Embed Size (px)
Citation preview

Learning Objectives✓ Differentiate between monosaccharides and polysaccharides in regard to
structure and function.
✓ Differentiate between proteoglycans and glycoproteins in regard to structureand function.
✓ What are the key chemical properties of fatty acids?
✓ What are the major lipids? Describe their biochemical functions.
Lipids and carbohydrates, along with proteins and nucleic acids, constitute thefour major classes of biomolecules. We have already looked at proteins,
emphasizing their roles as enzymes. Although carbohydrates and lipids play adiverse array of roles in living systems, their function as fuels is most obvious. Thisfunction is evident to anyone who examines the nutritional information on thelabels of containers of foods. In the parlance of such labels, lipids are designatedas fats, for reasons that we will discover in Chapter 10.
Lipids and carbohydrates provide the energy to power all biochemicalprocesses that take place inside a cell or organism. Their role as fuels is so para-mount that the taste and texture of these molecules elicits gustatory pleasure,and most animals, including human beings, are behaviorally motivated to seekout foods rich in lipids and carbohydrates. For reasons to be considered in later
SECTION
4Carbohydrates and Lipids

chapters, lipids provide much more usable energy per gram than do carbohy-drates. Yet, most organisms maintain supplies of both types of fuel. Why have twosources of fuel if one of them is so much more energy rich? The answer is that alllipids require oxygen to yield biologically useful energy. Although carbohydratescan release energy when they react with oxygen, they can also release energywhen oxygen is scarce, as would be the case for the leg muscles of a runner sprint-ing to the finish line or for a bacterium growing in an oxygen-free environment.Thus, the use of both lipids and carbohydrates as fuel provides biochemical flexi-bility for meeting the various biological needs of an organism. We will spend agood deal of time examining the biochemical processes by which these fuels areturned into useful cellular energy. In addition to serving as fuel molecules, carbo-hydrates and lipids serve as fuel deposits.
Carbohydrates and lipids also play important structural roles. For instance, carbohydrates provide the strength of plant cell walls, whereas lipids are ubiquitousin their role as membrane components. Indeed, carbohydrates and lipids can bejoined together to form particular membrane components called glycolipids. Finally,these two classes of molecules play essential roles in signal-transduction pathways.
In Chapter 9, we will examine the biochemical properties of carbohydrates, high-lighting some of their roles other than that of fuel. In Chapter 10, we will do the samefor lipids, paying particular to the hydrophobic properties of these molecules.
Chapter 9: Carbohydrates
Chapter 10: Lipids

122
CHAPTER
9 Carbohydrates9.1 Monosaccharides Are the Simplest
Carbohydrates
9.2 Monosaccharides Are Linked toForm Complex Carbohydrates
9.3 Carbohydrates Are Attached toProteins to Form Glycoproteins
9.4 Lectins Are Specific Carbohydrate-Binding Proteins
Carbohydrates are carbon-based molecules that are rich in hydroxyl ( )groups. Indeed, the empirical formula for many carbohydrates is ( )n—
literally, a carbon hydrate. Simple carbohydrates are called monosaccharides. Com-plex carbohydrates—polymers of covalently linked monosaccharides—are calledpolysaccharides. A polysaccharide can be as simple as one comprising two iden-tical monosaccharides. Or it can be as complex as one consisting of dozens of dif-ferent monosaccharides that are linked to form a polysaccharide composed ofmillions of monosaccharides. Monosaccharides are the monomers that make uppolysaccharides, just as amino acids are the monomers that make up proteins.However, the nature of the covalent bonds linking the monosaccharides in a poly-saccharide are much more varied than the canonical peptide bond of proteins.The variety of monosaccharides and the multiplicity of linkages forming polysaccha-rides mean that carbohydrates provide cells with a vast array of three-dimensionalstructures that can be used for a variety of purposes, as simple as energy storage oras complex as cell–cell recognition signals.
9.1 Monosaccharides Are the Simplest CarbohydratesWe begin our consideration of carbohydrates with monosaccharides, the simplestcarbohydrates. These simple sugars serve not only as fuel molecules but also asfundamental constituents of living systems. For instance, DNA is built on simple
C¬H2O¬OH
Grains or cereal crops are an abundant source of carbohydrates worldwide. Katherine LeeBates, a Wellesley College English professor, immortalized America’s grain-producing capacityin her words to “America the Beautiful.” Professor Bates was inspired to pen her words afterexperiencing the grandeur of America’s West while visiting Colorado College. [David NotonPhotography/Alamy.]

sugars: its backbone consists of alternating phosphoryl groups and deoxyribose,a cyclic five-carbon sugar.
Monosaccharides are aldehydes or ketones that have two or more hydroxylgroups. The smallest monosaccharides, composed of three carbons, are dihydrox-yacetone and D- and L-glyceraldehyde.
C
OHH
CH2OH
HO
D-Glyceraldehyde(an aldose)
C
HHO
CH2OH
HO
L-Glyceraldehyde(an aldose)
CH2OH
O
CH2OH
Dihydroxyacetone(a ketose)
C CC
Carbonyl group
O
C
Aldehyde
O
CR H R’RKetone
O
C
Monosaccharides and other sugars areoften represented by Fischer projections.Recall from Section 2, that, in a Fischerprojection of a molecule, atoms joined toan asymmetrical tetrahedral carbonatom by horizontal bonds are in front ofthe plane of the page, and those joinedby vertical bonds are behind the plane.
123
Dihydroxyacetone is called a ketose because it contains a keto group, whereas glycer-aldehyde is called an aldose because it contains an aldehyde group. They are referredto as trioses (tri- for three, referring to the three carbon atoms that they contain). Sim-ilarly, simple monosaccharides with four, five, six, and seven carbon atoms are calledtetroses, pentoses, hexoses, and heptoses, respectively. Perhaps the monosaccharides ofwhich we are most aware are the hexoses, glucose and fructose. Glucose is an essen-tial energy source for virtually all forms of life. Fructose is commonly used as a sweet-ener that is converted into glucose derivatives inside the cell.
Carbohydrates can exist in a dazzling variety of isomeric forms (Figure 9.1).Dihydroxyacetone and glyceraldehyde are called constitutional isomers becausethey have identical molecular formulas but differ in how the atoms are ordered.Stereoisomers are isomers that differ in spatial arrangement. Glyceraldehyde has asingle asymmetric carbon atom and, thus, there are two stereoisomers of thissugar: D-glyceraldehyde and L-glyceraldehyde. These molecules are a type ofstereoisomer called enantiomers, which are mirror images of each other.
ISOMERSHave the same molecular formula
but different structures
STEREOISOMERSAtoms are connectedin the same order but
differ in spatial arrangementC
OHH
CH2OH
CH2OH
CH2OH
HO
Glyceraldehyde(C3H6O3)
Dihydroxyacetone(C3H6O3)
CONSTITUTIONAL ISOMERSDiffer in the order of attachment of atoms
C C O
C
OHH
CH2OH
HO
D-Glyceraldehyde(C3H6O3)
ENANTIOMERSNonsuperimposable mirror images
C
C
HHO
CH2OH
HO
L-Glyceraldehyde(C3H6O3)
COHHO C
CHO
CH2OHD-Altrose
(C6H12O6)
DIASTEREOISOMERSIsomers that are not mirror images
OHH C
OHH C
OHH C
H OHC
CHO
CH2OHD-Glucose(C6H12O6)
HO HC
H OHC
H OHC
EPIMERSDiffer at one of several asymmetric
carbon atoms
OHH C
CHO
CH2OHD-Glucose(C6H12O6)
HHO C
OHH C
OHH C
HO HC
CHO
CH2OHD-Mannose(C6H12O6)
HO HC
H OHC
H OHC
ANOMERSIsomers that differ at a new
asymmetric carbon atom formed on ring closure
O
OH
OH
CH2OH
OH
HO
�-D-Glucose(C6H12O6)
O OH
OH
CH2OH
OH
HO
�-D-Glucose(C6H12O6)
Figure 9.1 Isomeric forms of carbohydrates.
