Embed Size (px)
Citation preview

1
1
Experimental evidences of human 2
coronary microvasculature and myocardial tissue 3
bacterial colonization during meningococcemia. 4
5
Jean Bergounioux1,2,3, Mathieu Coureuil1,4, Emre Belli5, Mohamed Ly5, Michelle 6
Cambillau6, Nicolas Goudin7, Xavier Nassif1,4,8and Olivier Join-Lambert1,4,8. 7
8 9 10 1 INSERM, U1151, Institut Necker Enfants Malades, Paris, France.; 2Université Versailles Saint Quentin en Yvelines, Versailles France; 3Service 11 de Neurologie et Réanimation pédiatrique, Hôpital Raymond Poincaré, APHP, Garches, France; 4Université Paris Descartes, Sorbonne 12 Paris Cité, Paris, France; 5Centre Chirurgical Marie Lannelongue, Clamard, France; 6Service de Biochimie, Hôpital Européen George 13 Pompidou, APHP, Paris, France 7SFR-Necker Cell Imaging Plateform, Imagine Institut, Paris France 8Laboratoire de Microbiologie 14 Hôpital Necker-Enfants malades, Assistance Publique-Hôpitaux de Paris, Paris France. 15
16
Corresponding Author : 17
Dr Jean Bergounioux 18
Réanimation et soins intensifs pédiatriques, Batiment Letulle, 3iém étage 19
Hôpital universitaire Raymond Poincaré, APHP, 104 Boulevard Raymond Poincaré, 92380 20
Garches, France 21
Phone: 01 47 10 79 00 DECT 41131 22
Fax: 01 47 10 68 93 23
Email: [email protected] 24
IAI Accepted Manuscript Posted Online 1 August 2016Infect. Immun. doi:10.1128/IAI.00420-16Copyright © 2016, American Society for Microbiology. All Rights Reserved.
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

2
The authors declare no funding, no conflict of interest and that no previous presentation of 25
this work was presented elsewhere. 26
27
E-mail addresses : 28
[email protected]; [email protected]; [email protected]; 29
[email protected]; [email protected]; [email protected]; 30
[email protected]; [email protected] 31
32
33
34
Keywords from your manuscript : 35
Neisseria meningitidis, Purpura fulminans, Septic shock, sepsis induced cardiac 36
dysfunction, xenograft, myocardial tissue 37
38
39
40
41
42
43
44
45
46
47
48
49
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

3
Abstract : 50
51
Background : Meningococcal septic shock is associated with profound vasoplegia, early and 52
severe myocardial dysfunction and extended skin necrosis responsible for a specific clinical 53
entity designated Purpura fulminans (PF). PF represents 90% of fatal meningococcal 54
infections. One characteristic of meningococcal PF is the myocardial dysfunction that occurs 55
in the early phase of sepsis. Furthermore, hemodynamic studies have shown that the 56
prognosis of meningococcal sepsis is directly related to the degree of impairment of cardiac 57
contractility during the initial phase of the disease. 58
Methods : To gain insight into a potential interaction of Neisseria meningitidis with the 59
myocardial microvasculature, we modified a previously described humanized mice model by 60
grafting to SCID mice human myocardial tissue. We then infected the grafted mice with N. 61
meningitides (Nm). 62
Results : Using the humanized-SCID mouse model we demonstrated that N. meningitidis 63
targets the human myocardial tissue vasculature, leading to the formation of blood thrombi, 64
infectious vasculitis and vascular leakage. 65
Conclusions : These results suggest a novel mechanism of myocardial injury in the course of 66
severe Nm Sepsis that is likely to participate to primary myocardial dysfunction. 67
68
69
70
71
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

4
72
Introduction: 73
74
Neisseria meningitidis is a human pathogen responsible for life-threatening infections 75
such as septic shock and meningitis, mainly in children and young adults [1]. Meningococcal 76
septic shock is associated with profound vasoplegia, early and severe myocardial dysfunction, 77
and extended skin necrosis responsible for a specific clinical entity designated Purpura 78
fulminans (PF). PF represents 90% of severe meningococcal infection fatal cases [2]. 79
80
Approximately 60% of patients admitted to intensive care units for severe sepsis or 81
septic shock present a clinical picture of cardiac dysfunction responsible for a mortality 82
ranging from 70 to 90%. On the other hand, mortality is 20% in septic patients without 83
cardiovascular involvement [3,4]. A specificity of meningococcal PF is the myocardial 84
dysfunction that occurs in the early phase of the sepsis and is often present from the start 85
[5,6]. By contrast, myocardial dysfunction appears during the late phase of sepsis with septic 86
shock caused by other Gram-negative infections. Furthermore, hemodynamic studies have 87
shown that the prognosis of meningococcal sepsis is directly related to the degree of 88
impairment of cardiac contractility during the initial phase of the disease [7,8]. 89
90
N. meningitidis (Nm) interacts with endothelial cells of the microvasculature 91
throughout the body [9,10]. Nm forms micro colonies on the apical surface of endothelial 92
cells and manipulates cell signaling pathways to disrupt cellular and endothelial tissue 93
architectures, eventually cross the vascular endothelium and colonize the surrounding tissue 94
or the adjacent organ [10-12]. A direct interaction of Nm with the myocardial tissue and 95
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

