Embed Size (px)
Citation preview

MG-132is a cell permeable proteasome inhibitorwith a Kd of 4nM.
5.08J Biological Chemistry II (2016)ANS to Problem Set 8 Chemistry 5.08 ThisPSisfocusedonHMGRregulation and revisits the PCSK-9paper.1. HMG-CoAreductase(HMGR)isa97kDaintegralmembraneglycoproteinlocalizedto the ER.The N-terminalmembranedomainhaseighttransmembranehelicesandis sufficienttoregulatetheenzyme’sstability.AtthetimeofExperiment1describedbelow, the signalingpathway or pathways for thedegradationofHMGRinresponsetometabolic cueswerenotunderstood.Experiment2describesexperimentstofurtheraddressthis issuethatwerecarriedoutin2014.ReviewthemodelforcontrollingHMGRactivity presented inyour class notes.
Experiment1 wascarriedouttodeterminethefateofHMGRinthepresenceandabsenceofsterolsandinthepresenceorabsenceofaproteasomeinhibitor,MG-132. TheresultsoftheseexperimentsareshowninFigures1and2andaredescribedinthefigurelegendsassociatedwitheachoftheexperiments.
Digression:
© American Society for Biochemistry and Molecular Biology. Ravid T., et al. "The ubiquitin-proteasome pathway mediates the regulated degradation of mammalian 3-hydroxy-3-methylglutaryl-coenzyme A reductase." J Biol Chem. 2000 Nov 17;275(46):35840-7. All rights reserved. This content is excluded from our Creative Commons license. For more information, see https://ocw.mit.edu/help/faq-fair-use.
Figure1:Effect of MG-132proteasome inhibitor on the amounts of HMGR in a pulse-chaseexperiment. LP-90cells were pulse-labeled for 30 min with 150 mCi of[35S]-Met and either lysed immediately (lane 18) or chased for 5 h in the absence ofsterols (lane 19) or in the presence ofsterols(lanes20-25) andthe indicatedconcentrationof the proteasome inhibitor. Cells were lysed in lysis buffer in the presence ofthe detergent deoxycholate and the lysates were centrifuged for 30min at 16,000 x g to remove cell debris. HMGR was immunoprecipitated from the supernatantfraction with an antibody (Ab) targeting the HMGR membrane domain and analyzed bySDSPAGE and fluorography(Phosphorimager technology where the phosphor detects radioactivity, seerecitation 2/3 notes).
1
© American Society for Biochemistry and Molecular Biology. Ravid T., et al. "The ubiquitin-proteasome pathway mediates the regulated degradation of mammalian 3-hydroxy-3-methylglutaryl-coenzyme A reductase." J Biol Chem. 2000 Nov 17;275(46):35840-7. All rights reserved. This content is excluded from our Creative Commons license. For more information, see https://ocw.mit.edu/help/faq-fair-use.
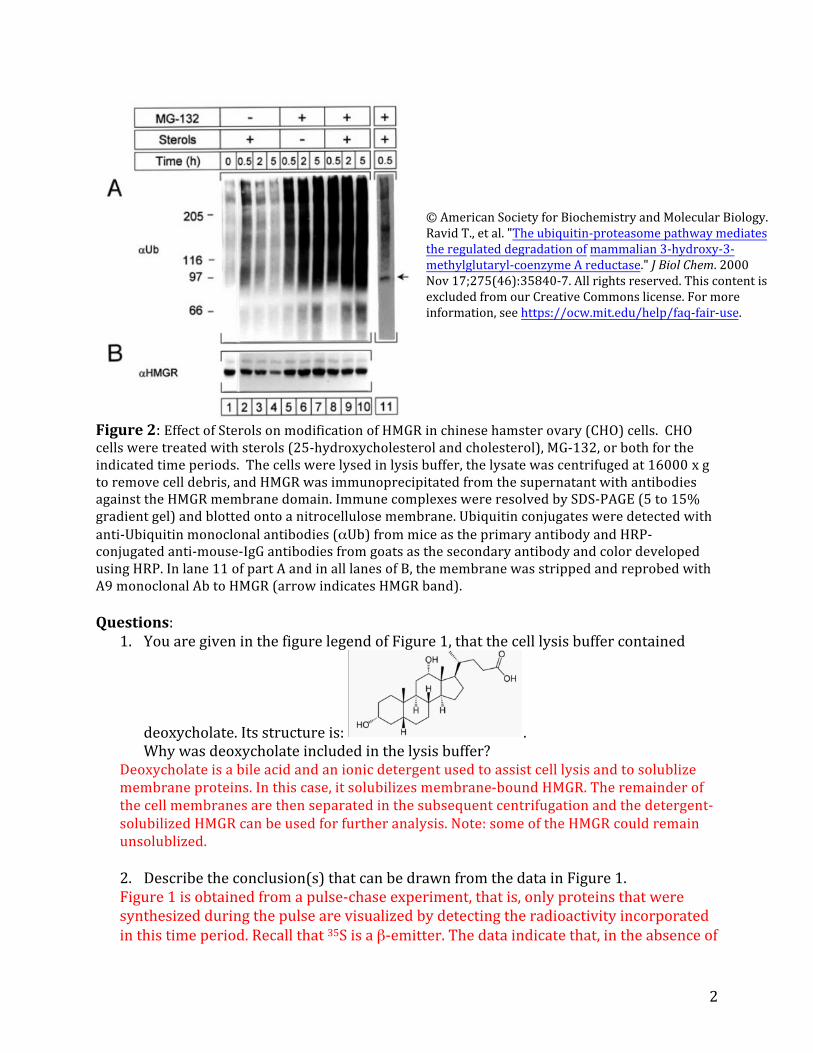
