispone en general de un ganglio por cada uno de los segmentos medulares. En la región medular los ganglios cervicales están fusionados en tres: superior, medio e inferior. Los ganglios prevertebrales están situados anteriores a la columna vertebral y cerca de las grandes arterias abdominales. Así, tenemos a los ganglios celiacos, situados a cada lado del tronco celiaco, justo por debajo del diafragma; los mesentéricos superiores y los mesentéricos inferiores. Como los ganglios se encuentran cerca de la médula espinal, los axones preganglionares son cortos. Cuando las fibras preganglionares llegan a los ganglios de la cadena paravertebral, la mayoría hacen sinapsis en ellos, ya sea al mismo nivel o dirigiéndose en sentido caudal o cefálico, siguiendo la cadena simpática para establecer sinapsis con un ganglio de otro segmento. Otras fibras preganglionares pasan a través de la cadena y por medio del nervio esplácnico se dirigen a un ganglio prevertebral donde establecen sinapsis. Hay fibras simpáticas preganglionares que inervan la médula suprarrenal, donde establecen sinapsis sobre las células cromafines (células ganglionares modificadas), las cuales sintetizan y secretan a la sangre adrenalina y noradrenalina. Los axones simpáticos postganglionares son fibras amielínicas que se originan en las neuronas, cuyos somas están situados en los ganglios simpáticos Un rasgo del SNS es la denominada divergencia que se caracteriza porque el número de fibras postganglionares que salen de los ganglios paravertebrales es muy superior al de las fibras preganglionares que salen de la médula espinal. Además hay interconexiones sinápticas entre los ganglios de la cadena simpática. La divergencia permite al SNS activar muchos efectores en el mismo momento y provocar una respuesta muy amplificada.

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
7
2.1 Distribución segmentaria de las fibras nerviosas simpáticas
Las vías simpáticas que nacen en los diversos segmentos de la médula espinal no tienen por qué distribuirse siguiendo la misma porción corporal que las fibras somáticas del nervio raquídeo correspondiente al mismo segmento. En su lugar, las fibras simpáticas del segmento medular T1 en general ascienden por la cadena simpática para acabar en la cabeza; las pertenecientes a T2 terminan en el cuello; las de T3, T4, T5 y T6 lo hacen en el tórax; las de T7, T8, T9, T10 y T11 en el abdomen, y las de T12, L1 y L2 en las piernas. Está distribución es aproximada y los solapamientos resultan abundantes. La distribución de los nervios simpáticos por cada órgano queda en parte determinada según el punto del embrión en el que se haya originado. Por ejemplo, el corazón recibe muchas fibras nerviosas simpáticas desde la porción cervical de la cadena simpática debido a que esta estructura surgió en el cuello del embrión antes de emigrar hacia el tórax. Análogamente, los órganos abdominales reciben la mayor parte de su inervación simpática desde los segmentos inferiores de la médula torácica, porque la mayor parte del intestino primitivo se origina en esta región.
Las fibras nerviosas simpáticas y parasimpáticas segregan básicamente una de las dos sustancias transmisoras de la sinapsis, acetilcolina o noradrenalina. Aquellas fibras que liberan acetilcolina se llaman colinérgicas, las que emiten noradrenalina se llaman adrenérgicas. Todas las neuronas preganglionares son colinérgicas tanto en el sistema nervioso simpático como en el parasimpático. La acetilcolina o las sustancias semejantes, al aplicarlas a los ganglios, excitarán las neuronas pre ganglionares tanto simpáticas como parasimpáticas, Todas o casi todas las neuronas postganglionares del sistema parasimpático también son colinérgicas. En cambio, la mayoría de las neuronas postganglionares simpáticas son adrenérgicas. Sin embargo, las fibras nerviosas simpáticas postganglionares dirigidas a las glándulas sudoríparas, los músculos piloerectores y un número muy escaso de vasos sanguíneos son colinérgicas. La estructura molecular de la acetilcolina y la noradrenalina es la siguiente:
2.2 Síntesis de noradrenalina, su eliminación y duración de su acción
La síntesis de noradrenalina comienza en el axosplasma de la terminación nerviosa de las fibras adrenérgicas, pero se completa en el interior de las vesículas secretoras. La secuencia de reacciones es la siguiente:

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
8
Después de la secreción de noradrenalina a través de la terminación nerviosa, se elimina de su punto de salida siguiendo tres vías:
Recaptación por las propias terminaciones nerviosas adrenérgicas mediante un proceso de transporte activo: se hace cargo de retirar el 50% al 80% de la noradrenalina segregada.
Difusión desde las terminaciones nerviosas hacia los líquidos corporales contiguos y a continuación hasta la sangre: explica la eliminación de la mayor parte de la noradrenalina restante.
Destrucción de pequeñas cantidades por parte de las enzimas tisulares (una de las cuales es la monoaminooxidasa, que está presente en las terminaciones nerviosas y otra es la catecol-O-metiltransferasa, distribuida de forma difusa por todos los tejidos).
Lo habitual es que la noradrenalina segregada directamente a un tejido se mantenga activa tan sólo unos pocos segundos, lo que manifiesta su recaptación y su difusión lejos de esta zona son rápidas. Sin embargo, la noradrenalina y la adrenalina liberadas a la sangre por la médula suprarrenal permanecen activas hasta que difunden hacia algún tejido, donde pueden resultar destruidas por la catecol-O-metiltransferasa; este proceso tiene lugar sobre todo en el hígado. Por tanto, cuando se segregan hacia la sangres, la noradrenalina y la adrenalina permanecen muy activas de 10 a 30 segundos, pero su funcionalidad disminuye hasta la extinción en 1 o varios minutos.
RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
9
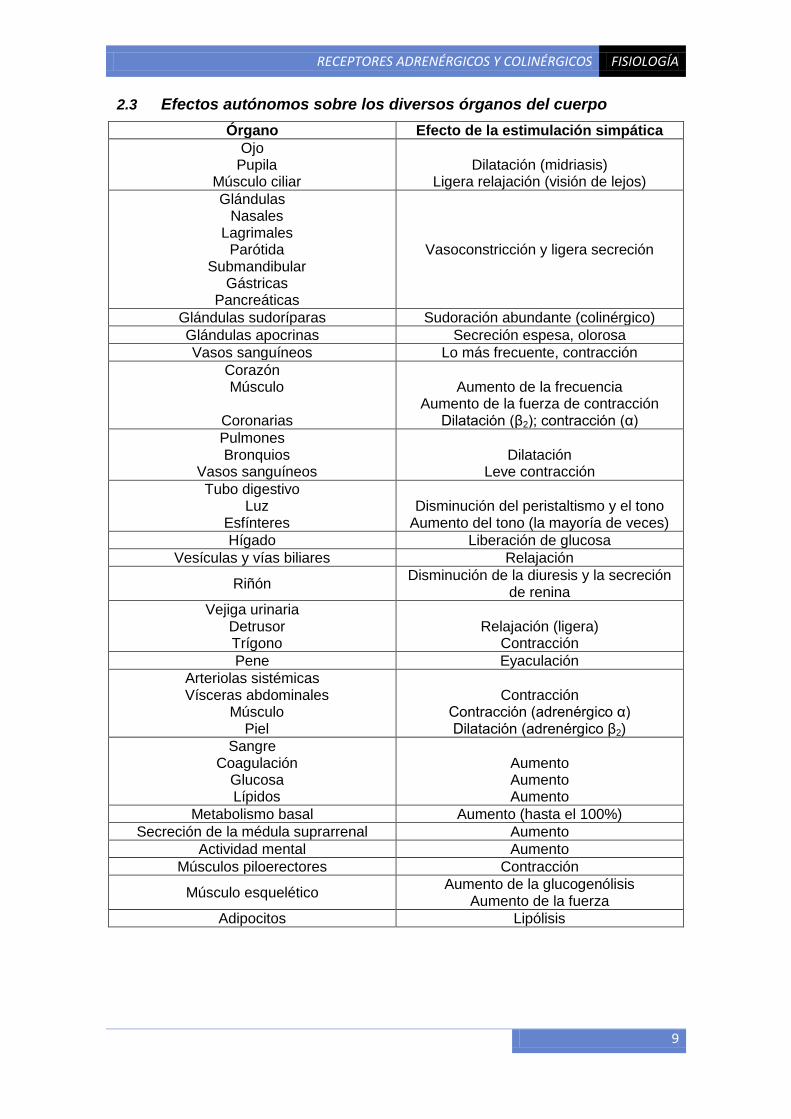
2.3 Efectos autónomos sobre los diversos órganos del cuerpo
Órgano Efecto de la estimulación simpática
Ojo Pupila
Músculo ciliar
Dilatación (midriasis)
Ligera relajación (visión de lejos)
Glándulas Nasales
Lagrimales Parótida
Submandibular Gástricas
Pancreáticas
Vasoconstricción y ligera secreción
Glándulas sudoríparas Sudoración abundante (colinérgico)
Glándulas apocrinas Secreción espesa, olorosa
Vasos sanguíneos Lo más frecuente, contracción
Corazón Músculo
Coronarias
Aumento de la frecuencia
Aumento de la fuerza de contracción Dilatación (β2); contracción (α)
Pulmones Bronquios
Vasos sanguíneos
Dilatación
Leve contracción
Tubo digestivo Luz
Esfínteres
Disminución del peristaltismo y el tono
Aumento del tono (la mayoría de veces)
Hígado Liberación de glucosa
Vesículas y vías biliares Relajación
Riñón Disminución de la diuresis y la secreción
de renina
Vejiga urinaria Detrusor Trígono
Relajación (ligera)
Contracción
Pene Eyaculación
Arteriolas sistémicas Vísceras abdominales
Músculo Piel
Contracción
Contracción (adrenérgico α) Dilatación (adrenérgico β2)
Sangre Coagulación
Glucosa Lípidos
Aumento Aumento Aumento
Metabolismo basal Aumento (hasta el 100%)
Secreción de la médula suprarrenal Aumento
Actividad mental Aumento
Músculos piloerectores Contracción
Músculo esquelético Aumento de la glucogenólisis
Aumento de la fuerza
Adipocitos Lipólisis