5
vasculature has not been demonstrated but strongly suspected. Indeed, postmortem 96
examination showed the presence of meningococci in the myocardial tissue associated with 97
histological features of myocarditis [13]. To gain insight into a potential interaction of Nm 98
with the myocardial microvasculature, we modified a previously described humanized mice 99
model by grafting to SCID mice human myocardial tissue [14]. Using this humanized SCID-100
mouse model we demonstrated that N. meningitidis targets the human myocardial tissue 101
vasculature, leading to the formation of blood thrombi, infectious vasculitis and vascular 102
leakage. These results suggest a novel mechanism of myocardial tissue injury in the course 103
of severe Nm Sepsis that is likely to participate to primary myocardial dysfunction. 104
105
106
Materials and Methods: 107
108
Bacterial Strains 109
We used the serogroup C meningococcal strain designated 2C4.3, and a nonpiliated isogenic 110
derivative (Nm ΔpilE) that was constructed by inserting a kanamycin resistance cassette in 111
the pilE gene locus [15]. The N. meningitidis strain 2C4.3, also known as clone 12 of the 112
clinical N. meningitidis strain LNP8013, is a naturally occurring pilin antigenic variant of the 113
original clinical isolate LNP8013, which expresses a pilin variant responsible for a high 114
adhesive phenotype (REF). N. meningitidis strains were stored frozen at −80°C in Giolitti-115
Cantoni broth (GCB) supplemented with 20% glycerol. 116
117
Mice 118
Six week-old CB17/Icr-Prkdcscid/IcrIcoCrl SCID female mice were purchased from Charles 119
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

6
River Laboratories (Saint Germain sur l’Arbresle, France). Experimental procedures were 120
performed in sterile conditions, and in accordance with the guidelines of the Institut 121
National de la Santé et de la Recherche Médicale (INSERM). The experimental protocol was 122
approved by the Animal Experimentation Ethics Committee of the Université Paris Descartes 123
(study registered under number 00309.0.02). 124
125
Graft Protocol 126
We used the experimental model described by Yan et al [14] that was initially designed to 127
study the expression and role of endothelial cell adhesion molecules for white blood cell 128
migration. Myocardial tissues were obtained from pediatric patients undergoing cardiac 129
surgery with myomectomy for left ventricular outflow tract obstruction in the Centre 130
Chirurgical Marie Lannelongue (Clamard, France). In accordance with French legislation, 131
human myocardial samples were obtained from patients whom legal tutors were informed 132
and did not refuse to participate in the study. The myocardial tissue was maintained on ice 133
cold cardioplegia medium (Custodiol® HTK Registered Trademark of Dr Franz Köhler Chemie 134
GmbH). Myocardial tissues were grafted under the back skin of SCID mice [16]. Briefly, mice 135
were prepared for transplantation by shaving the hair of the back and abdominal areas after 136
an intraperitoneal injection of ketamine 100 mg/kg and xylazine 10 mg/kg using a pair of 137
sponge forceps. Human Myocardial tissue was placed onto the wound bed. The transplants 138
were held in place with 6-0 non-absorbable monofilament suture materials, and the skin was 139
then sutured above the transplant. Grafted mice were used for N. meningitidis infection 140
experiments 15 days after human myocardial tissue transplantation. 141
142
Infection Protocol and Bacterial Counts 143
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

