Embed Size (px)
Citation preview

nucleus
lipid dropletGolgi
RasGAP
Ser338
Tyr341 Ser4
Ser62Raf
Tyr Gab2
PI3K
Ser
Thr SOS
Grb2
Grb2
Ser
Thr
SOSGrb2
Ser
Thr
SOS
Grb2
Ser
Thr
SOSGrb2
Ser
Thr
SOS
GTP
GDP
GDP
Ras
GDP
Ras
GTP
Ras
GTP
Ras
RasGAP
P Tyr Gab2
Tyr580
Tyr542SHP-2
P Tyr Gab2
Tyr580Tyr542SHP-2
P Tyr Gab2
Tyr580Tyr542SHP-2
Tyr544
Tyr559Tyr697
Tyr706Tyr721Tyr807
Tyr921Tyr974
M-CSFR
Tyr706 PP
Tyr706 PP
Tyr701 Ser727 STAT1
PTyr701
P Ser727 STAT1
PTyr701
P Ser727
PTyr701
P Ser727 STAT1
Tyr
Ser312(307:R) IRS
TyrIL-4R
Tyr JAK1
TyrJAK3
Tyr JAK1
TyrIL-4R
Tyr JAK1
TyrIL-4R
Tyr
common
chain
TyrJAK3
Tyr
common
chain
TyrJAK3
Tyr
common
chain
y yy yP
yP
yJ 3P
yP
yJ 3
SOCS1
/JAB
PTyr
Ser312(307:R) IRS
TyrSTAT6
P Tyr STAT6
P TyrP Tyr STAT6
P TyrP Tyr STAT6
PTyr701
P Ser727
PTyr701
P Ser727 STAT1
TyrTyr STAT6
Tyr440IFN R1
Tyr JAK1
Tyr1007JAK2
Tyr JAK1
Tyr440IFN R1
Tyr JAK1
Tyr440IFN R1
SHP-1
SOCS1/JAB
Tyr759
Tyr767Tyr814
Tyr905
Tyr915IL-6R
IL-6R
gp130
Tyr JAK1
TyrTyk2
Tyr JAK1
Tyr759
Tyr767Tyr814
Tyr905
Tyr915IL-6R
Tyr JAK1
Tyr759
Tyr767Tyr814
Tyr905
Tyr915IL-6R
TyrTyk2
Tyr759
Tyr767Tyr814
Tyr905
Tyr915IL-6R
TyrTyk2
Tyr759
Tyr767Tyr814
Tyr905
Tyr915IL-6R
Tyr JAK1 TyrTyk2Tyr JAK1 TyrTyk2
SOCS3
SHP-1
Tyr580
Tyr542SHP-2
PTyr580
PTyr542
SHP-2
Tyr STAT3 P Tyr STAT3
P TyrP Tyr STAT3
P TyrP Tyr STAT3
SOCS3
TyrTyk2TyrIL-10R
IL-10RIL-10R
Tyr JAK1
Tyr JAK1
TyrIL-10R
Tyr JAK1
TyrIL-10R
TyrTyk2
TyrIL-10R
TyrTyk2
TyrIL-10R
P PP P
TLR4 MD-2
IKK
TBK-1
P Ser386
P Ser385
IRF-3
Ser
Thr LysIRAK1
IRAK-M
SOCS1
/JAB
P Ser
P Thr Lys
IRAK1
Ubc13
Uev1A
Ubc13
Uev1A
TRIF/
TICAM-1
TBK-1
IKK
Ser386
Ser385 IRF-3
IFN-
CAPK
IKK
Ser176 Ser181 IKK
IKK
IKK
Ser176 Ser181 IKK
IKK
IKK
P Ser176
P
Ser181 IKK
IKK
IKK
P Ser176
P
Ser181 IKK
IKK
CAPK
SCF TrCP
P Thr183
PTyr185
ERK1
ERK2
P Ser276 Ser529
NF- B
p65+p50
P Ser276
P Ser529
NF- B
p65+p50
Ser276 Ser529
NF- B
p65+p50
Ser32
Ser36
Lys21
Lys22I B
P Ser32
P Ser36
UbLys21
UbLys22
I B
IL-10
IL-6
IL-1
p50
I B
TNFTNF
P Thr183
P
Tyr185
P Thr183
P
Tyr185
ERK1
ERK2
Ser338
Tyr341 Ser4
Ser62Raf
Tyr STAT5
GM-CSFR
GM-CSFR
TyrGM-CSFR
TyrJAK1
Tyr1007JAK2
P Tyr STAT5
P TyrP Tyr STAT5
P TyrP Tyr STAT5
Tyr701 Ser727 STAT1
IRF-2 IRF-1
IRF-9
Ser484
Ser485 IRF-7
PTyr701
P Ser727 STAT1
P Tyr STAT2
PTyr701
P Ser727
PTyr701
P Ser727 STAT1
IRF-1IRF-2
IFN-
IRF-1IRF-2
NOSII/iNOS
IRF-7
IRF-9
P Ser484
P Ser485
IRF-7
P Ser484
P Ser485
IRF-7
IFN-
IFN-
pyruvate pyruvate acetyl CoA
NAD+ NADH+H+
pyruvate
carboxylase
pyruvate
dehydrogenase
PDH kinase
pyruvate
carrier
P
P
Ppyruvate
dehydrogenase
PDH kinase
citrate
acyl-CoA
carnitineCoASH
acylcarnitine
fatty acid
malonyl CoA
carnitine acylcarnitine
acyl-CoA
CPT I
CPT II
CACT
CoASHacyl CoA
synthetase
fatty acid
synthetase
acetyl CoA
oxaloacetateCoASH
acetyl CoA
carboxylase
citrate
liase
malate
malate
dehydrogenase
malic enzyme
glycerol 3P
TG TG
O2
hypoxanthine
NADPH
xanthine
e-
NADP+
F
H2O2
Cl-
LOOH
Fe3+
e-
.O2-
HOCl
acyl CoA synthetase
27-hydroxyChol
LXR
9
r27-hydroxyChol
LXR
9
r
LXR
LXRLXR
R
SCAP
Site
Site-2 protease
?
acetyl CoA carboxylase
fatty acid synthetase
Ser383
Ser389 Elk-1
P Ser383
P Ser389 Elk-1
PKA
Ser276 Ser529
NF- B
p65+p50
Ser32
Ser36
Lys21
Lys22I B
PKA
Ser276 Ser529
NF- B
p65+p50
Ser32
Ser36
Lys21
Lys22I B
PKA
PKA
Ser276 Ser529
NF- B
p65+p50
P Ser32
P Ser36
Lys21
Lys22I B
PKA
Ser276 Ser529
NF- B
p65+p50
P Ser32
P Ser36
Lys21
Lys22I B
PKA
Ser276 Ser529
NF- B
p65+p50
P Ser32
P Ser36
Lys21
Lys22I B
PKA
Ser276 Ser529
NF- B
p65+p50
P Ser32
P Ser36
Lys21
Lys22I B
PKA
Ser276 Ser529
NF- B
p65+p50
P Ser32
P Ser36
UbLys21
UbLys22
I B
PKA
Ser276 Ser529
NF- B
p65+p50
P Ser32
P Ser36
UbLys21
UbLys22
I B
PKA Ser276 Ser529
NF- B
p65+p50PKA
Ser276 Ser529
NF- B
p65+p50
PKA
CK II
NOSII/iNOS
NADPH
oxidase
xanthine
oxidase
SOD
MPO
ATP
synthetase
ATP
ADP
O2
H+
e-
H+
PP2A
ys63TRAF6
Ser Thr LysIRAK1
ys63TRAF6
Ser Thr LysIRAK1
UbLys63TRAF6
P Ser
P Thr
UbLysIRAK1
UbLys63TRAF6
P Ser
P Thr
UbLysIRAK1
TAB2
Ser Thr TAB1 Ser192 Thr184 Thr187 TAK1
Ser Thr TAB1
Ser192 Thr184 Thr187 TAK1
TAB2
Ser Thr TAB1
Ser192 Thr184 Thr187 TAK1
TAB2
Ser Thr TAB1
Ser192 Thr184 Thr187 TAK1
TAB2
UbLys63TRAF6
P Ser
P Thr
UbLysIRAK1
Ser Thr TAB1
Ser192 Thr184 Thr187 TAK1
TAB2
UbLys63TRAF6
P Ser
P Thr
UbLysIRAK1
Ser Thr TAB1
Ser192 Thr184 Thr187 P
TAK1
PTAB2
UbLys63TRAF6
P Ser
P Thr
UbLysIRAK1
Ser Thr TAB1
Ser192 Thr184 Thr187 P
TAK1
PTAB2
UbLys63TRAF6
P Ser
P Thr
UbLysIRAK1
P Ser
P Thr
UbLysIRAK1
P Ser P
Thr TAB1
P Ser192
P Thr184
P Thr187 PTAK1
PTAB2
UbLys63TRAF6
P Ser P
Thr TAB1
P Ser192
P Thr184
P Thr187 PTAK1
PTAB2
UbLys63TRAF6
c-fos
c-jun
AP-1
c-Fos+c-Jun
Ser21 Ser32
Ser42 Ser70
Ser113
Ser374 c-Fos
Ser63 Ser73
c-Jun
P Ser63
P Ser73
c-Jun
P Ser21 P
Ser32
P Ser42
P Ser70 P
Ser113
P Ser374
c-Fos
P Ser63
P Ser73
c-Jun
p53
IL-4
IL-4
0
TRAF2 TRAF1 A20TRAF2 TRAF1 A20
IFN
IFN
A20
PAFR
calpain
TRAM
TRAMTRAM
Tyr580
Tyr542SHP-2
IL-1ra
IL-1ra
P Ser386
P Ser385
P Ser386
P Ser385
IRF-3
P Ser386
P Ser385
P Ser386
P Ser385
IRF-3
P Tyr STAT6
P Ser276
P Ser529
P Ser276
P Ser529
NF- B
p65+p50
P Ser276
P Ser529
P Ser276
P Ser529
NF- B
p65+p50
GM-CSF
GM-CSF
Tyr STAT3
P Tyr STAT3
-2
PTyr580
PTyr542
SHP-2
K
PI3K
Tyr STAT5
P Tyr STAT5
PTyr
PTyry
PTyr
PTyry
PTyr759
PTyr767
PTyr814 P
Tyr905
PTyr915
IL-6R
PTyr759
PTyr767
PTyr814 P
Tyr905
PTyr915
IL-6R
PTyr JAK1
PTyrTyk2
SOCS3
PTyr759
PTyr767
PTyr814 P
Tyr905
PTyr915
IL-6R
PTyr759
PTyr767
PTyr814 P
Tyr905
PTyr915
IL-6R
PTyr JAK1
PTyrTyk2
SOCS3
TyrIFN R2
PTyr440IFN R1
PTyr
IFN R2
PTyr JAK1
PTyr1007
JAK2
SOCS1
/JAB
PTyr440IFN R1
PTyr
IFN R2
PTyr JAK1
PTyr1007
JAK2
SOCS1
/JAB
Tyr1007JAK2
TyrIFN R2
Tyr1007JAK2
TyrIFN R2
y Tyr1007y Tyr1007P
y
PTyr1007
Py
PTyr1007
IL-4R
p38MAPKP
Ser473
P Thr38
Akt/PKB
IRF-9
P Tyr STAT2
PTyr701
P Ser727 STAT1
IRF-9
P Tyr STAT2
PTyr701
P Ser727 STAT1
IRF-9
P Tyr STAT2
PTyr701
P Ser727 STAT1
IRF-9
P Tyr STAT2
PTyr701
P Ser727 STAT1
PIAS3
PIAS3
P TyrP Tyr STAT3
PIAS3
P TyrP Tyr STAT3
Tyr701 Ser727
Tyr701 Ser727 STAT1
MKP
PIAS1
PIAS1
PTyr701
P Ser727
PTyr701
P Ser727 STAT1
PIAS1
PTyr701
P Ser727
PTyr701
P Ser727 STAT1
PTyrJAK1
PTyrJAK3
PTyrIL-4R
PTyr
common
chain
SOCS1
/JAB
PTyrJAK1
PTyrJAK3
PTyrIL-4R
PTyr
common
chain
SOCS1
/JAB
SHP-1
SHIP
Tyr Fyn
P Tyr Fyn
PI3K
PI3K
Ser473 Thr38 Akt/PKB
PTyr
P Thr
JNK
proteasome
P Ser21 P
Ser32
P Ser42
P Ser70 P
Ser113
P Ser374
c-Fos
PP2B
Thr183
Tyr185
Thr183
Tyr185
ERK1
ERK2
MKP
LXRRXR
CPT1
SREBP1c
/ bHLH
SREBP1c
/ bHLH
SREBP1c
/ bHLH
Tyrchain
TyrTyr
chain
Fc RIa
chain
Tyr518
Tyr519Syk
Tyr518
Tyr519Syk
PTyr518
PTyr519
Syk
PTyr771
PTyr783
PTyr1254
PLCTyr771
Tyr783 Tyr1254PLC
PP2APP2B
Pi
PTyr580
PTyr542SHP-2
PTyr580
PTyr542
SHP-2
P Tyr Gab2
PTyr580
PTyr542SHP-2
P Tyr Gab2
PTyr580
PTyr542SHP-2
P Ser63
P Ser73
P Ser63
P Ser73
c-Jun
P Ser369
P Thr577
P Ser386
P Ser227 RSK
P Ser369
P Thr577
P Ser386 Ser227
RSK
Ser133 CREB
P Ser133
CREB
Grb2
P Ser
P Thr
SOSGrb2
P Ser
P Thr
SOS
ASK
P MEKK
SEK1/MKK4
SEK2/MKK7
P
SEK1/MKK4
P
SEK2/MKK7
PTyr Thr
JNK
Tyr Thr
JNK
Ser473 Thr38
Akt/PKB
P Ser473
P Thr38
Akt/PKB
Tyr
P Ser312(307:R)
IRS
SOCS3
PTyr
P Thr
JNK
P Thr183
PTyr185
ERK1
ERK2
P Ser369
P Ser386
R
Src
P
UbcH5
p50
p60
TICAM-1TICAM-1TRIF/
TICAM-1
CHAPTER OUTLINE
589
Melanie H. Cobb and Elliott M. RossThe University of Texas Southwestern Medical Center at Dallas
IntroductionCellular signaling is primarily chemicalReceptors sense diverse stimuli but initiate a limitedrepertoire of cellular signalsReceptors are catalysts and amplifiersLigand binding changes receptor conformationSignals are sorted and integrated in signaling pathwaysand networksCellular signaling pathways can be thought of asbiochemical logic circuitsScaffolds increase signaling efficiency and enhancespatial organization of signalingIndependent, modular domains specify protein-proteininteractionsCellular signaling is remarkably adaptiveSignaling proteins are frequently expressed as multiplespeciesActivating and deactivating reactions are separate andindependently controlledCellular signaling uses both allostery and covalentmodificationSecond messengers provide readily diffusible pathwaysfor information transferCa2+ signaling serves diverse purposes in all eukaryoticcellsLipids and lipid-derived compounds are signalingmoleculesPI 3-kinase regulates both cell shape and the activationof essential growth and metabolic functionsSignaling through ion channel receptors is very fastNuclear receptors regulate transcriptionG protein signaling modules are widely used and highlyadaptable
Heterotrimeric G proteins regulate a wide variety ofeffectorsHeterotrimeric G proteins are controlled by a regulatoryGTPase cycleSmall, monomeric GTP-binding proteins are multiuseswitchesProtein phosphorylation/dephosphorylation is a majorregulatory mechanism in the cellTwo-component protein phosphorylation systems aresignaling relaysPharmacological inhibitors of protein kinases may beused to understand and treat diseasePhosphoprotein phosphatases reverse the actions ofkinases and are independently regulatedCovalent modification by ubiquitin and ubiquitin-likeproteins is another way of regulating protein functionThe Wnt pathway regulates cell fate during developmentand other processes in the adultDiverse signaling mechanisms are regulated by proteintyrosine kinasesSrc family protein kinases cooperate with receptorprotein tyrosine kinasesMAPKs are central to many signaling pathwaysCyclin-dependent protein kinases control the cell cycleDiverse receptors recruit protein tyrosine kinases to theplasma membraneWhat’s next?SummaryReferences
14.36
14.35
14.34
14.33
14.32
14.31
14.30
14.29
14.28
14.27
14.26
14.25
14.24
14.23
14.22
14.21
14.20
14.19
14.18
14.17
14.16
14.15
14.14
14.13
14.12
14.11
14.10
14.9
14.8
14.7
14.6
14.5
14.4
14.3
14.2
14.1
Principles of cell signaling
14
This image represents about 10% of the map of the known signaling interactions andreactions in the mouse macrophage. Preparing such a map in a computable format isthe first step in analyzing a large signaling network. This map was prepared by the groupled by Hiroaki Kitano at the Systems Biology Institute, Tokyo, using their CellDesignerprogram. Map courtesy of Kanae Oda, Yukiko Matsuoka, and Hiroaki Kitano (The SystemsBiology Institute).
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 589

590 CHAPTER 14 Principles of cell signaling
nearby), odors, molecules that regulate growthor differentiation, and proteins on the outsideof adjacent cells. A mammalian cell typicallyexpresses about fifty distinct receptors that sensedifferent inputs, and, overall, mammals expressseveral thousand receptors.
Despite the diversity of cellular lifestylesand the enormous number of substances sensedby different cells, the general classes of proteinsand mechanisms involved in signal transduc-tion are conserved throughout living cells, asshown in FIGURE 14.1.
• G protein-coupled receptors, composed of seven membrane-span-ning helices, promote activation of het-erotrimeric GTP-binding proteins calledG proteins, which associate with the in-ner face of the plasma membrane andconvey signals to multiple intracellularproteins.
• Receptor protein kinases are oftendimers of single membrane-spanningproteins that phosphorylate their in-tracellular substrates and, thus, changethe shape and function of the target pro-teins. These protein kinases frequentlycontain protein interaction domains thatorganize complexes of signaling pro-teins on the inner surface of the plasmamembrane.
• Phosphoprotein phosphatases re-verse the effect of protein kinases by re-moving the phosphoryl groups addedby protein kinases.
• Other single membrane-spanning en-zymes, such as guanylyl cyclase, havean overall architecture similar to the re-ceptor protein kinases but different en-zymatic activities. Guanylyl cyclasecatalyzes the conversion of GTP to 3′:5′-cyclic GMP, which is used to propagatethe signal.
• Ion channel receptors, although di-verse in detailed structure, are usuallyoligomers of subunits that each containseveral membrane-spanning segments.The subunits change their conforma-tions and relative orientations to per-mit ion flux through a central pore.
• Two-component systems may eitherbe membrane spanning or cytosolic. Thenumber of their subunits is also vari-able, but each two-component systemcontains a histidine kinase domain orsubunit that is regulated by a signalingmolecule and a response regulator that
IntroductionAll cells, from prokaryotes through plants andanimals, sense and react to stimuli in their en-vironments with stereotyped responses that al-low them to survive, adapt, and function inways appropriate to the needs of the organism.These responses are not simply direct physicalor metabolic consequences of changes in thelocal environment. Rather, cells express arraysof sensing proteins, or receptors, that recognizespecific extracellular stimuli. In response tothese stimuli, receptors regulate the activitiesof diverse intracellular regulatory proteins thatin turn initiate appropriate responses by thecell. The process of sensing external stimuli andconveying the inherent information to intra-cellular targets is referred to as cellular signaltransduction.
Cells respond to all sorts of stimuli. Microbesrespond to nutrients, toxins, heat, light, andchemical signals secreted by other microbes.Cells in multicellular organisms express recep-tors specific for hormones, neurotransmitters,autocrine and paracrine agents (hormone-like compounds from the secreting cell or cells
14.1
Responseregulator
SensorHistidinekinase( (
E1
E2
E1
E2Hetero-trimeric
G protein
(GPCR)G proteincoupledreceptor
Trans-membrane
scaffoldGuanylylcyclaseReceptor
proteinkinase
Ionchannel
Two-component
complex
Transcriptionfactor
NUCLEUS
Overview of major receptor types in a cell
FIGURE 14.1 Receptors form a rather small number of families that share com-mon mechanisms of action and overall similar structures.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 590

14.2 Cellular signaling is primarily chemical 591
contains a phosphorylatable aspartate(Asp) residue.
• Some receptors are transmembranescaffolds that change either the con-formation or oligomerization of theirintracellular scaffold domains in re-sponse to extracellular signaling mole-cules, or ligands, and, thus, recruitinteracting regulatory proteins to a com-mon site on the membrane.
• Nuclear receptors are transcriptionfactors, often heterodimers, that mayreside in the cytoplasm until activatedby agonists or may be permanently lo-cated in the nucleus.
The biochemical processes of signal trans-duction are strikingly similar among cells.Bacteria, fungi, plants, and animals use similarproteins and multiprotein modules to detectand process signals. For example, evolutionar-ily conserved heterotrimeric G proteins and Gprotein-coupled receptors are found in plants,fungi, and animals. Similarly, 3′:5′ cyclic AMP(cAMP) is an intracellular signaling moleculein bacteria, fungi, and animals; and Ca2+ servesa similar role in all eukaryotes. Protein kinasesand phosphoprotein phosphatases are used toregulate enzymes in all cells.
Although the basic biochemical componentsand processes of signal transduction are con-served and reused, they are often used in wildlydivergent patterns and for many different phys-iological purposes. For example, cAMP is synthe-sized by distantly related enzymes in bacteria,fungi, and animals, and acts on different pro-teins in each organism; it is a pheromone insome slime molds.
Cells often use the same series of signalingproteins to regulate a given process, such astranscription, ion transport, locomotion, andmetabolism. Such signaling pathways are as-sembled into signaling networks to allow thecell to coordinate its responses to multiple in-puts with its ongoing functions. It is now pos-sible to discern conserved reaction sequencesin and between pathways in signaling networksthat are analogous to devices within the circuitsof analog computers: amplifiers, logic gates,feedback and feed-forward controls, and mem-ory.
This chapter discusses the principles andstrategies of cellular signaling first and then dis-cusses the conserved biochemical componentsand reactions of signaling pathways and howthese principles are applied.
Cellular signaling isprimarily chemical
Most signals sensed by cells are chemical, and,when physical signals are sensed, they are gen-erally detected as chemical changes at the levelof the receptor. For example, the visual pho-toreceptor rhodopsin is composed of the pro-tein opsin, which binds to a second component,the colored vitamin A derivative cis-retinal (thechromophore). When cis-retinal absorbs aphoton, it photoisomerizes to trans-retinal,which is an activating ligand of the opsin pro-tein. (For more on rhodopsin signaling see 14.20G protein signaling modules are widely used andhighly adaptable). Similarly, plants sense red andblue light using the photosensory proteins phy-tochrome and cryptochrome, which detect pho-tons that are absorbed by their tetrapyrrole orflavin chromophores. Cryptochrome homologsare also expressed in animals, where they prob-ably mediate adjustment of the diurnal cycle.
A few receptors do respond directly to phys-ical inputs. Pressure-sensing channels, which ex-ist in one form or another in all organisms,mediate responses to pressure or shear by chang-ing their ionic conductance. In mammals, hear-ing is mediated indirectly by a mechanicallyoperated channel in the hair cell of the inner ear.The extracellular domain of a protein called cad-herin is pulled in response to acoustic vibration,generating the force that opens the channel.
Cells sense mechanical strain through anumber of cell surface proteins, including inte-grins. Integrins provide signals to cells based ontheir attachment to other cells and to molecu-lar complexes in the external milieu.
One major group of physically responsivereceptors is made up of channels that sense elec-tric fields. Another interesting group areheat/pain-sensing ion channels; several of theseheat-sensitive ion channels also respond tochemical compounds, such as capsaicin, the“hot” lipid irritant in hot peppers.
Whether a signal is physical or chemical, thereceptor initiates the reactions that change thebehavior of the cell. We will discuss how theseeffects are generated in the rest of the chapter.
Key concepts • Cells can detect both chemical and physical
signals.• Physical signals are generally converted to
chemical signals at the level of the receptor.
14.2
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 591

592 CHAPTER 14 Principles of cell signaling
Receptors sense diversestimuli but initiate alimited repertoire ofcellular signals
Receptors mediate responses to amazingly di-verse extracellular messenger molecules; hence,the cell must express a large number of recep-tor varieties, each able to bind its extracellularligand. In addition, each receptor must be ableto initiate a cellular response. Receptors, thus,contain two functional domains: a ligand-binding domain and an effector domain,which may or may not correspond to definablestructural domains within the protein.
The separation of ligand-binding and effec-tor functions allows receptors for diverse ligandsto produce a limited number of evolutionarilyconserved intracellular signals through the ac-tion of a few effector domains. In fact, there are
Key concepts • Receptors contain a ligand-binding domain and an
effector domain.• Receptor modularity allows a wide variety of
signals to use a limited number of regulatorymechanisms.
• Cells may express different receptors for the sameligand.
• The same ligand may have different effects on thecell depending on the effector domain of itsreceptor.
14.3only a limited number of receptor families, whichare related by their conserved structures and sig-naling functions (see Figure 14.1).
There are several useful correlates to thetwo-domain nature of receptors. For example,a cell can control its responsiveness to an extra-cellular signal by regulating the synthesis ordegradation of a receptor or by regulating thereceptor’s activity (see 14.10 Cellular signaling isremarkably adaptive).
In addition, the nature of a response is gen-erally determined by the receptor and its effec-tor domain rather than any physicochemicalproperty of the ligand. FIGURE 14.2 illustrates theconcept that a ligand may bind to more thanone kind of receptor and elicit more than onetype of response, or several different ligandsmay all act identically by binding to function-ally similar receptors. For example, the neuro-transmitter acetylcholine binds to two classesof receptors. Members of one class are ion chan-nels; members of the other regulate G proteins.Similarly, steroid hormones bind both to nu-clear receptors, which bind chromatin and reg-ulate transcription, and to other receptors inthe plasma membrane.
Conversely, when multiple ligands bind toreceptors of the same biochemical class, theygenerate similar intracellular responses. For ex-ample, it is not uncommon for a cell to expressseveral distinct receptors that stimulate produc-tion of the intracellular signaling molecule cAMP.The effect of the receptor on the cell will also bedetermined significantly by the biology of thecell and its state at any given time.
Ligand binding and effector domains mayevolve independently in response to varied se-lective pressures. For example, mammalian andinvertebrate rhodopsins transduce their signalthrough different effector G proteins (Gt andGq, respectively). Another example is calmod-ulin, a small calcium-binding regulatory pro-tein in animals, which in plants appears as adistinct domain in larger proteins.
The receptor’s two-domain nature allowsthe cell to regulate the binding of ligand andthe effect of ligand independently. Covalentmodification or allosteric regulation can al-ter ligand-binding affinity, the ability of the lig-and-bound receptor to generate its signal orboth. We will discuss these concepts further in14.13 Cellular signaling uses both allostery and co-valent modification.
Receptors can be classified either accord-ing to the ligands they bind or the way in whichthey signal. Signal output, which is character-
Ligand ALigand A
Output1
Output2
Output2
Output1
Output1
Ligand B Ligand C
LBD1 LBD1 LBD1 LBD2 LBD3
ED1 ED1 ED1ED2 ED2
Receptors have a ligand-binding domain and an effector domain
CHIMERICRECEPTOR
FIGURE 14.2 Receptors can be thought of as composed of two functional do-mains, a ligand-binding domain (LBD) and an effector domain (ED). The two-domain property implies that two receptors that respond to different ligands(middle) could initiate the same function by activating similar effector do-mains, or that a cell could express two receptor isoforms (left) that respond tothe same ligand with distinct cellular effects mediated by different effector do-mains. It also implies that one can create an artificial chimeric receptor withnovel properties.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 592

14.5 Ligand binding changes receptor conformation 593
istic of the effector domain, usually correlatesbest with overall structure and sequence con-servation. (Receptor families grouped by theirfunctions are the organizational basis of the sec-ond half of this chapter.) However, classifyingreceptors pharmacologically, according to theirspecificity for ligands, is particularly useful forunderstanding the organization of endocrineand neuronal systems and for categorizing themultiple physiological responses to drugs.
Expression of a receptor that is not nor-mally expressed in a cell is often sufficient toconfer responsiveness to that receptor’s ligand.This responsiveness often occurs because thecell expresses the other components necessaryfor propagating the intracellular signal from thereceptor. The precise nature of the response willreflect the biology of the cell. Experimentally,responsiveness to a compound can be inducedby introducing the cDNA that encodes the re-ceptor. For example, mammalian receptors maybe expressed in yeast, such that the yeast re-spond visibly to receptor ligands, thus provid-ing a way to screen for new chemicals (drugs)that activate the receptor.
Finally, it is possible to create chimeric re-ceptors by fusing the ligand-binding domainfrom one receptor with the effector domainfrom a different receptor (Figure 14.2). Suchchimeras can mediate novel responses to theligand. With genetic modification of the ligand-binding domain, receptors can be reengineeredto respond to novel ligands. Thus, scientists canmanipulate cell functions with nonbiologicalcompounds.
Receptors are catalystsand amplifiers
Receptors act to accelerate intracellular func-tions and are, thus, functionally analogous to en-zymes or other catalysts. Some receptors,including the protein kinases, protein phos-phatases, and guanylate cyclases, are themselvesenzymes and thus classical biochemical cata-lysts. More generally, however, receptors usethe relatively small energy of ligand binding toaccelerate reactions that are driven by alterna-tive energy sources. For example, receptors thatare ion channels catalyze the movement of ions
Key concepts • Receptors act by increasing the rates of key
regulatory reactions.• Receptors act as molecular amplifiers.
14.4
across membranes, a process driven by the elec-trochemical potential developed by distinct ionpumps. G protein-coupled receptors and otherguanine nucleotide exchange factors catalyzethe exchange of GDP for GTP on the G protein,an energetically favored process dictated by thecell’s nucleotide energy balance. Transcriptionfactors accelerate the formation of the transcrip-tional initiation complex, but transcription it-self is energetically driven by multiple steps ofATP and dNTP hydrolysis.
As catalysts, receptors enhance the rates ofreactions. Most signaling involves kinetic ratherthan thermodynamic regulation; that is, sig-naling events change reaction rates rather thantheir equilibria (see the next section). Thus, sig-naling is similar to metabolic regulation, inwhich specific reactions are chosen according totheir rates, with thermodynamic driving forcesplaying only a supportive role.
In all signaling reactions, receptors use theircatalytic activities to function as molecular am-plifiers. Directly or indirectly, a receptor gener-ates a chemical signal that is huge, bothenergetically and with respect to the numberof molecules recruited by a single receptor.Molecular amplification is a hallmark of recep-tors and many other steps in cellular signalingpathways.
Ligand binding changesreceptor conformation
A central mechanistic question in receptor func-tion is how the binding of a signaling moleculeto the ligand-binding domain increases the ac-tivity of the effector domain. The key to thisquestion is that receptors can exist in multiplemolecular conformations, some active for sig-naling and others inactive. Ligands shift theconformational equilibrium among these con-formations. The structural changes that occurduring the receptor’s inactive-active isomeriza-tion and how ligand binding drives thesechanges are exciting areas of biophysical re-search. However, the basic concept can be de-scribed simply in terms of coupling theconformational isomerizations of the ligand-binding and effector domains.
Key concepts • Receptors can exist in active or inactive
conformations.• Ligand binding drives the receptor toward the
active conformation.
14.5
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 593

