Embed Size (px)
Citation preview

35. POLLEN ANALYSIS OF SOME SAPROPEL LAYERS FROM THE DEEP-SEA FLOOROF THE EASTERN MEDITERRANEAN
Martine Rossignol-Strick, Laboratoire de Palynologie, Museum National d'Histoire Naturelle, Paris
INTRODUCTION
Presented here are the results of a palynologicalinvestigation of pollen and Dino flagellates in seven samplestaken from four different locations on the deep sea floor ofthe eastern Mediterranean during Leg 13 of the Deep SeaDrilling Project, August-October 1970. For this preliminarystudy of the cores of this leg, samples were chosen onlyfrom selected levels of dark, organic-rich mud. (Table 1).
Two types of lithologies were examined: one, consistingof redeposited clay and silt (terrigenous) from sedimentaryprovinces characterized by downslope and crossslopesediment distribution (that is, turbidity currents andgeostrophic circulation); and the other, containing pre-dominantly biogenic components derived from purelypelagic sedimentation. For the latter, we chose finelylaminated sapropels formed during periods of intermittentbasin-wide stagnation since these layers make excellentrepositories for organic trace material. In addition, theredeposited materials were selected at levels where the finaldeposition occurred either in an euxinic environment, orconsisted of components which had previously beendeposited in such an environment.
Pollen from the most recent ( 7000-9000 years B.P.)period of stagnation has been analysed in samples from theseabed near the Island of Crete (M. Rossignol and L.Pastouret, 1971). Investigations of surface sediments inshort cores have been reported by Koreneva (1969) andVronsky and Panov (1963).
Additional samples from deeper levels in Quaternarysections of piston cores from the Robert D. Conrad cruise 9in 1965 (Lamont-Doherty Geological Observatory ofColumbia University) are now being analysed.
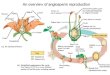
The materials discussed here come from four drillinglocations (Figure 1), namely Site 125 on the MediterraneanRidge in the Ionian Basin; Site 128 in the Hellenic Trench,Ionian Basin; Site 130 from the Mediterranean Ridge in theLevantine Basin; and Site 131A on the lower Nile Cone,Levantine Basin.
The Mediterranean Ridge sites are now isolated abovethe level of clastic sedimentation arriving onto the abyssalplains from the continents. Pelagic sedimentation at Site125 has been continuous throughout the last four millionyears (see Chapter 47), whereas, at Site 130, it has onlyreplaced predominantly terrigenous sedimentation in thelast few hundred thousand years (see Chapter 25).
The Nile Cone and Hellenic Trench sites are at localitiesdirectly reachable by downslope transport from Africa andEurope respectively.
SAMPLE PREPARATION
Moist samples of a few cubic centimeters volume werefirst weighed and then immersed in hydrochloric acid. Aftertwenty minutes the acid was diluted with distilled water,and.then after one and one half to two hours time forsettling, the liquid was decanted. The residue was sub-sequently soaked in 52 degree technical hydrofluoric acidand left to react for 24 hours. The preparation was thendiluted with distilled water and decanted. After this, 50percent HCI was added, left about half an hour, diluted andthen decanted again. The samples were washed once moreand then boiled for 3 to 5 minutes with 10 percentpotassium hydroxide. This was followed by washing in acentrifuge several times. This last reaction was repeated upto three times, depending upon the amount of aggregates ofamorphous organic matter still not dissolved. A Luberreaction was performed when the organic matter was tooabundant (mixture of 5 cc HN03 with a few drops of HCI,cold, for 2 or 3 minutes, and agitated with glass rods). Thesamples were finally washed by centrifuging, mounted inglycerin (to which was added a crystal of phenol) andsealed between slide and cover glass (22 X 40 mm.) withGlyceel.
DESCRIPTION OF THE POLLEN CATEGORIES
The pollen specimens recovered in the drill cores aresubdivided into several groups of trees, shrubs, grasses,hydrophytes, Pteridophytes, tropical trees and grasses, andothers undetermined. The raw pollen countings for eachsample investigated are listed in Table 2 and the percentagesof each group in the total pollen sum are indicated inTable 3.
Mediterranean Trees
An analysis of the pollen countings shows that two maintypes of trees, Quercus and Pinus are represented in similarrelative proportions. Along with Cedrus, Olea, and to alesser extent, Pistacia, these trees indicate a mediterraneanenvironment with a two-season year consisting of coolhumid winters and hot dry summers accompanied by arainfall of from 400 to 1200 or more millimeters per year.Rainfall is mostly limited to the winters with occasionalsummer rains at high altitudes.
Cedrus requires cold and sunny days in the winter,which often occur at high altitude. Cedrus is found inwidely separated locations: in Morocco in the Atlas, inLebanon, south Turkey and Cyprus, and in the Himalayas.The respective adaptations of Mediterranean Pinus, P.
971

M. ROSSIGNOL-STRICK
TABLE 1Sample Inventory
Location
SITE 125 - MediterraneanRidge
Ionian Basin
SITE 128 - HellenicTrench
Ionian Basin
SITE 130 - MediterraneanRidge
LevantineBasin
SITE 131A -Nile Cone
LevantineBasin
Latitude
34° 37.5'N
35° 42.6'N
33° 36.3'N
33° 06.3'N
Longitude
20° 25.8'E
22G 28.1'E
27°52.0'E
28°52.7'E
WaterDepth
(m)
2782
4640
2979
3037
SampleDepth
(m)
208.0
85.5
87.7
52.7
256.0
49.4
208.0
Core
3
3
3
2
5
1
4
Section
5
5
6
3
2
2
2
SampleInterval
(cm)
57-58
53-54
120-121
73-74
54-55
90-91
152-153
Blacksapropela
Redepo sitedsapropel b
Redepositedsapropelb
Jet blacksaρropela
Finely lam-inated blackclayb
Dark graysilty-claybBlack lami-nated silty-clay
Distance from Land
200 km N of CyrenaicanCoast, 260 km SW of Pelo-ponnese (Cape Matapan),300 km WSW of Crete, 520km SE of Reggio (Calabria).
110 km S of Peloponnese(Cape Malea), 60 km W ofCrete, 300 km N ofCyrenaican Coast
300 km NW of Alexandria,250 km N of African Coast,250 km S of Rhodes, 300 kmS of Turkish Coast
150 km NNW of Alexandriaand the Nile Branch ofRosetta
Pelagic sedimentation, predominantly biogenic components.bPredominantly terrigenous components.
Figure 1. Physiographic setting of the Mediterranean drill sites where pollen was investigated.
brutia, P. halepensis, P. nigra, and Quercus are dependentupon soil and bedrock conditions, altitude, and variationsof rainfall and temperature within the Mediterranean limits.Pinus here is mostly represented by P. diploxylon.However, in Sample 125-3-5, a low percentage (about 1%)of the Pinus group consists of P. haploxylon, considered asa Tertiary relic. The Mediterranean trees are most highlyrepresented in the samples close to Greece in the HellenicTrench, but are second in abundance at the MediterraneanRidge sites. They are poorly represented in samples fromthe Nile Cone.
Like the other ligneous groups described next, theMediterranean tree pollens were probably first blown in tothe sea from the Balkans and perhaps Italy, and thencarried by currents to their final resting place. Thedominant winds in the eastern Mediterranean basins arewesterly, but in the summer, strong Etesian winds comefrom the north.
Deciduous Thermophile Trees
These trees require the same conditions as those existingtoday in the mountain ranges of the Balkans, the Caucasus,
972

