Embed Size (px)
Citation preview

SD9900043
By:Elham Sharief Dawood
B.Sc. (HONS.)andM.Sc.
A thesis submitted for the fulfillment of the requirements for the degree ofdoctor of philosophy.
Department of BotanyFaculty of ScienceUniversity of KhartoumDecember 1997
4 •3 *
^ '3 0 - 4 4
'•-4.0

Dedication:
t o fye souls of roy father and n?y uncle Hassan.Dedication is extended to all my family members specially njy
sisters, brothers and to rpy aunts Samia and Vaf)eba.
u

, l
o ami, pamkjuloJwj.- majwvin, to- ma- MMWUMAM, i n . hA.dv'vaiwrh. f&\- ruAI f II ' II
rwrul MuwwiAi/yri, iAUKilujoM/b axhtk/b wrul (jswAW/rw vrwsu/vwmwnt.
Q / 9 9 / W J- A9T\ M / • -/ 9 •/' 9- -9 I0 ami q/vaMLul to Vnwjj^mlml (A/rwoAtmi' W\> w^ u/navvcxAil MMponl.
0 6 o S S II
hjMJual tna/rJnA to tfw ujola/rw, Sopo/wrrwrd', U/rwQAAiiu, <$l Jlka/ifowm \m>
9 -9999 -if0// OAHKMJPW UMmkWSi,f 6
"W/ alM- OUHJ to- tm SmoAA/rm/nl <al WWWQU-. Aaaih- mmlxhI 6 of'
9 / - •/ 9 /• 9 99 V J 9 99 • /J i f f I I I fi/ih inMmkjauMh op ty
I77) 19 9 9 f i l l 96 9 - 1 - 99 *)! 1 V//(//• uba/rJRA oho* al^& HMMUUHOL to- all vw> lAwrvaA \WttMMAi, lAMa/h Jlij^^ai/njP9 I • 9 °il I /P 9 I Q 9 J fj V I W 9 J 9 It •(MhanM', Ikhlajh [fiomal "Ja/ml OAUI Jj'i.jlvMuim llusfui/rmxi mn, itvu/i-
wrui/miwL mm-,
di/rbaEu< mu- Uva/wA, OM O/AG- axih to- //1/iA. Illaqwh UluMalxi l/y\< hipi/nn-
in

Sixty six bacteria and twenty fungi were isolated from various sources.
These varied from rotten fruits to local drinks and soil samples from
different parts of Sudan. On the basis of index of amylolytic activity, forty
one bacteria and twelve fungi were found to hydrolyse starch.
The best ten starch hydrolysing bacterial isolates were identified all as
bacilli {Bacillus licheniformis SUD-K,, SUD-K2, SUD-K^, SUD-O, SUD-
SRW, SUD-BRW, SUD-By, Bacillus subtilis SUD-K3 and Bacillus
circulans SUD-D and SUD-K7). Their amylase productivity was studied
with respect to temperature and time.
The highest activity was produced in around 24 hours of growth in all; six of
which gave the highest amylase activity at 50°C and the rest at 45°C.
Based on the thermal production six isolates were chosen for further
investigation. These were Bacillus licheniformis SUD-Ki, SUD-K2, SUD-
K4, SUD-O, Bacillus subtilis SUD-K3 and Bacillus circulans SUD-K7. The
inclusion of starch and Mg++ ions in the culture medium gave the highest
enzyme yield. The pH 9.0 was found to be the optimum for amylase
production for all isolates except Bacillus subtilis SUD-K3 which had an
optimum at pH 7.0. Three isolates {Bacillus licheniformis SUD-K 1, SUD-K4
and SUD-O) recorded highest amylase production in a medium
supplemented with peptone while the rest {Bacillus licheniformis SUD-K2,
Bacillus subtilis SUD-K3 and Bacillus circulans SUD-K7) gave highest
amylase productivity in a medium supplemented with malt extract. Four
isolates {Bacillus licheniformis SUD-K 1, SUD-K2 and Bacillus subtilis
SUD-K3) gave maximum amylase production in a medium containing 0.5%
iv

soluble starch while the rest (Bacillus lichenifonnis SUD-0 and Bacillus
circulans SUD-K7) gave maximum amylase production at 2%. Soluble
starch was found to be the best substrate among the different carbon sources
tested.
The maximum temperature for amylase activity ranged from 60-70°C and
1% starch concentration was optimum for all isolates. Addition of different
metal ions and different concentrations of sodium chloride separately to the
reaction mixture suppressed the enzyme activity.
Hydrolysis pattern of the substrate soluble starch by these amylases
indicated that higher ratio of oligosaccharides and maltose are the main
products and this indicated that these enzymes can be identified as a-
amylases. The a-amylases were partially purified up to 15-19 fold of their
initial activities. The Kin values of all isolates were in the range of 1.25-2.0
mg/ml.

Contents
1.
2.2.1.2.2.2.2.1.2.2.2.2.2.3.2.2.4.2.3.2.4.2.5.2.6.2.7.2.8.2.9.
3.3.1.3.1.1.3.1.2.3.2.
DedicationAcknowledgementsAbstractArabic abstract
Chapter One
Introduction
Chapter Two
Literature ReviewGeneral DescriptionSources of EnzymesAnimalsPlantsFungiBacteriaThe Genus BacillusOptimization of Amylase SynthesisPurificationHydrolytic Products of AmylasesStability and DenaturationAmylase KineticsApplication of Amylases in Industry
Chapter Three
Materials and MethodsMaterialsMicroorganismsChemicalsScreening of Microorganisms
Pageiiiiiivvi
1
33455679101213141517
1919191919
vi n

3.2.1.3.2.2.3.2.3.3.2.4.
3.2.4.1.3.2.4.2.3.2.4.3.3.2.4.3.1.3.2.4.3.2.3.2.4.3.3.3.2.4.3.4.3.2.4.3.5.3.2.4.3.6.3.2.4.3.7.3.2.4.3.8.3.2.4.3.9.3.2.4.3.10.3.2.4.3.11.3.2.4.3.12.3.2.4.3.13.3.2.4.3.14.3.2.4.3.15.3.2.4.3.16.3.2.5.3.2.5.1.3.2.5.2.3.2.5.3.3.2.5.4.3.2.5.5.3.2.5.6.3.2.6.
3.2.6.1.3.2.6.2.3.2.6.3.3.2.6.4.3.2.6.5.3.2.6.6.3.2.6.6.1.3.2.6.6.2.3.2.6.6.3.
Screening of Bacterial IsolatesIsolation of FungiSelection of IsolatesMorophological and BiochemicalCharacterization of Bacillus sp.Morophological TestsStaining of the SporeBiochemical TestsProduction of CatalaseVoges Proskauer TestProduction of Acid from CarbohydratesHydrolysis of StarchLiquefaction of GelatineReduction of Nitrate to NitriteDeamination of PhenylalanineProduction of IndoleProduction of Dihydroxy AcetoneUtilization of Citrate and PropionateGrowth in Sodium ChlorideGrowth at pH 5-7Determination of Optimum Growth TemperatureHydrolysis of CaseinAnaerobic GrowthEgg Yolk ReactionAmylase ProductionEffect of TemperatureEffect of pHEffect of Substrate ConcentrationEffect of Different Organic NitrogenEffect of Metal IonsEffect of Different Carbon SourcesDetermination of Amylolytic Activity(Enzyme Assays)Preparation of BufferPreparation of 3,5 Dinitrosalcylic Acid (DNS)Determination of Enzyme ActivityDetermination of ProteinsEnzyme Units and Specific ActivityAmylase ActivityEffect of Temperature on Amylase ActivitypH OptimaEffect of Substrate Concentration
192021
212222222223232424242525262627272727282829292930303031
31313132323333333333
IX

3.2.6.6.4. Effect of Reaction Time 343.2.6.6.5. Effect of Divalent Cation on Amylase Activity 343.2.6.6.6. Effect ofNaCl on Amylase Activity 343.2.7. Identification of Enzyme Products 353.2.8. Enzyme Purification 353.2.8.1. Column Chromatography 353.2.8.1.1. Packing and Equilibration of the Column 353.2.8.1.2. Application of the Enzyme on Sephadex A-25 363.2.9. Storage Stability 373.2.10. Determination of Km and Vinax. 37
Chapter Four
4.4.1.
4.1.1.4.2.4.3.4.3.1.
4.3.2.4.3.3.
4.3.4.
4.3.5.
4.3.6.
4.4.4.4.1.4.4.2.4.4.3.
4.4.4.
4.4.5.
4.4.6.
Results and DiscussionScreening of Bacterial and Fungal Isolates forAmylase ProductionSelection of IsolatesIdentification of Bacterial IsolatesAmylase ProductionEffect of Temperature and Time Intervals onAmylase ProductionEffect of pH on Amylase ProductionEffect of Substrate Concentration on AmylaseProductionEffect of Different Metal Ions on AmylaseProductionEffect of Different Organic Nitrogen on AmylaseProductionEffect of Different Carbon Sources on AmylaseProductionAmylase ActivityEffect of Temperature on Amylase ActivityEffect of pH on Amylase ActivityEffect of Different Time Intervals on AmylaseActivityEffect of Substrate Concentration on AmylaseActivityEffect of Different Metal Ions on AmylaseActivityEffect of NaCl on Amylase Activity
38
38384245
4546
47
48
49
51525253
53
54
5455

4.5.4.6.4.7.
4.8
4.9.
Identification of Enzyme ProductsPartial Purification of a-Ainylase EnzymeEffect of Storage Temperature on AmylaseActivityDetermination of Km and Vmax Valuesof a-AmylaseConclusionReferences
5556
58
586163
X]

Amylases are enzymes that hydrolyze starch to give oligosaccharides,
maltose or ultimately glucose.
They overshadow all others in industrial significance and utilization.
The use of amylases appears to be as ancient as civilization itself (Kiieen,
1949). It has been stated that the earliest known records of the preparation
of malt date back to 7000 B.C. and beer brewing was an established craft by
5000 B.C. (Lazer, 1938).
The real foundation for the scientific approach to the problem of
enzymatic starch degradation lies in the discovery of diastase by Kirchhoff
(1816) and Payen and Persoz (1833). Coinciding with utilization of cereal
malt amylases was an equally important discoveiy that certain types of
molds and bacteria produced starch degrading enzymes (Kneen, 1949).
In the first six decades of this century, the starch processing industry
employed dilute mineral acids to hydrolyse starch and the hydrolysate was
then neutralized and refined to a thick syrup. This syrup is widely used in
brewing, backing, food and confectionery and many other industries.
However, the use of acids is always accompanied by colour formation and
reversion products. Salt is also produced as a result of acid neutralisation
and would require an extra step to get rid of.
Nowadays the starch processing industry depends on the reactions of
amylase enzymes which include liquefaction or solublization of starch,
production of dextrins, or hydrolysis of starch to a high percentage of
reducing sugars such as maltose or dextrose (saccharogenic amylases). This

starch processing industry is considered as the largest user of enzymes,
mainly owing to the success of the a-amylase and amyloglucosidase-based
process for forming glucose syrup. These enzymes are relatively
inexpensive. However, the industry is always hungry for better enzymes
specially those that are relatively more stable at relatively elevated
temperature.
Objectives of this Work:
1. Screening of microbial species for the production of amylases.
2. Selection of the species that are thermophilic and determining their
optimum conditions for production of these enzymes.
3. Isolation, characterization and purification of these enzymes.

2.1. General Description:
Payen and Persoz (1833) demonstrated that an alcohol precipitate of
malt extract contained a thermostable substance capable of converting starch
into sugar. This substance was called diastase.
Amylases are hydrolytic enzymes which break down many
polysaccharides such as starch or glycogen to yield oligosaccharides or
disaccharides such as maltose and in some cases monosaccharides such as
glucose.
(Cr,H|2O6),, + nH2O • Amylase n(Ci2H22O,,)
starch maltose
The reaction is accompanied and characterized by four changes.
These are: (i) decrease in viscosity of the reaction mixture indicating the
cleavage of the polysaccharide chain (it) loss of the capacity to give a blue
colour with iodine (iii) appearance of reducing groups and (iv) formation of
maltose or glucose or oligosaccharides of varying chain length.
Among the known amylases there are three types designated a-
amylase, f5-amlyase and amyloglucosidase.
a-Ainylases (E.C.3.2.1.1, cc-1, 4-D glucon glucanohydrolase) are
endoenzymes catalyzing the conversion of starch to a range of
maltooligosaccharides by endo-acting mechanism (Fogarly and Kelly, 1980,
1990) and (Fogarty, 1983). These enzymes are, also, known as dextrino
genic or liquefying amylase.

a-Amylases hydrolyze the intenial a-1,4-glycosidic links in amylose
and amylopectin at random to yield soluble, less viscous, lower molecular
weight product in an a-configuration at the reducing glucose end. Some ot-
amylases produce moieties in the form of maltose and rarely traces of
glucose.
P-Amylase (E.C.3.2.1.2, cc-1,4 glucan malto-hydrolases) hydrolyzes
1 -4-oc glucan link in polysaccharides to remove maltose units from the non
reducing ends of the chain. This enzyme is an exoenzyme or exoglycosidase,
because it attacks the ends and produces maltose in its p-fonn. P-Amylase is
also known as saccharogenic amylase due to the early appearance of
reducing sugars. It does not act on the a - 1 , 6 bond in amylopectin and thus
form limit P-dextrin. As a result the production of maltose is limited (Peat, el
al, 1952; Hassid and Newfeld, 1955).
Amyloglucosidase (E.C.3.2.1.3, glucoamylase-a-amylase or a-1,4
glucan glucohydrolase) is an exoenzyme catalyzing the hydrolysis of a-1,4
bond in both starch and oligosaccharides releasing P-glucose from the non
reducing end of the molecules. This enzyme also hydrolyses the a-1,6 links
in amylopection to yield a glucose syrup containing 95-97% glucose, a
dextrose equivalent to 97-98% and 3-5% oligosaccharides (Bhuinibhamon,
1986).
Glucoamylases are isoenzymes occurring in two or three different
forms (Fleminig and Stone, 1965; Lineback and Aira, 1972).
2.2. Sources of Enzymes:
Amylases are widely distributed among animals, plants and
microorganisms. a-Amylases occur in microorganisms, plants and animal
tissues whereas P-Amylases occur predominantly in higher plants and

microorganisms (Karlson, 1974). Amyloglucosidase occur mainly in micro
organisms and in some animal tissues (Wiseman, 1985).
2.2.1. Animals:
a-Amylases of human saliva, human pancrease and pig pancrease
have been obtained in a crystalline form (Meyer et al. , 1951; MyrbSck and
Newmiiler, 1951 and Buisson et al. , 1987)
a-Amylases of saliva initiates the hydrolytic attack on the dietary
polysaccharides, producing maltooligosaccharides, while a-amylase of
pancreas is secreted as juice into the small intestine and completes the
digestion process yielding maltose which is hydrolysed by a- glucosidase to
glucose (Laner and Nickle, 1955). Amyloglucosidase hydrolyses longer
oligosaccharides and completes the degradation in the intestinal tract
producing glucose. This enzyme also occurs in muscle extract (Cori and
Laner, 1951).
2.2.2. Plants:
Studies on the occurrence, properties and purification of amylases
from higher plants are relatively few. Novellie (1982a) reported that
Sorghum sp. has the ability to produce amylases on germination.
a-Amylase of sorghum malt has been purified and characterized
(Botes et al. ,1976 ; Mundy, 1982 and Okon and Uwaifo, 1984). The
ungerminated barley grain contains a- atnylase whereas ungerminated
sorghum grain lacks it (Haas, 1976).
Amylases of barely and wheat are found in the aleurone layer of the
grain, while those of sorghum are formed in the germ and spread outward
through the endosperm (Daiber and Novellie, 1968). The a-amylase of

sorghum malt is acid resistant and active over a wide range of pH of 4-7
(Botes et al. , 1967). Budair( 1977) and Ahmed (1988) studied amylases of
Sudanese sorghum malt and showed that malt of feterita sorghum gave the
highest amylolytic activity when compared to malt of other sorghum types
The richest source of P-amylase, apart from sweet potato, are the
cereals, especially wheat and barely. Hams (1968) reported that P-amylases
together with other endogenous enzymes of barely malt, catalyzed the
conversion of starch to maltose which constitutes 45% of the total
carbohydrate content. The p-amylases of wheat and barely have been
isolated and characterized by several workers (Danielson and Sandegren,
1947; Danielson, 1948; Tipples and Tkachuk, 1965 and Tkachuk and
Tipples, 1966).At that time it was commonly accepted that sorghum malt
contained little or no p-amylase activity . However, contrary to this view,
Novellie (1960) reported the isolation of p-amylases from kafiircorn
(Sorghum caffrorum).
Sorghum p-amylases have been isolated ,purified and characterized
by several workers (Botes et al. 1967; Mundy, 1982 and Okon and Uwaifa,
1984). Sweet potato and Soya been P-amylases have been isolated and
obtained in a crystalline form by Balls et al. (1948) and Fukumoto and
Tsujsaka(1956).
2.2.3. Fungi:
Different types of amylases found in fungi were reviewed by
MacAllister (1979), Reichelt (1983) and Gupta and Gautan (1995).
a-Amylases have been isolated from Aspergillus foestidus,
Aspergilhis oryzae, and Aspergilhis mger (Michelena and Castillo, 1984;
Matsuura et al, 1984 and Boel et al., 1990 in order). Amylolytic activity of

different Rhizopus isolates has been determined by several authors (Lim et
al., 1987; Hang, 1989 and Roch- Chili and Hang, 1990).
Fungal P-amylases were isolated from Aspergillns niger and
Aspergillus oryzae (Svensson et al. , 1988).
Amyloglucosidases have, also, been isolated and purified from
Aspergillus niger (Lineback et al. , 1969 and Lineback and Aira, 1972),
Penicillium oxalicum (Yamasaki et al. , 1977a), Aspergillus oryzae (Mitsue
et al., 1979 and Kita et al., 1982) and, Aspergillus awamori (Bhumibhamon,
1983). Two forms or isoenzymes of glucoamylase have been isolated and
purified from Aspergillus niger (Flemingi and Stone, 1965; Pazur et al., 1971
and Lineback and Aira, 1972) and three isoenzymes of glucoamylase have
been isolated and separated from Aspergillus oryzae (Mitsue et al. , 1979)
and from Rhizopus sp. (Takahashi et al. , 1978 and Kanlayakrit et al.,
1987).
Glucoamylases of yeast and saacharomyces spp. have been reported
by Steverson et al. (1984), Pestana and Casstillo (1985) and Tani et al.
(1986).
2.2.4. Bacteria:
Many bacteria are known to produce extracelluar amylases.
a-Amylases from Bacillus sp. have found a considerable commercial
application for several decades ( Ingle and Boyer, 1976). Production of
amylases by Bacillus polymyxa have been reported by Robyt and French
(1964). Bacillus lichen (for mis is known as the best producer of thermostable

a-Amylase (Satio, 1973). Three theromstable and acidophilic a- amylases of
Bacillus acidocaldarius have been reported by Buonocore et al. (1976),
Ingle and Boyer (1976) and Uchino (1982). Another thermostable a-
amylase from Bacillus acidocaldarius strain A-2 was reported by Kanno
(1986). Yet another heat-stable a-amylase was isolated from Bacillus
subtilis by Yamane and Maruo (1974).
a-Amylases have been characterized and purified from Bacillus
stearothermophilus (Stark and Tetraut, 1951; Hartman et al. ,1955 and
Campbell and Manning, 1961), Bacillus coagulans (Campbell, 1954),
Bacillus subtilis (Toda and Narita, 1968), Bacillus lichewformis (Madsen et
al. , 1973 and Chiang et al. ,1979), Bacillus circulans (Takasaki, 1982 and
Taniguchi et al., 1982), Bacillus amyloliquefaciens (Norman, 1979;
Fogarty,1983 and Fogarty and Kelly, 1980 and 1990), Bacillus megaterium,
Bacillus cereus and Bacillus polymyxa (Hayashi et al. , 1988) and Bacillus
sp.E2(Goya\e(al., 1995).
Many bacteria other than the genus Bacillus, have also been reported
as a-amylase producers such as Pseudomonas stutzeri (Robyt and
Ackerman, 1971), Thermonospora sp. (Glymph and stutzenberger, 1977
and Upton and Fogarty, 1977) and Thermoactinomyces sp. (Kuo and
Hartman, 1966, Mai et al. , 1992). Kainuma et al. , (1975) isolated an
amylase that produces maltohexose from starch from Aerobacior aerogens.
High molecular weight a-amylase secreted by Lactobacillus amylovorus
was reported by Imam el. al. , (1991). Thennoactinomcyes thatpophilus F13
produce an amylase which digests raw starch (Okolo et al. , 1996). Bergman
et al. (1988) and Hayashida et al. (1988) reported that few microbial species
produce amylase that digest raw starch.
8

Bacterial p-amylases have been found in several genera such as
Pseudomonas, Streptomyces and Bacillus. Kneen and Beckord (1946) found
that the amylase of Bacillus polymyxa was of the P-type and described it as
being similar to the system in barley malt extract. Robyt and French (1964)
found that the starch-degrading enzyme system of Bacillus polymyxa was
capable of a 94% conversion of amylopectin into maltose.
P-Amylases have, also been found in various species such as Bacillus
cereus (Shinke et al. , 1975), Bacillus circulans (Kwan et al. , 1993),
Bacillus polymyxa (Murao et al. , 1979) and Bacillus megaferium (Rani et
al. , 1994). Alkaline p-amylase was isolated from Bacillus subtilis (Boyer
and Ingle, 1977), while acidic P-amylase had been obtained from
Streptomyces sp. (Koaze el al. ,1975). Bacillus cereus produced a
thermostable P-amylase that hydrolysed starch to 90.5% maltose, 7.5%
maltoseriose and 2% other oligosaccharides (Takasaki and Takahara, 1976).
Amyloglucosidases were produced by many species of bacilli
(Srivastava and Baruah, 1986). Ilori et al. (1995) characterized and purified
an amlyase that produces glucose from starch from Lactobacillus brevis.
2.3. The Genus Bacillus:
Bacillus sp. are gram positive, aerobic or facultatively anaerobic
bacteria characterized by having straight or nearly straight rod shaped cell
and possessing endospores which are very resistant to many adverse
conditions. Cells of Bacillus sp. may occur singly or in chain and may
contain parasporal bodies or protein crystals. This genus exhibits a wide
diversity of physiological chacteristies. Some are mesophilic while others

are thermophilic and some are acidiophilic while others are alkaliphilic.
Some strains are salt tolerant, while others have specific requirement for
salts . Most Bacillus spp. are characterized by being catalase positive (Claus
and Berkeley, 1986).
Gordon et al. (1973) classified bacilli into three groups on the basis
of the shape of endospore and sporangium. Group one has ellipsoidal spore
with unswollen sporangium, group two has ellipsoidal spore with swollen
sporangium while group three is charactized by spherical spore with swollen
sporangium. Bacilli produce different types of spores. These are,
cyclindrical, oval, kindney or banana-shaped. The location of the spore may
be central, paracentral, subterminal, terminal or lateral in the sporangium.
Bacillus colonies may be white or creamy coloured on nutrient agar
medium and the colony surface may be smooth or rough (Claus and
Berkeley, 1986).
2.4. Optimization of Amvlasc Synthesis:
Production of amylases has been investigated in microorganisms by
many authors Burbidge and Collier(1968) and Nyiri (1971 )reported that
higher yields of amylase were obtained on media composed of complex raw
materials containing starch rather than glucose. High production of Bacillus
siiblilis oc-amylase is accomplished by using high concentration of starch
(8-12%) (Nomura et al. , 1956). Khire and Pant (1992) reported that the
maximum production of amylase by Bacillus sp.64 was achieved using 0.5%
starch or dextrin as carbon source. Some strains of Bacillus sp. have been
reported to produce a-amylase when grown on media containing glucose
10

and other monosaccharides as the sole source of carbon. (Fukumoto et al.,
1957; Coleman and Elliot, 1962; Welker and Campbell, 1963 and Meers,
1972). Bacillus stearothermophilus produced oc-amylase after addition of
the inducer (maltose, methyle - p-D-maltoside, or phenyl a-D- glucoside at
10"3M) to the washed cell suspensions(Welker and Campbell, 1963). The
same authors reported that addition of casein hydrolysate replenished the
amino acid pools and restored the ability of the cell to produce a-amylase.
Also, addition of cylic adenosine 3',5 monophophate stimulated a-amylase
accumulation in growing cultures of the parent strains of Bacillus
licheniformis (Satio and Yamamoto, 1975). The p-amylases of Bacillus
circulans was produced when the bacterium was grown on a medium
containing 4% soluble starch (Kwan et al. , 1993).
Culture conditions, also, play a very important role on growth and
production of extracelluar glucoamylase in fungi (Windish and Mhatre,
1965; DeMot et al., 1984 and Ali el al. ,1989) and in bacteri (Burbidge and
Collier, 1968). Moreover, aminoacids and nucleotides (Futumoto et al. ,
1957 and Srivastava and Baruah, 1986), metal ions (Windish and Mhatra,
1965) and carbon and nitrogen (Manjunath et al 1983) also affect enzyme
production. Addition of a-methylglucoside doubled the glucoamylase
production in Aspergillus sp. K-27 (Ju-Nichi et al. , 1985). Also,
cyclodextrin had a stimulatory effect on amylase synthesis in yeast (Oten-
gyongefa/., 1980).
The pH value of the medium and the growth temperature were found to
affect the production of amylolytic enzymes. Bacillus sp. produce amylase
under different temperatures and pH values. Boyer and Ingle (1972) reported
that Bacillus spp. NRRLB-3881 produced (3-amylase maximally at 50°C and
pH 9, while Morgan and Priest, (1981) reported that the maximum
11

production of a-amylases from 10 strains of Bacillus lichenifonnis were at
70-90°C and pH 7. Amylases from Bacillus subtil is, Bacillus licheniformis
and Bacillus cereus were produced maximally at temperatures 30, 37 and
80°C and pH values of 7.0, 5.5 and 7.5 in order (Olukayode el al. ,1987).
Maximum production of glucoamylase of Aspergillus awamori was
found to be at 30°C and pH 4.5 (Pestana and Castillo, 1985), Flemingi and
Stone(1965) and Lineback and Aira (1972) reported that glucoamylase of
Aspergillus niger was produced maximally at 37°C and pH 4.3-4.5.
2.5. Purification:
Several procedures have been reported for the purification and
crystallization of amylases. a-Amylases from malt (Schwimmer and Balls,
1949), human pancrease, human saliva, and Bacillus subtil is (Myrback and
Neumiiller, 1951), Aspergillus sp. (Uuderkofler and Roy, 1951) and
Bacillus slearothermophilus (Manning and Campbell, 1961),.have been
purified and obtained in a crystalline forms.
a-Amylases from different species and strains of Bacillus have been purified
to different degrees of homogeneity using various types of ion exchange
chromatography. Morgan and Priest (1981) purified a-amylase from
Bacillus Ucheniformis-NCIB 634, thirty fold over that of the culture filterate
using DEAE-cellulose while Yoshiyuki (1982) purified a-amylases from
Bacillus circulans G6* by means of ammonium sulphate fractionation,
DEAE-Sepharose and DEAE-Sephadex G200 columns and gel electro-
phoresis. The a-amylase from Bacillus acidocaldarius and Bacillus sp.-JF
strain have been purified and characterized using ammonium-sulphate
12

fiactionation, SP-Sephadex 50, Ultra filtration and gel electrorophroesis
(Kanno, 1986 and Jin el al. , 1992).
As regards the P-amylases from sweet potato Balls et al.( 1948) and wheat
Meyer et al. (1951) have reported their purification to a crystalline form.
Also, two forms of P-amylase have been purified from Bacillus polymyxa
using DEAE- cellulose, CM- cellulose, and Biogel P-150 (Murao el al,
1979). From Bacillus megateium No 32, another P-amylases was purified
over 2600 fold using ammonium sulphate, SE-Sephadex-G50 and gel
filteration with Sephadex G100-(Higashihara and Okada, 1974).
Two forms of amyloglucosidase, have been purified from Aspergillus
niger using Sephadex G-25 gel filteration, DEAE Sephadex A-5 column
chromatography, Sephadex G100 gel filteration, Sephadex G200 gel
filteration and gel electrophoresis (Pazur et al. ,1971, Bhumibhanon, 1986).
2.6.Hvdrolvtic Products of Amylases:
The reaction products of amylases vary with species and strain used
(French and Knapp 1950; Robyt and French 1963 and 1964 and Robyt and
Whelan 1968).
The three types of amylases (a-amylases, P-amylases and glucoamylase)
hydrolyse amylose and amylopectin resulting in different products. Guilbot
and Merecier (1985) reported that a-amylases randomly cleave starch or
glycogen to give polysaccharides of various lengthes and, in some cases,
maltose and glucose as a final product. a-Amylases from Bacillus
licheniformis yield maltopentose as a major product (Morgan and Priest,
1981), while alkaline a-amylases from other strain of Bacillus licheniformis
13