594 CHAPTER 14 Principles of cell signaling
How do ligands activate (or not activate) areceptor? Most of the basic regulatory activitiesof receptors can be described by a simple schemethat considers the receptors as having two in-terconvertible conformations, inactive (R) andactive (R*). R and R* are in equilibrium, whichis described by the equilibrium constant J.
Because unliganded receptors are usuallyminimally active, J<<1 and an unliganded recep-tor spends most of its time in the R state. Whena signaling molecule (L) binds, it drives the re-ceptor toward the active conformation, R*, inwhich the effector domain is functional. The lig-and-bound receptor thus spends most of its timein the active R* state.
The mechanism whereby a ligand can ac-tivate receptor is a simple consequence of itsrelative affinities for the receptor’s active andinactive conformations. A ligand can bind tothe receptor in either of its conformations, de-scribed here by association constants K for theR state and K* for the R* state. Any ligand thatbinds with higher affinity for the R* conforma-tion than for R will be an activator. If K* is greaterthan K, the ligand is an agonist. According to theSecond Law of Thermodynamics, a system of
R + LJ
R*+ L
R LJ*
K*K
R* L
RJ
R*
coupled equilibria displays path independence:the net free energy difference between twostates is independent of which intermediary re-actions take place. For the receptor, any pathfrom R to R*L therefore has the same free en-ergy change, and the products of the equlib-rium constants along each path are equal. Forthe example above, path independence meansthat:
J•K* = K•J*Therefore, J* / J = K* / K.Thus, if binding to the R* configuration is
preferred (i.e., K*/K>>1), then ligand bindingwill shift the conformation to the R* state to anequivalent extent (i.e., J*/J>>1). The relativeactivation by a saturating concentration of lig-and, J*/J, will exactly equal the ligand’s relativeselectivity for the active receptor conformation,K*/K. This argument is generally valid for the reg-ulation of a protein’s activity by any regulatoryligand.
This model explains many properties of re-ceptors and their ligands both simply and quan-titatively.
• First, J must be greater than zero for theequilibrium to exist. Thus, even unli-ganded receptor has some activity.Overexpressed receptors frequently dis-play their intrinsic low activity.
• Because physiological receptors arenearly inactive in the absence of ligand,J must be much less than 1 and is prob-ably less than 0.01; most receptors areless than 1% active without agonist.
• Ligands can vary in their selectivitiesbetween R and R*. Their abilities to ac-tivate will also vary. Some ligands, re-ferred to as agonists, can drive formationof appreciable R*. Others, known as par-tial agonists, will promote submaxi-mal activation. Chemical manipulationof a ligand’s structure will often alter itsactivity as an agonist. These relation-ships are depicted graphically in FIGURE14.3.
• A ligand that binds equally well to boththe R and R* states will not cause acti-vation. However, such a ligand may stilloccupy the binding site and therebycompetitively inhibit binding of an ac-tivating ligand. Such competitive in-hibitors, referred to as antagonists, arefrequently used as drugs to block un-wanted activation of a receptor in var-ious disease states.
• A ligand that binds preferentially to R
0
1.0
0.8
0.6
0.4
0.2
0
0.012
0.010
0.008
0.006
0.004
0.002
Log [L]
Partialagonist
Log [L]
High affinityagonist
Lower affinityagonist
Fractional activityof receptor
Fractional activityof receptor
Inverseagonist
Receptor ligands can vary in their activities and potencies
FIGURE 14.3 The simple two-state model shown here can describe a wide va-riety of behaviors displayed by receptors and their various regulatory ligands.The left panel shows fractional activity of a receptor exposed to two agonistswith different affinities and one partial agonist. The right panel shows the ef-fect of an inverse agonist. If the low fractional activity of unliganded receptoris detected as significant biological activity, then its inhibition by the inverseagonist would be easily detectable.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 594

14.6 Signals are sorted and integrated in signaling pathways and networks 595
relative to R* will further shift the con-formational equilibrium to the inactivestate and cause net inhibition. Such lig-ands are called inverse agonists.Because J is already low, effects of in-verse agonists may only be noticeableif a receptor is overexpressed or if thereceptor is mutated to increase its in-trinsic activity (i.e., the mutation in-creases J).
• The extent to which an agonist stimu-lates a receptor is unrelated to its affin-ity. Both agonists and antagonists maybind with either high or low affinity.Affinity does determine the receptor’ssensitivity—that is, how low a concen-tration of ligand can the receptor detect.Affinities of receptors for natural regu-latory ligands vary enormously, withphysiologic Kd values ranging from<10-12 M for some hormones to about10-3 M for some bacterial chemoattrac-tants. Another aspect of sensitivity ishow abruptly or gradually the receptoris activated as the concentration of ag-onist increases. The above model pre-dicts that a receptor is activatedsignificantly at agonist concentrationsbetween 0.1 and 10 times its Kd. A va-riety of cellular mechanisms can con-vert such a conventional response rangeof about 100-fold to either a more grad-ual response or a very steep, switchlikeresponse.
• This model only describes equilibria. Itmakes no predictions about the rates ofligand binding or release, or of the con-formational isomerization that leads toactivation.
This model shows how three important as-pects of receptor action are independently de-termined. As mentioned above, affinity forligand, which determines the concentrationrange over which the ligand functions, is inde-pendent of the ligand’s net effectiveness at driv-ing receptor activation. The rate of response isalso largely independent of these other twoproperties. Each aspect of receptor function canthus be independently regulated in response toother incoming signals or by the metabolic ordevelopmental state of the cell. Such control ofsignal input is central to whole-cell coordina-tion of signal transduction. Examples and mech-anisms will recur throughout this chapter.
Signals are sorted andintegrated in signalingpathways and networks
Receptors rarely act directly on the intracellu-lar processes that they ultimately regulate.Rather, receptors typically initiate a sequence ofregulatory events that involve intermediaryproteins and small molecules. The use of mul-tistep signaling pathways allows cells to amplifysignals, adjust signaling kinetics, insert controlpoints, integrate multiple signals, and route sig-nals to distinct effectors.
Branched pathways give cells the ability tointegrate multiple incoming signals and to di-rect information to the correct control points.As FIGURE 14.4 illustrates, branching can be ei-ther convergent, with multiple signals regulat-ing common end points, or divergent, with asingle pathway branching to control more thanone process. In multicellular organisms, diver-gent branching allows a single hormone recep-tor to initiate distinct cell-appropriate patternsof responses in different cells and tissues.Divergent signaling also allows a receptor toregulate qualitatively different cellular responseswith quantitatively distinct intensities, each de-pendent on signal amplification in the interme-diary pathway.
Convergent branching—when several re-ceptors activate the same pathway to elicit thesame regulatory responses—is also common.Convergent branching allows multiple incom-ing signals, both stimulatory and inhibitory, tobe integrated and coordinately regulated at acommon site downstream of the receptors.Receptors for several different hormones fre-quently initiate similar or overlapping patternsof signaling in a single target cell.
Overlapping converging and diverging sig-naling pathways create signaling networks withincells that coordinate responses to multiple in-puts (Figure 14.4). Typically, such pathways arecomplex in the number and diversity of theircomponents and in the topology of their circuit
Key concepts • Signaling pathways usually have multiple steps
and can diverge and/or converge.• Divergence allows multiple responses to a single
signal.• Convergence allows signal integration and
coordination.
14.6
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 595

596 CHAPTER 14 Principles of cell signaling
maps. Signaling networks are also spatially com-plex. They may include components in varioussubcellular locations, with initial receptors andassociated proteins in the plasma membrane, butwith downstream proteins in the cytoplasm or in-tracellular organelles. Such complexity is neces-sary to allow the cells to integrate and sortincoming signals and to regulate multiple intra-cellular functions simultaneously.
The complexity and adaptability of signal-ing networks, like the one shown in the lowerhalf of Figure 14.4, make their dynamics at thewhole-cell level difficult or impossible to graspintuitively. Signaling networks resemble large
analog computers, and investigators are increas-ingly depending on computational tools to un-derstand cellular information flow and itsregulation. First, many signaling interactionsthat include only two or three proteins exertfunctions analogous to traditional computa-tional logic circuits (see the next section). Thetheory and experience with such circuits in elec-tronics facilitate understanding biological sig-naling functions as well.
The enormous complexity of cellular signal-ing networks can be simplified by consideringthem to be composed of interacting signalingmodules, i.e., groups of proteins that process sig-nals in well-understood ways. A cellular signal-ing module is analogous to an integrated circuitin an electronic instrument that performs aknown function, but whose exact componentscould be changed for similar use in another de-vice. The concept of modular construction facil-itates both qualitative and quantitativeunderstanding of signaling networks. We will re-fer to many standard signaling modules later inthe chapter. Examples include monomeric andheterotrimeric G protein modules, MAPK cas-cades, tyrosine (Tyr) kinase receptors and theirbinding proteins, and Ca2+ release/uptake mod-ules. In each case, despite the numerous phylo-genetic, developmental, and physiologicvariations, understanding the basic function ofthat class of module conveys understanding of allits incarnations. Last, the evolutionary impor-tance of modules is significant; once the architec-ture of a module is established it can be reused.
For larger-scale networks, multiplexed,high-throughput measurements on living cellshave been combined with powerful kinetic mod-eling strategies to allow an increasingly accuratequantitative depiction of information flowwithin signaling modules or entire networks.Such models, with sound and experimentallybased parameter sets, can describe signalingprocesses in systems too complex for intuitiveor ad hoc analysis. They are also vital as tests ofunderstanding because they can predict exper-imental results in ways that can be used to testthe validity of the model. Well-grounded mod-els can then be used (cautiously) to suggest themechanisms of systems for which data sets re-main unattainable. At even greater levels ofcomplexity, the theories and tools of computerscience are increasingly giving useful systems-level analyses of signal flow in cells. Using com-putational tools to analyze large arrays ofquantitative data allows us to understand cel-lular information flow and its regulation.
Linear,parallel
Convergent Divergent Multiplybranched
RECEPTORS
TRANSDUCERS
EFFECTORS
Convergent and divergent signaling pathways
FIGURE 14.4 Signaling pathways use convergent and divergent branching to co-ordinate information flow. The diagrams at top show how even a simple, three-level signaling network can sort information. Convergence or divergence cantake place at multiple points along a signaling pathway. As an example of com-plexity, the lower portion of the figure shows a small segment (~10%) of the Gprotein-mediated signaling network in a mouse macrophage cell line. It omitsseveral interpathway regulatory mechanisms and completely ignores inputs fromnon-G protein-coupled receptors. Pathway map courtesy of Lily Jiang, Universityof Texas Southwestern Medical Center.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 596

14.7 Cellular signaling pathways can be thought of as biochemical logic circuits 597
Developing quantitative models of signalingnetworks is a frontier in signaling biology. Thesemodels both help describe network functionand pinpoint experiments to clarify mechanism.
Cellular signalingpathways can be thoughtof as biochemical logiccircuits
As introduced in the preceding section, processesthat signaling pathways use to integrate and directinformation to cellular targets are strikingly anal-ogous to the mathematical logic functions that areused to design the individual circuits of electroniccomputers. Indeed, there are biological equiva-lents of essentially all of the functional compo-nents that computer scientists and engineersconsider in the design of computers and electroniccontrol devices. To understand signaling path-ways, it is, therefore, useful to consider groups ofreactions within a pathway as constituting logic cir-cuits of the sort used in electronic computing, asillustrated in FIGURE 14.5. The simplest example iswhen two stimulatory pathways converge. If suf-ficient input from either is adequate to elicit theresponse, the convergence would constitute an“OR” function. If neither input is sufficient by it-self but the combination of the two elicits the re-sponse, then the converging pathways wouldcreate “AND” functions. AND circuits are also re-ferred to as coincidence detectors—a responseis elicited only when two stimulating pathwaysare activated simultaneously.
AND functions can result from the combi-nation of two similar but quantitatively inade-quate inputs. Alternatively, two mechanisticallydifferent inputs might both be required to elicita response. An example of the latter would bea target protein that is allosterically activatedonly when phosphorylated, or that is activatedby phosphorylation but is only functional whenrecruited to a specific subcellular location.
The opposite of an AND circuit is a NOTfunction, where one pathway blocks the stim-
Key concepts • Signaling networks are composed of groups of
biochemical reactions that function asmathematical logic functions to integrateinformation.
• Combinations of such logic functions combine assignaling networks to process information at morecomplex levels.
14.7
ulatory effect of another. Simple logic gates areobserved at many locations in cellular signalingpathways.
We can also think about convergent signal-ing in quantitative rather than Boolean termsby considering the additivity of inputs to a dis-tinct process (see Figure 14.5, right). The ORfunction referred to above can be considered tobe the additive positive inputs of two pathways.Such additivity could represent the ability ofseveral receptors to stimulate a pool of a partic-ular G protein or the ability of two protein ki-nases to phosphorylate a single substrate.Additivity may be positive, as in the examplesabove, or negative, such as when two inhibitoryinputs combine. Inhibition and stimulation mayalso combine additively to yield an algebraicallybalanced output. Alternatively, multiple inputscan combine with either more or less than anadditive effect. The NOT function, discussedabove, is analogous to describing a blockade ofstimulation. The AND function describes syn-ergism, where one input potentiates anotherbut alone has little effect.
Even simple signaling networks can displaycomplex patterns of information processing. One
Additive
Logical (Boolean) Quantitative (Analog)
Response
A + B Response
A OR B
A NOT B
A AND B
B
A Response
Response
BA
A + fixed [B]
A + B
A
Response
Response
A + B
B
A
Less than additive
More than additive
A + B Response
B
A
A + B
B
A Response
log (agonist concentration)
log (agonist concentration)
log (agonist concentration)
B
Simple logic circuits
FIGURE 14.5 Signaling networks use simple logic functions to processinformation. Boolean OR, AND, and NOT functions (left) correspond tothe quantitative interactions between converging signals that are shownon the right.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 597

598 CHAPTER 14 Principles of cell signaling
good example is the creation of “memory”: mak-ing the effect of a transient signal more or lesspermanent. Signaling pathways have multipleways of setting memories, and of forgetting. Onemechanism, common in protein kinase path-ways, is the positive feedback loop, illustratedin the top panel of FIGURE 14.6. In a positive feed-back loop, the input stimulates a transducer (T),which in turn stimulates the effector protein (E)to create the output. If the effector can also ac-
tivate the transducer, sufficient initial signal canbe fed back to the transducer that it can main-tain the effector's full signal output even wheninput is removed. Such systems typically displaya threshold behavior, as shown on the right.
A positive feed-forward loop can generatememory of another type (Figure 14.6, middlepanel), indicating the duration of input. In suchcircuits, the effector requires simultaneous in-put from both the receptor and from the inter-mediary transducer. If the pathway fromreceptor through transducer is relatively slow,or if it requires the accumulation of a substan-tial amount of transducer, only a prolonged in-put will trigger a response, as shown in thetime-base output diagram at the right.
A third way to establish memory is to allowone input to control the reversibility of a sec-ond regulatory event (Figure 14.6, bottom panel).WASP, a protein that initiates the polymerizationof actin to drive cellular motion and shapechange, is activated both by phosphorylationand by the binding of Cdc42, a small GTP-bind-ing protein (G). However, the phosphorylationsite on WASP is only exposed when WASP isbound to Cdc42. Phosphorylation thus requiresboth activated Cdc42 and activated protein ki-nase. If Cdc42 dissociates, the phosphorylatedstate of WASP persists until another signalingmolecule, whose identity remains uncertain,binds again to expose the site to a protein phos-phatase. As shown in the time-base graph, ex-posure to Cdc42 will activate, but exposure tokinase alone will not. If Cdc42 is present, thenthe kinase can activate WASP. Phospho-WASPis relatively insensitive to protein phosphatase(P) alone, but can be dephosphorylated if Cdc42or another G protein binds to expose the site tophosphatase.
Scaffolds increasesignaling efficiency andenhance spatialorganization of signaling
Key concepts • Scaffolds organize groups of signaling proteins and
may create pathway specificity by sequesteringcomponents that have multiple partners.
• Scaffolds increase the local concentration ofsignaling proteins.
• Scaffolds localize signaling pathways to sites ofaction.
14.8
Positive feedback loop : irreversible ON switch
Positive feed-forward loop : responds to prolonged input
Conformational lock - Dual control switch
Input
Input strength
Output
Output
T
Input OutputT
Output
Time
Time
Output
+
+
input
Kinase
Phosphatase
OH
G
OH
G
P
G
P
G
P
G K PG K G P
E
E
OHE E
E E
E E
Signal processing circuits
FIGURE 14.6 Relatively complex signal processing can be executed by simplemulti-protein modules. The figure depicts three types of signaling modules(left) and their behavior in response to agonist (right). (top) In a positivefeed-back module, a transducer protein (T) stimulates an effector (E) to pro-duce a cellular output, but the effector also stimulates the activity of the trans-ducer. The result can be an all-or-none switch, where input up to a thresholdhas little effect, but then becomes committed when feedback from the effec-tor is sufficient to maintain transducer activity even in the absence of contin-ued input from the receptor. (center) In a positive feed-forward module, theeffector requires input both from the transducer and from upstream in the path-way. When stimulation is brief (short horizontal bar under trace at right), sig-nificant amounts of active transducer do not accumulate and output is minimal.When stimulation is prolonged (longer bar), signal output is substantial. (bot-tom) In some dual-control switching modules, the binding of one regulator (G)can both activate the effector and expose another regulatory site, shown hereas a Ser substrate site (-OH) for a protein kinase. The effector can only be phos-phorylated or dephosphorylated when G is bound. Therefore, as shown at theright, addition of G alone will activate but activation of the kinase (K) alonewill not. If kinase is active while G is bound, phosphorylation is resistant tophosphatase activity unless G is again present to reexpose the phosphoserineresidue (shown on the graph at the right as a bold P).
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 598
14.8 Scaffolds increase signaling efficiency and enhance spatial organization of signaling 599
The proteins in a signaling pathway are fre-quently colocalized within cells such that theirmutual interactions are favored and their in-teractions with other proteins are minimized.Many signaling pathways are organized on scaf-folds. Scaffolds bind several components of asignaling pathway in multiprotein complexesto enhance signaling efficiency. Scaffolds pro-mote interactions of proteins that have a lowaffinity for each other, accelerate activation (andoften inactivation) of the associated compo-nents, and localize the signaling proteins to ap-propriate sites of action. Colocalization may betonic or regulated, and stimulus-dependent scaf-folding often determines signaling outputs.
The binding sites on a scaffolding proteinare often localized in distinct modular protein-binding domains, giving the impression that theprotein is designed simply to hold the compo-nents of the pathway together. Many scaffold-ing proteins do lack intrinsic enzymatic activity,but some signaling enzymes also act as scaffolds.
Binding to a scaffold facilitates signaling byincreasing the local concentrations of the com-ponents, so that diffusion or transport of mol-ecules to their sites of action is not necessary. Inthe photoreceptor cells of Drosophila, scaffold-ing of signaling components is critical for rapidsignal transmission. These cells contain the InaD
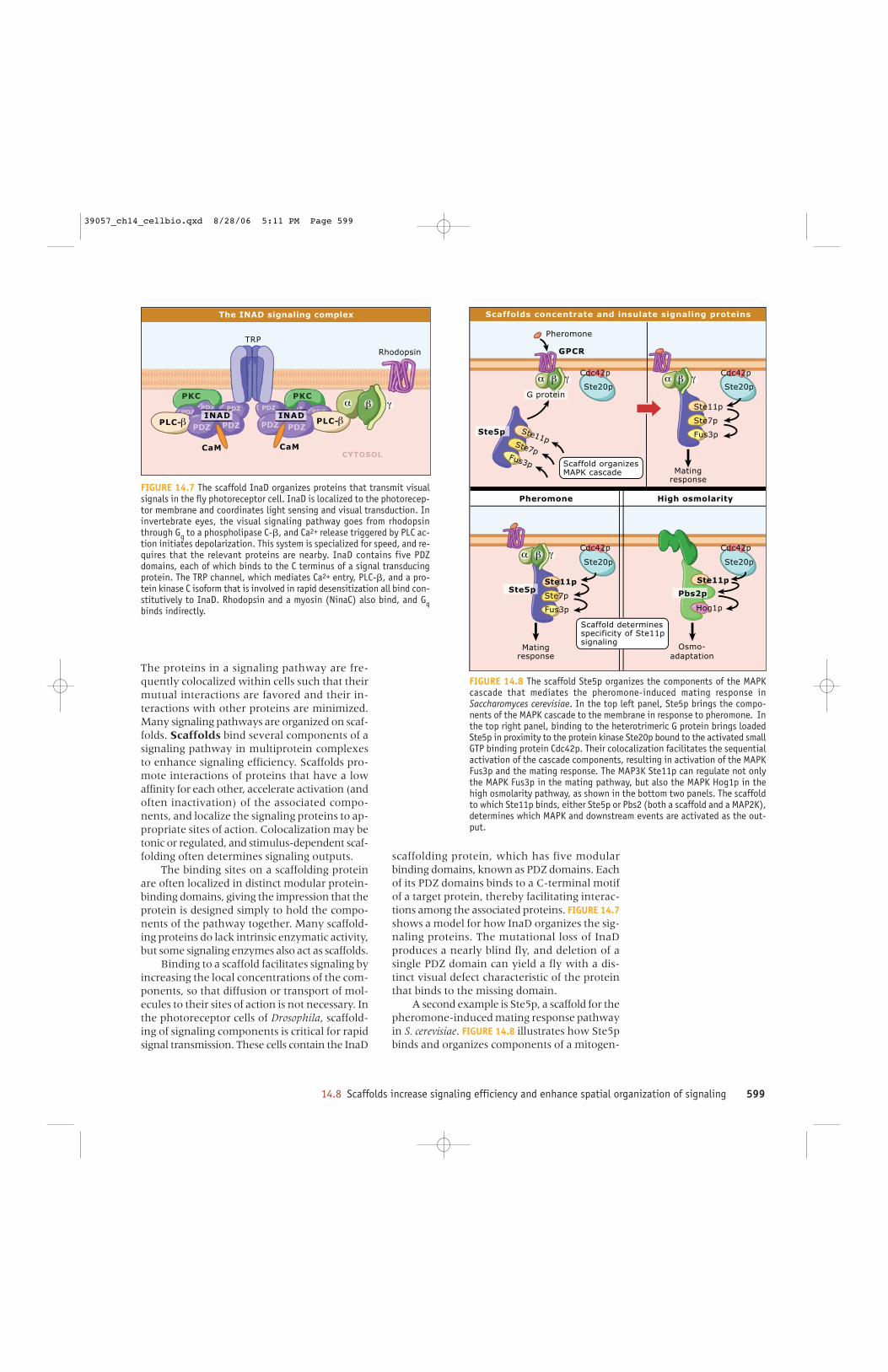
scaffolding protein, which has five modularbinding domains, known as PDZ domains. Eachof its PDZ domains binds to a C-terminal motifof a target protein, thereby facilitating interac-tions among the associated proteins. FIGURE 14.7shows a model for how InaD organizes the sig-naling proteins. The mutational loss of InaDproduces a nearly blind fly, and deletion of asingle PDZ domain can yield a fly with a dis-tinct visual defect characteristic of the proteinthat binds to the missing domain.
A second example is Ste5p, a scaffold for thepheromone-induced mating response pathwayin S. cerevisiae. FIGURE 14.8 illustrates how Ste5pbinds and organizes components of a mitogen-
PKC
CYTOSOL
TRP
Rhodopsin
CaMCaM
PDZPDZ
-
PDZ
PKC
PDZ
-PDZ
INADPDZZ
INADPDZPDZ
PDZ PDZ
PDZ
The INAD signaling complex
FIGURE 14.7 The scaffold InaD organizes proteins that transmit visualsignals in the fly photoreceptor cell. InaD is localized to the photorecep-tor membrane and coordinates light sensing and visual transduction. Ininvertebrate eyes, the visual signaling pathway goes from rhodopsinthrough Gq to a phospholipase C-�, and Ca2+ release triggered by PLC ac-tion initiates depolarization. This system is specialized for speed, and re-quires that the relevant proteins are nearby. InaD contains five PDZdomains, each of which binds to the C terminus of a signal transducingprotein. The TRP channel, which mediates Ca2+ entry, PLC-�, and a pro-tein kinase C isoform that is involved in rapid desensitization all bind con-stitutively to InaD. Rhodopsin and a myosin (NinaC) also bind, and Gqbinds indirectly.
Scaffold determines specificity of Ste11p signaling
Scaffold organizes MAPK cascade
Ste11p
Ste11p
Ste20p
Ste7p
Ste7p
Fus3p
Fus3p
Ste20p
Ste20pSte20p Ste20pSte20p
Ste7p
Fus3p
Ste11p
Pheromone
Cdc42pCdc42p
Cdc42pCdc42p
Matingresponse
G protein
Ste5p
Ste5p Pbs2p
Pheromone High osmolarity
Osmo-adaptation
Matingresponse
Ste11p
Hog1p
Scaffolds concentrate and insulate signaling proteins
GPCR
FIGURE 14.8 The scaffold Ste5p organizes the components of the MAPKcascade that mediates the pheromone-induced mating response inSaccharomyces cerevisiae. In the top left panel, Ste5p brings the compo-nents of the MAPK cascade to the membrane in response to pheromone. Inthe top right panel, binding to the heterotrimeric G protein brings loadedSte5p in proximity to the protein kinase Ste20p bound to the activated smallGTP binding protein Cdc42p. Their colocalization facilitates the sequentialactivation of the cascade components, resulting in activation of the MAPKFus3p and the mating response. The MAP3K Ste11p can regulate not onlythe MAPK Fus3p in the mating pathway, but also the MAPK Hog1p in thehigh osmolarity pathway, as shown in the bottom two panels. The scaffoldto which Ste11p binds, either Ste5p or Pbs2 (both a scaffold and a MAP2K),determines which MAPK and downstream events are activated as the out-put.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 599