35. POLLEN ANALYSIS OF SAPROPELS
and Iran (on the northern slopes of the Elburz), and in theHyrcanian forest) facing the Caspian Sea (the Balkano-Caucasian group). Most of them extend today to the forestsof temperate European lowlands further North (Acer,Fagus, Tilia, Ulmus, Castanea, Fraxinus, most Betulaceaé)and do not require as much warmth. Others were naturallyrestricted to southern European and S.W. Asian mountains,which appear to be a refuge for a Tertiary relict flora:Liquidambar, Zelkova, Juglans, Ostrya. Some are Tertiaryrelics which disappeared sometime during the Quaternaryfrom this area and are to be found now either further eastin Asia, {Pterocarya), or in America (Carya).
Pterocarya is found today both in the Far East and inthe Near East: in the Caucasus, East Turkey, and NorthIran, facing the Caspian Sea, It grows on river banks in low-lands, rarely above 300 meters (Tralau, 1963). Liquidambarstill exists on the south shore of Turkey, as well as in Chinaand North America (Kuprianova, 1960). Zelkova is nowendemic in Crete, and occurs in the Caucasus, in Iran in theHyrcanian forest, and in the Far East (Tralau, 1963). In theCaucasus, it does not grow higher than 300 meters abovesea level, but in Iran, it reaches 1500 meters; the averagetemperature in the Caucasus is lower than in the Hyrcanianforest.
The requirements of these species are primarily, rainfallevenly distributed throughout the year, cold winters, andwarm summers. The chronological implications of thepresence of these Tertiary relics—Cedrus, Pterocarya, Carya,Liquidambar, Zelkova-will be discussed in further detail.They appear up to the lower Pleistocene in Italy.
In the pollen record, Pterocarya appears with 5, 6, or 7pores, size 25 to 37 µm.
Carya has the typical 3-sub-equatorial pores in the samehemisphere.
The distinction between Zelkova and Ulmus is based ona thicker pore annulus and more conspicuous rugulatepattern in Zelkova than in Ulmus.
This group is never dominant, nor even second inimportance, but is highly valuable for the purpose ofdating. Its greatest abundance occurs in samples close toGreece, and it is almost completely absent in samples closeto the Nile Delta.
Other TreesThe trees grouped here are neither deciduous (Tsuga,
Abies) nor thermophiles (Betula). Tsuga is another Tertiaryrelic of the early Pleistocene of southern Europe. Abiesexists today in the mountains of Greece and Turkey. Betulais found further north in Europe.
This group is present in very low percentages. However,it is highest in representation near Greece.
Shrubs
Ericaceae are common in the area north of theMediterranean. Among Thymeleaceae, the most common,Thymelea hirsuta, is a desert shrub in sandy soil. It occurshere (128-3-6) in a station where the desert and steppegrasses are dominant.
Other prevalent shrubs include Ilex in Sample 131 A-1-2on the Nile Cone, Buxus cf. sempervirens, and Poteriumspinosum. A single specimen of Myrica appears in Sample
131A-4-2 and may have come from Uganda by way of theNile. The highest representation of this group of shrubsoccurs in the samples of pelagic sediment from theMediterranean Ridge (130-2-3 and 125-3-5).
Grasses: abundant
The dominant grasses comprise four types: Graminae,Cyperaceae, Chenopodiaceae, and Artemisia. Theyrepresent steppe, saline desert, littoral sand belt and marshyenvironments.
Comparisons of the percentages of the grass group in thepollen sum, with the percentages of the other groups, andwith those of each grass type in the group and in the pollensum, reveal the following (Table 4):
1) The percentage of grasses in the pollen sum is usuallyhighest in samples taken close to the Nile Delta, where theMediterranean tree element is lowest. They are lowest,although still above 30 per cent, in samples from theMediterranean Ridge where the Mediterranean tree elementis highest.
2) Within the group, Graminae and Artemisia arepresent in the highest percentages. A comparison of thepercentages of these two types in the pollen sum show thatGraminae are highest in the samples of Nile River origin(131A-1-2, 131A-4-2, 130-5-2).
Artemisia is the most abundant grass in the pollen sumin the pelagic sediments of the Mediterranean Ridge and inthe redeposited sapropels of the Hellenic Trench. In all ofthese, the Mediterranean arboreal element also reaches itsmaximum. In one of the northern samples, Artemisia evendetermines a strong dominance of the grass group in thepollen sum (128-3-6).
The high percentages of Graminae in the Nile trans-ported sediment brings the grass group as a whole to muchhigher percentages than does Artemisia in most northsamples, where it is counterbalanced by a large Mediter-ranean tree group.
3) There are two secondary members of the grass group,Chenopodiaceae and Cyperaceae. The distribution ofCyperaceae parallels that of Graminae: high in the samesamples—and low in all the northern samples.
Chenopodiaceae variations are not as clear if only theirpercentages in the grass group are considered. However,their percentages in the pollen sum appear fairly constant inall the samples investigated, with the narrowest range ofvariation of all the grass types, this being from 4.4 to 13.5per cent. Since their deposition does not seem linked to theNile sedimentation, nor to the northern vegetation element,an independent origin for this family is suggested, namely,wind-transport from the North African or Middle Easterndeserts and/or from the Mediterranean shores.
These comparisons of percentages establish the geo-graphical origin of each grass type. Graminae and Cyper-aceae are transported by the Nile either from the Sudanesemarshes of Bahr-el-Ghazal, or from the marshes of theDelta, or both. However, it is possible that part of theGraminae in the samples from the North, also have anorthern origin. Artemisia, similar to the Mediterraneantree element, has a northern origin in the Balkanic andTurkish areas. Within this northern vegetation group, theimportance of Artemisia appears to be inversely related to
973

M. ROSSIGNOL-STRICK
TABLE 2Pollen Counts in the Seven Samples from the Eastern Mediterranean
Pollens
TreesMediterranean
QuercusOleaPhillyreaOleaceaePistacia spPistacia lentiscusPinusCedrus
Total
Deciduous thermophilesAcerFagusTiliaUlmusZelkovacf EngelhardtiaLiquidambarCaryaPterocaryaJuglansAlnusCarpinusCarpinus caucasicaOstryaCorylusCastaneaFraxinus
Total
OthersBetulaAbiesTsuga
Total
ShrubsEricaceaeThymeleaceaeIlexBuxux sempervirensPoterium spinosumMyrica
Total
GrassesDominant (Steppic, deserticor littoral)
GraminaeCyperaceaeChenopodiaceaeArtemisia
Total
FrequentCompositae:
LigulifloresTubiflores
EphedraPlant ago
Total
Samples
131A1-2
2
1
3
0
0
1
1
247235
731
556
12
5
8
131A4-2
2
1
3
1
1
0
1
1
2137117
3
304
2523
12
1302-3
32652
9
14569
11
981
2
124
9
331
22
18
19
3
96
411
6
78
14
92
30674
235623
1238
13801525
133
1305-2
221
7
30
1
2
6
9
1
1
1
1
28512758
8
478
937
19
1283-5
15011
9
899
268
2
611
10
12
3
3
1
4
111
4589
146
4821
15
1283-6
170313
24112
430
246
1
20
24
57
84
12
1
6
7
580
29812261582
122443
7
86
1253-5
14113
222
18014
372
33
2011
51026
734
1424
11
132
1842
24
36
2
38
14144
116214
515
9241333
19
974

35. POLLEN ANALYSIS OF SAPROPELS
TABLE:
Pollens
OthersBorraginaceaeCampanula ceaeCaryophyllaceaeCentaur eaCistaceaeConvolvulaceaeCruciferaeDipsaceaeGeraniaceaecf. Galiumcf. GuttifereHelianthemumLabiataeLythraceaePlumbaginaceaePolygonaceaeRanunculaceaeRosaceaeRumexSanguisorbaSolanaceaeUmbelliferaeUrticeae
Total
HydrophytesNympheaceaeTyphaceaeJuncaginaceae
Total
PteridophytesSpores monoletesSpores triletesRiccia
Total
Tropical Trees and GrassesPodocarpusA caciaAcanthaceae
ctDiclipteracLHypoestes
cf. CapparidaceaeCombretaceaeEuphorbiaceaeEuphorbiaAlchornea ctcordifolia
cf. TamarixMoraceaeFicusOthersTotal
Undetermined and undeterminable
Total Counted (Pollen Sum)
. - Continued
Samples
131A1-2
1
1
181
10
1024
8
42
21
1
3
2
9
5
635
131A4-2
1
1
1
2
5
5
5
327
2
32
11
11
3
7
14
384
1302-3
17
5
1
2
1
1
1111
13
35
2
2
1
1
4
3
1
1
9
6
2599
1305-2
1
1
1
1
367
2
72
1
1
2
20
634
1283-5
2
3
1
3
9
1
1
11
2
0
21
475
1283-6
17
1
1
1
1
12
24
2
2
0
11
2
6
2208
1253-5
51
2
1
14
76
27
22
4
11
2
1
1
12
1224
that of the Mediterranean trees. Chenopodiaceae, as anisolated group, seems to originate from the surroundingdeserts.
Grasses: frequentThe group of grasses frequently encountered includes
the Compositae (mostly Tubiüoxes), Ephedra andPlantago.
975