1249 Carbohydrates
CHO
C
C
H OH
CH2OH
OHH
CHO
C
C
HO H
CH2OH
OHH
CHO
C
C
H OH
C
OHH
OHH
C OHH
CH2OH
CHO
C
C
HO H
C
OHH
OHH
C OHH
CH2OH
CHO
C
C
H OH
C
HHO
OHH
C OHH
CH2OH
CHO
C
C
HO H
C
HHO
OHH
C OHH
CH2OH
CHO
C
C
H OH
C
OHH
HHO
C OHH
CH2OH
CHO
C
C
HO H
C
OHH
HHO
C OHH
CH2OH
CHO
C
C
H OH
C
HHO
HHO
C OHH
CH2OH
CHO
C
C
HO H
C
HHO
HHO
C OHH
CH2OH
CHO
C
CH2OH
H OH
D-Glyceraldehyde
D-Erythrose D-Threose
CHO
C
C
H OH
C
OHH
OHH
CH2OHD-Ribose
CHO
C
C
HO H
C
OHH
OHH
CH2OHD-Arabinose
CHO
C
C
H OH
C
HHO
OHH
CH2OHD-Xylose
CHO
C
C
HO H
C
HHO
OHH
CH2OHD-Lyxose
D-Allose D-Altrose D-Glucose D-Mannose D-Gulose D-Idose D-Galactose D-Talose
11
22
33
11
22
33
44
11
22
33
44
55
11
22
33
44
55
66
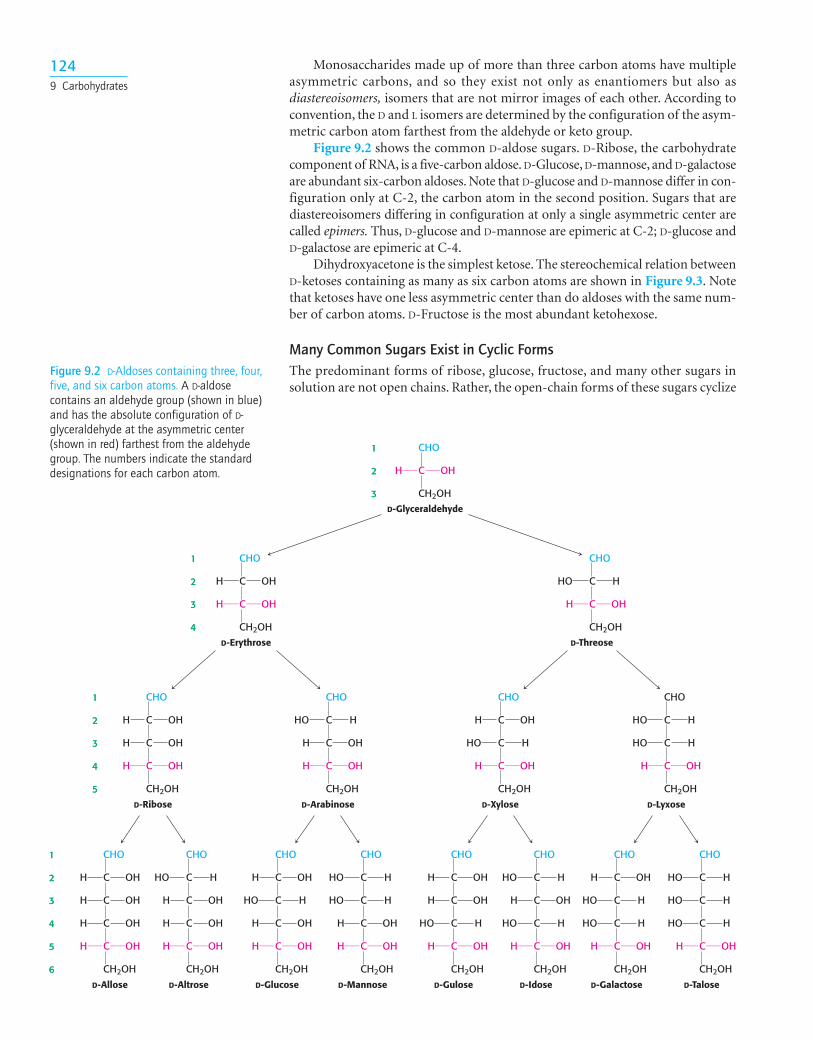
Figure 9.2 D-Aldoses containing three, four,five, and six carbon atoms. A D-aldosecontains an aldehyde group (shown in blue)and has the absolute configuration of D-glyceraldehyde at the asymmetric center(shown in red) farthest from the aldehydegroup. The numbers indicate the standarddesignations for each carbon atom.
Monosaccharides made up of more than three carbon atoms have multipleasymmetric carbons, and so they exist not only as enantiomers but also asdiastereoisomers, isomers that are not mirror images of each other. According toconvention, the D and L isomers are determined by the configuration of the asym-metric carbon atom farthest from the aldehyde or keto group.
Figure 9.2 shows the common D-aldose sugars. D-Ribose, the carbohydratecomponent of RNA, is a five-carbon aldose. D-Glucose, D-mannose, and D-galactoseare abundant six-carbon aldoses. Note that D-glucose and D-mannose differ in con-figuration only at C-2, the carbon atom in the second position. Sugars that arediastereoisomers differing in configuration at only a single asymmetric center arecalled epimers. Thus, D-glucose and D-mannose are epimeric at C-2; D-glucose andD-galactose are epimeric at C-4.
Dihydroxyacetone is the simplest ketose. The stereochemical relation betweenD-ketoses containing as many as six carbon atoms are shown in Figure 9.3. Notethat ketoses have one less asymmetric center than do aldoses with the same num-ber of carbon atoms. D-Fructose is the most abundant ketohexose.
Many Common Sugars Exist in Cyclic FormsThe predominant forms of ribose, glucose, fructose, and many other sugars insolution are not open chains. Rather, the open-chain forms of these sugars cyclize
O
Pyran
1259.1 Monosaccharides
+R
C
O
HOR�H R
C
HO OR�
HAldehyde Alcohol Hemicetal
C
CO
C
OHH
OHH
C
CO
C
HHO
OHH
C
CO
C
OHH
C
OHH
OHH
CH2OH CH2OH CH2OH CH2OH
CH2OH CH2OH
CH2OH CH2OH
CH2OH
CH2OH
CH2OH
CH2OH
CH2OHCH2OH
CH2OH CH2OH
C
CO
C
HHO
C
OHH
OHH
C
CO
C
OHH
C
HHO
OHH
C
CO
C
HHO
C
HHO
OHH
CO
Dihydroxyacetone
11
22
33
44
55
66
11
22
33
44
55
11
22
33
44
11
22
33
C
CO
OHH
D-Erythrulose
D-Ribulose D-Xylulose
D-Psicose D-Fructose D-Sorbose D-Tagatose
Figure 9.3 D-Ketoses containing three,four, five, and six carbon atoms. The ketogroup is shown in blue. The asymmetriccenter farthest from the keto group,which determines the D designation, isshown in red.
C
C
C
H OH
C
HHO
OHH
C OHH
CH2OH
CC
CH
OH
OH
C
C OH
H
OH
HO
H
CH2OH
H
HO
=
D-Glucose(open-chain form)
�-D-Glucopyranose
O H
OH
CH2OH
H
OH
OH
H
H
HO
H
�-D-Glucopyranose
O OH
H
CH2OH
H
OH
OH
H
H
HO
H
1
2
6
5
4
3 2
1
3
4
5
6
Figure 9.4 Pyranose formation. The open-chain form of glucose cyclizes when theC-5 hydroxyl group attacks the oxygenatom of the C-1 aldehyde group to form anintramolecular hemiacetal. Two anomericforms, designated � and �, can result.
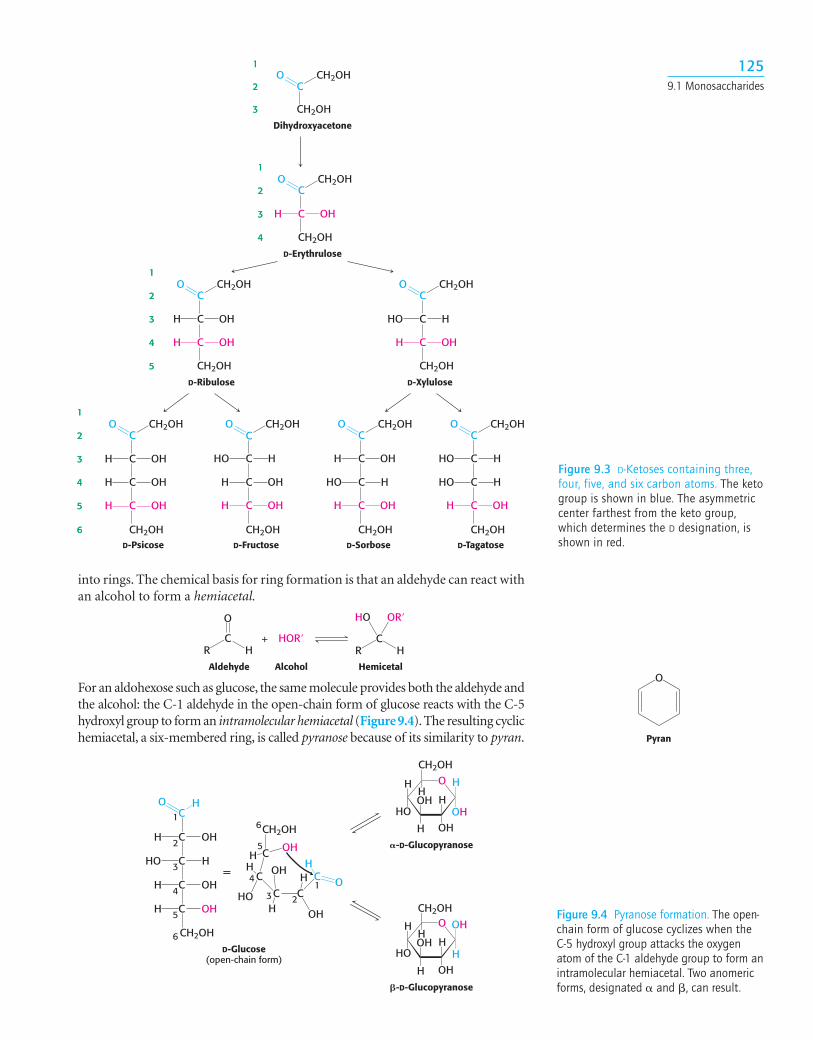
For an aldohexose such as glucose, the same molecule provides both the aldehyde andthe alcohol: the C-1 aldehyde in the open-chain form of glucose reacts with the C-5hydroxyl group to form an intramolecular hemiacetal (Figure 9.4).The resulting cyclichemiacetal, a six-membered ring, is called pyranose because of its similarity to pyran.
into rings. The chemical basis for ring formation is that an aldehyde can react withan alcohol to form a hemiacetal.

1269 Carbohydrates
C
CO CH2OH
C
HHO
C
OHH
OHH
CH2OH
C
O
CH2OHOHH
HOH
H
HOH2COH
=
OCH2OH
OH
HO
H
H
OH
HOH2C
H
D-Fructose(open-chain form)
�-D-Fructofuranose(a cyclic form of fructose)
C
CC
1
2
3
4
5
6
6
5
43 2
1
Figure 9.5 Furanose formation. The open-chain form of fructose cyclizes to a five-memberedring when the C-5 hydroxyl group attacks the C-2 ketone to form an intramolecularhemiketal. Two anomers are possible, but only the � anomer is shown.
C
C
C
H OH
C
HHO
OHH
C OHH
CH2OH
HOC
C
C
H OH
C
HHO
OHH
C OHH
CH2OH
O OH
O OH
H
CH2OH
H
OH
OH
H
H
HO
H
Cu2+ Cu+ Cu2O
H2O, HO–
O
Furan
+R
C
O
HOR�R� R
C
HO OR�
R�
Ketone Alcohol Hemiketal
Similarly, a ketone can react with an alcohol to form a hemiketal.
The C-2 keto group in the open-chain form of a ketohexose, such as fructose, canform an intramolecular hemiketal by reacting with either the C-6 hydroxyl groupto form a six-membered cyclic hemiketal or the C-5 hydroxyl group to form afive-membered cyclic hemiketal (Figure 9.5). The five-membered ring is calleda furanose because of its similarity to furan.