7
In vivo, iron-binding proteins such as transferrin and lactoferrin restrict the amount of ferric 144
iron available in body fluids to a level that does not support meningococcal growth [17]. To 145
counteract these iron-deficient conditions, N. meningitidis has several iron acquisition 146
systems, such as transferrin binding proteins A and B that are expressed when bacteria are 147
grown under iron-deficient conditions [18]. These receptors are specific of human 148
transferrin, therefore, bacterial strains were grown under iron-deficient conditions before 149
each infectious challenge, and human holotransferrin was added to the bacterial inoculum 150
to optimize bacterial growth in vivo [19, 20]. N. meningitidis strains were grown overnight at 151
37°C on GCB agar plates prepared without iron and supplemented with deferoxamine 152
(Desferal, Novartis) at a final concentration of 15 μM. Kanamycin (100 μg/mL) was added to 153
the medium to grow the kanamycin resistant ΔpilE strain. Bacterial colonies were harvested 154
and cultured in RPMI with 1% bovine serum albumin medium and 0.06 μM deferoxamine 155
with gentle agitation to reach the exponential growth phase. Bacteria were then 156
concentrated by centrifugation and resuspended in physiological saline containing 20 mg/mL 157
of human holotransferrin to promote bacterial growth in vivo (2914HT, R&D systems) [18]. 158
Preliminary experiments showed that a 100-fold-higher inoculum was necessary to obtain a 159
sustained bacteremia by intravenous injection of bacteria, as compared to intraperitoneal 160
route (data not shown). Mice were infected intraperitoneally with 0.5 mL of this bacterial 161
suspension. To assess bacteremia in infected animals, 10 μL of blood was sampled using a 162
heparinized hematocrit glass tube after puncture or the lateral tail vein, or at the time of 163
death by intracardiac puncture after intraperitoneal injection of a lethal dose of ketamine 164
and xylazine. 165
166
Bacterial counts were performed by plating serial dilutions of blood or of myocardial tissue 167
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

8
graft homogenates on GCB agar plates. Results were expressed in colony-forming units (CFU) 168
per mL of blood. 169
170
Histology and immunofluoresence 171
172
Mice were killed 24 hours after infection. Mouse myocardial tissue grafts and mouse heart 173
were removed, fixed in 10% buffered formalin and embedded in paraffin. Hematoxylin, 174
eosin, and safranin staining and immunofluorescence labeling were performed on 5-μm 175
tissue sections using the Leica BOND-MAX automated system (Leica-microsystems, Wetzlar, 176
Germany). Tissues were fixed overnight in PBS 4% PFA. Cells were incubated with 177
primary antibodies at the recommended concentration in PBS containing 0.3% BSA for 1 178
hr. After 3 washings in the same buffer, Alexa-conjugated phalloidin and DAPI (0.5 179
μg/ml) were added to Alexa-conjugated secondary antibodies for 1 hr. After additional 180
washings, coverslips were mounted in mowiol. 181
For fluorescence microscopy the following reagents were used: Polyclonal anti PilE 182
antiserum. Secondary antibodies used were: antirabbit Alexa 56. All samples were mounted 183
in Vectashield mounting reagent (Vector Laboratories, Eurobio ABCys, France). 184
Image acquisition was performed on a structured illumination microscope (Apotome 2 185
Zeiss) with a 63x (NA 1.4) oil objectives. Images were collected using the Zen 2012 186
(Zeiss) software. Figures were performed using Fiji (v 1.43) software and FigureJ (v 1.) 187
plugin. Final figures were created in Photoshop (Adobe, USA). 188
189
190
191
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

9
Results: 192
193
Human Myocardial tissue Xenograft in SCID mice is revascularized by mouse’s vessels: 194
195
N. meningitidis does not interact with non-human cells. In order to study the consequences 196
of a potential interaction between N. meningitidis and myocardial microvasculature we 197
modified a previously described humanized mouse model by grafting human myocardial 198
tissue to SCID mice. Human myocardial tissues were obtained from pediatric patients 199
undergoing cardiac surgery with myomectomy for left ventricular outflow tract obstruction 200
and were grafted less than 2 hours after surgical removal. Our first goal was to assess 201
whether the vascularization of human myocardial tissues connect with that of the mouse as 202
it is the case for human skin graft. As shown in Figure 1a the macroscopic aspect of the graft 203
at day 14 was satisfying showing a well-vascularized tissue. Histological microscopic analysis 204
of the graft showed a normal vascularization with healthy cardiomyocytes surrounding the 205
vessels. The architecture of cardiac cells and cardiac tissue were preserved and there was no 206
evidence of inflammatory cells infiltrates into the graft tissue (Figure 1b). The vessels 207
present in the graft are all microvessels with a diameter inferior to 300µm and all of them 208
presented the structure of a vein or a capillary with thin vascular wall without layer of 209
smooth muscle cells. We conclude that human myocardial tissue xenotransplanted into SCID 210
mice are efficiently revascularized tissue displays characteristics of human myocardial tissue. 211
212
Neisseria meningitidis colonizes the myocardium venous and capillary microvessels 213
214
Grafted mice were subsequently infected using 106 CFU IP as this inoculum produced 215
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