M. ROSSIGNOL^TRICK
TABLE 3Percentages of Each Group in the Pollen Sum
Pollens
TreesMediterraneanDeciduousthermophileOthers
Shrubs
GrassesDominantFrequentOthers
Hydrophytes
Pteridophytes
Tropical trees and grasses
Undetermined andundeterminable
A P
N A P
Pollen Sum
Samples
131A1-2
0.50.0
0.0
0.1
87.51.20.1
1.6
6.6
1.4
1.0
0.6
99.4
635.0
131A4-1
0.80.3
0.0
0.3
79.53.11.3
1.3
8.3
1.8
3.6
1.4
98.6
384.0
1302-3
37.83.6
0.2
3.5
47.75.11.3
0
0
0.3
0.2
45.1
54.9
2599.0
1305-2
4.71.4
0.1
0.1
75.43.00.1
0.1
11.3
0.3
3.1
6.3
93.7
634.0
1283-5
56.41.3
0.6
0.8
30.73.11.9
0.2
0.4
0.0
4.4
59.1
40.9
475.0
1283-6
19.52.6
0.5
0.3
71.63.91.0
0.2
0.1
0
0.3
22.9
77.1
2208.0
1253-5
30.412.1
2.0
3.1
42.06.42.2
0.3
0.2
0.1
1.0
47.7
52.3
1224.0
TABLE 4Percentages of Grass Types in the Grass Group and in the Pollen Sum
Pollens
Percentages of each grasstype in the grass group
GraminaeCyperaceaeChenopodiaceaeArtemisia
Sum of grasses
Percentage in totalpollen sum
Percentages of each grasstype in total pollen sum
GraminaeCyperaceaeChenopodiaceaeArtemisia
Note from Table 2:Per Cent of Mediterraneantrees in pollen sum
Samples
131A1-2
44.0%42.013.0
0.2
556
87.5%
38.9%37.011.50.1
0.5
131A4-2
70.0%23.0
5.61.0
304
79.2%
55.5%18.54.40.8
0.8
1302-3
25.0%6.0
19.050.0
1238
47.7%
11.8%2.89.0
24.0
37.8
1305-2
60.0%26.012.0
2.0
478
75.4%
45.7%20.3
9.11.3
4.7
1283-5
7.5%0.7
31.061.0
146
30.7%
2.3%0.29.4
18.6
56.4
1283-6
4.0%0.0
19.077.0
1582
71.6%
2.6%0.0
13.555.5
19.5
1253-5
27.0%8.5
22.541.5
515
42.0%
11.5%3.69.5
17.5
30.4
Their representation is greatest in the pelagic sapropelsfrom the Mediterranean Ridge and the redepositedsapropels from the Hellenic Trench. They may belong to abelt of vegetation along the northern Mediterranean shore.Grasses: others
As a whole this diverse group is also best represented inthe sediments derived from Europe. However, two families,
Borraginaceae and partly Umbelliferae, are probably ofsouthern origin.
Hydrophytes
These ubiquitous plants are more highly represented inNile Cone samples.
976

Pteridophytes
Pteridophytes are abundant only in the samples from theNile Cone and the terrigenous black clays from Site 130.The trilete spores belong to 12 types, the monolete, to 2.There are Pteris, Cyatheaceae, Selaginellaceae, Osmon-daceae, Polypodiaceae. No attempt has been made to makemore precise identifications. These same types, with about30 more, have already been found off Palestine, in siltsfrom the Nile Delta, and from Abou Simbel (M. Rossignol,1969). Therefore, in our samples they can also beconsidered as Nile-transported. It is uncertain whether thesespores come from the present fern vegetation around LakeVictoria (where the substratum is granitic) and were carriedby the White Nile, or whether they are redeposited fromthe Mesozoic marine strata which, underlying the BasaltTrap in Ethiopia, are deeply cut by the Nile Gorge and itstributaries. The spores would then have been carried by theBlue Nile. The second hypothesis seems more probable,although comparative data are lacking.
Tropical Trees and GrassesAlthough numerically not significant, the group of
tropical trees and grasses is qualitatively very interesting. Itappears to be more highly represented in the terrigenoussamples from the Nile Cone: Podocarpus already has beenfound rather frequently off Palestine and in its Pleistocenecoastal plain substratum (M. Rossignol, 1961, 1969). Thissmall bladdered pollen has good buoyancy and may comefrom the forest belt of the east African equatorialmountains. Podocarpus pollen as found in the northernsamples is probably a local Tertiary relic.
The Acanthaceae are also found off Palestine. They areabundant in the African savanna. Two genera are recordedhere: cf. Dicliptera and cf. Hypoestes. The Euphorbiaceaehave appeared here only as non-Mediterranean, tropicalspecies: Alchornea cf. cordifolia (Van Campo, PalynologieAfricaine, IV, PI. 96), and two tropical tricolporateEuphorbiaceae with strongly developed sunken rolled poremargins in equatorial contour; one of them is probably anEuphorbia.
The Capparidaceae are identified by their granulousendoaperture membrane.
The Ficus and Moraceae are triporate with a thin exineand no differentiated aperture margins, about 20 µm.Among the other tropical pollens is a stephanocolpate (IIcolpi) sub-prolate, 39 X 36 µm, with a tectate scabrateexine and thick ectexine provided with large isolatedcolumellae: cf. Bignoniaceae or Rubiaceae.
Undetermined and Undeterminable Pollens
Some small triporate pollens with a thin exine andnon-annulated pores could not be satisfactorily attributedto Betulaceae, Halorhagaceae, Myricaceae, Casuarinales,Juglandaceae, Moraceae, Urticeae, Symplocaceae.
A very conspicuous tricolporate pollen, frequent in twosamples, could not be identified (Plate 3, Figures 1-6, 10).It is sub-oblate having a 42 to 37 µm equatorial diameterand a 39 to 35 µm polar axis. The equatorial contour isangular, hexagonal with V-shaped planaperturate colpi andslightly convex, bow-shaped intercolpial planes. It has alarge polar triangle (side, 10 µm). Exine pattern: theectexine is composed of columellae united by their heads,
35. POLLEN ANALYSIS OF SAPROPELS
and this tectum has a scabrate surface. The columellae havevarying lengths and thicknesses. They are long and thick atthe angles in equatorial contour and in the polar areas andare short and much thinner in an aureole around theapertures and in the middle of the intercolpial planes. Inthis latter location they delineate a circular area of thinexine easily folded on itself in equatorial contour, thusaccentuating the angular outline. A very conspicuouscharacteristic is the apparent absence of any endexine inthe white-light microscope; the foot of the columellaeappears to rest on nothing. The colpi are slit-shapedwithout incrassate margins. There is a barely visibleendoaperture: oval-shaped, lalongate, ill-defined at itsmeridian extremities; its existence would tend to prove,however, that of an endexine, at least in the endoaperturearea. This very typical pollen appears overall 23 times: 4 inSample 130-2-3, 19 in Sample 125-3-5. Both these sampleshave a northern pollen record, among which the deciduousthermophile trees and the Tertiary relics reach their highestpercentages. This suggests that the pollen could also be aTertiary ligneous relic. It is not impossible that it belongs tothe Verbenaceae family, which is essentially tropical andsub-tropical.
ANALYSIS OF THE NORTHERN POLLEN GROUPAND OF THE NILE POLLEN GROUP
The seven samples are arranged in two groups accordingto the dominant origin of their pollens: northern or Nile.Only the pollens of northern origin are considered in thenorthern dominated samples, and of Nilotic origin for theNile dominated samples. This allows us to establish moreclearly the composition of each group at the time of itstransport, before it became mixed with some elements ofthe other group during its final moment of deposition.
The Northern PollenFor the group of northern pollen, we consider the
arborescent pollen (AP) and the non-arborescent pollen(NAP) as two subdivisions. The NAP includes Artemisia, allthe frequent grasses, and all the "other" grasses. We notethat the Graminae and Cyperaceae have not been includedin the northern group, although some of them may wellhave had an origin in Europe. The AP division includes theMediterranean trees, the deciduous thermophile trees, andthe "other" trees and shrubs. The countings and relativepercentages of AP and NAP in the northern group are givenin Table 5. The ratio of the AP to the sum AP + NAP isgreater than 60 per cent in all samples but the redepositedsapropel of 128-3-6, and suggests a dense Mediterraneanforest not far from the coast. The deciduous thermophileelement may grow either in an upper altitudinal belt, orfurther to the north. Artemisia represents the highestpercentage of the NAP: 79 per cent in 130-2-3, 79 per centin 128-3-5, 92 per cent in 128-3-6, 67 per cent in 125-3-5,and the following percentages of the northern pollen sum,in the same order: 31 per cent, 23 per cent, 67 per cent,and 24 per cent. In Sample 128-3-6, a heavy increase ofArtemisia (67%) is coupled with a sharp decrease of the AP(27%). In the two samples of Core 128, there is clearly atrade off between Artemisia and the AP, their sumreaching 94 per cent in both samples. If this is not linked
977