600 CHAPTER 14 Principles of cell signaling
activated protein kinase (MAPK) cascade, in-cluding a MAP3K (Ste11p), a MAP2K (Ste7p)and a MAPK (Fus3p). (The MAPK cascade willbe discussed in 14.32 MAPKs are central to manysignaling pathways). The function of Ste5p is par-tially retained even if the positions of its bind-ing sites for the kinases are shuffled in the linearsequence of the protein, indicating that a majorrole is to bring the enzymes into proximity, ratherthan to precisely orient them. Ste5p also bindsto the �� subunits of the heterotrimeric G pro-tein that mediates the actions of matingpheromones, linking the membrane signal tothe intracellular transducers. Yeast that lackSte5p cannot mate, demonstrating that Ste5p isrequired for this biological function (but not allfunctions) carried out by the pathway.
In addition to facilitating signaling in theirown pathways, scaffolds can enhance signalingspecificity by limiting interactions with othersignaling proteins. Scaffolds thus insulate com-ponents of a signaling pathway both from acti-vation by inappropriate signals and fromproducing incorrect outputs. For example, themating and osmosensing pathways in yeastshare several components, including the MAP3KSte11p, but each pathway maintains specificitybecause it employs different scaffolds that restrictsignal transmission.
In contrast, the presence of excess scaffoldcan inhibit signaling because the individual sig-naling components will more frequently bindto distinct scaffold proteins rather than forminga functional complex. Such dilution among scaf-folds causes separation rather than concentra-tion of the components, preventing theirproductive interaction.
Independent, modulardomains specify protein-protein interactions
Modular protein interaction domains or motifsoccur in many signaling proteins and confer theability to bind structural motifs in other mole-cules, including proteins, lipids, and nucleic
Key concepts • Protein interactions may be mediated by small,
conserved domains.• Modular interaction domains are essential for
signal transmission.• Adaptors consist exclusively of binding domains or
motifs.
14.9
acids. Some of these domains are listed in FIGURE14.9. In contrast to scaffolds, which bind spe-cific proteins with considerable selectivity, mod-ular interaction domains generally recognizenot a single molecule but a group of targets thatshare related structural features.
Modular interaction domains important forsignal transduction were first discovered in theprotein tyrosine kinase proto-oncogene Src,which contains a protein tyrosine kinase do-main and two domains named Src homology(SH) 2 and 3 domains. The modular SH2 andSH3 domains were originally identified by com-parison of Src to two other tyrosine kinases, Fpsand Abl. One or both of these domains appearin numerous proteins and both are critically in-volved in protein-protein interactions.
SH3 domains, which consist of approxi-mately 50 residues, bind to specific short pro-line-rich sequences. Many cytoskeletal proteinsand proteins found in focal adhesion complexescontain SH3 domains and proline rich se-quences, suggesting that this targeting motifmay send proteins with these domains to thesesites of action within cells. In contrast to phos-photyrosine-SH2 binding, the proline-rich bind-ing sites for SH3 domains are present in restingand activated cells. However, SH3-proline inter-actions may be negatively regulated by phospho-rylation within the proline-rich motif.
SH2 domains, which consist of approxi-mately 100 residues, bind to Tyr phosphory-lated proteins, such as cytoplasmic tyrosinekinases and receptor tyrosine kinases. Thus, Tyrphosphorylation regulates the appearance ofSH2 binding sites and, thereby, regulates a setof protein-protein interactions in a stimulus-dependent manner.
A clever strategy was used to identify thebinding specificity of SH2 domains. An isolatedrecombinant SH2 domain was incubated withcell lysates and then recovered from the lysatesusing a purification tag. The proteins associatedwith the SH2 domain were some of the sameproteins that were recognized by antiphospho-tyrosine antibodies. By this and other methods,it was discovered that SH2 domains recognizesequences surrounding Tyr phosphorylationsites and require phosphorylation of the in-cluded Tyr for high affinity binding.
Information on specific amino acid se-quences that recognize and bind to modularbinding domains is being accumulated as theseindividual interactions are identified. In addi-tion, screening programs using cDNA and/orpeptide libraries to assess binding capabilities
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 600
14.9 Independent, modular domains specify protein-protein interactions 601
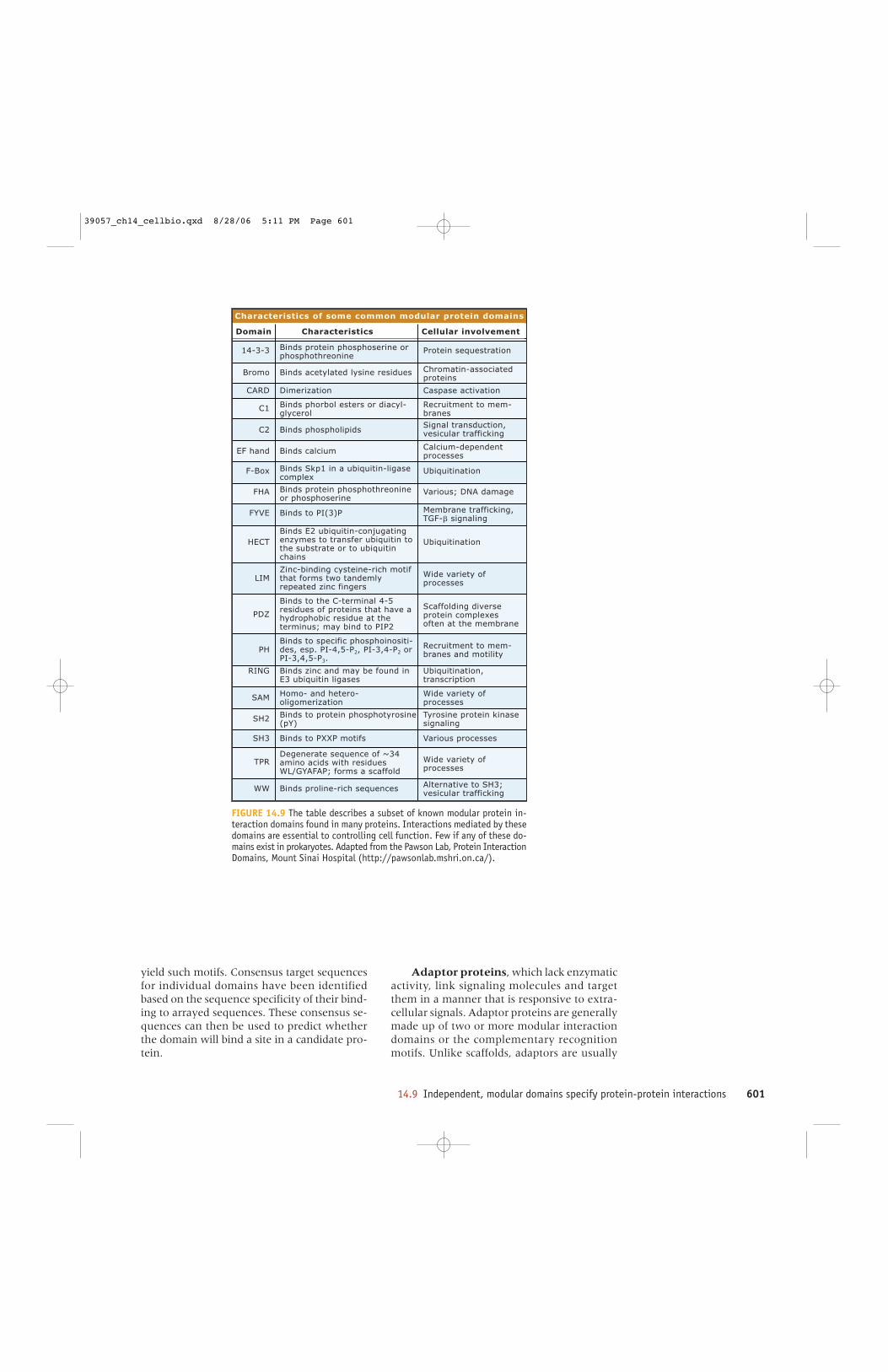
Characteristics of some common modular protein domains
14-3-3 Binds protein phosphoserine or phosphothreonine
Protein sequestration
Domain Characteristics Cellular involvement
WW Binds proline-rich sequences Alternative to SH3; vesicular trafficking
TPRDegenerate sequence of ~34 amino acids with residues WL/GYAFAP; forms a scaffold
Wide variety of processes
SH3 Binds to PXXP motifs Various processes
SH2 Binds to protein phosphotyrosine (pY)
Tyrosine protein kinase signaling
SAM Homo- and hetero-oligomerization
Wide variety of processes
RING Binds zinc and may be found in E3 ubiquitin ligases
Ubiquitination,transcription
PHBinds to specific phosphoinositi-des, esp. PI-4,5-P2, PI-3,4-P2 or PI-3,4,5-P3.
Recruitment to mem-branes and motility
PDZ
Binds to the C-terminal 4-5 residues of proteins that have a hydrophobic residue at the terminus; may bind to PIP2
Scaffolding diverse protein complexes often at the membrane
LIMZinc-binding cysteine-rich motif that forms two tandemly repeated zinc fingers
Wide variety ofprocesses
HECTBinds E2 ubiquitin-conjugating enzymes to transfer ubiquitin to the substrate or to ubiquitin chains
Ubiquitination
FYVE Binds to PI(3)P Membrane trafficking, TGF-� signaling
FHA Binds protein phosphothreonine or phosphoserine
Various; DNA damage
F-Box Binds Skp1 in a ubiquitin-ligase complex
Ubiquitination
EF hand Binds calcium Calcium-dependent processes
C2 Binds phospholipids Signal transduction, vesicular trafficking
C1 Binds phorbol esters or diacyl-glycerol
Recruitment to mem-branes
Dimerization Caspase activation
Bromo
CARD
Binds acetylated lysine residues Chromatin-associated proteins
FIGURE 14.9 The table describes a subset of known modular protein in-teraction domains found in many proteins. Interactions mediated by thesedomains are essential to controlling cell function. Few if any of these do-mains exist in prokaryotes. Adapted from the Pawson Lab, Protein InteractionDomains, Mount Sinai Hospital (http://pawsonlab.mshri.on.ca/).
yield such motifs. Consensus target sequencesfor individual domains have been identifiedbased on the sequence specificity of their bind-ing to arrayed sequences. These consensus se-quences can then be used to predict whetherthe domain will bind a site in a candidate pro-tein.
Adaptor proteins, which lack enzymaticactivity, link signaling molecules and targetthem in a manner that is responsive to extra-cellular signals. Adaptor proteins are generallymade up of two or more modular interactiondomains or the complementary recognitionmotifs. Unlike scaffolds, adaptors are usually
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 601

602 CHAPTER 14 Principles of cell signaling
multifunctional because their modular interac-tion domains and motifs are not as highly spe-cific. Adaptors bind to two or more othersignaling proteins via their protein-protein in-teraction domains to colocalize them or to fa-cilitate additional interactions.
Grb2 is a prototypical adaptor protein thatwas identified as a protein that bound to the C-terminal region of the EGF receptor. Grb2 hasone SH2 and two SH3 domains. It binds consti-tutively to specific proline-rich segments of pro-teins through its SH3 domain, although thisbinding can be negatively regulated. One targetof Grb2 is SOS, a guanine nucleotide exchangefactor that activates the small GTP-binding pro-tein Ras in response to EGF signaling. Throughits SH2 domain, Grb2 binds Tyr-phosphorylatedproteins, including the receptors themselves ina stimulus-dependent manner. Thus, Tyr phos-phorylation of these receptors in response toligand will enable the binding of Grb2 to the re-ceptors, which, in turn, will recruit SOS to themembrane-localized receptor. Once at the mem-brane, SOS can activate its target, Ras.
Cellular signaling isremarkably adaptive
A universal property of cellular signaling pathwaysis adaptation to the incoming signal. Cells contin-uously adjust their sensitivity to signals to main-tain their ability to detect changes in input. Typically,when a cell is exposed to a new input, it initiatesa process of desensitization that dampens the cel-lular response to a new plateau lower than the ini-tial peak response, as illustrated in FIGURE 14.10.When the stimulus is removed, the desensitizedstate can persist, with sensitivity slowly returningto normal. Similarly, the removal of a tonic stim-ulus can hypersensitize signaling systems.
Key concepts • Sensitivity of signaling pathways is regulated to
allow responses to change over a wide range ofsignal strengths.
• Feedback mechanisms execute this function in allsignaling pathways.
• Most pathways contain multiple adaptive feedbackloops to cope with signals of various strengths anddurations.
14.10
Initialresponse
Heterologousdesensitization
Homologousdesensitization
Time
Time
R1 R2 R2
X1 X2
Y
Z
Response
R esponse
ab
K
a
R1 R2
Z
X1 X2
Agonist Agonist Agonist
Desensitization
Agonist a for R1
Reapplya or b
Agonist a for R1
Reapplya or b
Time
Response
R1R1R2R1 or
a
Y
Patterns of adaptation in signaling networksFIGURE 14.10 Top: Upon exposure toa stimulus, signaling pathways adjusttheir sensitivities to adapt to the newlevel of input. Thus, the response de-cays after initial stimulation. A sec-ond similar stimulus will elicit a smallerresponse unless adequate time is al-lowed for recovery. Bottom: Some adap-tation mechanisms feed back only onthe receptor that is stimulated and donot alter parallel pathways. Such mech-anisms are referred to as homologous.At left, agonist a for receptor R1 caninitiate either of two feedback eventsthat desensitize R1 alone. In othercases, a stimulus will also cause par-allel or related systems to desensitize.At the right, agonist a initiates desen-sitization of both R1 and R2. The re-sponse to agonist b, which binds toR2, is also desensitized. Such heterol-ogous desensitization is common.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 602
14.10 Cellular signaling is remarkably adaptive 603
Adaptation in signaling is one of the best ex-amples of biological homeostasis. The adaptabil-ity of cellular signaling can be quite impressive.Cells commonly regulate their sensitivity to phys-iological stimuli over more than a 100-fold range,and the mammalian visual response can adapt toincoming light over a 107-fold range. This re-markable ability allows a photoreceptor cell todetect a single photon, and allows a person toread in both very dim light and intense sunlight.Adaptability is observed in bacteria, plants, fungi,and animals. Many of its properties are conservedthroughout biology, although the most complexadaptive mechanisms are found in animals. Thegeneral mechanism for adaptation is the nega-tive feedback loop, which biochemically samplesthe signal and controls the adaptive process.
Adaptation varies with both the intensity andthe duration of the incoming signal. Stronger ormore persistent inputs tend to drive greater adap-tive change and, often, adaptation that persistsfor a longer time. Cells can modulate adaptationin this way because adaptation is exerted by asuccession of independent mechanisms, each withits own sensitivity and kinetic parameters.
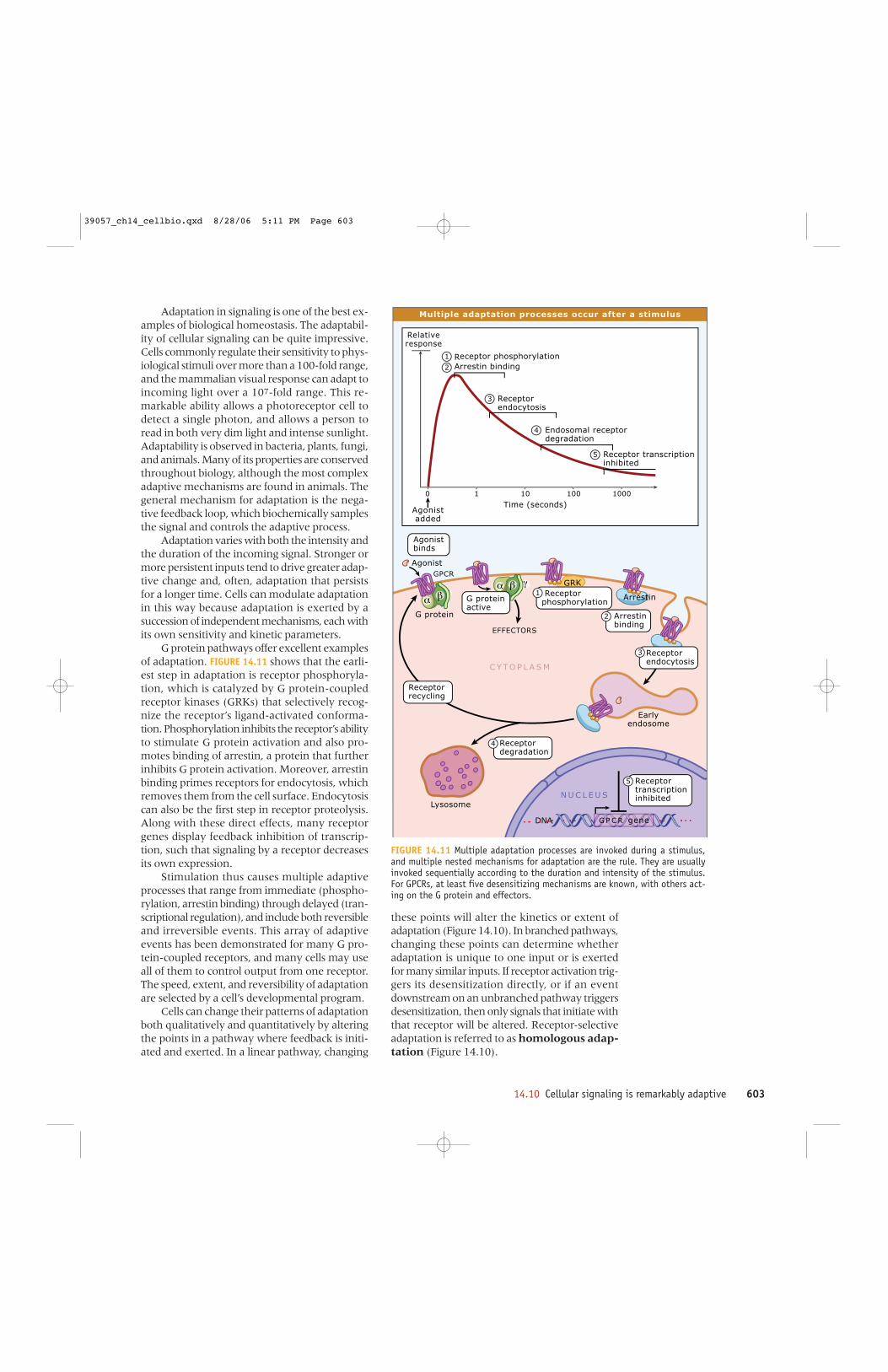
G protein pathways offer excellent examplesof adaptation. FIGURE 14.11 shows that the earli-est step in adaptation is receptor phosphoryla-tion, which is catalyzed by G protein-coupledreceptor kinases (GRKs) that selectively recog-nize the receptor’s ligand-activated conforma-tion. Phosphorylation inhibits the receptor’s abilityto stimulate G protein activation and also pro-motes binding of arrestin, a protein that furtherinhibits G protein activation. Moreover, arrestinbinding primes receptors for endocytosis, whichremoves them from the cell surface. Endocytosiscan also be the first step in receptor proteolysis.Along with these direct effects, many receptorgenes display feedback inhibition of transcrip-tion, such that signaling by a receptor decreasesits own expression.
Stimulation thus causes multiple adaptiveprocesses that range from immediate (phospho-rylation, arrestin binding) through delayed (tran-scriptional regulation), and include both reversibleand irreversible events. This array of adaptiveevents has been demonstrated for many G pro-tein-coupled receptors, and many cells may useall of them to control output from one receptor.The speed, extent, and reversibility of adaptationare selected by a cell’s developmental program.
Cells can change their patterns of adaptationboth qualitatively and quantitatively by alteringthe points in a pathway where feedback is initi-ated and exerted. In a linear pathway, changing
DNA
GRK
Relative response
Agonistadded
Endosomal receptor degradation
Receptor transcription inhibited
Receptor phosphorylationArrestin binding
Receptor endocytosis
G protein
Earlyendosome
Lysosome
Time (seconds)0 1 10 100 1000
Agonist binds
G proteinactive
EFFECTORS
Arrestin
GPCR
G P C R gene
1
2
3
5
4
5 Receptor transcription inhibited
C Y TO P L A S M
N U C L E U S
Agonist
Receptordegradation
4
Receptor endocytosis
3
Receptorrecycling
1 Receptorphosphorylation
Arrestin binding
2
Multiple adaptation processes occur after a stimulus
FIGURE 14.11 Multiple adaptation processes are invoked during a stimulus,and multiple nested mechanisms for adaptation are the rule. They are usuallyinvoked sequentially according to the duration and intensity of the stimulus.For GPCRs, at least five desensitizing mechanisms are known, with others act-ing on the G protein and effectors.
these points will alter the kinetics or extent ofadaptation (Figure 14.10). In branched pathways,changing these points can determine whetheradaptation is unique to one input or is exertedfor many similar inputs. If receptor activation trig-gers its desensitization directly, or if an eventdownstream on an unbranched pathway triggersdesensitization, then only signals that initiate withthat receptor will be altered. Receptor-selectiveadaptation is referred to as homologous adap-tation (Figure 14.10).
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 603

604 CHAPTER 14 Principles of cell signaling
Alternatively, feedback control can initiatedownstream from multiple receptors in a con-vergent pathway and thus regulate both theinitiating receptor and the others. Such het-erologous adaptation regulates all the possi-ble inputs to a given control point. A commonexample is the phosphorylation of G protein-coupled receptors by either protein kinase A orprotein kinase C, which are activated by down-stream signals cAMP or Ca2+ plus the lipid dia-cylglycerol, respectively. Like GRK, these kinasesboth attenuate receptor activity and promotearrestin binding.
Cells also alter their responses to incomingsignals for homeostatic reasons. These consid-erations include phase of the cell cycle, meta-bolic status, or other aspects of cellular activity.Again, all these adaptive processes may be dis-played to a greater or lesser extent in differentcells, different pathways within a cell or differ-ent situations during the cell’s lifetime.
Signaling proteins arefrequently expressed asmultiple species
Key concepts • Distinct species (isoforms) of similar signaling
proteins expand the regulatory mechanismspossible in signaling pathways.
• Isoforms may differ in function, susceptibility toregulation or expression.
• Cells may express one or several isoforms to fulfilltheir signaling needs.
14.11
Cells increase the richness, adaptability, andregulation of their signaling pathways by ex-pressing multiple species of individual signal-ing proteins that display distinct biochemicalproperties. These species may be encoded bymultiple genes or by multiple mRNAs derivedfrom a single gene by alternative splicing ormRNA editing. The numerical complexity im-plicit in these choices is impressive. Considerthe neurotransmitter serotonin: In mammals,there are thirteen serotonin receptors, each ofwhich stimulates a distinct spectrum of G pro-teins of the Gi, Gs, and Gq families. (A four-teenth serotonin receptor is an ion channel.)FIGURE 14.12 shows the relationship of serotoninreceptors to these G protein families.
There is also tremendous diversity amongthe G proteins and adenylyl cyclases. There arethree genes for Gαi and one each for the closelyrelated Gαz and Gαo. Furthermore, the GαomRNA is multiply spliced. There are four Gqmembers. In addition, there are five genes forGβ and twelve for Gγ, and most of the possibleGβγ dimers are expressed naturally. There areten genes for adenylyl cyclases, which are directtargets of Gs and either direct or indirect targetsof the other G proteins. While all nine mem-brane-bound adenylyl cyclase isoforms are stim-ulated by Gαs, they display diverse stimulatoryand inhibitory responses to Gβγ, Gαi, Ca2+,calmodulin, and several protein kinases, as il-lustrated in FIGURE 14.13. Thus, stimulation byserotonin can lead to diverse responses depend-ing upon the various forms of the proteins thatare engaged at a particular time and location.
FIGURE 14.12 Receptors for serotonin haveevolved in mammals as a family of 13 genes thatregulate three of the four major classes of G pro-teins. While all respond to the natural ligandserotonin, the binding sites have evolved suf-ficient differences that drugs have been devel-oped that specifically target one or moreisoforms. The type 3 serotonin receptors, notshown here, are ligand-gated ion channels andare not obviously related to the others.
1B
Gi
Gs
Gs
Gq
1D
1E
1F
1A
7
5A
5B
4
2A
2C
2B
6
120 100 80 60 40 20 0
Isoforms
Nucleotide substitution distance
G protein
Evolutionary relationship of serotonin receptor isoforms
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 604

14.12 Activating and deactivating reactions are separate and independently controlled 605
Sometimes isoforms of a signaling proteinare subject to quite different kinds of inputs.For example, all of the members of the phospho-lipase C family (PLC) hydrolyze phosphatidyli-nositol-4,5-bisphosphate to form two secondmessengers, diacylglycerol and inositol-1,4,5trisphosphate (see 14.16 Lipids and lipid-derivedcompounds are signaling molecules). The distinctisoforms may be regulated by diverse combina-tions of Gαq, Gβγ, phosphorylation, monomericG proteins, or Ca2+.
Because a cell has multiple options whenexpressing a form of a signaling protein, it canuse expression of particular isoforms to alterhow it performs otherwise identical signalingfunctions. Different cells express one or moreisoforms to allow appropriate responses, and ex-pression can vary according to other inputs orthe cell’s metabolic status. In addition, signalingpathways are remarkably resistant to mutationalor other injuries because loss of a single speciesor isoform of a signaling protein can often becompensated for by increased expression or ac-tivity of another species. Similarly, engineered
overexpression can result in the reduced expres-sion of endogenous proteins. The existence ofmultiple receptor species can, thus, substantiallyadd to adaptability and the consequent resist-ance of signaling networks to damage.
Activating anddeactivating reactions are separate andindependently controlled
In signaling networks, individual proteins arefrequently activated and deactivated by distinctreactions, a feature that facilitates separate reg-ulation. Common examples include using pro-tein kinases and phosphoprotein phosphatases
Key concepts • Activating and deactivating reactions are usually
executed by different regulatory proteins.• Separating activation and inactivation allows for
fine-tuned regulation of amplitude and timing.
14.12
CaM
CaMK
Gαs
Gαi
Gβγ
PKA
PKC
inhibit
activate
Regulators
Ca2+ NO
Different isoforms of adenylyl cyclase are regulated differently
FIGURE 14.13 All of the mammalian membrane-bound adenylyl cyclases arestructurally homologous and catalyze the same reaction, and all are stimulatedby G�s. Their responses to other inputs (protein kinases CaMK, PKA and PKC;Ca2+; calmodulin (CaM); NO•) are specific to each isoform, allowing a rich com-binatoric input to cellular cAMP signaling.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 605