10
reproducible high and sustained bacteremia in infected animals. Twenty-four hours after 216
infection, animals were sacrificed. At this time, the explanted grafts were colored and well 217
perfused with a healthy aspect of the tissue. 218
Human myocardial tissue grafts were sampled and analyzed. All the vessels were colonized 219
by N. meningitidis, while extracellular bacteria adhering to the intraluminal surface of 220
endothelial cells of myocardial tissue microcirculation were seen in all human myocardial 221
tissue graft sampled from infected animals (3/3 myocardial tissue grafts obtained from 3 222
different experiments) (Figure 2,4 and 5). All the colonized vessels were capillaries or 223
venules with a thin wall. N. meningitidis is disposed as a layer of several strata of bacteria 224
uniformly distributed on the inner face of the vessel. Bacteria underlined the entire 225
endoluminal surface of vessels, or in some circumstances completely occluded the vascular 226
lumen (Figure 2a and 2b). 227
It should be pointed out that Hematoxylin, eosin, and safranin staining of sections containing 228
both human myocardial tissue graft and subcutaneous mouse paniculus carnosus 229
demonstrated that N. meningitidis interacted specifically with human graft vessels (Figure 3). 230
Furthermore, hearts of infected animals were sampled at the same time as the graft and 231
prepared following the same protocol and did not show any vascular colonization by Nm, as 232
expected (data not shown). Altogether these data demonstrated a specific interaction of Nm 233
with the human myocardial vessels. 234
235
236
N. meningitidis myocardium vascular colonization induced intravascular thrombosis and 237
vascular endothelial destruction as well as vasculitis. 238
239
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

11
240
Two different types of vascular lesions were observed in the myocardial tissue xenograft. 241
(i) The most frequent type of vascular lesion (type I) were vessel occlusions by infectious 242
clots (75% on a count of 100 vessels) which was mainly observed in vessels having an 243
average diameter of 20µm. These clots are composed by the aggregation of fibrin, blood 244
cells and bacteria (Fig 2). Only few vessels of the graft did not present any histological lesion 245
(8% on a count of 100 vessels). This septic clot had a very peculiar organization. The red 246
blood cells and fibrin clots were central to the vessel’s lumen and surrounded by a layer of 247
bacteria adhering to the vascular endothelium (Figure 4). The clots structural organization 248
suggests that the clot formation followed a centripetal process from the endothelium to the 249
center of the vessel. This organization suggests that the bacterial interaction with the 250
endothelium triggered the clotting process leading to the blood clot formation. 251
(ii) Another type of vascular lesion (type II) was observed less frequently (17% on a count of 252
100 vessels), was characterized by smaller vessels with an average size of 15 µm (count of 253
100 vessels). These lesions corresponded to vasculitis with massive infiltration by 254
polymorphonuclear cells (PMN), with a destruction of the vascular wall and disappearance 255
of the vascular lumen (Figure 5a and 5b). Unlike what was seen in the type I lesions, few 256
bacteria or bacterial clots are visible in these lesions. This loss of vascular wall integrity 257
results in vascular leakage leading to red cells getting in the perivascular space (Figure 4b). 258
It should be pointed out that in only 8% of the vessels no lesions was observed. 259
260
In many occurrences but specially in type II lesions, N. meningitidis was found outside of the 261
vascular lumen, in the myocardial tissue surrounding the vessels accompanied by red blood 262
cells, which indicated an endothelial rupture (Figure 4a and 4b). These images resembled 263
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