M. ROSSIGNOL-STRICK
TABLE 5Percentages of Arborescent Pollen (AP) and Non-arborescent
Pollen (NAP) in the Northern Pollen Group
Northern Pollens
Sum of AP:Medit. + Decid. therm. + others+ shrubs
Sum of NAP:Artemisia
+ grass frequent+ grass others
Sum of AP + NAP
AP X 100AP + NAP
Samples
1302-3
1175
791
1964
60%
1283-5
281
113
394
71%
1283-6
506
1336
1842
27%
1253-5
584
320
904
64%
only to deposition conditions, it could indicate a climaticvariation resulting in the increase of the territory ofArtemisia at the expense of the Mediterranean forest. Itmay be relevant here to recall that, in the pollen analysis ofa Würmian core in Thrace, northern Greece, Wijmstra(1969) found that the cold phases were similarly marked byan increase of Artemisia at the expense of the Mediter-ranean forest. This suggests that the Pleni-Glacial Würm waslocally a cool and dry phase rather than cool and humid. Asimilar conclusion has been reached by Van Zeist (1967)concerning a palynological investigation in the Zagrosmountains of western Iran, where local conditions wereconsidered as having a dominant influence. We suggest thatthe climatic limiting factor was humidity, rather thantemperature.
It is quite apparent from this grouping of the northernpollen that the pelagic sapropel from the MediterraneanRidge north of the Nile Cone at Site 130, in Core 2, Section3, owes more than 75 per cent of its pollen content totransport from the north. A very similar arborescent pollenratio is found at the other Mediterranean Ridge location inthe Ionian Basin (125-3-5) and again in a pelagic sapropellayer. In 125-3-5, the abundance of the now locally extinctCarya, Pterocarya, Tsuga points to a very early Pleistoceneage.
The high pollen counts of the northern pollen group atthese two topographically isolated locations argues for anefficient pelagic transport (wind and oceanic circulation)over moderately long distances in the open ocean.
The Southern (Nilotic) Pollen
The southern group, of dominant Nile origin, isrepresented in the finely laminated black and dark grayterrigenous clays from the Nile Cone (131 A-1-2; 131A-4-2)and the lower part of the sediment column from theMediterranean Ridge (130-5-2) directly north of the NileCone. Because of the absence in these samples ofarborescent pollen, the AP/NAP ratio is not significant. Thepollen sum (see Table 6) only includes the pollen from Nileorigin: Graminae, Cyperaceae, frequent grasses, hydro-phytes, Pteridophytes, tropical trees and grasses.
The decreasing order of importance of the pollen groupsremains constant in all three samples. Graminae and
Cyperaceae together account for 81 to 86 per cent of theentire southern group. The order of magnitude of eachgroup does vary, but not very widely, except forCyperaceae (from 21 to 42 per cent). This suggests, mostlikely, a fairly constant input to the sea by the Nile River.
MECHANISMS OF TRANSPORT AND DISPERSALOF THE POLLEN
Three different processes involving the transport, dis-persal and eventual sedimentation of the pollen aresuggested from the observed distribution pattern in the fourdrillsites.
1. The first involves transport in a large stream such asthe Nile River where the pollen becomes thoroughly mixedwith terrigenous and other organic materials. The Gra-minae, Cyperaceae, Pteridophytes, hydrophytes, tropicaltrees and grasses enter the sea in suspension with thesediment discharge of the Delta. As observed by Emery etal. (1966) and Venkatarathnam and Ryan (1971), the Nilesediment crosses the Nile Cone and reaches the presentHerodotus Abyssal Plain in nearbottom flows. Similarly,the pollen behaves as a benthic element.
The characteristic blue black Nile muds extend only asfar as the northern terminus of the abyssal plain, and areabsent in surface sediments on isolated topographicelevations on the lower Nile Cone.
The river-transport process is very efficient for longdistances provided there is a pollen abundance in thecatchment area. The pollen can be either contemporarywith its transport, or it can be reworked from thesedimentary substratum cut by the upper valley erosion.Much pollen is probably destroyed during the transport.Upon reaching the open sea, the initial discharge maytemporarily come to rest on the shallow shelf beforeeventual flushing downslope by slumping and turbiditycurrents. Thus, redeposited pollen of different stages iscommon in the southern (Nilotic) pollen group.
TABLE 6Percentages of Graminae, Cyperaceae, Pteridophytes, Tropical
Trees and Grasses, and Frequent Grasses in theSouthern (Nilotic) Pollen Group
Nile Pollens
Sum of Nile pollens:Gram. + Cyper. + grass frequent+ Pteridoph. + tropic.
Percentages:Gram. X 100
Sum
Cyper. X 100Sum
Pteridoph. X 100Sum
Tropical X 100Sum
Grass frequent X 100
Sum
Samples
131A1-2
541
45.0%
43.0%
8.0%
2.0%
2.0%
131A4-2
335
64.0%
21.0%
9.0%
2.0%
4.0%
1305-2
505
56.0%
25.0%
14.0%
0.4%
3.7%
978

35. POLLEN ANALYSIS OF SAPROPELS
2. A second mechanism primarily involves short-distance wind-transport to the sea from the area of originalvegetation, and then long-distance dispersal within theoceanic circulation system, mainly in surface currents. Herewe surmise that the vegetation supplying the pollenprobably lies within a few tens of kilometers from thecoast. This pollen behaves essentially as a. pelagic element; itfloats longer and further if it has a high surface tension, anda low density. The northern pollen group, Mediterraneantrees, deciduous thermophile trees, shrubs, Artemisia, andsome others, are carried in this way. The eveningland-breezes of the coastal belt, the westerly winds and thenorthern Etesian winds, probably play an important role.This transport pattern results in a decreasing gradient ofsedimentation from the area of origin. Because there are nolarge streams from Greece, southern Italy or southernTurkey, the probability of reworked pollen into thenorthern group is low.
3. The uniform distribution of certain pollen types overlarge areas of the open-sea seems best explained by a thirdmechanism primarily involving long-distance wind-transportin the upper atmosphere. Such aeolian transport is invokedin order to provide the wide uniform dispersal of smallsized non-bladdered pollens. It also happens to beresponsible for the relatively high amounts of kaolinitesediments derived from the North African deserts and nowfound across the western Mediterranean Ridge and westernNile Cone (Venkatarathnam and Ryan, 1971). Only pollenwhich is very abundant in its area of production can beexpected to be recognized as having this type of dispersal.This concerns the Chenopodiaceae, which probably comefrom the deserts of North Africa and the Middle East. It isthe only pollen transport which can provide a similardeposition pattern on land.
Except in the case of bladdered pollens, the process oftransport and deposition of pollen grains does not seem tobe determined primarily by their intrinsic characteristicsother than abundant production and a rather small size (thequasi-totality of non-bladdered recovered pollens are below30 to 35 µm). Rather, they depend on the environment ofthe plants which produce them: geographical and topo-graphical location, proximity of a stream, and atmosphericcirculation.
The resultant marine pollen spectrum has the advantageof eliminating the distortions caused by the immediatevegetal environment in the pollen spectrum of a small bodyof water.
ABSOLUTE FREQUENCIES
Absolute frequencies represent the number of micro-fossils, pollens or Dinoflagellate cysts (Hystrichospheres),by gram of sediment. They are calculated as follows: P isthe number of microfossils counted on a surface of Shorizontal lines of the microscopic field at low scanningmagnification (wide-field eye-piece 10X, objective 16X).Each line is 1 mm in diameter; there are 20 lines in eachpreparation (cover-glass of 22 X 40 mm). N is thenumber of microscopic preparations which may be madewith all the residue of the sample. It is evaluated by thenumber of drops in the tube of residue (two drops arenecessary for one preparation). This is measured by
comparing levels of liquid in two similar tubes, one aswitness, and the other with the residue. W is the weight ingrams of the sediment sample. The absolute frequency Fa iscalculated:
FaSW
For the three samples of Nilotic origin (131A-1-2,131A-4-2, 130-5-2), the pollan and Hystrichospheres abso-lute frequencies are rather low: between 800 and 3100pollens, and 4 to 120 Hystrichospheres per gram of sediment.
This can be interpreted as the result of the fastaccumulation rate of Nile mineral sediments. In fact, thetwo samples at Site 131A are very sandy. The benthicpollen input is apparently diluted and moreover, may nothave been very abundant originally. (This last concept isnot easy to define; that is the number of pollen grainsreaching a still surface unit of the water, stream or sea, pertime unit).
For the other samples except Sample 128-3-5, theabsolute frequencies of pollen and Hystrichospheres aremuch higher, being from 15200 to 32000 for pollen and1250 to 5350 for Hystrichospheres. These high frequenciesresult mainly from a low sedimentation rate for its matrix,both mineral and organic; good conditions of preservationfor pollen on the sea bottom after it settled there; andmaybe also a more abundant pollen input. Pelagic polleninput is concentrated and also abundant (see aboveremark). All these values are much higher than thoserecorded in the Nile silt deposited offshore of Palestineduring Pleistocene and Recent times (M. Rossignol, 1961,1969) where the still high mineral sedimentation rate wascombined with non-stagnant bottom conditions.
The absolute frequencies of Hystrichospheres, expressedas percentages of the absolute frequencies of pollen in thesame sample, are relatively much lower in the sediments ofNile origin than in the pelagic sediments. This results fromthe conditions of formation of the Hystrichospheres. Theplanktonic motile stage of the Dinoflagellates lives in thesuperficial waters, where the waters of the Nile flood, lessdense, less saline, than normal sea water, but richer innutrients, circulate. The formation of the cyst is induced inthese superficial waters, when the conditions of life are nolonger favorable to the motile stage: less nutrients, changeof salinity, overgrowth of the population; all events linked,in the south, to the periodicity of the Nile Flood. Geneticdeterminism may also have a role. Then, the cyst settles onthe sea floor. Therefore, the induction of the cystformation is linked to sea surface conditions, but theirfrequency per gram of bottom sediment, although notdirectly related to bottom topography, is influenced by therate of mineral accumulation. This explains why the lessdiluted pelagic sediment of 130-2-3, which is rich inNorthern pollen, is also the richest in Dinoflagellate cysts.
In the sediments of Nile origin (131 A-1-2, 4-2, 130-5-2),the Dinoflagellate absolute frequencies increase with thedistance from the Nile mouth, corresponding with lessdilution of the cysts by mineral sedimentation. The Nilepollen absolute frequencies present a parallel trend in thesame sediments. It is suggested that at the time ofdeposition of 131A-4-2, the mineral sedimentation rate was
979