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
10
3. RECEPTORES ADRENÉRGICOS
3.1 Bases moleculares de la función de los receptores adrenérgicos
Las reacciones que ocurren después de la activación de todos los tipos de receptores adrenérgicos parecen resultado de los efectos mediados por la proteína G sobre la generación de segundos mensajeros y sobre la actividad de los canales de iones. Estos sistemas abarcan tres proteínas de interacción: El receptor, la proteína G de acoplamiento y las enzimas efectoras o canales de iones.
3.2 Estructura de los receptores adrenérgicos
Los receptores adrenérgicos constituyen una familia de proteínas sumamente relacionadas tanto en estructura como en función, se relacionan también con los receptores de gran variedad de otras hormonas y neurotransmisores que se acoplan a la proteína G. Esta familia amplia de receptores incluye a los colinérgicos muscarínicos.
3.3 Clasificación
Pueden clasificarse en los siguientes grupos: Receptores adrenérgicos alfa: α1 postsináptico, α2 pre y
postsinápticos. Receptores adrenérgicos ß (beta): ß1 cardioselectivos, ß2
broncodilatadores, ß3 lipolíticos y ß2-presinápticos. Receptores dopaminérgicos: En general son postsinápticos, aunque
pueden localizarse presinápticamente en algunas regiones. Los D1 y D2 están localizados en SNC y a nivel periférico.3,4
3.4 Receptores adrenérgicos alfa
3.4.1 RECEPTORES α1: localización postsináptica. Mecanismo de acción: La activación del receptor α1 provoca una redistribución e incremento del calcio citosólico lo cual desencadena los efectos fisiofarmacológicos. La formación del complejo agonista-receptor inicia los siguientes mecanismos:
Se activa la fosfolipasa C (fosfodiesterasa de membrana) a través de la intervención de la proteína G (reguladora de nucleótidos de guanina). La fosfolipasa activada actúa sobre fosfoinositoles de membrana.
La proteína G es un trímero de 3 subunidades: α, ß y gamma, las subunidades beta-gamma en general actúan unidas. La subunidad alfa puede ser estimulatoria (as) o alfa inhibitoria (αi), lo que determina a su vez la existencia de proteína Gs o Gi de acuerdo con la subunidad alfa que posea. La proteína G por medio de la subunidad a se halla unida a una molécula de GDP.
La estimulación del receptor a1 por un agonista provoca la activación de la fosfolipasa C a través de la intervención de la proteína Gs.
Por la acción enzimática de la fosfolipasa C el fosfatidilinositol bifosfato (IP2) origina 1,4, 5, inositol trifosfato (IP3) y diacilglicerol (DAG) que actúan como segundos mensajeros del agonista.
El IP3 moviliza calcio desde depósitos no mitocondriales, principalmente del retículo endoplásmico, incrementando la concentración de calcio. El calcio es fundamental en el desarrollo de importantes funciones celulares dependientes del mismo, y en la activación de calmodulinas, etc.

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
11
El DAG activa la proteinkinasa C, esta enzima promueve fosforilaciones de otras proteínas específicas, enzimas, proteínas ligadas a canales iónicos, etc. que actúan en la secreción celular, contracción de músculos lisos, secreción de hormonas y autacoides, etc.
α1 activados por: NA, metaraminol, fenilfedrina, nafazolina, xilometazolina.
Principales efectos farmacológicos: Contracción de músculos lisos, vasoconstricción (hipertensión), estímulo de algunas secreciones exocrinas (salivales, sudoríparas), disminución de la secreción de insulina y jugo pancreático.3
3.4.2 RECEPTORES α2: Localización pre y postsináptica.
Mecanismo de acción: La activación de receptores α2 presinápticos produce la inhibición de la adenilciclasa y disminución del AMPc intracelular a través de la proteína Gi (inhibitoria) ligada al GTP a través de la subunidad αi de la misma.
α2 activados por: Clonidina, α-metil-noradrenalina, α-metil-DOPA. Principales efectos α2: Inhibición por un mecanismo de
autorregulación de la liberación de NA en la terminal adrenérgica,. Efecto simpaticolítico principalmente central. Los agentes activadores de receptores a2 actúan principalmente en el núcleo tracto solitario bulbo protuberancial, originando un reflejo simpaticolítico inhibitorio del centro vasomotor (hipotensión, bradicardia). También existen receptores a2 postsinápticos, demostrados principalmente en arteriolas, venas y bronquios, con funciones similares a los α1. Estos receptores a2 están acoplados a canales de calcio operados por cambios de voltaje, la activación de estos a2 postsinápticos explicaría los efectos vasoconstrictores iniciales del agente antihipertensivo clonidina.
3.5 Receptores adrenérgicos beta
3.5.1 RECEPTORES ß1: localización postsináptica, están en corazón principalmente.
Mecanismo de acción: La formación del complejo agonista receptor activa la enzima adenilciclasa, estimulando la formación de AMPc. El AMPc activa reacciones catalíticas estimulando proteinkinasas y proteinfosfatasas que agregan o ligan grupos fosfatos o sustratos claves (enzimas y otras macromoléculas) responsables finales de los efectos fisiofarmacológicos.
Receptores ß1 activados por: dobutamina. Principales efectos cardíacos: Producen estímulos de las
propiedades fundamentales del corazón. Aumento de la frecuencia cardíaca, aumento de la contractilidad y velocidad de conducción en aurículas y ventrículos. Aumento del automatismo en el nódulo AV, haz de Hiss y sistema de Purkinge, con posibilidad de extrasístoles por aparición de marcapasos ectópicos.
3.5.2 RECEPTORES ß2: se localizan en músculo liso bronquial, en arteriolas, en arteriolas, en músculo liso de venas, estómago e intestino (motilidad y tono), en útero, en músculo ciliar, en células ß de los islotes de Langerhans, en hepatocitos, en aparato yuxtaglomerular (estimulan la secreción de renina) son postsinápticos.
Mecanismo de acción: idem ß1.