12
those described in skin purpuric lesions characteristic of the meningococcal purpura 264
fulminans [9]. Moreover, some bacteria were localized between the vessels and most 265
frequently in close proximity to polymorphonuclear cells (Fig 4c and Fig 5b). The colonization 266
of the surrounding tissues by N. meningitidis is clearly visible adjacent to the vessels (Figures 267
4c and 5b). 268
269
N. meningitidis adhesion to endothelial cells of the coronary microvascularisation and 270
infectious vasculitis require Type IV pilus 271
272
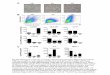
Type IV pili have a central role in meningococcal pathogenesis [15]. In vitro and in 273
vivo meningococcal adhesion to human endothelial cells has been shown to require Type IV 274
Pili. We subsequently tested the ability of a capsulated non-piliated derivative of N. 275
meningitidis 2C4.3 (ΔpilE) to target the human myocardial tissue graft vasculature. As shown 276
Figure 6, the non-piliated strain failed to target the human myocardial tissue graft as 277
demonstrated by the absence of bacteria sticking to the vessels endothelium. In addition, no 278
thrombosis nor vascular lesion was observed in the graft microvasculature when infected by 279
the non-piliated strain, thus demonstrating that the histologic lesions observed in the 280
vasculature of grafts infected with wild type N. menigitidis are type IV pilus dependent. 281
282
Discussion: 283
284
The model allowed, for the first time, an analysis of the intimate relations between N. 285
meningitidis, coronary microcirculation and myocardial tissue in vivo. Our results showed 286
that N. meningitidis adhere to the myocardium micro-vessel’s wall and induced a septic 287
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

13
vasculitis of the coronary microcirculation. Moreover, N. meningitidis induced septic 288
thrombosis of the micro vessels followed by a destruction of the vascular endothelium. The 289
damages observed resembled those observed in the skin of patients with meningococcemia. 290
Vascular lesions were similar to those described in patient’s skin purpuric lesions specific of 291
Purpura Fulminans, including thrombosis of vessels, perivascular infiltrates, and vascular 292
leakage (Figure 4) [10]. Inflammatory cells, including mononuclear cells and neutrophils 293
surrounded capillaries located in the superficial and mid dermis and infiltrated their walls 294
(Figure 2A and 2B). A mutant of N. meningitidis, defective for the pilin PilE, did not cause any 295
vasculitis, demonstrating that Type IV pilin is required for vasculitis. 296
297
Our results give experimental evidences that a direct interaction between N. 298
meningitidis and the heart takes place. To our knowledge, this represents the first report of 299
myocardial tissue colonization by bacteria. Moreover, our findings corroborate the 300
observations made by Monsalve et al. obtained from postmortem examination of three 301
patients suffering of meningococcemia showing the presence of myocarditis associated with 302
the presence of Gram-negative diplococci [13]. From this study it was hypothesized that N. 303
meningitidis colonize the myocardial tissue and induce tissue inflammation. Considering our 304
results, it is likely that the mechanism of this colonization is an infectious vasculitis of the 305
coronary microvascularisation. Destruction of the vascular endothelium allowed N. 306
meningitidis to enter the myocardial tissue generating neutrophils inflow and subsequent 307
local inflammation. 308
309
Diverse mechanisms underlying septic myocardial depression have been described 310
[21,22]. Sepsis-induced cardiomyopathy appears to be an excessive physiological adaptation 311
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

14
of the heart function to the sepsis and not a myocardial tissue injury. Indeed, clinical studies 312
have shown a specific hemodynamic profile of the sepsis induced cardiac depression, i.e. a 313
transient and global defect of the heart function, sometimes profound, followed by full 314
recovery [23]. It was shown that myocardial ischemia and necrosis are not the mechanisms 315
of myocardial septic dysfunction, the main hypothesis being a massive signaling to the heart 316
muscle. This signaling would be responsible for hibernation-like conditions where the 317
myocardial muscle down regulates its function in order to reduce energy expenditure, thus 318
preventing activation of cell death pathways of cardiomyocytes [22,24]. Endotoxins, Toll like 319
receptors (TLR) and cytokines mediate this signaling. Cytokines are produced and liberated 320
from activated immune cells after contact with bacterial compounds. The signaling is 321
systemic, via cytokine, as well as local, via TLR4 activation, leading to myocardial septic 322
dysfunction [22,24]. TLR4 must be present on macrophage and neutrophils to cause myocyte 323
dysfunction during endotoxemia [25,27]. Interleukin 6 has been shown to have an important 324
role in myocardial depressant activity in children with meningococcal septic shock [6]. Our 325
results, showing a massive presence of N. meningitidis in the coronary microvascularisation 326
as well as in myocardial tissue, suggest that N. meningitidis septicemia may be associated 327
with strong local signaling. Massive stimulation of the TLRs by N. meningitidis emphasizes by 328
the local septic thrombi and increased concentration of N. meningitidis LPS analog due to 329
endothelium adherence of the bacteria could generate such antigenic stimulation resulting 330
in strong innate immune signaling and myocardial dysfunction. Polynuclear infiltration of the 331
myocardial muscle has been demonstrated of critical importance in the TLR4 mediated 332
contractile dysfunction induced by sepsis [27]. Polynuclear infiltration is present in almost 333
20% of the vessels analyzed in our model and may also participate to myocardial dysfunction. 334
This model will help to provide a better understanding of the septic myocardial depression 335
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