Figure2:Effect of Sterolson modification of HMGR in chinese hamster ovary (CHO)cells. CHOcells were treated with sterols (25-hydroxycholesterol and cholesterol), MG-132, or bothfor the indicated time periods. The cells were lysed in lysis buffer,the lysate was centrifuged at 16000 x gto remove cell debris, and HMGR was immunoprecipitated from the supernatant with antibodiesagainst the HMGR membrane domain. Immune complexes were resolved by SDS-PAGE(5 to 15% gradient gel) and blotted ontoanitrocellulose membrane. Ubiquitin conjugates were detected withanti-Ubiquitin monoclonal antibodies (αUb) from mice as the primary antibody and HRP-conjugated anti-mouse-IgG antibodies from goats as the secondary antibody and color developedusing HRP. In lane 11 of part A and in all lanes of B, the membrane was stripped and reprobed withA9 monoclonal Ab to HMGR(arrow indicates HMGR band).
Questions:
deoxycholate. Its structureis:Why was deoxycholate included inthelysisbuffer?
Deoxycholateis a bileacid and an ionic detergent used to assist cell lysis and to solublizemembrane proteins. In this case, it solubilizes membrane-bound HMGR. The remainder ofthe cell membranes are then separated in the subsequentcentrifugation and the detergent-solubilized HMGR can be used for further analysis. Note: some of the HMGR could remainunsolublized.
2. Describetheconclusion(s)thatcanbedrawnfromthedatainFigure1.Figure1isobtainedfromapulse-chaseexperiment,thatis,onlyproteinsthatweresynthesizedduringthepulsearevisualizedbydetectingtheradioactivityincorporatedinthistimeperiod.Recallthat35Sisaβ-emitter.Thedataindicatethat,intheabsenceof
1. Youare giveninthe figurelegendof Figure1, thatthe celllysis buffer contained
.
2

sterols,HMGRisstable,asitslevelsdecreaseonlymarginallyduringthe5hour chase.Inthepresenceofsterols,almostalloftheHMGRsynthesizedduringthepulseisdegradedafter5hours,indicatingthat itshalf-life is significantly shorter.The presence oftheMG-132,aproteasomeinhibitor,preventsthedegradationof HMGRatconcentrationsaslow as1µM,indicatingthat HMGRisusuallydegradedbytheproteasomeinthepresenceofsterols.
3. InFigure2 lanes 1-4(noMG-132)provideanexplanationforwhytherearesomanyhighmolecularweightspeciesobserved?
HMGRisamembraneproteinthatisglycosylatedandgetsubiquitinatedinthepresenceofsterolstotargetitfordegradation.Allthree of these factors could contributetotheheterogeneityinthegel:Membraneproteinscanoftenexhibitsignificant heterogeneityingel electrophoresisbecauseofvariableloadingwithSDS.Differentextentsofglycosylationcanalterthetotalmolecular weightof aproteinand alsoaffectitsloadingwithSDS.Finally,ubiquitinationchangesthetotalmolecularweightoftheprotein,dependingonthenumberofubiquitinmoleculesthatareattached.Whichofthesefactorscontributesmostwilldependontheexperimentalconditions,e.g.howthesampleswereprepared.