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
12
Activados por: salbutamol, orciprenalina, terbutalina, fenoterol, clembuterol, procaterol (broncodilatadores principalmente); isoxuprina, ritodrina (útero -inhibidores principalmente).
Principales efectos: Broncodilatación, (antiasmáticos), vasodilatación (hipotensión), incremento de liberación de insulina, estímulo a la glucogenólisis y gluconeogénesis, relajación del músculo uterino (útero -inhibidores).
3.5.3 RECEPTORES ß3: Ubicados principalmente en adipocitos estos recientemente descriptos receptores ß3 incrementan la lipólisis por activación de una lipasa específica e incrementan la lipemia en numerosas especies. Son postsinápticos.
Mecanismo de acción: idem a los beta 1. Estimulan adenilciclasa y aumentan AMPc.
Bloqueador ß total: PROPRANOLOL: Este agente no discrimina entre receptores ß1, ß2 o ß3, bloquea a todos por igual.
Receptores ß presinápticos: Intervienen en el mecanismo de autorregulación. Cuando se activan favorecen la entrada de calcio y el proceso de exocitosis. Cambios conformacionales en los canales pueden ser la explicación de la apertura de los mismos.4
4. RECEPTORES DOPAMINÉRGICOS
Son pre y postsinápticos. Se localizan en SNC y a nivel periférico. Se demostró que los receptores centrales (D1 y D2) son virtualmente idénticos a los periféricos (DA1 y DA2), es por ello que tratando de unificar la denominación se utilizará D1 y D2 para describir tanto los centrales como los periféricos.
4.1 Receptores dopaminérgicos d1:
D1 centrales: Son postsinápticos. Se localizan en núcleo caudado, putamen, sustancia negra, tubérculo olfatorio, núcleo amigdalino, corteza cerebral, sistema límbico, hipotálamo, tálamo. La activación de los D1 en los núcleos de la base por agonistas selectivos como el SKF 38393, droga de uso experimental, produce estimulación de la actividad locomotora extra piramidal.
D1 periféricos: Son postsinápticos, están localizados en músculo liso arteriolar de vasculatura renal, mesentérica, coronaria y cerebral. Son principalmente vasodilatadores, incrementan los flujos sanguíneos.
Mecanismo de acción: La interacción agonista receptor D1 activa la enzima adenilciclasa con intervención de la proteína Gs {estimulatoria} e incrementa la formación de AMPc.
Activados por: el agonista D1 selectivo SKF 38393 y también por la dopamina.
4.2 Receptores dopaminérgicos d2:
D2 centrales: Son principalmente postsinápticos, están localizados en los núcleos de la base, cuerpo estriado, sustancia negra, globus pallidus, bulbo olfatorio, en zona quimiorreceptora gatillo del bulbo, en hipotálamo e hipófisis posterior. Serían periféricos, se ha postulado además que los receptores D2 al ser activados podrían incrementar la conductancia al potasio (K+).
D2 periféricos: Se localizan en terminales nerviosas autonómicas e inhiben la liberación de catecolaminas de las terminales simpáticas.
Mecanismo de acción: Los receptores D2 postsinápticos a través de la proteína Gi {inhibitoria} inhiben adenilciclasa y disminuyen los niveles intracelulares de AMPc. También se observó que inhiben fosfoinositoles

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
13
disminuyendo la producción de IP3 y DAG. Además inhiben canales de Ca++ y activan canales de K+.
Principales efectos: Los receptores D2 centrales modulan inhibitoriamente la actividad locomotora extrapiramidal (efecto antiparkinsoniano). También están involucrados en procesos de consolidación de la memoria sobre todo los receptores ubicados en la región ventrolateral y posteroventral del núcleo caudado. Los receptores de hipotálamo e hipófisis posterior inhiben la secreción de prolactina, los de la ZQG producen efectos emetisantes y nauseosos.
Activados por: Quinpirole y apomorfina en forma selectiva y también son activados por el agonista total dopamina.
Los receptores dopaminérgicos D1 y D2 forman en realidad 2 subfamilias de receptores, cada una de ellas con propiedades muy similares. La subfamilia D1 estaría formada por los receptores D1 (o D1a) y D5 (o D1b) y la subfamilia D2 estaría formada por los receptores D2 (o D2a), D3 (o D2b) y D4 (o D2c).
Subfamilia D1: En tal sentido, evidencias experimentales indicarían que los D5 (o D1b) tendrían algunas acciones propias ya que pre dominan en hipocampo e hipotálamo y el neurotransmisor endógeno dopamina, en estas zonas demuestra mucho mayor afinidad (10 veces más) para los receptores D5 con respecto a los D1. Se ha postulado que la activación de los D5 sería importante para el mantenimiento del tono dopaminérgico y el estado de alerta.
Subfamilia D2: También los receptores D2 demuestran heterogeneidad en diferentes regiones del SNC. Los receptores D3 (o D2b) son insensibles a los nucleótidos de guanina y activarían canales de K para ejercer sus efectos. Predominan en núcleo acumbens y sistema límbico (hipocampo, septum). Estarían relacionados con funciones cognoscitivas y emocionales. Sobre estos receptores actuarían con más afinidad el antipsicótico atípico clozapina que justamente por este mecanismo produciría menores efectos extrapiramidales. También se ha demostrado que la activación de receptores D3 en terminales corticoestriatales inhiben la liberación del neurotransmisor excitatorio ácido glutámico en núcleo estriado. Se ha postulado que en la corea de Hungtington hay una disminución o pérdida de estos receptores D3 lo que explicaría parcialmente la fisiopatología de este padecimiento.
También los D4 (o D2c) localizados preferentemente en corteza frontal, cerebro medio, amígdala y en menor concentración en striatum y tubérculo olfatorio desarrollan acciones similares a los D3. En general la distribución de los receptores de la subfamilia D2 es similar aunque existen variaciones importantes en la densidad o cantidad de receptores en algunas áreas del SNC. Así por ejemplo el bloqueo de los D3 y D4 por agentes antipsicóticos como clozapina produce mucho menos efectos extrapiramidales ya que su densidad o concentración es mucho menor que la de los D2 en núcleo de la base.
5. SISTEMA NERVIOSO PARASIMPÁTICO:
El sistema nervioso parasimpático tiene su origen en el mesencéfalo (cerebro medio), médula oblongata y en la porción sacra de la médula espinal, se le denomina sistema nervioso cráneo-sacral. Actúa retornando al organismo condiciones de normalidad, relajo, contrarresta y vuelve a su sitio la actividad del organismo.

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
14
5.1 Distribución de las fibras del Sistema Nervioso Parasimpatico:
Las fibras nerviosas parasimpáticas salen directamente del sistema nervioso central a través de los núcleos de los pares craneales III (oculomotor), VII (facial), IX (glosofaríngeo) y X (vago), estas fibras conforman las denominadas neuronas preganglionares. Otras fibras salen de la parte más inferior de la médula espinal (sacra), por medio de las astas posteriores y anteriores del segundo y tercer nervios raquídeos sacros (S2 y S3) y ocasionalmente por el primero y cuarto (S1 y S4). Desde los núcleos anteriormente mencionados las fibras viajan por los correspondientes nervios craneales y raquídeos sacros para sinaptar en ganglios parasimpáticos periféricos, desde ahí se encuentran las neuronas posganglionares.
Fibras parasimpáticas del III par craneal (oculomotor): El III par craneal tiene dos componentes, uno motor somático que inerva cuatro músculos extraoculares y otro motor visceral, el cual inerva a los músculos oculares intrínsecos (constrictor de la pupila y músculo ciliar). Las fibras motoras viscerales son las eferentes parasimpáticas y salen del mesencéfalo por el núcleo de Edinger-Westphal, estas fibras tienen la función de dar inervación parasimpática de los músculos constrictor de la pupila y ciliar. Los axones motores viscerales se encuentran sobre la cara ventral del nervio y discurren con las fibras motoras somáticas del nervio hasta el músculo oblicuo inferior por una distancia corta, luego lo abandonan para terminar en el ganglio ciliar. Las neuronas posganglionares abandonan el ganglio ciliar como ocho a diez nervios ciliares cortos y entran en el ojo en la cara posterior, cerca de la salida del nervio óptico. Dentro del globo ocular, los nervios discurren hacia adelante, entre la coroides y la esclerótica y terminan en el cuerpo ciliar y el músculo constrictor de la pupila. Ya que estas fibras controlan los músculos constrictor de la pupila y ciliar, controlan el tamaño de la pupila y forma del cristalino.5
Fibras parasimpáticas del VII par craneal (facial): El VII par craneal tiene cuatro componentes, las fibras motoras branquiales, las sensitivas generales, las sensitivas especiales y las motoras viscerales, las tres últimas mencionadas están unidas en una vaina de fascia y se denominan nervio intermedio. Las fibras motoras viscerales son las eferentes parasimpáticas y salen por el núcleo salivar superior (lagrimal), estas fibras tienen la función de estimular las glándulas lagrimales, submandibulares y sublinguales, las glándulas mucosas de la nariz, los senos paranasales y los paladares duro y blando. El núcleo salivar superior está principalmente bajo la influencia del hipotálamo. Los impulsos del sistema límbico (conducta emocional) y el área olfatoria (área sensitiva especial para el olfato) entran al hipotálamo y así median reflejos como la salivación, llanto, etc. Las fibras parasimpáticas pertenecientes al nervio intermedio se dividen en dos grupos en el ganglio geniculado en el conducto facial y se convierten en el nervio petroso mayor (para glándulas lagrimales y nasales) y nervio cuerda del tímpano (para glándulas submandibulares y sublinguales). El nervio petroso mayor se une con el nervio petroso profundo (simpático) y se convierte en el nervio del conducto pterigoideo, luego las fibras parasimpáticas de este nervio hacen sinapsis en el ganglio pterigopatalino parasimpático. Las fibras posganglionares continúan hacia adelante y llegan a la glándula lagrimal y a las glándulas mucosas en la mucosa de las cavidades nasal y oral.