606 CHAPTER 14 Principles of cell signaling
to catalyze protein phosphorylation and de-phosphorylation; using adenylyl cyclase to cre-ate cAMP while using phosphodiesterases tohydrolyze it or anion transporters to pump itout of the cell; or using GTP/GDP exchange fac-tors (GEFs) to activate G proteins and GTPase-activating proteins (GAPs) to deactivate them.Depending on stoichiometry and detailed mech-anism, these strategies can convey either addi-tive or nonadditive inputs while maintainingfine control over the kinetics of activation anddeactivation of a signaling pathway. The use ofdistinct reactions for activation and deactiva-tion is analogous to the use of distinct anabolicand catabolic enzymes in reversible metabolicpathways.
Cellular signaling usesboth allostery andcovalent modification
Cellular signaling uses almost every imagina-ble mechanism for regulating the activities ofintracellular proteins, but most can be describedas either allosteric or covalent. Individual sig-naling proteins typically respond to multiple al-losteric and covalent inputs.
Allostery refers to the ability of a moleculeto alter the conformation of a target proteinwhen it binds noncovalently to that protein.Because a protein’s activity reflects its confor-mation, the binding of any molecule that altersconformation can change the target protein’sactivity. Any molecule can have allosteric ef-fects: protons or Ca2+, small organic molecules,or other proteins. Allosteric regulation can beboth inhibitory or stimulatory.
Covalent modification of a protein’s chem-ical structure is also frequently used to regulateits activity. The change in the protein’s chemi-cal structure alters its conformation and, thus,its activity. Most regulatory covalent modifica-tion is reversible. The classic and most commonregulatory covalent event is phosphorylation,in which a phosphoryl group is transferred fromATP to the protein, most often to the hydroxylgroup of serine (Ser), threonine(Thr), or tyro-sine (Tyr). Enzymes that phosphorylate proteins
Key concepts • Allostery refers to the ability of a molecule to alter
the conformation of a target protein when it bindsnoncovalently to that protein.
• Modification of a protein’s chemical structure isalso frequently used to regulate its activity.
14.13
are known as protein kinases. Their actions areopposed by phosphoprotein phosphatases, whichcatalyze the hydrolysis of the phosphoryl groupto yield free phosphate and restore the unmod-ified hydroxyl residue. Other forms of covalentmodification are also common and will be ad-dressed throughout the chapter.
Second messengersprovide readily diffusiblepathways for informationtransfer
Signaling pathways make use of both proteinsand small molecules according to their distinc-tive attributes. A small molecule used as an in-tracellular signal, or second messenger, has anumber of advantages over a protein as a sig-naling intermediary. Small molecules can besynthesized and destroyed quickly. Because theycan be made readily, they can act at high con-centrations so that their affinities for target pro-teins can be low. Low affinity permits rapiddissociation, such that their signals can be ter-minated promptly when free second messengermolecules are destroyed or sequestered. Becausesecond messengers are small, they also can dif-fuse quickly within the cell, although many cellshave developed mechanisms to spatially restrictsuch diffusion. Second messengers are, thus,superior to proteins in mediating fast responses,particularly at a distance. Second messengersare also useful when signals have to be addressedto large numbers of target proteins simultane-ously. These advantages often overcome theirlack of catalytic activity and their inability tobind multiple molecules simultaneously.
FIGURE 14.14 lists intracellular second mes-sengers developed through evolution. This num-ber is surprisingly low. Several are nucleotidessynthesized from major metabolic nucleotideprecursors. They include cAMP, cyclic GMP,ppGppp, and cyclic ADP-ribose. Other solublesecond messengers include a sugar phosphate,inositol-1,4,5-trisphosphate (IP3), a divalent metalion Ca2+, and a free radical gas nitric oxide (NO•).Lipid second messengers include diacylglyceroland phosphatidylinositol-3,4,5-trisphosphate,
Key concepts • Second messengers can propagate signals between
proteins that are at a distance.• cAMP and Ca2+ are widely used second messengers.
14.14
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 606
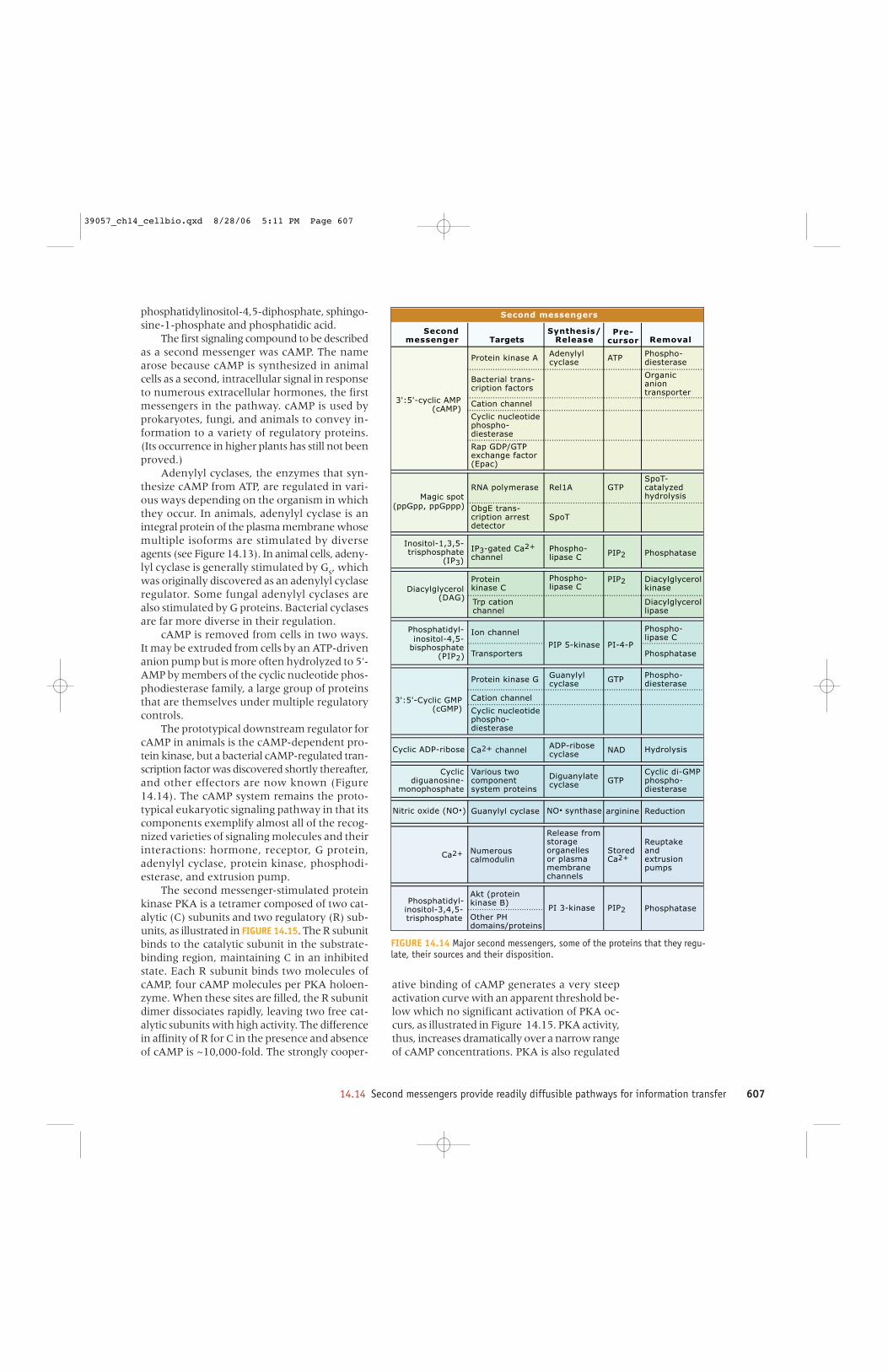
14.14 Second messengers provide readily diffusible pathways for information transfer 607
phosphatidylinositol-4,5-diphosphate, sphingo-sine-1-phosphate and phosphatidic acid.
The first signaling compound to be describedas a second messenger was cAMP. The namearose because cAMP is synthesized in animalcells as a second, intracellular signal in responseto numerous extracellular hormones, the firstmessengers in the pathway. cAMP is used byprokaryotes, fungi, and animals to convey in-formation to a variety of regulatory proteins.(Its occurrence in higher plants has still not beenproved.)
Adenylyl cyclases, the enzymes that syn-thesize cAMP from ATP, are regulated in vari-ous ways depending on the organism in whichthey occur. In animals, adenylyl cyclase is anintegral protein of the plasma membrane whosemultiple isoforms are stimulated by diverseagents (see Figure 14.13). In animal cells, adeny-lyl cyclase is generally stimulated by Gs, whichwas originally discovered as an adenylyl cyclaseregulator. Some fungal adenylyl cyclases arealso stimulated by G proteins. Bacterial cyclasesare far more diverse in their regulation.
cAMP is removed from cells in two ways.It may be extruded from cells by an ATP-drivenanion pump but is more often hydrolyzed to 5′-AMP by members of the cyclic nucleotide phos-phodiesterase family, a large group of proteinsthat are themselves under multiple regulatorycontrols.
The prototypical downstream regulator forcAMP in animals is the cAMP-dependent pro-tein kinase, but a bacterial cAMP-regulated tran-scription factor was discovered shortly thereafter,and other effectors are now known (Figure14.14). The cAMP system remains the proto-typical eukaryotic signaling pathway in that itscomponents exemplify almost all of the recog-nized varieties of signaling molecules and theirinteractions: hormone, receptor, G protein,adenylyl cyclase, protein kinase, phosphodi-esterase, and extrusion pump.
The second messenger-stimulated proteinkinase PKA is a tetramer composed of two cat-alytic (C) subunits and two regulatory (R) sub-units, as illustrated in FIGURE 14.15. The R subunitbinds to the catalytic subunit in the substrate-binding region, maintaining C in an inhibitedstate. Each R subunit binds two molecules ofcAMP, four cAMP molecules per PKA holoen-zyme. When these sites are filled, the R subunitdimer dissociates rapidly, leaving two free cat-alytic subunits with high activity. The differencein affinity of R for C in the presence and absenceof cAMP is ~10,000-fold. The strongly cooper-
ative binding of cAMP generates a very steepactivation curve with an apparent threshold be-low which no significant activation of PKA oc-curs, as illustrated in Figure 14.15. PKA activity,thus, increases dramatically over a narrow rangeof cAMP concentrations. PKA is also regulated
Protein kinase A
Bacterial trans-cription factors
Cation channel
Cyclic nucleotidephospho-diesterase
Rap GDP/GTPexchange factor(Epac)
RNA polymerase
ObgE trans-cription arrestdetector
IP3-gated Ca2+
channel
Proteinkinase C
Trp cationchannel
Ion channel
Transporters
Protein kinase G
Cation channel
Cyclic nucleotidephospho-diesterase
Ca2+ channel
Various twocomponentsystem proteins
Guanylyl cyclase
Numerouscalmodulin
Akt (proteinkinase B)
Other PHdomains/proteins
Adenylylcyclase
Rel1A
SpoT
Phospho-lipase C
Phospho-lipase C
PIP 5-kinase
PI 3-kinase
Guanylylcyclase
ADP-ribosecyclase
Diguanylatecyclase
NO. synthase
ATP
GTP
PIP2
PIP2
PI-4-P
GTP
NAD
GTP
arginine
StoredCa2+
PIP2
Phospho-diesterase
Organicaniontransporter
SpoT-catalyzedhydrolysis
Phosphatase
Diacylglycerolkinase
Diacylglycerollipase
Phospho-lipase C
Phosphatase
Phospho-diesterase
Hydrolysis
Cyclic di-GMP phospho-diesterase
Reduction
Reuptakeandextrusionpumps
Phosphatase
Magic spot
3':5'-cyclic AMP(cAMP)
(ppGpp, ppGppp)
Inositol-1,3,5-trisphosphate
(IP3)
Diacylglycerol(DAG)
Phosphatidyl-inositol-4,5-
bisphosphate(PIP2)
3':5'-Cyclic GMP (cGMP)
Cyclic ADP-ribose
Cyclicdiguanosine-
monophosphate
Nitric oxide (NO.)
Ca2+
Phosphatidyl-inositol-3,4,5-trisphosphate
Secondmessenger
Synthesis/ReleaseTargets
Pre-cursor Removal
Release fromstorage organellesor plasma membrane channels
Second messengers
FIGURE 14.14 Major second messengers, some of the proteins that they regu-late, their sources and their disposition.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 607

608 CHAPTER 14 Principles of cell signaling
by phosphorylation of its activation loop.Phosphorylation occurs cotranslationally, andthe activation loop phosphorylation is requiredfor assembly of the R2C2 tetramer.
The PKAs are mostly cytosolic and are alsotargeted to specific locations by binding or-ganelle-associated scaffolds (A-kinase anchor-ing proteins, or AKAPs). These AKAPs facilitatephosphorylation of membrane proteins includ-ing GPCRs, transporters, and ion channels.AKAPs can also target PKA to other cellular lo-cations including mitochondria, the cytoskele-ton, and the centrosome. AKAPs often harborbinding sites for other regulatory moleculessuch as phosphoprotein phosphatases and ad-ditional protein kinases, which allows for co-ordination of multiple signaling pathways andintegration of their outputs.
PKA generally phosphorylates substrateswith a primary consensus motif of Arg-Arg-Xaa-Ser-Hydrophobic, placing it in a large groupof kinases that recognize basic residues preced-ing the phosphorylation site. PKA regulates pro-
teins throughout the cell ranging from ion chan-nels to transcription factors, and its conservedsubstrate preference frequently permits predic-tion of substrates by sequence analysis. ThecAMP response element binding protein CREBis phosphorylated by PKA on Ser 133 and islargely responsible for the impact of cAMP ontranscription of numerous genes.
Ca2+ signaling servesdiverse purposes in alleukaryotic cells
Ca2+ is used as a second messenger in all cells,and is, thus, an even more widespread secondmessenger than cAMP. Many proteins bind Ca2+
with consequent allosteric changes in their en-zymatic activities, subcellular localization, orinteraction with other proteins or with lipids.Direct targets of Ca2+ regulation include almostall classes of signaling proteins described in thischapter, numerous metabolic enzymes, ionchannels and pumps, and contractile proteins.Most noteworthy may be muscle actomyosinfibers, which are triggered to contract in re-sponse to cytosolic Ca2+ (see 8.21 Myosin-II func-tions in muscle contraction).
Although free Ca2+ is found at concentra-tions near 1 mM in most extracellular fluids, in-tracellular Ca2+ concentrations are maintainednear 100 nanomolar levels by the combined ac-tion of pumps and transporters that either ex-trude free Ca2+ or sequester it in the endoplasmicreticulum or mitochondria. Ca2+ signaling is ini-tiated when Ca2+-selective channels in the en-doplasmic reticulum or plasma membrane areopened to allow Ca2+ to enter the cytoplasm.The most important entrance channels includeelectrically gated channels in animal plasmamembranes; a Ca2+ channel in the endoplasmicreticulum that is opened by another second mes-senger, inositol 1,4,5-trisphosphate (see below);and an electrically gated channel in the endo-plasmic (sarcoplasmic) reticulum of musclethat opens in response to depolarization of nearbyplasma membrane, a process known as excita-tion-contraction coupling (see 2.9 Plasma mem-
Key concepts • Ca2+ serves as a second messenger and regulatory
molecule in essentially all cells.• Ca2+ acts directly on many target proteins and also
regulates the activity of a regulatory proteincalmodulin.
• The cytosolic concentration of Ca2+ is controlled byorganellar sequestration and release.
14.15
[cAMP]
Kinase activity as a function of [cAMP] (%)
100
80
60
40
20
2 x 10-9 2 x 10-8 2 x 10-7
10%
90%
(R) R egulatory subunits
4 cAMP(C)Catalyticsubunits
CR
RC
- cAMP- cAMP
- cAMP- cAMP
C R
R C
R2C2 + 4 cAMP R2 . cAMP4 + 2C
PKAActivated
PKA
Activation of PKA by cAMP
FIGURE 14.15 PKA is a heterotetramer composed of two catalytic (C) andtwo regulatory (R) subunits. Binding of four molecules of cAMP to the reg-ulatory subunits induces dissociation of two molecules of C, the active formof PKA, from the cAMP-bound regulatory subunit dimer. In the bottom panel,the cooperative binding of four molecules of cAMP generates a steep acti-vation profile. Activity increases from approximately 10% to 90% as thecAMP concentration increases only 10-fold. An apparent threshold is intro-duced because there is little change in activity at low concentrations ofcAMP.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 608

14.16 Lipids and lipid-derived compounds are signaling molecules 609
brane Ca2+ channels activate intracellular functions).In addition to the proteins that are regulated
by binding Ca2+ directly, many other proteins re-spond to Ca2+ by binding a widespread Ca2+ sen-sor, the small, ~17 kDa protein calmodulin.Calmodulin requires the binding of four mole-cules of Ca2+ to become fully active, and bind-ing is highly cooperative, generating a sigmoidactivation profile illustrated in FIGURE 14.16.Calmodulin generally binds its targets in a Ca2+-dependent manner, but Ca2+-free calmodulinmay remain bound but inactive in some cases.For example, calmodulin is a constitutive sub-unit of phosphorylase kinase that is activatedupon Ca2+ binding. Higher plants again makemajor modifications to this paradigm. Calmodulinis not expressed as a distinct protein but, instead,is found as a domain in Ca2+-regulated proteins.In yet another variation, the adenylyl cyclase se-creted by the pathogenic bacterium Bordetella per-tussis is inactive outside cells but is activated byCa2+-free calmodulin in animal cells, where itsrapid production of cAMP is highly toxic.
Lipids and lipid-derivedcompounds are signalingmolecules
Signals that originate at the plasma membranemay have soluble regulatory targets in the cy-toplasm or intracellular organelles, but integralplasma membrane proteins are also subject toacute controls. For these targets, lipid secondmessengers may be primary inputs. Lipids de-rived from membrane phospholipids or other
Key concepts • Multiple lipid-derived second messengers are
produced in membranes.• Phospholipase Cs release soluble and lipid second
messengers in response to diverse inputs.• Channels and transporters are modulated by
different lipids in addition to inputs from othersources.
• PI 3-kinase synthesizes PIP3 to modulate cellshape and motility.
• PLD and PLA2 create other lipid secondmessengers.
14.16
Calcium-freecalmodulin
calmodulinfree + 4 Ca2+ (Ca2+)4 . calmodulin . active target
Calcium-bound calmodulin bound to
target peptide of CaMK
Ca2+ target
Calcium binding causes a conformational change in calmodulin
100
80
60
40
20
3 x 10-8 3 x 10-7 3 x 10-6
10%
90%
[Ca2+]
Activation of targetby calmodulin (%)
FIGURE 14.16 Ribbon diagrams represent-ing the crystal structures of calmodulin freeof Ca2+ and bound to four Ca2+ ions revealthe huge conformational change thatcalmodulin undergoes upon Ca2+ binding.Ca2+-calmodulin causes activity changes intarget proteins. The bottom panel showsthe activation of a target by calmodulin asa function of the intracellular free Ca2+ con-centration. The requirement for bindingfour Ca2+ ions to induce the conformationaltransition results in cooperative activationof targets. Activity increases from 10% to90% as the Ca2+ concentration increasesonly 10-fold. Structures generated fromProtein Data Bank files 1CFD and 1MXE.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 609

610 CHAPTER 14 Principles of cell signaling
lipid species play numerous roles in cell signal-ing. Because their analysis has been more dif-ficult than for soluble messengers, manyprobably remain to be discovered and under-stood. FIGURE 14.17 shows the structure of someof these lipids.
Phospholipase Cs (PLCs) are the prototyp-ical lipid signaling enzymes. PLC isoforms cat-alyze the hydrolysis of phospholipids betweenthe 3-sn-hydroxyl and the phosphate group toyield a diacylglycerol and phosphate ester. Inanimals and fungi, PLCs specific for the substrate
23
4 56 OH
1
OH
OH
O
O
O
O
OP
O
O-
23
4 56 OH
1
OH
O
O
O
O
O
O
HOOH
OH
OP
O
O-
O-
O
O
O
O
OP
O
O-
O
O
O
O
OPO3H-
OPO3H-
OPO3H-
OPO3H-H-O3PO
HO
23
4 56 OH
1
OH
OPO3H-
Phosphatidic acid (PA)
Diacylglycerol (DAG)
Phosphatidylinositol (PI)
Phosphatidylinositol-3,4,5-trisphosphate (PIP3)
Inositol trisphosphate (IP3)
Structures of some lipid second messengers
FIGURE 14.17 Structures of some lipid second messengers and the common precursor phosphatidylinositol.The acyl side chain structures shown here are the most common for mammalian PI lipids. Much of the PA incells is derived from PC, and its acyl chains may differ from those shown.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 610

14.16 Lipids and lipid-derived compounds are signaling molecules 611
phosphatidylinositol-4,5-bisphosphate (PIP2)hydrolyze PIP2 to form two second messengers:1,2-sn-diacylglycerol (DAG) and inositol-1,4,5-trisphosphate (IP3). The PLC substrate PIP2 is it-self an important regulatory ligand thatmodulates the activity of several ion channels,transporters, and enzymes. Thus, PLC alters con-centration of three second messengers; its neteffect depends on the net turnover of the sub-strate and products.
DAG is probably the best known lipid sec-ond messenger; its hydrophobicity limits it to ac-tion in membranes. DAG activates some isoformsof protein kinase C (PKC), modulates the ac-tivity of several cation channels and activates atleast one other protein kinase. DAG can be fur-ther hydrolyzed to release arachidonic acid,which can regulate some ion channels.Arachidonic acid is also the precursor of oxida-tion products, such as prostaglandins and throm-boxanes, which are potent extracellular signalingagents. In addition to DAG, PKCs require inter-action with Ca2+ and an acidic phospholipid,such as phosphatidylserine, to become activated.Thus, activation of PKC requires the coincidenceof multiple inputs both to generate DAG and toincrease intracellular Ca2+. There are more thana dozen PKCs, classified together according tohighly conserved sequences in the catalytic do-main. Three subgroups of PKCs, also identifiableby sequence, share different patterns of regu-lation. Their regulation provides examples ofmany ways in which other mammalian proteinkinases are regulated.
The first of these groups, canonical PKCs,are generally soluble or very loosely associatedwith membranes prior to the appearance ofDAG. DAG causes their association with mem-branes and permits activation upon binding ofother regulators. The second group of PKCs re-quires similar lipids but not Ca2+, and the thirdgroup requires other lipids but neither DAG norCa2+ for activation.
The N-terminal region of PKCs contains apseudosubstrate domain, a sequence that re-sembles that of a typical substrate except thatthe target Ser is replaced with Ala. The pseu-dosubstrate region binds to the active site to in-hibit the kinase. Activators cause thepseudosubstrate domain to flip out of the ac-tive site. PKCs are also activated by proteoly-sis, as are many protein kinases with discreteautoinhibitory domains. Proteases clip a flex-ible hinge region, which results in loss of theregulatory domain and consequent activationof the kinase.
PKC is the major receptor for phorbol esters,a class of powerful tumor promoters. Phorbolesters mimic DAG and cause a more massiveand prolonged activation than physiologicalstimuli. This massive stimulation can induceproteolysis of PKC, resulting in downregula-tion, or loss of the kinase. (For a personal de-scription on the discovery of protein kinase Csee )
IP3, the second product of the PLC reaction,is a soluble second messenger. The most signif-icant IP3 target is a Ca2+ channel in the endo-plasmic reticulum. IP3 causes this channel toopen and release stored Ca2+ into the cytoplasm,thereby rapidly elevating the cytosolic Ca2+ over100-fold and, in turn, causing the activation ofnumerous targets of Ca2+ signaling.
There are at least six families of PIP2-selec-tive PLC enzymes, defined by their distinct formsof regulation, domain compositions, and over-all sequence conservation. Their catalytic do-mains are all quite similar. The PLC-βs arestimulated primarily by Gαq and Gβγ (to individ-ually varying extents). Several are also modu-lated by phosphorylation. PLC-γ isoforms arestimulated by phosphorylation on Tyr residues,frequently by receptor tyrosine kinases. ThePLC-ε isoforms are regulated by small,monomeric G proteins of the Rho family. Theregulation of the PLC-δs is still incompletely un-derstood. Two other classes similar to the PLC-δs, PLC-η and -ζ, have also been defined recently.(There is no PLC-α.) In addition to their distinctmodes of regulation, all of the PLCs are stimu-lated by Ca2+, and Ca2+ often acts synergisti-cally with other stimulatory inputs. This synergyunderlies the intensification and prolongationof Ca2+ signaling observed in many cells.
Phospholipases A2 and D (PLA2 and PLD)also hydrolyze glycerol phospholipids in cellmembranes to form important signaling com-pounds. PLA2 hydrolyzes the fatty acid at the sn-2 position of multiple phospholipids to producethe cognate lysophospholipid and the free fattyacid, which is generally unsaturated. The freefatty acid is often arachidonic acid, a precursorof extracellular signals. The biological roles offree lysophospholipids are not understood indetail but have been linked to effects on thestructure of the membrane bilayer.
PLD catalyzes a reaction much like that ofPLC but instead hydrolyzes the phosphodiesteron the substituent side of the phosphate groupto form 3-sn-phosphatidic acid. Cellular PLDs acton multiple glycerol phospholipid substrates,but phosphatidylcholine is probably the sub-
EXP : 14-0001
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 611

612 CHAPTER 14 Principles of cell signaling
strate most relevant to signaling functions. Thefunctions of the phosphatidic acid product,which is also formed by phosphorylation ofDAG, remain poorly understood but appear toinclude a role in secretion and the fusion of in-tracellular membranes.
PI 3-kinase regulatesboth cell shape and theactivation of essentialgrowth and metabolicfunctions
Lipid second messengers may also be modifiedby phosphorylation. PI 3-kinase phosphorylatesPIP2 on the 3-position of the inositol ring toform PI 3,4,5-P3, another lipid second messen-ger. The total activity of PI 3-kinase is too lowto significantly deplete total PIP2, but forma-tion of small amounts of PIP3 in localized mem-brane domains is vital for altering cell shapeand cellular motility.
PIP3 acts by recruiting proteins that con-tain PIP3 binding domains, including pleckstrinhomology (PH) and FYVE domains, to siteswhere they regulate cytoskeletal remodeling,contractile protein function, or other regula-tory events. These proteins anchor and/or ori-ent the structural or motor proteins involvedin cellular movement and localize signaling pro-teins to sites of action at the membrane. PIP3
Key concepts • Phosphorylation of some lipid second messengers
changes their activity.• PIP3 is recognized by proteins with a pleckstrin
homology domain.
14.17
signaling can be fast and dramatic; it largely ac-counts for directing the mobility of motile mam-malian cells.
Lipid mediators are essential in the insulinsignaling pathway. The binding of insulin stimu-lates the Tyr autophosphorylation of its receptorand the activation of effectors through insulin re-ceptor substrate (IRS) proteins (see 14.30 Diversesignaling mechanisms are regulated by protein tyro-sine kinases). PI 3-kinase is activated when its p85subunit binds to IRS1. The PIP3 generated by PI3-kinase binds the protein kinases Akt and phospho-inositide-dependent kinase-1 (PDK-1) via their PHdomains. This interaction results in the localiza-tion of Akt to the membrane where it is activatedby PDK1, as illustrated in FIGURE 14.18. Akt phos-phorylates downstream targets, including pro-tein kinases, GAPs, and transcription factors.Activation of Akt, specifically Akt-2, is requiredfor the hallmark actions of insulin including reg-ulation of glucose transporter translocation, en-hanced protein synthesis, and expression ofgluconeogenic and lipogenic enzymes.
Signaling through ionchannel receptors is veryfast
Ligand-gated ion channels are multisubunit,membrane-spanning proteins that create andregulate a water-filled pore through the mem-brane, as illustrated in the X-ray crystal struc-ture of the nicotinic acetylcholine receptor inFIGURE 14.19. When stimulated by extracellularagonists, the subunits rearrange their conforma-tions and orientations to open the pore and, thus,connect the aqueous spaces on either side of themembrane. The pore has a diameter that allowsions to diffuse freely from one side of the mem-brane to the other, driven by the electrical andchemical gradients that have been establishedby ion pumps and transporters. (For more aboutchannel, pump and transporter mechanics see 2Transport of ions and small molecules across mem-branes.) Channels maintain selectivity amongions by regulating the pore diameter precisely
Key concepts • Ion channels allow the passage of ions through a
pore, resulting in rapid (microsecond) changes inmembrane potential.
• Channels are selective for particular ions or forcations or anions.
• Channels regulate intracellular concentrations ofregulatory ions, such as Ca2+.
14.18
PIP2 PIP3
p85 p110
Akt and PDK1 bind PIP
PI 3-kinase
3 through PH domains
Akt is activated by phosphorylation
PHdomains Other
kinase
Akt PDK1
Akt PDK1
PIP2 phosphorylated
Glucose uptakeGlycogen synthesisAntilipolysisAntiapoptosis
PIP3 binding brings Akt and PDK1 to the membrane
FIGURE 14.18 Activated PI 3-kinase phosphorylates PIP2 to pro-duce PIP3. The PH domain-contain-ing protein kinases PDK1 and Aktbind to PIP3 at the plasma mem-brane. Their colocalization facili-tates the phosphorylation of Aktby PDK1. A second phosphoryla-tion within a hydrophobic motif re-sults in Akt activation by one ofseveral candidate protein kinases.The Akt-2 isoform is required toelicit hallmark actions of insulin.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 612

14.18 Signaling through ion channel receptors is very fast 613
and by lining the walls of the pore with appro-priate hydrophilic residues. Receptor ion chan-nels can, thus, provide a diffusion path for onlycations or anions, or select among different ions.
Ligand-gated ion channels provide thefastest signal transduction mechanism found inbiology. Upon binding an agonist ligand, chan-nels open within microseconds. At synapses,where neurotransmitters need to diffuse lessthan 0.1 micron, a signal in the postsynapticcell can be generated in 100 microseconds. Incontrast, receptor-stimulated G proteins requireabout 100 milliseconds to exchange GDP forGTP, and the action of receptor protein kinasesis even slower. Ligand-gated ion channels areimportant receptors in many cells in additionto neurons and muscle, and other ion channelsplay equally vital roles in signaling pathwaystriggered by other classes of ligands.
Ion channel signaling differs from that ofthe other receptors mentioned in this chapterin that there is no immediate protein target nor,in most cases, is there a specific second messen-ger involved. In most cases, channel-mediatedion flow acts to increase or decrease the cell’smembrane potential and, thus, modulates alltransport processes for metabolites or ions thatare electrically driven.
Animal cells maintain an inside-negativemembrane potential by pumping out Na+ ionsand pumping in K+ ions (for more on mem-brane potential see 2.4 Electrochemical gradientsacross the cell membrane generate the membrane po-tential). The opening of a channel selective forNa+ will thus depolarize cells, and the openingof a channel for K+ will hyperpolarize cells.Similarly, because Cl- is primarily extracellular,opening Cl- channels will also cause hyperpo-larization. These electrical effects convey infor-mation to effector proteins that are energeticallycoupled to the membrane potential, or to spe-cific ion gradients, or that bind a specific ion(such as Ca2+) whose concentration changesupon channel opening.
The nicotinic acetylcholine receptor is the pro-totypical receptor ion channel and was the firstreceptor that was shown to be a channel. It is a rel-atively unselective cation channel that causes de-polarization of the target cell by allowing Na+
influx. It is best known as the excitatory receptorat the neuromuscular synapse, where it triggerscontraction, but alternative isoforms are also ac-tive in neurons and many other cells. In muscles,nicotinic depolarization acts via a voltage-sensitiveCa2+ channel to allow Ca2+ release from the sar-coplasmic reticulum into the cytosol. Calcium acts
Pore
CYTOSOL
CLOSED OPEN
Pore
Nicotinic acetylcholine receptor structure
FIGURE 14.19 The nicotinic cholinergic receptor is a cation-selective channelthat is composed of five homologous but usually nonidentical subunits thatoligomerize to form a primarily �-helical membrane-spanning core. The chan-nel itself is created within this core, and its opening and closing are executedby cooperative changes in subunit arrangement. Structure generated fromProtein Data Bank file 2BG9.
as a second (or third) messenger to initiate con-traction (see 2.13 Cardiac and skeletal muscles are ac-tivated by excitation-contraction coupling). Nicotinicreceptors promote exocytosis in some secretorycells by a similar mechanism, where Ca2+ triggersthe exocytic event. In neurons, where nicotinicstimulation causes an action potential (depo-larization that is rapidly propagated along the neu-ron), the initial depolarization is sensed byvoltage-sensitive Na+ channels. Their opening(along with the action of other channels) propa-gates the action potential along the neuron.
The nervous system is rich in receptor cationchannels that respond to other neurotransmit-ters, the most common of which is the aminoacid glutamate (Glu). The three different fam-ilies of glutamate receptors share the propertyof cation conductance, but each family has itsown spectrum of drug responses. All operate asneuronal activators, with one interesting twist:The NMDA family of receptors, named for theirresponse to a selective drug, is permeant to Ca2+
in addition to Na+. A significant component ofits activity is to permit the inward flow of Ca2+,which acts as a second messenger on a wide va-riety of targets. Persistant stimulation of NMDAchannels by glutamate released during injury,or by drugs, can cause toxic amounts of Ca2+ toenter, resulting in neuronal death.
A second functional group of receptor chan-nels is selective for anions and, by allowing in-ward flux of Cl-, hyperpolarizes the target cell.Anion-selective receptors include those for γ-aminobutyric acid (GABA) and glycine (Gly). Inneurons, hyperpolarization can inhibit the ini-tiation of an action potential and/or neurotrans-mitter release.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 613