Onewaytoexaminethebasisfortheheterogeneity,forexamplemodificationwithsugars,istotreatthesamplewithadeglycosylase thatremovesallthesugarsattachedtoanasparagineinadefinedmotif aminoacidmotif onthecell surface.Toperhapsremoveheterogeneityduetoincompleteunfolding,onecouldboilthesamplelongerinSDS.Toremoveanyheterogeneityassociatedwith ubiquitination,youcould treatthe samplewithdeubiquitinases.NotethattheprimaryantibodyusedintheWesternBlotwas ananti-ubiquitinantibody,soonlyubiquitinated HMGRis detected.
4. Describeconclusionsthatcanbedrawnfromthisexperiment.Inthisexperiment,HMGRwasimmunoprecipitatedandanalyzedbyWesternBlotsafter cellgrowth with and withoutsterols and with and withoutMG-132.Withoutsterolsandwithout MG-132(lane1),thereareafewhighmolecularweightbandsthatreact with the ubiquitinantibody.Uponadditionof sterols (lanes 2-4),highermolecularweightbandsforubiquitinappearthatthenloseintensityovertime.Thehighestlevelsofubiquitinationappeartobeatthe30mintimepoint.AscanbeseeninpartB,HMGRgetsmodifiedand/ordegradedoverthistimeperiod,asthelevelsforthe97kDaversiondecrease.Likely,HMGRisgettingrapidlyubiquitinateduponadditionofsterolsandisthendegradedbytheproteasome.InthepresenceofMG-132,but intheabsenceofsterols(lanes5-7),thereisastrongaccumulationofhighmolecularweightbandsthatreactwiththeubiquitinantibody.Atthesametime,levelsofHMGRincrease(partB,lanes 5-7).WhensterolsareaddedinthepresenceofMG-132(lanes8-10),theintensityofthehighmolecularweightbandsincreasesevenfurther.Undertheseconditions,however,thereisnoincreaseinlevelsofHMGR(part B,lanes8-10).
Thereareanumberofconclusionstobedrawnfromthisexperiment.First,additionofsterolsleadstopolyubiquitinationofHMGRanddegradationbytheproteasome.InhibitingtheproteasomeleadstosignificantaccumulationofubiquitinatedHMGR
3

(comparelanes2-4and8-10).Second,itseemsthatHMGRalsogetspolyubiquitinatedintheabsenceofsterols(lanes5-7),butthesespeciesonlyaccumulatewhentheproteasomeisblocked.Becausethehalf-life of HMGRinthe absence of sterols is long(seeFigure1),maybethereisaproteasome-dependent deubiquitinationactivitythat prevents HMGRdegradationintheabsenceofsterols.Inhibitingtheproteasomeleadstoaccumulationoftheubiquitinatedspecies.
Thirdandmuchmoresubtly,HMGRistranscriptionallyregulatedaswell:intheabsence of sterols,its levels increase whenthe proteosomeisblocked(part B,lanes5-7),butitslevelsdonotincreaseinthepresenceofsterolswhentheproteasomeisblocked (partB,lanes 8-10).NotetheimagesinFigure2cannotbequantifiedbythe“eyeball”method.Itisthusverydifficulttotellaboutsmallchangeswithoutanalyzingtheintensityofthebandsmorequantitatively.
5. Describehow theαUb-HRP-anti-mouseantibodysandwichworkstoallowvisualizationoftheproteinsofinterest,inthiscaseubiquitin.
Theprimaryantibody,αUb,bindstotheubiquitinonthenitrocellulosemembrane.αUbwasraisedinmice.ThesecondaryantibodybindsmouseIgGandisconjugatedtoHRP.HRPcan oxidizeprecursorsubstratesintocoloreddyesthat deposit wheretheyaregeneratedandallowfor visualizationofubiquitin.
6. Givenwhatyouhavelearnedaboutthemechanismoftheproteasome,proposeamechanismbywhichMG-132inhibitionmightoccur.WhatmaybetheissueswithusinganaldehydeasaninhibitoringeneralinhumansandwithMG-132specifically?
MG-132isacompetitivereversible,covalentinhibitoroftheproteasome.Thealdehydegetsattackedbytheactivesitehydroxylofthethreoninenucleophilewiththehelpofitsα-aminogroupthatfunctions as a generalbasecatalyst.Recallthattheactivesitethreonine is atthe N-terminusinthehumanproteasome.Thereactionmayalsobetime-dependent,ifthehemiacetalformedbytheattackbythreonineisstabilized.Thepeptidepartof theinhibitor provides thespecificity.