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
15
El nervio cuerda del tímpano se une con la rama lingual del nervio mandibular (V3), viajan juntos hacia el borde lateral del piso de la boca y hacen sinapsis en el ganglio submandibular. Las fibras posganglionares continúan hasta las glándulas submandibular y sublingual y hasta otras en el piso de boca.5
Fibras parasimpáticas del IX par craneal (glosofaríngeo): El IX par craneal tiene cinco componentes, las fibras sensitivas generales, sensitivas viscerales, sensitivas especiales, motoras branquiales y motoras viscerales. Las fibras motoras viscerales son las eferentes parasimpáticas emergen del bulbo raquídeo del tronco encefálico, entre la oliva y el pedúnculo cerebeloso inferior, por los núcleos salivar inferior y ambiguo y tiene como función estimular la glándula parótida y controlar los vasos sanguíneos en el cuerpo carotídeo. Las neuronas preganglionares parasimpáticas de este par craneal son influidas por estímulos del hipotálamo y el sistema olfatorio y se unen con los demás componentes del par craneal en el bulbo raquídeo y luego se vuelve a separar de estas en el ganglio inferior como un componente de la rama timpánica, desde el plexo timpánico, las fibras parasimpáticas forman el nervio petroso menor, el cual hará sinapsis en el ganglio ótico, desde el cual las fibras posganglionares se unen al nervio auriculotemporal (rama de V3) para aportar fibras secretomotoras a la glándula parótida.5
Fibras parasimpáticas del X par craneal (vago): Contiene la mayoría de fibras parasimpáticas. Tienen cuatro componentes que abandonan el bulbo raquídeo: las fibras sensitivas generales, sensitivas viscerales, motoras branquiales y motoras viscerales. Las fibras motoras viscerales son las parasimpáticas y abandonan al bulbo raquídeo mediante el núcleo motor dorsal del nervio vago, el cual inerva ganglios del intestino y sus derivados (pulmones, hígado, páncreas) y el núcleo ambiguo, el cual inerva ganglios del plexo cardiaco. Estas neuronas son influidas por el hipotálamo, el sistema olfatorio, la formación reticular y el núcleo del tracto solitario. Los axones preganglionares se distribuyen en el plexo faríngeo a través de ramas faríngea y laríngea interna. Dentro del tórax hay ramas pulmonares que producen broncoconstricción y ramas esofágicas que aceleran el peristaltismo en el esófago (activan el músculo liso), estos axones hacen sinapsis en los ganglios localizados en las mismas paredes de los órganos individuales. Los nervios gástricos derecho e izquierdo emergen del plexo esofágico y estimulan la secreción de las glándulas gástricas y son motores para el músculo liso del estómago. Las ramas intestinales también son motores en el intestino delgado, el ciego, el apéndice vermiforme, el colon ascendente y la mayor parte del colon transverso. Con respecto a los axones preganglionares cardiacos se sitúan en el núcleo ambiguo medial y terminan sobre ganglios pequeños asociados con el corazón y disminuyen la velocidad del ciclo cardiaco.5
Fibras parasimpáticas de los nervios raquídeos sacros: S2: aparece en el intervalo de los dos fascículos superiores del piramidal y se divide en dos ramas, una superior para S1 y otra inferior para S3. S3: transversal, se adosa a la rama inferior de S2 (nervio bigémino de Jehring). Los dos nervios se distribuyen por el colon descendente, recto, vejiga urinaria y porciones inferiores de los uréteres. Suministra señales nerviosas a los genitales externos para provocar la erección.5
Neuronas parasimpáticas pre y posganglionares:

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
16
Excepto en el caso de algunos pocos nervios parasimpáticos craneales, las fibras preganglionares recorren sin interrupción todo el trayecto hasta el órgano que vayan a controlar, el cual contiene las fibras posganglionares en su pared. Las fibras pre y posganglionares hacen sinapsis, estas fibras posganglionares son cortísimas, su longitud puede ir desde una fracción de milímetro hasta varios centímetros y son estas las que finalmente dan inervación a los tejidos de los órganos. Esta organización es completamente distinta a la del sistema nervioso simpático, ya que la longitud de las fibras posganglionares es mayor y no se hallan en las paredes de los órganos.
5.2 Neurotransmisores del Sistema Nervioso Parasimpático:
Aquellas fibras que segregan acetilcolina son las colinérgicas. Todas las neuronas preganglionares son colinérgicas, tanto para el sistema nervioso parasimpático como para el simpático. La acetilcolina cumple la función de, al aplicarlas en los ganglios, excitar las neuronas posganglionares simpáticas y parasimpáticas. Casi todas las neuronas posganglionares parasimpáticas son colinérgicas, esa es la razón por la que la acetilcolina es conocida como transmisor parasimpático, por el contrario, las neuronas posganglionares simpáticas son adrenérgicas, a excepción de las dirigidas a las glándulas sudoríparas, los músculos piloerectores y a pocos vasos sanguíneos, las cuales son colinérgicas.
Secreción y destrucción de la acetilcolina:
Algunas terminaciones nerviosas autónomas posganglionares parasimpáticas son muy parecidas a las de la unión neuromuscular esquelética. Muchas de estas fibras solo rozan a las células efectoras de los órganos o terminan en tejido conectivo que es adyacente a las células que van a ser activadas. El punto donde estos filamentos pasan sobre las células estimuladas suelen presentar dilataciones bulbosas o varicosidades, en las cuales se sintetizan y almacenan las vesículas transmisoras de acetilcolina. En las varicosidades también se encuentran mitocondrias que darán la energía (ATP) para activar la síntesis de acetilcolina.
Potencial de acción: 1) El potencial de acción llega a la sinapsis. 2) Se libera acetilcolina a través de los canales protéicos. 3) La acetilcolina se une al receptor, causando la excitación de la
membrana postsináptica. 4) Potencial de acción postsináptica. 5) La acetilcolina se desactiva por la acción de la acetilcolinesterasa. 6) La acetilcolina se resintetiza, entra en el citoplasma y se almacena
parcialmente en las vesículas.
El potencial de acción llega al terminal presináptico y como consecuencia del incremento de voltaje, los canales de Ca++ se abren. Los Ca++ se difunden dentro de la célula y causan que las vesículas sinápticas segreguen acetilcolina. Las moléculas de acetilcolina se difunden desde el terminal presináptico hasta la hendidura sináptica y se unen a sus receptores en los canales ligando-dependientes de Na+. Esto causa que los canales ligando-dependientes de Na+ se abran y que los Na+ se difundan en toda la célula, haciendo más positivo al potencial de membrana. Si es que el potencial de membrana alcanza los niveles del umbral, un potencial de acción se producirá. La reacción química básica de la síntesis de acetilcolina es:

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
17
La Acetil-CoA se deriva del piruvato por vía de la reacción del piruvato deshidrogenasa de etapa múltiple o por efecto de la tioacetatocinasa, la cual cataliza la reacción del acetato con ATP para formar un aciladenilato ligado a enzima (acetil-AMP), la síntesis te Acetil-CoA se realiza en las mitocondrias. La colina se capta desde el líquido extracelular hacia el axoplasma por transporte activo. La etapa final de la síntesis se realiza en el citoplasma. Luego de que la acetilcolina se segrega a un tejido, se queda ahí mientras transmite la señal nerviosa. Posteriormente, la Acetilcolina se descompone de la siguiente manera:
La acetilcolinesterasa está unida al colágeno y a los glucosaminoglucanos en el tejido conjuntivo local. Luego, la colina formada vuelve a la terminación nerviosa, en la cual se volverá a utilizar para la síntesis de una nueva acetilcolina.6
5.3 Características de la transmisión colinérgica en diferentes sitios:
Músculo estriado: La combinación de ACh con los receptores colinérgicos nicotínicos a nivel de la superficie externa de membrana posináptica incrementa la permeabilidad de los cationes. La ACh activa al receptor y abre su canal intrínseco e ingresan 50000 Na2+. El proceso es la base para el potencial despolarizante en la placa motora que desencadena el potencial muscular de acción, el cual produce contracción. Luego de la degeneración del nervio motor del músculo estriado o de las fibras preganglionares que van hacia los efectores autonómicos, se produce una reducción notable en las dosis umbral de los transmisores, este cambio se logra por diseminación de las moléculas del receptor desde la región de la placa terminal hacia las porciones adyacentes de la membrana sarcoplásmica y abarca toda la superficie muscular.
Efectores autonómicos: La estimulación o la inhibición de las células efectoras autonómicas se produce con la activación de los receptores colinérgicos muscarínicos. Aquí, el efector se acopla al receptor mediante la proteína G. El sistema de conducción del músculo liso y del corazón manifiesta normalmente una actividad intrínseca eléctrica y mecánica que se modifica, pero no es iniciada por impulsos nerviosos. En el estado basal, el músculo liso manifiesta espigas de despolarización que se propagan de célula a célula a ritmos mucho más lentos que los del potencial de axones o de músculo estriado. La aplicación de ACh al músculo liso intestinal aislado produce disminución de potencial en reposo, es decir, el potencial de membrana se vuelve menos negativo e incrementa la frecuencia de la producción de espigas, lo cual aumenta la tensión. La ACh puede producir contracción de los músculos lisos cuando se ha despolarizado por completo la membrana por concentraciones altas de K+ y si se encuentra presente Ca2+. En el sistema de conducción cardiaco, la estimulación de la inervación colinérgica produce inhibición aunada a hiperpolarización de membrana y disminución de despolarización.
Ganglios autonómicos: Semejante a la unión neuromuscular del m. estriado. La despolarización inicial es el resultado de activación de los receptores colinérgicos nicotínicos. Varios moduladores secundarios fomentan o