614 CHAPTER 14 Principles of cell signaling
Perhaps the most diverse family of ligand-gated channels is that of the TRP and TRP-likefamily, of which about 30 have been found inmammals. Distinct forms are found in inverte-brates. The TRP channels are Ca2+-selectivechannels that are formed by tetramers of iden-tical subunits that surround the central chan-nel. Each subunit is composed of a homologousbundle of six membrane-spanning helices, butthe N and C termini contain a diverse collec-tion of regulatory and protein interaction do-mains, including protein kinase domains (whosesubstrates are currently unknown).
All TRP channels allow transmembrane fluxof Ca2+ to permit its action as a second messen-ger, but different TRP isoforms serve numerousphysiological functions. The prototypical TRP,found in invertebrate photoreceptors, gates Ca2+
flow from intracellular stores into the cytoplasmto initiate visual signaling. Others admit Ca2+
from outside the cell, and still others allow Ca2+
to enter the endoplasmic reticulum virtually di-rectly from the extracellular space because theyform a bridge between the plasma membraneand channels in the endoplasmic reticulum atpoints where the membranes abut each other.
Regulation of TRP channels is perhaps evenmore diverse. Various TRP channels respond toheat, cold, painful stimuli, pressure, and highor low osmolarity. Many TRPs are regulated ei-ther positively or negatively by lipids, such aseicosanoids, diacylglycerol, and PIP2. For ex-ample, capsaicin, the hot compound in chilis, isan agonist for some vanilloid receptors (TRPVs).Still other TRP channels are mechanosensorsthat allow cilia to sense fluid flow. The most fa-mous of these is the sensory channel of the haircell of the inner ear. This channel opens whenthe apical cilia on the hair cell are bent in re-sponse to sound-driven fluid flow.
Nuclear receptorsregulate transcription
Nuclear receptors are unique among cellular
Key concepts • Nuclear receptors modulate transcription by
binding to distinct short sequences inchromosomal DNA known as response elements.
• Receptor binding to other receptors, inhibitors, orcoactivators leads to complex transcriptionalcontrol circuits.
• Signaling through nuclear receptors is relativelyslow, consistent with their roles in adaptiveresponses.
14.19
receptors in that their ligands pass unaidedthrough the plasma membrane. These recep-tors, when complexed with their ligands, en-ter the nucleus and regulate gene transcription.Ligands for nuclear receptors include sex steroids(estrogen and testosterone) and other steroidhormones, vitamins A and D, retinoids andother fatty acids, oxysterols, and bile acids.
Nuclear receptors are structurally conserved.They consist of a C-terminal ligand binding do-main, an N-terminal interaction region that rec-ognizes components of the transcriptionalmachinery and acts as a transactivation domain,a centrally located zinc finger domain that bindsDNA, and, often, another transactivation do-main nearer the C-terminus. In the absence ofligand, these receptors are bound to corepres-sor proteins that suppress their activity. Uponhormone binding, corepressors dissociate andthe receptors are assembled in multiproteincomplexes with coactivators that modulate re-ceptor action and facilitate transcriptional reg-ulation. As illustrated in FIGURE 14.20, agonistsand antagonists bind to distinct receptor confor-mations (see 14.5 Ligand binding changes receptorconformation). Receptor agonists favor the bind-ing of receptors to coactivators and DNA, andantagonists favor conformations that block coac-tivator-receptor binding.
Nuclear receptors bind with high specificityto hormone response elements in the 5’ un-transcribed region of regulated genes. Responseelements are typically short direct or invertedrepeat sequences, and a gene may contain re-sponse elements for several different receptorsin addition to binding sites for other transcrip-tional regulatory proteins.
The sex steroid estrogen can bind to twodifferent nuclear receptors, the estrogen recep-tors ER� and ER�. Coactivator and corepressorproteins differentially regulate ER� and ER� intranscriptional complexes that are expressed inspecific cell types. Other ligands that bind tothese receptors include valuable therapeuticagents. For example, 4 hydroxy-tamoxifen isan estrogen receptor antagonist used in the ther-apy of estrogen-receptor-positive breast cancerto inhibit growth of residual cancer cells.However, unlike its antagonistic effects on theestrogen receptor in breast, 4 hydroxy-tamox-ifen displays weak partial agonist activity inuterus. In the estrogen receptor system, partialagonists are known as selective estrogen recep-tor modulators (SERMs). Properties that con-tribute to partial agonist activity include therelative expression of the two estrogen recep-
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 614

14.20 G protein signaling modules are widely used and highly adaptable 615
tors, ER� and ER�, as well as the expression ofrepressors and coactivators that interact witheach receptor type. Thus, the behavior of nu-clear receptor ligands must be considered in thetissue, cellular, and signaling context.
G protein signalingmodules are widely usedand highly adaptable
Activation of G protein-coupled receptors(GPCRs) and their associated heterotrimeric Gproteins is one of the most widespread mecha-nisms of communicating extracellular signalsto the intracellular environment. G protein sig-naling modules are found in all eukaryotes.Depending on the species, mammals express500-1000 GPCRs that respond to hormones,neurotransmitters, pheromones, metabolites,
Key concepts • The basic module is a receptor, a G protein and an
effector protein.• Cells express several varieties of each class of
proteins.• Effectors are heterogeneous and initiate diverse
cellular functions.
14.20
local signaling substances, and other regulatorymolecules. Essentially all chemical classes arerepresented among the GPCR ligands. In addi-tion, a roughly equal number of olfactory GPCRsare expressed in olfactory neurons and work incombination to screen compounds in the ani-mal’s environment via the sense of smell.Because GPCRs are involved in many kinds ofphysiologic responses, they are also one of themost widely used targets for drugs.
A minimal G protein signaling module con-sists of three proteins: a G protein-coupled re-ceptor, the heterotrimeric G protein, and aneffector protein, as illustrated in FIGURE 14.21. Thereceptor activates the G protein on the inner faceof the plasma membrane in response to an ex-tracellular ligand. The G protein then activates (oroccasionally inhibits) an effector protein thatpropagates a signal within the cell. Thus, signalconduction in the simplest G protein module islinear. However, as depicted in FIGURE 14.22, atypical animal cell may express a dozen GPCRs,more than six G proteins, and a dozen effectors.Each GPCR regulates one or more G proteins,and each G protein regulates several effectors.Moreover, distinct efficiencies and rates governeach interaction. Thus, a cell’s G protein networkis actually a signal-integrating computer whose
545
545
542
542
538
538
Agonist-boundconformation
Antagonist-boundconformation
NN
H11
H11
H5 H5
H6
H6H3
H3
CK362
K362
Estrogen receptor conformation depends on which ligand is bound
FIGURE 14.20 The estrogen receptor adopts different conformations whenbound to agonists and antagonists. The ligand-binding domain of the estro-gen receptor is bound to the agonist estradiol on the left and to the antago-nist raloxifene on the right. Note the marked difference in position of helix 12,shown in blue in the active structure and green in the inhibited structure.Reproduced from Brzozowski, A. M., et al. 1997. Molecular basis of agonism andantagonism in the oestrogen receptor. Nature. 389: 753–758. Photo courtesyof M. Brzozowski, University of New York.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 615

616 CHAPTER 14 Principles of cell signaling
output is a spectrum of cellular signals that iscomplex in both amplitude and kinetics. Becauseof their conserved parts list, G protein modulesare well suited to initiating a wide variety of in-tracellular signals in response to diverse molec-ular inputs and can do so over a wide range oftime scales (milliseconds to minutes).
GPCRs are integral plasma membrane pro-teins composed of a bundle of seven hydropho-bic membrane-spanning helices with anextracellular N terminus and cytosolic C termi-nus, as depicted in FIGURE 14.23. Based on thethree-dimensional structure of rhodopsin andon copious biochemical and genetic data, it islikely that all GPCRs share the same basic mech-anism of conformational activation and deacti-vation in response to activating ligands (see 14.5Ligand binding changes receptor conformation).Binding of agonist ligand on the extracellularface of the receptor drives realignment of the he-lices to alter the structure of a binding site for theheterotrimeric G protein on the cytoplasmic face,and this altered conformation of the G protein-binding surface promotes G protein activation.
Ca2+
Ca2+pump
Agonist C5a ISO PGE S1P UDP UTP PAF LPA
GPCR
G Protein
Effector
E2RC5aR EDG EDGP2YR P2YR PAFR
Gi Gs GqG12 G12
IP3 IP2 + Pi
PIP2
PIP3
AMP
PDE
??ATP
cAMP
cAMP
DAG +
GRK
IP3R
β2AR
Phos-phatase
PI 3-Kinase PLC-β
Inactivationmechanisms
AdCyc
Partial G protein signaling network in mouse macrophagesFIGURE 14.22 A portion of the G pro-tein-mediated signaling network inmacrophages highlights some of thecomplexity of interactions possible insuch systems. Several receptors and Gprotein subunits are omitted. Where anamed G protein is shown, its signalingoutput is probably mediated by its G�subunit. Activation of any G protein alsoactivates its G�� subunit, although G��-mediated signaling is usually most promi-nent from Gi trimers. In addition, severalG proteins modulate the activities ofothers through poorly understood path-ways. Only a small sampling of effectorsis shown, and the only adaptive mech-anism shown is GRK-catalyzed phospho-rylation of receptors. Data from PaulSternweis, Alliance for Cellular Signaling.
Ca2+
CYTOSOL
ENDOPLASMICRETICULUM
PIP2
IP3
GPCR
TrimericG protein
IP3 -gatedCa2+
channel
Receptoractivated
Hydrolysis of PIP2 to IP3 and DAG
Release of Ca2+
DAG
Activated G proteindissociates
-
Agonist
Heterotrimeric G protein signaling
FIGURE 14.21 G protein-mediated signal transduction follows a path of ago-nist to receptor to heterotrimeric G protein to effector to the effector's out-put. Both G� and G�� subunits regulate distinct effectors. In the exampleshown here, Gq regulates a phospholipase C-� to produce two second messen-gers, diacyglycerol (DAG) and inositol-trisphosphate (IP3). IP3 triggers Ca2+ re-lease from the endoplasmic reticulum.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 616

14.20 G protein signaling modules are widely used and highly adaptable 617
The heterotrimeric G proteins to whichGPCRs are coupled are composed of a nu-cleotide-binding Gα subunit and a Gβγ subunitdimer, as illustrated in FIGURE 14.24. The struc-ture of the trimer and each subunit is known forseveral states of activation and in complex withseveral interacting proteins. A Gαβγheterotrimeris named according to its α subunit, which largelydefines the G protein’s selectivity among recep-tors. Each subunit also regulates a distinct groupof effector proteins.
Gα subunits are globular, two-domain pro-teins of 38-44 kDa. The GTP-binding domainbelongs to the GTP-binding protein superfam-ily that includes the small, monomeric G pro-teins (such as Ras, Rho, Arf, Rab; see 14.23 Small,monomeric GTP-binding proteins are multiuseswitches) as well as the GTP-binding translationalinitiation and elongation factors. A second do-main modulates GTP binding and hydrolysis.Gα subunits are only slightly hydrophobic, butthey are predominantly membrane-associated
because of constitutive N-terminal fatty acyla-tion and because they bind to the membrane-attached Gβγ subunits. Mammals have 16 Gαgenes that are grouped in subfamilies accordingto similar sequence and function (e.g., s, i, q,and 12). These subfamilies are listed in FIGURE14.25.
Gβ and Gγ subunits associate irreversiblysoon after translation to form stable Gβγ dimers,which then associate reversibly with a Gα. Gβsubunits are 35 kDa proteins composed of seven
Retinal
MEMBRANE
CYTOPLASM
Structure of rhodopsin
FIGURE 14.23 The figure shows the crystal structureof the GPCR rhodopsin. Each membrane-spanning he-lix is a different color; most structures on the cytoplas-mic face are not shown. The retinal chromophore isshown within the helix bundle. GPCR sequence similar-ity separates the mammalian GPCRs into at least fourstructural families that are so diverse that there maybe little sequence similarity among the classes. Withina family, similarity is greatest in the membrane-span-ning helices, less in the interhelical loops, and least inthe N- and C-terminal domains and in the cytoplasmicloop that connects spans five and six. Regardless, thegeneralizations about functional domains in receptorsseem to hold true within different families. GPCRs fre-quently form dimers, occasionally heterodimers, anddimerization can be crucial for function. Structure gen-erated from Protein Data Bank file 1F88.
Heterotrimeric G protein structure
FIGURE 14.24 The structure of the nonactivated Gi het-erotrimer, the G protein that is responsible for inhibition ofadenylyl cyclase and for most G��-mediated signaling, isshown with each subunit colored as shown. GDP is shownbound to the G�i subunit. Structure generated from ProteinData Bank file 1GP2.
EFFECTOR PROTEIN
GsGolf
Adenylyl cyclase
Adenylyl cyclaseK+channel, PI 3-kinase
Other cation channel
Rho GEF
Gi (3)GoGz
G12G13
Gt (2)
Ggus
Gq (4)
StimulatedG
protein Inhibited
Cyclic GMP phospho-diesterase
Phospholipase-Cβ
G protein targets
FIGURE 14.25 G protein-regulated effectors do not share struc-tural similarities. They may be ion channels or membrane span-ning enzymes in the plasma membrane, peripheral proteins on theinner face of the membrane, or fundamentally soluble proteinsthat can bind to G� subunits. The chart shows the major groupsof G proteins, sorted according to sequence similarity, and someof the effectors that they are known to regulate.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 617

618 CHAPTER 14 Principles of cell signaling
�-strand repeats that form a cylindrical structureknown as a � propeller. There are five Gβ genesin mammals. Four encode strikingly similar pro-teins that naturally dimerize with the twelve Gγsubunits (Figure 14.24). The fifth, Gβ5, is lessclosely related to the others and interacts prima-rily with a Gγ-like domain in other proteins ratherthan with Gγ subunits themselves.
Gγ subunits are smaller (~7 kDa) and farmore diverse in sequence than are the Gβ’s. Thelast three amino acid residues of Gγ subunitsare proteolyzed to leave a conserved C-termi-nal cysteine that is irreversibly S-prenylatedand carboxymethylated, helping to anchor Gβγto the membrane. Gβ and Gγ subunits can as-sociate in most possible combinations. Becausealmost all cells express multiple Gβ and Gγ sub-units, it has been difficult to assign specific rolesto individual Gβγ combinations. The best rec-ognized interactions of Gβγ subunits occur atsites on Gβ, although distinct functions of Gγhave also been supported.
Heterotrimeric G proteinsregulate a wide variety ofeffectors
G protein-regulated effectors include enzymesthat create or destroy intracellular second messengers (adenylyl cyclase, cyclic GMP phos-phodiesterase, phospholipase C-β, phosphatidyli-nositol-3-kinase), protein kinases, ion channels(K+, Ca2+) and possibly membrane transportproteins (see Figure 14.25). Effectors may beintegral membrane proteins or intrinsically sol-uble proteins that bind G proteins at the mem-brane surface. No conserved G protein-bindingdomain or sequence motif has been identifiedamong effector proteins, and most effectors arerelated to proteins that have similar functionsbut that are not regulated by G proteins.Sensitivity to G protein regulation, thus, evolvedindependently in multiple families of regula-tory proteins.
Key concepts • G proteins convey signals by regulating the
activities of multiple intracellular signalingproteins known as effectors.
• Effectors are structurally and functionally diverse.• A common G-protein binding domain has not been
identified among effector proteins.• Effector proteins integrate signals from multiple G
protein pathways.
14.21
Because they can respond to a variety ofGα and Gβγ subunits, effector proteins can in-tegrate signals from multiple G protein path-ways. The different Gα or Gβγ subunits mayhave opposite or synergistic effects on a giveneffector. For example, some of the membrane-bound adenylyl cyclases in mammals are stim-ulated by Gαs and inhibited by Gαi (see Figure14.13). Many effectors are further regulated byother allosteric ligands (e.g., lipids, calmodulin)and by phosphorylation, contributing even moreto integration of information.
Effectors are usually represented as multi-ple isoforms, and each isoform may be regu-lated differently, adding to the complexity of Gprotein networks. For example, some isoformsof adenylyl cyclase are stimulated by Gβγ,whereas others are inhibited. All phospholipaseC-βs are stimulated both by Gαq family mem-bers and by Gβγ, but the potency and maximaleffect of these two inputs vary dramaticallyamong the four PLC-β isoforms.
Heterotrimeric G proteinsare controlled by aregulatory GTPase cycle
The key event in heterotrimeric G protein sig-naling is the binding of GTP to the Gα subunit.GTP binding activates the Gα subunit, whichallows both it and the Gβγ subunit to bind andregulate effectors. The Gα subunit remains ac-tive as long as GTP is bound, but Gα also hasGTPase activity and hydrolyzes bound GTP toGDP. Gα-GDP is inactive. G proteins thus traversea GTPase cycle of GTP binding/activation and hy-drolysis/deactivation, as depicted in FIGURE 14.26.Therefore, the control of G protein signaling isintrinsically kinetic. The relative signal strength,or amplitude, is proportional to the fraction ofG protein that is in the active, GTP-bound form.This fraction equals the balance of the rates ofGTP binding and GTP hydrolysis, the activating
Key concepts • Heterotrimeric G proteins are activated when the
Gα subunit binds GTP.• GTP hydrolysis to GDP inactivates the G protein.• GTP hydrolysis is slow, but is accelerated by
proteins called GAPs.• Receptors promote activation by allowing GDP
dissociation and GTP association; spontaneousexchange is very slow.
• RGS proteins and phospholipase C-βs are GAPs forG proteins.
14.22
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 618

14.22 Heterotrimeric G proteins are controlled by a regulatory GTPase cycle 619
and deactivating arms of the GTPase cycle. Bothlimbs are highly regulated over a range of ratesgreater than 1000-fold.
Receptors promote G protein activation byopening the nucleotide-binding site on the Gprotein, thus accelerating both GDP dissociationand GTP association. This process is referred toas GDP/GTP exchange catalysis. Exchange pro-ceeds in the direction of activation because theaffinity of G proteins for GTP is much higher thanthat for GDP and because the cytosolic concen-tration of GTP is about 20-fold higher than thatof GDP. Spontaneous GDP/GTP exchange is veryslow for most G proteins (many minutes), whichmaintains basal signal output at a low level. Incontrast, receptor-catalyzed exchange can takeplace in a few tens of milliseconds, which allowsrapid responses in cells such as visual photore-ceptors, other neurons, or muscle.
Because receptors are not directly requiredfor a G protein’s signaling activity, a receptor candissociate after GDP/GTP exchange and catalyzethe activation of additional G protein molecules.In this way, a single receptor may maintain theactivation of multiple G proteins, providing mo-lecular amplification of the incoming signal.Other receptors may remain bound to their Gprotein targets, which means that they do notact as amplifiers. However, more tightly boundreceptors can initiate signaling more quickly andpromote G protein reactivation when hydroly-sis of bound GTP is rapid.
In the absence of stimulus, Gα subunitshydrolyze bound GTP slowly. The average ac-tivation lifetime of the Gα-GTP complex isabout 10-150 seconds, depending on the Gprotein. This rate is far slower than rates of de-activation often observed in cells when an ag-onist is removed. For example, visual signalingterminates in about 10 ms after stimulation bya photon, and many other G protein systemsare almost as fast. GTP hydrolysis is acceler-ated by GTPase-activating proteins (GAPs),which directly bind Gα subunits. In some casesacceleration exceeds 2000-fold. Such speed isnecessary in systems like vision or neurotrans-mission, which must respond to quickly chang-ing stimuli. Because G protein signaling is abalance of activation and deactivation, GAPsdeplete the pool of GTP-activated G proteinand can thereby also act to inhibit G proteinsignaling. GAPs can thus inhibit signaling,quench output upon signal termination, orboth. What behavior predominates dependson the GAP’s intrinsic activity and its regula-tion.
There are two families of GAPs for het-erotrimeric G proteins. The RGS proteins (reg-ulators of G protein signaling) are a family ofabout 30 proteins, most or all of which haveGAP activity and regulate G protein signalingrates and amplitudes. The role of RGS proteinsin terminating the G protein signal can be seenin FIGURE 14.27. Some proteins with RGS do-mains also act as G protein-regulated effectors.These include activators of the Rho family ofmonomeric GTP-binding proteins (see below)and GPCR kinases, which are feedback regula-tors of GPCR function. The second group of Gprotein GAPs are phospholipase C-βs. These en-zymes are effectors that are stimulated by bothGαq and by Gβγ, but they also act as Gq GAPs,probably to control output kinetics.
Effector protein
G A P
GTPGDP
Pi
R e c e p t o r- a g o n i s t
Receptor + agonist
G protein
G protein-GTP*ACTIVE*
G protein-GTP-Effector protein
*ACTIVE*G protein-GDP
The regulatory GTPase cycle
FIGURE 14.26 G proteins are activated when GTP binds to the G� subunit, suchthat both G�-GTP and G�� can bind and regulate the activities of appropriateeffector proteins. G� subunits also have intrinsic GTPase activities, and the pri-mary deactivating reaction is hydrolysis of bound GTP to GDP (rather than GTPdissociation). Thus, the steady-state signal output from a receptor-G proteinmodule is the fraction of the G protein in the GTP-bound state, which reflectsthe balance of the activation and deactivation rates. Both GTP binding and GTPhydrolysis are intrinsically slow and highly regulated. GDP binds tightly to G�,such that GDP dissociation is rate-limiting for binding of a new molecule ofGTP and consequent reactivation. Both GDP release and GTP binding are cat-alyzed by GPCRs. Hydrolysis of bound GTP is accelerated by GTPase-activatingproteins (GAPs). Receptors and GAPs coordinately control both the steady-statelevel of signal output and the rates of activation and deactivation of the mod-ule.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 619

620 CHAPTER 14 Principles of cell signaling
While the GTPase cycle described in Figure14.26 is general, it is highly simplified. Interactionsamong receptor, Gα, Gβγ, GAP, and effector arefrequently simultaneous and often demonstratecomplex cooperative interactions. For example,Gβγ inhibits the release of GDP (to minimizespontaneous activation), promotes the exchangecatalyst activity of the receptor, inhibits GAP ac-tivity, and helps initiate receptor phosphoryla-tion that leads to desensitization. The othercomponents can be nearly this multifunctional.In addition, inputs from other proteins can alterthe dynamics of the GTPase cycle at several points.The core G protein module is, thus, functionallyversatile as a signal processor in addition to be-ing versatile in the scope of its targets.
Small, monomeric GTP-binding proteins aremultiuse switches
Monomeric GTP-binding proteins, which areencoded by about 150 genes in animals, mod-ulate a wide variety of cellular processes includ-ing signal transduction, organellar trafficking,intra-organellar transport, cytoskeletal assem-bly, and morphogenesis. The small GTP-bind-ing proteins that most clearly function in signaltransduction are the Ras and Ras-related pro-teins (Ral, Rap) and the Rho/Rac/Cdc42 pro-teins, about 10-15 in all. They are usually about20-25 kDa in size and are homologous to theGTP-binding domains of Gα subunits.
The regulatory activities of the small GTP-binding proteins are controlled by a GTP bind-ing and hydrolysis cycle like that of theheterotrimeric G proteins, with similar regula-tory inputs. They are activated by GTP, and hy-drolysis of bound GTP to GDP terminatesactivation. GDP/GTP exchange catalysts, knownas GEFs (guanine nucleotide exchange factors,functionally analogous to GPCRs) promote ac-tivation, and GAPs accelerate hydrolysis andconsequent deactivation. In addition, GDP dis-sociation inhibitors (GDIs) slow spontaneousnucleotide exchange and activation to dampenbasal activity, an activity shared by Gβγ subunitsfor the heterotrimeric G proteins.
While the underlying biochemical regula-tory events are essentially identical formonomeric and heterotrimeric G proteins,monomeric G proteins use the basic GTPase cy-cle in additional ways. Signal output by het-erotrimeric G proteins and many monomericG proteins is usually thought to reflect a bal-ance of their active (GTP-bound) and inactive(GDP-bound) states in a rapidly turning-overGTPase cycle. GEFs favor formation of more ac-tive G protein, and GAPs favor the inactive state.In contrast, probably an equal number of themonomeric G proteins behave as acute on-offswitches. Upon binding GTP, they initiate aprocess (regulation, recruitment of other pro-
Key concepts • Small GTP-binding proteins are active when bound
to GTP and inactive when bound to GDP.• GDP/GTP exchange catalysts known as GEFs
(guanine nucleotide exchange factors) promoteactivation.
• GAPs accelerate hydrolysis and deactivation.• GDP dissociation inhibitors (GDIs) slow
spontaneous nucleotide exchange.
14.23Current (pA)
Time (s)
heterozygouswild-type
knockout0.50
0.25
0.00
0 2 4
Light flash
Single photon responses of GAP-deficient mice
FIGURE 14.27 G protein GAPs can accelerate signal termi-nation upon removal of agonist, and often do not act as in-hibitors during the response to receptor. The figure showsthe electrical response of a mouse photoreceptor (rod) cellto a single photon of light. In mice that lack RGS9, the GAPfor the photoreceptor G protein Gt, the signal is prolongedfor many seconds because hydrolysis of GTP bound to Gt isslow. In wild-type or heterozygous mice, hydrolysis takesplace in about 15 milliseconds, and the decay of the signalis much faster. Note that the maximal output is similar inwild-type and mutant mice, indicating that the GAP doesnot act as an inhibitor in rod cells. In humans, genetic lossof RGS9 leads to severe loss of vision that is particularlymarked in bright light. Reproduced from Chen et al. Nature.2000. 403:557–560. Permission also granted by Ching-KangJason Chen, Virginia Commonwealth University.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 620

14.24 Protein phosphorylation/dephosphorylation is a major regulatory mechanism in the cell 621
teins). They then maintain this activity, some-times for many seconds or minutes, until theyare acted upon by a GAP. For example, themonomeric G protein Ran regulates nucleocy-toplasmic trafficking of protein and RNA in bothdirections, cooperating with carrier proteinsknown as karyopherins (see 5.15 The Ran GTPasecontrols the direction of nuclear transport). In thenucleus, high Ran GEF activity promotes GTPbinding. Nuclear Ran-GTP then binds importkaryopherins to drive dissociation of newly ar-rived cargo and promote return of the karyo-pherin to the cytoplasm. It also binds exportkaryopherins to permit binding of outgoingcargo. Outside the nucleus, high Ran GAP ac-tivity promotes GTP hydrolysis. CytoplasmicRan-GDP dissociates from both the export karyo-pherins to allow dissociation of outgoing cargoand from the import karyopherins to allow themto bind cargo for import. Thus, for monomericG proteins such as Ran, each phase of the GTPasecycle determines a specific, coupled step in aparallel regulatory cycle.
A second major difference between themonomeric and the heterotrimeric G proteinsis the structures of the GEFs, GAPs, and GDIs.Both GEFs and GAPs for monomeric GTP-bind-ing proteins are structurally heterogeneous (al-though some clearly related families areevident). In addition, mechanisms for regulat-ing these GEFs and GAPs are equally diverse.They include phosphorylation by protein ki-nases; allosteric regulation by heterotrimericand/or monomeric G proteins, by second mes-sengers and by other regulatory proteins; sub-cellular sequestration or recruitment to scaffolds;and assorted other mechanisms.
The Ras proteins were the first small GTP-binding proteins to be discovered. They wereidentified as oncogene products because theycause malignant growth if they are either over-expressed or persistently activated by mutation;they are among the most commonly mutatedgenes in human tumors. Several viral ras genesfigure prominently as oncogenes.
Mammalian cells contain three ras genes (H,N, and K). They may share inputs and outputsto varying extents, and they can compensate foreach other in some genetic screens. It has beendifficult to assign unique functions to the indi-vidual Ras proteins. Inputs to the Ras proteinsare diverse and speak to the importance of Rasproteins as a crucial node in signaling.
Ras GEFs and GAPS are regulated by bothreceptor and nonreceptor Tyr kinases throughdirect phosphorylation and by recruitment ofthe regulators to the plasma membrane. Other
cytoplasmic serine/threonine kinases also con-verge on Ras activation. Rap1, another mem-ber of the Ras family, may also fit directly intothis network because it is suspected of compet-ing with Ras proteins for protein kinase targets;in vivo it can suppress the oncogenic activity ofRas. Rap1 is regulated independently, however,and acts on independent signaling pathways aswell. One of its GAPs is stimulated by the Giclass of G proteins, for example, and its severalGEFs are stimulated by Ca2+, diacylglycerol, andcAMP.
Ras proteins generally regulate cell growth,proliferation, and differentiation by modulat-ing the activities of multiple effector proteins.The best known and best studied Ras effector isthe protein kinase Raf, which initiates a MAPKcascade. FIGURE 14.28 shows well established Raseffectors.
Rho, Rac, and Cdc42 are related monomericGTP-binding proteins that are involved in gen-erating signals that affect cell morphology. Eachclass of proteins regulates its own array of effec-tors and is controlled by separate groups of GEFs,GAPs, and GDIs. Effectors regulated by this fam-ily include phospholipases C and D, multipleprotein and lipid kinases, proteins that nucle-ate or reorganize actin filaments, and compo-nents of the neutrophil oxygen activatingsystem, among others (see 8.14 Small G proteinsregulate actin polymerization).
Protein phosphorylation/dephosphorylation is amajor regulatorymechanism in the cell
Key concepts • Protein kinases are a large protein family.• Protein kinases phosphorylate Ser and Thr, or Tyr,
or all three.• Protein kinases may recognize the primary
sequence surrounding the phosphorylation site.• Protein kinases may preferentially recognize
phosphorylation sites within folded domains.
14.24
Function Effector
Ras has three main effectors
Target
Protein kinase cascade Raf MAPK
Lipid kinase PI 3-kinase Akt
Exchange factor RalGDS Exocyst
FIGURE 14.28 Ras-GTP bindsto many proteins. Three wellestablished effectors includeRaf, PI 3-kinase, and RalGDS.Activation of these effectorsactivates a MAPK pathway, in-creases PI 3-kinase activity,and promotes assembly of aprotein complex involved inexocytosis of secretory vesi-cles.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 621