hydrolyse starch to glucose, maltose, maltotriose and maltotetrose rather
than higher oligosacchorides (Medda and Chandra, 1980). On the other hand,
a-amylase from Bacillus amyloliqiiefaciens and Bacillus caldovelox yield
maltohexose as a major product (Norman, 1981 and Bealin-Kelly el al. ,
1990).
However, glucoamylase always gives glucose as the only and final
product (Mac Allister, 1979 and Reichelt, 1983).
2.1. Stability and Denaturation:
Amylases are generally influenced by the presence of anions such as
chloride which activate the enzyme (Muss et al., 1956). Khire and Pant
(1992) reported that divalent cations such as Mg** and Ca++stablized the
activity of amylase of Bacillus sp. 64.
Purified a-amylase obtained from Bacillus coagulans was still active
after 12hr at 90°C (Stark and Tetraut, 1951) and Hartman et al., 1955).
Boyer and Ingle (1972) reported that the alkaline amylase of Bacillus sp.
NRRLB-3881 and the a-amylase of Bacillus amyloliquefaciem were stable
at pH 9.2 for 1 hr at 50°C. Murao et al. , (1979) reported that p-amylases
from Bacillus polymyxa No. 72 were most active at pH 7.5 and at 45°C and
stable between pH 4 and 9 for 15 hr at 37°C.
Amyloglucosidase of Aspergillus niger and Aspergillus oryzae are
stabilized by calcium ions against heat and alkali denaturation (Wiseman,
1985). Pestana and Castillo (1985) showed that glucoamylase of Aspergillus
awamori was stable at temperature up to 45°C and it's activity at 50°C was
protected by the presence of raw starch. A similar protective effect by
14

substrate lias been reported for the glucoamylases of Aspergillus niger
(Solomon and Levin 1974) and Humicola lanuginose (Taylor el al. , 1978).
a-Amylase activity of Bacillus sfearothermophilus is inhibited by
many factors such as chloroamphenicol, 8-azaguanine, actionoinycine C and
D and some amino-acids analogues, such as norvaline, norleucine and
ethoine (Welker and Campbell, 1963). Nivva el al (1970) found that
nojirimycine (5-amino-5 deoxy glucopyranose) has an inhibitory effect on
some ainylases of various microorganisms. Also, Jin el al. (1992) reported
that some metal ions such as Fe++, Cu4^, Zn++ and Ag*4" have an inhibitory
effect on a-amylase activity of Bacillus spp J.F strain.
p-Amylase activity of Bacillus megalerium strain No 32 was inhibited
by p-chloromecuribenzoate (PCM13C) (Iligashihara and Okada, 1974).
Also, Kwan el al (1993) reported that some carbohydrates such as glycogen,
dextran, inulin, raffinose, melibiose, trehalose, maltose, glucose, sorbitol and
mannitol inhibited f3-amylase activity of Bacillus circulans. Murao el al.
(1977) reported that an amylase inhibitor-producing microorganism was
identified as a subspecies of sfreplomyces diaslaiicas sub sp anilyfoslalicus
No.2476 (S-AI) has an inhibitory activity on a-amylase and glucoamylase
but not on [3-amylase.
Yamasaki and Kanno (1991) showed that 5mM Hg+reduced gluco-
amylase activity of Mucov javanicus to 30% of it's original activity.
2.8. Amylasc Kinetics:
Allen and Thoina (1978) reported that a-amylase of Aspergillus
oryzae, which can hydrolyse maltotriose, does not follow Michaelis-Menten
15

kinetics on this substrate and thus comparison by such means as Km values
may, for specific amylases be very misleading. Me concluded that Michaelis-
Menten constant is not always as reliable as is expected.
The affinity for starch of the Klehsiella pnewnoniae exo-amylase is
twice that of Bacillus caldovelox a-amylase and the affinity of these
enzymes for the smallest substrate that each can hydrolyse (maltotetraose for
Klehsiella pneumoniae and maltoheptaose for Bacillus caldovelox ) showed
that Bacillus caldovelox enzyme has twice the affinity of the other (Monma
et at. ,1983 and Fogarty, et at. ,1991). Michelena and Castillo (1984),
estimated the Km values of the a-amylase of Aspergillus foesiidus for
amylopectin, soluble starch and amylose. He found that the affinity for
amylopectin was two fold lower than that for soluble starch and 7 fold lower
than for amylose. Kanno (1986), studied the properties of a-amylase of
Bacillus acidocafdrius and estimated the Km value of this enzyme, which
was 1.6 mg starch/ml. Fogarty et al.(!99l) studied the kinetics of substrate
hydrolysis of the a-amylase of Bacillus caldovelox and found that the
affinity of the a-amylase of this microorganism for starch is considerably
lower than the affinity of other maltohexose producing amylases, towards
the same substrate. Okon and Uwaifo (1984) reported that a km value for
the P-amylase of Sorghum hi color was 1.272 mg starch/L.
llori el at. (1995) estimated the Km value of glucoamylase of
iMclohahillus hrevis and found that it's affinity for soluble starch is higher
than that of the enzyme from Aspergillus oryzae (Razzaque and Ueda,
1978).
16

2.9. Application of Amylascs in Industry:
There is a large variety of commercially available enzymes differing
in biological source, activity, physical form and characteristics such as pH
and temperature optima.
It is advantageous to use thermostable enzymes, as by carrying out the
reaction at higher temperature, faster reaction rate can be achieved. Also,
increased reactant solubilities and decreased viscosities and microbial
contamination can be obtained.
Organisms suitable for use in the production of enzymes should be
characterized by easy and rapid growth in large fennenters on comparatively
cheap and simple nutrients without the need for inducers. A high yield of
enzyme should be obtained in a form that is easy to isolate, purify and
concentrate without the formation of toxic or immunogenic metabolites. The
organisms should have stable physiological characteristics and be readily
acceptable to the food and drug authorities (Barfoed, 1981).
Most commercially available amylases are produced by many
microorganisms especially, Aspergilhis oryzae. Bacillus amyloliqiiefaciens
and Bacillus lichenifonnis (Priest, 1984; Linardi and Machado, 1990 and
Bezbaruch el al , 1991).
Amylases have been used in a variety of industrial processes, which
require efficient saccharification of raw starch (Fogarty, 1983). This
enzymatic hydrolysis of starch is important from the point of view of energy
saving and effective utilization when compared to acid hydrolysis.
Amylases are, also, used in food processing, Pharmaceuticals, starch derived
adhesives, grain alcohol, production of moist cakes and fruit cakes. They are
17

also used in animal feeds, sewage treatment, detergents and confectionery
(Koniaki, 1956).
In addition, bacterial a-amyiases are used in the textile industry
because they produce adhesive starch which is important in fabric sizing and
designing. This process is accelerated at elevated temperature, so Bacillus
iichenifonius oc-amylases is considered the best one because it is active at
temperature 105-110°C and the reaction time is 1-2 minutes (Wiseman,
1985). Bacterial and fungal a-amylases are, also, used in paper manufacture.
It is known that liquefying starch is used as a coating binder and this
liquefying process is achieved by a-amylase (Clayton el ai ,1984).
a-Amylases, P amylases and amyloglucosidase are used in ethanol
fermentation from grains. The process is started by the addition of a-
amylases during the cooking stage to reduce viscosity, followed by addition
of saccharogenic a-amylase or f3-amylase and finally addition of amylo-
glucosidase and yeast to complete fermentation of starch to ethanol
(Godfrey and Reichelt, 1983). ot-and P Amylases are also used in baking.
P Amylases are mainly used in the production of maltose syrup. Two
principal types of maltose syrup are produced. One contained 30-50%
maltose, 6-10% glucose and 42-49% dextrose equivalent which is used in
jams and confectionery, because it is resistant to colour formation and
crystallization. The second contains 30-40% maltose, 3-5% glucose and 63-
70% dextrose equivalent, which is used in bread-making and brewing of
beer due to it's high content of ferementable sugars (Maeda and Tsao, 1979).
The industrial use of amyloglucosidase is mainly for the production of
glucose. The glucose is either used as a syrup or crystallized to give pure
solid glucose (MacAllister, 1979 and Reichelt, 1983).
18

3.1. Materials:
3.1.1. Microorganisms: Bacteria and fungi were isolated from various
Sudanese soils and different food materials.
3.1.2. Chemicals: All general chemicals were of the Analar grade or
equivalent. 3,5 dinitrosalicylic acid (DNS) and DEAE Sephadex were from
Sigma Chemical Company (UK). All media from Oxoid Chemical Company
(U.K) and were prepared according to Harrigan and McCance (1966) and
Peter eta! (1986).
3.2. Screening of Microorganisms:
3.2.1. Screening of Bacterial Isolates:
Soil samples were obtained from different areas namely, Atbara, Khartoum
and Kassala cities. Food materials were obtained from Khartoum local
market and various homes.
Ten grams of soil and food samples were suspended in saline phosphate
bulYer pH 7.0. The suspension were pasteurized at 80°C for 10 minutes to
kill the vegetative forms of the bacteria. One ml. of this suspension was
then added to 20 ml. of melted nutrient agar medium in a Petri-plate. This
medium was prepared by dissolving 1 g. of meat extract, 2g. yeast extract,
5g. peptone, 5g. sodium chloride and 15g. agar in 1 liter distilled water.
After adjusting the pH to 7.5 using either dilute HC1 or NaOH the medium
was then sterilized by autoclaving at 121 °C and 15 lb/square inch for 15
19

minutes. The Petri-plates were incubated at 37°C in a Gallenkamp (U.K.)
cooled incubator. The growth of the bacteria was observed periodically and
colonies were then picked off and subcultured on a fresh nutrient agar
medium to obtain a pure culture . The bacteria were examined using a light
microscope.
3.2.2. Isolation of Fungi:
Healthy grains of different cereals were surface sterilized with 0.01%
mercuric chloride solution for 3 minutes. They were then washed several
times with sterile distilled water in sterilized Petri-plates. The sterilized
grains were sown on three layers of sterile moistened filter paper in glass
Petri-plates. The plates were incubated at 30°C for 7 days. The grains were
then examined for seed-bom fungi using a stereoscopic binocular
microscope.
For culturing these seed-bom fungi malt extract medium was prepared by
dissolving 5g. mycological peptone, and 15g. agar in 1 liter distilled water.
This medium was sterilized by autoclaving at 121°C and 15 Ib/square inch
for 15 minutes. After sterilization the medium was poured aspectically in
Petri-plates and left to solidify.A single spore of fungi was picked off and
cultured on this malt extract media. The plates were incubated at 30°C for 7
days.
For isolation of yeast lg. of food material was suspended in 10 ml sterile
distilled water and streaked on malt extract agar. The plates were also
incubated at 30°C but for 48 hours. The resulting fungi were examined using
a light microscope.
20

3.2.3. Selection of Isolates:
The bacterial isolates were grown on nutrient agar and the fungal isolates on
malt extract agar media. Both media contained 0.2% starch as the major
carbon and energy source. The bacterial plates were incubated at 37°C for 2
days, while the fungal plates incubated at 30°C for 2-7 days.
Amylase activity was detected by the formation of a clear zone around the
colonies when flooded with Gram's iodine solution(l% iodine in 2%
potassium iodide). The index of amylolytic activity was determined
according to the method of Hankin and Anagnostakis (1975) using the
following formula:
Index of amylolytic activity =
The diameter of the clear zone - The diameter of the culture zone
The diameter of the culture zone
Based on this index of amylolytic activity ten bacteria were chosen for
further studies. As judged by microscopic study they were all bacilli.
For routine keeping of these isolates, nutrient agar medium was prepared (as
in expt. 3.2.1). This medium was distributed in test tubes which were
sterilized by autoclaving at 121°C and 15 Ib/square inch for 15 minutes.
After cooling in a slanted way the tubes were inoculated aspetically with the
bacillus isolates in a Laminair cabinet; each isolate being inoculated in
several tubes. These were kept in a refrigerator and were recultured eveiy
month. These stocks were then used in all experiments.
3.2.4. Morouhological and Biochemical Characterization of Bacillus sp.
The ten Bacillus isolates were characterized using Bergy's Manual of
Systematic Bacteriology (Peter et a! 1986).
21

3.2.4.1. Morphological Tests:
The bacillus cultures were grown on nutrient broth medium containing lg.
meat extract, 2g. yeast extract, 5g. sodium chloride in 1 liter distilled water.
The medium was distributed into 250 ml. conical flasks (100 ml in each) and
autoclaved at 121°C for 15 minutes. The flasks were inoculated with a
loopfiil of Bacillus isolates and incubated in an incubator shaker
(Gallenkamp) at 37°C. General morphological characteristics of the
vegetative cells and sporangia were determined as described by Claus and
Berkeley (1986).
3.2.4.2. Staining of the Spore:
Staining of the spore was carried out according to the method of
Bartholomew and Mittwer's (1950). A thin smear of Bacillus isolate was
taken from the broth culture, spread over a slide and fixed with heat using a
Bunzen flame. The slide was then flooded with Malachite green (0.5%
solution) and heated gently over a Bunzen flame until steam appeared. This
process is continued for 5 minutes, until the stain is nearly dry. The excess
stain was removed by rinsing the slide with tap water and the smear was
flooded with the counter stain safranin solution (0.25%) for 30 seconds.
Again the slide was washed with tap water and dried either by blotting with
a filter paper or in the air. The colour of the spore was observed using a light
microscope.
3.2.4.3.Biochemical Tests:
3.2.4.3.1 Production of Catalase:
Bacillus cultures (from the stock) were grown on nutrient agar slants (as in
expt. 3.2.1) for one or two days. They were then flooded with 0.5 ml of 10%
22

hydrogen peroxide. Production of air bubbles indicated a positive test as
described by Whittenburry (1964).
3.2.4.3.2. Voges Proskauer Test:
Voges Proskauer test was carried out according to the method of Levin
(1916 b). Voges Proskauer broth was prepared as follows:
lOg. tryptone, 5g. disodium hydrogen phosphate, lg. potassium dihydrogen
phosphate, 2g. sodium chloride, O.lg magnesium sulphate 7H2O and 2g.
glucose were dissolved in 1 liter distilled water. After adjusting its pH to
6.5, the medium was distributed in 5 ml. portions in test tubes and
autoclaved at 121°C for 20 minutes. After cooling the test tubes were
inoculated with Bacillus isolates in triplicates (as in expt. 3.2.4.1) and
incubated at 37°C for 3, 5 and 7 days. Three ml of 40% (w/v) sodium
hydroxide were mixed with the culture, followed by addition of 1 mg
creatine. Production of a red colour after 30-60 minutes at room temperature
is an indication of formation of acetyl methyl carbinol.
3.2.4.3.3. Production of Acid from Carbohydrates:
Bacillus cultures were inoculated on acid production medium, containing 1 g.
of diammonium hydrogen phosphate, 0.2g yeast extract, 0.2g potassium
chloride, 0.2g. magnesium sulphate.7H2O and 15g agar in 1 litre distilled
water . The pH of the medium was adjusted to 7.0 followed by the addition
of few drops of 0.04%(w/v) solution of bromocresol purple. The medium
was distributed into 3 sets of test tubes and sterilized by autoclaving at
121 °C for 20 minutes. Sterilized D-glucose, L-arabinose and D-mannitol
each was added aseptically to each tube to make a final concentration of
10% (w/v) and the tubes were allowed to cool in a slanted position.
23

The slants were inoculated with the Bacillus isolates and incubated at 37°C
for 7 days. Production of acid and gas was checked by the change of
bromocresol purple from purple to yellow colour and appearance of gas
bubbles, respectively.
3.2.4.3.4.Hydrolysis of Starch:
Duplicate plates of starch agar were inoculated with the Bacillus culture.
The medium was prepared from nutrient agar supplemented with 0.2%
soluble starch, autoclaved and distributed aspectically into Petri-plates. After
incubation at 37°C for 3 to 5 days the plates were flooded with 95% ethanol.
Hydrolysis of starch was detected by the formation of a clear zone
underneath (after the growth is scraped off) and around the growth.
3.2.4.3.5. Liquefaction of Gelatine:
Nutrient agar supplemented with 0.4% gelatine was prepared, autoclaved at
121 °C for 15 minutes and distributed aspetically into Petri-plates. The
plates were then inocubated with the Bacillus cultures and incubated at 37°C
for 3 to 5 days. They were men flooded with 1N sulphuric acid saturated
with sodium sulphate. Hydrolysis of gelatine is indicated by formation of a
clear zone under and around the growth as described by Cruickshank
(1960).
3.2.4.3.6. Reduction of Nitrate to Nitrite:
Nitrate broth containing 3g. beef extract, lg. potassium nitrate, 5g. peptone
in 1 litre distilled water was prepared. After the pH was adjusted to 7.0, the
medium was distributed into test tubes containing Durham's tubes and
autoclaved at 121°C for 20 minutes .After cooling, the tubes were inoculated
24

with the Bacillus isolates and incubated at 37°C for 3-7 days. The cultures
were then tested with filter paper strips dipped in potassium iodide, followed
by addition of a few drops of IN hydrochloric acid. Production of a purple
colour in the filter paper indicated the presence of nitrite and accumulation
of nitrogen gas in the Durham's tubes was observed, according to Peter ei al
(1986).
3.2.4.3.7. Deamination of Phcnvlalanine;
Agar slants of phenylalanine was prepared by dissolving 3g. yeast extract,
2g. DL-phenylalanine and 1 g. agar in 1 litre distilled water. After adjusting
the pH of the medium to 7.3, it was distributed in test tubes, autoclaved at
121 °C for 20 minutes and then cooled in a slanted position. The slants were
then inoculated with the Bacillus cultures and incubated at 37°C for 7 days.
The deamination of phenylalanine was tested by the addition of 5 drops of
10%(w/v) ferric chloride solution over the growth zone. Production of green
colour beneath the colony indicates the formation of phenylpyruvic acid.
Report, (1958).
3.2.4.3.8. Production of Indolc:
Indole production medium was prepared by dissolving lg commercial
tryptone broth in 100 ml distilled water. (This broth was prepared by
dissolving 1 Og. trypton and 5g. sodium chloride in 1 Liter distilled water).
The medium was adjusted to pH 7.5 and then distributed in 5 ml portion in
test tubes and autoclaved at 121°C for 15 minutes. The tubes were
inoculated with Bacillus cultures and incubated at 37°C for 14 days. The
cultures were then tested by the addition of 2 ml. of a test solution which
contained 5g p-dimethyl ammo benzaldehyde, 75 ml isoamyl alcohol and 25
25

ml. hydrochloric acid. Production of indole is indicated by the formation of a
pink to red colour in the alcoholic layer, as described by APHA (1955).
3.2.4.3.9 Production of Dihydroxy Acetone:
One hundred ml. of nutrient agar medium containing lg. yeast extract, and 2
ml. glycerol was sterilized by autoclaving at 121 °C for 20 minutes and
poured aspetically into Petri-plates. After solidifying the medium was
streaked once across the plate with Bacillus culture and incubated at 37°C
for 10 days. The plates were then flooded with a mixture of two solutions in
a ratio of 1:1. The first solution contained 34.66g hydrous copper sulphate.
5H2O in 500 ml. distilled water and the second solution contained 173g.
potassium sodium tartarate and 50g. sodium hydroxide in 500 ml distilled
water. Fonnation of a red halo around the growth indicates a positive test.
3.2.4.3.10. Utilization of Citrate and Propionate:
Citrate and propionate media containing either 1 g. trisdium citrate or 2g.
sodium propionate, 1.2g magnesium sulphate.7H2O, 0.5g diammonium
hydrogen phosphate, lg. potassium chloride, 15g agar, 2 ml of 0.04%
phenyl red and 40 ml trace elements solution in 920 ml distilled water were
prepared. (The trace elements solution contained 500 mg ethylene diamine
tetra acetic acid, 200 mg FeSO4 , 10 mg ZnSO4.7H2O, 3 mg MNC12.4H2O,
30 mg H3BO3, 20 mg CoCl2.6H2O, 1 mg CuCl2.2H2O, 2 mg NiCl2.6H2O
and 3 mg Na2MoO4.2H2O in 1 liter distilled water). After adjusting the pH
of the medium to 6.8, it was distributed into test tubes and autoclaved at
121°C for 20 minutes. After cooling, the slants were inoculated with
Bacillus isolates and incubated at 37°C for 14 days. Production of a red
26

colour indicates utilization of the organic acid as described by Crawford
(1962).
3.2.4.3.11. Growth in Sodium Chioridc:
Bacillus cultures were inoculated in nutrient broth medium and grown for
one day at 37°C. Three ml of these broth culture were then inoculated in
fresh sterile nutrient broth media containing 0, 5, 7 or 10% sodium chloride
in test tubes and grown for 7-14 days at 37°C. The growth of the bacterial
was followed during this period.
3.2.4.3.12. Growth at pH 5-7:
Sabouraud dextrose agar slant was prepared by the addition of lOg.
neopeptone and 40g. dextrose to 1 liter distilled water. The pH of the
medium was adjusted to different values (5, 6 and 7) using either dilute HCL
or NaOH. The medium was then distributed into three sets of test tubes and
autoclaved at 37°C for 20 minutes. After cooling the slants were inoculated
with Bacillus isolates. At the same time, tubes of sterilized nutrient agar
were inoculated with the same isolates as controls. The tubes were all
incubated at 37°C for two weeks. The growth of the culture was observed
periodically.
3.2.4.3.13.Determination of Optimum Growth Temperature :
Slants of nutrient agar were prepared as described previously (expt. 3.2.3),
these were inoculated with Bacillus cultures and incubated at different
temperatures namely, 5°C, 10°C, 30°C, 40°C, 50°C, 55°C, 60°C and 65°C.
The growth of the culture were observed after 3 days for high temperatures
and after 7-14 days for low temperatures.
3.2.4.3.14. Hydrolysis of Casein:
Milk agar medium was prepared as follows:
27

5g. milk powder were dissolved in 50 ml distilled water and lg. agar in
another 50 ml distilled water. The two media were autoclaved separately at
121°C for 20 minutes, mixed aspetically and then poured into the Petri-
plates. After cooling the media were streaked across once with the inocula
from the stock. The Petri-plates were incubated at 37°C for 7-14 days and
examined. A clear zone around and below the growth was taken as a
positive result as described by Davis (1959).
3.2.4.3.15. Anaerobic Growth:
Anaerobic medium was prepared as follows:
20g. thioglycolate, 2g. sodium formaldehyde-sulfoxylate, 15g. agar, 20g.
trypticase, lOg. glucose and 15g. sodium chloride were dissolved in 1 liter
distilled water. The medium was adjusted topH 7.2, distributed into test
tubes which were autoclaved at 121°C for 20 minutes. After cooling the
anaerobic agar was inoculated with a loopful of nutrient broth culture by
stabbing into the bottom of the culture tube. The tubes were incubated at
45°C for 3-7 days and the growth was observed.
3.2.4.3.16. Egg Yolk Reaction:
Egg yolk broth was prepared by adding lOg. tryptone 5g. disodium sulphate.
7H2O, 2g. glucose and lg. potassium dihydrogen phosphate to 1 liter
distilled water. The pH of the medium was adjusted to 7.6 and the
autoclaved at 121°C for 20 minutes. 1.5 ml of sterilized commercial milk
was added aspetically to 100 ml. of the basal medium. The medium was
allowed to stand in a refrigerator overnight and the supernatant broth was
distributed in sterile tubes in 2.5 ml amounts. The tube were inoculated with
Bacillus cultures and incubated at 37°C for 1, 3, 5 and 7 days. The
28

appearance of a heavy white precipitation or on the surface of the egg yolk
medium indicates a positive result, Chu (1949) and Knight and Proom
(1950).
3.2.5. Amyiasc Production:
3.2.5.1. Effect of Temperature:
A nutrient broth medium containing (g/1) peptone 10; dipotassium hydrogen
phosphate 3; magnesium sulphate. 7H2O 1 and starch 5 in 1 liter distilled
water was prepared. The pH of the medium was adjusted to 7.5 and
distributed in 500 ml Erlenmyer flasks; 100 ml to each. The media were
autoclaved at 121°C for 20 minutes . After cooling the flasks were
inoculated with a loopful of the chosen isolates (from a 48 hours old culture
grown on a nutrient agar slant). The cultures were allowed to grow on a
rotary shaker (200 rev/min) at 37°C for 24 hours. Ten ml of these vegetative
inocula were then inoculated in 100 ml of fresh medium also on a rotaiy
shaker (200 rev/min)at different temperatures namely 25°C, 30°C, 35°C
,40°C, 45°C 50°C and 55°C. Twenty ml samples were collected for each
temperature at different time intervals namely 18, 24 and 48 hours. The
samples were centrifuged to remove the bacterial cells, using a Sigma
Laboratory refrigerated centrifuge Gmbh (Germany model 2K15withNr
12139 rotor).The supernatants were collected for assaying amylase activity
(see expt. 3.2.6.3). Based on the result of this experiment further work was
performed on six isolates only (see result section expt.4.3.2.).
3.2.5.2. Effect of pH;
Nutrient broth media as described above (expt,3.2.5.1.) was prepared and
the pH was adjusted to different values namely, 5, 6, 7, 8, 9 and 10 using
29

using dilute HC1 or NaOH solutions. Each isolate was inoculated into a set
of these media and grown at 50°C for 24 hours. 20 ml of each sample were
collected and treated as above (expt. 3.2.5.1). The amylolytic activity was
determined as described in expt. 3.2.6.3.
3.2.5.3. Effect of Substrate Concentration:
The Bacillus isolates were grown on nutrient broth medium at pH 9.0 for all
isolates except Bacillus subtilis SUDKj which was grown on nutrient broth
medium at pH 7.0. Fresh nutrient broth media containing different soluble
starch quantities to give a final concentration of 0.1%, 0.2%, 0.3% 3%
were prepared . The six bacillus isolates were then inoculated, each in a set
of these media and grown at 50°C. After 24 hours the amlolytic activity was
determined as described above (expt. 3.2.6.3.)
3.2.5.4. Effect of Different Organic Nitrogen:
The Bacillus cultures were inoculated in nutrient broth media containing
different nitrogen sources such as peptone, tryptone, malt extract and meat
extract at a concentration of 1 % (w/v), at 50°C and pH as in experiment
3.2.5.3. After 24 hours, samples were taken for determination of amyloltic
activity.
3.2.5.5. Effect of Metal Ions:
Seven metal ions i.e. Mg^, Ca^, Zii"", Fe^, Mil**, Li"" and Cu4* in their
sulphate form except Ca** which was in the chloride form, were added to
nutrient broth media to make a final concentration of 5 mM. The media were
autoclaved at 121°C for 15 minutes,. After cooling, they were inoculated
30

with Bacillus isolates and incubated at 50°C and pH as above (expt 3.2.5.3)
for 24 hours. The amylolytic activity was determined.
3.2.5.6. Effect of Different Carbon Sources:
Six different carbon sources namely, starch, sucrose, maltose, lactose,
fructose and glucose were added separately to the basal nutrient broth media
at 0.05%(w/v) for isolates Bacillus licheniformis SUDK,, SUDK2, SUDK4
and Bacillus subtilis SUDK3 and 2% for Bacillus lichen (form is SUDO and
Bacillus circulans SUDK7. The pH as described in experiment 3.2.5.3.
After sterilization the media were inoculated with bacillus culture and
incubated at 50°C for 24 hours. Samples were then taken for determination
of amyloltic activity.
3.2.6 Determination of Amyloltic Activity (Enzyme Assays):
3.2.6.1 Preparation of Buffer:
The buffer used for the enzyme assay was 0.05M sodium phosphate buffer
pH 7.0. This buffer was prepared by adding 390 ml of 0.05 M monosodium
phosphate soultion to 610 ml of 0.05 M disodium phosphate solution, the pH
was adjusted to 7.0 using an electronic pH media.
3.2.6.2 Preparation of 3,5 Dinitrosalcvlic Acid (DNS):
3,5 Dinitrosalcylic acid (DNS) was prepared by dissolving in distilled water,
4g. DNS, 4g. sodium hydroxide, 0.8g phenol, 0.5g. sodium sulphite and 80g
sodium potassium tartarate in a total volume of 400 ml.
31