M. ROSSIGNOL-STRICK
higher than for 131A-1-2, and maybe also the distance fromthe Nile mouth was smaller. In the south the absolutefrequency of Dinoflagellate cysts results from the combinedinfluences of superficial Nile waters and bottom-transported Nile sediments. In the northern samples of theMediterranean Ridge (Site 125) and Hellenic Trench (Site128), the presence of Dioflagellate cysts could be linked tothe arrival of part of the less saline and nutrient-loadedwaters of the Adriatic and Black Seas. It should be notedthat the absolute frequencies of both pollen and Hystricho-spheres in 128-3-5 are about 5 times lower than in 128-3-6.The ratios of Hystrichospheres to pollen are similar,however, being 8 and 9 per cent. This suggests that themineral dilution is 5 times higher in 128-3-5 than in128-3-6, a fact probably linked to the sedimentationcondition of 128-3-5.
TABLE 7Absolute Frequencies of Pollen and Hystrichospheres
FaMicrofossils
Fa pollen
FaHystricho-spheres
FaHystr.× 100/
Fa pollen
131A1-2
2600
60
2%
131A4-2
800
4
0.5%
1302-3
32000
5350
17%
Samples
1305-2
3100
120
4%
1283-5
3300
300
9%
1283-6
15200
1250
8%
1253-5
21200
2500
12%
POLLEN SEDIMENTARY CONDITIONS
This paragraph summarizes for each sample, informationfrom the previous paragraphs (Table 8).
Samples 131A-1-2 and 131A-4-2The two samples from Site 131 A, on the lower Nile
Cone, have a distinct pollen content of Nile origin. Itsultimate source has not been ascertained. Grasses andCyperaceae may come mostly from the Delta marshes, butalso from the Bahr el Ghazal marshes further upstream.Pteridophytes may be recent, from the Lake Victoria area,or more likely, reworked from the Mesozoic substratum ofthe Ethiopian plateau. Also, they may be from any othereroded sedimentary area in the past catchment area. Only,the tropical element points clearly to a tropical Africanorigin.
The pelagic transport of the northern pollens in thestation area is prevented or extremely diluted by theenormous influx of the Nile waters. This should cast lighton the water circulation in this area. Even some of thepollens which are northern in the pelagic samples from Site131A are more likely of Nile origin (Oleaceae,//ex).
Samples 130-2-3 and 130-5-2Site 130 is located within a deformed belt of the
Mediterranean Ridge, north of the Herodotus Abyssal Plain(see Chapters 11 and 25 of this volume). The two samplestaken from the sediment column show a striking difference.The deeper sample (130-5-2) taken from the laminatedblack terrigenous clay, displays a dominant Nile influence,only slightly weaker than in the samples of Core 131 A. The"northern" pollens, although very low, are higher than inCore 131 A, and their variety points clearly to a northernorigin. The upper sample (130-2-3) from the pelagicsapropel, shows on the contrary, a weak Nile influence anda much higher northern pelagic influence. The main part ofthis northern pollen fraction is composed of Mediterraneantrees, with a very significant abundance of deciduousthermophile trees. The different pollen pattern of the twolevels indicates that an important geological event has takenplace at this drill site. It is suggested that the lower level,rich in terrigenous mud, shared the same sediment source asthe Herodotus Abyssal Plain, namely, the Nile. Subse-quently, an uplifting of the area prevented the arrival of thebulk of the Nile sediments and pollens, which apparentlyare only transported in deep water masses as benthicelements. On the subsequent uplifted zone, the pelagicpollen sedimentation of northern origin became dominant,as evidenced in the upper level. (For a further discussion seeChapter 48 of this volume). The sedimentation of thewind-transported Chenopodiaceae was not affected bychanges of the bottom topography, nor by the origin of thewater masses.
Table 9 compares the composition of the northern group(main components) in both samples from Site 130 andfrom Site 125, also on the Mediterranean Ridge. All threesamples have the same dominant element (Mediterraneantrees). The relative abundance of the deciduous ther-mophile tree group in relation to the northern pollen groupis higher in the lower sample from Site 130, and very closeto that observed at Site 125.
The absolute frequencies also reflect the environmentchange. The lower level has a low absolute frequencysimilar to that at Site 131A where it was attributed mostlyto a high mineral rate of accumulation. The upper level has
TABLE 8Percentages of the Pollen Geographical Groups in the Pollen Sum
Percent in Pollen Sum of:
Nile pollensNorthern pollensChenopodiaceaeUndetermined + undeterminableAbsolute frequency
Samples
131A-1-2
86.61.0
11,51.0
2600
131A-4-2
88.53.44.43.6
800
130-2-3
14.975.5
9.00.2
32000
130-5-2
80.07.69.13.1
3100
128-3-5
3.183.0
9.44.4
3300
128-3-6
2.983.013.50.3
15200
125-3-5
15.7?74.0
9.51.0
21200
980

35. POLLEN ANALYSIS OF SAPROPELS
TABLE 9Percentage Composition of the Components of the Northern
Pollen Group as Related to the Pollen Sum and to theNorth Pollen Group Itself
MainNorthern
Components
DeciduousThermophileTrees
MediterraneanTrees
Artemisia
% Total
130-2-3
%ofPollenSum
3.6
37.8
24.0
65.4
%ofNorthPollen
5.5
57.8
36.7
100.0
Samples
130-5-2
%ofPollenSum
1.4
4.7
1.37.4
%ofNorthPollen
19.0
63.5
17.5
100.0
125-3-5
%ofPollenSum
12.2
30.4
17.5
60.1
%ofNorthPollen
20.3
50.6
29.1100.0
a very high absolute frequency which is common to almostall samples with dominant pelagic pollen.
The upper level is dated as early Pleistocene by someTertiary relics in a mostly Pleistocene pollen record(Pterocarya, Carya, Tsuga). This post-dates the uplifting.The lower level is, therefore, either even earlier Pleistoceneor late Pliocene.
It should be added that the three Nile-influencedsamples have a common palynofacies: a very cleanbackground, abundance of ligneous debris, and no amor-phous organic matter left after the technical preparation.This is not the case of the other sapropels.
Samples 128-3-5 and 128-3-6
These two samples, which come from the floor of theHellenic Trench west of Crete, have a very similar pollenspectrum. The dominant group is naturally the northernone with the same percentages in both samples. A verysmall group of Graminae and Cyperaceae appears. The term"Nile" pollens, as applied to them by analogy with theprevious samples, could well be misleading because of theubiquity of these families. But the tropical element isclearly missing, and the Pteridophytes are hardly repre-sented. Variation in the Chenopodiaceae is only very slight.In the northern pollens, however, a significant shift hasalready been noticed in the percentages of Artemisia andthe Mediterranean forest in the two samples. The phyto-geographical and climatic interpretations will be discussedwhen more data will become available.
Sample 125-3-5
Site 125 was drilled in an elevated area of theMediterranean Ridge providing undisturbed conditions ofsedimentation for pelagic and aeolian elements. This samplehas an original pollen spectrum with the Northern groupdominating; the pelagic nature of this pollen group is thusconfirmed.
SURVEY OF THE UPPER TERTIARY THERMOPHILERELICS IN SOUTH EUROPE (SPAIN, ITALY,
THE BALKANS)
The elements of middle-late Tertiary in northern Italy,which disappeared more or less early in Pleistocene or in
late Pliocene (Ricciardi, 1965a), include: Symplocos,Sequoia, Nyssa, Pinus haploxylon, Cedrus, Picea, Tsuga,Abies, Taxodium, Sciadopitys, Podocarpus, Liquidambar,Carya, Pterocarya, Juglans, cf. Engelhardtia, Myrica. Lona(1963a) has discovered Cedrus, Sciadopitys, Podocarpus,Pinus, Keteleeria, Abies, in a middle-upper Pliocene marineclay near Parma. In the Tertiary of Boca-Maggiora,Piemonte, Lona (1963b) has also described Sequoia, Pinushaploxylon, Symplocaceae, Cyrillaceae, Clethraceae, Celas-traceae, Sapindus, Castanea.
In northern Greece, the upper Pliocene flora ofAlmopias (Central Macedonia) (Mercier and Sauvage, 1965)includes Pinus haploxylon, Picea, Pseudolarix, Taxodiaceae,Pterocarya, Nyssapollenites, and deciduous thermophiletrees, still existing in the Balkans, along with Mediterraneanelements.
In Portugal (Diniz, 1967) the Pliocene appears with adominant Mediterranean character (Quercus cf. ilex,Oleaceae, Ericaceae, Cistaceae), a dominance of Pinusdiploxylon, and a well diversified herbaceous element,although the typically Tertiary trees and tropical elementsare also present {Liquidambar, Carya, Sapotaceae, Sym-plocaceae, Palmae, Keteleeria, Pseudolarix, Tsuga, Cedrus,Podocarpus).
Typically Tertiary pollens persist in Italy during thelower to middle Pleistocene. This is later than those foundin the Netherlands. In the Netherlands (Zagwijn, 1960), thecomplex Tsuga-Pterocarya-Carya has disappeared by theCromerian which is correlated in Italy with the Gunz-Mindel Interstadial. In Italy, in the upper part of the Leffeseries, near Bergamo at the foothills of the Alps, thecomplex Carya-Pterocarya-Tsuga occurs as a whole until theGunz MI Interstadial (Lona and Follieri, 1958) after whichit is dissociated. Carya and Pterocarya persist thereabundantly through the Gunz-Mindel Interglacial and thensporadically until the end of the Mindel II-III Interstadial.Tsuga is found intermittently from the Gunz II-IIIInterstadial to the Mindel HI Interstadial, where it becomesquite abundant again.
In the lower Leffe series (Lona, 1950), Liquidambar andNyssa are very rare. They do not appear in the upper Leffeseries. Zelkova does not range beyond the Gunz—MindelInterglacial in the upper Leffe series.
In central Italy, some trees which disappeared earlier innorthern Italy, persist until a later time (Zelkova, Liqui-dambar). Zelkova appears until perhaps the Riss inCimini-Sabatini, (Ricciardi, 1961b), or even the Riss-WürmInterglacial near Mazzano, Rome (Napoleone and Follieri,1967).
In the Leonessa basin, central Italy (Ricciardi, 1965b),Tsuga, Carya, Ptercocarya, Cedrus, and Pinus haploxylonare found in lower Pleistocene sediments and are correlatedwith the upper Leffe series. Liquidambar is present, butonly in the lower quarter of the diagram.
In the basin of Gubbio, central Italy (Lona andRicciardi, 1961), the sedimentary sequence is correlatedwith the upper Leffe series, and elements of the end of theTertiary also persist: Pinus haploxylon, Picea type orienta-lis, Tsuga, Cedrus, Pterocarya, Zelkova, Liquidambar. Thelatter is much more abundant in Gubbio than in Leffe; itappears with a later local expansion. In Pietrafitta near
981