622 CHAPTER 14 Principles of cell signaling
Protein phosphorylation is the most commonform of regulatory posttranslational modifica-tion. It occurs in all organisms, and it is estimatedthat about one-third of proteins in animals areat some time phosphorylated. Phosphorylationcan stimulate or inhibit the catalytic activity ofan enzyme, the affinity with which a proteinbinds other molecules, its subcellular localiza-tion, its ability to be further covalently modified,or its stability. Single phosphorylations may cause500-fold or greater changes in activity, and pro-teins are often phosphorylated on multipleresidues in complex and interacting patterns.
Most protein phosphorylation in eukary-otes, and essentially all in animals, is catalyzedby protein kinases; dephosphorylation is cat-alyzed by phosphoprotein phosphatases. Bothclasses of enzymes are controlled by diversemechanisms. In addition, proteins are oftenphosphorylated by multiple protein kinases, re-sulting in the generation of a range of activitystates. This complexity allows inputs from dif-ferent signaling pathways to be integrated intothe resulting activity of the target.
In bacteria, plants, and fungi, an additionalprotein phosphorylating system known as two-component signaling is vital. The protein ki-
nases involved in two-component signaling areunrelated to the eukaryotic protein kinase su-perfamily and phosphorylate aspartate residuesrather than serine, threonine, or tyrosine.
Protein kinases transfer a phosphoryl groupfrom ATP to Ser, Thr, and Tyr residues of pro-tein substrates to form chemically stable phos-phate esters, as shown in FIGURE 14.29. Inanimals, the distribution of phosphate amongthese three amino acid residues is uneven:~90%-95% is on Ser, 5%-8% on Thr, and lessthan 1% on Tyr residues. The human genomecontains approximately 500 genes that encodeprotein kinases, and many protein kinasemRNAs undergo alternative splicing. This makesthe protein kinase gene superfamily one of thelargest functional gene groups. The number anddiversity of these enzymes emphasize the greatand varied uses of protein kinases to regulate cel-lular functions. Although some protein kinaseshave a limited tissue and/or developmental dis-tribution, many are ubiquitously expressed.
Protein kinases are grouped according totheir residue specificity. Protein kinases thatphosphorylate Ser will usually also recognizeThr, hence the name protein Ser/Thr kinase.Multicellular organisms have protein Tyr ki-nases, which only recognize Tyr. Dual speci-ficity protein kinases can phosphorylate Ser,Thr, and Tyr in the appropriately restricted sub-strate conformational context and are gener-ally the most selective of the protein kinases.
The analysis of the kinomes of several or-ganisms has led to a more elaborate groupingderived from sequence relationships, shown inFIGURE 14.30, that also reflects to some extenton regulatory mechanisms and substrate speci-ficity. For example, the AGC group is namedfor its founding members, cAMP-dependentprotein kinase (PKA), cyclic GMP-dependentprotein kinase (PKG), Ca2+, and phospholipid-dependent protein kinase (PKC). These proteinkinases are regulated by second messengers andprefer substrates that contain basic residues nearthe phosphorylation site.
In addition to substrate specificity for aminoacid residues, most protein kinases are also selec-tive for local sequence surrounding the substratesite. Screening strategies have resulted in meth-ods to predict if proteins contain consensus sub-strate sites for a wide variety of protein kinases.Antibodies can be used to identify and roughlyquantitate protein phosphorylation at specificsites in proteins. Beyond local recognition, pro-tein kinases may display marked substrate selec-tivity among similar proteins based on overall
Mg2+
(Mg2+. ATP)SUBSTRATE 2
(Mg2+. ADP)
SUBSTRATE 1(Protein SER)
CH2OHO-
O
O
PO-
O
O
PO-
O
O
P
Adenine
(Mg2+.Triphosphate)
PRODUCT 2PRODUCT 1
(Phosphorylated protein)
O-
O-
O
P
PROTEIN KINASE
O-
O-
Mg2+
CH2
O-
O
O
PO-
O
O
P
Adenine
Rib
Ribose
O
CH2CH
C
HN
HN
O
C O
CH2CH
C
HN
HN
O
C O
+
Protein kinases are two substrate enzymes
(Mg2+.Diphosphate)
FIGURE 14.29 Protein kinases transfer the �-phosphoryl group from ATP toserine, threonine, or tyrosine residues in protein substrates.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 622

14.24 Protein phosphorylation/dephosphorylation is a major regulatory mechanism in the cell 623
three-dimensional structure, for example, oramong proteins that have been differentially co-valently modified by phosphorylation or ubiqui-tination.
In animal cells, some protein kinases arehormone receptors that span the plasma mem-brane. Some protein kinase receptors are pro-tein serine/threonine kinases, such as thetransforming growth factor-� (TGF-�)receptor,but the majority are protein tyrosine kinases, in-cluding receptors for insulin, epidermal growthfactor (EGF), platelet-derived growth factor(PDGF), and other regulators of cell growth anddifferentiation. Other protein kinases are in-trinsically soluble intracellular enzymes, al-though they may bind to one or more organellarmembranes.
X-ray crystallographic structures of proteinkinases have revealed a wealth of informationabout their mechanism of activation. The con-served minimum catalytic core of a protein ki-nase contains about 270 amino acids, yieldinga minimum molecular mass of about 30,000Da. Within this core, there are two folded do-mains that form the active site at their inter-face, as shown in FIGURE 14.31. One or both ofthe conserved lysine (Lys) or aspartate (Asp)residues that are required for phosphoryl trans-fer are frequently mutated to disrupt kinase ac-tivity. A sequence near the active site, referredto as the activation loop, often undergoes a con-formational rearrangement to generate activeforms of the protein kinases and is the mostcommon site of regulatory phosphorylation in
Human kinome tree FIGURE 14.30 The protein kinases inthe human genome can be grouped ac-cording to sequence relationships thatreveal seven major branches. The tyro-sine kinases are contained within onemajor branch. The others are Ser/Thr-specific or dual specificity, and are namedfor the best described members: AGCfrom PKA, PKG, and PKC; CAMK from thecalcium, calmodulin-dependent kinases;CMGC from CDKs, MAPKs, GSK3, Clks; CK1from casein kinase 1; STE from Ste20,Ste11, and Ste7, the MAP4K, MAP3K,and MAP2K in the yeast mating path-way; and TKL, the Tyr kinase-like en-zymes. Reproduced with permission fromG. Manning, et al. 2002. Science. 298:1912-1934. © 2002 AAAS. Photo cour-tesy of Gerard Manning, Salk Institute,and reprinted with permission of CellSignaling Technology, Inc. (www.cellsig-nal.com).
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 623
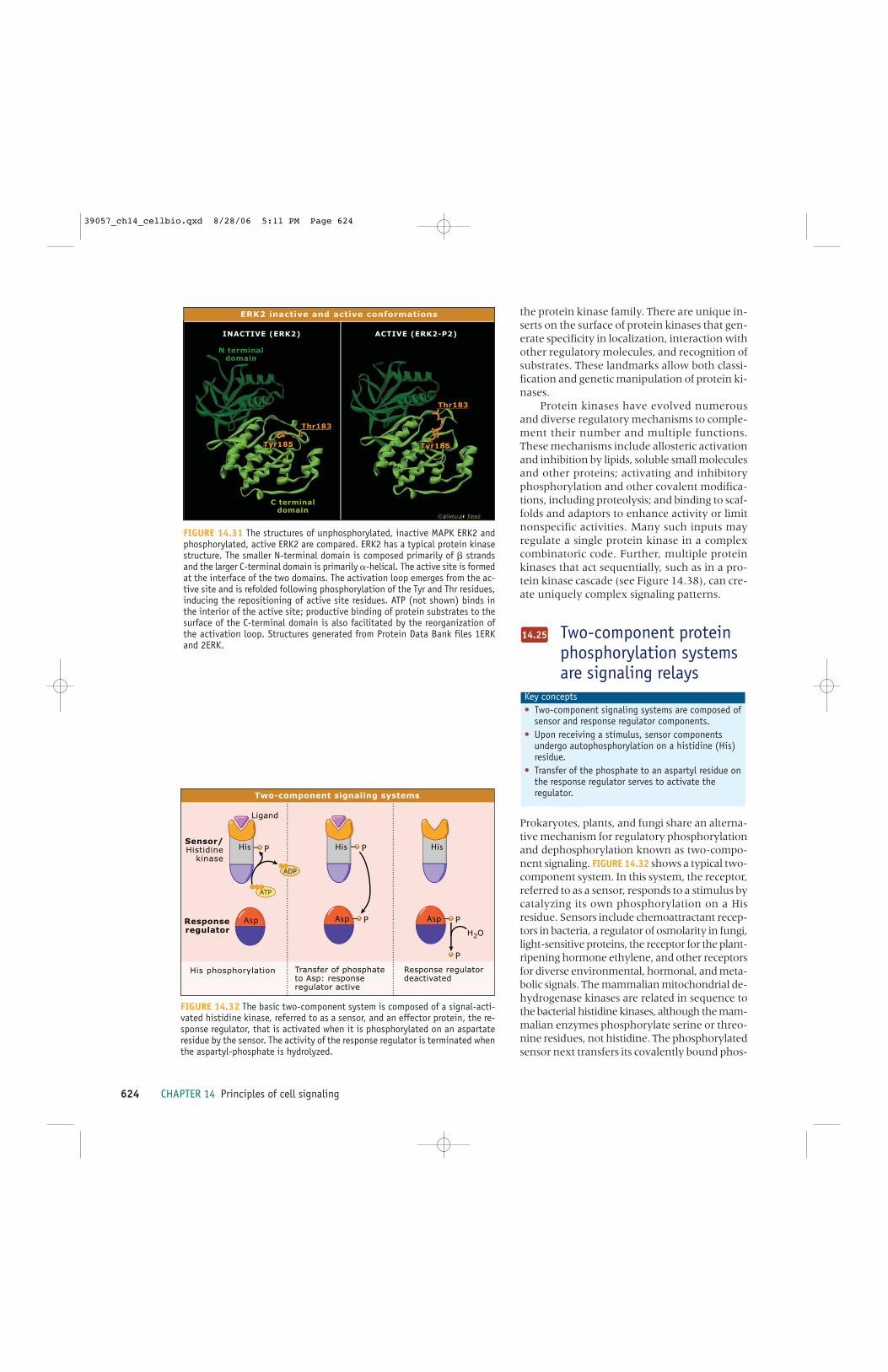
624 CHAPTER 14 Principles of cell signaling
the protein kinase family. There are unique in-serts on the surface of protein kinases that gen-erate specificity in localization, interaction withother regulatory molecules, and recognition ofsubstrates. These landmarks allow both classi-fication and genetic manipulation of protein ki-nases.
Protein kinases have evolved numerousand diverse regulatory mechanisms to comple-ment their number and multiple functions.These mechanisms include allosteric activationand inhibition by lipids, soluble small moleculesand other proteins; activating and inhibitoryphosphorylation and other covalent modifica-tions, including proteolysis; and binding to scaf-folds and adaptors to enhance activity or limitnonspecific activities. Many such inputs mayregulate a single protein kinase in a complexcombinatoric code. Further, multiple proteinkinases that act sequentially, such as in a pro-tein kinase cascade (see Figure 14.38), can cre-ate uniquely complex signaling patterns.
Two-component proteinphosphorylation systemsare signaling relays
Prokaryotes, plants, and fungi share an alterna-tive mechanism for regulatory phosphorylationand dephosphorylation known as two-compo-nent signaling. FIGURE 14.32 shows a typical two-component system. In this system, the receptor,referred to as a sensor, responds to a stimulus bycatalyzing its own phosphorylation on a Hisresidue. Sensors include chemoattractant recep-tors in bacteria, a regulator of osmolarity in fungi,light-sensitive proteins, the receptor for the plant-ripening hormone ethylene, and other receptorsfor diverse environmental, hormonal, and meta-bolic signals. The mammalian mitochondrial de-hydrogenase kinases are related in sequence tothe bacterial histidine kinases, although the mam-malian enzymes phosphorylate serine or threo-nine residues, not histidine. The phosphorylatedsensor next transfers its covalently bound phos-
Key concepts • Two-component signaling systems are composed of
sensor and response regulator components.• Upon receiving a stimulus, sensor components
undergo autophosphorylation on a histidine (His)residue.
• Transfer of the phosphate to an aspartyl residue onthe response regulator serves to activate theregulator.
14.25
INACTIVE (ERK2) ACTIVE (ERK2-P2)
N terminaldomain
N terminaldomain
C terminaldomain
C terminaldomain
Tyr185Tyr185Tyr185Tyr185
Thr183Thr183
Thr183Thr183
ERK2 inactive and active conformations
FIGURE 14.31 The structures of unphosphorylated, inactive MAPK ERK2 andphosphorylated, active ERK2 are compared. ERK2 has a typical protein kinasestructure. The smaller N-terminal domain is composed primarily of � strandsand the larger C-terminal domain is primarily �-helical. The active site is formedat the interface of the two domains. The activation loop emerges from the ac-tive site and is refolded following phosphorylation of the Tyr and Thr residues,inducing the repositioning of active site residues. ATP (not shown) binds inthe interior of the active site; productive binding of protein substrates to thesurface of the C-terminal domain is also facilitated by the reorganization ofthe activation loop. Structures generated from Protein Data Bank files 1ERKand 2ERK.
His P
Asp P
His
Asp P
P
H2OResponseregulator
Sensor/Histidine
kinaseHis P
Asp
Ligand
ADP
ATP
His phosphorylation Transfer of phosphate to Asp: response regulator active
Response regulator deactivated
Two-component signaling systems
FIGURE 14.32 The basic two-component system is composed of a signal-acti-vated histidine kinase, referred to as a sensor, and an effector protein, the re-sponse regulator, that is activated when it is phosphorylated on an aspartateresidue by the sensor. The activity of the response regulator is terminated whenthe aspartyl-phosphate is hydrolyzed.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 624

14.27 Phosphoprotein phosphatases reverse the actions of kinases and are independently regulated 625
phate to an aspartyl residue on a second proteinknown as a response regulator. Response regu-lators initiate cellular responses, usually by bind-ing to other cytoplasmic proteins and allostericallyregulating their activities.
Although all two-component systems fol-low this same general pattern, their structuresand precise reaction pathways vary enormously.Some two-component systems are composedof only one protein (sensor and response reg-ulator in a single polypeptide chain). Others arecomposed of a sensor protein and two aspartyl-phosphorylated proteins, in which the first orthe second may display response regulatory ac-tivity. Finally, two-component systems usuallylack conventional protein phosphatases.Hydrolysis of the aspartyl-phosphate bond maybe spontaneous or regulated by the responseregulator itself.
Pharmacologicalinhibitors of proteinkinases may be used tounderstand and treatdisease
Many inhibitors have been developed for basicresearch purposes to explore the functions ofprotein kinases. The importance of these en-zymes in disease processes has also made themtargets of drug screening projects yielding in-hibitors for many protein kinases. The major-ity of pharmacological inhibitors of proteinkinases compete with ATP binding. Because ofthe huge number of ATP-binding proteins in acell, there are inevitable concerns about in-hibitor specificity not only with respect to theother protein kinases but also to the other pro-teins that bind nucleotides. This problem hasbeen mitigated with variable success throughchemical library screening, structure-based mod-ification of lead compounds, and inhibitor test-ing against panels of protein kinases.
Many inhibitors with actions on PKA orPKCs, for example, have effects on several othermembers of the AGC family. Although phar-macological inhibitors with effects on PKAabound, the most selective are derived from the
Key concepts • Protein kinase inhibitors are useful both for
signaling research and as drugs.• Protein kinase inhibitors usually bind in the ATP
binding site.
14.26
naturally occurring small inhibitory proteinknown as PKI or the Walsh inhibitor. In vitroand cell-based screens have identified muchmore selective inhibitors for MAP2Ks in theERK1/2 pathway. These inhibitors have fewerknown protein kinase cross reactivities, prob-ably due to the fact that they do not bind in theATP site. Among inhibitors that have progressedin the clinic, compounds developed against theEGF receptor and certain other protein tyro-sine kinases have had considerable success.
Phosphoproteinphosphatases reverse theactions of kinases and areindependently regulated
Protein phosphorylation is reversed by phospho-protein phosphatases. These enzymes display dis-tinct specificities and modes of regulation.Phosphoprotein phosphatases can be consideredin two broad groups based on their specificity andsequence relationships: protein-serine/threoninephosphatases and protein-tyrosine phosphatases.
Most protein-serine/threonine phosphatasesare regulated by association with other proteins.Targeted localization is the major determinant ofsubstrate specificity. Phosphoprotein phosphatase1 (PP1) associates with a variety of regulatorysubunits that specifically direct it to relevant or-ganelles. One subunit (known as the G subunit),for example, specifies association with glycogenparticles. The interaction with this subunit is it-self regulated by phosphorylation. Small proteininhibitors can suppress PP1 activity.
Phosphoprotein phosphatase 2A (PP2A) iscomposed of a catalytic subunit, a scaffoldingsubunit, and one of a large number of regulatorysubunits. The regulatory subunit modulates ac-tivity and localization of the phosphatase. Someviruses alter the behavior of the cells they infectby interfering with phosphatase activity. For ex-ample, cells transformed with the SV40 virusexpress a viral protein known as small t anti-
Key concepts • Phosphoprotein phosphatases reverse the actions
of protein kinases.• Phosphoprotein phosphatases may
dephosphorylate phosphoserine/threonine,phosphotyrosine, or all three.
• Phosphoprotein phosphatase specificity is oftenachieved through the formation of specific proteincomplexes.
14.27
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 625

626 CHAPTER 14 Principles of cell signaling
gen. Small t displaces the regulatory subunitfrom PP2A and alters the activity and the sub-cellular localization of the phosphatase. In ad-dition, natural toxins such as okadaic acid,calyculin, and microcystin inhibit PP2A and PP1to varying extents both in vitro and in intact cells.
Another major protein-serine/threoninephosphatase, called calcineurin (also known asphosphoprotein phosphatase 2B), is regulatedby Ca2+-calmodulin (see 14.15 Ca2+ signalingserves diverse purposes in all eukaryotic cells) andplays essential roles in cardiac development andT cell activation, among other events. The ma-jor mechanism of action of the immunosup-pressants cyclosporin and FK506 is to inhibitcalcineurin.
The protein tyrosine phosphatases (PTPs)are cysteine-dependent enzymes that utilize aconserved Cys-Xaa-Arg motif to hydrolyze phos-phoester bonds in their substrates. The PTPs areencoded by over 100 genes in humans and areclassified in four subfamilies: the phosphotyro-sine-specific phosphatases, the Cdc25 phos-phatases, the dual specificity phosphatases(DSPs), and the low molecular weight phos-phatases.
Thirty-eight of the PTPs are highly selec-tive for phosphotyrosine residues within sub-strates. Some of the phosphotyrosine-selectivephosphatases are transmembrane proteins,whereas others are membrane associated. Themost obvious function of the PTPs is to reversethe functions of tyrosine kinases; however, somehave primary functions in transducing tyrosinekinase signals. For example, the protein tyro-sine phosphatase SHP2 (also known as SHPTP2),binds to certain tyrosine kinase receptorsthrough its SH2 domain and is itself tyrosinephosphorylated, thereby creating a binding sitefor the SH2 domain-containing adaptor pro-tein, Grb2, which leads to activation of Ras (see14.32 MAPKs are central to many signaling path-ways).
The Cdc25 phosphatases recognize cyclin-dependent kinase (CDK) family members assubstrates and play a critical role in increasingCDK activity at key junctures of the cell cycle(see Figure 14.39 and 11.4 The cell cycle is a cycleof CDK function). Similar to the dual specificitykinases, the dual specificity phosphatases arespecific for a restricted number of substrates. Anumber of DSPs dephosphorylate MAPKs; theseDSPs are called MAP kinase phosphatases, orMKPs. Several of these have been implicatedin MAPK nuclear entry and exit. Some MKPsare encoded by early response genes, whose
products are active near the initiation of the cellcycle (see 11.7 Entry into cell cycle and S phase istightly regulated).
Substrates of other PTP family members,such as the tumor suppressor PTEN, includephosphoinositides, which are phosphory-lated derivatives of the glycerolipid phos-phatidylinositol that serve as second messengers(see 14.16 Lipids and lipid-derived compounds aresignaling molecules). Removal of the phosphategroup inactivates the second messenger. It re-mains unclear whether members of this groupwork exclusively on phophoinositides or alsoon protein tyrosine phosphate.
Covalent modification byubiquitin and ubiquitin-like proteins is anotherway of regulating proteinfunction
An important mechanism for control of proteinfunction is through covalent modification withsmall proteins of the ubiquitin family. Ubiquitinis one of a family of proteins referred to as ubiq-uitin-like (Ubl) proteins. Ubiquitin itself is highlyconserved among species, suggesting the func-tional importance of all of its 76 residues. In ad-dition to the long-established role of ubiquitinin initiating protein degradation, ubiquitin mod-ification also has a variety of functions in signaltransduction.
Ubl proteins are conjugated to the substrateprotein by an isopeptide bond between an aminogroup on the substrate, usually from a Lys sidechain, and the C-terminal Gly residue of theprocessed Ubl protein. E1, E2, and E3 proteinsare required to catalyze conjugation to Ubl pro-teins (see Biochem 4.3 Ubiquitin attachment to sub-strates requires multiple enzymes). Several Ublproteins may be attached to one substrate, of-ten by serial formation of a polyubiquitin chain.Mono- and polyubiquitination both change the
Key concepts • Ubiquitin and related small proteins, may be
covalently attached to other proteins as atargeting signal.
• Ubiquitin is recognized by diverse ubiquitinbinding proteins.
• Ubiquitination can cooperate with other covalentmodifications.
• Ubiquitination regulates signaling in addition toits role in protein degradation.
14.28
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 626

14.28 Covalent modification by ubiquitin and ubiquitin-like proteins is another way of regulating protein function 627
protein’s behavior to induce downstream signals.Monoubiquitination is a significant regulatorymodification in vesicular trafficking and DNA re-pair. For example, the monoubiquitinated formof the FANCD2 protein becomes associated withthe repair protein BRCA1 at sites of DNA re-pair. Modification by the Ubl protein SUMO hasroles in nuclear transport, transcription, andcell cycle progression.
Polyubiquitin chains are formed when theLys residues of ubiquitin itself, particularly K48and K63, are ubiquitinated. Addition of polyu-biquitin with a K48 linkage generally directs pro-teins to the proteasome for degradation, whereasconjugation to polyubiquitin chains with a K63linkage promotes signal transmission, not prote-olysis. Protein-bound ubiquitin is recognized bya variety of ubiquitin binding domains, includ-ing UIM (ubiquitin-interacting motif), UBA (ubiq-uitin association), and certain zinc finger domains.Such domains have the capacity to act as recep-tors for ubiquitin within modified proteins.
Activation of the transcription factor NF-κBoccurs by a mechanism dependent on modifica-tion both by the addition of Ubl proteins and phos-phorylation. This fascinating example of regulationby ubiquitin is depicted in FIGURE 14.33. Prior tostimulation, NF-κB is retained in the cytoplasmin an inactive form by binding to its inhibitor, IκB.Phosphorylation of IκB by the IκB kinase (IKK)complex promotes its recognition by a multisub-unit E3 ligase, which directs its ubiquitinationand subsequent proteasomal degradation.Destruction of IκB allows NF-κB to move to thenucleus to mediate changes in transcription.
IκB can be stabilized in response to certainsignals through covalent attachment of the Ubl,SUMO. Sumoylation occurs on the same Lysresidues that must be conjugated to ubiquitinto achieve IκB degradation. Thus, SUMO at-tachment stabilizes IκB and attenuates NF-κB ac-tion. This is one of numerous examples ofcrosstalk between Ubl conjugates.
A key regulatory event in NF-κB signalingis activation of the IKK complex. IKK is itselfregulated by ubiquitination and phosphoryla-tion. The cytokine interleukin-1β (IL-1�) causesassociation of adaptor proteins with its recep-tor to create a receptor activation complex. Theinterleukin-1β receptor activation complex re-cruits another adaptor complex containingTRAF6. A phosphorylation event releases aTRAF6 complex from the receptor activationcomplex into the cytoplasm.
TRAF6 contains a RING domain, and is anE3 ubiquitin ligase that catalyzes formation of
K63 polyubiquitin chains on the protein kinaseTAK1. Polyubiquitinated TAK1 can then recruitTAB2 and TAB3, which are adaptor proteinswith conserved zinc finger domains. These par-ticular zinc finger domains bind to polyubiqui-tinated TAK1 and enhance its activity. TAK1,thus activated, phosphorylates and activatesIKK, which then phosphorylates IκB, targetingit for degradation. Thus, ubiquitin-binding do-mains, such as the TAB2 and TAB3 zinc fingers,may selectively recognize K63 polyubiquitinchains to promote signal transmission.
Naturally occurring small molecules maycontrol ubiquitin ligase activity directly. Auxin(indole 3-acetic acid) is a plant hormone that reg-ulates development by promoting the transcrip-tion of a large number of genes. Rather than
NUCLEUS
DNA
IKKIKK
Nemo
IL-1β
TAK1
TRAF6 TRAF6
K48 ubiquitination
Degradation
TRAF6 TRAF6
TRAF6 TRAF6
TAB2
K63 ubiquitination
Complex formation
ATP
ADP
ATP
ADP
CYTOPLASM
Modification with Ubl proteins plays multiple roles in IL-1β signaling
FIGURE 14.33 Activation of NF-�B involves steps dependent on the interaction of pro-teins attached to ubiquitin through ubiquitin-binding proteins, competition by sumoy-lation, phosphorylation, and ubiquitin-mediated protein degradation.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 627

628 CHAPTER 14 Principles of cell signaling
stimulating transcription factors, however, auxinaccelerates the degradation of several specifictranscriptional repressors. The auxin receptor isin fact a ubiquitin ligase complex that targetsthe auxin-regulated transcriptional repressorsfor proteolysis. F-box proteins account for allof the auxin binding activity in plant extracts.
The Wnt pathwayregulates cell fate duringdevelopment and otherprocesses in the adult
Wnt pathways function during embryonic de-velopment and in the adult in morphogenesis,body patterning, axis formation, proliferation,and cell motility. The classical Wnt signalingmechanism was uncovered largely throughstudies of Drosophila and Xenopus development,as well as by analyzing genetic alterations incancer.
Wnt proteins are unusual extracellular lig-ands. In addition to carbohydrate, they containcovalently bound palmitate that is essential fortheir biological activity. Wnts transduce signalsby binding to multiple distinct receptors. Themost significant are members of the Frizzledfamily of seven-transmembrane-spanning recep-tors.
Wnts regulate the stability of β-catenin,which either is rapidly degraded or, in responseto Wnt, is stabilized to enter the nucleus andinduce transcription by interacting with TCF(T-cell factor). Genes induced include c-jun, cy-clin D1, and many others.
The coordinated activities of the protein ki-nases glycogen synthase kinase 3 (GSK3) andcasein kinase 1(CK1), the scaffolding proteinsaxin and adenomatous polyposis coli (APC), andthe protein disheveled (DSH) are key to β-cateninstability. In the absence of Wnt, phosphoryla-tion of β-catenin by CK1 and GSK3 promotesits ubiquitination and subsequent destructionby the proteasome. Axin and APC are requiredfor phosphorylation of β-catenin by GSK3.
In contrast to most seven transmembrane-
Key concepts • Seven transmembrane-spanning receptors may
control complex differentiation programs.• Wnts are lipid-modified ligands.• Wnts signal through multiple distinct receptors.• Wnts suppress degradation of �-catenin, a
multifunctional transcription factor.
14.29
spanning receptors, the Frizzled family has notyet been shown to have significant functionsmediated by a heterotrimeric G protein, and Gproteins may not be central to this pathway.Instead a proximal step in signaling by Frizzledinvolves binding to DSH, which inactivates theβ-catenin destruction mechanism.
Mutations that cause changes in theamounts of components of the classical pathwayare common in a wide variety of cancers. BothWnts and β-catenin may be viewed as proto-oncogenes. APC is a tumor suppressor and ismutated in the majority of human colorectalcancers, for example. Either too little or toomuch axin can also disrupt Wnt signaling, andaxin, like APC, is a tumor suppressor.
Wnts utilize additional signaling mecha-nisms. The receptor proteins Lrp5/6 (which arerelated to the low-density lipoprotein receptor)are Wnt receptors and also bind axin. Wnts bindto tyrosine kinase receptors to influence axonguidance and to other proteins that inhibit theirfunction. Through DSH, Wnts can regulate theJNK MAPK pathway and Rho family G proteinsto control planar cell polarity. Certain Wnts in-crease intracellular calcium to activate calcium-dependent signaling pathways.
Diverse signalingmechanisms are regulatedby protein tyrosine kinases
A large group of protein tyrosine kinases arereceptors that span the plasma membrane andbind extracellular ligands, as shown in FIGURE14.34. The receptors are generally activated bygrowth factors whose normal physiological func-tions are to promote growth, proliferation, de-velopment, or maintenance of differentiatedproperties. This group includes receptors for in-sulin, epidermal growth factor (EGF), andplatelet derived growth factor (PDGF). Thesereceptors both control the activities of manyother protein kinases of all families and directlyregulate other classes of signaling proteins.
Key concepts • Many receptor protein tyrosine kinases are
activated by growth factors.• Mutations in receptor tyrosine kinases can be
oncogenic.• Ligand binding promotes receptor oligomerization
and autophosphorylation.• Signaling proteins bind to the phosphotyrosine
residues of the activated receptor.
14.30
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 628

14.30 Diverse signaling mechanisms are regulated by protein tyrosine kinases 629
Because of their physiologic roles as growthregulators, mutations that activate receptor ty-rosine kinases are often oncogenic. For exam-ple, the oncogene erbB results from themutational loss of the extracellular ligand-bind-ing domain of a kinase closely related to theEGF receptor. This mutation causes constitu-tive activation of the protein kinase domain.Point mutations that affect the transmembranedomain can also cause oncogenic activation, asis found in the EGF receptor-related neu/HER2oncogene (see 13.8 Cell growth and proliferationare activated by growth factors).
Receptor tyrosine kinases are diverse bothin their extracellular ligand-binding domainsand, with the exception of a conserved tyrosineprotein kinase domain, their intracellular reg-ulatory regions. These receptors usually haveone membrane span per monomer but some,such as the insulin receptor, which is a disulfide-bonded heterotetramer, have two. Ligand bind-ing to receptor tyrosine kinases favors receptoroligomerization and enhances kinase activityleading to increased Tyr phosphorylation of theintracellular domain of the receptor and of as-sociated molecules. These tyrosine-phosphory-lated motifs create docking sites for additionalsignal transducers and adaptors.
A comparison of the PDGF and insulin re-ceptors reveals common themes and a range ofbehaviors of receptor tyrosine kinases. The twoPDGF receptors are monomeric receptor tyro-sine kinases. The insulin receptor exists in twoalternatively spliced forms each of which is aheterotetramer of two � and two � subunits. Ineach case, the receptor isoforms utilize someunique signaling mechanisms.
PDGF and insulin each stimulate the kinaseactivity of their receptors, causing oligomeriza-tion and autophosphorylation. Seven or moresites are phosphorylated on the PDGF receptor,and each phosphotyrosine residue generates abinding site for one or more SH2 domain-con-taining proteins as illustrated in FIGURE 14.35. ThePDGF receptor binds PI 3-kinase, p190 Ras GAP,phospholipase C-�, Src (which may catalyze ad-ditional Tyr phosphorylation of the receptor),and the SHP2 tyrosine phosphatase which itselfbinds the adaptor Grb2 (see 14.32 MAPKs are cen-tral to many signaling pathways). With the excep-tion of Src, all of these proteins are also receptorsubstrates. Thus, substrates are recruited to thereceptor as a consequence of specific interactionsof substrate SH2 domains with receptor phos-photyrosine producing changes in activities anddistributions of numerous intracellular signal
Src
Shp2CYTOPLASM
PDGFreceptors
PI 3-kinase
p190RasGAP
PDGF
PLC-γ SOS
Grb2
Activation of the PDGF receptor leads to many outputs
p85 p110
FIGURE 14.35 PDGF binds to its receptor and induces receptor au-tophosphorylation. The autophosphorylated receptor binds target pro-teins that contain SH2 domains.
EGFreceptor
Insulinreceptor
PDGFreceptor
FGFreceptor
KINASEDOMAINS
Kinaseinserts
Receptor protein tyrosine kinase families
FIGURE 14.34 The monomeric tyrosine kinase receptors consist of aglobular extracellular domain that binds ligand, a single transmembranespan, and a globular intracellular region containing the protein kinasedomain. The intracellular regions contain additional sequences preced-ing, following, and, in the case of the PDGF and FGF receptor groups,inserted into the protein kinase domain. These regions contain sites oftyrosine phosphorylation-dependent interactions. The insulin receptoris encoded by a single gene. The precursor is proteolyzed into � and �subunits, which are disulfide bonded to each other. Disulfide bonds alsolink two � subunits, yielding an obligate heterotetramer.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 629