3.2.6.3. Determination of Enzyme Activity:
Amylolytic activity was measured by the method of Bernfeld (1955) as
modified by Miller( 1959).The assay mixture contained 2 ml of a solution of
1% starch in 50 mM sodium phosphate buffer (pH 7.0) and 0.1 ml enzyme
solution. After 10 minutes of incubation at 40°C, the reaction was stopped
by adding 2 ml of 3,5 dinitrosalicylic acid reagent and the tubes were heated
at 100°C for 5 minutes. The absorbance was measured spectro-
photometrically at 540 nm against a blank containing buffer instead of the
culture supernatant. Another blank was prepared using part of the
supernatant which was heated at 100°C for 5 minutes and used in the
reaction in place of buffer. No difference in absorbance between the two
blanks was observed. The amount of reducing sugars was calculated from a
standard curve constructed by using maltose (Fig 1)
3.2.6.4. Determination of Proteins:
Protein was determined using the dye- binding method of Bradford( 1976).
Commassie Brilliant Blue G250 (100 ing) was dissolved in 500 ml of 95%
ethanol. To this solution 100 ml of 85% (w/v) phosphoric acid was added
and the mixture was diluted with distilled water to a final volume of 1 liter.
This solution was then filtered and kept in a brown bottle. Five ml of
Commassie Brilliant Blue G250 solution were added to 0.1 ml protein
sample (containing 0.1-1 mg protein/ml). After mixing the absorbance was
read at 595 nm after two minutes and before one hour against a blank
prepared using 0.1 ml of phosphate buffer and 5 ml. of the Commassie
Brilliant Blue G250 reagent. A standard curve was constructed using bovin
serum albumin (Fig 2).
32
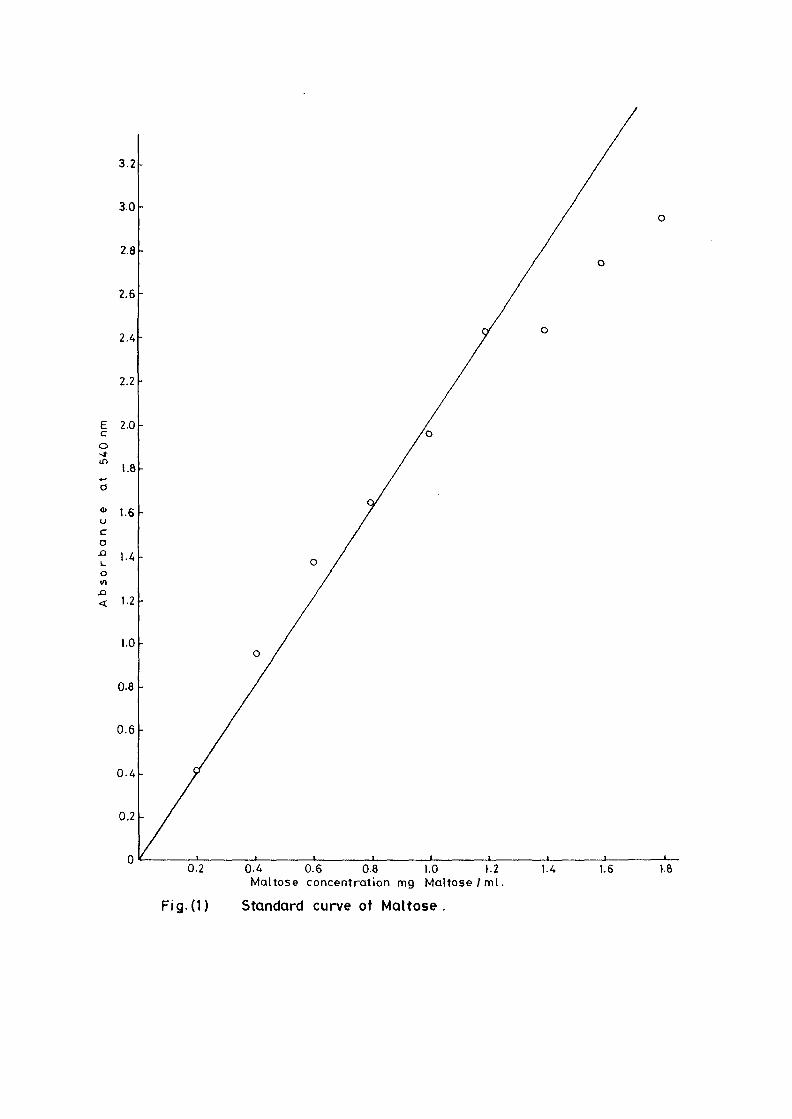
3.2
3.0
2.8
2.6
2. A
2.2
E 2.0c
1.8
01 1.6
ca•? 1-4on
1.2
1.0
0-8
0.6
0.4
0.2
0.2 0.4 0.6 0.8 1.0 1.2Maltose concentrat ion mg M a l t o s e / m l .
Fig( i ) Standard curve of Maltose.
14 1.6 1.8
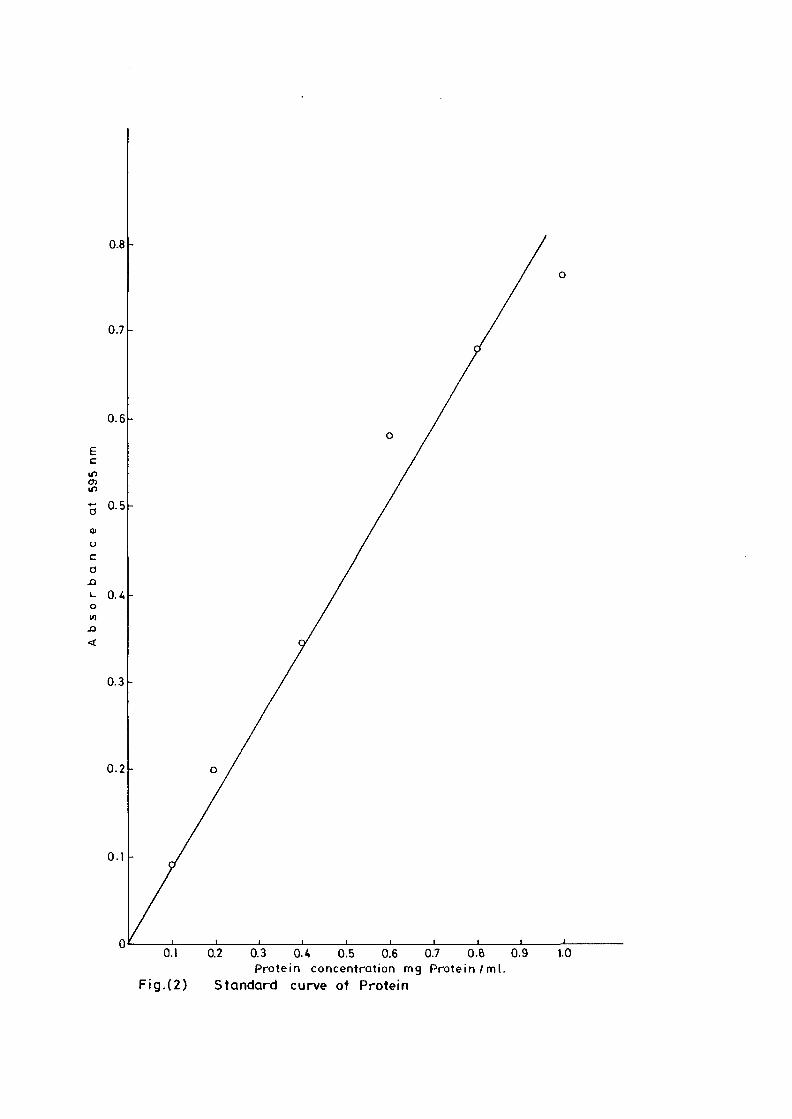
0.8
0.7
0.6
Ecin
en
uca
X>L.O
0.5
0.4
0.3
0.2
0.1
0.1 0.2 0.3 0.4 0.5 0.6 0.7 O.ft 0.9Protein concentration mg Protein /m l .
Fig.(2) Standard curve of Protein
1.0

3.2.6.5 Enzyme Units and Specific Activity:
One enzyme unit is defined as the amount of enzyme that catalyzes the
liberation of 1 nig. maltose per minute under the assay conditions. The
specific activity was taken as unit per milligram protein under the defined
conditions. It was calculated from the equation:
_ . „ . . mg maltose produced / mlSpecific activity - incubation time in minutes X mg. protein / ml
3.2.6.6 Amylase Activity:
3.2.6.6.1 Effect of Temperature on Amylase Activity:
Enzyme assays were carried out at pH 7.0 and at different temperatures
namely 40°C, 50°C, 60°C, 70°C, 80°C and 90°C. The amylase specific
activities for the six isolates were determined at each temperature.
3.2.6.6.2. pH Optima:
0.05M Sodium phosphate buffers with pH values of 4.0, 5.0, 6.0, 7.0, 8.0,
9.0, 10.0, 11.0 and 12.0 were prepared. In each buffer was dissolved starch
to make a solution of 1 % and the activity was determined at 70°C for
Bacillus lichemformis SUDK, SUDK4 and SUDO and at 60°C for Bacillus
lichenifonnis SUDK2, Bacillus subtilis SUDK3 and Bacillus circulans
SUDK7.
3.2.6.6.3. Effect of Substrate Concentration:
Enzyme assays were earned out at different soluble starch concentration
(0.5%, 1.0%, 1.5%.2.0%,2.5% and 3.0%) dissolved in 0.05M sodium
33

phosphate buffer pH 7.0. The amylase activities were determined for all
isolates at each substrate concentration and at temperature as above (expt.
3.2.6.6.2).
3.2.6.6.4. Effect of Reaction Time:
Enzyme assays were carried out at different reaction times namely, 10 min,
20 min, 30 min, 40 min, 50 min and 60 minutes. The amylase activities were
determined at each time interval. 0.05 M phosphate buffer pH 7.0 containing
1% starch was used and the temperature being as above (expt. 3.2.6.6.2).
3.2.6.6.5. Effect of Divalent Cation on Amylase Activity:
Seven dialvent ions namely Ca", Mg++, Mn^, Zn"", Cu++, Li++ and Fe^ in
their sulphate form except Ca4^ which was in it's chloride form (each at a
concentration of 5mM) were prepared in 0.05M phosphate buffer pH 7.0.
Amylase activity was measured for each cation after 10 minutes of
incubation, the conditions for substrate concentration and temperature were
as described above (expt 3.2.6.6.3).
3.2.6.6.6. Effect of NaCi on Amvlase Activity:
The activities of the amylases of the six isolates were determined using
0.05M phosphate buffer pH 7.0 containing various concentrations of sodium
chloride (0-5M). The reaction mixture was incubated for 1 hour and the
amylase activities were measured at each concentration as described above
(expt 3.2.6.6.3).
34

3.2.7. Identification of Enzyme Products:
Thin layer plates (Silica gel 0.5 cm in thickness) which were readily coated
and commercially available were used. A strip of 25 x 10 cm was cut using a
pair of scissors.
A reaction mixture of 2 ml 1% starch in sodium phosphate buffer (0.05 M,
pH 7) and 1 ml crude enzyme was incubated as above (expt.3.2.6.6.6) for 1
hour. From each reaction mixture 50 u.1 were spotted in portions on the thin
layer plate. Together and alongside these reaction spots were spotted similar
quantities of 0.01% of glucose and maltose. After drying, the plate was
developed in a solvent mixture composed of n-butanol: ethanol: water in a
ratio of 4: 2.2: 2 in a chromatographic tank. When the front of the solvent
reached nearly the top of the plate; the plate was taken out and dried in the
air. Sugars were detected according to the method of Trevelyan etal (1950)
as follows:
The plates was first passed rapidly through a reagent solution of 0.1 ml
saturated aqueous silver nitrate solution in 20 ml acetone. After drying in air
it was sprayed with alcoholic sodium hydroxide solution (0.5N solution of
sodium hydroxide in aqueous ethanol ) and finally washed with 0.5M
sodium thiosulphate solution to remove the brown silver oxide background.
Reducing sugars appeared as black or dark brown spots on white
background.
3.2.8. Enzyme Purification:
3.2.8.1. Column Chromatography:
3.2.8.1.1. Packing and Equilibration of the Column:
DEAE- Sephadex A-25 was washed several times with distilled water,
always decanting the fines until the supernatant was clear. It was then
35

washed with 0.05 M sodium phosphate buffer pH 7.0 and the slurry was
poured into a chromatographic column (1.8 x 20 cm).The suspension was
allowed to settle and the excess buffer was run off. More buffer was passed
into the column using a peristaltic pump, until the pH of the effluent was 7.0.
3.2.8.1.2. Application of the Enzyme on DEAE-
Sephadex A-25:
The crude enzyme preparations of the six culture filtrate {Bacillus
lichenifonnis SUDK^SUDK^UDIQ and SUDO, Bacillus subtilis SUDK3
and Bacillus circulans SUDK7), were applied separately to a column of
DEAE-Sephadex A-25, equilibrated as described above (expt.3.2.8.1.1).
After all the sample had entered the resin, one bed volume of the
equilibrating buffer was passed through until the unbound proteins were
removed. No activity in the washing was detected. The enzyme was eluted
with a linear gradient of sodium chloride (0-0.4 M) in 200 ml of sodium
phosphate buffer (0.05M and pH 7.0) with the aid of gradient mixer. The
first reservoir of the gradient mixer contained 100 ml of 0.05 M sodium
phosphate buffer pH 7.0 while the second one contained 0.4 M NaCl in 100
ml of the same buffer. The two reservoir were connected through their bases
by a narrow opening controlled by small valve. The first one which was
stirred magnetically was connected to the top of the column by a rubber
tubing. The flow rate was adjusted to 1 ml per 1 minute and the 200 ml of
eluents were collected into 40 tubes (1 x7 cm) using an automatic circular
fraction collector.
Enzyme activity and protein concentration were determined in each fraction
as described in the assay method. Fractions of the highest specific activity
were pooled together and kept for further studies.
36

3.2.9. Storage Stability:
Six fractions of 5 ml. Each from the pooled partially purified enzyme were
taken in sample bottles; three of these were stored at 4°C in a refrigerator,
the rest were stored in a freezer at -20°C . Every week the enzyme activity
was assayed, both for the frozen enzyme and the enzyme in the refrigerator.
The change in absorbance was measured and the residual activity was
calculated.
3.2.10. Determination of Km and Vmax:
Maximum velocity and Michaelis constant were determined using the
Bernfeld Method (1955). Initial velocities at different starch concentrations
were measured (see expt. 3.2.6.6.3) and the results were plotted by the
method of Lineweaver and Burk(l 934) as modified by Dixon (1953). The
intercepts on the 1/v axis and the slopes of the lines were further analyzed to
evaluate the maximum velocity and Michaelis constant.
37

4.1. Screening of Bacterial and Fungal Isolates for
Amylase Production:
Eighty six isolates of bacteria and fungi were isolated from food and soil
materials in Sudan (Table 1 and 2). Of these, sixty six were bacteria, forty
two from various food sources and twenty four from different soils. The
remainder were fungi obtained from food materials.
4.1.1. Selection of Isolates:
When bacterial isolates were grown on nutrient agar plates containing 0.2%
soluble starch as a major carbon source, forty one isolates formed clear
zones around the colonies when treated with iodine as described in the
materials and methods section.
On the basis of the index of amylolytic activity ten isolates were chosen for
further investigation. All these gave an index in the range of 2.1-4.5 cm
(Table 1).
The twenty fungal isolates were also tested for the production of amylolytic
activity. Eleven gave positive result but with lower level of amylolytic
activity relative to the bacterial isolates (Table 2).
Similar findings have been published on bacteria (Mai, et al, 1992) and
Rhizopus sp. (Hesseltine et al., 1963; Ellis et al, 1974 and Lim et al, 1987).
However, the fungal isolates were dropped out because of their lower
amylolytic activities.
38
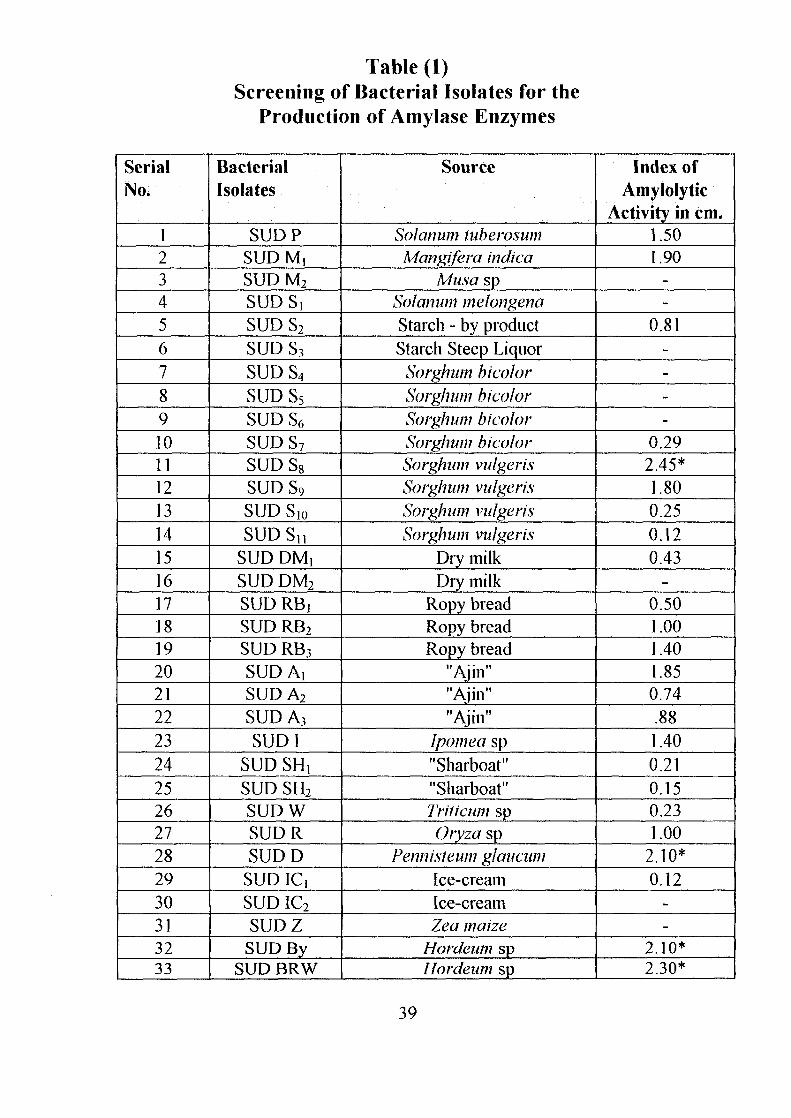
Table (1)Screening of Bacterial Isolates for the
Production of Amylase Enzymes
SerialNo.
123456789101112131415161718192021222324252627282930313233
BacterialIsolates
SUDPSUDM,SUD M2
SUDS!SUDS2
SUD S3
SUDS4
SUDS5
SUD S6
SUDS7
SUD S8
SUDS9
SUD Slo
SUDSnSUD DM,SUD DM2
SUD RBjSUD RB2
SUD RB3
SUD A,SUDA2
SUD A3
SUD1SUD SHt
SUD SH2
SUDWSUDRSUDD
SUDlCiSUD IC2
SUDZSUD By
SUD BRW
Source
Solarium luberosumMangifera indica
Musa spSolatium melongenaStarch - by productStarch Steep Liquor
Sorghum bicolorSorghum bicolorSorghum bicolorSorghum bicolorSorghum vulgerisSorghum vulgerisSorghum vulgerisSorghum vulgeris
Dry milkDry milk
Ropy breadRopy breadRopy bread
"Ajin""Ajin""Ajin"
Ipomea sp"Sharboat""Sharboat"Triiicum spOryza sp
Fennisieum glaucumIce-creamIce-creamZea maize
Hordeum spHordeum sp
Index ofAmylolytic
Activity in cm.1.501.90
--
0.81----
0.292.45*1.800.250.120.43
-0.501.001.401.850.74.881.400.210.150.231.00
2.10*0.12
--
2.10*2.30*
39
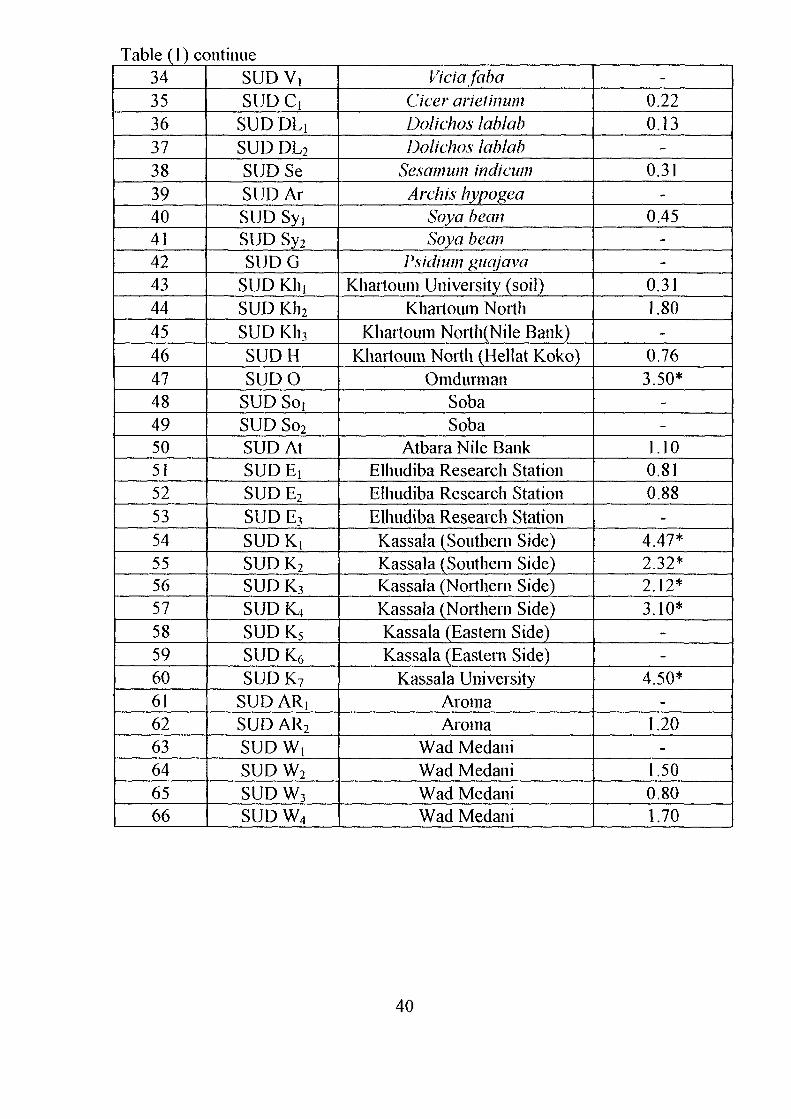
Table (1) continue343536373839404142434445464748495051525354555657585960616263646566
SUDV,SUDCi
SUD DLiSUD DL2
SUD SeSUDArSUD SyiSUD Sy2
SUDGSUD KinSUD Kh2
L_ SUD Kh3
SUDHSUDO
SUD So,SUD So2
SUD AtSUD EiSUDE2
SUDE3
SUDK!SUDK2
SUDK3
SUDK4SUDK5
SUD K6
SUDK7
SUD ARjSUD AR2
SUDW,SUD W2
SUDW3
SUDW4
ViciafabaCicer arieiinumDolichos lablabDolichos lablab
Sesamum indicumArch is hypogea
Soya beanSoya bean
Psidium giiajavaKhartoum University (soil)
Khartoum NorthKhartoum North(Nile Bank)
Khartoum North (Hellat Koko)Omdurman
SobaSoba
Atbara Nile BankElhudiba Research StationElhudiba Research StationElhudiba Research StationKassala (Southern Side)Kassala (Southern Side)Kassala (Northern Side)Kassala (Northern Side)Kassala (Eastern Side)Kassala (Eastern Side)
Kassala UniversityAromaAroma
Wad MedaniWad MedaniWad MedaniWad Medani
-0.220.13
-0.31
-0.45
--
0.311.80
-0.763.50*
--
1.100.810.88
-4.47*2.32*2.12*3.10*
--
4.50*-
1.20-
1.500.801.70
40
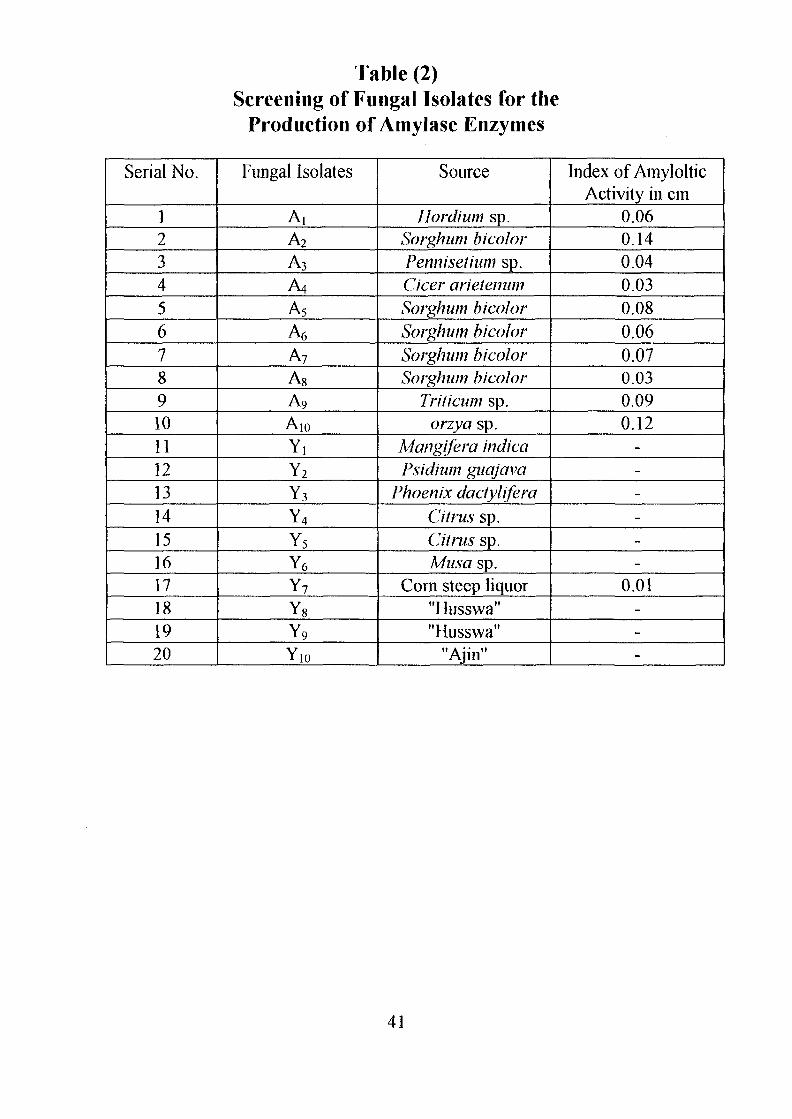
Table (2)Screening of Fungal Isolates for the
Production of Amylase Enzymes
Serial No.
1234567891011121314151617181920
Fungal Isolates
A,A2
A3
A4
A5
A6
A7
A8
A9
A10
YiY2
Y3
Y4
Y5
Y6
Y7
Y8
Y9
Y,o
Source
Hordium sp.Sorghum hi col orPennisethim sp.deer arielemimSorghum bicoiorSorghum bicoiorSorghum bicoiorSorghum bicoior
Trilicum sp.orzya sp.
Mangifera indicaPsidium guajava
Phoenix dactyliferaCitrus sp.Citrus sp.Musa sp.
Cora steep liquor"Husswa""Husswa"
"Ajin"
Index of AmylolticActivity in cm
0.060.140.040.030.080.060.070.030.090.12
------
0.01---
41