M. ROSSIGNOL-STRICK
Perugia (Ricciardi, 1961a), Liquidambar, Zelkova, Ptero-carya, Tsuga are met in the Villafranchian. Indeed,Liquidambar has been with Zelkova, the most persistentTertiary relic in Italy.
Some typically Tertiary elements do not appear in Italyin the lower Pleistocene. Nyssa shows only few pollens inthe lower Leffe series and neither Sequoia nor Sciadopitysoccur at all. The latest occurrence of Sciadopitys seems tobe in the Apennines (Castell 'Arquato) in the Plio-Pleistocene marine deposits below the Calabrian and at theboundary between the Amphistegina layers and theirunderlying sandy clay layer, called Arquatian. At this level,it reaches an abundance of 6 per cent (Lona, 1962).Podocarpus appears last in the middle-upper Pliocene ofEmilia (Lona, 1963a).
CHRONOLOGY OF THE MEDITERRANEANNORTHERN SAMPLES
We can now attempt to date our Mediterranean samplesof northern influence on the basis of their percentages ofTertiary relics. We must keep in mind that the further southin Europe we go, the later Tertiary thermophile relicspersist. We must also take into account the distancebetween the north Mediterranean shores, the supposedplace of origin of the pollens, and the location where thepollens finally settled.
The Tertiary relics in our Mediterranean samples appearwith the percentages shown in Table 10. They include:Cedrus, Zelkova, cf. Engelhardtia, Liquidambar, Carya,Pterocarya, Juglans, Tsuga, and Podocarpus (north). Cedrushas been evaluated separately because its high percentagestend to mask the other trees.
Sample 125-3-5
The flora of Sample 125-3-5 is rich in Tertiary relicsincluding: Carya, Pterocarya, Tsuga, Liquidambar, Zelkova,Pinus haploxylon, and cf. Engelhardtia. However, sometypically Tertiary elements are missing, these are: Sequoia,Nyssa, Sciadopitys, and Picea of antique type, Taxodium.Therefore, a Tertiary age (Pliocene) is excluded. TheTertiary relics point to a lower Pleistocene age (Villa-franchian). This is not incompatible with the abundance ofthe Mediterranean forest and the dominance of Artemisia.It also agrees with the biostratigraphic zonation based onnannofossils (see Chapter 33) and Foraminifera (seeChapter 46).
The Villafranchian age makes it very likely that thePodocarpus pollen comes from the north and not from theNile.
Sample 130-2-3
Further to the east and slightly more to the south,Sample 130-2-3 shows the same Tertiary relics, althoughthey are much less abundant. They are Tsuga, Pterocarya,Carya, Liquidambar, and Zelkova. The four Podocarpuspollens are probably also of northern origin, rather thanNilotic. The pollen spectrum has the same structure as125-3-5, with fewer deciduous thermophile trees. Therelative frequencies of the other groups are rather simi-lar. The probable main source is the southern coast ofTurkey.
TABLE 10Percentage of Tertiary Relics in the Pollen Count of the
Eastern Mediterranean
Per Cent ofTertiary
Relics
In the T o t a l
Pollen _ ,Sum
Portion
In the TotalNorthernPollen CedrusGroup Portion
131A1-2
0.0
131A4-2
0.0
1302-3
1.2
0.4
1.6
0.6
Samples
1305-2
0.0
1283-5
1.9
1.9
2.3
2.3
1283-6
0.9
0.5
1.1
0.6
1253-5
7.1
1.1
9.6
1.5
The variety of Tertiary relics would suggest a similarlower Pleistocene age for this sample, but its percentage,much lower than in 125-3-5, may point to a somewhat laterperiod. It could then be either late lower Pleistocene orearly middle Pleistocene. The probability of a later age isalso increased by the fact that Turkey is closer thansouthern Italy or Greece to the last and still existing refugesof the Tertiary thermophile relics, that is the Caucasus andnorthern slopes of the Elbourz facing the Caspian Sea.Therefore, the Tertiary relics may have remained later inTurkey than further west, and do support a more recentdating than would be probable in the west.
Sample 130-5-2If there is no stratigraphical disturbance in Site 130
between Samples 5-2 and 2-3, an older age-maybe lowerVillafranchian-should be attributed to the deeper 130-5-2sample (its dominant Nile spectrum could not (yet) providea dating clue). Table 9 shows, among the northernelements, the same percentages of deciduous thermophiletrees as Sample 125-3-5.
Sample 128-3-6
The Teritiary element is greatly reduced in thissample, and half of it is represented by Cedrus, even thoughthe station is located closer to the northern shore than Site125. Besides Cedrus, only Zelkova and Pterocarya remain.The suggested age is middle or late Pleistocene. A post-Wurm sapropel, previously studied near southeast Crete anddated 7900 BP. (M. Rossignol and L. Pastouret, 1971), alsoshows Cedrus, and a few pollens of Zelkova, Peterocarya,and Liquidambar.
Therefore, these trees persisted very late in thisarea. Their higher abundance in 128-3-6 than offsoutheast Crete suggests an earlier age — possibly theMindel-Riss Interglacial.
Sample 128-3-5
Only Cedrus appears here. This sample is onlyslightly higher in the sequence than the previous oneand therefore has an equivalent age. The absence hereof the Tertiary relics, which still persist in theHolocene off southeast Crete, may not have a chron-ologic implication, but may only reflect sedimentmixing during redeposition.
982