The depictions of glucopyranose (glucose) and fructofuranose (fructose) shownin Figures 9.4 and 9.5 are Haworth projections. In such projections, the carbon atomsin the ring are not written out. The approximate plane of the ring is perpendicularto the plane of the paper, with the heavy line on the ring projecting toward the reader.
We have seen that carbohydrates may contain many asymmetric carbonatoms. An additional asymmetric center is created when a cyclic hemiacetal isformed, creating yet another diastereoisomeric form of sugars called anomers. Inglucose, C-1 (the carbonyl carbon atom in the open-chain form) becomes anasymmetric center. Thus, two ring structures can be formed: �-D-glucopyranoseand �-D-glucopyranose (see Figure 9.4). For D sugars drawn as Haworth projec-tions in the standard orientation as shown in Figure 9.4, the designation � meansthat the hydroxyl group attached to C-1 is below the plane of the ring; b means thatit is above the plane of the ring. The C-1 carbon atom is called the anomeric carbonatom, and the � and � forms are called anomers. An equilibrium mixture ofglucose contains approximately one-third � anomer, two-thirds � anomer, and�1% of the open-chain form.
Because the � and � isomers of glucose are in an equilibrium that passesthrough the open-chain form, glucose has some of the chemical properties of freealdehydes, such as the ability to react with oxidizing agents. For example, glucosecan react with cupric ion (Cu2�).

OCH2OH
OH
HO
H
H
OH
HOH2C
H
-D-Fructofuranose
O CH2OH
OH
H
HO
H
H
OH
H
HO
H
-D-Fructopyranose
OOH
CH2OH
HO
H
H
OH
HOH2C
H
-D-Fructofuranose
O OH
CH2OH
H
HO
H
H
OH
H
HO
H
-D-Fructopyranose
Figure 9.6 Ring structures of fructose. Fructose can form both five-membered furanose andsix-membered pyranose rings. In each case, both � and � anomers are possible.
1279.1 Monosaccharides
�-D-Pyranose, found in honey, is one of the sweetest chemicals known. The�-D-furanose form is not nearly as sweet. Heating converts �-pyranose into the�-furanose form, reducing the sweetness of the solution. For this reason, cornsyrup with a high concentration of fructose in the �-D-pyranose form is usedas a sweetener in cold, but not hot, drinks.
Monosaccharides are Joined to Alcohols and Amines Through Glycosidic BondsThe biochemical properties of monosaccharides can by modified by reactionwith other molecules. These modifications increase the biochemical versatility ofcarbohydrates, enabling them to serve as signal molecules or rendering themmore susceptible to combustion. Three common reactants are alcohols, amines,and phosphates. A bond formed between the anomeric carbon atom of glucoseand the oxygen atom of an alcohol is called a glycosidic bond—specifically, anO-glycosidic bond. O-glycosidic bonds are prominent when carbohydrates arelinked together to form long polymers and when they are attached to proteins.In addition, the anomeric carbon atom of a sugar can be linked to the nitrogen
Solutions of cupric ion (known as Fehling’ solution) provide a simple test for thepresence of sugars such as glucose. Sugars that react are called reducing sugars;those that do not are called nonreducing sugars. Reducing sugars can often non-specifically react with other molecules. For instance, as a reducing sugar, glucosecan react with hemoglobin to form glycosylated hemoglobin. Changes in theamount of glycosylated hemoglobin can be used to monitor the effectiveness oftreatments for diabetes mellitus, a condition characterized by high levels of bloodglucose (p. 386). Although the glycosylation of hemoglobin has no effect on oxy-gen binding and is thus benign, similar reducing reactions between sugars andother proteins are often detrimental to the body. Modifications known asadvanced glycolsylation end products (AGE) have been implicated in aging, arte-riosclerosis, and diabetes, as well as other pathological conditions. AGE is thename given to a series of reactions between an amino group not participating ina peptide bond in a protein and the aldehyde form of a carbohydrate.
The furanose-ring form of fructose also has anomeric forms, in which � and� refer to the hydroxyl groups attached to C-2, the anomeric carbon atom (see Fig-ure 9.5). Fructose forms both pyranose and furanose rings. The pyranose form pre-dominates in fructose free in solution, and the furanose form predominates inmany fructose derivatives (Figure 9.6).
An ester
R’R
O
C O
A phosphoester
PR O–
O–
O O
C O
OCH2OH
HH
HOOH
H
H
OH
H
O
OCH2OH
HH
OH
H
H
OH
H
OHα α1 4
α-1,4-Glycosidic bond Figure 9.8 Maltose, a disaccharide. Twomolecules of glucose are linked by an �-1,4-glycosidic bond to form thedisaccharide maltose. The angles in thebonds to the central oxygen atom do notdenote carbon atoms. The angles areadded only for ease of illustration.
Glucose 6-phosphate(G6P)
Dihydroxyacetonephosphate
(DHAP)
2OPO32�
O
HO
OH
CH
OH
OH
CH2OPO32�
HHO
H
C
CO
C OHH
C OHH
CH2OH
R =
O H
OH
H
HO
H
H
OH
H
HO
CH3
�-L-Fucose(Fuc)
O OH
H
CH2OH
H
HN
OH
H
HOH
CH3
H
�-D-Acetylgalactosamine(GalNAc)
CH3
O OH
H
CH2OH
H
HN
OH
H
HH
HO
�-D-Acetylglucosamine(GlcNAc)
O COO–
H
H
H
H
OH
OH
NHR
H
H3C
Sialic acid (Sia)(N-Acetylneuraminate)
C
O
C
O
C
O
Figure 9.7 Modified monosaccharides.Carbohydrates can be modified by theaddition of substituents (shown in red)other than hydroxyl groups. Such modifiedcarbohydrates are often expressed on cellsurfaces.
atom of an amine to form an N-glycosidic bond. Carbohydrates can also form anester linkage to phosphates, one of the most prominent modifications incarbohydrate metabolism. Examples of modified carbohydrates are shownin Figure 9.7.
9.2 Monosaccharides Are Linked to Form ComplexCarbohydrates
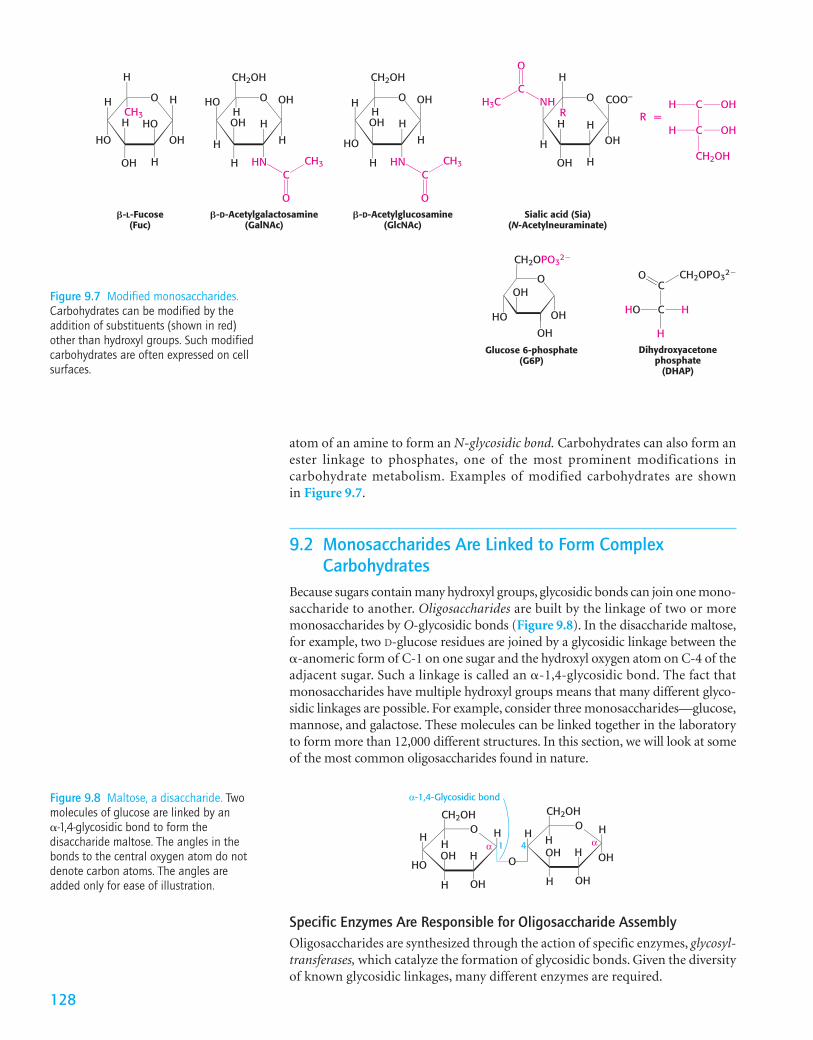
Because sugars contain many hydroxyl groups, glycosidic bonds can join one mono-saccharide to another. Oligosaccharides are built by the linkage of two or moremonosaccharides by O-glycosidic bonds (Figure 9.8). In the disaccharide maltose,for example, two D-glucose residues are joined by a glycosidic linkage between the�-anomeric form of C-1 on one sugar and the hydroxyl oxygen atom on C-4 of theadjacent sugar. Such a linkage is called an �-1,4-glycosidic bond. The fact thatmonosaccharides have multiple hydroxyl groups means that many different glyco-sidic linkages are possible. For example, consider three monosaccharides—glucose,mannose, and galactose. These molecules can be linked together in the laboratoryto form more than 12,000 different structures. In this section, we will look at someof the most common oligosaccharides found in nature.
128
Specific Enzymes Are Responsible for Oligosaccharide AssemblyOligosaccharides are synthesized through the action of specific enzymes, glycosyl-transferases, which catalyze the formation of glycosidic bonds. Given the diversityof known glycosidic linkages, many different enzymes are required.