4.2. Identification of Bacterial Isolates:
All the ten Bacterial isolates were found to be Gram-positive, rod shaped,
endospore forming and catalase producers. They also formed pellicle in
liquid medium. All these characters suggest that these isolates belong to the
genus Bacillus as described by Gordon el al (1973). Table (3) represents the
morphological and biochemical characteristics of the ten isolates, which
were carried out according to Peter el al (1986).
The isolates SUD SRW, SUD BRW, SUD By, SUD K,, SUD K2, SUD K4
and SUD O resemble each other in most morphological and biochemical
characteristics. They all have white, lobate, large and flat colonies. The cells
are short motile rods. The spores are cylindrical or oval and central in non-
swollen sporangia. They are also catalase positive, facultatively anaerobic
and produce acid from mannitol and acid and gas from glucose. Also, they
reduce nitrate to nitrite, produce acetyl methyl carbinol (Voges-Proskaeur
test), grow at pH 5-7 and in 7% and 10% NaCl. They also grow at
temperature up to 65°C. It was, thus, concluded these organisms are all
identified as Bacillus licheniformis.
Isolates SUD D and SUD K7 resemble each other in all morphological and
biochemical characteristics. The two have creamy opaque colonies. The
cells are short motile rods and the spores are cylindrical and subtenninal in
non-swollen sporangia. The isolates are catalase positive, facultatively
anaerobic, produce acid from mannitol and acid and gas from glucose, grow
at pH 5-7 and at 60°C.
42
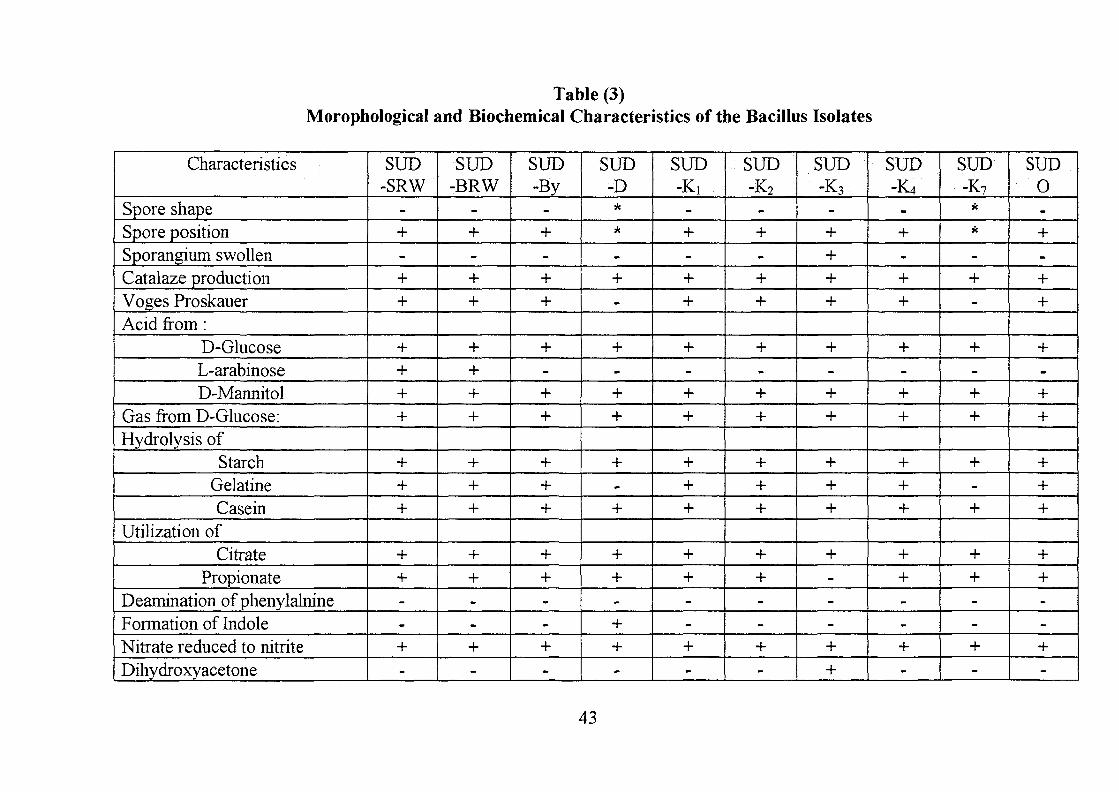
Table (3)Morophological and Biochemical Characteristics of the Bacillus Isolates
Characteristics
Spore shapeSpore positionSporangium swollenCatalaze productionVoges ProskauerAcid from :
D-GlucoseL-arabinoseD-Mannitol
Gas from D-Glucose:Hydrolysis of
StarchGelatineCasein
Utilization ofCitrate
PropionateDeamination of phenylalnineFormation of IndoleNitrate reduced to nitriteDihydroxyacetone
SUD-SRW
-+-++
++++
+++
++--+-
SUD-BRW
-+-++
++++
+++
++--+-
SUD-By
-+-++
+-++
+++
++--+-
SUD-D*
-+-
+-++
+-+
++-++-
SUD-K,
-+-++
+-++
+++
++--+-
SUD-K2
-+-++
+-++
+++
++--+-
SUD-K3
-++++
+-++
+++
+---++
SUD-K4
-
+-++
+-++
+++
++--+-
SUD-K7
*-+-
+-++
+-+
++--+-
SUD0-+-++
+-++
+++
++--+-
43
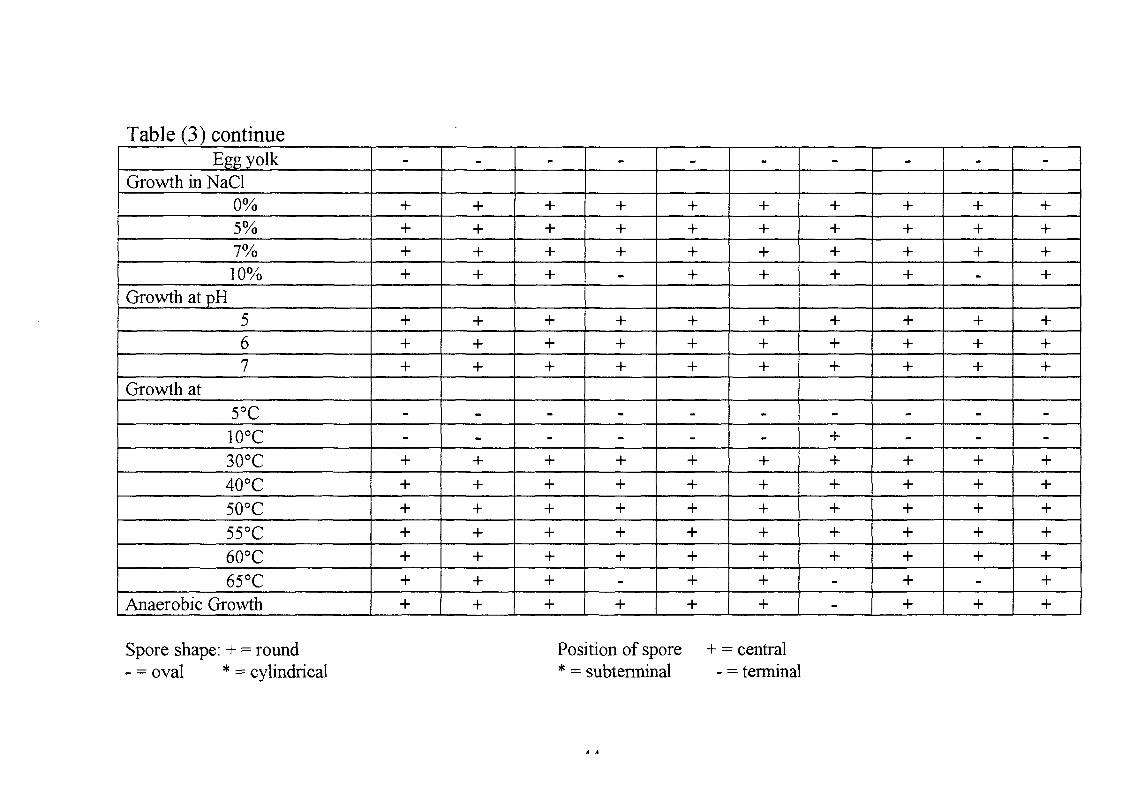
Table (3) continueEgg yolk
Growth in NaCl0%5%7%10%
Growth at pH567
Growth at5°C10°C30°C40°C50°C55°C60°C65°C
Anaerobic Growth
-
++++
+++
--+++++++
_
++++
+++
--+++++++
-
++++
+++
--+++++++
-
+++-
+++
--+++++-+
-
++++
+++
--+++++++
-
++++
+++
--+++++++
-
++++
+++
-++++++--
-
++++
+++
--+++++++
-
+++-
+++
--+++++-+
-
++++
+++
--+++++++
Spore shape: + = round- = oval * = cylindrical
Position of spore* = subterminal
+ = central- = terminal

These isolates give negative results with Voges-Proskaeur test and can not
hydrolyze gelatine. They are therefore classified as Bacillus circulans.
Isolates SUD K.i has some different morphological and biochemical
characteristics than the rest. It has a creamy, small, flat and round colony.
The cells are short motile rods occurring in chains. The spores are oval and
central in swollen sporangia. The organism produces catalase and gives a
positive test with Voges-Proskaeur test. It utilizes citrate and propionate,
oxidises glucose to produce acid and gas, produces acid from mannitol and
dihydroxyacetone from glycerol agar. This isolate is strictly aerobic. It
grows at 7% NaCl and at pH 5-7, but does not grow at 65°C. It was thus
identified as Bacillus subtilis.
4.3. Amvlase Production:
4.3.1 Effect of Temperature and Time Intervals on Amvlase:
Production:
The selected isolates were grown on nutrient broth media containing 0.5%
soluble starch as the major carbon source at pH 7.5. Samples were taken as
described in section three (Materials and Methods).
All the isolates showed maximum amylase production after 24 hour growth
(Fig3-Figl2).
Bacillus licheniformis SUD SRW, SUD BRW, SUD By and Bacillus
circulans SUD D showed maximum amylase production at 45°C (specific
activities at 45°C were 2.0, 1.7, 1.7 and 2.84 U/mg protein in order)
retaining about 23%, 66%, 37% and 9% in order of their maximum specific
activities at 50°C. (Fig.3-Fig 6). These isolates seem to be mesophilic and
are similar to the bacilli tested by (Khire and Pant 1992; Kwan et al, 1993
and Patel et al, 1993) using Bacillus sp. 64, Bacillus circulans, and
45
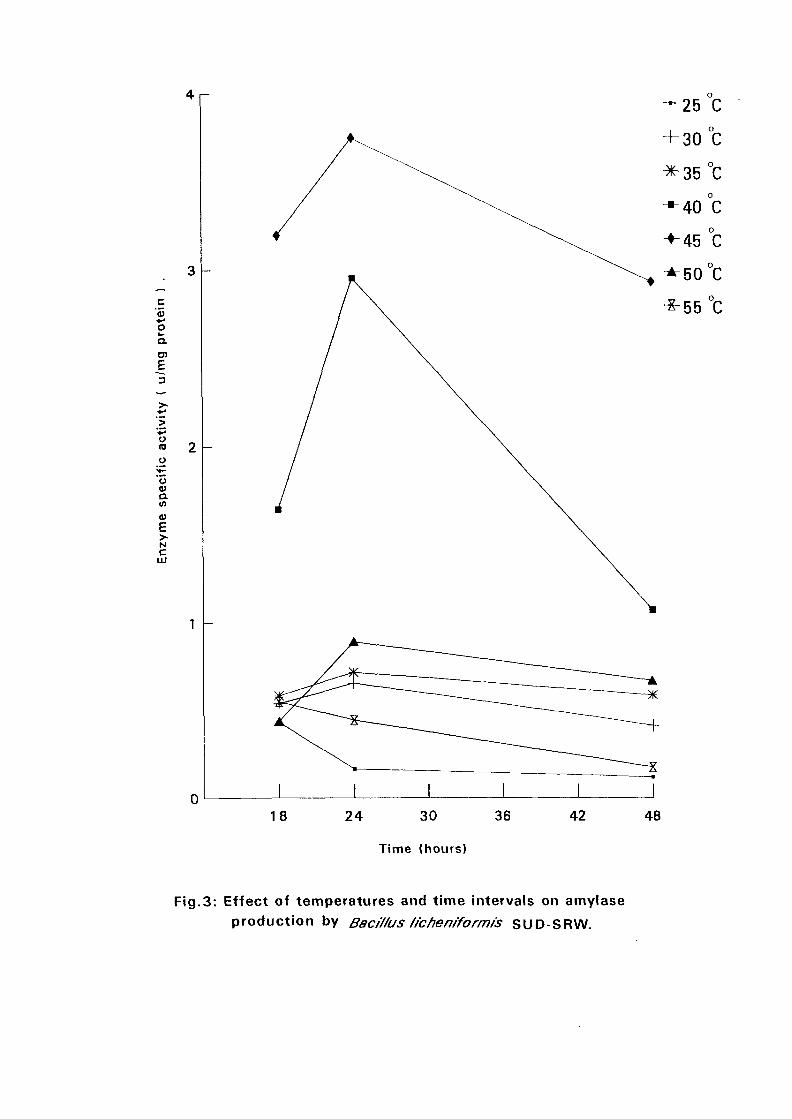
c'5•#-»oa.
£:
+
o
oaV)
Nc
LU
018 24 30
Time (hours)
36 42 48
Fig.3: Effect of temperatures and time intervals on amylase
production by Baci/fus /fc hen/form is SUD-SRW.
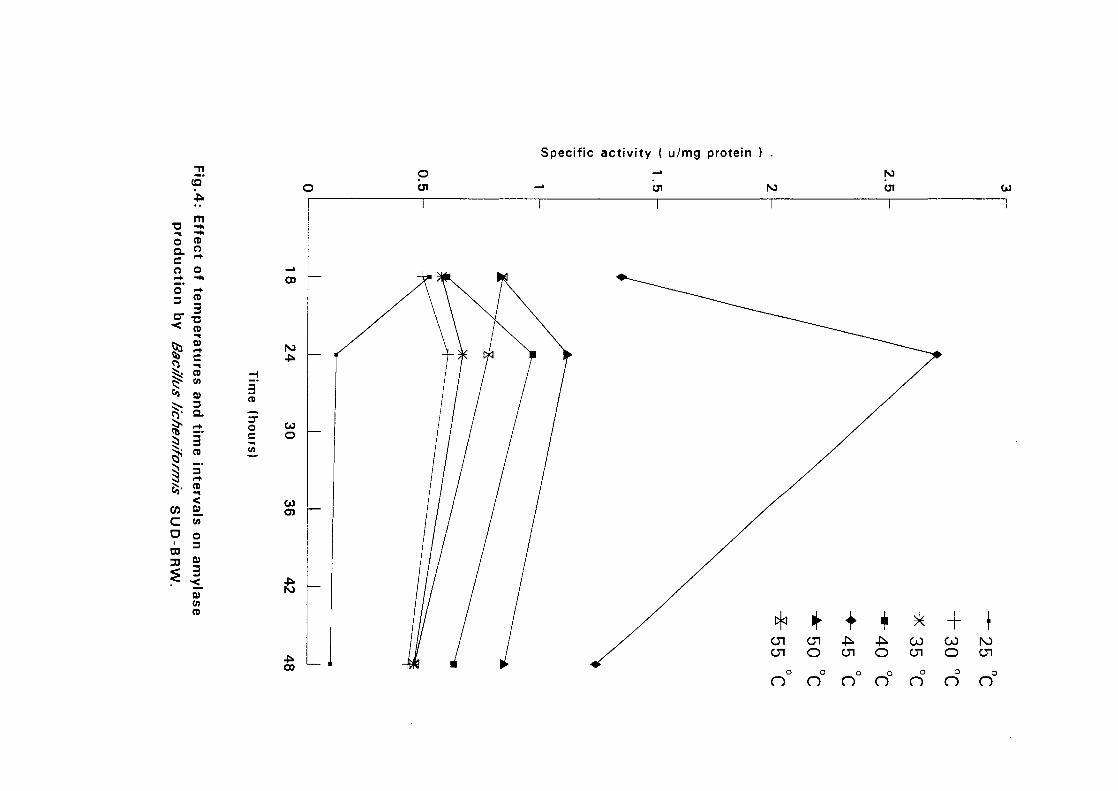
Specific activity ( u/mg protein ) .
•aoQ .Co
o'3
or• <
8&|$
I5i55-COcoCO
:>
r45.
m^^CDO
o^ <CD
3CD
C
CDCA
Q>3a
3CDint
CD
<
CA
o3
0)
3•<_
0)CACD
||B
3CD
ocCA
pen NJ
ro
CO
o
CO
oo
n n n n n no
n
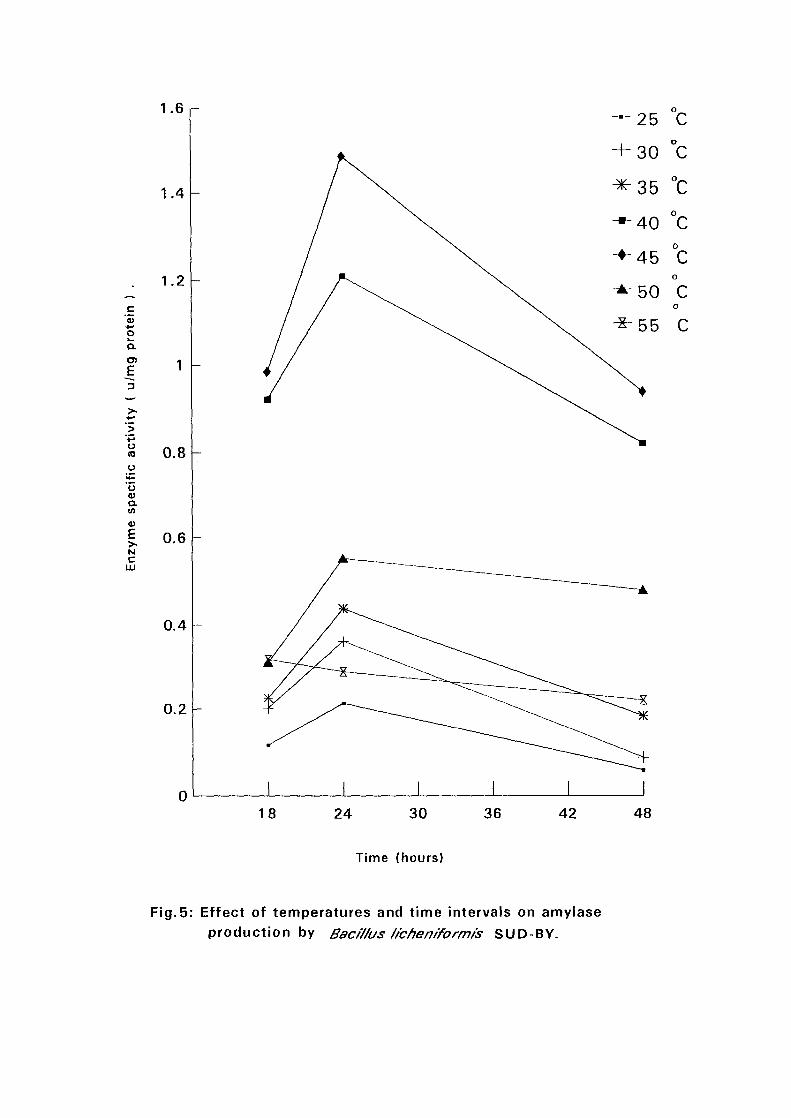
1.6
c+-•oQ.
E
u(0
oa.m
Nc
LU
1.4 -
1.2
0.8
0.6
0.4
0.2
018 24 30 36 42 48
Time (hours)
Fig.5: Effect of temperatures and time intervals on amylaseproduction by Bac/V/as /ichen/formis SUD-BY.
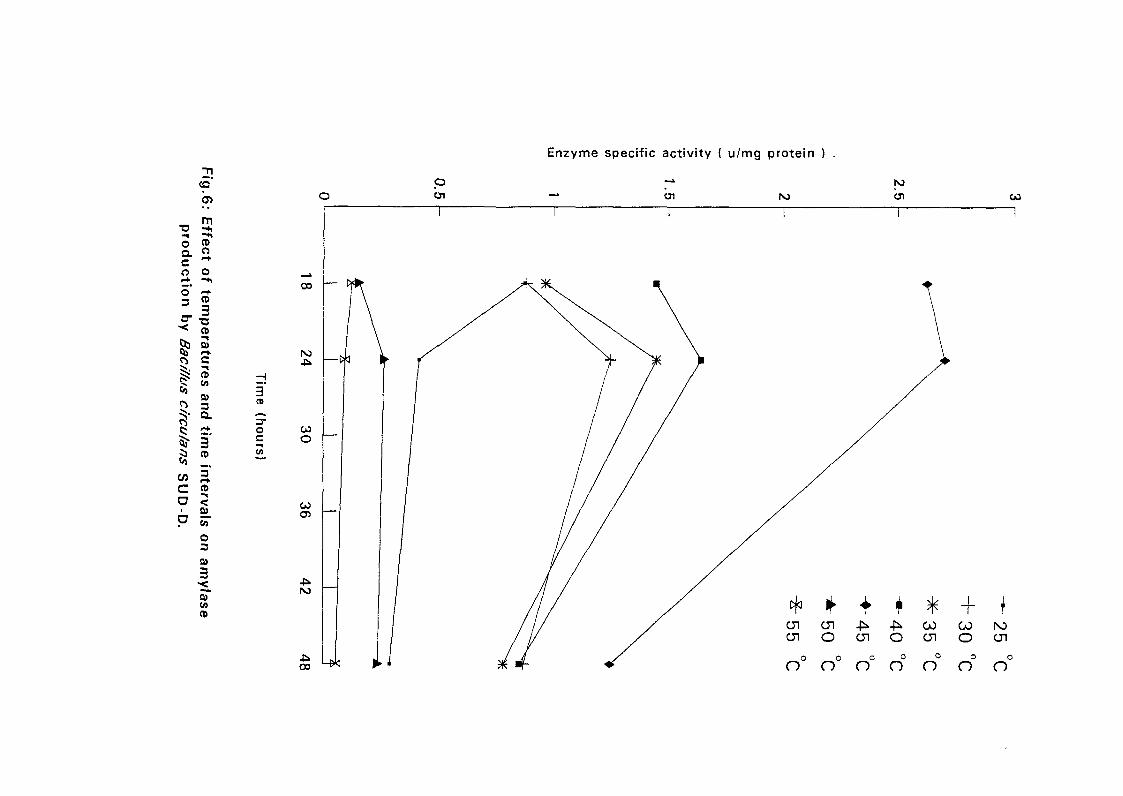
Enzyme specific activity ( u/mg protein )
<a OCJ1 CJ1 Ol
oaco5"3
CDO
O
CD
3
3
03
ands cir 1 tim
ecu/ani
w 3C" CDrvals
ID-D
.
o3am
yfase
3CDhours)
CO
o
CO
en
-IS.00