35. POLLEN ANALYSIS OF SAPROPELS
Dinoflagellates CystsTwenty-three Dinoflagellate cysts species have been
recorded. Their absolute frequency has been discussedabove.
The most important group is Spiniferites furcata (Ehr.)O. Wetzel, S. mirabilis Rossignol. Operculodinium Wallappears with 5 species. Hystrichokolpoma rigaudaeDeflandre and Cookson appears only in the two oldestsamples of pelagic sediment from the Mediterranean Ridge(125-3-5 and 130-2-3), but Tuberculodinium van campoae(Rossignol) Wall is absent there.
A number of cysts of unknown systematic positionappeared also. A detailed study of all the cysts is underway.
Some fresh water microfossils—Algae and uncertainaffinities—occurred only in samples of Nilotic influence(Pediastrum [131A-1-2: 17; 131A-4-2: 32; 130-5-2: 12],Botryococcus [131A-1-2: 8; 131A4-2: 3; 130-5-2: 11]Concentricystes rubinus Rossignol [131 A-1-2: 2; 131A-4-2:
SUMMARY
Seven samples from four deep sea drilling sites in theeastern Mediterranean have been analysed for their pollen:two cores south and southwest of Greece, two north ofEgypt.
The pollen spectra belong to two categories. One isdominated by the pollens coming from the northern shoresof the Mediterranean; the other one is strongly character-ized by pollen carried into the sea by the Nile River. In thefirst group, appear the following phytogeographical types:Mediterranean trees {Pinus, Quercus, Cedrus, Olea), decid-uous thermophile trees (Fagus, Acer, Castanea, Alnus,Ostrya, Corylus, Carpinus, Carya, Pterocarya, Juglans,Engelhardtia, Liquidambar, Tilia, Ulmus, Zelkova, Frax-inus), other trees (Tsuga, Abies, Be tula), shrubs (Ericaceae,Thymeleaceae, Buxus, Poterium, Myrica), and, among theherbaceous, Artemisia and an array of other families. Thesecond group includes Graminae, Cyperaceae, Pterido-phytes, hydrophytes, tropical trees and grasses. All thesamples close to Greece and the upper sample of one of thecores from the Mediterranean Ridge north of the Nile Conebelong to the first group. The three other samples from theRidge and the Nile Cone belong to the second group.Chenopodiaceae are equally present in all samples. Theyseem to result from a rather constant pollen rain on a largesea area after a long wind transport in the upperatmosphere. They may come from the steppe and desertregions of Africa and the Middle East.
The north-Mediterranean pollen may have been trans-ported from their source area first by wind for a shortdistance to the nearby sea surface. Indeed, it is suggestedthat their source area is in the littoral zone including thefirst range of mountains in the back-country of southernItaly, Greece, and Turkey. Once arriving in the sea, theyfloat in the superficial waters, and behave as pelagicelements. Their distribution in the sediments does notdepend upon the bottom topography, but rather upon thedistance from the shores and direction and magnitude ofthe currents and winds.
Conversely, distribution in the sediments of pollens fromNilotic origin is markedly influenced by the bottomtopography. This suggests these pollens behave as benthicelements; transported in density currents of the deep watermasses with the mineral-rich Nilotic sediments.
The number of pollens per gram of sediment is muchhigher for the pelagic pollens (up to 32000) than for thebenthic pollens (average 2000).
A chronology has been made possible by the presence—or absence—in the samples of Northern origin, of pollens oftrees which belong to a Tertiary relictual flora still living inEurope at the beginning of the Pleistocene: Pterocarya,Carya, Tsuga, Juglans, Liquidambar, Zelkova, Cedrus, Pinushaploxylon.
Nearly all of this flora progressively became extinctduring the Quaternary. A Pliocene age is excluded by theabsence of Nyssa and Sciadopitys. The proposed datationsfor the samples of northern influence extend from lowerVillafranchian to the Mindel-Riss Interglacial. At least oneof the Tertiary relics, Cedrus, and maybe also Zelkova,disappeared from the continent only in Holocene times,probably under the influence of man. The vegetal landscapein southern Italy, Greece and southern Turkey coastal areaswas modified during the Pleistocene by the vertical shiftingof the vegetal belts which appear in the pollen spectra. TheArtemisia steppe, at the expense of the Mediterraneanforest, is attributed mostly to an increased dryness, as is theprogressive disappearance of the deciduous thermophiletrees which did not find the refuge they would have neededto survive in the vicinity. High mountains further south,provided both humidity and warmth. A return of theMediterranean forest in the youngest sample is attributed toa more humid phase. The previous disappearance of thedeciduous thermophile trees prevented them fromspreading again. The temperature increased slightly.
The pollen analysis of these Quaternary marine sapropelsyields information about transport and sedimentation ofpollen in the sea, and the geographic origin of such pollen.It is an excellent tool for datation which also allowsreconstruction of the history of the original vegetation inthe coastal areas, the history of the vegetal extinctionsduring the Quaternary, and the climatic evolution. Thepursuit of these investigations appears extremely promising.
CONCLUSIONS
It is hoped that this preliminary study will show thegreat impact of marine palynology on studies of thisparticular geographical area. Thanks to the geobotanicaldifferences between the northern and southern shores ofthe eastern Mediterranean the origin of the pollen canindeed be pointed out. The distribution of the pollenprovides information on the processes of pollen transportand sedimentation. This, in turn, is related to the tracing ofthe terrigenous sediments once they reached the sea, to themovements of the water masses, and to the tectonic historyof the sea-bottom.
Palynological long-distance correlations appear possiblethanks to the widespread, non-localized character of themarine pollen spectra, even though the variety of places oforigin of the pollen results in a recognizable sedimentationgradient. The marine pollen spectra is believed to represent
983

M. ROSSIGNOL-STRICK
partly the vegetation of the littoral areas. It keeps track ofthe global variations in altitude of the different vegetalbelts, allowing one to attempt to describe the climatichistory. The progressive extinctions of the Tertiary relicsthrough the Quaternary finally provides a chronology fordating even isolated samples such as those studied here.Indeed, this preliminary study calls for further research inthis field, and already gives assurance of great interest.
ACKNOWLEDGMENTS
The author, who has been supported by the CentreNational de la Recherche Scientifique, Paris, France, wishesto thank the Lamont-Doherty Geological Observatory ofColumbia University, Palisades, New York, and the Depart-ment of Geology, Imperial College, London, for providingher with their laboratory facilities. She thanks also W. B. F.Ryan for reading and editing the manuscript, and providingthe available sedimentology and lithology information onthe samples used in this study.
RESUME
Sept niveaux très riches en matière organique provenantde quatre forages profonds en Méditerranée Orientale ontété analyses: deux au Sud et Sud-ouest de la Grèce, deux auNord de 1'Egypte.
Les spectres polliniques se rangent en deux categories,où dominent, dans Tune, les apports des rivages Nord de laMéditerranée, dans 1'autre, les apports du Nil.
La premiere categorie groupe les types phytogeo-graphiques suivants: arbres mediterraneens (Pinus, Cedrus,Quercus, Olea), arbres décidus thermophiles (Fagus, Acer,Castanea, Alnus, Ostrya, Corylus, Carpinus, Carya, Ptero-carya, Juglans, Engelhardtia, Tilia, Liquidambar, Ulmus,Zelkova, Fraxinus), arbres divers (Tsuga, Abies, Betula),buissons (Ericacees, Thymele'acees, Buxus, Poterium, Ilex,Myrica) et parmi les herbacees, Artemisia, et un cortege defamilies diverses. Les apports du Nil comprennent Gram-inees, Cyperacees, Pteridophytes, hydrophytes, arbres etherbacees tropicales. Tous les echantillons proches de laGrèce, et le niveau superieur d'un des deux forages prochesde 1'Egypte, appartiennent á la premiere categorie. Lesapports du Nil se trouvent dans les trois autres echantillonsproches de FEgypte.
Les Chenopodiacees, egalement representees partout,semblent resulter d'une pluie pollinique assez constante,repartie sur toute la surface de la mer, sans doute après unlong transport aerien dans la haute atmosphere. Ellespeuvent provenir des zones steppiques et de'sertiquesd'Afrique, ou du Moyen-Orient.
Les pollens nord-mediterrane'ens auraient subi d'abordun bref transport aerien depuis leur vege'tation d'origine,situee, semble-t-il,dans la region còtière dTtalie du Sud, deGrèce et de Turquie, y compris les premieres zonesmontagneuses de l'arrière-pays, jusqu'á la surface de la merà proximite du rivage. Us flottent ensuite á la surface, et secomportent alors comme des elements pèlagiques. Leursedimentation ne depend pas de la topographie du fondmarin, mais plutot de la distance des cßtes, des courants,des vents.
Au contraire, la sedimentation des pollens d'originenilotique depend de la topographie du fond marin. Celasuggère qu'ils se comportent comme des e'le'ments ben-thiques, transported peu au-dessus du fond marin avec lacharge sedimentaire miner ale du fleuve. Le nombre depollens par gramme de se'diment est beaucoup plus eleve'pour les echantillons á dominante pelagique (jusqu'á32000), que pour les benthiques (en moyenne 2000).
Une chronologie est rendue possible par la presence—ou1'absence—dans les echantillons d'influence nordique, depollens d'arbres appartenant á une flore relictuelle tertiaire,vivant encore en Europe du Sud, Italie surtout, Grèce,Espagne, au debut du Pleistocene: Pterocarya, CaryaK Tsuga,Juglans, Liquidambar, Zelkova, Cedrus, Pinus haploxylon.Presque toute cette flore s'est eteinte progressivement aucours du Quaternaire. Un age Pliocene est exclu par1'absence de Nyssa, Sciadopitys. Les ages proposes pour lesdivers echantillons d'influence nordique vont du Villa-franchien inferieur á FInterglaciaire Mindel-Riss. Au moinsune des reliques tertiaires, Cedrus, et peut-ëtre aussiZelkova, n'ont disparu du continent qu'á 1'Holocène, sansdoute sous 1'influence de 1'Homme.
Le paysage vegetal dans la region cotière dTtaliemeridionale, Grèce, et Turquie, s'est modifie par undeplacement vertical des zones de vegetation qui apparais-sent dans les spectres polliniques: la steppe á Artemisia aubord du rivage, dans les plaines cótières et sur les plateauxde l'arrière-pays, la forèt mediterraneenne sur les pentesbasses et moyennes, dont les elements les plus humides semelent vers les sommets avec les arbres décidus ther-mophiles qui necessitent des pluies d'e'te'. Dans ces regionsmediterraneennes, le facteur climatique limitant est lapluviosite'. Les variations de temperature ont une moindreinfluence.
L'extension de la steppe á Artemisia est ainsi attribuéesurtout á une diminution de la pluviosite; de méme, ladisparition progressive des arbres décidus thermophiles. Unretour de la forèt mediterraneenne dans le niveau le plusrecent est attribue á une phase plus humide. La disparitk>nanterieure des arbres décidus thermophiles les empéche deregagner du terrain.
L'analyse pollinique de ces sediments organiques marinsQuaternaires fournit done des renseignements, á la fois surle transport et la sedimentation polliniques en mer, surForigine geographique du pollen et sur celle des sedimentsqui les recèlent; c'est un excellent outil de datation, quipermet en outre de retracer Fhistoire de la vegetationoriginelle dans les regions cótières, celle des extinctionsvegetales au cours du Quaternaire, ainsi que Involutionclimatique. La poursuite de ces recherches apparait commeextrëmement prometteuse.
REFERENCES
Diniz, F., 1967. Une flore tertiaire de caractère Mediter-ranean au Portugal. Rev. Palaeobot. Palynol. 5, 263.
Emery, K. O., Heezen, B. C, and Allan, T. D., 1966.Bathymetry of the eastern Mediterranean Sea. Deep SeaRes. 13, 173
Koreneva, E. V., 1969. Palynological investigations ofsediment cores from the Mediterranean Sea. In Generalproblems of Micropaleontology and organic sedimentsin Oceans and Seas, Nauka, Moscow, 268, (in RussianEngl. Summary).
Kuprianova, L. A., 1960. Palynological data contributing tothe history of Liquidambar. Pollen Spores. 11 (1), 71.
Lona, F., 1950. Contributo alia storia della vegetazione edel clima nella Val Padana. Analisi polliniche delgiacimento Villafranchiano di Leffe (Bergamo). Atti Soc.Ital. Sci. Nat. 89, 123.
, 1962. Prime analisi pollinologiche sui depositiTerziari-Quaternari di Castell 'arquato: reperti di vegeta-zione da clima freddo sotto le formazioni calcaree adAmphistegina, Boll. Soc. Geol. Ital. 81 (1), 89, (Res.Engl).
984