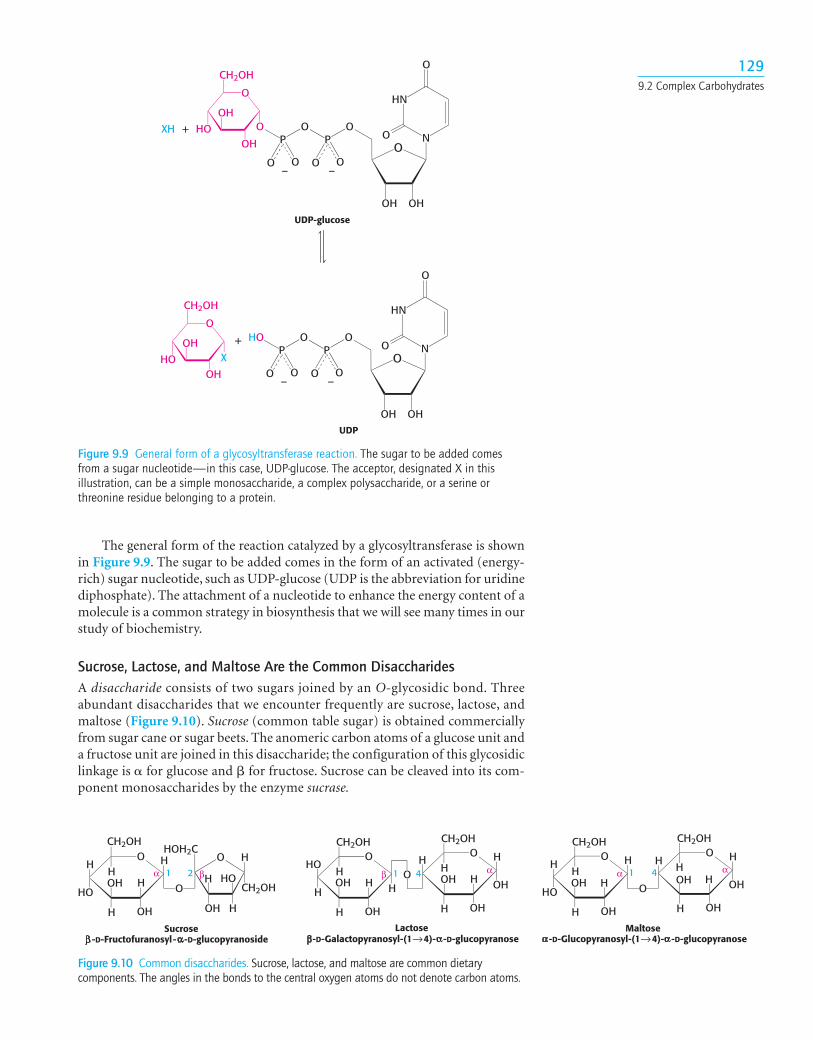
The general form of the reaction catalyzed by a glycosyltransferase is shownin Figure 9.9. The sugar to be added comes in the form of an activated (energy-rich) sugar nucleotide, such as UDP-glucose (UDP is the abbreviation for uridinediphosphate). The attachment of a nucleotide to enhance the energy content of amolecule is a common strategy in biosynthesis that we will see many times in ourstudy of biochemistry.
Sucrose, Lactose, and Maltose Are the Common DisaccharidesA disaccharide consists of two sugars joined by an O-glycosidic bond. Threeabundant disaccharides that we encounter frequently are sucrose, lactose, andmaltose (Figure 9.10). Sucrose (common table sugar) is obtained commerciallyfrom sugar cane or sugar beets. The anomeric carbon atoms of a glucose unit anda fructose unit are joined in this disaccharide; the configuration of this glycosidiclinkage is � for glucose and � for fructose. Sucrose can be cleaved into its com-ponent monosaccharides by the enzyme sucrase.
O
O
CH2OH
OH
OH
HOP
O
O O
P
O O
O
O
OHOH
HN
N
N
O
O
– –
+XH
OHO
CH2OH
OH
OH
HOP
O
O O
P
O O
O
O
OHOH
HN
O
O
– –
X
+
UDP-glucose
UDP
Figure 9.9 General form of a glycosyltransferase reaction. The sugar to be added comesfrom a sugar nucleotide—in this case, UDP-glucose. The acceptor, designated X in thisillustration, can be a simple monosaccharide, a complex polysaccharide, or a serine orthreonine residue belonging to a protein.
OH
H
HOOH
H
H
OH
H
O
O
HO
H
H
OH
H
CH2OH
Sucrose-D-Fructofuranosyl- -D-glucopyranoside
HOH2C
1 2α β
CH2OHO
CH2OH
HHO
HOH
H
H
OH
H
OCH2OH
HH
OH
H
H
OH
H
OHO
Lactose-D-Galactopyranosyl-(1→4)- -D-glucopyranose
β α1 4
OCH2OH
HH
HOOH
H
H
OH
H
O
OCH2OH
HH
OH
H
H
OH
H
OH
Maltose-D-Glucopyranosyl-(1→4)- -D-glucopyranose
α α1 4
Figure 9.10 Common disaccharides. Sucrose, lactose, and maltose are common dietarycomponents. The angles in the bonds to the central oxygen atoms do not denote carbon atoms.
1299.2 Complex Carbohydrates

Figure 9.11 Electron micrograph of amicrovillus. Lactase and other enzymesthat hydrolyze carbohydrates are presenton microvilli that project from the outerface of the plasma membrane of intestinalepithelial cells. [From M. S. Mooseker and L. G. Tilney, J. Cell. Biol. 67:725–743, 1975.]
Lactose, the disaccharide of milk, consists of galactose joined to glucose by a�-1,4-glycosidic linkage. Lactose is hydrolyzed to these monosaccharides bylactase in human beings and by b-galactosidase in bacteria. In maltose, two glu-cose units are joined by an �-1,4-glycosidic linkage. Maltose comes from thehydrolysis of large polymeric oligosaccharides such as starch and glycogen and isin turn hydrolyzed to glucose by maltase. Sucrase, lactase, and maltase are locatedon the outer surfaces of epithelial cells lining the small intestine (Figure 9.11). The
cleavage products of sucrose, lactose, and maltose can be furtherprocessed to provide energy in the form of ATP.
Glycogen and Starch Are Storage Forms of GlucoseLarge polymeric oligosaccharides, formed by the linkage of multiple monosaccha-rides, are called polysaccharides and play vital roles in energy storage and in main-taining the structural integrity of an organism. The variety of differentmonosaccharides can be put together in any number of arrangements, creating ahuge array of possible polysaccharides. If all of the monosaccharide units in apolysaccharide are the same, the polymer is called a homopolymer. The most com-mon homopolymer in animal cells is glycogen, the storage form of glucose. Glyco-gen is present in most of our tissues but is most common in muscle and liver. Aswill be considered in detail in Chapters 23 and 24, glycogen is a large, branchedpolymer of glucose residues. Most of the glucose units in glycogen are linked by�-1,4-glycosidic bonds. The branches are formed by �-1,6-glycosidic bonds, pre-sent about once in 10 units (Figure 9.12).
The nutritional reservoir in plants is the homopolymer starch, of whichthere are two forms. Amylose, the unbranched type of starch, consists of glu-cose residues in �-1,4 linkage. Amylopectin, the branched form, has about 1 �-1,6 linkage per 30 �-1,4 linkages, in similar fashion to glycogen except for itslower degree of branching. More than half the carbohydrate ingested by humanbeings is starch, found in wheat, potatoes, and rice, to name just a few sources(Figure 9.13). Amylopectin, amylose, and glycogen are rapidly hydrolyzed bya-amylase, an enzyme secreted by the salivary glands and the pancreas.
Cellulose, a Structural Component of Plants, Is Made of Chains of GlucoseCellulose, the other major polysaccharide of glucose found in plants, serves astructural rather than a nutritional role as an important component of the plantcell wall. Cellulose is among the most abundant organic compounds in the biosphere.Some 1015 kg of cellulose is synthesized and degraded on Earth each year, anamount 1000 times as great as the combined weight of the human race. Cellu-lose is an unbranched polymer of glucose residues joined by �-1,4 linkages, incontrast with the �-1,4 linkage seen in starch and glycogen. This simple differ-ence in stereochemistry yields two molecules with vastly different propertiesand biological functions. The � configuration allows cellulose to form very long,straight chains. Fibrils are formed by parallel chains that interact with one anotherthrough hydrogen bonds, generating a rigid, supportive structure. The straight
OCH2OH
HH
OOH
H
H
OH
H
O
OCH2
HH
OH
H
H
OH
H
Oα α1 4
OCH2OH
HH
OOH
H
H
OH
H
O
α 1
6
α-1,6-Glycosidic bond
Figure 9.12 Branch point in glycogen.Two chains of glucose molecules joined by�-1,4-glycosidic bonds are linked by an �-1,6-glycosidic bond to create a branchpoint. Such an �-1,6-glycosidic bond formsat approximately every 10 glucose units,making glycogen a highly branchedmolecule.
Figure 9.13 Pasta is a common source ofstarch. Pasta is made with starch, as flour,from Durham wheat. [Superstock.]