halobhim in order. The rest showed maximum amylase production at 50°C
(Fig.7 - Fig 12). Bacillus licheniformis SUD K,, SUD K2, SUD K4 and SUD
O retained about 34% , 2 1 % , 24% and 33% of their maximum specific
activities in order at 55°C (specific activities at 50°C were 3.5, 1.9, 3.4 and
1.95 U/mg protein in order), while Bacillus circulans, SUD K7 and Bacillus
subtilis SUD K3 retained about 23% and 58% of their maximum specific
activities respectively at 55°C (specific activities at 50 were 3.0 and 2.35
U/mg protein respectively). These isolates, all from soil, appear to be
thermophilic bacilli. The former ones (the mesophilic isolates) were isolated
from food materials. Similar results were reported by Satio and Yamanoto
(1975) using Bacillus licheniformis.
4.3.2. Effect of pH on Amylase Production:
The six bacillus isolates namely Bacillus licheniformis. SUD Ki, SUD K2,
SUD JQ, SUD O, Bacillus subtilis SUD K3 and Bacillus circulans SUD K7,
which grow maximally at 50°C, were chosen for further investigation. From
the results (Fig 13) it is seen that all the Bacillus isolates gave maximum
amylase productivity at pH 9.0 except Bacillus subtilis SUD K3 which
produced amylase maximally at pH 7.0. It is seen that amylase production
occurs over a wide range of pH values (5-10) for Bacillus licheniformis
SUD K, and SUD K4.
In this respect Burgess-Cassler and Imam(1991) and Mai el at (1992),
reported maximum amylase production at pH 7.0 by Lactobacillus
amyloverus and Bacillus stearothermophilus respectively, while Boyer and
Ingle (1972) and Medda and Chandra (1980) reported maximum amylase
production at alkaline pH 9.0 using Bacillus licheniformis. Bacillus
acidocladarius, on the other hand, showed maximum amylase production at
acidic pH (3.5) as reported by Kanno (1986).
46
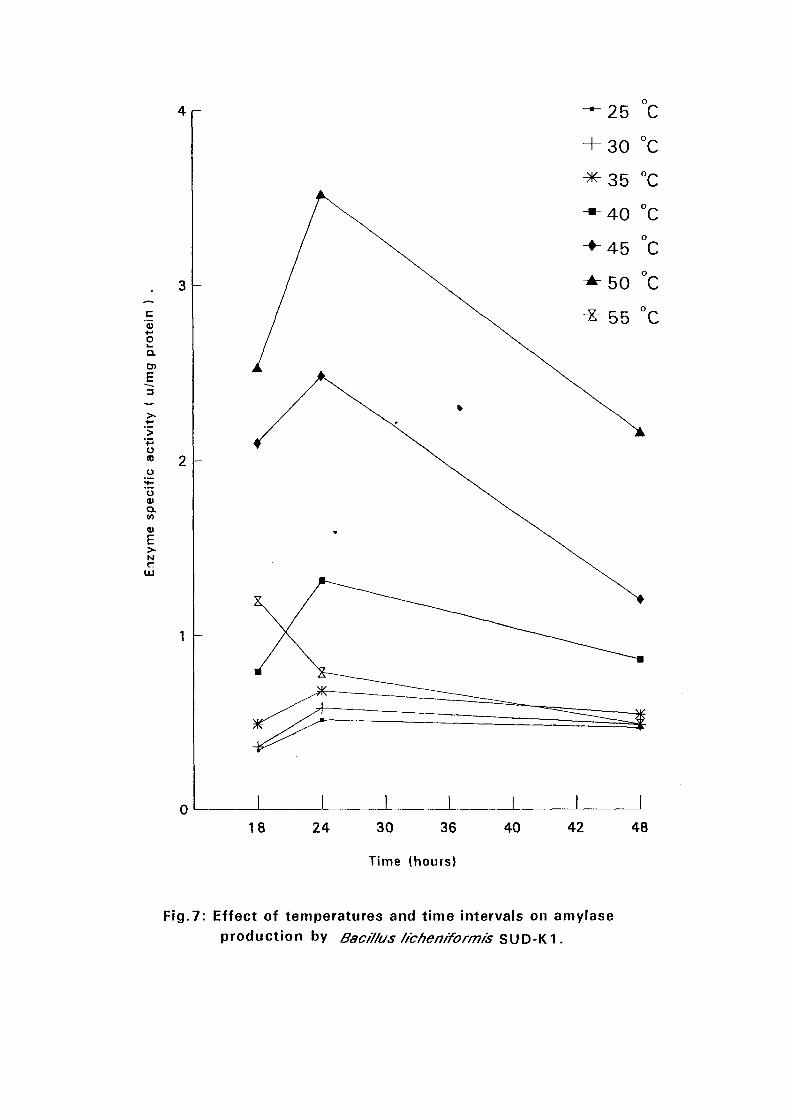
4 r -
oQ.
E
uid
ua>a.to
a>E
018 24 30 36
Time (hours)
40 42 48
Fig.7: Effect of temperatures and time intervals on amylase
production by Bac/7/us /ichen/formr's SUD-K1.
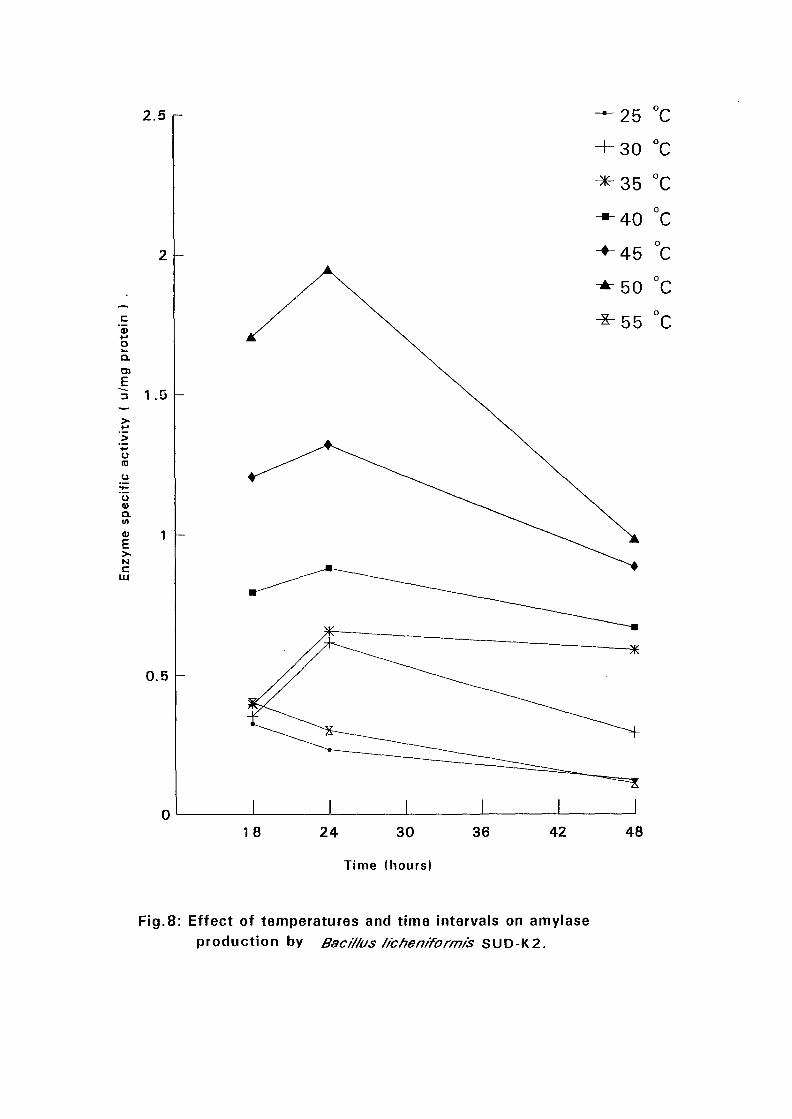
2.5 r
oQ.
3
>«+-•'>
O<0
oQ.(0
Nc
Ul
1.0
0.5
0
— 25 °C
+ 30 °C
45 °C
50 °C
55 °C
18 24 30
Time (hours)
36 42 48
Fig.8: Effect of temperatures and time intervals on amyiaseproduction by Bac/7/us f/cheniformfs SUD-K2.
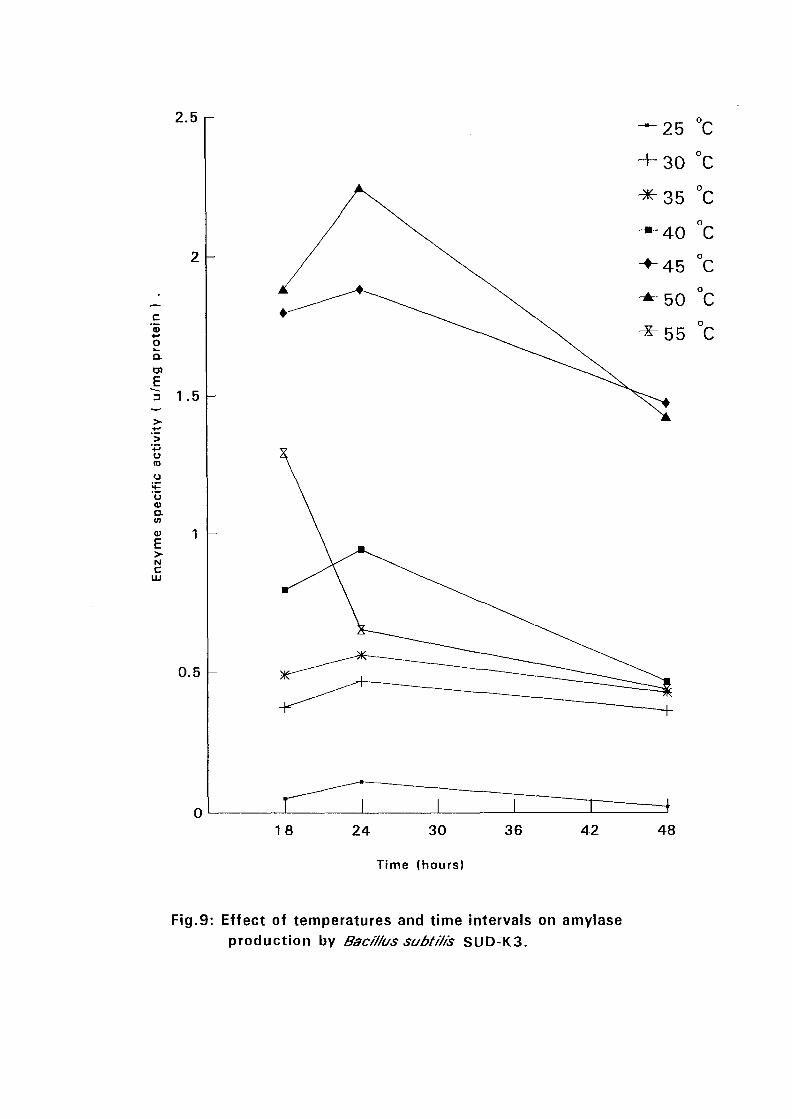
2.5
c'5+-»oha
Q.
E
u(0
oo.
Nc
LU
1.5
0.5
018 24 30
Time (hours)
36 42 48
Fig.9: Effect of temperatures and time intervals on amylaseproduction by ffac/7/us subt/ft's SUD-K3.
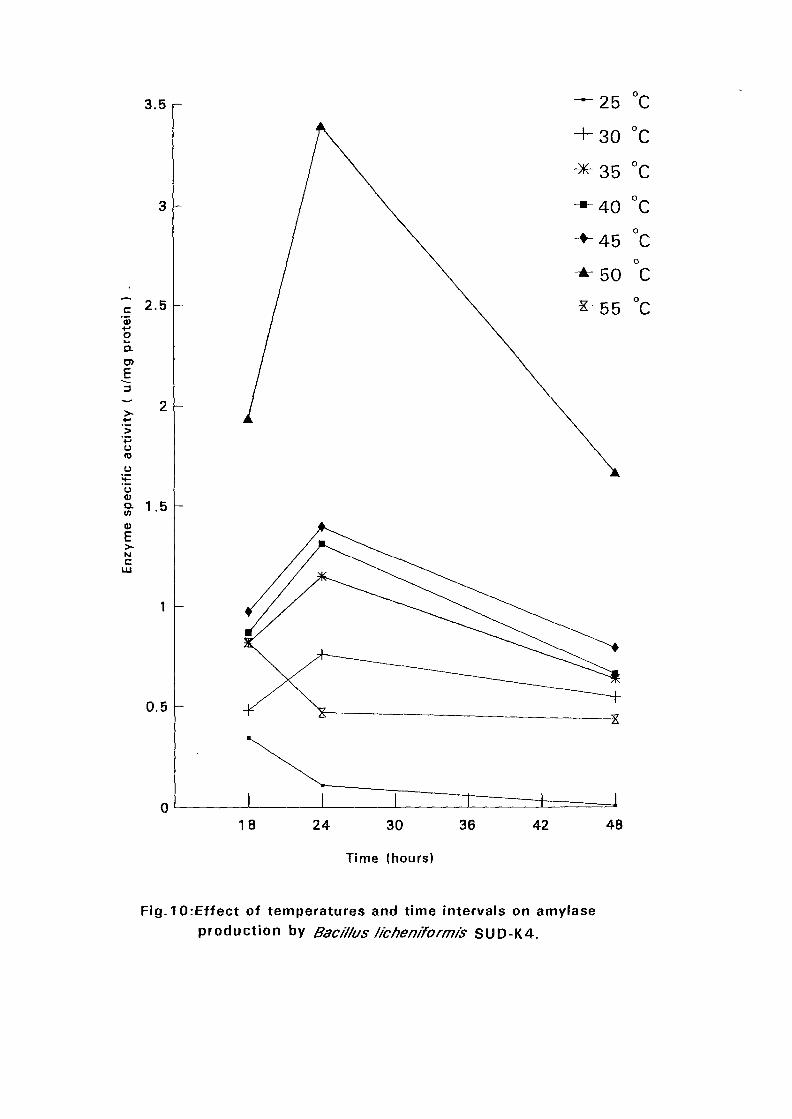
3.5 r
Q —
c'55oa
E
o(0o
o0)
0)E>-NC
2.5
0.5
0
Fig. 10rEffect of temperatures and time intervals on amylaseproduction by Bacillus /ichen/form/'s SUD-K4.
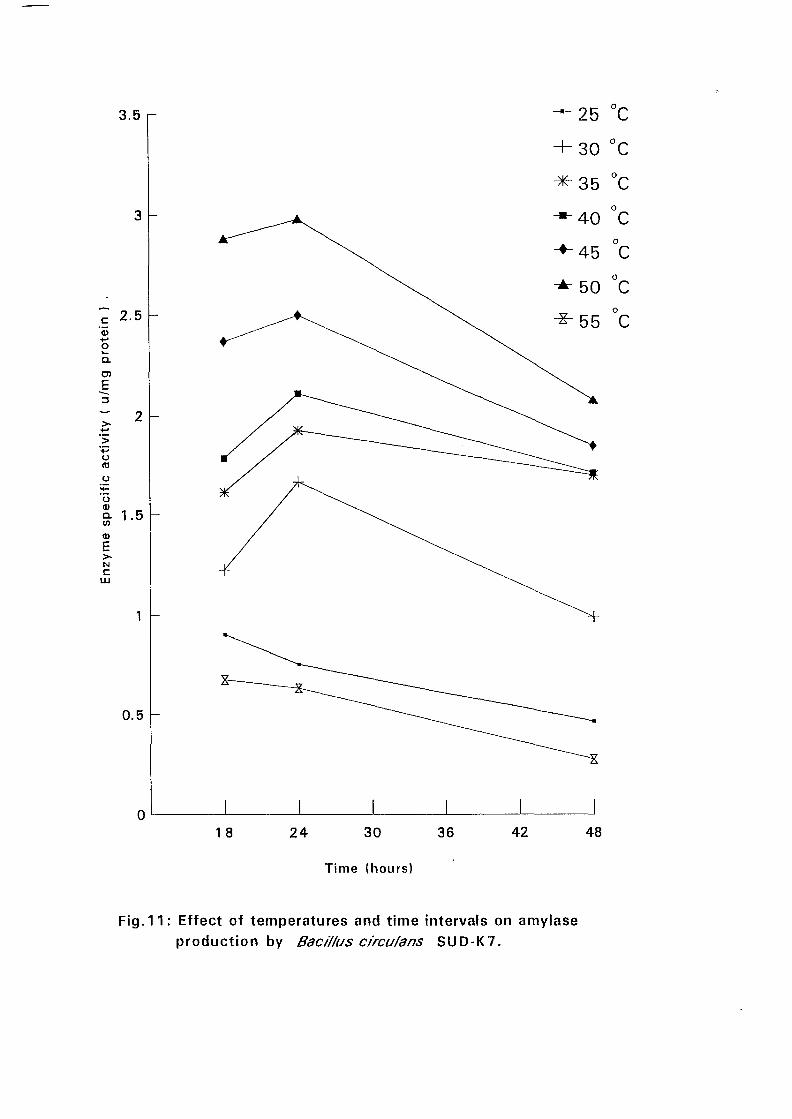
3.5 r
c'55+-»ow.Q.
E
o
o
(I)
ENc
UJ
2.5
I - 5
0.5
0
25 v^
18 24 30
Time (hours)
36 42 48
Fig.11: Effect of temperatures and time intervals on amylaseproduction by Bac/ffus circu/ans SUD-K7.
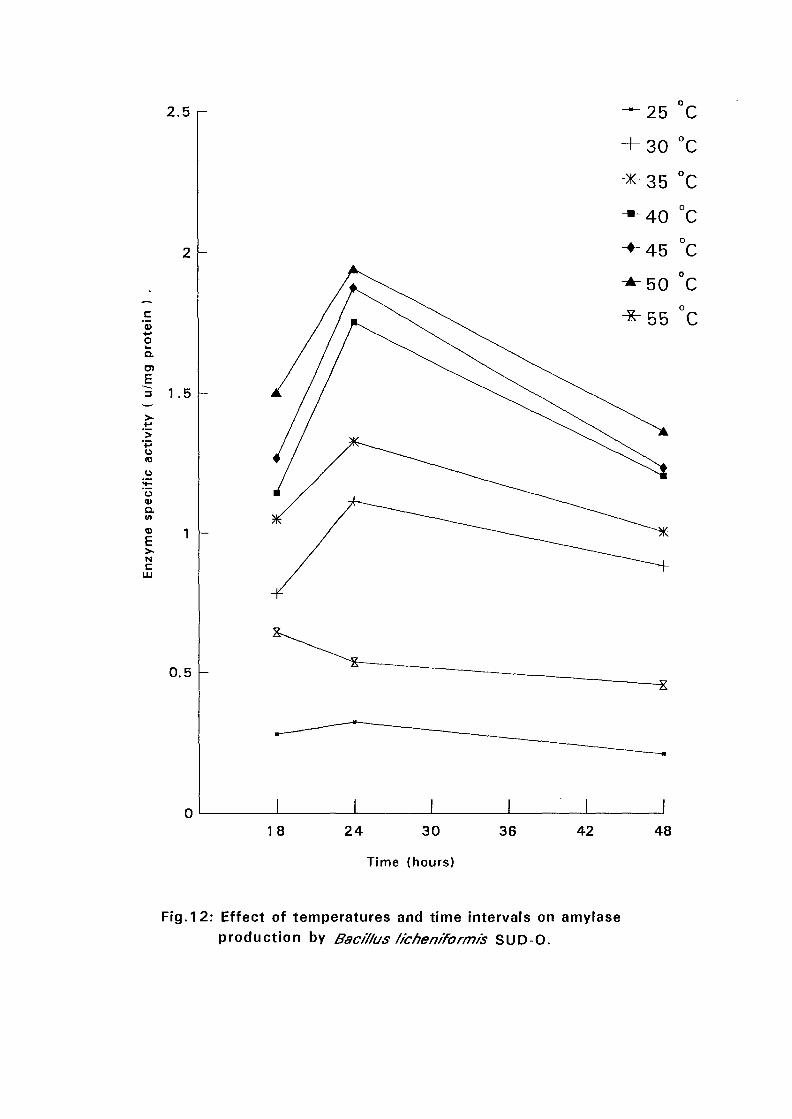
2.5 r
c'55oQ.
O)ji3
O(0
uVQ.tf>43
E>•
N
c
1 .5
0.5
0
25 C
30 °C
°C
°C
°C
°C
18 24 30
Time (hours)
36 42 48
Fig.12: Effect of temperatures and time intervals on amylaseproduction by Bacif/us f/cheniformis SUD-O.
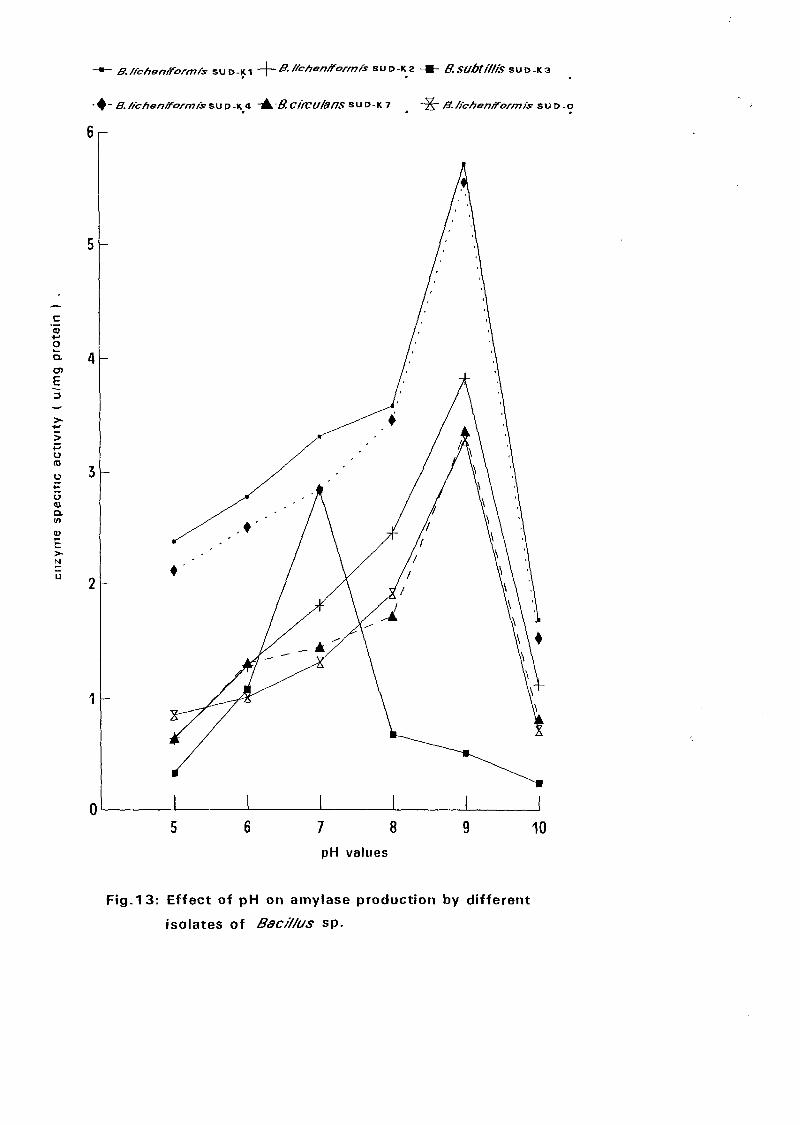
B./icfion/formis SUD-KI -\-B.ffchenxrorm/s SUD-KZ - » - B.SUbtiffiS SUD-K3
/S". /fc/ionrform/s sup-K4 -tk.B.Ctncufans SUD-K 7 -Jj- B./fchenfform/s su D-o
Fig.13: Effect of pH on amylase production by different
isolates of Bac/7/us sp.

4.3.3. Effect of Substrate Concentration on Amylase
Production:
Fig 14 shows the result of the effect of substrate concentration on amylase
production. It is seen that Bacillus licheniformis SUD K]? SUD K2, SUD K4
and Bacillus subtil is SUD K3 gave maximum amylase production at 0.5% of
soluble starch with a maximum specific activities of 5.72, 3.80, 5.50 and
2.86 U/mg protein in order. Bacillus licheniformis SUD O and Bacillus
circulam SUD K7, on the other hand, showed maximum amylase production
at 2% soluble starch with maximum specific activity of 4.3 and 4.2 U/mg.
protein respectively.
Substrate concentration appeared to be directly correlated with
saccharification of soluble starch. Production of reducing sugars increase
with the increase in substrate concentration in the growth medium. Above
0.5% no further increase in reducing sugars was detected by Bacillus
licheniformis SUD K,, SUD K2, SUDK4 and Bacillus subtilis SUDK3, but
there was a very slight decrease and this could be due to the product
inhibition of the enzyme. Similar result was obtained by Rani el al (1994)
who tested tapioca as a substrate for production of P-amylase by Bacillus
megaterium. For Bacillus licheniformis SUD O and Bacillus circulans SUD
K7 it was observed that there was a gradual increase in enzyme production
with increase in substrate concentration. Maximum specific activity was
obtained at 2% starch (4.5 and 4.3 U/mg protein respectively). Above this
concentration no increase in the production of reducing sugar was detected.
Bajpai et al (1992) reported that 2% com starch together with 3% com
gluten in addition to different mineral salt produced maximum yield of oc-
amylase from Bacillus sp. TCRDC-M. Rani et £7/(1994), concluded that
maltose production increased with increase in substrate concentration in the
growth medium of Bacillus.megaterium.
47
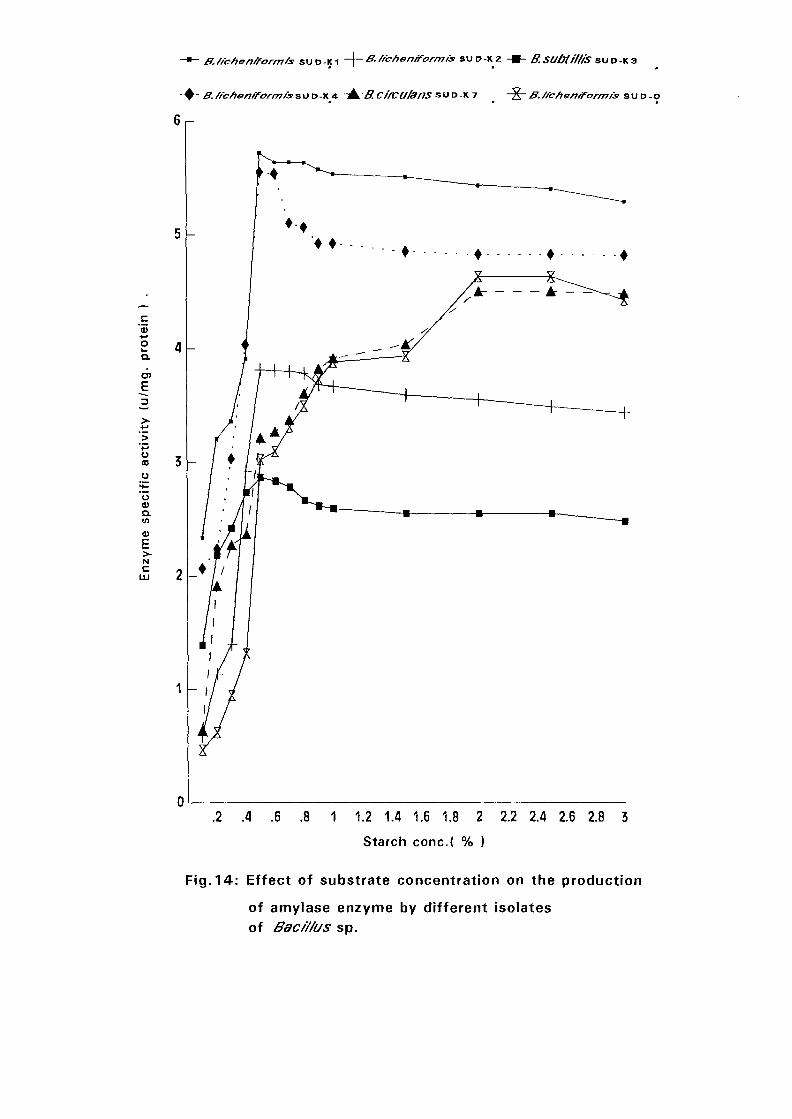
-*- B./fchen/form/s SUD-K1 "4~s- ffchonfTormts SUP-K2 -B- B.SUbtt'//fS SUD-K3
• - B.//c/70n/formts&uv-Kji -± B.CffCU/anS SUD-K7 _ - ^ - i?. //c/ien/formfs s u D - o
c'55••-•oQ.
- l>-[>'<-•U
Od)a.
vENc
LLI
.2 .4 .6 .8 1 1.2 1.4 1.6 1.8 2 2.2 2.4 2.6 2.8 3
Starch concl % )
Fig. 14: Effect of substrate concentration on the production
of amylase enzyme by different isolatesof Bac/"//us sp.