Aldehydescanbeproblematicasinhibitorsforseveralreasons.Theyareoftenhydratedandsincetheactiveformoftheinhibitormustbethealdehyde,thereactiveconcentrationislowerthanthe“weighed”amountofaldehydedependingonthelevelofhydration.Insidethecellaldehydesareingeneralrapidlymetabolizedtoacidsoralcoholsthatwillbemuchpoorer, non-covalent inhibitors.
7. ArethedatapresentedinFigures1and2consistentwiththemodelforHMGRregulation that you learnedabout inclassandyourreading?
Thedatapresentedhereareinprincipleconsistentwiththemodeldescribedinclassandpresented onthe PP slides.Note,however,thatthereisnoinformationontheother
4

proteins involved inthis process.Thedataonlysuggest that HMGRisindeedubiquitinated and degraded inthepresenceof sterols.
2. Fromthedata describedabovethereareNOmoleculardetails,thatis, informationabouttheproteinsE1,E2andE3thatmustbeinvolvedinthisprocess.Infact,themodeldiscussedinclass shouldincludegp78andUbc7whichareE3andE2proteinsinvolved inthe ERAD(endoplasmicreticulumassociateddegradation process).
Experiment 2: StudiesinthelastdecadehaveshownthattherearethreeE3proteincomplexesinvolvedinERAD,oneofwhichincludesgp78.Songetalreportedthatgp78,which they showed was associated with Insig1,is responsible for degradationof HMGRinresponsetosterols.TheyalsoreportedthatanotherERADE3,TRC8,interactswithInsig1andalsoplaysaroleinHMGRdegradation.
InitiallytheycarriedouttheexperimentshowninFigure3.
© The American Society for Cell Biology. Tsai,Y.C., G.S. Leichner, et al. "Differential regulation of HMG-CoA reductase and Insig-1 by enzymes of the ubiquitin-proteasome system." Mol Biol Cell. 2012 Dec 1; 23(23): 4484– 4494. All rights reserved. This content is excluded from our Creative Commons license. For more information, see https://ocw.mit.edu/help/ faq-fair-use.
RecentlyTsaietalstudiedmouseembryonicfibroblasts(MEFs)fromaliver-specificgp78knockoutmouse.
Figure3: Levels andlengthof gp78in twodifferent sets of primaryMEFs (from twoseparatepregnancies of mice) were examined because of the complexity of the targeting strategy for theknockout of gp78inorder to determine ifthe knockout experiment was successful. Ignore the blackarrow inthe Figure. The β-actin serves as aloadingcontrol (actin should be, and is, present at the same levels in WT and KO cells).
Theythenused35Spulse-chasemetaboliclabelingandimmunoprecipitationtoquantitativelyassessthe degradationoftheendogenousHMGRintheseMEFs. Thecellsweregrownundercomplexconditionstomaintaintheirviability(detailshavebeenomitted)andthenpulselabeledfor30minwith[35S]-Metand Cys.This labelingwas followedbya chasein“cold”mediumintheabsenceorpresenceofsterols.Atindicatedtimes(Figures4AandB),theendogenousHMGRwasimmunoprecipitatedwithantibodiestothetransmembranedomainofHMGR(asdescribedabove).
5

Figure4AandB. Resultsofpulse-chaseexperimentsdescribedabove.
Figure 4 A, B, and C © The American Society for Cell Biology. Tsai,Y.C., G.S. Leichner, et al. "Differential regulation of HMG-CoA reductase and Insig-1 by enzymes of the ubiquitin-proteasome system." Mol Biol Cell. 2012 Dec 1; 23(23): 4484–4494. All rights reserved. This content is excluded from our Creative Commons license. For more information, see https://ocw.mit.edu/help/faq-fair-use.
Figure4C.(IP=Immunoprecipitation;IB=Immunoblot/Westernblot)
IntheexperimentwhoseresultsareshowninFigure4C,thecellswereallowedtoaccumulateHMGRbymediamanipulation,followedbyadditionofsterols(whereindicated)inthepresenceorabsenceoftheproteasomeinhibitorMG-132.After60min,the cellswerelysedandHMGRwasimmunoprecipitatedwithHMGRantibodiesraisedinrabbit.Theimmunoprecipitatesweresequentiallyimmunoblottedwithmousemonoclonalantibodiestoubiquitin(toppart)andHMGR(bottompart).