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
18
disminuyen la sensibilidad de la célula posganglionar a la ACh, la cual parece estar relacionada con el potencial de membrana del cuerpo de la célula posináptica o de sus dendritas.7
6. RECEPTORES DEL SISTEMA NERVIOSO PARASIMPÁTICO:
En el Sistema nervioso parasimpático, el neurotransmisor usado es la acetilcolina y sus receptores son llamados colinérgicos. Estos receptores colinérgicos se dividen en dos tipos con respecto a sus propiedades farmacológicas: muscarínicos y nicotínicos.
6.1 Receptores muscarínicos:
La muscarina es una amina cuaternaria y compuesto natural alcaloide que se obtienen del hongo Amanita muscaria. Este agente tiene escaso efecto en los receptores de los ganglios autónomos pero, posee la propiedad de producir acciones parasimpaticomiméticas, es decir, mimetiza la acción estimuladora de la acetilcolina en el musculo liso y las glándulas; por lo tanto estas acciones en la acetilcolina son llamadas acciones muscarínicas y los receptores correspondientes son los receptores muscarínicos, que tienen como principal antagonista a la atropina.8
FIG”3”. Estructura del receptor muscarínico
Los receptores muscarínicos pertenecen a la familia de receptores metabotrópicos acoplados a la Proteína G. Se caracterizan por estimularse lentamente y porque al ser activados pueden provocar respuestas de tipo excitatorio o inhibitorio. Estos receptores son proteínas glucosiladas con tamaños moleculares que oscilan entre 51 y 66 kD, codificadas en secuencias de entre 460(M1) y 590(M3) aminoácidos. Su extremo aminoterminal se encuentra en el exterior y el carboxiloterminal en el interior citoplasmático, y tiene un gran tercer bucle intercitoplasmático que es el responsable del reconocimiento y activación de las proteínas G, los determinantes para la especificidad del agonista estarán distribuidos en los diferentes segmentos transmembranales presentando una perfecta conservación en todos los subtipos y conformando tridimensionalmente una rueda (Fig.3), donde estos residuos conservados se dispondrán hacia el interior, mientras que los no conservados se situarán hacia el exterior en contacto con la bicapa lipídica. De este modo, los 30-47 residuos con potencialidad de formar enlaces por puentes de hidrogeno se situarán uno frente a otro en torno a un gran hueco central. Análogamente a lo que sucede con el receptor adrenérgico, los

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
19
residuos implicados en la fijación del ligando podrían ser Tyr529 y la Tyr533 del VII segmento transmembranal, y la Tyr148 y acido Asp147 del III.9
6.1.1 TIPOS DE RECEPTORES MUSCARÍNICOS:
La existencia de la diversidad de los receptores muscarínicos se manifestó con las acciones cardioselectivas de la gallamina; y, luego, en los años 70 con la introducción del antagonista pirezepina, se indica la selectividad de algunos receptores en territorios específicos y se inició un proceso de diferenciación de subtipos de receptores que culminó con la clonación de 5 subtipos moleculares.10
Las diferencias en la estructura primaria de cada subtipo generan diferencias funcionales que se reflejan en la afinidad ante diferentes ligandos. De la misma forma, el subtipo determina la Proteína G con la que el receptor interacciona. Y, la respuesta muscarínica generada en una célula particular depende, tanto del fenotipo celular como del subtipo del receptor que se exprese.11
Receptor M1: se localiza en los ganglios autónomos, en la corteza cerebral y en glándulas secretorias como las glándulas salivales. Se acopla a la proteína Gq lo que da lugar a la activación de la Fosfolipasa C. Esta enzima cataliza la hidrólisis del fosfatidil-4,5-bifosfato (PIP2) produciendo segundos mensajeros: trifosfato de inositol (IP3) y diacilglicerol (DAG):
El DAG actúa sobre la Proteinokinasa C, que actúa fosforilando proteínas a nivel intracelular.
El IP3 aumenta los niveles de calcio en el citosol a partir de los depósitos intracelulares; por lo tanto, estimula procesos calcio dependientes como las secreciones y la contracción del musculo liso.
En el cerebro, la activación de estos receptores M1 postsinápticos inhibe canales de K+ que median el nivel basal de excitabilidad neuronal.
Receptor M2: predominan en el miocardio, nódulo aurículo sinusal y ventrículoauricular, autorreceptores y en mucho menor grado en zonas musculares lisas. Funciona a través de la proteína Gi que inhibe a la adenilciclasa, disminuyendo la síntesis de AMPc. Además promueve la apertura de los canales de K+ (este fenómeno se observa en las células marcapasos del nódulo sinoauricular), lo que es responsable de las acciones inotropas (fuerza de contracción) cronotropas (frecuencia cardiaca) y dromotropas(velocidad de conducción) negativas(disminuye) de la acetilcolina. En las neuronas, está presente en los axones e inhiben la excitabilidad neuronal generando una retroalimentación negativa para la liberación del neurotransmisor. En el músculo liso, genera respuestas opuestas a los receptores adrenérgicos Su agonista es la metocramina.
Receptor M3: se sitúan en glándulas secretorias, musculo liso, endotelio. Activa a la proteína Gq. En el musculo liso vascular produce un aumento en la concentración intracelular de Ca2+, lo cual induce la contracción, mientras que el mismo receptor, cuando se ubica en la zona del endotelio, va a causar la activación de la enzima sintetasa de oxido nítrico, el cual difunde y causa la relajación de las células musculo liso adyacentes. En las glándulas induce secreción de hormonas e incrementa las secreciones digestivas, bronquiales y lagrimales. En el musculo liso intestinal y genitourinario incrementa el peristaltismo,
RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
20
defecación y micción. A nivel bronquial causará broncoconstricción. En el musculo de la pupila causa miosis.
Receptor M4: se acopla a la proteína Gi y están presentes en neuronas ganglionares, vasos deferentes, útero y también se han descrito en glándulas secretoras y musculo liso. Tiene una ubicación específica en el conducto de Schlemm en donde disminuye la presión intraocular. Y esta implicado en procesos de memoria y analgesia a nivel del SNC.
Receptor M5: activan a la proteína Gq cumpliendo con la activación de
la fosfolipasa C. Está compuesto por 532aa. No se ha estudiado bien sus funciones.
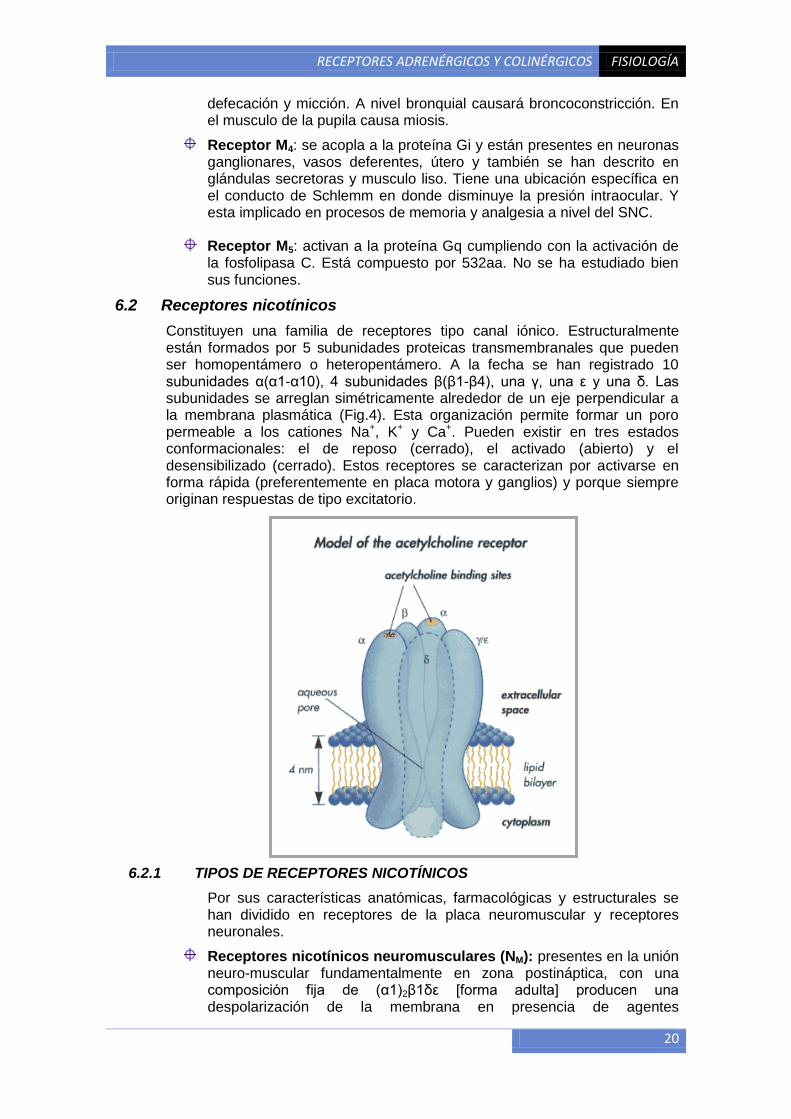
6.2 Receptores nicotínicos
Constituyen una familia de receptores tipo canal iónico. Estructuralmente están formados por 5 subunidades proteicas transmembranales que pueden ser homopentámero o heteropentámero. A la fecha se han registrado 10 subunidades α(α1-α10), 4 subunidades β(β1-β4), una γ, una ε y una δ. Las subunidades se arreglan simétricamente alrededor de un eje perpendicular a la membrana plasmática (Fig.4). Esta organización permite formar un poro permeable a los cationes Na+, K+ y Ca+. Pueden existir en tres estados conformacionales: el de reposo (cerrado), el activado (abierto) y el desensibilizado (cerrado). Estos receptores se caracterizan por activarse en forma rápida (preferentemente en placa motora y ganglios) y porque siempre originan respuestas de tipo excitatorio.
6.2.1 TIPOS DE RECEPTORES NICOTÍNICOS
Por sus características anatómicas, farmacológicas y estructurales se han dividido en receptores de la placa neuromuscular y receptores neuronales.
Receptores nicotínicos neuromusculares (NM): presentes en la unión neuro-muscular fundamentalmente en zona postináptica, con una composición fija de (α1)2β1δε [forma adulta] producen una despolarización de la membrana en presencia de agentes