130
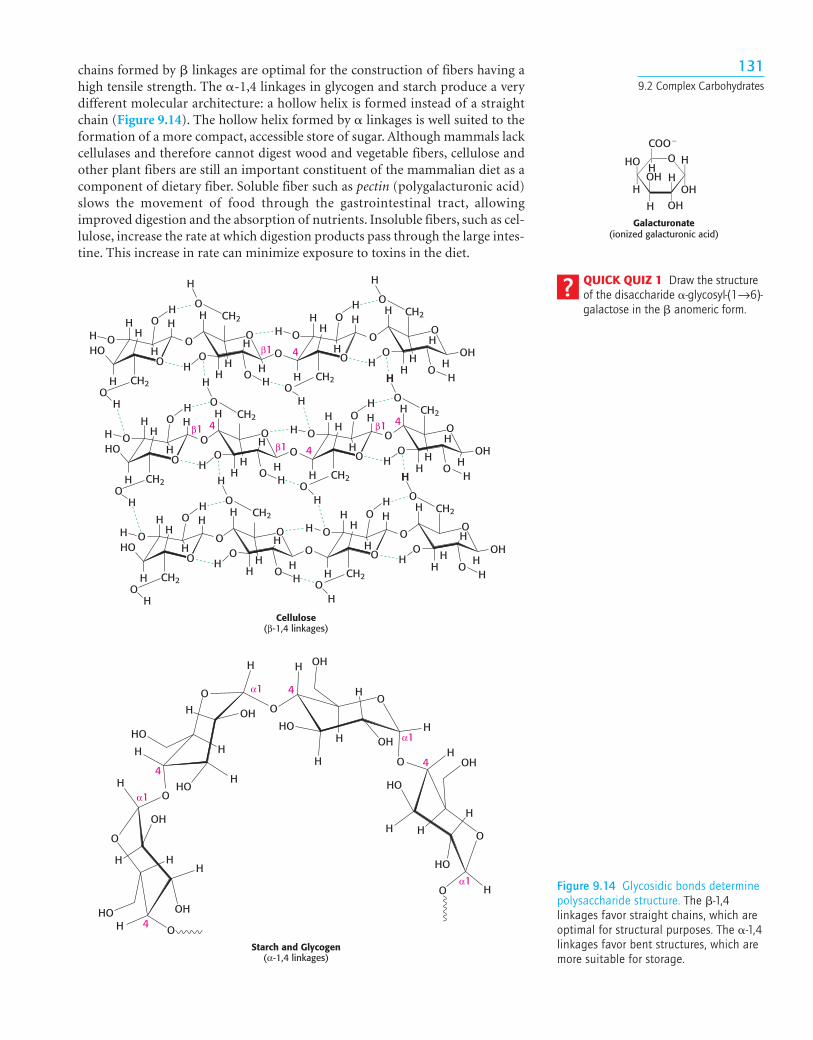
1319.2 Complex Carbohydrates
Galacturonate(ionized galacturonic acid)
HHO
H
O
H
OHOH
OH
H
H
COO�
QUICK QUIZ 1 Draw the structureof the disaccharide �-glycosyl-(1→6)-galactose in the � anomeric form.
Cellulose(β-1,4 linkages)
β1 4
4
44
HCH2
CH2CH2
CH2
OH
H
HH
HH
H
H
H
HH
H
H
HH
H
H
H
HH
HH
H
H
H
HH
HH
HH
HHO OH
O
OO
O
O
O
OO
O
O
O
O
OO
O O
O O
HCH2
CH2CH2
CH2
OH
H
HH
HH
H
H
H
HH
H
H
HH
H H
H
HH
HH
H
H
H
HH
HH
HH
HHO OH
O
OO
O
O
O
OO
O
O
O
O
OO
O O
O O
HCH2
CH2CH2
CH2
OH
H
HH
HH
H
H
H
HH
H
H
HH
H H
HH
HH
H
H
H
HH
HH
HHHO OH
O
OO
O
O
O
OO
O
O
O
O
OO
O O
O O
H
β1
β1
β1
Starch and Glycogen(α-1,4 linkages)
OH
OH
OH
OH
OH
OH
HO
HO
HOHO
HO
HO
H
H
O
O
O
OO
O
O
H
H
H
H H
HH
H
H
H
HH
H H
H
H
H
H
O
O
α1
α1
α1
α1
4
44
4
Figure 9.14 Glycosidic bonds determinepolysaccharide structure. The �-1,4linkages favor straight chains, which areoptimal for structural purposes. The �-1,4linkages favor bent structures, which aremore suitable for storage.
chains formed by � linkages are optimal for the construction of fibers having ahigh tensile strength. The �-1,4 linkages in glycogen and starch produce a verydifferent molecular architecture: a hollow helix is formed instead of a straightchain (Figure 9.14). The hollow helix formed by � linkages is well suited to theformation of a more compact, accessible store of sugar. Although mammals lackcellulases and therefore cannot digest wood and vegetable fibers, cellulose andother plant fibers are still an important constituent of the mammalian diet as acomponent of dietary fiber. Soluble fiber such as pectin (polygalacturonic acid)slows the movement of food through the gastrointestinal tract, allowingimproved digestion and the absorption of nutrients. Insoluble fibers, such as cel-lulose, increase the rate at which digestion products pass through the large intes-tine. This increase in rate can minimize exposure to toxins in the diet.
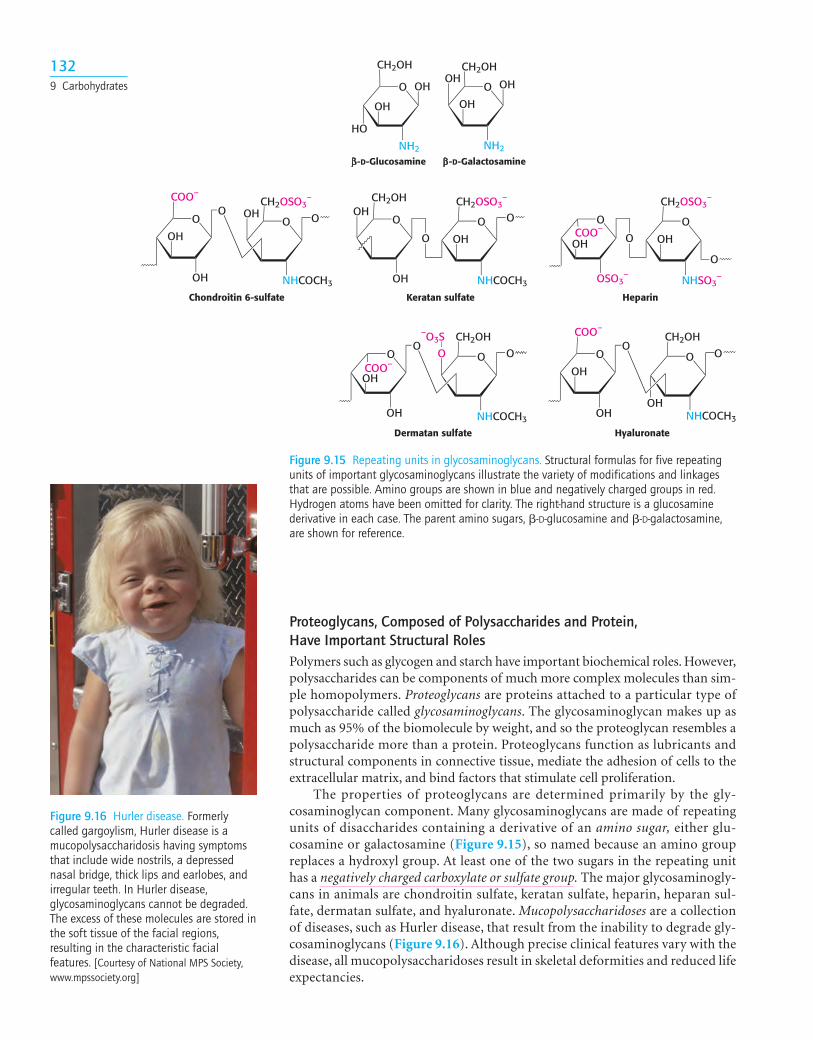
Chondroitin 6-sulfate Keratan sulfate Heparin
Dermatan sulfate Hyaluronate
O
COO–
OH
OHO
CH2OH
NHCOCH3
OO
OH
O
OH
OH
O
CH2OH
NHCOCH3
OOO
COO–
–O3S
CH2OSO3–
NHSO3–
O
OSO3–
OH
O
OHO
O
COO–O
COO–
OH
OHO
CH2OSO3–
NHCOCH3
OOHOCH2OSO3
–
NHCOCH3
O
O O
CH2OH
OH
O
OH
O
O
OH
CH2OH
NH2
CH2OH
NH2
OH
OH
OH
OH
HO
OH
-D-Glucosamine -D-Galactosamine
Figure 9.15 Repeating units in glycosaminoglycans. Structural formulas for five repeatingunits of important glycosaminoglycans illustrate the variety of modifications and linkagesthat are possible. Amino groups are shown in blue and negatively charged groups in red.Hydrogen atoms have been omitted for clarity. The right-hand structure is a glucosaminederivative in each case. The parent amino sugars, �-D-glucosamine and �-D-galactosamine,are shown for reference.
Figure 9.16 Hurler disease. Formerlycalled gargoylism, Hurler disease is amucopolysaccharidosis having symptomsthat include wide nostrils, a depressednasal bridge, thick lips and earlobes, andirregular teeth. In Hurler disease,glycosaminoglycans cannot be degraded.The excess of these molecules are stored inthe soft tissue of the facial regions,resulting in the characteristic facialfeatures. [Courtesy of National MPS Society,www.mpssociety.org]
Proteoglycans, Composed of Polysaccharides and Protein, Have Important Structural RolesPolymers such as glycogen and starch have important biochemical roles. However,polysaccharides can be components of much more complex molecules than sim-ple homopolymers. Proteoglycans are proteins attached to a particular type ofpolysaccharide called glycosaminoglycans. The glycosaminoglycan makes up asmuch as 95% of the biomolecule by weight, and so the proteoglycan resembles apolysaccharide more than a protein. Proteoglycans function as lubricants andstructural components in connective tissue, mediate the adhesion of cells to theextracellular matrix, and bind factors that stimulate cell proliferation.
The properties of proteoglycans are determined primarily by the gly-cosaminoglycan component. Many glycosaminoglycans are made of repeatingunits of disaccharides containing a derivative of an amino sugar, either glu-cosamine or galactosamine (Figure 9.15), so named because an amino groupreplaces a hydroxyl group. At least one of the two sugars in the repeating unithas a negatively charged carboxylate or sulfate group. The major glycosaminogly-cans in animals are chondroitin sulfate, keratan sulfate, heparin, heparan sul-fate, dermatan sulfate, and hyaluronate. Mucopolysaccharidoses are a collectionof diseases, such as Hurler disease, that result from the inability to degrade gly-cosaminoglycans (Figure 9.16). Although precise clinical features vary with thedisease, all mucopolysaccharidoses result in skeletal deformities and reduced lifeexpectancies.
1329 Carbohydrates

Clinical Insight
Proteoglycans Are Important Components of CartilageThe proteoglycan aggrecan and the protein collagen are key components of carti-lage. The triple helix of collagen (p. 51) provides structure and tensile strength,whereas aggrecan serves as a shock absorber. The protein component of aggre-can is a large molecule composed of 2397 amino acids (Figure 9.17). Manyaggrecans are linked together by hyaluronan, a glycosaminoglycan. This combi-nation of glycosaminoglycan and protein is especially suited to function as ashock absorber. Water is absorbed on the glycosaminoglycans, attracted by themany negative charges. Aggrecan can cushion compressive forces because theabsorbed water enables the proteoglycan to spring back after having beendeformed. When pressure is exerted, such as when the foot hits the ground inwalking, water is squeezed from the glycosaminoglycan, cushioning the impact.When the pressure is released, the water rebinds. Osteoarthritis can result from theproteolytic degradation of aggrecan and collagen in the cartilage.