15
induced by N. meningitidis but mouse and human differences in complement, binding 336
receptors, and signaling receptors would probably limit its exploitation at the molecular level. 337
338
In the case of severe meningococcal infection, our results showed vascular damages 339
of the coronary microcirculation, presence of massive amount of LPS and infiltration of 340
polymorphonuclear cells and bacteria in the myocardial tissue. Indeed, the histological 341
lesions of the coronary microcirculation observed in our model are mimicking the lesions 342
observed at the skin level in purpura fulminans. The results of our ex vivo experiments 343
suggest that these damages participate to the specificity of the N. meningitidis associated 344
sepsis-induced cardiomyopathy that take place early in the course of the disease and is 345
usually severe. The exact nature of the signaling and its consequences would need further 346
investigation but the trigger of the vascular damages appears to be linked to the presence of 347
type IV pili. Indeed the role of type IV pili in the disorganization of the cell-cell junctions and 348
in the opening of paracellular route in the brain capillaries has already been shown to allow 349
N. meningitidis to cross the blood brain barrier and invade the meninges and in the skin 350
capillaries to induce the skin lesions typical of the disease (Figure 6) [10,12]. All together, 351
these results may explain the early and profound septic cardiac dysfunction associated to N 352
meningitidis septic shock. 353
354
355
Conclusion: 356
357
We describe for the first time a model of human cardiac muscle tissue xenograft in 358
SCID mice providing a new tool to study intimate interaction of human pathogens with the 359
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

16
human heart. Our work demonstrates the tropism of N. meningitidis for human coronary 360
microcirculation in vivo, and its pilus-mediated specificity. This interaction with human 361
coronary microcirculation may play a role in the profound and early cardiac dysfunction that 362
classically goes with severe meningococcal sepsis. Our model will allow a better 363
understanding of the pathogenesis of septic heart dysfunction and will open the path to 364
specific treatments of this major complication of severe meningococcal sepsis. 365
366
367
Acknowledgments: We thank Virginie Lambert, Catherine Rucker, Gérard Pivert, Omar Aimer, 368
John Rohde and Isabelle Le Maner for technical assistance and advice. 369
370
371
372
Bibliography: 373
374
1- Stephens DS, Greenwood B, Brandtzaeg P. Epidemic meningitis, meningococcaemia and Neisseria 375
meningitidis. Lancet 2007; 369:2196–210. 376
2- De Greeff SC, de Melker HE, Schouls LM, Spanjaard L, van Deuren M. Pre-admission clinical course of 377
meningococcal disease and opportunities for the earlier start of appropriate intervention: a prospective 378
epidemiological study on 752 patients in the Netherlands, 2003-2005. Eur J Clin Microbiol Infect Dis. 2008 379
Oct;27(10):985-92 380
3- Vieillard-Baron A, Caille V, Charron C, Belliard G, Page B, Jardin F. Actual incidence of global left ventricular 381
hypokinesia in adult septic shock. Crit Care Med. 2008 Jun;36(6):1701-6. 382
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