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
21
biscuaternarios como el decametonium, lo que genera una excitación y aumento de la permeabilidad catiónica del Na+ y K+; tienen como agonista al feniltrimetilamonio y como antagonista competitivo al d-tubocurarina, pero son bloqueados irreversiblemente por α-toxinas de serpiente. Provoca la contracción muscular.
Receptores nicotínicos neuronales (NN): se localizan en los ganglios autonómicos, medula SR y SNC. Se han identificado homopentámeros de la subunidad α7, heteropentámero α2 y β4, y heteropentámeros de α4,β2 y α5. Se caracterizan por ser estimulados preferencialmente por 1,1-dimetil-4-fenilpiperazino, por ser bloqueados competitivamente por trimetafan y no competitivamente por agentes biscuaternarios; y, ser resistentes a las toxinas de serpientes. La activación de estos receptores por acetilcolina es responsable de la transmisión en el sistema nervioso autónomo. En los ganglios, los nervios presinápticos liberan acetilcolina, la cual induce una despolarización y genera u nuevo impulso en el nervio postsináptico. Esta estimulación ganglionar genera a su vez la liberación de acetilcolina de terminaciones parasimpáticas y de noradrenalina en las terminaciones adrenérgicas. En el SNC, la estimulación de receptores nicotínicos también genera despolarizaciones. Sin embargo, la presencia de receptores muscarínicos en el SNC y la complejidad de los efectos producidos, dificultan la identificación del componente nicotínico.

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
22
III. CONCLUSIONES
El sistema nervioso simpático se caracteriza por presentar una fibra nerviosa
preganglionar corta y una fibra nerviosa postganglionar larga, por lo tanto el
ganglio donde se realiza la última sinapsis está lejos del órgano efector.
Las neuronas que originan las fibras preganglionares se ubican en las
columnas intermedio lateral de la médula espinal.
Todas las fibras preganglionares del sector simpático liberan como
neurotransmisor acetilcolina (Ach).
La mayoría de las fibras postganglionares liberan como neurotransmisor a la
Noradrenalina (NA).
Gracias al sistema nervioso simpático, el organismo se encuentra mejor
preparado para luchar o huir.
Se entiende por receptor adrenérgico, el lugar donde van a actuar las
catecolaminas que han sido sintetizadas y almacenadas en forma de granulo,
en las terminaciones nerviosas postganglionares. El receptor en si no tiene
representación morfológica y su activación se realiza por mecanismos de
interacción enzimática.
El sistema nervioso parasimpático tiene origen en el mesencéfalo, mediante los
pares craneales III, VII, IX y X, también tiene origen en la porción sacra de la
médula espinal, mediante S2 y S3 principalmente, por ello se le denomina
sistema nervioso cráneo-sacral.
El sistema nervioso parasimpático actúa en condiciones de normalidad, relajo,
contrarresta y vuelve a su sitio la actividad del organismo.
Las fibras preganglionares parasimpáticas recorren casi todo el trayecto hasta
el órgano que van a controlar, el cual contiene las fibras posganglionares en su
pared.
Las fibras que segregan acetilcolina son colinérgicas y todas estas son
preganglionares simpáticas, preganglionares parasimpáticas y posganglionares
parasimpáticas.
En todo potencial de acción se sintetiza y se degrada acetilcolina para que
vuelva a ser usado en el mismo proceso.
Las características de transmisión colinérgica son diferentes dependiendo del
sitio del que suceda la sinapsis.
Los receptores colinérgicos se dividen receptores nicotínicos que tienen acción
en la fibra preganglionar; y, en receptores muscarínicos que actúan a nivel
postganglionar. Ambos grupos tienen subdivisiones a sus propiedades
farmacológicas y anatómicas.
Los receptores muscarínicos pertenecen a la familia de receptores acoplados a
la Proteína G, y presenta 5 subtipos: M1, M2, M3, M4, M5.
Los receptores nicóticos constituyen el grupo de receptores tipo canal iónico y
se clasifican en receptores nicotínicos neuronales (NM) y nicotínicos
neuromusculares (NM).

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
23
IV. COMENTARIOS DE LOS ARTICULOS
Artículo 1: RESPUESTA TERAPEUTICA DE LA PILOCARPINA EN RELACION A LA XEROSTOMIA INDUCIDA POR RADIOTERAPIA.
Comentado por: Katherine Parra Gálvez
Este artículo hace referencia al efecto que tiene el alcaloide Pilocarpina, como terapia
para contrarrestar el daño causado en las glándulas salivales, principalmente en la
Parótida, como consecuencia del sometimiento de los pacientes con cáncer, a
sesiones de radioterapia. La Pilocarpina tiene efectos parasimpáticos de estimulación
salival acondicionando a los receptores muscarínicos que actúan en glándulas
salivales.
Fernández y col., en su estudio tuvieron una muestra dividida en un grupo control (10
personas) a el que se le dio placebo y otro grupo de estudio (10 personas) al que se le
administró clorhidrato de pilocarpina al 1% 5mg cada 8horas al día por 7 semanas. En
ambas muestras se exceptuaron personas con enfermedades como diabetes,
pacientes que presenten reacción adversa a la pilocarpina y a aquella población que
consumía fármacos como atropina, etc.
El análisis que se realizo fue mediante la cuantificación de la saliva en tres fechas, la
primera cuantificación fue a la primera semana, observándose una diferencia no
significativa con respecto a la variación del flujo salival tanto en el grupo control como
en el grupo de estudio; la segunda, a la quinta, obteniendo resultados similares a la
primera pero destacando una disminución del flujo; y, la tercera, a la séptima semana
de administración de pilocarpina, evidenciándose diferencia significativa en ambos
grupos. Así, con el resultado de la segunda cuantificación se concluye que los efectos
degenerativos en las glándulas salivales se manifiestan a la 5ta semana, justificándose
en la disminución del flujo salival en ambas muestras.
En el tiempo de investigación no se presentaron casos de reacciones adversas a
comparación con otros estudios en los que si se dio los efectos aplicando dosis
similares. Finalmente, se demuestra la acción de la pilocarpina de manera positiva
como terapia para la xerostomía, basándose en la acción muscarínica sobre las
glándulas salivales serosas.