630 CHAPTER 14 Principles of cell signaling
transducers. This array of signaling events leadsto increased proliferation of connective tissueduring development and in wound healing.
Autophosphorylation also occurs on the in-sulin receptor to stabilize the active state and togenerate a smaller number of binding sites, as il-lustrated in FIGURE 14.36. A key event is the Tyrphosphorylation of insulin receptor substrate(IRS) proteins, notably IRS1, on as many as adozen sites. IRS1 takes over interactions with sev-eral signaling effectors that, in the case of PDGF,bind directly to the receptor. Among these tar-gets is PI 3-kinase which leads to activation ofAkt-2 and several essential metabolic actions ofinsulin (see 14.16 Lipids and lipid-derived compoundsare signaling molecules). IRS proteins are also phos-phorylated by serine/threonine protein kinases tomodulate their signaling capability.
Tyr phosphorylation often enhances the en-zymatic activity of the associated proteins. Otherproteins gain enhanced function primarily as aconsequence of greater proximity to targetsachieved by binding through their SH2 domainsto phosphotyrosine sites either on the recep-tors or IRS adaptors. The precise actions of themany tyrosine kinase receptors are determinedby the overlapping sets of signal transducerswith which they interact, as well as by detaileddifferences in amounts of signal transducers,adaptor accessory proteins and receptor expres-sion patterns (see Figure 14.43).
Src family protein kinasescooperate with receptorprotein tyrosine kinases
The first protein tyrosine kinase to be discoveredwas Src, which was identified as the transform-ing entity in the Rous sarcoma virus. Src is theprototype of a number of related enzymes, theSrc family kinases. It participates in signalingpathways regulated by numerous cell surfacereceptors, including those that lack their ownkinase domain (see in 14.34 Diverse receptors re-cruit protein tyrosine kinases to the plasma mem-brane). Src is bound to the plasma membranevia an N-terminal myristoyl group. In the in-active state, Src is phosphorylated on Tyr527, C-terminal to its catalytic domain, by CSK(C-terminal Src kinase).
The structure and regulation of Src is de-picted in FIGURE 14.37. Phosphorylation of Tyr527causes it to bind to its own SH2 domain. TheSH2 and SH3 domains suppress the kinase ac-tivity through interactions on the surface of theprotein. The SH3 domain binds to an SH3 bind-ing site distant from the active site. Activationof Src by dephosphorylation of Tyr527 causesits SH2 to dissociate; this causes a conforma-tional change in the SH3 domain to dissociateit from the binding site. Viral isolates of Src areoften truncated prior to Tyr527, which increasestheir activity.
Conformational changes in the kinase do-main resulting from dissociation of the SH3 pro-mote Src autophosphorylation on Tyr416 in itsactivation loop and further increase protein ki-nase activity. An important consequence of theinteraction of Src with its own SH2 and SH3domains is that these domains cannot bind any-thing else when in the autoinhibited state; there-fore, other interactions are promoted when theSH2 and SH3 domains are released from theirassociations with the Src kinase domain. Theheterologous interactions of the SH2 and SH3domains contribute to Src localization and sig-naling.
Key concepts • Src is activated by release of intrasteric inhibition.• Activation of Src involves liberation of modular
binding domains for activation-dependentinteractions.
• Src often associates with receptors, includingreceptor tyrosine kinases.
14.31
PIP2 PIP3
Aktsignaling
CYTOPLASM
Insulinreceptor
p85 p110
IRS1PI 3-kinase
Insulin signaling through IRS1
Insulin
FIGURE 14.36 Insulin binding to its receptor causes activation of thereceptor tyrosine protein kinase and autophosphorylation. The receptorkinase also phosphorylates IRS1, a large adaptor with many potentialphosphorylation sites. IRS1 is an essential intermediate in insulin ac-tion. PI 3-kinase binds to IRS1 via the SH2 domain within its p85 sub-unit. Akt and PDK1 bind to PIP3 produced by activated PI 3-kinase so thatPDK1 can phosphorylate and activate Akt (see Figure 14.18).
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 630

14.32 MAPKs are central to many signaling pathways 631
MAPKs are central tomany signaling pathways
Mitogen-activated protein kinases (MAPKs) arepresent in all eukaryotes. They are among themost common multifunctional protein kinasesmediating cellular regulatory events in responseto many ligands and other stimuli. MAPKs areactivated by protein kinase cascades consistingof at least three protein kinases acting sequen-tially, as illustrated in FIGURE 14.38. Activationof a MAPK is catalyzed by a MAPK kinase(MAP2K), which is itself activated by phospho-rylation by a MAPK kinase kinase (MAP3K).MAP3Ks are activated by a variety of mecha-nisms including phosphorylation by MAP4Ks,oligomerization, and binding to activators suchas small G proteins.
Key concepts • MAPKs are activated by Tyr and Thr
phosphorylation.• The requirement for two phosphorylations creates
a signaling threshold.• The ERK1/2 MAPK pathway is usually regulated
through Ras.
14.32MAP2Ks are activated by phosphorylation
on two Ser/Thr residues; MAP2Ks then acti-vate MAPKs by dual phosphorylation on Tyrand Thr residues (Figure 14.30). Each MAP2Kphosphorylates a limited set of MAPKs and fewor no other substrates. The great specificity ofMAP2Ks is one means of insulating MAPKsfrom activation by inappropriate signals. BothTyr and Thr phosphorylations are required formaximum MAPK enzymatic activity.
Studies on the MAPK ERK2 led to an un-derstanding of the events induced by phospho-rylation that are important for increased activity.Conformational changes include refolding ofthe activation loop to improve substrate posi-tioning and realignment of catalytic residues;this is most obvious in the repositioning of �helix C, which contains a Glu involved in phos-phoryl transfer.
Amplification occurs moving down the cas-cade from the MAP3K to the MAP2K step be-cause the MAP2Ks are much more abundantthan the MAP3Ks. The MAP2K to MAPK stepmay also amplify the signal if the MAPK is pres-ent in excess of the MAP2K. In addition, thephosphorylation of a MAPK by a MAP2K on a
SH3
SH2
KINASEDOMAIN
Tyr527Tyr527
Structure and regulation of Src
P P
INACTIVE ACTIVE
FIGURE 14.37 The structures of inactive and active Src are compared. The in-active protein is autoinhibited by binding to its own SH2 and SH3 domains.The SH2 domain binds to phosphorylated Tyr527. The SH3 domain binds to anoncanonical SH3-binding motif on the opposite side of the kinase domainactive site. In contrast to the steric inhibition of PKA caused by its R subunit,inhibition of Src by its SH2 and SH3 domains is allosteric. In the active struc-ture the SH2 and SH3 domains are not bound to the kinase domain and areavailable for heterologous interactions. Structures generated from Protein DataBase files 1FMK and 1Y57.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 631

632 CHAPTER 14 Principles of cell signaling
Tyr and a Thr residue creates cooperative acti-vation of the MAPK; this is another mechanism,in addition to those described for PKA andcalmodulin, to introduce a threshold and ap-parently cooperative behavior into the path-way over a narrow range of input signal. Thismultistep cascade provides multiple sites formodulatory inputs from other pathways.
Stabilized interactions between componentsare also important. MAP2Ks, as well as MAPKsubstrates and MAPK phosphatases, generallycontain a basic/hydrophobic docking motif thatinteracts with acidic residues and binds in a hy-drophobic groove on the MAPK catalytic do-main. Additional components including scaffoldsare necessary for the efficient activation ofMAPK cascades in cells and usually have addi-
tional functions. Several scaffolds have beenidentified that bind to two or more componentsfor each of the three major MAPK cascades, theERK1/2, JNK1-3, and p38 α, β, γ, and δ cascades.
The ERK1/2 pathway is regulated by mostcell surface receptors, including receptors thatemploy tyrosine kinases, GPCRs, and others. ThePDGF receptor, like most receptor systems, acti-vates the ERK1/2 cascade through Ras. PDGFstimulates autophosphorylation of its receptorand the subsequent association of effectors withits cytoplasmic domain (see 14.30 Diverse signal-ing mechanisms are regulated by protein tyrosine ki-nases). In response to PDGF, ERK1/2 promotes cellproliferation and differentiation by phosphory-lation of membrane enzymes, proteins involvedin determining cell shape and motility, and alsoby concentrating in the nucleus to phosphory-late regulatory factors that control transcription.
Cyclin-dependent proteinkinases control the cellcycle
Cell division is regulated positively and nega-tively by factors that stimulate proliferation andinputs that monitor cell state. The sum of thesefactors is integrated in the regulation of cyclin-dependent protein kinases (CDKs). CDKs areprotein serine/threonine kinases that are ma-jor regulators of cell cycle progression. MostCDKs are regulated both by kinases and phos-phatases and by association with other proteinscalled cyclins. Cyclins are synthesized and de-graded every cell cycle. Because most CDKs aredependent upon cyclin binding for activation,the timing of the synthesis and degradation ofindividual cyclins determines when a CDK willfunction. The most notable noncycling memberof the CDK family is Cdk5, which is highly ex-pressed in terminally differentiated neurons.Cdk5 binds the non-cyclin protein p35 as its ac-tivating subunit.
We will briefly examine the regulation ofCdc2, a major CDK in both mammals and yeast.The first step in regulation of Cdc2 is the asso-ciation with cyclin. A second step required foractivation of Cdc2 is phosphorylation of a Thr
Key concepts • The cell cycle is regulated by cyclin-dependent
protein kinases (CDKs).• Activation of CDKs involves protein binding,
dephosphorylation, and phosphorylation.
14.33
MAP4K
G Protein
MAP3K
MAP2K
MAPK
Ste20p Ste20family
Ste20family
Ste11p
Ste7p
Fus3p
Ste12p
Mating
Ras ?
Output
PAK/PKC ?
S.cerevisiae MammalsGeneric
small orheterotrimer
Gβγ
Major targets:
ProliferationDevelopment
Differentiation(and other processes)
Rac/Cdc42 Rac
RafMEKK2MEKK3
many many
MEK5
ERK5
MEK1MEK2
MEK4MEK7
MEK3MEK6
ERK1ERK2
JNK1JNK2JNK3
p38αp38βp38γp38δ
Elk-1
Rsk Rsk
c-JunATF2
MEF2 MEF2
MAPKAPK2Proteinkinase
Trans-criptionfactor
MAPK pathways
FIGURE 14.38 MAPK pathways can be regulated by a diverse group of upstreamregulatory mechanisms that often include adaptors, small G proteins, and MAP4Ks.These molecules impinge on the activities of MAP3Ks. MAP3Ks regulate one ormore MAP2Ks depending on localization and scaffolding. The MAP2Ks displaygreat selectivity for a single MAPK type. MAPKs have overlapping and uniquesubstrates and participate in signaling cascades leading to many cellular responses.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 632

14.34 Diverse receptors recruit protein tyrosine kinases to the plasma membrane 633
residue in its activation loop by another CDKtype kinase. In spite of its association with cy-clin, this form of Cdc2 is not yet active due toinhibitory phosphorylation of Tyr and Thrresidues in the ATP binding pocket. Release ofinhibition by dephosphorylation of the residuesin the ATP pocket is catalyzed by the Cdc25 fam-ily of phosphoprotein phosphatases, resulting inactivation of Cdc2. The proximity of the Tyrresidue to catalytic residues is shown in FIGURE14.39. The complexity of activation of CDKsmakes possible the imposition of cell cycle check-points. For more on CDKs and cyclins see 11.4The cell cycle is a cycle of CDK function.
Diverse receptors recruitprotein tyrosine kinasesto the plasma membrane
Key concepts • Receptors that bind protein tyrosine kinases use
combinations of effectors similar to those used byreceptor tyrosine kinases.
• These receptors often bind directly to transcriptionfactors.
14.34
Many receptors act through protein tyrosinekinases, but their cell surface receptors lack ki-nase activity. Instead, these receptors act by re-cruiting and activating protein tyrosine kinasesat the plasma membrane. In this group of recep-tors are integrins, which are key molecules in-volved in cell adhesion, growth hormonereceptors, and receptors that mediate inflam-matory and immune responses. While theirstructures vary enormously, their mechanismsof action are related.
Integrins are receptors whose major functionis to attach cells to the extracellular matrix. Theyalso mediate some interactions with proteins onother cells, as depicted in FIGURE 14.40. Ligands forintegrins include a number of extracellular ma-trix proteins, such as fibronectin, as well as cellsurface proteins that cooperate in cell-cell inter-actions. Integrin ligation provides cells with infor-mation about their environment that influencescell behavior. Ligation of integrins initiates signalsthat control cell programs, including cell cycle en-try, proliferation, survival, differentiation, changesin cell shape, and motility, as well as fine-tuningresponses to other ligands. For more details on in-tegrins see 15.13 Most integrins are receptors for extra-cellular matrix proteins and 15.14 Integrin receptorsparticipate in cell signaling.
Lys33Lys33
Cdk2Cdk2
Cyclin ACyclin A
Tyr15Tyr15
Glu51Glu51
CDKs require cyclin binding for activation
FIGURE 14.39 The view of the crystal structure of CDK2 bound to cyclin Ashows residues in the ATP binding site. The enlargement on the right showsthe interaction between Lys33 and Glu51, catalytic residues that interact withATP to promote phosphoryl transfer. Tyr15 is phosphorylated in inactive formsof CDK2. A phosphoryl group on Tyr15 inhibits CDK activity by interfering withATP binding. Structure generated from Protein Data Bank file 1JST.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 633

634 CHAPTER 14 Principles of cell signaling
Talin and α-actinin are among cytoskeletalproteins that interact directly with certain inte-grin subunits. These cytoskeletal proteins linkintegrins to complex cytoskeletal structuresknown as focal adhesions.
Focal adhesions connect the cytoskeleton tosignal transduction cascades that communicatestates of cellular attachment to the regulation ofcellular responses. Focal adhesion complexes con-tain the focal adhesion kinase FAK, which is ac-tivated by integrin ligation. Autophosphorylationof FAK recruits signaling proteins containing SH2domains, especially the p85 subunit of PI 3-ki-nase and Src family protein kinases. The signal-ing molecules associated with the integrin-boundcytoskeletal proteins, whether focal adhesions orother structural complexes, mediate the diverseactions of integrins. The association of cytoskele-tal proteins with integrin receptors also causesfunctional changes to the receptors.
Signals that act over a distance, such as hor-mones, can also employ nonreceptor tyrosinekinases to transmit their message inside a cell.Growth hormone (GH) is a protein hormonesecreted by the anterior pituitary gland that reg-ulates bone growth, fat metabolism, and other
cellular growth phenomena. Absence of growthhormone results in short stature, whereas hy-persecretion causes acromegaly, a form of gi-gantism. The GH receptor is a member of thecytokine receptor family, which includes recep-tors for prolactin, erythropoietin, leptin, andinterleukins. All these receptors display similarbiochemical functions, such as association withmembers of the JAK/TYK family of protein ty-rosine kinases, but select for different but over-lapping sets of cytoplasmic signaling proteins.Signal transduction by the GH receptor providesa model for receptors that lack enzymatic func-tion and act as agonist-promoted scaffolds forintracellular signaling proteins.
FIGURE 14.41 shows the structure of growthhormone bound to the extracellular domain ofits receptor. The majority of binding energycomes from only a small number of residues inthe binding interface. Inside the cell, signalingby the GH receptor depends significantly on itsassociation with the cytoplasmic tyrosine pro-tein kinase Janus kinase 2 (JAK2). FIGURE 14.42shows that JAK2 binds to a proline-rich regionof the receptor. Ligand binding induces recep-tor dimerization, which then promotes activation of JAK2 through intermolecular auto-phosphorylation.
GH signaling is thus mediated primarily byinducing Tyr phosphorylation. In addition toJAK2 autophosphorylation, the receptor itselfbecomes Tyr phosphorylated. As is true for recep-tor tyrosine kinases, Tyr phosphorylation of thegrowth hormone receptor creates binding sitesfor signaling proteins that contain phosphotyro-sine-binding domains. Primary targets are tran-scription factors known as signal transducers andactivators of transcription, or STATs. STATs con-tain SH2 domains and bind Tyr-phosphorylatedmotifs on the growth hormone receptor. Whilereceptor bound, STATs are Tyr phosphorylatedby JAK2 and then released to travel to the nu-cleus to mediate changes in transcription.
The growth hormone receptor and the as-sociated JAK2 also activate other signaling path-ways. For example, the adaptor Shc is Tyrphosphorylated by JAK2. Engagement of Shcleads to activation of Ras and the ERK1/2 MAPKpathway. Adaptors specialized for insulin-signal-ing pathways, insulin receptor substrates (IRS)1, 2, and 3, are also growth hormone targets, per-haps reflecting the ability of growth hormoneto induce certain insulin-like metabolic actions.
Feedback circuits are also engaged duringGH signaling. The growth hormone receptorcomplex binds the adaptor SH2-B, which has a
CYTOPLASM
ECM
Paxillin
Actinfilament
INTEGRINS
VinculinVinculin
TalinTalin
Tensin
TalinTalinPI 3-
Kinase
ILK
p85
p110
Src
Grb2
SOS
FAKCASCrk
Crk
Integrin signaling
FIGURE 14.40 Integrins bind to an array of cytoplasmic proteins to regulatethe cytoskeleton and intracellular signaling pathways. The associated cytoskele-tal elements include actin filaments and focal adhesion proteins �-actinin, vin-culin, paxillin, and talin. Signaling molecules include the focal adhesion kinaseFAK; the adaptors Cas, Crk and Grb2; Src and CSK (see 14.31 Src family proteinkinases cooperate with receptor protein tyrosine kinases), PI 3-kinase (see 14.16Lipids and lipid-derived compounds are signaling molecules); and the Ras ex-change factor SOS. Stimulation of GTP binding of Ras by SOS leads to activa-tion of the MAPK pathway (see 14.32 MAPKs are central to many signalingpathways).
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 634

14.34 Diverse receptors recruit protein tyrosine kinases to the plasma membrane 635
stimulatory effect on growth hormone signal-ing. On the other hand, suppressors of cytokinesignaling (SOCS proteins) are among the geneswhose transcription is induced by growth hor-mone. As the name indicates, SOCS proteins in-hibit cytokine signaling in some if not all cases byinhibiting the activity of JAK2. SOCS proteinscontain an SH2 domain that facilitates their bind-ing either to phosphorylated JAK2 or cytokinereceptors. The mechanism of signaling inhibi-tion may differ among SOCS proteins becausesome require the GH receptor to interfere withJAK2 signaling. SOCS-1, on the other hand, bindsdirectly to the JAK2 activation loop and does notrequire a receptor to inhibit JAK2 activity. Thismechanism may be particularly important in GHsignaling because, in contrast to the ligand-in-
duced down regulation mechanisms controllingmany receptors, the GH receptor is degraded ina ligand-independent manner.
Receptors for cytokines also act by recruit-ing tyrosine kinases. The cytokines—signalingproteins that modulate inflammation and cellgrowth and differentiation—include interleukins,leukemia inhibitory factor, oncostatin M, car-diotrophin-1, cardiotrophin-like cytokine, andciliary neurotrophic factor (CNTF). Each cy-tokine binds a unique receptor, but each recep-tor binds a transmembrane protein called gp130.Mechanisms of signaling by gp130 involve in-teractions with tyrosine kinases of the JAK/TYKtypes and transcription factors in the STAT fam-ily. This mechanism is similar to those employedby the growth hormone receptor.
Growth hormonereceptor
hGH
ΔΔG (kcal/mol)
> 1.50.5 to 1.5-0.5 to 0.5< -0.5untested
Growth hormone structure
FIGURE 14.41 Proteins often interact over a large surface area. Growthhormone binding to its receptor is an example of the energy of bind-ing coming primarily from a small number of the contacts betweenthe two proteins, creating an interaction hot spot. The complex ofgrowth hormone bound to the growth hormone receptor-binding do-main determined by crystallography has been peeled apart in this fig-ure to show the binding energy associated with residues in the bindinginterface from each protein determined by mutagenesis and bindingstudies. Fewer than half of the residues in the interface contributethe majority of binding energy. Reproduced with permission from T.Clackson and J. A. Wells. 1995. Science. 267: 383–386. © AAAS. Photoscourtesy of Tim Clackson, ARIAD Pharmaceuticals, Inc.
NUCLEUS
CYTOPLASM
Phosphorylated STATs bind DNA
STATSTAT
JAK2JAK2
Dimerization
STATs bind andare phosphorylated
JAK2s bind and phosphorylate receptor
Growth hormone signaling is transduced by JAK2
Growthhormone
FIGURE 14.42 The growth hormone receptor binds to JAK2. Many GH signalsare mediated by Tyr phosphorylation of the receptor by JAK2, which createsbinding sites for signaling molecules with SH2 domains, notably STATs. STATsthen enter the nucleus to cause changes in gene transcription.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 635
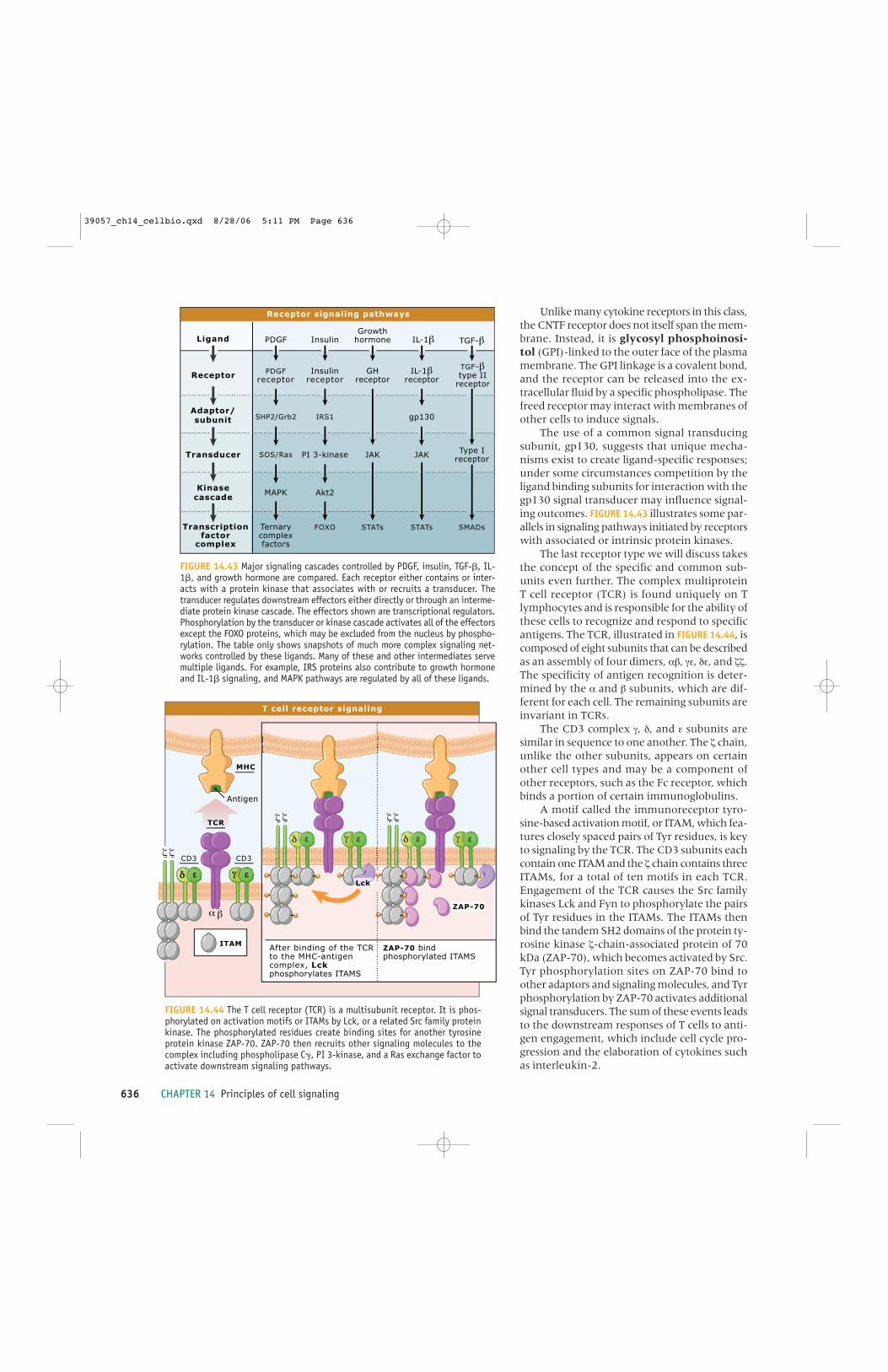
636 CHAPTER 14 Principles of cell signaling
Unlike many cytokine receptors in this class,the CNTF receptor does not itself span the mem-brane. Instead, it is glycosyl phosphoinosi-tol (GPI)-linked to the outer face of the plasmamembrane. The GPI linkage is a covalent bond,and the receptor can be released into the ex-tracellular fluid by a specific phospholipase. Thefreed receptor may interact with membranes ofother cells to induce signals.
The use of a common signal transducingsubunit, gp130, suggests that unique mecha-nisms exist to create ligand-specific responses;under some circumstances competition by theligand binding subunits for interaction with thegp130 signal transducer may influence signal-ing outcomes. FIGURE 14.43 illustrates some par-allels in signaling pathways initiated by receptorswith associated or intrinsic protein kinases.
The last receptor type we will discuss takesthe concept of the specific and common sub-units even further. The complex multiproteinT cell receptor (TCR) is found uniquely on Tlymphocytes and is responsible for the ability ofthese cells to recognize and respond to specificantigens. The TCR, illustrated in FIGURE 14.44, iscomposed of eight subunits that can be describedas an assembly of four dimers, αβ, γε, δε, and ζζ.The specificity of antigen recognition is deter-mined by the α and β subunits, which are dif-ferent for each cell. The remaining subunits areinvariant in TCRs.
The CD3 complex γ, δ, and ε subunits aresimilar in sequence to one another. The ζ chain,unlike the other subunits, appears on certainother cell types and may be a component ofother receptors, such as the Fc receptor, whichbinds a portion of certain immunoglobulins.
A motif called the immunoreceptor tyro-sine-based activation motif, or ITAM, which fea-tures closely spaced pairs of Tyr residues, is keyto signaling by the TCR. The CD3 subunits eachcontain one ITAM and the ζ chain contains threeITAMs, for a total of ten motifs in each TCR.Engagement of the TCR causes the Src familykinases Lck and Fyn to phosphorylate the pairsof Tyr residues in the ITAMs. The ITAMs thenbind the tandem SH2 domains of the protein ty-rosine kinase ζ-chain-associated protein of 70kDa (ZAP-70), which becomes activated by Src.Tyr phosphorylation sites on ZAP-70 bind toother adaptors and signaling molecules, and Tyrphosphorylation by ZAP-70 activates additionalsignal transducers. The sum of these events leadsto the downstream responses of T cells to anti-gen engagement, which include cell cycle pro-gression and the elaboration of cytokines suchas interleukin-2.
After binding of the TCR to the MHC-antigen complex, Lck phosphorylates ITAMS
MHC
Antigen
CD3 CD3
TCR
Lck
ITAM
ZAP-70
T cell receptor signaling
FIGURE 14.44 The T cell receptor (TCR) is a multisubunit receptor. It is phos-phorylated on activation motifs or ITAMs by Lck, or a related Src family proteinkinase. The phosphorylated residues create binding sites for another tyrosineprotein kinase ZAP-70. ZAP-70 then recruits other signaling molecules to thecomplex including phospholipase C�, PI 3-kinase, and a Ras exchange factor toactivate downstream signaling pathways.
Ligand
Receptor
Adaptor/subunit
Kinasecascade
Transducer
Transcriptionfactor
complex
PDGF IL-1β TGF-βGrowth
hormone
PDGFreceptor
IL-1βreceptor
TGF-βtype II
receptor
GHreceptor
SHP2/Grb2 gp130
Type IreceptorSOS/Ras
MAPK
SMADsSTATs STATs
Insulin
Insulinreceptor
IRS1
PI 3-kinase
Akt2
FOXOTernarycomplexfactors
JAK JAK
Receptor signaling pathways
FIGURE 14.43 Major signaling cascades controlled by PDGF, insulin, TGF-�, IL-1�, and growth hormone are compared. Each receptor either contains or inter-acts with a protein kinase that associates with or recruits a transducer. Thetransducer regulates downstream effectors either directly or through an interme-diate protein kinase cascade. The effectors shown are transcriptional regulators.Phosphorylation by the transducer or kinase cascade activates all of the effectorsexcept the FOXO proteins, which may be excluded from the nucleus by phospho-rylation. The table only shows snapshots of much more complex signaling net-works controlled by these ligands. Many of these and other intermediates servemultiple ligands. For example, IRS proteins also contribute to growth hormoneand IL-1� signaling, and MAPK pathways are regulated by all of these ligands.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 636