Although glycosaminoglycans may not seem to be familiar molecules, theyare common throughout the biosphere. Chitin is a glycosaminoglycan found inthe exoskeleton of insects, crustaceans, and arachnids and is, next to cellulose, thesecond most abundant polysaccharide in nature (Figure 9.18). ■
9.3 Carbohydrates Are Attached to Proteins to Form Glycoproteins
Carbohydrates can be covalently linked to proteins to form glycoproteins.Carbohydrates are a much smaller percentage of the weight of glycoproteins than thatof proteoglycans.Whereas proteoglycans serve mainly structural roles, glycoproteinsare more versatile. Many glycoproteins are components of cell membranes, wherethey play a variety of roles. The carbohydrate parts of membrane glycoproteins areon the outer surface of the cell membrane. Many secretory proteins are glycoproteins.
300 nm
HyaluronanKeratan sulfate
Aggrecan
Chondroitin sulfate
G3
G2G1 G3
G2
G1
G3 G2 G1
G3
G3
G2 G1
G2
G1
(A) (B)
Figure 9.17 The structure of proteoglycan from cartilage. (A) Electron micrograph of aproteoglycan from cartilage (with color added). Proteoglycan monomers emerge laterally at regularintervals from opposite sides of a central filament of hyaluronan. (B) Schematic representation inwhich G stands for globular domain. [(A) Courtesy of Dr. Lawrence Rosenberg. From J. A. Buckwalter andL. Rosenberg. Collagen Relat. Res. 3:489–504, 1983.]
Figure 9.18 Chitin, a glycosaminoglycan,is present in insect wings and theexoskeleton. Glycosaminoglycans arecomponents of the exoskeletons of insects,crustaceans, and arachnids. [FLPA/Alamy.]
1339.3 Glycoproteins
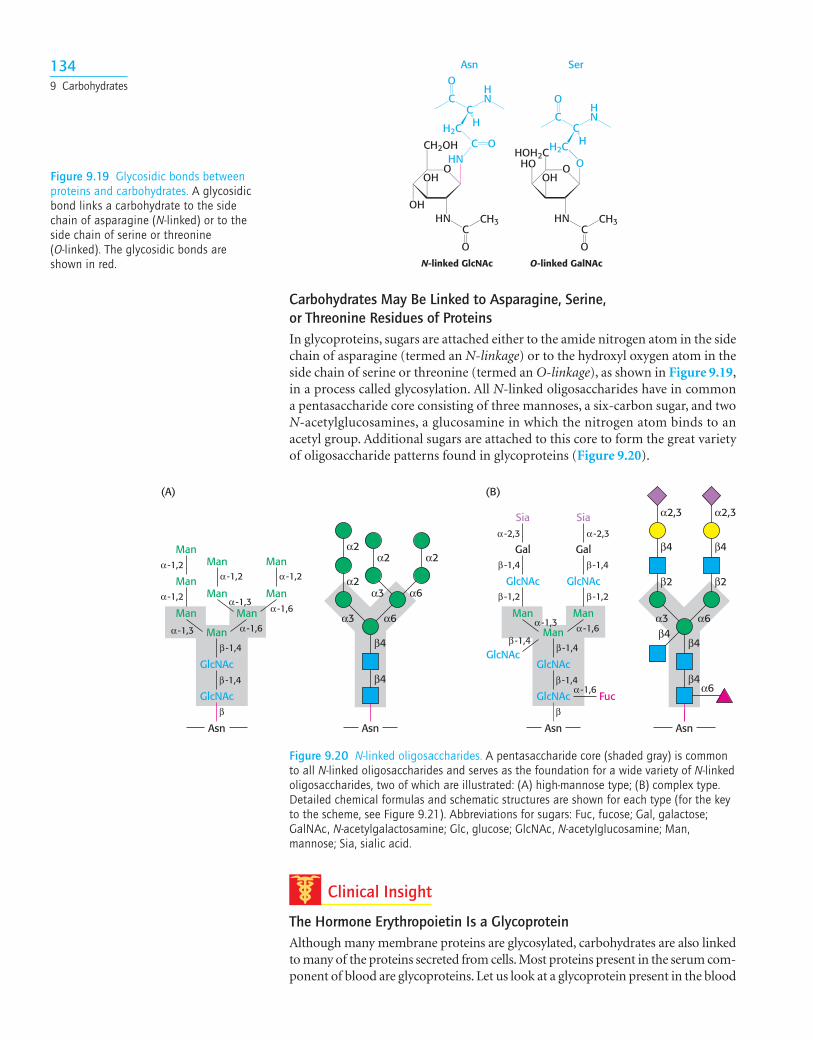
Carbohydrates May Be Linked to Asparagine, Serine, or Threonine Residues of ProteinsIn glycoproteins, sugars are attached either to the amide nitrogen atom in the sidechain of asparagine (termed an N-linkage) or to the hydroxyl oxygen atom in theside chain of serine or threonine (termed an O-linkage), as shown in Figure 9.19,in a process called glycosylation. All N-linked oligosaccharides have in commona pentasaccharide core consisting of three mannoses, a six-carbon sugar, and twoN-acetylglucosamines, a glucosamine in which the nitrogen atom binds to anacetyl group. Additional sugars are attached to this core to form the great varietyof oligosaccharide patterns found in glycoproteins (Figure 9.20).
α6
β4
β4
α6α3
α3
α2α2α2
α2
β4α6
β4β4
α6α3
α2,3
β2
α2,3
β2
β4 β4
Asn Asn Asn
GlcNAc
GlcNAc
Man
Man Man
Man
Man
Man
Man
β
β -1,4
β -1,4
α -1,3 α -1,6
α -1,2 Man
Manα -1,2
α -1,3α -1,6
α -1,2 α -1,2
(A)
Gal
Fuc
GlcNAc
GlcNAc
Sia
Asn
GlcNAc
GlcNAc
Man
Man Man
β
β -1,4
β -1,4
α -1,6
α -1,3 α -1,6
β -1,2
α -2,3
β -1,4
Gal
GlcNAc
Siaα -2,3
β -1,2
β -1,4
β -1,4
(B)
Figure 9.20 N-linked oligosaccharides. A pentasaccharide core (shaded gray) is commonto all N-linked oligosaccharides and serves as the foundation for a wide variety of N-linkedoligosaccharides, two of which are illustrated: (A) high-mannose type; (B) complex type.Detailed chemical formulas and schematic structures are shown for each type (for the keyto the scheme, see Figure 9.21). Abbreviations for sugars: Fuc, fucose; Gal, galactose;GalNAc, N-acetylgalactosamine; Glc, glucose; GlcNAc, N-acetylglucosamine; Man,mannose; Sia, sialic acid.
N-linked GlcNAc O-linked GalNAc
C
O O
H2CC
H
HN
HNC
O
OH OH
OH
CH2OHHOH2CHN
O
C
O
C
H2C H
HNC
O
O
CH3 HNC
O
CH3
HO
Asn Ser
Figure 9.19 Glycosidic bonds betweenproteins and carbohydrates. A glycosidicbond links a carbohydrate to the sidechain of asparagine (N-linked) or to theside chain of serine or threonine (O-linked). The glycosidic bonds areshown in red.
Clinical Insight
The Hormone Erythropoietin Is a GlycoproteinAlthough many membrane proteins are glycosylated, carbohydrates are also linkedto many of the proteins secreted from cells. Most proteins present in the serum com-ponent of blood are glycoproteins. Let us look at a glycoprotein present in the blood
1349 Carbohydrates

serum that has dramatically improved treatment for anemia, particularly thatinduced by cancer chemotherapy. The glycoprotein hormone erythropoietin (EPO)is secreted by the kidneys and stimulates the production of red blood cells. EPO iscomposed of 165 amino acids and is N-glycosylated at three asparagine residues andO-glycosylated on a serine residue (Figure 9.21), making the mature EPO 40%carbohydrate by weight. Glycosylation enhances the stability of the protein in theblood; unglycosylated protein has only about 10% of the bioactivity of the glycosy-lated form because the protein is rapidly removed from the blood by the kidney. Theavailability of recombinant human EPO has greatly aided the treatment of anemias.Some endurance athletes have used recombinant human EPO to increase thered-blood-cell count and hence their oxygen-carrying capacity, a practice prohib-ited by most professional sports organizations. One means of detecting the presenceof prohibited human recombinant EPO in athletes is based on the detection ofdifferences in the glycosylation patterns of recombinant and natural EPO. ■
Biological Insight
Blood Groups Are Based on Protein Glycosylation PatternsThe human ABO blood groups illustrate the effects of glycosyltransferases.Each blood group is designated by the presence of one of the three differentcarbohydrates, termed A, B, or O, on the surfaces of red blood cells (Figure 9.22).These structures have in common an oligosaccharide foundation called the O (orsometimes H) antigen. The A and B antigens differ from the O antigen by theaddition of one extra monosaccharide, either N-acetylgalactosamine (for A) orgalactose (for B) through an �-1,3 linkage to a galactose moiety of the O antigen.
Specific glycosyltransferases add the extra monosaccharide to the O antigen.Each person inherits the gene for one glycosyltransferase of this type from each par-ent. The type A transferase specifically adds N-acetylgalactosamine, whereas the typeB transferase adds galactose. These enzymes are identical in all but 4 of 354 positions.The O phenotype is the result of a mutation that leads to premature termination oftranslation and, hence, to the production of no active glycosyltransferase.
Asn 83
Asn 38
Asn 24
Ser126Key
Galactose
N-Acetylgalactosamine
N-Acetylglucosamine
Mannose
N-Acetylmannosamine
Fucose
Sialic acid
Figure 9.21 Oligosaccharidesattached to erythropoietin.