4.3.4. Effect of Different Metal Ions on Amylase Production:
The results of the effect of divalent ions namely, Mg4"4", Ba4^, Mn++ ^a4^
and Fe++ are presented in Figs (15-20).
It is seen that Bacillus licheniformis SUD K| gave maximum production
(5.7 U/mg. protein) in a medium containing Mg++ions. Addition of any of
the ions Ca++, Fe++ Mn++ and Ba++ to the broth media decrease the aniylase
production. In presence of these ions specific activities of 3.7, 3.1, 3.2 and
3.3 U/mg protein in order were obtained (Fig 15).
Ca++ and Mg++ ions enhanced aniylase production by Bacillus licheniformis
SUD K2 with specific activities of 3.0 and 3.7 U/mg protein respectively. At
the same time and under the same conditions the presence of Ba++, Mn++ and
Fe++ decreased the enzyme specific activity to 3.0,2.4 and 1.9 U/mg protein
in order (Fig 16).
In Fig. 17 it is seen that Mg4"1" stimulates amylase production by Bacillus
licheniformis SUD IC| with maximum specific activity of 5.5 U/mg protein
while Ca++ ,Ba++, Mil4"4" and Fel+ gave comparable specific activities which
were 3.4, 3.4, 3.2 and 3.1 U/mg protein in order. Also, Bacillus
licheniformis SUD O gave maximum amylase production in a medium
containing Mg++ with specific activity of 4.2 U/mg protein. Ca4^ and Ba++
reduced the enzyme specific activity to 3.6 and 3.5U/mg protein
respectively, while Fe4"1 and Mn4^ gave the lowest specific activities which
were 1.33 and 1.054 U/mg protein respectively (Fig. 18).
In Fig. 19 it is seen that the maximum amylase production by Bacillus
circulans SUD K7 was obtained in presence of Mg++ with specific activity
of 4.3 U/mg protein. Fe++ and Ba++decreased the amylase specific activity to
2.9-2.8U/mg protein respectively, while Ca++ and Mn^ ions gave an even
48
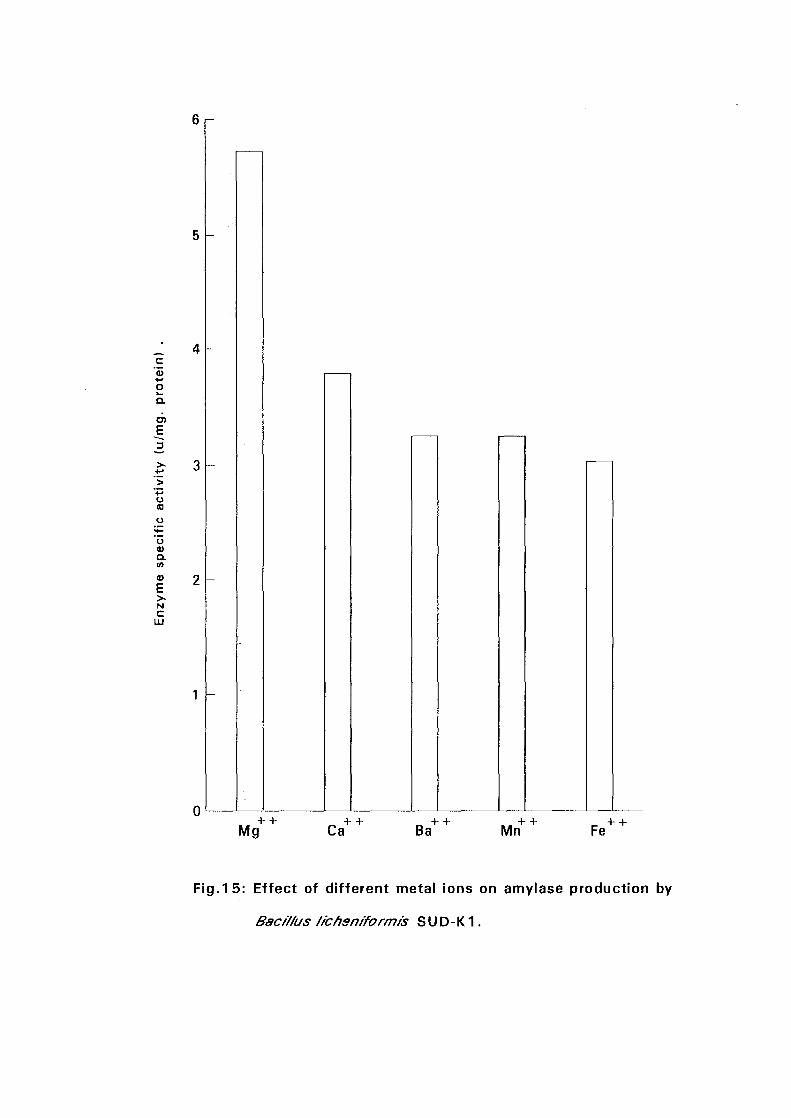
^ 4c
'55+-•oQ.
o(0
a.v>
NC
Mg Ca Ba Mn Fe
Fig.15: Effect of different metal ions on amylase production by
Bacif/us /fchsn/formis S U D - K1.
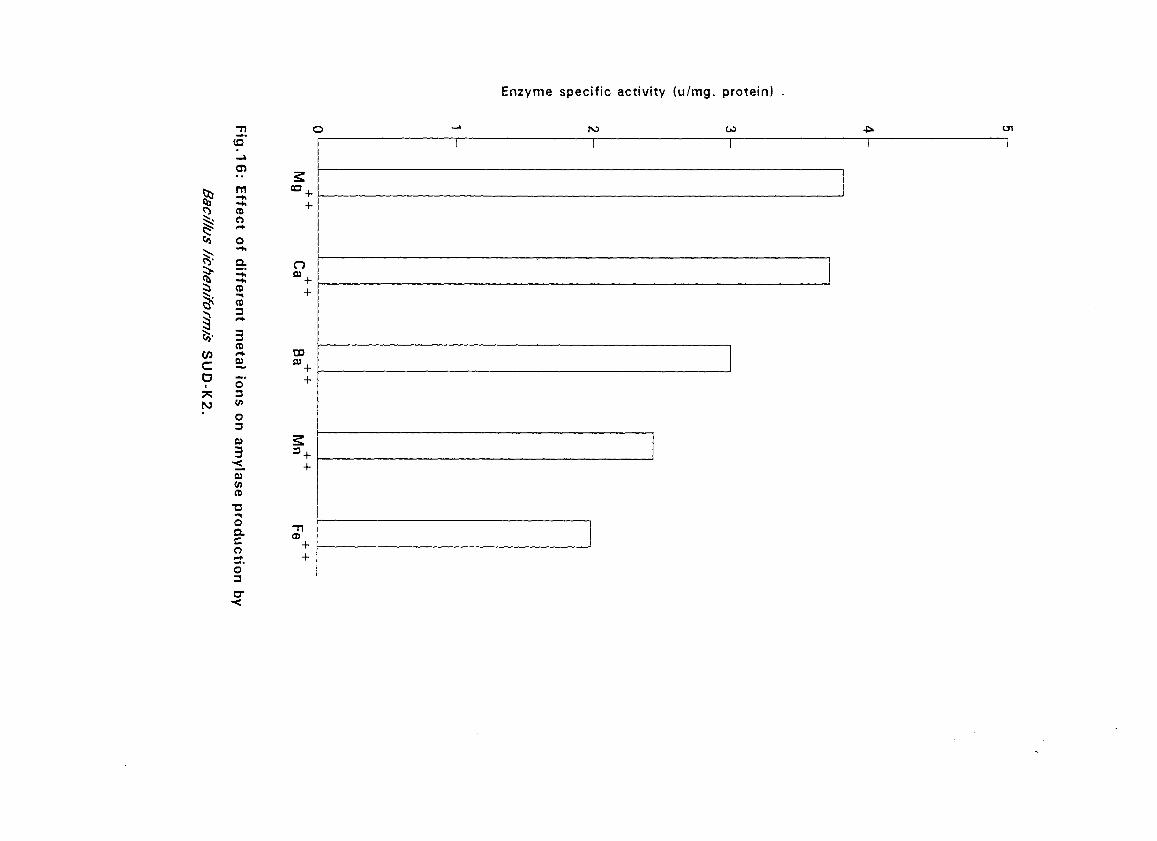
Enzyme specific activity (u/mg. protein)
3
?
!cDI
m
CD
«s 2
CDen
3CDr*
O3<J>
O30)
30)
CD
oa.co5'3
o0) ,
CO
-nCD
(V) CO
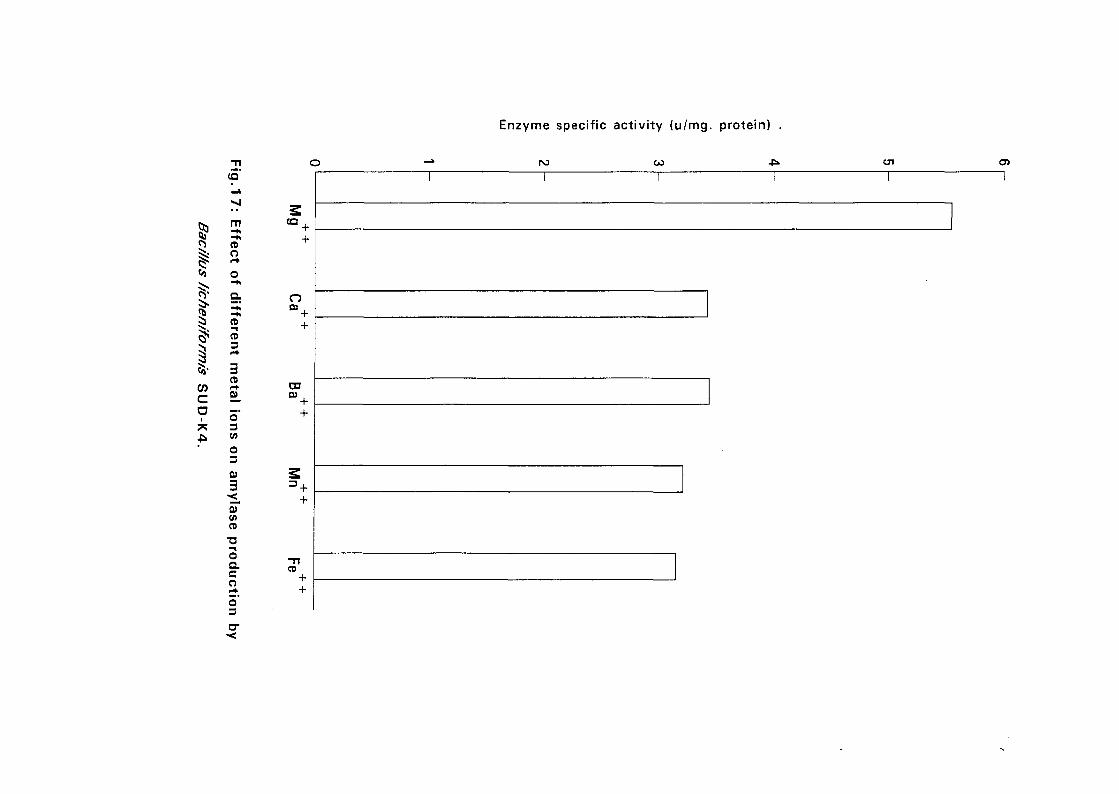
(Q
Enzyme specific activity (u/mg. protein) .
m-hCDO
++
ItoCo
CD
CD
3CD
0)
O3
o03
D301
O3B)
3
5TCOCD
' ++
oaco5'3
CD++
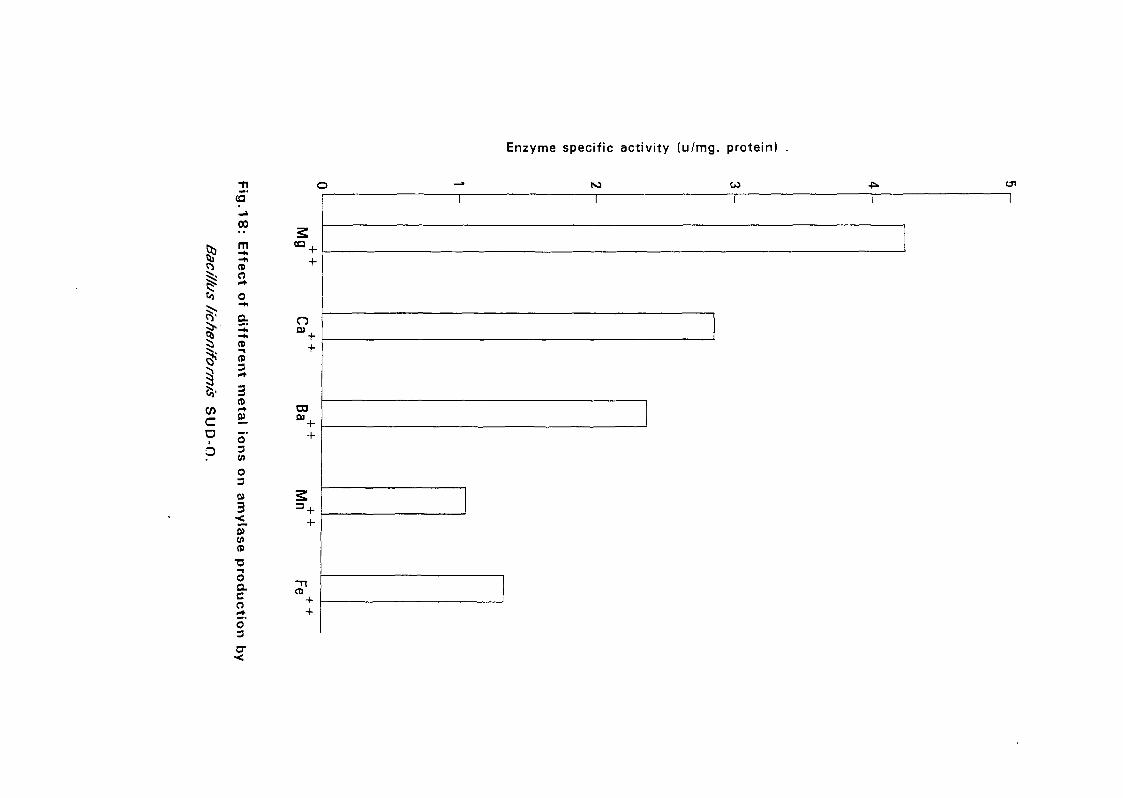
Enzyme specific activity (u/mg. protein)
<5"N3
00
m-*,-hCD
a
<a
wca
(0
CD
3CD
O3
n
CO
O3
0)(ACD
Oaco5"3
CD
+
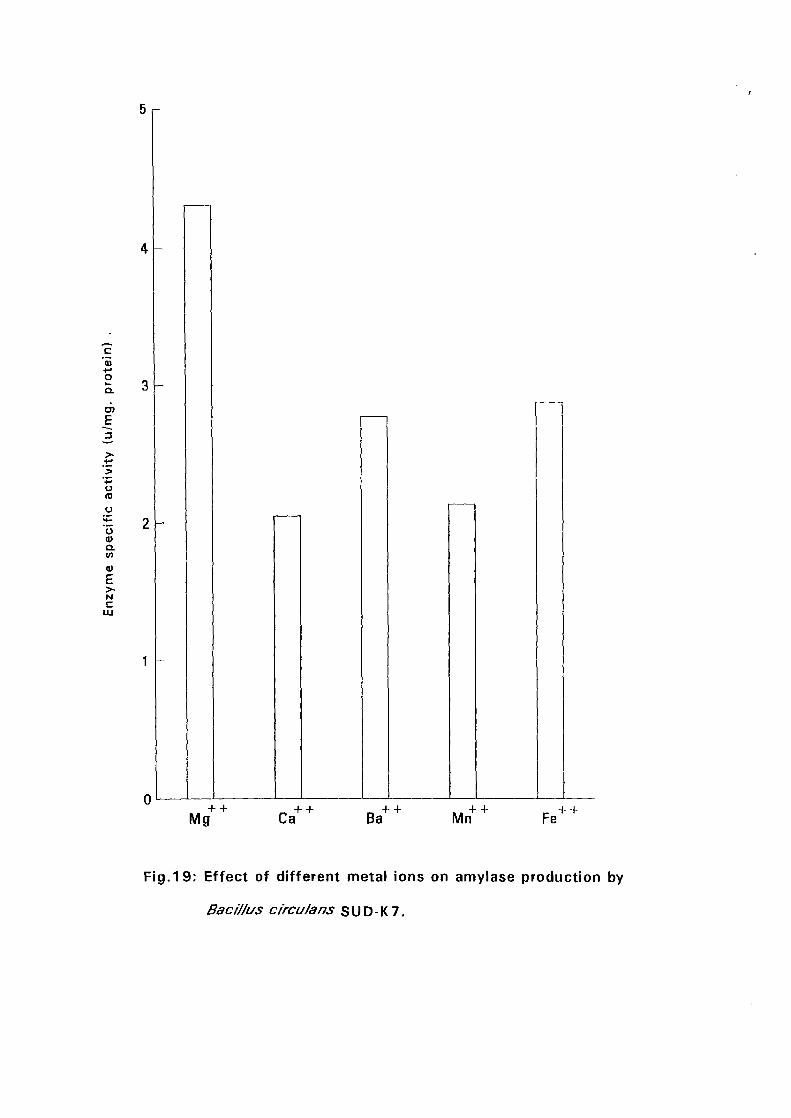
c'55oaa>E
u(0
oa>aV)
a>£>-Mc
LU
Mg Ca Mn Fe
Fig.19: Effect of different metal ions on amylase production by
Baci/fus c/rcu/ans SUD-K7.

lower specific activities to less than 50% (2.05 and 2.1 U/mg protein
respectively).
In Fig 20 it is seen that Bacillus subtilis SUD K3 produced maximum
amylase activity, also, in a medium containing Mg++ ion, the specific activity
being 2.9 U/mg protein. A slight decrease in enzyme specific activity was
obtained in the presence of Ca++ ion with specific activity 2.0 U/mg protein.
Fe^, Mii^ and Ba^ gave an even lower level of specific activities which
were 1.1, 1.2 and 1.3 U/mg protein in order.
All the data in Figs.( 15-20) was subjected to a two way analysis of variance
at p = 0.05 using Statistix 3.1 program and then followed by LSD.
From the results it is concluded that Mg++ ions gave the highest amylase
productivity for all isolates and showed significant difference from the rest
of the ions for isolates Bacillus•licheniformis SUD K,, SUD K4, SUD O
and Bacillus circulans SUD K7. On the other hand Bacillus licheniformis
SUD K2 and Bacillus subtilis SUD K3 showed no significant difference in
thier production of amlyase between Mg^ and Ca^ ions, but these ions
significantly differ in these effects from the rest of the ions.
The rest of the ions, generally had the same effect on all isolates. These
results are generally in agreement with that obtained by Mai et a I (1992) and
Khire and Pant (1993) using Bacillus steorothermophilus and Bacillus sp.64
respectively.
4.3.5. Effect of Different Organic Nitrogen on Amylasc
Production:
Among the different nitrogen sources tried, peptone proved to be the best for
amylase production by Bacillus licheniformis SUD K], SUD K4, SUD O,
resulting in a maximum enzyme specific activity of 5.7, 5.5 and 4.2 U/mg
protein in order (Fig. 21, Fig 24 and Fig 26). Yeast extract, tryptone and
49
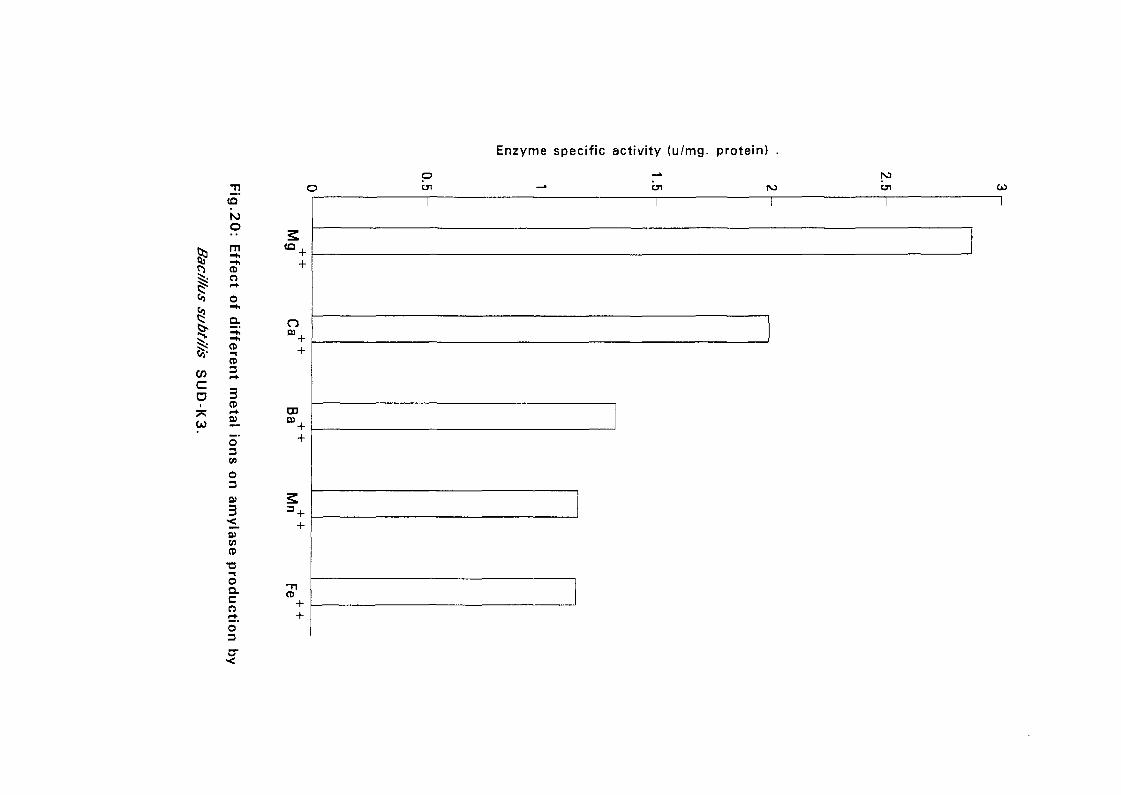
O
Qj -h^ CD5i: O
Enzyme specific activity (u/mg. protein)
pen
I
c
o-hCD
CD3
3CD
o'3V)
o30)
3
CD
0)enCD
OQ .CO
CD
o3

malt extract gave similar results which were significantly lower than the
activity in presence of peptone. On the other hand meat extract gave the
lowest specific activities which were 3.5, 2.9 and 1.6 U/mg protein in order.
These results were generally similar to that obtained by Bajpai el al (1992)
(Bacillus sp. TCRDC-M) and Patel el al (1993) (Halobacterium.halohium).
The effect of organic nitrogen on amylase production by Bacillus
lichenifonnis SUD K2> Bacillus subtilis SUD K? and Bacillus circulans
SUD K7 are presented in Fig 22, Fig 23 and Fig 25 in order. It is observed
that malt extract produced the maximum activity in all species. Their specific
activities were 3.8, 2.9 and 4.3 U/mg protein in order. Peptone showed a
slight decrease in amylase production by Bacillus licheniformis SUD K2 and
Bacillus circulans SUD Kybut there was a clear decrease in case of Bacillus
subtilis SUD K3. Tryptone and meat extract recorded the lowest specific
activities.
Statistical analysis of the effect of organic nitrogen on activity showed a
poor performance of meat extract for all isolates.
Kwan et a! (1993) tested various nitrogenous compounds for production of
P-amylase by Bacillus circulans and reported that a 3%(w/v) beef extract
supported the highest specific activity while l%(w/v) of peptone and yeast
extract supported a good specific activity. Okolo el al (\996) tested the
effect of various organic nitrogenous compounds on amylase production by
Thermoactinomyces thalpophilus F13 and showed that the maximum
enzyme activity was obtained in presence of malt extract as the major
nitrogen source.
50
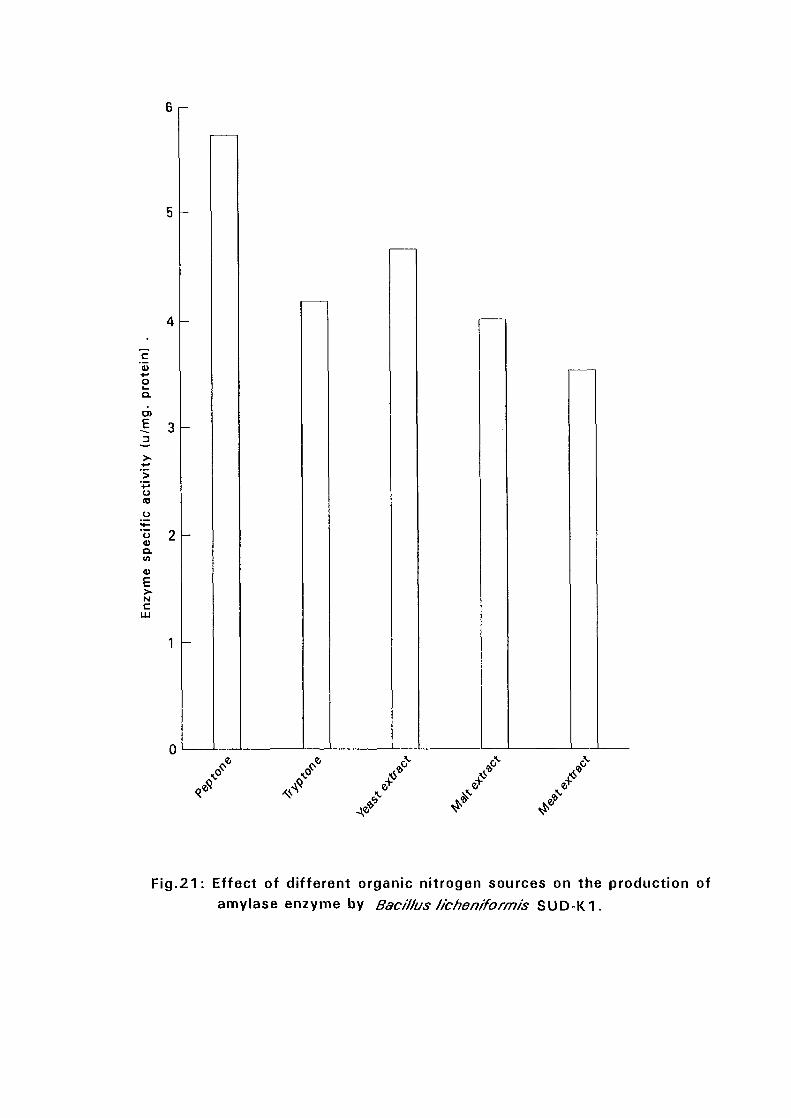
6 i-
cto
>•-»oQ.
E
o
a)E>-N
c
fJ-J
•«•
„ *
Fig.21: Effect of different organic nitrogen sources on the production ofamylase enzyme by Bacillus lie he niform Is SUD-K1.
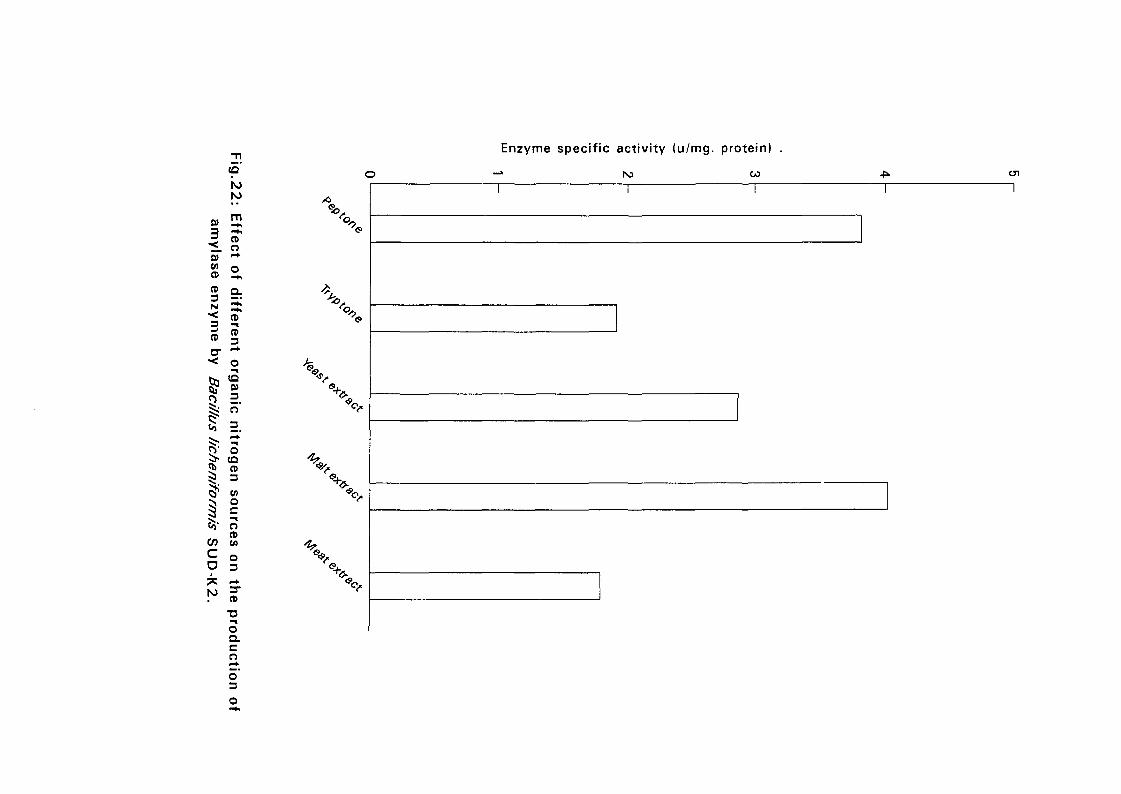
I 31 sCD S ,CD o .
l -3 5CD CB
3
JV O
$ CD
^ =
(J CO
to' o00 Z
s °D 3
^ cT
oQ.C
oo'3
Enzyme specific activity (u/mg. protein)
•» ro co enT T
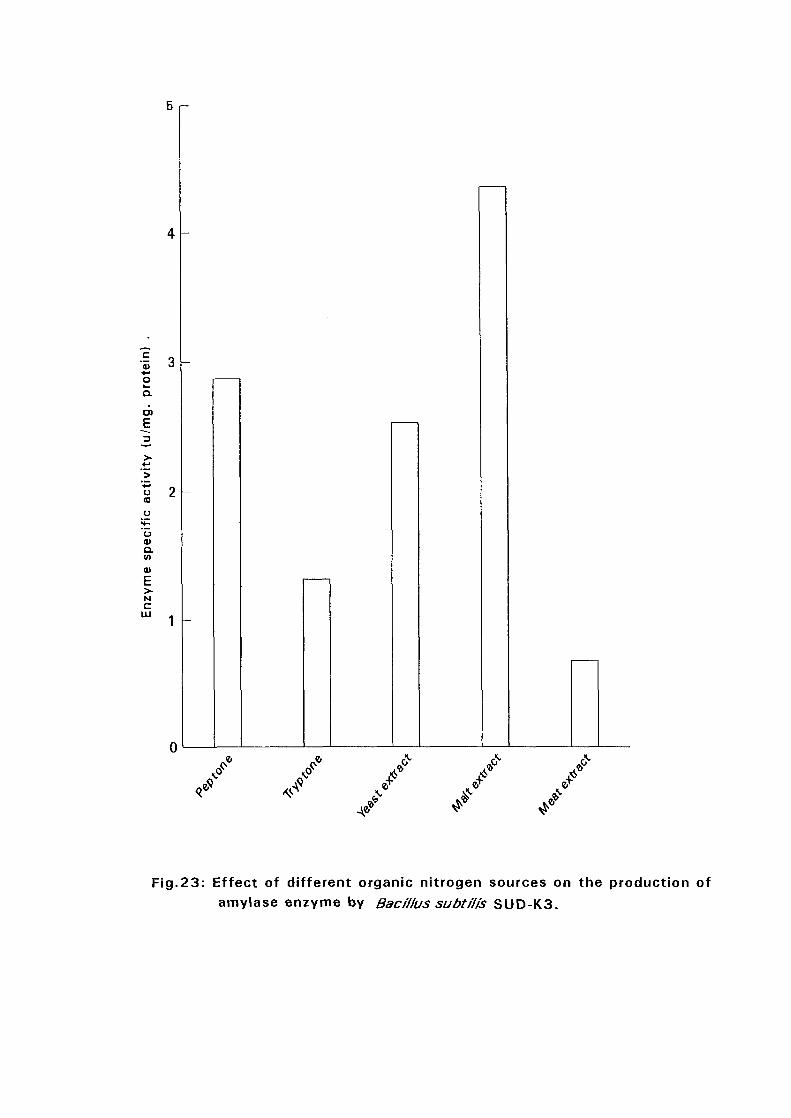
5 i-
<= „"55 •i+-•oLa
aO)
E
U(0
oa.
c
/ / / / .f/ / / / /
Fig.23: Effect of different organic nitrogen sources on the production of
amylase enzyme by BacH/us subtil is SUD-K3.
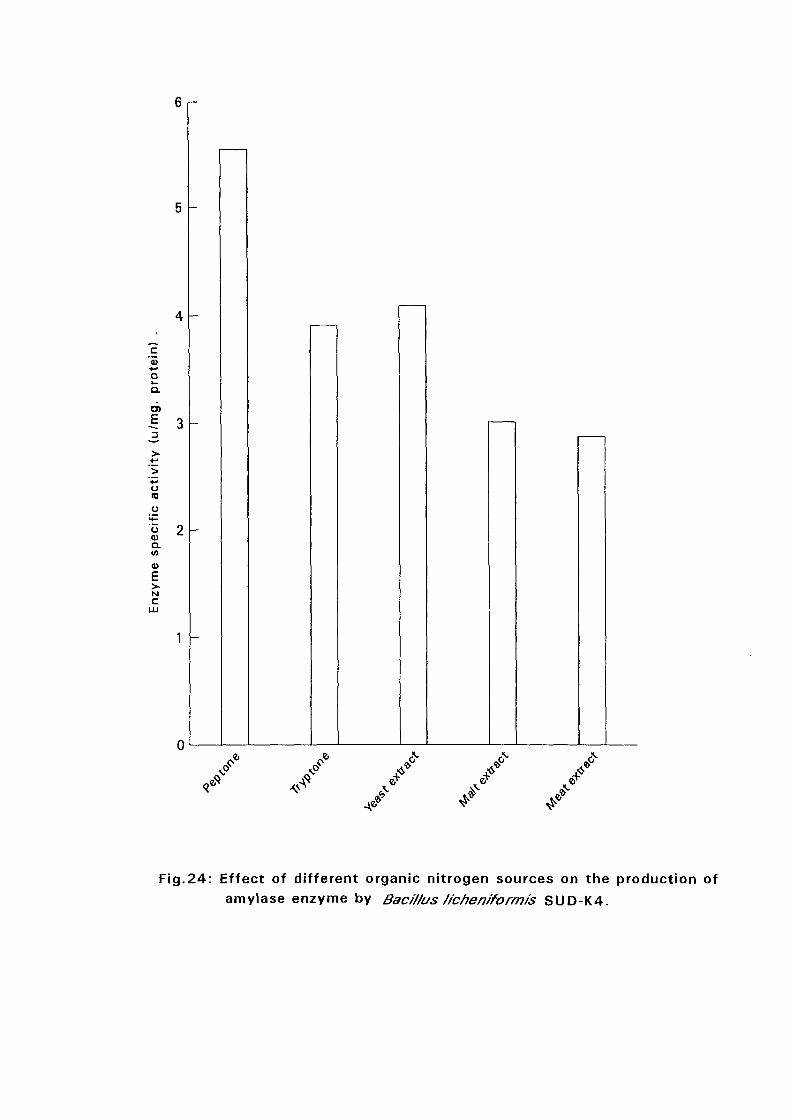
c<o+->oQ .
CT)
E
u(D
a.
Nc
UJ
0
. *
Fig.24: Effect of different organic nitrogen sources on the production of
amylase enzyme by Bac/f/us t/chen/form/s SUD-K4.
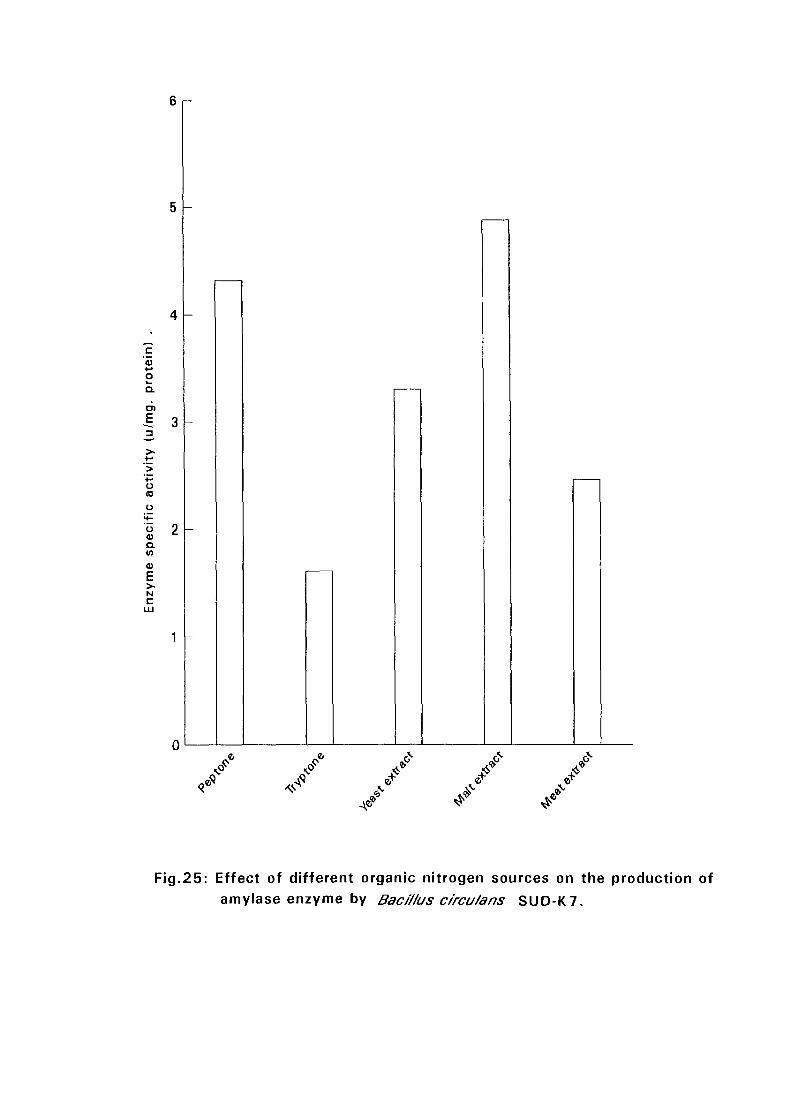
6 r -
oQ.
vo(0
uuQ.V)0)E>.NC
Fig.25: Effect of different organic nitrogen sources on the production ofamylase enzyme by Bacillus circu/ans SUD-K7.
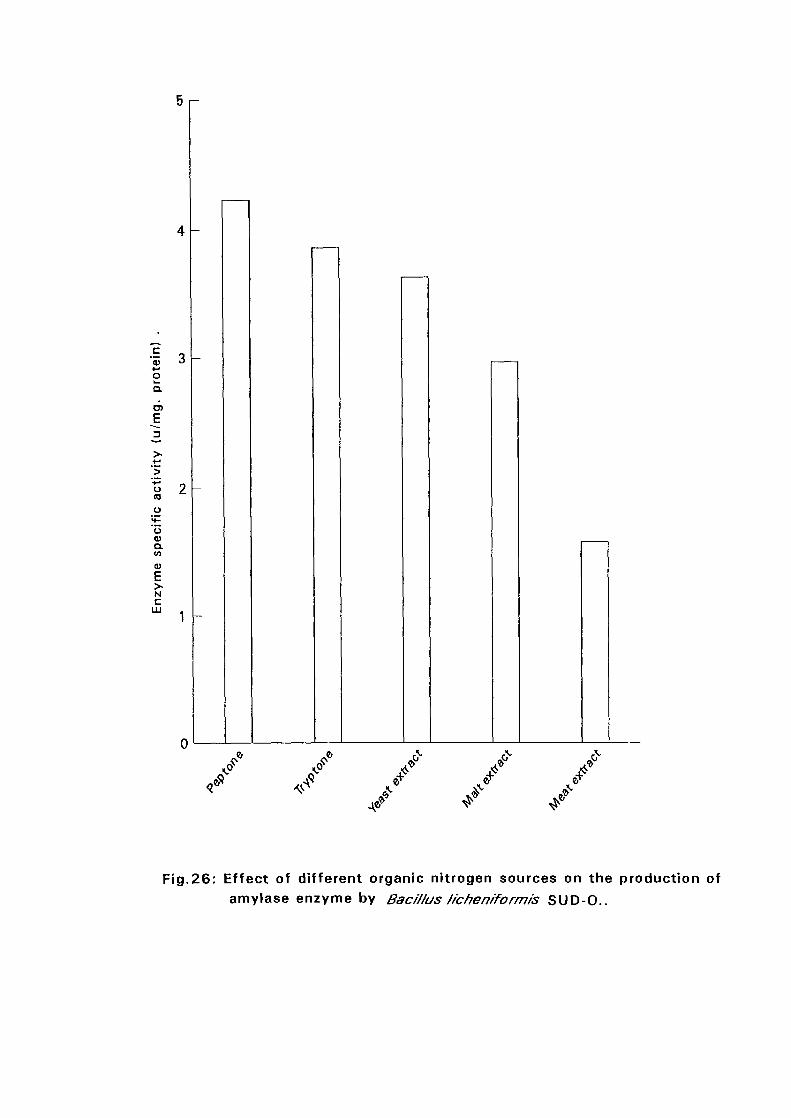
•I 3oa.en
(0
a.CO
N
c
/
Fig.26: Effect of different organic nitrogen sources on the production of
amylase enzyme by Bacillus f/cheniform/'s SUD-O..