Questions: 1. Describewhat thedata inFigure3tellyouaboutthegp78knockoutmouse.Figure3indicatesthatgp78isindeedknockedoutinthemouse.Thereisnobandforgp78intheWesternBlot,whiletheβ-Actincontrollaneshowsnochange.
2. Describewhat theresultsin Figure4A-CtellyouaboutthestudieswithembryonicfibroblastsrelativetothemodelforHMGRregulation.
ThedatainFigure4Aand4BindicatethatthelevelsofHMGRinthegp78knockoutare thesame as inthe wild-type inthe presence or the absence of sterols.TheresultsinFigure4Cfurtherindicatethat HMGR in thegp78knockout isstill getting ubiquitinatedinthepresenceofsterols.Thus,thegp78knockout behaveslikeWTunderall conditionstested,indicatingeitherthat gp78isnotinvolvedinubiquitinatingHMGRor thatthere is analternative pathway.
6

3. Theresults givenabove aredifferentfromtheearlierreportedstudiesofSongetal. Howmightthedifferencesbetweenthisstudyandtheearlierstudyberesolved? I amnotaskingfordetails,butfor howyouwould goaboutdetermining the basis for the differences betweenthe two studies, which would inturn, providetheimpetus forfutureexperiments.
4. TheexperimentaldatainthisPSweretakeninpartfromMolecular Biology of the Cell 234484(2012).Inthispaper,whichisverythoughtful,theygostep bystep throughthedifferencesintheirexperimentalprotocolincomparisonwiththose reportedbySong et al. However, theyalsoofferexplanationsforhow theearlier experimentscouldhavebeencarriedoutmorecarefully.Thecomplexityofthe experimentsandthemodelcellsoccurinmanyexperimentsdonetoday.Thusthis paperisimportanttothinkabout,asmanyissuesthatmustbeconsideredin experimentaldesignarearticulated.Think aboutthedetailsofhowSongetal.determinedthatgp78andTRC8are involvedinmediatingHMGRdegradation.Werethereexperimentsdoneinvitroor invivo? What werethetypesofcellsused? Didtheyhavegoodantibodiestothe proteins or weretheantibodiestotheepitopetags(e.g.myc)?Invitroandafter HMGRdetergentsolubilization,maybegp78canmediateubiquitinationofHMGR. Etc
3. Inrecitation8,youread thepaper inwhich Crispr-cas9wasusedtoremovethePCSK-9proteinfrom3T3L1cellsandfrommouselivercells.Thisquestionisfocusedontheexperimentaldetailsdescribedinthatpaper.Thegeneforpcsk-9 isshowninFigure5.
Figure5. pcsk-9 gene where the black rectangles are the targeted exons within this gene: exon oneand exontwo are atthe farleftand farrightof the gene.
© source unknown. All rights reserved. This content is excluded from our Creative Commons license. For more information, see https://ocw.mit.edu/help/faq-fair-use.
© source unknown. All rights reserved. This content is excluded from our Creative Commons license. For more information, see https://ocw.mit.edu/help/faq-fair-use.
Figure6. The target for the gRNA from exon1
Both exon1 and exon2 of pcks-9 (Figure5)weretargetedforguide RNAdesignwiththechoiceforexon1targetingshowninFigure6.ShowninFigure7aretheresultsoftheexperimenttodeterminewhichexon (onevstwo) would be besttarget.
7

© American Heart Association, Inc. “Permanent Alteration of PCSK9 with in Vivo CRISPR-Cas9 Genome Editing” Ding, Strong, Patel et al. Circulation Research 115, 488-492 2014. All rights reserved. This content is excluded from our Creative Commons license. For more information, see https://ocw.mit.edu/help/faq-fair-use.
Figure7Target effects in mousecells and livers subsequent to CRISPR-cas9 expression.A.Surveyor assaysperformed with genomic DNA from 3T3-L1cells transfectedwithCas9andguide RNA targetingeither exon 1(gRNA-1) or exon 2(gRNA-2). B. Surveyor assays performed with genomic DNA from liver samples takenfrom mice from mice 3 days after receiving a controladenovirus expressing GFP or adenovirus expressing Cas9andgRNA-1. In panel B, there are two GFP controls (A and B) and two mouse liver tissue experiments(C and D).
. Finally, PCRamplificationandSangersequencingofthetargetsiteofmanysamples of mouselivergenomicDNA subsequent totheCRISPR-cas9studiesgaveresults,someofwhich are shownin Figure8.