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
24
Artículo: 2: EXPRESIÓN DE RECEPTORES COLINÉRGICOS MUSCARÍNICOS EN PLACENTA HUMANO A TÉRMINO
Comentado por: María Huapaya Pardavé
El presente artículo trata de determinar la expresión de los receptores colinérgicos
muscarínicos M1 y M2 en la placenta humana a término.
Gracias a las investigaciones, sabemos que el transporte de micronutrientes al feto a
través de la placenta está regulado por el sistema colinérgico placentario, el cual está
conformado por la acetilcolina, las enzimas acetilcolinesterasa y la colina
acetiltransferasa y los receptores colinérgicos muscarínicos y nicotínicos. La activación
de estos receptores por la acetilcolina hace que se libere el factor de relajación del
músculo liso.
Existen estudios en ratas y humanos que han demostrado que la placenta expresa
receptores colinérgicos específicamente se ha demostrado que los receptores
colinérgicos muscarínicos M1 al M4 se encuentran localizados en el sincitiotrofoblasto,
la membrana basal y el membrana con borde en cepillo de las microvellosidades.
Además debido a la exposición de plaguicidas se ha demostrado una disminución en
la expresión de los receptores colinérgicos muscarínicos M1 y M2.
El objetivo del trabajo ya mencionado en líneas anteriores es determinar la expresión
de los receptores colinérgicos muscarínicos M1 y M2 en la placenta humana a término,
para lo cual se empleó una muestra de 20 placentas a término tomadas de pacientes
del Instituto Mexicano del Seguro Social de la Ciudad de Chihuahua, México.
Se excluyeron las pacientes con diagnóstico de diabetes mellitus pregestacional y
gestacional, hiper o hipotiroidismo, enfermedades renales, anomalías uterinas y
embarazo gemelar, también a pacientes con exposición a plaguicidas debido a que
ocasionaría una disminución en la expresión de los receptores M1 y M2. Se detectó la
expresión de los receptores mediante inmunohistoquímica y antisueros policlonales Al
analizar la expresión de los receptores colinérgicos muscarínicos M1 y M2, se
encontró una mayor expresión de los receptores colinérgicos muscarínicos M2.
Se concluyó que la expresión de los receptores M1 y M2 en la placenta humana a
término se localizó en el sincitiotrofoblasto de las vellosidades secundarias y terciarias
de la placenta a término.

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
25
Artículo 3: GREATER NICOTINIC ACETYLCHOLINE RECEPTOR DENSITY IN SMOKERS THAN IN NONSMOKERS: A PET STUDY WITH 2-18F-FA-85380
Comentado por: Daniella Núñez Díaz
El propósito del estudio fue determinar si el método utilizado es lo suficientemente
sensible para detectar diferencias entre los receptores nicotínicos de los fumadores y
no fumadores y si lo es, para determinar la magnitud de la diferencia observada in
vivo.
Las imágenes de los estudios PET de los receptores nicotínicos de acetilcolina fueron
reportados usando el subtipo específico α4β2 de radioligando 2-18-F-FA-85380 (2FA).
Se utilizó ese método para medir el volumen total de 2FA libre en plasma (Vt/fp) en 10
no fumadores y en 6 fumadores.
El estudio PET es in vivo y muestra la distribución de los efectos al fumar en la
densidad de los receptores nicotínicos en todo el cerebro, mientras que en los estudios
in vitro solo se muestran regiones seleccionadas.
Ya que las pruebas in vivo tienen algunas limitaciones, la sensibilidad del PET y 2FA
para observar las diferencias de densidad entre fumadores y no fumadores debió ser
realizada primero.
Según el estudio, la densidad de estos receptores es mayor en fumadores y en
animales tratados crónicamente con nicotina que en no fumadores o ex fumadores y
en animales tratados con solución salina. Los ratones fueron utilizados para cuantificar
los receptores después y antes de la administración de nicotina. En los humanos,
durante un prolongado tiempo de abstinencia, los estudios determinaron el tiempo del
retorno de las densidades del receptor y también determinaron la correlación de estos
efectos dinámicos con los síntomas al dejar de fumar. La radioactividad del plasma en
fumadores fue mayor que la de los no fumadores. Mientras tanto, la fracción de 2FA
libre fue similar en ambos grupos.
De acuerdo a las regiones en donde se evaluaron los niveles de 2FA, se observó que
en el tálamo los niveles fueron más altos, mientras que los más bajos se encontraron
en el cuerpo calloso. En todas las regiones se encontraron valores más altos en todas
las regiones cerebrales de los fumadores que en las de los no fumadores, a excepción
del tálamo. La región en donde los niveles no fueron tan diferentes fue en el cuerpo
calloso.
Finalmente, se encontró que los niveles en fumadores que excedían entre 25% a
200% a los niveles de los no fumadores, y que fumar un solo cigarro reducía los

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
26
receptores disponibles en un 88%. Los resultados fueron muy similares a otros
estudios anteriormente realizados.
El incremento se debe a que la nicotina interactúa con los receptores nicotínicos de
acetilcolina en las neuronas centrales y periféricas, estos receptores son canales
iónicos pentoméricos regulados por ligandos, que existen en varias formas
homoméricas y heteroméricas en el cuerpo mamilar. Los receptores nicotínicos
heteroméricos que contienen las subunidades α4 y β2, posiblemente con otras
subunidades, son los subtipos predominantes en el cuerpo mamilar y funcionan como
sustratos para los efectos adictivos de la nicotina. Este tipo de receptores tienen una
afinidad mayor por la nicotina.
Los efectos en el mesencéfalo son intrigantes, ya que incluye el tegumento ventral y la
sustancia negra, las cuales son regiones que segregan dopaminérgicos al estriado
dorsal y ventral. Por otra parte, el mesencéfalo cumple un papel importante en las
reacciones a abuso de drogas.
En conclusión, el presente artículo sirve de gran ayuda para los posteriores estudios
que se enfoquen en encontrar diversas curas de adicción al tabaco de acuerdo a la
densidad de los receptores nicotínicos, es decir, probablemente logrando que la
densidad de estos receptores sea menor, se reduciría la adicción por la nicotina que
contiene el tabaco. Es evidente que el propósito de este artículo fue realizado.

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
27
Artículo 4: INTERACCION ENTRE LOS POLIMORFISMOS DEL RECEPTOR β1 Y β2 ADRENÉRGICO COMO PREDICTOR DE RIESGO DE INSUFICIENCIA CARDIACA CRONICA
Comentado por: Karina Huamani Chipana
Se ha propuesto que los polimorfismos genéticos se asocian a las génesis o desarrollo
de diversas enfermedades cardiovasculares y a la variabilidad individual a la respuesta
farmacológica. Los receptores β-adrenérgicos (β-RA) son reguladores altamente
polimórficos e importante en la homeostasis cardiovascular.
El propósito de este trabajo es evaluar el genotipo y la interacción gen-gen de los
polimorfismos β1-AR Arg389Gly y β2-AR Arg16Gly, Gln27Glu y Thr164Ile, como
factores de riesgo de insuficiencia cardiaca (IC).
En el corazón normal, los ß1-RA predominan. En cambio, en el corazón disfuncional la
sobreactivación simpática lleva a una selectiva regulación negativa del ß1-RA y un
incremento relativo en los ß2-RA por sobre 40%. Dada la propiedad de los
polimorfismos ß-RA, en particular ß2-RA Arg16Gly y ß2-RA Gln27Glu de regular
negativa o positivamente al ß-RA, las interacciones entre los polimorfismos del ß-RA
son importantes en el desarrollo del síndrome de IC. Por lo tanto, para determinar si
las variantes ß1-RA Arg389Gly, ß2-RA Arg16Gly, ß2-RA Gln27Glu y ß2-RA Thr164Ile
solas o en combinación representan un factor de riesgo para desarrollar IC, se
procedió a evaluar 80 pacientes con IC crónica y 88 pacientes controles sanos.
Para poder determinar el ADN genómico, se obtuvo muestras de sangre periférica por
lisis hipotónica y extracción con fenol/cloroformo. Los polimorfismos ß2-RA Arg16Gly y
ß2-RA Gln27Glu se determinaron amplificando con una sola reacción de PCR el gen
del ß2-RA y posterior digestión con las enzimas de restricción NcoI y BbvI,
respectivamente.
Según este estudio se observó que en el análisis de frecuencias alélicas de los
polimorfos, el alelo ß2-RA Glu27 era más común en los pacientes con IC que en los
controles. Cuando se analizaron los tres genotipos por polimorfismo en forma
conjunta, la asociación del polimorfismo ß2-AR Gln27Glu con IC se mantuvo
significativa.
Puesto que los ß1 y ß2-RAs coexisten en el corazón, se evaluó si el polimorfismo ß2-
RA Gln27Glu combinado ya sea con el polimorfismo de ß2-RA Arg16Gly o ß1-RA
Gly389Arg aumentaban o podían influir el riesgo de IC.
Se encontró que la presencia adicional del alelo ß2-RA Arg16 o ß1-RA Gly389
aumentaba el riesgo relativo de IC que se atribuía al alelo ß2-RA Glu27.