4.3.6. Effect of Different Carbon Sources on Amylase
Production:
Different carbon sources in a concentration of 0.5%(w/v) were tested for the
production of amylase by Bacillus lichenifonnis SUD Ki, SUD K2, SUD K4
and Bacillus subtil is SUD K?. For Bacillus lichemformis SUD O and
Bacillus circulans SUD K7 a concentration of 2% was used.
Soluble starch was found to be the best inducer for amylase production by
all isolates (Fig 27-Fig 32). The production was substantially higher than
that reported for Bacillus stearothermophilus (Jaeyong et al, 1989). When
sucrose, fructose, maltose, lactose and glucose were used as growth
substrate there was lower amylase production, although all the isolates
utilized these soluble carbohydrates and grew to approximately the same
extent, than in the medium containing soluble starch. This indicates the
inducible nature of the amylase and that all strains can use carbohydrates
other than starch for their growth. Statistical analysis of the data showed no
significant difference in amylase production by the different isolates using
soluble carbohydrates except when glucose is used. This latter case gave the
lowest specific activity for all isolates. This result agrees with finding of
Yamasaki et al (1977b); All et al (1990) and Achi and Njoka (1992) using
Aspergillus awamori, Aspergillus terms and Bacillus alvei in order, but
differ from the finding of Satio and Yamamoto (1975) and Olukayode et al
(1987) using Bacillus licheniformis and different species of bacteria
respectively. They reported that no significant difference in amylase activity
induced either by glucose or maltose or others polysaccharides.
51
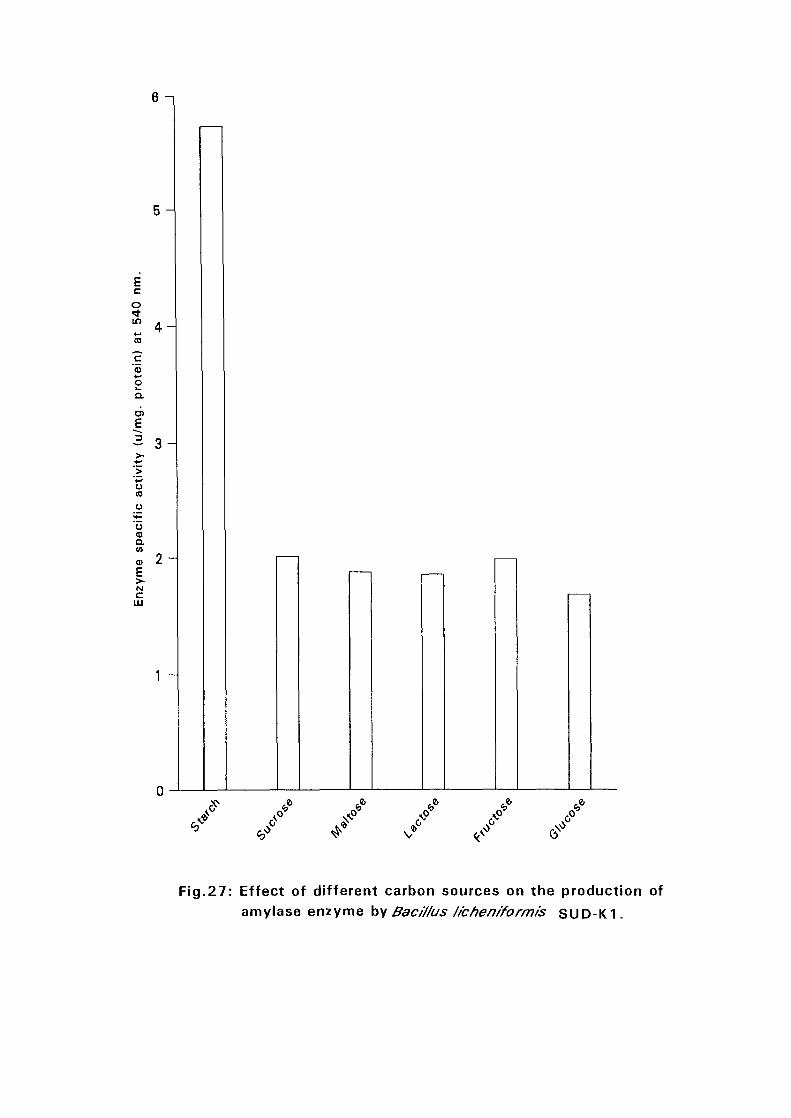
6 - i
5 -
Ecoin
«-*CO"c
oa.
O)
E
4 -
3 -
a(A
a)E>~N
HI
2 -
1 -
v
Fig.27: Effect of different carbon sources on the production ofamylase enzyme by Bac///us /ichen/formis SUD-K1.
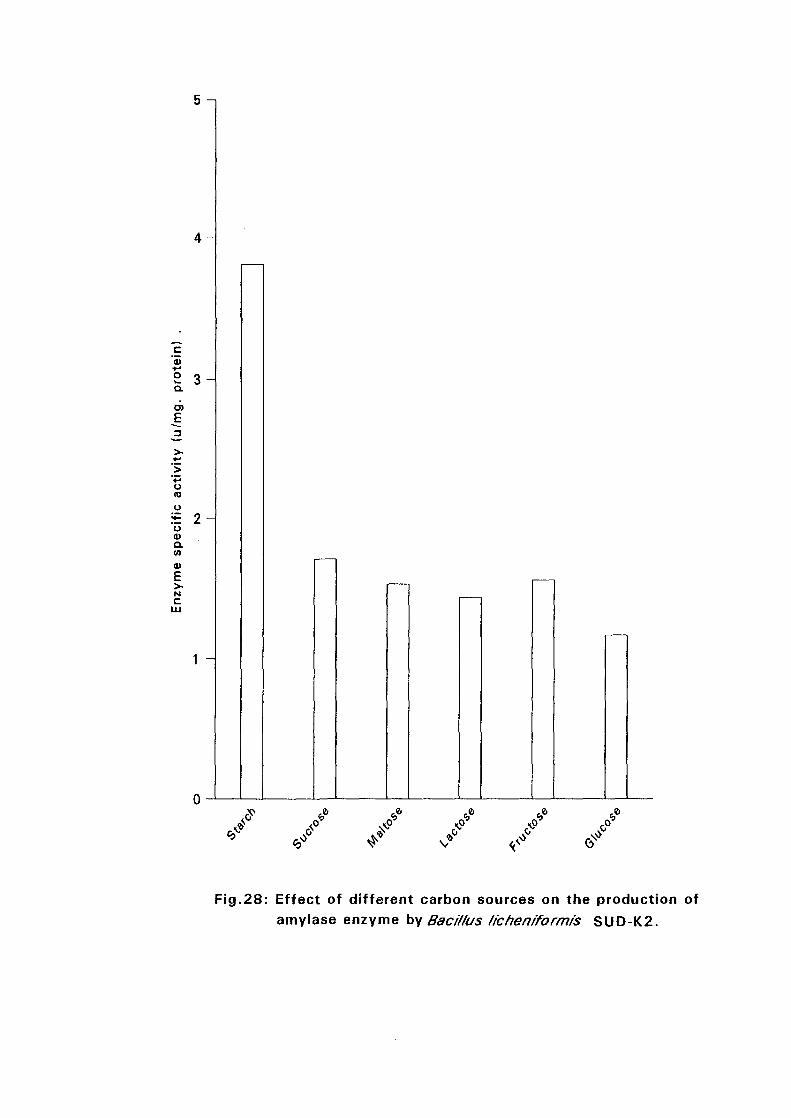
5 -i
c'3
• * -
O o
a 3
a>E
o(0
uE 2u<uaMa>ENC
1 -
* *
Fig.28: Effect of different carbon sources on the production ofamylase enzyme by Baci/fus ficheniformis SUD-K2.

Enzyme specific activity (u/mg. protein)
p
21
up
3•<0)0)CD
CD3N
3CD
CT• <
8"|
1c
00•
mCDOr*
oa.<̂
CD
CD3
O
cro3(AOcoCD(A
O3
3"CD
T3
3acoo'3
%
X^̂*•*s.
ICJ1I
CO
I
toOl
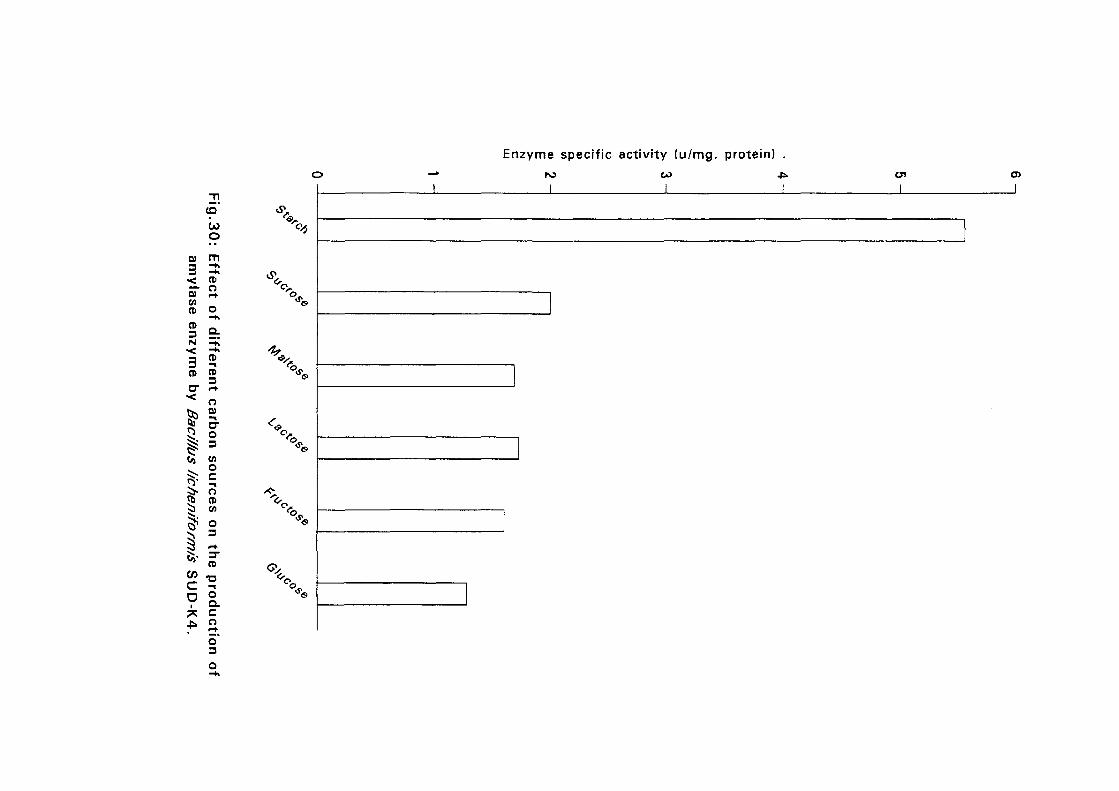
CO
oOJ rn
< CD
s "CD O
s I.3 5CD CDO " <•+* * O
^ D -
3 og 3Co M
Jb CD
C? °
CDto-coc ^O °a.
co
Enzyme specific activity (u/mg. protein) .CO CJI
Ia >
I
o3
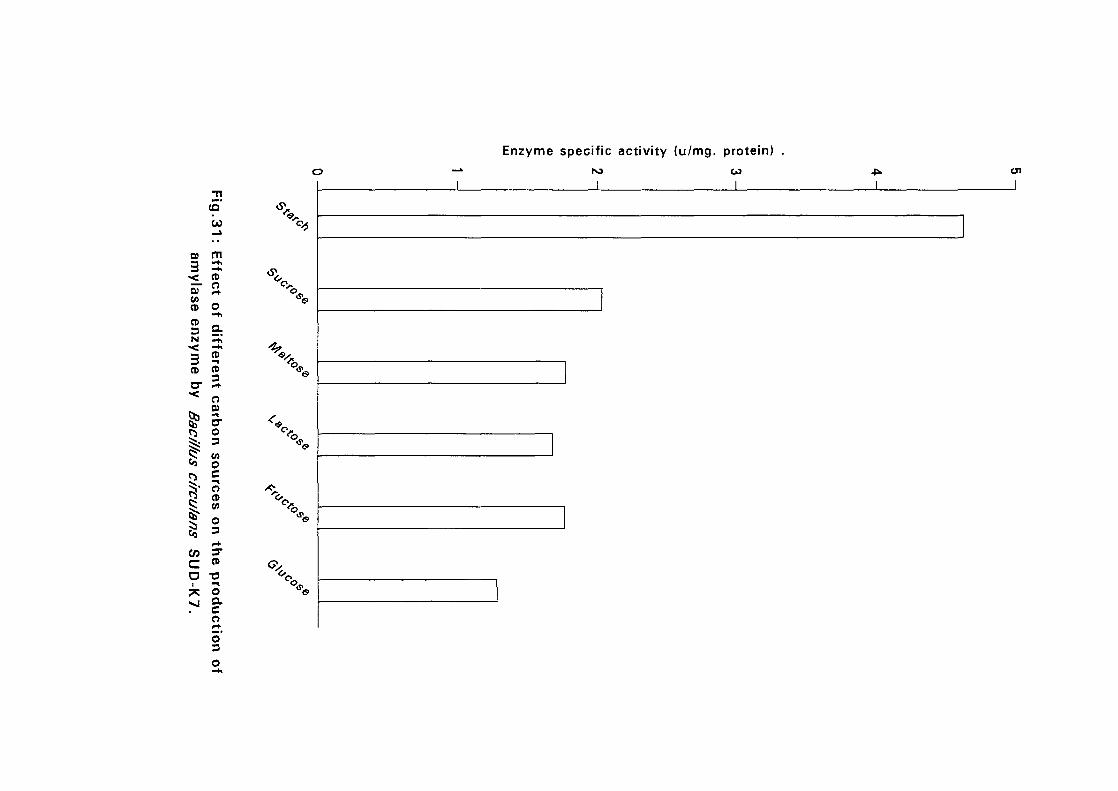
Enzyme specific activity (u/mg. protein)
01
3•<_0)
u>CDenz
3CD
<
t*i
toca
,.31
m—h—hCDOr*O
a
fer
CD3
O01
S-o3CO
ocoCDV)
o3
3"CD"OOacoo'3
ICO
Ion
I
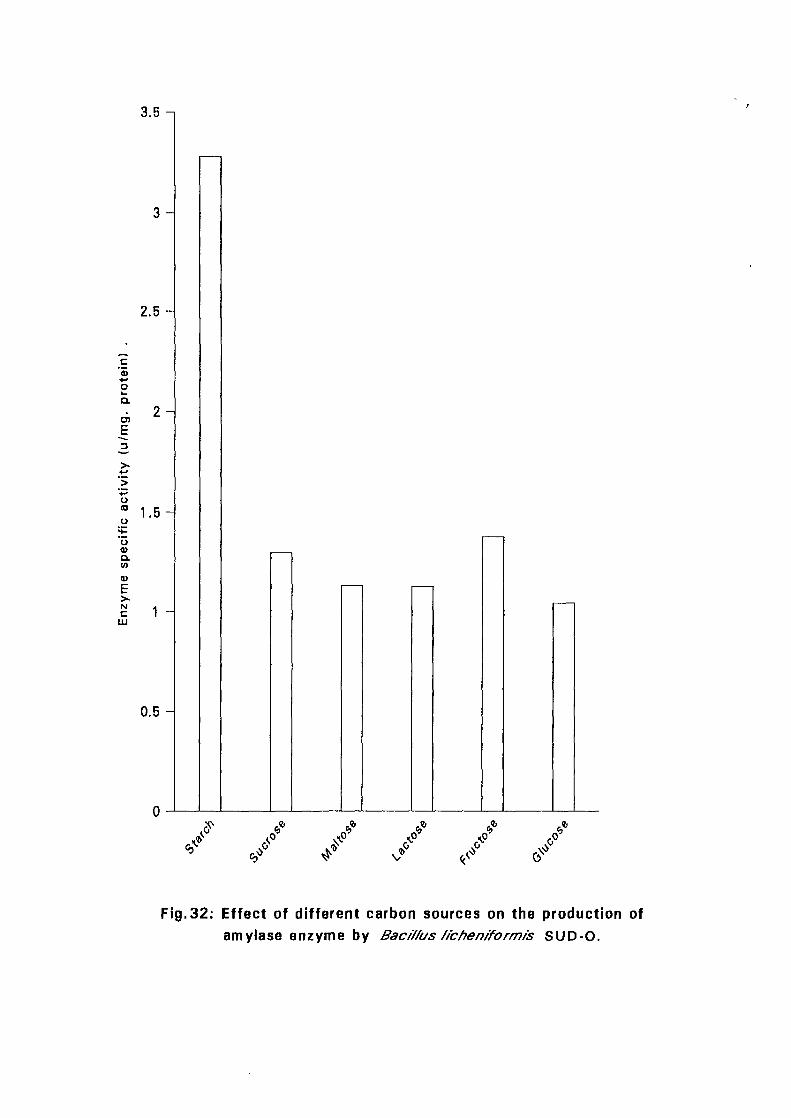
3.5 -i
3 -
2 . 5 -
c'5•-»oa.enE
o
o0)Q.V)
1 . 5 -
Nc
LU
•t _
0.5 -
. 0 *V
Fig.32: Effect of different carbon sources on the production ofamylase enzyme by Bac/7/us //chen/form/s SUD-0.

4.4. Amylase Activity:
4.4.1. Effect of Temperature on Amylase Activity:
Activity measurements of the crude enzyme preparations from the six
Bacillus isolates were carried out over a temperature range of 40°-90°C
using 1% starch in 0.05M phosphate buffer at pH 7.0. The results are
presented in Fig 33a and Fig 33b. It is seen that amylases from Bacillus
licheniformis SUD K2, Bacillus sublilis SUD K3 and Bacillus circulans
SUD K7 are active over a wide range of temperatures (40-90) with a
maximum activity at 60°C (specific activities at 60°C were 4.60, 3.90 and
7.05 U/mg protein in order). At 90°C the enzymes still retained some
activity which were 37.55, 30.55 and 50.5% of the original activity in order.
These results are in agreement with the results reported by Hayashida et al
(1988), Takasaki et al (1991) and Burgess-Cassler and Imam (1991) using
different bacilli. Satio (1973) reported a heat-stable a-amylase from
Bacillus licheniformis 584, but that enzyme lost it's stability rapidly at 76°C.
Bacillus licheniformis SUD Ki , SUDK4 and SUD O gave a maximum
amylase activity at 70°C with specific activities of 7.25, 6.97 and 5.24U/mg
protein in order. The amylases of these isolates were active up to 90°C with
residual activity of only 42.8%, 40.55% and 30.4% in order. Welker and
Campbell (1967) reported that the amylases from the thermophilic Bacillus
amyloliquefaciens and Bacillus sublilis has a maximum activity at 65°C.
Morgan and Priest (1981) reported an amylase from Bacillus licheniformis
NCSB 6346 with maximum activity at a higher temperature (85°C) than the
amylases from several thermophilic bacilli including Bacillus coagulans
(Campbell, 1955), Bacillus stearothermophilus (Ogashara et al 1970) and
Bacillus caldolyticus (Heinen and Heinen, 1972), all of which showed a
maximum activity at 70°C. Mai et al (1992), Khire and Pant (1993) and
52
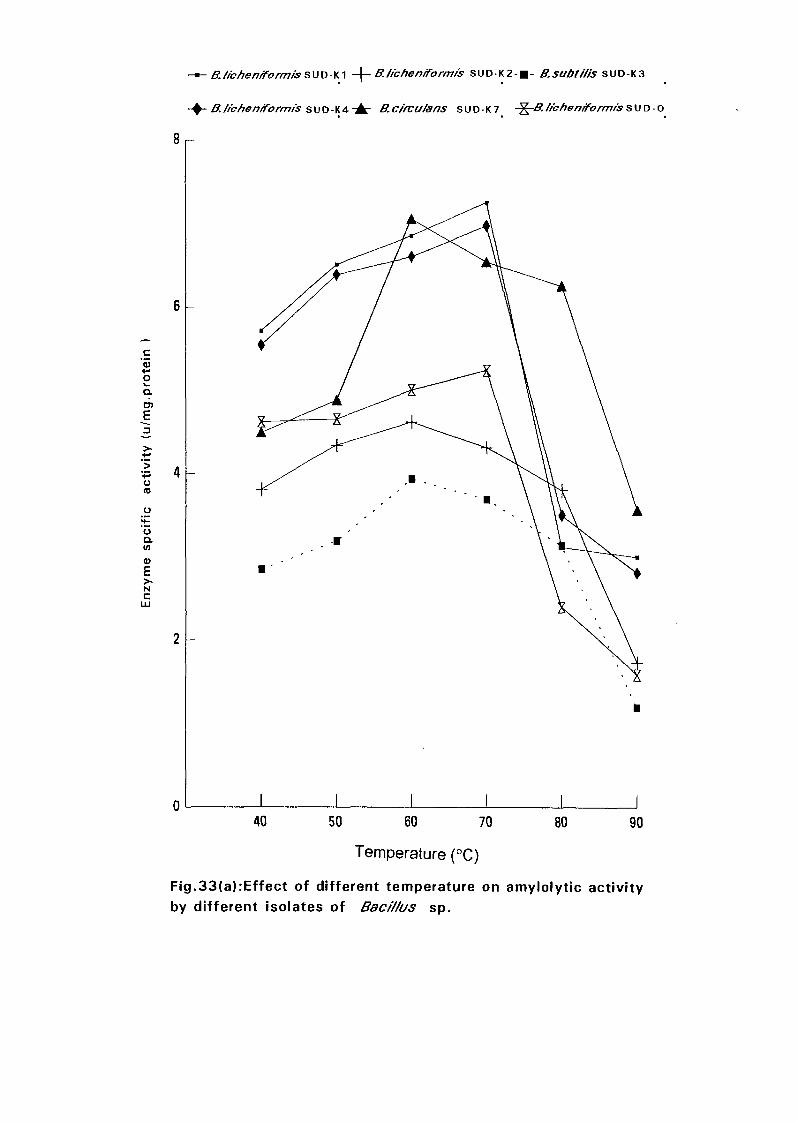
B./ichenfformisSUD-K1 —|— B./ichenfformis SUD-K2-H- B.subti/is SUD-K3
B./ichenfformis suD-K4-Ar B.cfrcu/ans sUD-K7 -%-B./ichenfformisSUD-O
f
oQ.
OQ.inD
ENc
LU
40 50 60 70
Temperature (°C)
80 90
Fig.33(a):Effect of different temperature on amylolytic activityby different isolates of Bacf/fus sp.
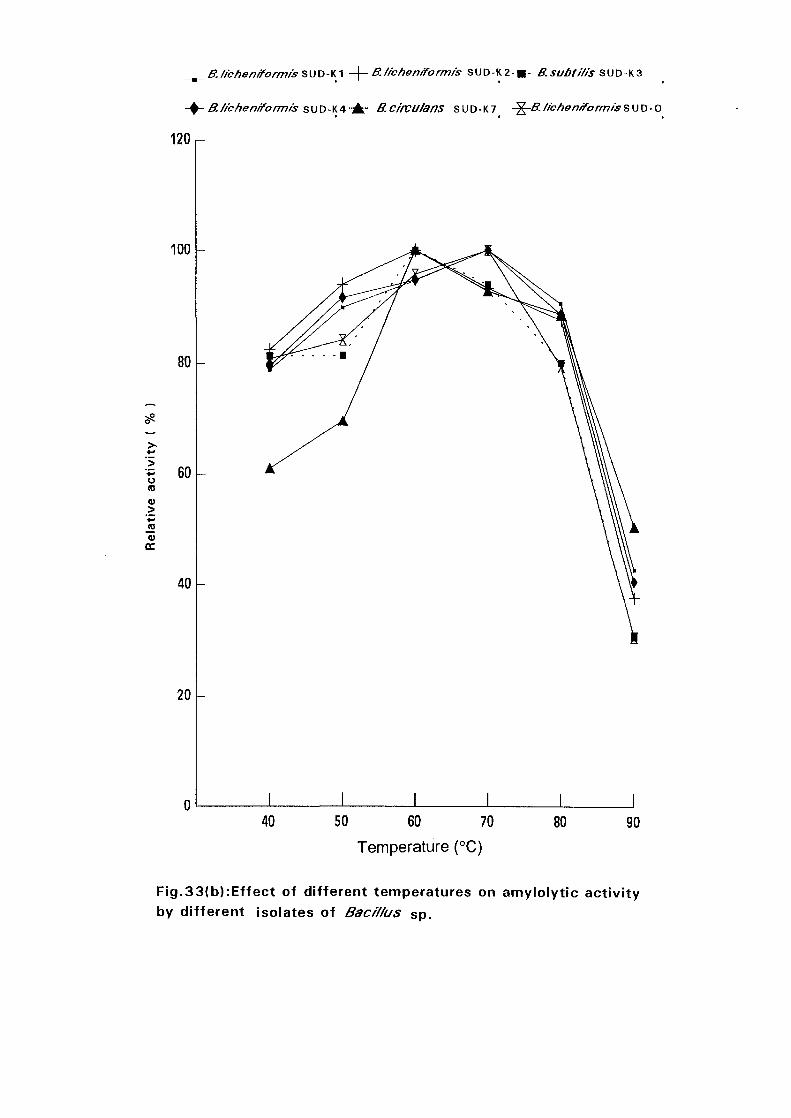
B./tchenfformis SUD-K1 -\- B./ichenfformis SUD-K2-H- B.subti/is SUD-K3
• B./ichenrformfs SUD-K4-A" B.Circufans SUD-K7 -%-&• /f'chenrformisSUD-0
120
100
80
O60
40
20
40 50 60 70
Temperature (°C)
80 90
Fig.33(b):Effect of different temperatures on amylolytic activity
by different isolates of Baci/fus sp.