Figure8.Sequencingof liver genomic DNA based on the target from the gRNA sequence.
© source unknown. All rights reserved. This content is excluded from our Creative Commons license. For more information, see https://ocw.mit.edu/help/faq-fair-use.
Questions: 1. Why were the exons and notthe intronstargetedbyDinget al ?
TheexonsendupinthemRNAandultimatelycodefortheprotein.Alteringtheintronsthat areremovedbysplicingwilllikelystillresult in protein beng present.
2. Thetargetedregionofexon1isshowninFigure6. TherearethreefeaturesofthistargetDNAthatarehighlighted:thelarge rectangularbox,thesmallboxandthearrow.Givenyour understandingthe Crispr-cas9technologydescribewhythesethree features are highlighted and why they are essentialforthesuccessful targetingpcks-9.
8

GGT (GGN) in the small rectangle is the required PAM sequence, that is the protospaceradjacent motif that is required for Cas9 (with the two nuclease domains) to bind the targetDNA. It must immediately follow the target sequenceinthelargerectangleonthe3’ sideinthe non-target strand of the DNA. gRNAtargeting sequence is in the large rectangle and experimentation in this systemhas shown that 20 nucleotides,inthiscase, precedingthePAM sequence in the genomicDNA was effective for obtainingthe required double strand DNAcutting. The two strands of ds DNAtargeted must be separated (see your recitation handout). Remarkably this systemdoes not require a helicase or ATP to effect this strand separation.
3. The first experiments described in the paper determined whether exon 1 or exon 2would be the target of choice. An agarose gel is shown in Figure 7 describing theresults. Describewhat you observein Figure 7 Aand B and the how theseobservationsledtothe next sets of experiments.
The experiments in Aare carried out onthe tissue culture cells (invitro) and the experiments in B are carried out on the mice. The results may or may not be the same if thetissue culture cells are not good models for the animal. (Note, mice not be good models for humans). The results use PCR and agarose gels to determine if the sequence targetedremains intact or if it is degraded. In the tissue culture cells (3T3-L1)onecan seein thecontrol mock infection that the sequence remains intact. However in the adjacent twolanes in the experiment targeting exon 1 the sequence has been extensively degraded,while in the experiment targeting exon 2, the sequence remains intact. This type of experiment provide the impetus to focus on gRNAtargeting exon 1. gRNAtargeting exon 2 would have no effect on exon 1. Similar experiments were carried out with mice. Theadenovirus targets the liver and DNAfromthe liver was examined. Acontrol used GFP. The results fromtwo separate experiments on mice are shown in B. Both samples are extensivelydegraded,but todifferent extents.ThustheCrispr-cas9targetingappearstobeworkingwhenexon1 is targeted.
4. Ontargetand off-target DS DNAcleavage is a major concern with the Crispr-cas9technology. What experiments using methods similar to those described in Figure 7were carried out to determine off-targetcleavage?
Off target cleavage is apparently still a problemwith this technology. They thus sought the 10siteswithin the genome most closely matched to the on-target site and most likely to give rise to off target cleavage. They examined as stated in the paper 6 sites with 3mismatches and some with 4 mismatches. Using the Surveyor assay they found and the method describedinFigure7theyfoundnoevidenceofofftarget cleavage.
5. TheCrisprcas9 technology leads to ds DNAcleavage that can be repaired in anumber of ways. What are the two general mechanisms of repair? Explain how thedata shown in Figure 8 was obtained and what it tells you about which mechanismof repair predominates in this particular set of experiments based onthe datashown.
As shown below, taken fromyour recitation handout, theirare two methods for repair ofDS breaks. Oneisnon template dependent, non homologous end (NHEJ)repairwerethebluntended cleavage isrepairedvia insertionsordeletions ofa few nts,orvia frame shifts.
9

The second method HDR involves template dependent repair and results in insertion of asequence of interest, among other outcomes. The PCR reactions used in Figure 7 was used to acquire the datain Figure 8. One can see that most of the examples shown reveal deletionsofinsertionsofa few ntsandthustheNHEJrepairappearstobeoccurring.
10

MIT OpenCourseWare https://ocw.mit.edu
5.08J Biological Chemistry II Spring 2016
For information about citing these materials or our Terms of Use, visit: https://ocw.mit.edu/terms





