Erythropoietin has oligosaccharides linkedto three asparagine residues and oneserine residue. The structures shown areapproximately to scale. The carbohydratestructures represented in the amino acidresidues are depicted symbolically byemploying a scheme (shown in the key,which also applies to Figures 9.20 and9.22) that is becoming widely used. [Drawnfrom 1BUY.pdf.]
1359.3 Glycoproteins

These structures have important implications for blood transfusions andother transplantation procedures. If an antigen not normally present in a personis introduced, the person’ immune system recognizes it as foreign. Adversereactions can ensue, initiated by the intravascular destruction of the incompatiblered blood cells.
Why are different blood types present in the human population? Suppose thata pathogenic organism such as a parasite expresses on its cell surface a carbohy-drate antigen similar to one of the blood-group antigens. This antigen may not bereadily detected as foreign in a person with the blood type that matches the par-asite antigen, and the parasite will flourish. However, other people with differentblood types will be protected. Hence, there will be selective pressure on humanbeings to vary blood type to prevent parasitic mimicry and a corresponding selec-tive pressure on parasites to enhance mimicry. The constant “arms race” betweenpathogenic microorganisms and human beings drives the evolution of diversityof surface antigens within the human population. ■
Clinical Insight
Lack of Glycosylation Can Result in Pathological ConditionsAlthough the role of carbohydrate attachment to proteins is not known indetail in most cases, data indicate that this glycosylation is important for theprocessing and stability of these proteins, as it is for EPO. For instance, certaintypes of muscular dystrophy can be traced to improper glycosylation of mem-brane proteins. Indeed, an entire family of severe inherited human diseasecalled congenital disorders of glycosylation has been identified. These patholog-ical conditions reveal the importance of proper modification of proteins bycarbohydrates and their derivatives.
An especially clear example of the role of glycosylation is provided by I-celldisease (also called mucolipidosis II), a lysosomal storage disease. Normally, acarbohydrate marker directs certain digestive enzymes from the Golgi complexto lysosomes where they normally function. Lysosomes are organelles thatdegrade and recycle damaged cellular components or material brought into thecell by endocytosis. In patients with I-cell disease, lysosomes contain large inclu-sions of undigested glycosaminoglycans and glycolipids (p. 149) the “I” in thename of the disease. These inclusions are present because the enzymes normallyresponsible for the degradation of glycosaminoglycans are missing from
GalNAc
α3
β3
β3
α2
β3
β3
α2α3
β3
β3
α2
Fucα -1,2 Gal
Gal
GlcNAcβ -1,3
β -1,3
Fucα -1,2 Gal
Gal
GlcNAcβ -1,3
β -1,3
α -1,3
GalFucα -1,2 Gal
Gal
GlcNAcβ -1,3
β -1,3
α -1,3
A antigenO antigen B antigen
Figure 9.22 Structures of A, B, and Ooligosaccharide antigens. Thecarbohydrate structures represented inthe upper part of this illustration aredepicted symbolically (refer to the key in Figure 9.21).
1369 Carbohydrates
QUICK QUIZ 2 Which amino acidsare used for the attachment of
carbohydrates to proteins?

affected lysosomes. Remarkably, the enzymes are present at very high levels inthe blood and urine. Thus, active enzymes are synthesized, but, in the absenceof appropriate glycosylation, they are exported instead of being sequestered inlysosomes. In other words, in I-cell disease a whole series of enzymes is incorrectlyaddressed and delivered to the wrong location. Normally, these enzymes containa mannose 6-phosphate residue, but, in I-cell disease, the attached mannoselacks a phosphate (Figure 9.23). Mannose 6-phosphate is in fact the marker thatnormally directs many hydrolytic enzymes from the Golgi complex to lyso-somes. I-cell patients are deficient in the phosphotransferase catalyzing the firststep in the addition of the phosphoryl group; the consequence is the mistarget-ing of eight essential enzymes. I-cell disease causes the patient to suffer severepsychomotor retardation and skeletal deformities, similar to those in Hurlerdisease (p. 132). ■
9.4 Lectins Are Specific Carbohydrate-Binding ProteinsGlycoproteins contain multiple types of monosaccharides joined by differentkinds of glycosidic linkages. An enormous number of patterns in the compositionand structure of sugars composing the carbohydrate units of glyoproteins are pos-sible.The diversity and complexity of the carbohydrate units suggest that they arefunctionally important. Nature does not construct complex patterns when sim-ple ones suffice. Why all this intricacy and diversity? It is becoming evident thatcarbohydrates are information-rich molecules that guide many biologicalprocesses. The diverse carbohydrate structures displayed on cell surfaces are wellsuited to serving as sites of interaction between cells and their environments. Thecarbohydrate structures are the keys to protein locks. Such proteins, termed lectins(from Latin legere, “to select”), are the partners that bind specific carbohydratestructures on neighboring cell surfaces. Lectins are ubiquitous: they are found inanimals, plants, and microorganisms.
Lectins Promote Interactions Between CellsCell–cell contact is a vital interaction in a host of biochemical functions, frombuilding a tissue from isolated cells to facilitating the transmission of informa-tion. The chief function of lectins, carbohydrate-binding proteins, is to facilitatecell–cell contact. A lectin usually contains two or more binding sites for carbohy-drate units. These carbohydrate-binding sites on the surface of one cell interactwith arrays of carbohydrates displayed on the surface of another cell. Lectins andcarbohydrates are linked by a number of weak noncovalent interactions thatensure specificity yet permit unlinking as needed.
Clinical Insight
Lectins Facilitate Embryonic DevelopmentOne class of lectins are proteins termed selectins, which bind immune-systemcells to sites of injury in the inflammatory response (Figure 9.24). The L, E, andP forms of selectins bind specifically to carbohydrates on lymph-node vessels,endothelium, or activated blood platelets, respectively. L-Selectin, originallythought to participate only in the immune response, is produced by an embryowhen it is ready to attach to the endometrium of the mother’ uterus. For a shortperiod of time, the endometrial cells present an oligosaccharide on the cellsurface. When the embryo attaches through lectins, the attachment activatessignal pathways in the endometrium to make implantation of the embryopossible. ■
137
O
OR
CH2
HOOH
HO
HO
Mannose residue
UDP-GlcNAc
UMP
Phospho-transferase
O
OR
CH2
HOOH
HO
OP
O
OO
GlcNAc
–
O
OR
CH2
HOOH
HO
OP
O
OO
2–
Mannose 6-phosphate residue
H2O
GlcNAc
Phospho-diesterase
Figure 9.23 The formation of a mannose6-phosphate marker. A glycoproteindestined for delivery to lysosomes acquiresa phosphate marker in the Golgicompartment in a two-step process. First, aphosphotransferase adds a phospho-N-acetylglucosamine unit to the 6-OH groupof a mannose residue; then, aphosphodiesterase removes the addedsugar to generate a mannose 6-phosphateresidue in the core oligosaccharide.
Figure 9.24 Selectins mediate cell–cellinteractions. The scanning electronmicrograph shows lymphocytes adheringto the endothelial lining of a lymph node.The L selectins on the lymphocyte surfacebind specifically to carbohydrates on thelining of the lymph-node vessels. [Courtesyof Dr. Eugene Butcher.]
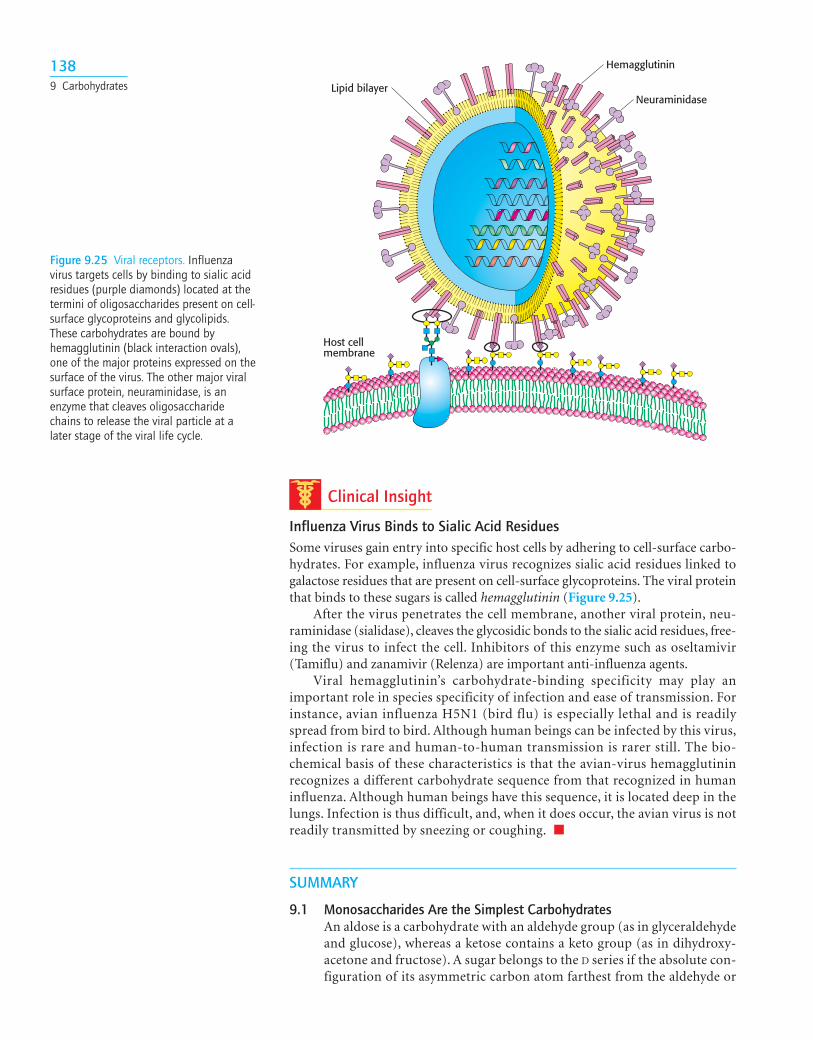
Clinical Insight
Influenza Virus Binds to Sialic Acid ResiduesSome viruses gain entry into specific host cells by adhering to cell-surface carbo-hydrates. For example, influenza virus recognizes sialic acid residues linked togalactose residues that are present on cell-surface glycoproteins. The viral proteinthat binds to these sugars is called hemagglutinin (Figure 9.25).