35. POLLEN ANALYSIS OF SAPROPELS
, 1963a. Boschi di Cedrus e Sciadopitys in Emiliadurante il Pliocene (Riassunto). Nuovo Giorn. Botan.Ital. 70,573.
., 1963b. Alcune tipi di pollini fossili rinvenuti indepositi terziari del Piemonte (Riassunto). Nuovo Giorn.Botan. Ital. 70, 574.
Lona, F. and Follieri, M., 1958. Successione pollinica dellaserie superiore (Günz-Mindel) di Leffe (Bergamo).Veröff. Geobot. Inst. Rubel Zurich. 34, 86.
Lona, F. and Ricciardi, E., 1961. Studio pollinologicostratigrafico su una serie lacustre pleistocenica dell 'Italiacentrale (Bacino di Gubbio, Perugia). Pollen Spores, Fr.1,93 (Res Fr.).
Mercier, J. and Sauvage, J., 1965. Sur la geologie de laMace'doine centrale: les tufs volcaniques et les forma-tions volcano-detritiques plioènes á pollens et sporesd'Almopias (Grèce). Ann. Geol. Pays Helléniques. 16,188.
Napoleone, I. and Follieri, M., 1967. Pollen analysis of adiatomite near Mazzano (Rome). Rev. Palaeobotan.Palynol. 4(1-4), 143.
Ricciardi, E., 1961a. Analisi pollinologica dei sedimentilacustri lignitiferi di Pietrafitta (Perugia, Italia). PollenSpores, Fr. 3 (2), 325 (Res. Fr.).
, 1961b. Considerazioni preliminari sui pollinifossili rinvenuti in un deposito lacustre Quaternariocontenente materiali vulcanici (Gruppo Cimini-Sabatini).Boll. Soc. Geol. Ital. 80 (4), 129 (Res. Engl).
., 1965a. Pollini fossili terziari nell 'Italia del nord(Piemonte) e loro significato paleoclimatico. Bull. Soc.Geol. Ital. 82 (1), 111 (Res. Engl.).
1965b. Analisi polliniche di una serie strati-grafica dei sedimenti lacustri del Pleistocene inferiore nelbacino di Leonessa (Rieti, Italia centrale). B. Bot. Ital.72(1), 62 (Res. Engl.).
Rossignol, M., 1961. Analyse pollinique de sedimentsmarins quaternaires en Israėl-1: se'diments recents. PollenSpores. 3 (2), 303.
, 1969. Sedimentation palynologique dans ledomaine marin quaternaire de Palestine: etude depaleo-environement. Notes Mém. Moyen-Orient, Fr., X,273 p.
Rossignol, M. and Pastouret, L., 1971. Analyse polliniquede niveaux sapropeliques post-glaciaires dans une carotteen Me'diterranee Orientale, Rev. Palaeobotan. Palynol.11,227.
Ryan, W. B. F., 1969. The deep-sea floor of theMediterranean, PhD. Thesis, Columbia University, NewYork.
Tralau, H., 1963. Asiatic dicotyledonous affinities in theCainozoic flora of Europe. Handl. Kgl. Svenska Veten-skapsakad. 4 (9, 3), 87 p.
van der Hammen, T., Wijmstra, T. A., and van der Molen,W. H., 1965. Palynological study of a very thick peatsection in Greece, and the Würm-glacial vegetation in theMediterranean region. Geol. en Mijnbouw. 44 37.
van Zeist, W., 1967. Late Quaternary vegetation history ofwestern Iran.i?ev. Palaeobot. Palynol., Nederl, 301.
Venkatarathnam, K. and Ryan, W. B. F., 1971. Dispersalpattern of clay minerals in the sediments of the easternMediterranean, Marine Geol. 11, 261.
Vronsky, V. A. and Panov, D. G., 1963. Composition andrepartition of pollens and spores in the superficial layerof marine sediments of the Mediterranean Sea (inRussian). Dokl. Akad. Nauk, SSSR. 153 (2) 447.
Wijmstra, T. A., 1969. Palynology of the first 30 meters ofa 120 meter deep section in Northern Greece. Acta. Bot.Neerl. 18(4), 511, 527.
Zagwijn, W. H., 1960. Aspects of the Pliocene and earlyPleistocene vegetation in the Netherlands. Mededelingenvan de Geolog. Sticht. C, 111, 1, 5, Maastricht. 78 p.
Zohary, M., 1963. On the geobotanical structure of Iran.Bull. Res. Counc. Israel. 11 (D), (suppl.) 113 p.
985

M. ROSSIGNOL-STRICK
PLATE 1
Locations: Figures 1-6, 10, 11 (Sample 125-3-5); Figure 7 (Sample128-3-6); Figures 8, 9 (Sample 130-2-3); Figures 12, 13 (Sample131A-4-2). Scales: 10 microns: A for Figures 1-4, 6, 11; B for Figures5,7, 10, 12, 13; C for Figures 8, 9.
Figures 1, 2, 5. Zelkova
Figures 3, 4, 6, 7. Ulmus
Figures 8, 9. Olea
Figure 10. Cyatheaceae
Figure 11. Riccia
Figures 12, 13. cf Hypoestes (Acanthaceae)
986

35. POLLEN ANALYSIS OF SAPROPELS
PLATE 1
987

M. ROSSIGNOL-STRICK
PLATE 2
Locations: Figures 1, 13, 16, 17 (Sample 130-2-3); Figure 3 (Sample128-3-6); Figures 2, 4, 5, 6, 7, 8, 9, 10, 11, 12, 14, 15, 18 (Sample125-3-5). Scales: 10 microns: A for Figures 2, 5, 6, 7, 8, 9, 15, 17; Bfor Figures 10, 11, 16, 18; C for Figures 12, 13; D for Figures 1, 3, 4,14.
Figures 1,8.
Figure 2.
Figures 3, 4.
Figures 5,10.
Figure 6.
Figure 7.
Figure 9.
Figure 11.
Figure 12.
Figure 13.
Figure 14.
Figures 15,17. Tsuga
Figure 16. Pinus haploxylon
Figure 18. Ostrya
Corylus
Carpinus caucasica
Betula
Liquidambar
Ericaceae
Alnus
Ostrya
Rumex
Plantago
Poterium spinosum
Quercus
988

35. POLLEN ANALYSIS OF SAPROPELS
PLATE 2
989

M. ROSSIGNOL-STRICK
PLATE 3
All pollens are from Sample 125-3-5, except 5, 6 and 10 which are fromSample 130-2-3. Scale bar represents 10 microns.
Figures 1-6, 10. Unknown tricolporate pollen (see p. 10).
Figure 1. Polar contour.
Figure 2. Equatorial contour.
Figure 3. Polar exine.
Figure 4. Exo-and endoapertures.
Figures 5,6,7. Three different focusings of the same pollen, inoblique view.
Figure 5. Polar area, lower surface, with a colpus (vertical).
Figure 6. Mesocolpial and pericolpial areas, with a colpus(oblique).
Figure 7. Optical section. The three optical sections, Figures 1,2, 10 show the strong columellation and the apparentabsence of an endexine.
Figures 7, 8. Carya
Figures 9, 11. Pterocarya
Figure 12. Tilia
990

35. POLLEN ANALYSIS OF SAPROPELS
PLATE 3
991