17
4- Makwana N, Baines PB. Myocardial dysfunction in meningococcal septic shock. Curr Opin Crit Care. 2005 383
Oct;11(5):418-23. Review. 384
385
5- Thiru Y, Pathan N, Bignall S, Habibi P, Levin M. A myocardial cytotoxic process is involved in the cardiac 386
dysfunction of meningococcal septic shock. Crit Care Med. 2000 Aug;28(8):2979-83. 387
6- Pathan N, Hemingway CA, Alizadeh AA, Stephens AC, Boldrick JC, Oragui EE, McCabe C, Welch SB, Whitney A, 388
O'Gara P, Nadel S, Relman DA, Harding SE, Levin M. Role of interleukin 6 in myocardial dysfunction of 389
meningococcal septic shock. Lancet. 2004 Jan 17;363(9404):203-9. 390
7- Boucek MM, Boerth RC, Artman M, Graham TP Jr, Boucek RJ Jr. Myocardial dysfunction in children with 391
acute meningococcemia. J Pediatr. 1984 Oct;105(4):538-42. 392
8- Pathan N, Faust SN, Levin M. Pathophysiology of meningococcal meningitis and septicaemia. Arch Dis Child. 393
2003 Jul;88(7):601-7. Review. 394
9- Bernard SC, Simpson N, Join-Lambert O, Federici C, Laran-Chich MP, Maïssa N, Bouzinba-Ségard H, Morand 395
PC, Chretien F, Taouji S, Chevet E, Janel S, Lafont F, Coureuil M, Segura A, Niedergang F, Marullo S, Couraud PO, 396
Nassif X, Bourdoulous S. Pathogenic Neisseria meningitidis utilizes CD147 for vascular colonization. Nat Med. 397
2014 Jul;20(7):725-31. doi: 10.1038/nm.3563. Epub 2014 Jun 1. 398
10- Join-Lambert O, Lecuyer H, Miller F, Lelievre L, Jamet A, Furio L, Schmitt A, Pelissier P, Fraitag S, Coureuil M, 399
Nassif X. Meningococcal interaction to microvasculature triggers the tissular lesions of purpura fulminans. J 400
Infect Dis. 2013 Nov 15;208(10):1590-7. doi: 10.1093/infdis/jit301. Epub 2013 Jul 9. 401
11- Coureuil M, Lécuyer H, Scott MG, Boularan C, Enslen H, Soyer M, Mikaty G, Bourdoulous S, Nassif X, 402
Marullo S. Meningococcus Hijacks a β2-adrenoceptor/β-Arrestin pathway to cross brain microvasculature 403
endothelium. Cell. 2010 Dec 23;143(7):1149-60. doi: 10.1016/j.cell.2010.11.035. 404
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

18
12- Coureuil M, Mikaty G, Miller F, Lécuyer H, Bernard C, Bourdoulous S, Duménil G, Mège RM, Weksler BB, 405
Romero IA, Couraud PO, Nassif X. Meningococcal type IV pili recruit the polarity complex to cross the brain 406
endothelium. Science. 2009 Jul 3;325(5936):83-7. doi: 10.1126/science.1173196. Epub 2009 Jun 11. 407
13- Monsalve F, Rucabado L, Salvador A, Bonastre J, Cuñat J, Ruano M. Myocardial depression in septic shock 408
caused by meningococcal infection. Crit Care Med. 1984 Dec;12(12):1021-3. 409
14- Yan HC, Juhasz I, Pilewski J, Murphy GF, Herlyn M, Albelda SM. Human/severe combined immunodeficient 410
mouse chimeras. An experimental in vivo model system to study the regulation of human endothelial cell-411
leukocyte adhesion molecules. J Clin Invest 1993; 91:986–96. 412
15- Nassif X, Lowy J, Stenberg P, O’Gaora P, Ganji A, So M. Antigenic variation of pilin regulates adhesion of 413
Neisseria meningitidis to human epithelial cells. Mol Microbiol 1993; 8:719–25. 414
16- Barrandon Y, Li V, Green H. New techniques for the grafting of cultured human epidermal cells onto 415
athymic animals. J Invest Dermatol 1988; 91:315–8. 416
17- Calver GA, Kenny CP, Kushner DJ. Inhibition of the growth of Neisseria meningitidis by reduced ferritin and 417
other iron-binding agents. Infect Immun 1979; 25:880–90. 418
18- Simonson C, Brener D, DeVoe IW. Expression of a high-affinity mechanism for acquisition of transferrin iron 419
by Neisseria meningitidis. Infect Immun 1982; 36:107–13. 420
19- Holbein BE. Iron-controlled infection with Neisseria meningitidis in mice. Infect Immun 1980; 29:886–91. 421
20- Holbein BE. Enhancement of Neisseria meningitidis infection in mice by addition of iron bound to 422
transferrin. Infect Immun 1981; 34:120–5. 423
424
21- Zaky A1, Deem S, Bendjelid K, Treggiari MM. Characterization of cardiac dysfunction in sepsis: an ongoing 425
challenge. Shock. 2014 Jan;41(1):12-24. 426
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