After the virus penetrates the cell membrane, another viral protein, neu-raminidase (sialidase), cleaves the glycosidic bonds to the sialic acid residues, free-ing the virus to infect the cell. Inhibitors of this enzyme such as oseltamivir(Tamiflu) and zanamivir (Relenza) are important anti-influenza agents.
Viral hemagglutinin’s carbohydrate-binding specificity may play animportant role in species specificity of infection and ease of transmission. Forinstance, avian influenza H5N1 (bird flu) is especially lethal and is readilyspread from bird to bird. Although human beings can be infected by this virus,infection is rare and human-to-human transmission is rarer still. The bio-chemical basis of these characteristics is that the avian-virus hemagglutininrecognizes a different carbohydrate sequence from that recognized in humaninfluenza. Although human beings have this sequence, it is located deep in thelungs. Infection is thus difficult, and, when it does occur, the avian virus is notreadily transmitted by sneezing or coughing. ■
SUMMARY
9.1 Monosaccharides Are the Simplest CarbohydratesAn aldose is a carbohydrate with an aldehyde group (as in glyceraldehydeand glucose), whereas a ketose contains a keto group (as in dihydroxy-acetone and fructose). A sugar belongs to the D series if the absolute con-figuration of its asymmetric carbon atom farthest from the aldehyde or
1389 Carbohydrates Lipid bilayer
Hemagglutinin
Neuraminidase
Host cellmembrane
Figure 9.25 Viral receptors. Influenzavirus targets cells by binding to sialic acidresidues (purple diamonds) located at thetermini of oligosaccharides present on cell-surface glycoproteins and glycolipids.These carbohydrates are bound byhemagglutinin (black interaction ovals),one of the major proteins expressed on thesurface of the virus. The other major viralsurface protein, neuraminidase, is anenzyme that cleaves oligosaccharidechains to release the viral particle at alater stage of the viral life cycle.

keto group is the same as that of D-glyceraldehyde. Most naturally occur-ring sugars belong to the D series. The C-1 aldehyde in the open-chainform of glucose reacts with the C-5 hydroxyl group to form a six-memberedpyranose ring. The C-2 keto group in the open-chain form of fructosereacts with the C-5 hydroxyl group to form a five-membered furanosering. Pentoses such as ribose and deoxyribose also form furanose rings.An additional asymmetric center is formed at the anomeric carbon atom(C-1 in aldoses and C-2 in ketoses) in these cyclizations. Sugars arejoined to alcohols and amines by glycosidic bonds at the anomeric carbon atom.
9.2 Monosaccharides Are Linked to Form Complex CarbohydratesSugars are linked to one another in disaccharides and polysaccharides byO-glycosidic bonds. Sucrose, lactose, and maltose are the common disac-charides. Sucrose (common table sugar) consists of �-glucose and �-fructosejoined by a glycosidic linkage between their anomeric carbon atoms.Lactose (in milk) consists of galactose joined to glucose by a �-1,4 link-age. Maltose (in starch) consists of two glucoses joined by an �-1,4 link-age. Starch is a polymeric form of glucose in plants, and glycogen servesa similar role in animals. Most of the glucose units in starch and glycogenare in �-1,4 linkage. Glycogen has more branch points formed by �-1,6linkages than does starch, and so glycogen is more soluble. Cellulose, themajor structural polymer of plant cell walls, consists of glucose unitsjoined by �-1,4 linkages. These � linkages give rise to long straight chainsthat form fibrils with high tensile strength. In contrast, the � linkages instarch and glycogen lead to open helices, in keeping with their roles asmobile energy stores. Cell surfaces and the extracellular matrices of ani-mals contain polymers of repeating disaccharides called glycosaminogly-cans. Proteoglycans consist largely of a glycosaminoglycan covalentlylinked to protein.
9.3 Carbohydrates Are Attached to Proteins to Form GlycoproteinsSpecific enzymes link the oligosaccharide units on proteins either to theside-chain hydroxyl oxygen atom of a serine or threonine residue or to theside-chain amide nitrogen atom of an asparagine residue.
9.4 Lectins Are Specific Carbohydrate-Binding ProteinsCarbohydrates on cell surfaces are recognized by proteins called lectins.In animals, the interplay of lectins and their sugar targets guides cell–cellcontact. The viral protein hemagglutinin on the surface of the influenzavirus recognizes sialic acid residues on the surfaces of cells invaded bythe virus.
139Key Terms
Key Terms
monosaccharide (p. 122)ketose (p. 123)aldose (p. 123)triose (p. 123)tetrose (p. 123)pentose (p. 123)hexose (p. 123)heptose (p. 123)enantiomer (p. 123)diastereoisomer (p. 124)epimer (p. 124)
hemiacetal (p. 125)pyranose (p. 125)hemiketal (p. 126)furanose (p. 126)anomer (p. 126)reducing sugar (p. 127)nonreducing sugar (p. 127)glycosidic bond (p. 128)oligosaccharide (p. 128)glycosyltransferase (p. 129)disaccharide (p. 129)
polysaccharide (p. 130)glycogen (p. 130)starch (p. 130)cellulose (p. 130)proteoglycan (p. 132)glycosaminoglycan (p. 132)glycoprotein (p. 133)lysosome (p. 136)lectin (p. 137)selectin (p. 137)

140
Answers to QUICK QUIZZES
1. 2. Asparagine, serine, and threonine.O
O
HCH2OH
CH2H
OH
OH
H
H
OH
H
O
H
OH
OH
H
H H
OH OHH
�-Glucosyl-(1S 6)-galactose
1. Word origin. Account for the origin of the term carbohydrate.
2. Diversity. How many different oligosaccharides can bemade by linking one glucose, one mannose, and one galac-tose residue? Assume that each sugar is in its pyranose form.Compare this number with the number of tripeptides thatcan be made from three different amino acids.
3. Couples. Indicate whether each of the following pairs ofsugars consists of anomers, epimers, or an aldose–ketose pair:
(a) D-glyceraldehyde and dihydroxyacetone(b) D-glucose and D-mannose(c) D-glucose and D-fructose(d) �-D-glucose and �-D-glucose(e) D-ribose and D-ribulose(f) D-galactose and D-glucose
4. Mutarotation. The specific rotations of the � and �anomers of D-glucose are �112 degrees and �18.7 degrees,respectively. Specific rotation, [�]D, is defined as theobserved rotation of light of wavelength 589 nm (the D lineof a sodium lamp) passing through 10 cm of a 1 g ml�1
solution of a sample. When a crystalline sample of �-D-glucopyranose is dissolved in water, the specific rotationdecreases from 112 degrees to an equilibrium value of 52.7degrees. On the basis of this result, what are the proportionsof the � and � anomers at equilibrium? Assume that theconcentration of the open-chain form is negligible.
5. Telltale marker. Glucose reacts slowly with hemoglobinand other proteins to form covalent compounds. Why isglucose reactive? What is the nature of the adduct formed?
6. Periodate cleavage. Compounds containing hydroxylgroups on adjacent carbon atoms undergo carbon–carbonbond cleavage when treated with periodate ion (IO4�). Howcan this reaction be used to distinguish between pyrano-sides and furanosides if cis-glycans are cleaved more rapidlythan trans-glycols?
7. Oxygen source. Does the oxygen atom attached to C-1in methyl �-D-glucopyranoside come from glucose ormethanol?
8. Sugar lineup. Identify the following four sugars.
Problems
OOH
OH
HHH
H
H
HOHO
HOH2C
OH
OH
HOHH
OH
H
HOH
HOH2C
OH
OH
HNH2
H
H
H
HOHO
HOH2C
HO
OH
OH
HH
H
OHOH2C
HOH2C
(a)
(c)
(b)
(d)
9. Cellular glue. A trisaccharide unit of a cell-surfaceglycoprotein is postulated to play a critical role in mediat-ing cell–cell adhesion in a particular tissue. Design a simpleexperiment to test this hypothesis.
10. Mapping the molecule. Each of the hydroxyl groups ofglucose can be methylated with reagents such as dimethyl-sulfate under basic conditions. Explain how exhaustivemethylation followed by the complete digestion of a knownamount of glycogen would enable you to determine thenumber of branch points and reducing ends.
11. Component parts. Raffinose is a trisaccharide and aminor constituent in sugar beets.
O
OHO
HO
HOH2CHO
OH2C
HOHO OH
O
O
HO OHCH2OH
CH2OHRaffinose
(a) Is raffinose a reducing sugar? Explain.
(b) What are the monosaccharides that compose raffinose?
(c) �-Galactosidase is an enzyme that will remove galac-tose residues from an oligosaccharide. What are the prod-ucts of �-galactosidase treatment of raffinose?

Problems 141
12. Anomeric differences. �-D-Mannose is a sweet-tastingsugar. �-D-Mannose, on the other hand, tastes bitter. A puresolution of �-D-mannose loses its sweet taste with time as itis converted into the � anomer. Draw the � anomer andexplain how it is formed from the � anomer.
14. Carbohydrates and proteomics. Suppose that a proteincontains six potential N-linked glycosylation sites. Howmany possible proteins can be generated, depending onwhich of these sites is actually glycosylated? Do not includethe effects of diversity within the carbohydrate added.
15. Many possibilities. Why are polysaccharides consideredinformation-rich molecules?
Chapter Integration Problem
16. Stereospecificity. Sucrose, a major product of photosyn-thesis in green leaves, is synthesized by a battery ofenzymes. The substrates for sucrose synthesis, D-glucoseand D-fructose, are a mixture of � and � anomers as well asacyclic compounds in solution. Nonetheless, sucrose con-sists of �-D-glucose linked by its C-1 atom to the C-2 atomof �-D-fructose. How can the specificity of sucrose beexplained in light of the potential substrates?
O
OH
CH2OH
HOOH
HO
�-D-Mannose
13. A taste of honey. Fructose in its �-D-pyranose formaccounts for the powerful sweetness of honey. The �-D-furanose form, although sweet, is not as sweet as thepyranose form. The furanose form is the more stable form.Draw the two forms and explain why it may not always bewise to cook with honey.
Selected readings for this chapter can be found online at www.whfreeman.com/Tymoczko





