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
28
Los estudios in vivo han demostrado que las personas que son homocigóticas para el
alelo ß2-RA Glu27 tiene una mayor respuesta al agonista que los sujetos homocigotos
para el alelo Gln27.
Se observa que hay un aumento en los niveles de catecolaminas en pacientes con
disfunción ventricular, en un primer momento es un mecanismo de compensación pero
progresivamente tiene un efecto crónico sobre el miocardio; como resultado hay un
incremento en la relación de los ß2-RA/ß1-RA. Por lo tanto, polimorfismos que
aumenten la actividad del β1-RA, o disminuya en el β2 podrían tener efectos
protectores sobre el miocardio.
En resumen, el alelo Glu27 del ß2-AR es un predictor de riesgo de IC, la cual aumenta
con la presencia adicional de los alelos ß2-RA Gly16 o ß1-RA Arg389. Por lo tanto el
genotipo y frecuencia del alelo ß2-RA Glu27Glu se relaciona a la etiología de la IC
crónica y también a la prevalencia de infarto al miocardio.
Concluyendo, este trabajo de investigación resulta ser un atractivo modelo para tener
un conocimiento sobre las interacciones entre los polimorfos de los receptores β1 y β2
adrenérgicos con la respuesta a fármacos, su relación con la susceptibilidad o
progresión de las enfermedades cardiovasculares; siendo estos receptores
importantes en la contractibilidad, en la frecuencia cardiaca y también como
importantes blancos farmacológicos.

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
29
Artículo 5: SINDROME ANTICOLINÉRGICO PERIFÉRICO
Comentado por: Marylin Herrera Ccoicca
El síndrome anticolinérgico es el conjunto de manifestaciones clínicas ocasionadas por
el antagonismo de la acetilcolina en el receptor muscarínico esto sucede por la
administración de fármacos .Sus signos y síntomas son variables y se hacen notorios
luego de la administración del fármaco y desaparecen después de algunos días de la
suspensión del fármaco. Este síndrome implica cambios a nivel central (en el estado
de animo, alteraciones en la marcha, memoria y desorientación, colapso
cardiorespiratorio y muerte; a nivel periférico (xerostomía, taquicardia, dolor
abdominal, visión borrosa, vomito, retención urinaria, íleo dinámico, entre otros.
Existen fármacos y sustancias con características anti colinérgicas y competitivas que
hacen que los receptores muscarínico funcionen de forma contraria a la habitual , esto
produce inhibición de la neurotransmisión colinérgica .Los antihistamínicos son uno de
los muchos fármacos que pueden inducir el síndrome anticolinérgico.
El caso es de una paciente de 38 años ocupación costurera, presenta síntomas de
congestión nasal, estornudo frecuente, otalgia, prurito en el paladar, que fueron
concomitantes con rinitis alérgica e infección de la faringe.
Se le realizaron diversos exámenes de laboratorio, el análisis de química sanguínea
detecto disminución de acido úrico en 1.78 mg/dL, el diagnostico de los senos
paranasales fue sinusitis maxilar bilateral, hipertrofia de los cornetes inferiores y rinitis
crónica.
Luego se efectuaron pruebas para diagnosticar que clases de alérgenos ocasionaban
los síntomas de irritación, prurito y rinitis alérgica, con estos resultados se le administro
vacunas de hiposensibilización y se le prescribieron los siguientes fármacos para
eliminar las molestias loratadina y ketotifeno .
En las siguientes visitas la paciente reporta la disminución de los síntomas, debido a
eso solo se le prescribió ebastina y furoato de mometazona ,se le recomendó a la
paciente a asistir a revisiones esporádicas para control. Luego de un tiempo la
ebastina produjo en la paciente disurea, alteraciones digestivas y vértigo; sin embargo
ya no mostraba alteraciones alérgicas pero perdió peso deforma injustificada. Se le
practicó la prueba de ELISA se obtuvieron resultados negativos, se le realizaron
estudios complementarios para diagnosticar una posible infección ,también se le
realizó un estudio de perfil tiroideo y química sanguínea, dando resultados normales
se descartaron alteraciones del metabolismo e insuficiencia renal. Luego de unos días

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
30
la paciente tuvo retención urinaria , alteraciones emocionales y caquexia .Debido a
esto la derivaron al servicio de psiquiatría .Se le siguió administrando antihistamínicos
para controlar su alergia .De acuerdo con el reporte neurológico se sospecho de
síndrome anti colinérgico periférico ,Se la derivo a urología para realizarle estudios
urodinámicos ,se le diagnostico obstrucción urinaria baja y disfunción vesiculo-
esfinteriana de esta forma se comprobó el síndrome anticolinérgico y se le
suspendieron los antihistamínicos .Se sometió ala paciente a tratamiento de
inmunoterapia con el cual desaparecieron la retención urinaria ,la perdida de peso
,las molestias gastrointestinales .Después de unos días la paciente tuvo síntomas
alérgicos y se auto prescribió antihistamínicos con los cuales volvió a tener problemas
para miccionar.
Existe una homología del receptor H-1 con el receptor muscarínico esto va a permitir
que los antihistamínicos funcionen utilizando estos receptores muscarínicos (M1-M5)
entre estos el M3 es el estimula parasimpáticamente las glándulas, el musculo liso del
endotelio vascular ,bronquial , gastrointestinal y urinario .En el caso de la paciente
originaron alteraciones en la digestión ,perdida de peso, y retención urinaria baja
,todos estos síntomas desaparecieron cuando se le suspendieron los
antihistamínicos.

RECEPTORES ADRENÉRGICOS Y COLINÉRGICOS FISIOLOGÍA
31
V. BIBLIOGRAFIA
1. CAMPBELL, AMG, ER WILLIAMS, D BARLTROP.: “Motor neuron disease and exposure to lead”. J Neurol Neurosurg Psychiatry 1970. 33:877-885.
2. HART, DE. 1988. “Neuropsychological Toxicology:Identification and Assessment of Human Neurotoxic Syndromes”. Nueva York: Pergamon Press.
3. Dr Eduardo Fernández Manchón pdf-Receptores Farmacológicos –capitulo 6 .
4. GOODMAN Y GILMAN: “Las bases farmacológicas de la terapéutica”. Capitulo 6 - pág. 134-138.
5. JUAN .C ALVARADO ALVA: “Manual de farmacología”.
6. WILSON, PAUWELS, AKESSO, STEWART, SPACEY. “Nervios craneales en la salud y la enfermedad”. Buenos Aires: Ed. Médica panamericana, 2006, 262 pp. (49-68), (115-140), (163-202).
7. GUYTON & HALL. “Tratado de fisiología médica”. Barcelona: Elservier, 2006, 1115 pp. (751).
8. HARDMAN, JOE. “Las bases farmacológicas de la terapéutica”. Madrid: McGraw-Hill, 2001, 1997 pp. (125-126).
9. GANONG, WILLIAM F.: “Fisiología Medica” – 18va
edición. Editorial “El Manual Moderno”, México. Pág.: 83-129.
10. FLORES, JESUS: “Farmacología Humana” - 4ta edición. Editorial
MASSON S.A., Barcelona – España, 2003. Pág.: 197-244.
11. RIVAS MURTON , CARLOS A.: “Farmacología vol. I” 1ª edición 2002 pág. 133-153
12. MENDOZA PATIÑO, NICANDRO: “Farmacología Medica”. Editorial Médica Panamericana, México - 2008. Pág.: 222-230.
13. RANG Y DALE: “Farmacología” – 6ta edición. Editorial ELSERVIER
S.L. , España – 2008. Pág.: 144-162.
14. http://www.iqb.es/cbasicas/farma/farma03/parte03/anticolinergicos01.htm
15. http://www.medigraphic.com/pdfs/revmexneu/rmn-2005/rmn054f.pdf





