19
22- Rudiger A, Singer M. Mechanisms of sepsis-induced cardiac dysfunction. Crit Care Med. 2007 427
Jun;35(6):1599-608. Review. 428
23- Parker MM, Shelhamer JH, Bacharach SL, Green MV, Natanson C, Frederick TM, Damske BA, Parrillo JE. 429
Profound but reversible myocardial depression in patients with septic shock. Ann Intern Med. 1984 430
Apr;100(4):483-90. 431
24- Rudiger A, Singer M. The heart in sepsis: from basic mechanisms to clinical management. Curr Vasc 432
Pharmacol. 2013 Mar 1;11(2):187-95. Review. 433
25- Arslan F, de Kleijn DP, Pasterkamp G. Innate immune signaling in cardiac ischemia. Nat Rev Cardiol. 2011 434
May;8(5):292-300. doi: 10.1038/nrcardio.2011.38. Epub 2011 Mar 29. Review. 435
26- Baumgarten G, Knuefermann P, Schuhmacher G, Vervölgyi V, von Rappard J, Dreiner U, Fink K, Djoufack C, 436
Hoeft A, Grohé C, Knowlton AA, Meyer R. Toll-like receptor 4, nitric oxide, and myocardial depression in 437
endotoxemia. Shock. 2006 Jan;25(1):43-9. 438
27- Tavener SA, Long EM, Robbins SM, McRae KM, Van Remmen H, Kubes P. Immune cell Toll-like receptor 4 is 439
required for cardiac myocyte impairment during endotoxemia. Circ Res. 2004 Oct 1;95(7):700-7. Epub 2004 Sep 440
Figure legends: 441
Figure 1: (a) viability of the myocardial tissue xenograft macroscopic aspect. (b): viability of the myocardial 442
tissue xenograft microscopic aspect. Eosin-hematoxilin coloration of the control graft showing normal 443
myocardial muscular cells and normal myocardial muscle architecture. Graft vessels of human origin are 444
perfused by red cells of mouse origin. 445
Figure 2: (a) Control graft of the non-infected animal showing a normal vessel of human origin in the graft tissue 446
with circulating red blood cells of mice origin in its lumen. (b) This image shows graft vessel from infected 447
animal with central infectious clot occupying the entire lumen of the vessel and large amount of bacteria 448
localized inside the clot and on the vessel internal surface. 449
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

20
Figure 3: the arrow 1 is in the human graft showing a human vessel infiltrated by polynuclear cells and a total 450
loss of the vessel architecture; the arrow 2 is in mouse surrounding tissue showing a mouse vessel of normal 451
aspect emphasizing the human specificity of Nm infection and vasculitis. 452
Figure 4: (a) This image shows a vessel occluded by a septic thrombus composed of meningococcus and a clot 453
of fibrin and platelets. The arrow 1 shows the central clot mainly composed of red cells; the arrow 2 shows the 454
ring of meningococci surrounding the clot; the arrow 3 shows meningococci outside of the vascular lumen. (b) 455
This image shows a vessel occluded by a septic thrombus composed of meningococcus and a clot of fibrin and 456
platelets with vascular lumen filed by the meningococci. The arrow 1 shows the central clot mainly composed of 457
red cells; the arrow 2 shows the ring of meningococci surrounding the clot; the arrow 3 shows a rupture in the 458
vascular endothelium with red cells and bacteria passing in the surrounding tissue. (c) This image shows 459
multiple fields reconstruction of apotome acquisitions (4 x 2); scale bare is 20 μm; upper left image: tissue 460
imaging using transmitted light; upper right image: circling of the blood vessels; lower left image: Apotome 461
imaging of the DAPI (blue) and 2C43 antibody (red); White arrows show N meningitidis present in the 462
surrounding tissue; lower right image showing a magnification of the lower left image rectangular selection with 463
central clot circled by a ring of N. meningitidis. 464
Figure 5: (a) This image shows massive infiltration of two vessels by polynuclear cells that results into the loss 465
of their architecture and the surrounding tissue. (b) This image shows multiple fields reconstruction of apotome 466
acquisitions (4 x 2); scale bare is 20 μm; upper left image: tile scan tissue imaging using transmitted light; 467
upper right image: circling of the blood vessels; lower left image: Apotome imaging of the DAPI (blue) and 468
2C43 antobody (red); lower right image showing a magnification of the lower left image rectangle with a 469
massive infiltration of the vessel by the PMN. White arrows show N meningitidis present in the surrounding 470
tissue. 471
Figure 6: Upper figure shows control graft with no infection. Central figure shows infection with isogenic 472
mutant (Nm ΔpilE) showing normal graft vessels and no inflammatory cells. Lower figure showing infection 473
with wild strain 2C4.3 showing N. meningitidis underlining the vascular endothelium (see black arrow). Graph a 474
showed similar levels of bacteremia in mice at H1 and H6 after infection with the wild-type 2C4.3 N. 475
meningitidis strain or with the Δ pilE derivative of 2C4.3 (p = ns) (2 mice per group). Graph b showed the 476
bacterial load of myocardial graft was lower in mice infected with the non piliated strain 24 hours after infection 477
(1 sample of myocardial tissue analyzed for both condition). 478
on May 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from