References 637
What’s next?New signaling proteins and new regulatory in-teractions seem to show up every day. The chal-lenge now is to understand how cells organizethese proteins and their individual interactionsto create adaptable information-processing net-works. How do cells use simple chemical reac-tions to sort and integrate multiple simultaneousinputs and then direct this information to di-verse effector machinery? How do they inter-pret the inputs in the context of their growth andmetabolic activities? In principle, three areas ofresearch have to contribute to allow us to un-derstand integrative cellular signaling.
First, we need real-time, noninterferingbiosensors to measure intracellular signalingreactions. Most current sensors use combina-tions of fluorescent moieties and signal-bind-ing protein domains to provide fast opticalreadouts. For many pathways, several reactionscan be monitored within cells over subsecondtime scales. We need more, better, and fastersensors and sensors that can report with single-cell and subcellular resolution. Genetically en-coded sensors will be complemented by syntheticmolecules.
Our ability to manipulate signaling net-works is also improving dramatically but stillfalls short. We can manipulate signaling net-works by overexpression, knockout, and knock-down of genes, but signaling pathways arewonderfully adaptive and frequently circum-vent our best efforts to control them. We stillneed chemical regulators that can act promptlyin cells. Structure-based design of such regula-tory molecules will be vital.
Last, our ability to analyze the behavior ofsignaling networks depends on our ability tomeasure and interpret signaling quantitatively.It is ironic but true that really complex systemscannot be described without explicit quantita-tive models for how they work. Computationalmodeling and simulation of signaling networksrequires both better theoretical understandingof network dynamics and better algorithmic im-plementation.
The goal is to understand how cells think.
SummarySignal transduction encompasses mechanismsused by all cells to sense and react to stimuli intheir environment. Cells express receptors thatrecognize specific extracellular stimuli, includ-
14.36
14.35ing nutrients, hormones, neurotransmitters,and other cells. Upon receptor binding, signalsare converted to well-defined intracellular chem-ical or physical reactions that change the activ-ities and the organization of protein complexeswithin cells. The changes directed by the stim-uli lead to altered cell behavior. The behavior ofthe cell is determined then by its intracellularstate and the integrated information from ex-tracellular stimuli so that the appropriate re-sponses are achieved.
The basic biochemical components andprocesses of signal transduction are conservedthroughout biology. Families of proteins areused in a variety of ways for many differentphysiological purposes. Cells often use the sameseries of signaling proteins to regulate multipleprocesses, such as transcription, ion transport,locomotion, and metabolism.
Signaling pathways are assembled into sig-naling networks to allow the cell to coordinateits responses to multiple inputs with its ongo-ing functions. It is now possible to discern con-served reaction sequences in and betweenpathways in signaling networks that are anal-ogous to devices within the circuits of analogcomputers: amplifiers, logic gates, feedback andfeed-forward controls, and memory.
ReferencesIntroduction
ReviewSauro, H. M. and Kholodenko, B. N., 2004.
Quantitative analysis of signaling net-works. Prog. Biophys. Mol. Biol. v. 86 p.5–43.
ResearchMilo, R., Shen-Orr, S., Itzkovitz, S., Kashtan,
N., Chklovskii, D., and Alon, U., 2002.Network motifs: simple building blocks ofcomplex networks. Science v. 298 p.824–827.
Cellular signaling is primarily chemical
ReviewArshavsky, V. Y., Lamb, T. D., and Pugh, E. N.,
Jr., 2002. G proteins and phototransduc-tion. Annu. Rev. Physiol. v. 64 p. 153–187.
Caterina, M. J. and Julius, D., 2001. Thevanilloid receptor: a molecular gatewayto the pain pathway. Annu. Rev. Neurosci.v. 24 p. 487–517.
Gillespie, P. G. and Cyr, J. L., 2004. Myosin-1c, the hair cell’s adaptation motor. Annu.Rev. Physiol. v. 66 p. 521–545.
14.2
14.1
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 637

638 CHAPTER 14 Principles of cell signaling
Lin, C. and Shalitin, D., 2003. Cryptochromestructure and signal transduction. Annu.Rev. Plant Biol. v. 54 p. 469–496.
Rockwell, N.C., Su, Y.-S., and Lagarias, J.C.,2006. Phytochrome structure and signal-ing mechanisms. Annu. Rev. Plant Biology,v. 57 p. 837–858.
Sancar, A., 2000. Cryptochrome: the secondphotoactive pigment in the eye and itsrole in circadian photoreception. Annu.Rev. Biochem. v. 69 p. 31–67.
Receptors sense diverse stimuli but initi-ate a limited repertoire of cellular signals
ResearchKlein, C., Paul, J. I., Sauvé, K., Schmidt, M.
M., Arcangeli, L., Ransom, J., Trueheart,J., Manfredi, J. P., Broach, J. R., andMurphy, A. J., 1998. Identification of sur-rogate agonists for the human FPRL-1 re-ceptor by autocrine selection in yeast.Nat. Biotechnol. v. 16 p. 1334–1337.
Ligand binding changes receptor confor-mation
ReviewRoss, E. M., and Kenakin, T. P., 2001.
Pharmacodynamics: Mechanisms of drugaction and the relationship between drugconcentration and effects. In Goodmanand Gilmans’s The Pharmacological Basis ofTherapeutics, 10th. Ed., J. G. Hardman andL. E. Limbird, eds., New York: McGraw-Hill, p. 31–43.
Signals are sorted and integrated in sig-naling pathways and networks
ResearchItzkovitz, S., Milo, R., Kashtan, N., Ziv, G.,
and Alon, U., 2003. Subgraphs in randomnetworks. Phys. Rev. E. v. 68 p. 126–127.
Cellular signaling pathways can bethought of as biochemical logic circuits
ReviewMilo, R., Shen-Orr, S., Itzkovitz, S., Kashtan,
N., Chklovskii, D., and Alon, U., 2002.Network motifs: Simple building blocks ofcomplex networks. Science v. 298 p.824–827.
ResearchTorres, E. and Rosen, M. K., 2003. Contingent
phosphorylation/dephosphorylation pro-vides a mechanism of molecular memoryin WASP. Mol. Cell v. 11 p. 1215–1227.
Scaffolds increase signaling efficiencyand enhance spatial organization of sig-naling
14.8
14.7
14.6
14.5
14.3
ReviewElion, E. A., 2001. The Ste5p scaffold. J. Cell
Sci. v. 114 p. 3967–3978.Kholodenko, B. N., 2003. Four-dimensional
organization of protein kinase signalingcascades: the roles of diffusion, endocyto-sis and molecular motors. J. Exp. Biol. v.206 p. 2073–2082.
O’Rourke, S. M., Herskowitz, I., and O’Shea,E. K., 2002. Yeast go the whole HOG forthe hyperosmotic response. Trends Genet.v. 18 p. 405–412.
Pawson, T. and Nash, P., 2003. Assembly ofcell regulatory systems through proteininteraction domains. Science v. 300 p.445–452.
Tsunoda, S. and Zuker, C. S., 1999. The or-ganization of INAD-signaling complexesby a multivalent PDZ domain protein inDrosophila photoreceptor cells ensuressensitivity and speed of signaling. CellCalcium v. 26 p. 165–171.
ResearchLevchenko, A., Bruck, J., and Sternberg, P.
W., 2000. Scaffold proteins may biphasi-cally affect the levels of mitogen-acti-vated protein kinase signaling and reduceits threshold properties. Proc. Natl. Acad.Sci. USA v. 97 p. 5818–5823.
Rensland, H., Lautwein, A., Wittinghofer, A.,and Goody, R. S., 1991. Is there a rate-limiting step before GTP cleavage by H-ras p21? Biochemistry v. 30 p. 11181–11185.
Shenker, A., Goldsmith, P., Unson, C. G., andSpiegel, A. M., 1991. The G protein cou-pled to the thromboxane A2 receptor inhuman platelets is a member of the novelGq family. J. Biol. Chem. v. 266 p. 9309–9313.
Independent, modular domains specifyprotein-protein interactions
ReviewPawson, T. and Nash, P., 2003. Assembly of
cell regulatory systems through proteininteraction domains. Science v. 300 p.445–452.
Turk, B. E. and Cantley, L. C., 2003. Peptidelibraries: at the crossroads of proteomicsand bioinformatics. Curr. Opin. Chem. Biol.v. 7 p. 84–90.
ResearchGinty, D. D., Kornhauser, J. M., Thompson,
M. A., Bading, H., Mayo, K. E.,Takahashi, J. S., and Greenberg, M. E.,1993. Regulation of CREB phosphoryla-tion in the suprachiasmatic nucleus bylight and a circadian clock. Science v. 260p. 238–241.
14.9
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 638

References 639
Cellular signaling is remarkably adaptive
ReviewPerkins, J. P., Hausdorff, W. P., and Lefkowitz,
R. J., 1990. Mechanisms of ligand-in-duced desensitization of �-adrenergic re-ceptors. In The Beta-Adrenergic Receptors, J.P. Perkins, ed., Clifton, NJ: Humana Press,p. 73–124.
Signaling proteins are frequently ex-pressed as multiple species
ReviewBarnes, N. M., and Sharp, T., 1999. A review
of central 5-HT receptors and their func-tion. Neuropharmacology v. 38 p. 1083–1152.
Gilman, A. G., 1987. G proteins: transducersof receptor-generated signals Annu. Rev.Biochem. v. 56 p. 615–649.
Rebecchi, M. J., and Pentyala, S. N., 2000.Structure, function, and control of phos-phoinositide-specific phospholipase C.Physiol. Rev. v. 80 p. 1291–1335.
Sunahara, R. K. and Taussig, R., 2002.Isoforms of mammalian adenylyl cyclase:Multiplicities of signaling. Mol.Interventions v. 2 p. 168–184.
Second messengers provide readily dif-fusible pathways for information transfer
ReviewBeavo, J. A., Bechtel, P. J., and Krebs, E. G.,
1975. Mechanisms of control for cAMP-dependent protein kinase from skeletalmuscle. Adv. Cyclic Nucleotide Res. v. 5 p.241–251.
Kobe, B., Heierhorst, J., and Kemp, B. E.,1997. Intrasteric regulation of protein ki-nases. Adv. Second Messenger PhosphoproteinRes. v. 31 p. 29–40.
Wong, W. and Scott, J. D., 2004. AKAP sig-nalling complexes: focal points in spaceand time. Nat. Rev. Mol. Cell Biol. v. 5 p.959–970.
ResearchRall, T. W. and Sutherland, E. W., 1958.
Formation of a cyclic adenine ribonu-cleotide by tissue particles. J. Biol. Chem. v.232 p. 1065–1076.
Ca2+ signaling serves diverse purposes inall eukaryotic cells
ReviewNewton, A. C., 2001. Protein kinase C: struc-
tural and spatial regulation by phospho-rylation, cofactors, and macromolecularinteractions. Chem. Rev. v. 101 p.2353–2364.
14.15
14.14
14.11
14.10ResearchClapperton, J. A., Martin, S. R., Smerdon, S.
J., Gamblin, S. J., and Bayley, P. M.,2002. Structure of the complex ofcalmodulin with the target sequence ofcalmodulin dependent protein kinase I:Studies of the kinase activation mecha-nism. Biochemistry. v. 41 p. 14669–14679.
Kuboniwa, H., Tjandra, N., Grzesiek, S., Ren,H., Klee, C.B., Bax, A., 1995. Solutionstructure of calcium-free calmodulin. Nat.Struct. Biol. v. 2 p. 768–776.
Lipids and lipid-derived compounds aresignaling molecules
ReviewRebecchi, M. J., and Pentyala, S. N., 2000.
Structure, function, and control of phos-phoinositide-specific phospholipase C.Physiol. Rev. v. 80 p. 1291–1335.
Yang, C. and Kazanietz, M. G., 2003.Divergence and complexities in DAG sig-naling: looking beyond PKC. TrendsPharmacol. Sci. v. 24 p. 602–608.
PI 3-kinase synthesizes a lipid activatorof cell motion and shape change
ReviewDownward, J., 2004. PI 3-kinase, Akt and cell
survival. Semin. Cell Dev. Biol. v. 15 p.177–182.
Lawlor, M. A. and Alessi, D. R., 2001.PKB/Akt: a key mediator of cell prolifera-tion, survival and insulin responses? J.Cell Sci. v. 114 p. 2903–2910.
Van Haastert, P. J. and Devreotes, P. N. 2004.Chemotaxis: signaling the way forward.Nat. Rev. Mol. Cell Biol. v. 5. p. 626–634.
Signaling through ion channel receptorsis very fast
ReviewClapham, D. E., 2003. TRP channels as cellu-
lar sensors. Nature v. 426 p. 517–524.Corey, D. P., 2003. New TRP channels in
hearing and mechanosensation. Neuron v.39 p. 585–588.
Hille, B, 1992. Ionic channels of excitablemembranes. Sinauer Associates.
Siegelbaum, S. A. and Koester, J., 2000. Ionchannels and Membrane potential, inPrinciples of Neural Science, 4th. Ed., E. R.Kandel, J. H. Schwartz, and T. M. Jessell,eds., New York: McGraw-Hill. p.105–139.
ResearchUnwin, N., 2005. Refined structure of the
nicotinic acetylcholine receptor at 4Å res-olution. J. Mol. Biol. v. 346 p. 967.
Nuclear receptors regulate transcription14.19
14.18
14.17
14.16
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 639

640 CHAPTER 14 Principles of cell signaling
ReviewMangelsdorf, D. J. et al., 1995. The nuclear
receptor superfamily: the second decade.Cell v. 83 p. 835–839.
Smith, C. L. and O’Malley, B. W., 2004.Coregulator function: a key to under-standing tissue specificity of selective re-ceptor modulators. Endocr. Rev. v. 25 p.45–71.
ResearchBrzozowski A. M., Pike, A. C., Dauter, Z.,
Hubbard, R. E., Bonn, T., Engstrom, O.,Ohman, L., Green, G. L., Gustafsson, J. A.,and Carlquist, M., 1997. Molecular basisof agonism and antagonism in the oestro-gen receptor. Nature v. 389 p. 753–758.
G protein signaling modules are widelyused and highly adaptable
ReviewClapham, D. E. and Neer, E. J., 1997. G pro-
tein �� subunits. Annu. Rev. Pharmacol.Toxicol. v. 37 p. 167–203.
Filipek, S., Teller, D. C., Palczewski, K., andStenkamp, R., 2003. The crystallographicmodel of rhodopsin and its use in studiesof other G protein-coupled receptors.Annu. Rev. Biophys. Biomol. Struct. v. 32 p.375–397.
Ross, E. M. and Wilkie, T. M., 2000. GTPase-activating proteins for heterotrimeric Gproteins: Regulators of G protein signal-ing (RGS) and RGS-like proteins. Annu.Rev. Biochem. v. 69 p. 795–827.
Ross, E. M., 1989. Signal sorting and amplifi-cation through G protein-coupled recep-tors. Neuron v. 3 p. 141–152.
Sprang, S. R., 1997. G proteins, effectors andGAPs: structure and mechanism. Curr.Opin. Struct. Biol. v. 7 p. 849–856.
Sprang, S. R., 1997. G protein mechanisms:Insights from structural analysis. Annu.Rev. Biochem. v. 66 p. 639–678.
ResearchBenke, T., Motoshima, C. A., Fox, H., Le
Trong, B. A., Teller, I., Okada, D. C.,Stenkamp, T., Yamamoto, R. E., andMiyano, M., 2000. Crystal structure ofrhodopsin: A G protein-coupled receptor.Science v. 289 p. 739–745.
Chen, C. -K., Burns, M. E., He, W., Wensel, T.G., Baylor, D. A., and Simon, M. I., 2000.Slowed recovery of rod photoresponse inmice lacking the GTPase acceleration pro-tein RGS9-1. Nature, v. 403 p. 557–560.
Palczewski, K., Kumasaka, T., Hori, T.,Behnke , C. A., Motoshima, H., Fox, B.A., Le Trong, I., Teller, D. C., Okada, T.,Stenkamp, R. E., Yamamoto, M., andMiyano, M., 2000. Crystal structure of
14.20
rhodopsin: a G protein-coupled receptor.Science v. 289 p. 739–745.
Wall, M. A., Coleman, D. E., Lee, E., Iniguez-Lluhi, J. A., Posner, B. A., Gilman, A. G.,and Sprang, S. R., 1995. The structure ofthe G protein heterotrimer Gi�1�1�2. Cellv. 83 p. 1047–1058.
Small, monomeric GTP-binding proteinsare multiuse switches
ReviewBishop, A. L. and Hall, A., 2000. Rho GTPases
and their effector proteins. Biochem. J. v.348 pt. 2 p. 241–255.
Kuersten, S., Ohno, M., and Mattaj, I. W.,2001. Nucleocytoplasmic transport: Ran,beta and beyond. Trends Cell Biol. v. 11 p.497–503.
Takai, Y., Sasaki, T., and Matozaki, T., 2001.Small GTP-binding proteins. Physiol. Rev.v. 81 p. 153–208.
Protein phosphorylation/dephosphoryla-tion is a major regulatory mechanism inthe cell
ReviewCohen, S., 1983. The epidermal growth factor
(EGF). Cancer v. 51 p. 1787–1791.Fischer, E. H., 1997. “Protein phosphorylation
and cellular regulation, II,” in NobelLectures, Physiology or Medicine 1991–1995,N., Ringertz, Ed., Singapore: WorldScientific Publishing Co.
Krebs, G., 1993. Protein phosphorylation andcellular regulation. Bioscience Reports v. 13p. 127–142.
Manning, G., Whyte, D. B., Martinez, R.,Hunter, T., and Sudarsanam, S., 2002.The protein kinase complement of thehuman genome. Science v. 298 p.1912–1934.
Newton, A. C., 2003. Regulation of the ABCkinases by phosphorylation: protein ki-nase C as a paradigm. Biochem. J. v. 370 p.361–371.
Nolen, B., Taylor, S., and Ghosh, G., 2004.Regulation of protein kinases; controllingactivity through activation segment con-formation. Mol. Cell v. 15 p. 661–675.
Turk, B. E. and Cantley, L. C., 2003. Peptidelibraries: at the crossroads of proteomicsand bioinformatics. Curr. Opin. Chem. Biol.v. 7 p. 84–90.
ResearchCanagarajah, B. J., Khokhlatchev, A., Cobb,
M. H., and Goldsmith, E. J., 1997.Activation mechanism of the MAP kinaseERK2 by dual phosphorylation. Cell v. 90p. 859–869.
Ginty, D. D., Kornhauser, J. M., Thompson,
14.24
14.23
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 640

References 641
M. A., Bading, H., Mayo, K. E.,Takahashi, J. S., and Greenberg, M. E.,1993. Regulation of CREB phosphoryla-tion in the suprachiasmatic nucleus bylight and a circadian clock. Science v. 260p. 238–241.
Knighton, D. R., Zheng, J. H., Ten Eyck, L. F.,Ashford, V. A., Xuong, N. H., Taylor, S. S.,and Sowadski, J. M., 1991. Crystal struc-ture of the catalytic subunit of cyclicadenosine monophosphate-dependentprotein kinase. Science v. 253 p. 407–414.
Taylor, S. S., Radzio-Andzelm, E., and Hunter,T., 1995. How do protein kinases discrim-inate between serine/threonine and tyro-sine? Structural insights from the insulinreceptor protein-tyrosine kinase. FASEBJ. v. 9 p. 1255–1266.
Zhang, F., Strand, A., Robbins, D., Cobb, M.H., and Goldsmith, E. J., 1994. Atomicstructure of the MAP kinase ERK2 at 2.3Å resolution. Nature v. 367 p. 704–711.
Two-component protein phosphorylationsystems are signaling relays
ReviewHoch, J. A, and Silhavy, T. J., eds., 1995. Two-
component signal transduction.Washington, D. C.:American Society forMicrobiology.
Stock, A. M., Robinson, V. L., and Goudreau,P. N., 2000. Two-component signal trans-duction. Annu. Rev. Biochem. v. 69 p.183–215.
Pharmacological inhibitors of protein ki-nases may be used to understand andtreat disease
ReviewBlume-Jensen, P, and Hunter, T., 2001.
Oncogenic kinase signalling. Nature v.411 p. 355–365.
Cherry, M. and Williams, D. H., 2004. Recentkinase and kinase inhibitor X-ray struc-tures: mechanisms of inhibition and se-lectivity insights. Curr. Med. Chem. v. 11 p.663–673.
Cohen, P., 2002. Protein kinases—the majordrug targets of the twenty-first century?Nat. Rev. Drug Discov. v. 1 p. 309–315.
Davies, S. P., Reddy, H., Caivano, M., andCohen, P., 2000. Specificity and mecha-nism of action of some commonly usedprotein kinase inhibitors. Biochem. J. v.351 p. 95–105.
Tibes, R., Trent, J., and Kurzrock, R., 2005.Tyrosine kinase inhibitors and the dawnof molecular cancer therapeutics. Annu.Rev. Pharmacol. Toxicol. v. 45 p. 357–384.
14.26
14.25
Phosphoprotein phosphatases reverse theactions of kinases and are independentlyregulated
ReviewAramburu, J., Heitman, J., and Crabtree, G.
R., 2004. Calcineurin: a central controllerof signalling in eukaryotes. EMBO Rep. v.5 p. 343–348.
Dounay, A. B. and Forsyth, C. J., 2002.Okadaic acid: the archetypal serine/thre-onine protein phosphatase inhibitor. Curr.Med. Chem. v. 9 p. 1939–1980.
Neel, B. G., Gu, H., and Pao, L., 2003. The‘Shp’ing news: SH2 domain-containingtyrosine phosphatases in cell signaling.Trends Biochem. Sci. v. 28 p. 284–293.
Neely, K. E. and Piwnica-Worms, H., 2003.Cdc25A regulation: to destroy or not todestroy—is that the only question? CellCycle v. 2 p. 455–457.
Olson, E. N. and Williams, R. S., 2000.Remodeling muscles with calcineurin.Bioessays v. 22 p. 510–519.
Virshup, D. M., 2000. Protein phosphatase2A: a panoply of enzymes. Curr. Opin. CellBiol. v. 12 p. 180–185.
ResearchSun, H., et al. 1993. MKP-1 (3CH134), an im-
mediate early gene product, is a dualspecificity phosphatase that dephospho-rylates MAP kinase in vivo Cell v. 75 p.487–493.
Terrak, M., Kerff, F., Langsetmo, K., Tao, T.,and Dominguez, R., 2004. Structural ba-sis of protein phosphatase 1 regulation.Nature v. 429 p. 780–784.
Covalent modification by ubiquitin andubiquitin-like proteins is another way ofregulating protein function
ReviewGill, G., 2004. SUMO and ubiquitin in the nu-
cleus: different functions, similar mecha-nisms? Genes Dev. v. 18 p. 2046–2059.
Pickart, C. M. and Eddins, M. J., 2004.Ubiquitin: structures, functions, mecha-nisms. Biochim. Biophys. Acta v. 1695 p.55–72.
ResearchDharmasiri, N., Dharmasiri, S., and Estelle,
M., 2005. The F-box protein TIR1 is anauxin receptor. Nature v. 435 p. 441–445.
Kanayama, A. et al. 2004. TAB2 and TAB3activate the NF-�B pathway throughbinding to polyubiquitin chains. Mol. Cellv. 15 p. 535–548
Kepinski, S. and Leyser, O., 2005. TheArabidopsis F-box protein TIR1 is anauxin receptor. Nature v. 435 p. 446–451.
14.28
14.27
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 641

642 CHAPTER 14 Principles of cell signaling
The Wnt pathway regulates cell fate dur-ing development and other processes inthe adult
ReviewLogan, C. Y. and Nusse, R., 2004. The WNT
signaling pathway in development anddisease. Annu. Rev. Cell. Dev. Biol. v. 20, p.781–810.
Tolwinski, N. S., Wieschaus, E., 2004.Rethinking WNT signaling. Trends Genet.v. 20 p. 177–81.
Diverse signaling mechanisms are regu-lated by protein tyrosine kinases
ReviewBirge, R. B., Knudsen, B. S., Besser, D., and
Hanafusa, H., 1996. SH2 and SH3-con-taining adaptor proteins: redundant orindependent mediators of intracellularsignal transduction. Genes Cells v. 1 p.595–613.
Blume-Jensen, P. and Hunter, T., 2001.Oncogenic kinase signalling. Nature v.411 p. 355–365.
Cohen, P., 2002. Protein kinases—the majordrug targets of the twenty-first century?Nat. Rev. Drug Discov. v. 1 p. 309–315.
Pawson, T. and Nash, P., 2003. Assembly ofcell regulatory systems through proteininteraction domains. Science v. 300 p.445–452.
Tallquist, M. and Kazlauskas, A., 2004. PDGFsignaling in cells and mice. CytokineGrowth Factor Rev. v. 15 p. 205–213.
Taniguchi, C. M., Emanuelli, B., and Kahn, C.R., 2006. Critical nodes in signallingpathways: insights into insulin action.Nat. Rev. Mol. Cell Biol. v. 7, p. 85–96.
Src family protein kinases cooperate withreceptor protein tyrosine kinases
ReviewBoggon, T. J. and Eck, M. J., 2004. Structure
and regulation of Src family kinases.Oncogene v. 23 p. 7918–7927.
Frame, M. C., 2004. Newest findings on theoldest oncogene; how activated src doesit. J. Cell Sci. v. 117 p. 989–998.
Research
Cowan-Jacob, S. W., Frendrich, G., Manley, P.W., Jahnke, W., Fabbro, D.,Liebetanz, J., Meyer, T. 2005. Thecrystal structure of a c-Src complex inan active conformation suggests pos-sible steps in c-Src activation Structurev. 13 p. 861–871.
Xu, W., Harrison, S. C., and Eck, M. J., 1997.Three-dimensional structure of the tyro-
14.31
14.30
14.29sine kinase c-Src. Nature v. 385 p.595–602.
MAPKs are central to many signalingpathways
ReviewChen, Z., Gibson, T. B., Robinson, F.,
Silvestro, L., Pearson, G., Xu, B., Wright,A., Vanderbilt, C., and Cobb, M. H., 2001.MAP kinases. Chem. Rev. v. 101 p.2449–2476.
Lewis, T. S., Shapiro, P. S., and Ahn, N. G.,1998. Signal transduction through MAPkinase cascades. Adv. Cancer Res. v. 74 p.49–139.
Morrison, D. K. and Davis, R. J., 2003.Regulation of MAP kinase signaling mod-ules by scaffold proteins in mammals.Annu. Rev. Cell Dev. Biol. v. 19 p. 91–118.
Sharrocks, A. D., Yang, S. H., and Galanis, A.,2000. Docking domains and substrate-specificity determination for MAP ki-nases. Trends Biochem. Sci. v. 25 p.448–453.
ResearchAnderson, N. G., Maller, J. L., Tonks, N. K.,
and Sturgill, T. W., 1990. Requirementfor integration of signals from two dis-tinct phosphorylation pathways for acti-vation of MAP kinase. Nature v. 343 p.651–653.
Cyclin-dependent protein kinases controlthe cell cycle
ReviewDorée, M. and Hunt, T., 2002. From Cdc2 to
Cdk1: when did the cell cycle kinase joinits cyclin partner? J. Cell Sci. v. 115 p.2461–2464.
Hartwell, L. H., 1991. Twenty-five years ofcell cycle genetics. Genetics v. 129 p.975–980.
Neely, K. E. and Piwnica-Worms, H., 2003.Cdc25A regulation: to destroy or not todestroy—is that the only question? CellCycle v. 2 p. 455–457.
Nurse, P., 2000. A long twentieth century ofthe cell cycle and beyond. Cell v. 100 p.71–78.
ResearchPavletich, N. P., 1999. Mechanisms of cyclin-
dependent kinase regulation: structuresof Cdks, their cyclin activators, and Cipand INK4 inhibitors. J. Mol. Biol. v. 287 p.821–828.
Russo, A. A., Jeffrey, P. D., and Pavletich, N.P., 1996. Structural basis of cyclin-de-pendent kinase activation by phosphory-lation. Nat. Struct. Biol. v. 3 p. 696–700.
14.33
14.32
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 642

References 643
Diverse receptors recruit protein tyrosinekinases to the plasma membrane
ReviewErnst, M. and Jenkins, B. J., 2004. Acquiring
signalling specificity from the cytokinereceptor gp130. Trends Genet. v. 20 p.23–32.
Herrington, J. and Carter-Su, C., 2001.Signaling pathways activated by thegrowth hormone receptor. TrendsEndocrinol. Metab. v. 12 p. 252–257.
Levy, D. E. and Darnell, J. E., 2002. Stats:transcriptional control and biological im-pact. Nat. Rev. Mol. Cell Biol. v. 3 p.651–662.
14.34Miranti, C. K. and Brugge, J. S., 2002.
Sensing the environment: a historicalperspective on integrin signal transduc-tion. Nat. Cell Biol. v. 4 p. E83–E90.
Palacios, E. H. and Weiss, A., 2004. Functionof the Src-family kinases, Lck and Fyn, inT-cell development and activation.Oncogene v. 23 p. 7990–8000.
ResearchClackson, T. and Wells, J. A., 1995. A hot spot
of binding energy in a hormone-receptorinterface. Science v. 267 p. 383–386.
39057_ch14_cellbio.qxd 8/28/06 5:11 PM Page 643





