Goyal et al (1995) also showed a heat stable amylase enzyme of different
bacilli with maximum activity at 70°C.
4.4.2. Effect of pH on Amylase Activity:
pH profiles of the amylase activity for all six isolates is shown in Fig. 34a
and Fig. 34b. It is seen that all the isolates were found to be active over a
wide range of pH (4-10). Morgan and Priest (1981) reported maximum
activity of Bacillus lichemformis between pH 7.0 and 9.0 and a marked
reduction at lower and higher pHs was observed.
Amylases produced by the tested isolates in this study, seems to be generally
similar in their pH activity profiles to these reported by Satio and Yamamoto
(1975) and Chiang et al (1979). However, they differ from the enzymes of
Bacillus amyloliquefaciens and Bacillus subtilis which possess maximum
activity at pH 6.5 and 5.5 respectively. Welker and Campbell; 1967 and
Fogarty; 1983, reported that amylases from bacilli were found to be active in
the pH range of 5-8.
4.4.3. Effect of Different Time Intervals on Amylase Activity:
The effect of different incubation periods on amylase activity is presented in
Fig. 35a and Fig. 35b.
Dextrinization or saccharolytic activity was calculated as U/mg protein/10
min, /20 min., ,/60 min.
Amylases of all isolates showed an increase in specific activity up to 1 hour.
When the activity was calculated as per minute it is seen that the activity
showed gradual decrease (Fig. 35b); the rate of the reaction is highest at the
start of the reaction and it gradually decreases. This is quite a common
phenomenon in enzyme kinetics and it is almost certain that it is due to
product inhibition of the enzyme activity. This result would indicate that if
53
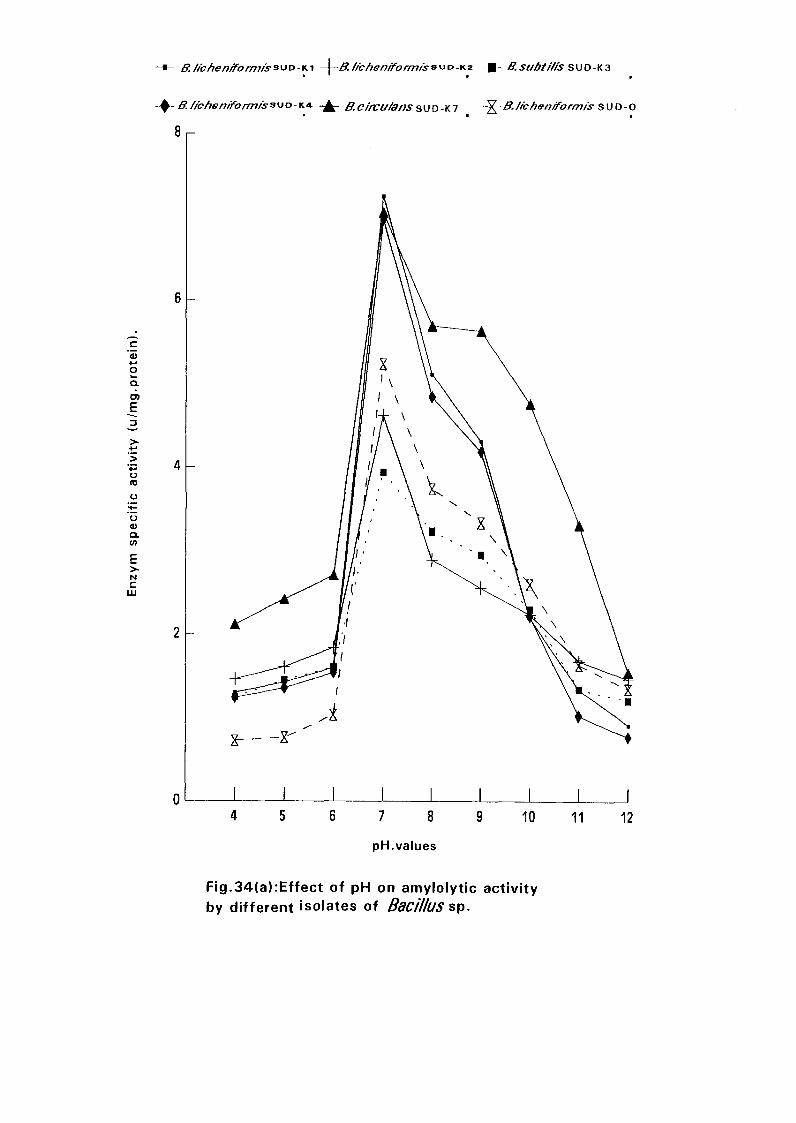
£.//b/re/7/jfo/777/s3UD-Ki B.subti/is SUD-K3
B./fcften/Form/s3uo-K4 - ^ - B.c/rcu/ansSUD-K7 -^B.//c/ien/form/s s\JO-o
c'55+-»owQ.6)
oto
o0)Q.V)
Nc
LLJ
7 8
pH.values
10 11
Fig.34(a):Effect of pH on amylolytic activityby different isolates of Bacillus sp.
12
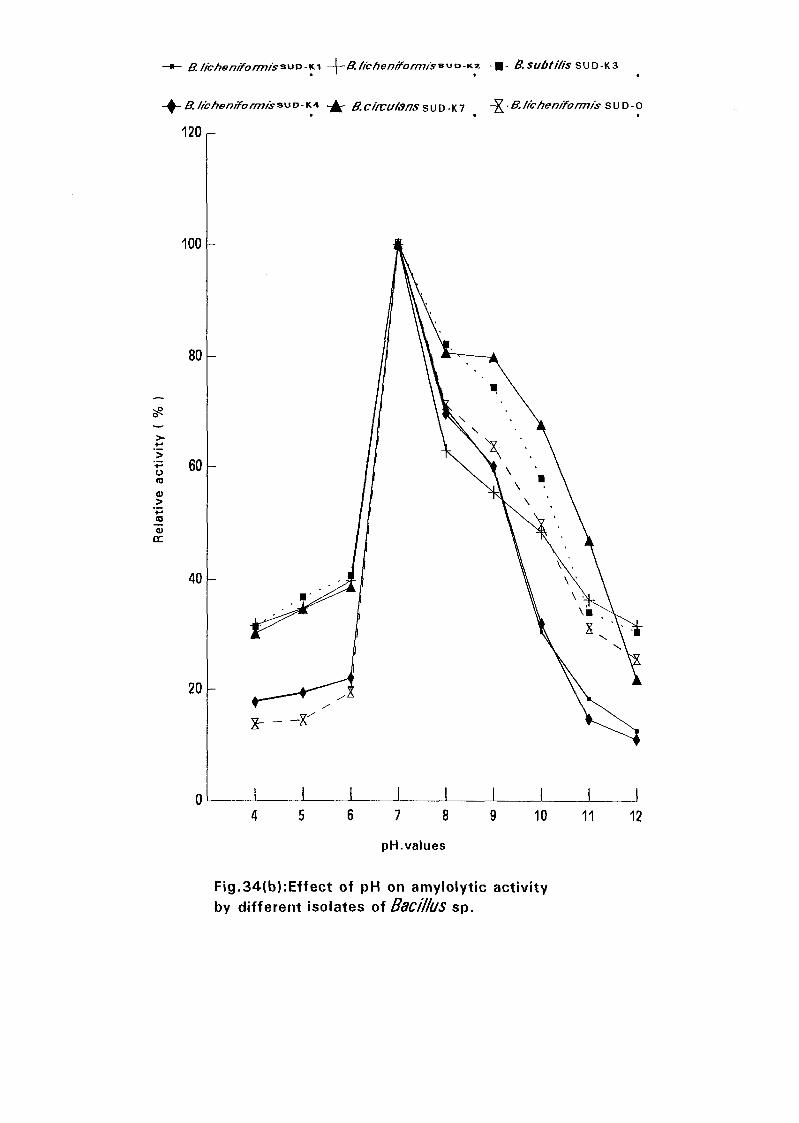
-Ki -\~B./f - B.subti/is SUD-K3
B.//c/ien/f0rm/s3\jD-K4 -fa- B.Circu/ansSUD-K7 ~X'BJ/cheniform/s SUD-0• • •
120 ^
100
80
\ 60(a(D
cc
40
20
6 7 8
pH.values
10 11
Fig.34(b):Effect of pH on amylolytic activityby different isolates of Bacillus sp.
12
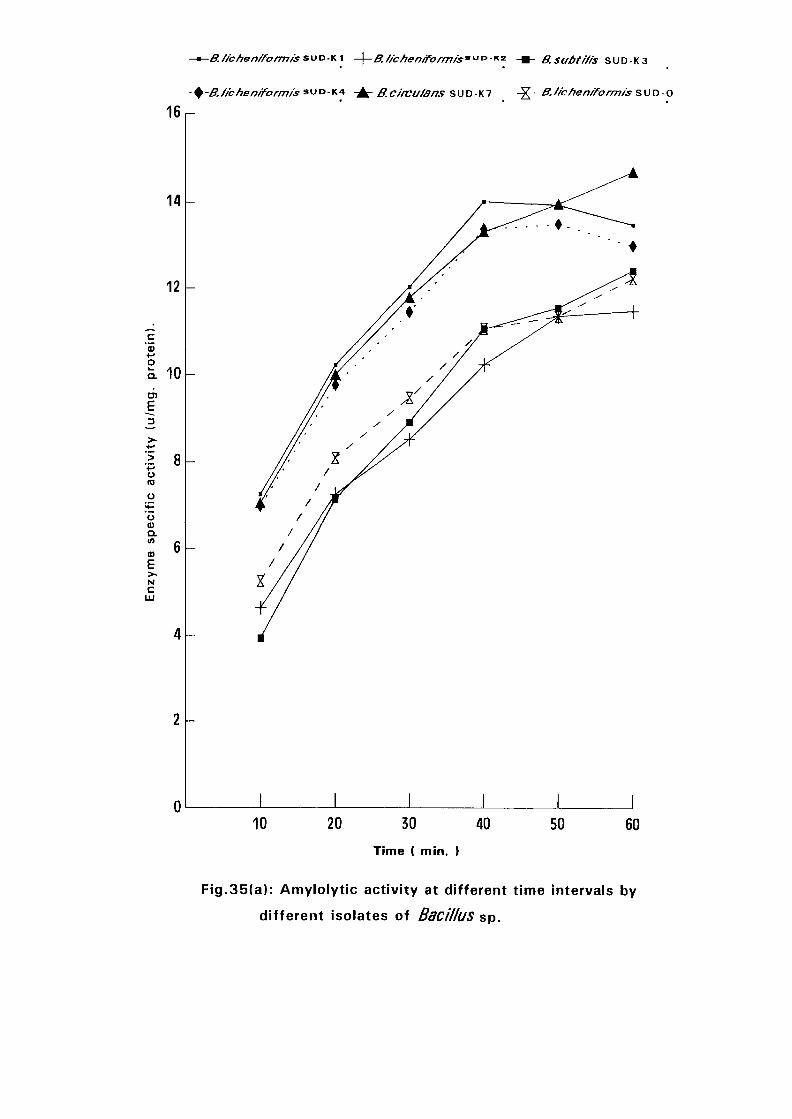
—0.//cf?en/fcir/T7fs&vo-*.i -\-BJ/chenff:ormis9Vi0-v-'2- -m- B.subtr/is SUD-K3
-B./icheni'formis suD-K4 -±- g_circu/ans SUD-K7 -X" &• tichen/formis SUD-O
16
14
12
c'53oa 10
> 8u(0
oa>o.V)
0)E>-Nc
LU
10 20 30 40
Time ( min. )
50 60
Fig.35(a): Amylolytic activity at different time intervals by
different isolates of Bacillus sp.
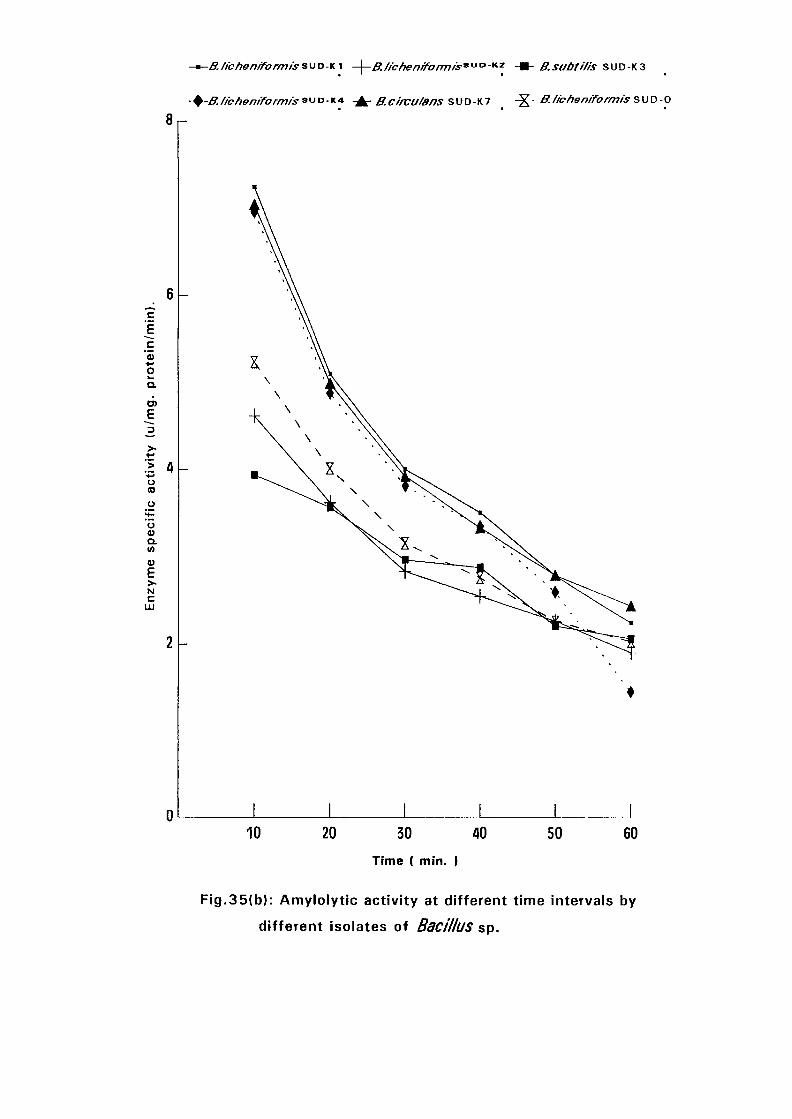
-*-B./fchenffonnisSUDKI -\-B.//c/re/7/fo/777/s"tJOK2 -Wh B.subti/is SUD-K3
-+-B.//cfien/form/s SUD-K* - ^ B.c/rvufans SUD-K7 -X B./rcften/form/s s\)D-Q
8r-
«oQ.
O
U0)
aM
Nc
LU
110 20 30 40
Time ( min. I
50 60
Fig.35{b): Amylolytic activity at different time intervals by
different isolates of Bacillus sp.

that if the enzyme is used in a batch wise manner, the reaction would slow
with time and might stop before complete hydrolysis of the starch. It, also,
indicates that the enzyme is best be used in an immobilised form to
overcome this problem. Similar results was reported by Ogashara et a\
(1970) using Bacillus slearofherniophilus and by Medda and Chandra
(1980) using Bacillus licheniformis and Bacillus coagulans.
4.4.4. Effect of Substrate Concentration on Amylasc
Activity:
The effect of substrate concentration on amylase activity is shown in Fig. 36.
All the isolates showed maximum amylase activity at 1% soluble starch.
Above this concentration no change in the enzyme activity was detected
indicating enzyme saturation. This result generally agrees with the finding oi'
Pestana and Castillo (1985) and Kanno(1986) who tested the effect of
substrate concentration on amylase activity by Aspergilius awamori and
Bacillus acidocaldarius respectively.
4.4.5. Effect of Different Metal Ions on Ainylase
Activity:
The effect of different metal ions on amylase activity is shown in Fig. 37-42.
All the metal ions showed an inhibitory effect on amylase activity except
Mn++ which had little or no stimulatory effect on amylase activity of
Bacillus licheniformis SUD Kl. For the rest of the isolates Mn4"1" ions
showed a very slight decrease in amylolytic activity when compared with the
control. However, in case of Bacillus licheniformis SUD O there was a
marked inhibitory effect by Mn++. It is concluded that amylase of the
54
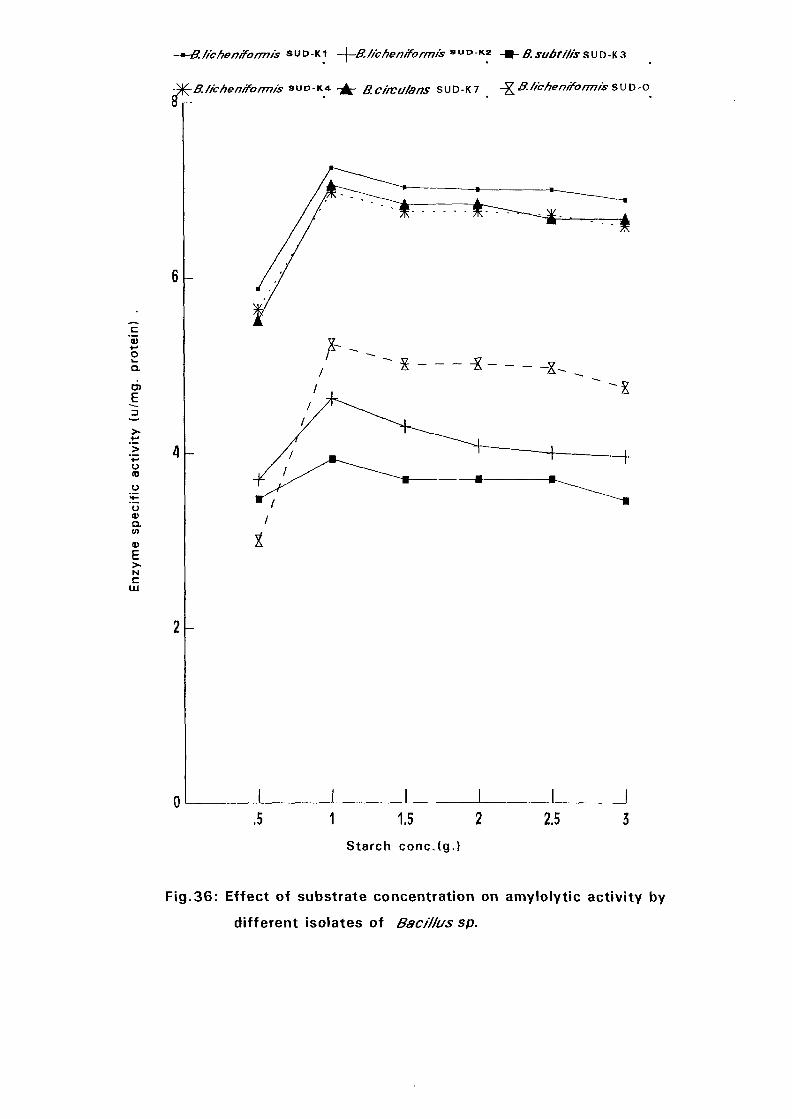
c'55+-'o
a
E
oCO
Ooa>a.tn
a>E>~Nc
LU
-*-B./fehenrformrs SUD-KI -\-BJfchenrformis
-^BJ/chenfformis SUD-K* - ^ B.Cf'fVU/ans SUD-K7 -X-S/fchenrformis SUD-0
.5 1 1.5 2
Starch conc.(g.)
2.5
Fig.36: Effect of substrate concentration on amylolytic activity by
different isolates of Bacillus sp.
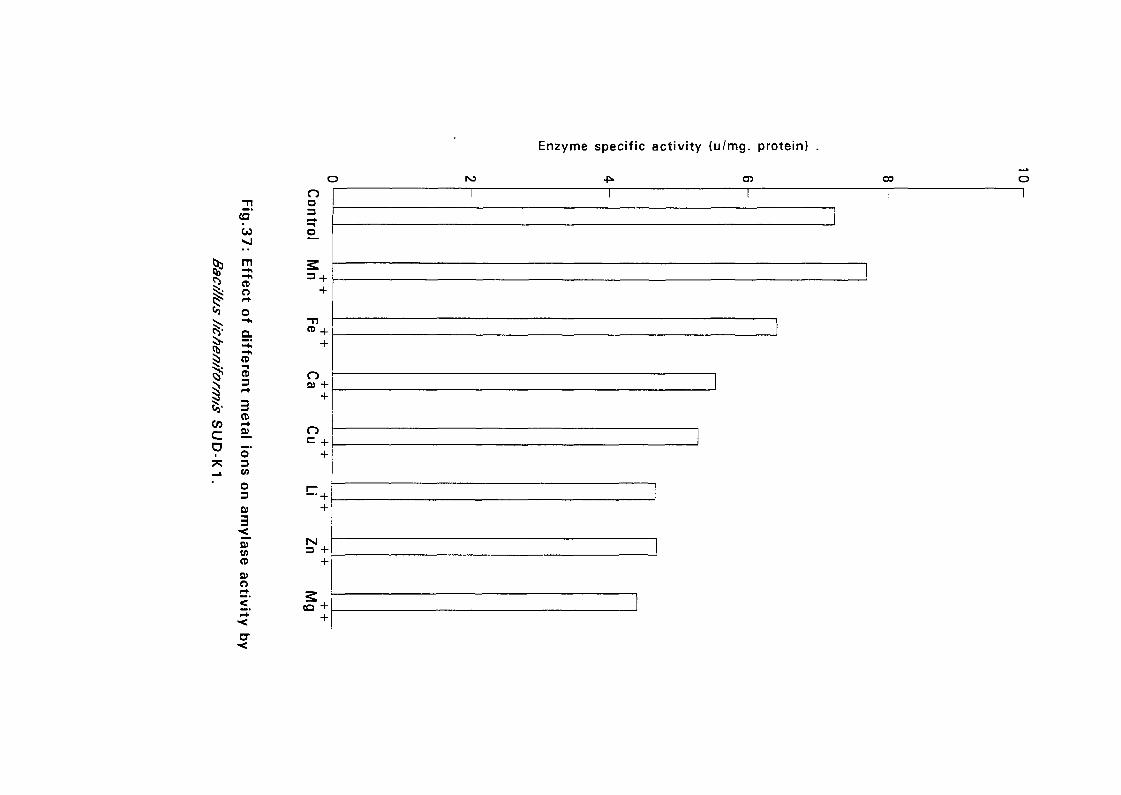
Enzyme specific activity (u/mg. protein)
00
IICOcD
CD
W
m- h-+iCDO
oQ.
(D"i
CD3
3CD
5"3CA
o30)|
5TCftCD0)O
oo
nta +
O+
—+
++
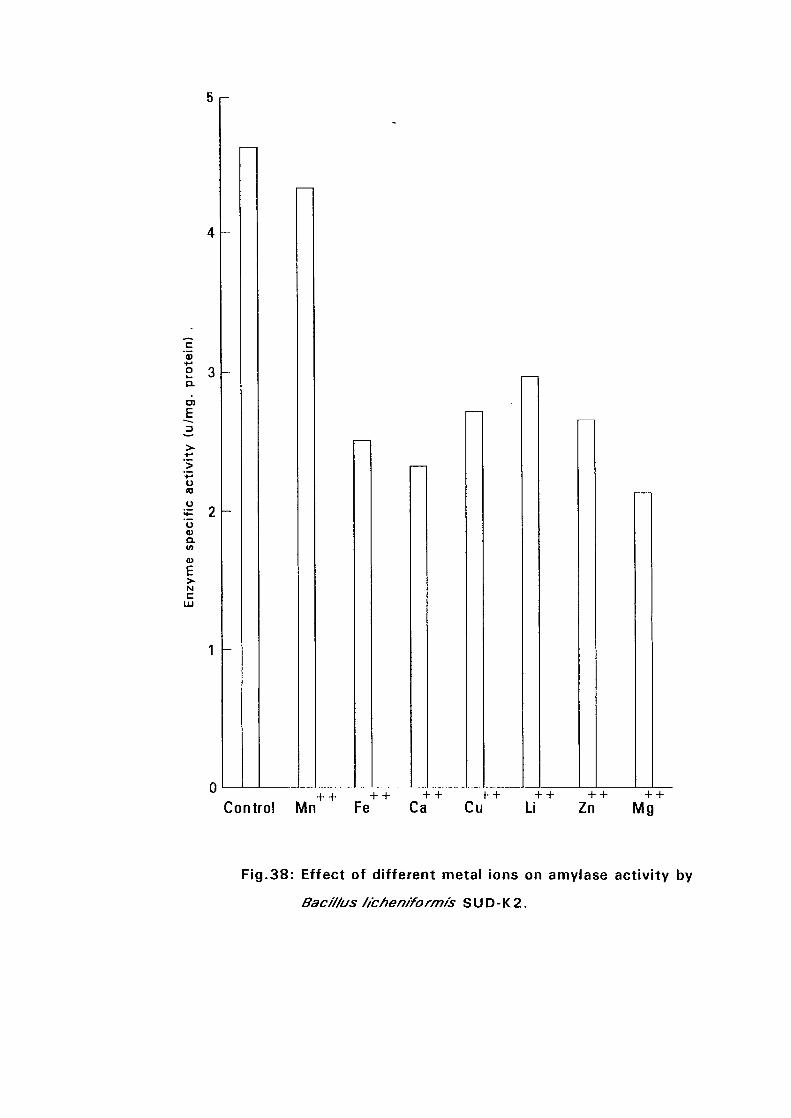
5 i -
c'53•-»
S 3a.b>E
>•
• >
u(D
I 2u<u
Q.
0>
EN
c
Control Mn Fe Ca Cu Li Zn Mg
Fig.38: Effect of different metal ions on amylase activity by
Bac///us //cheniform/'s S U D - K 2.
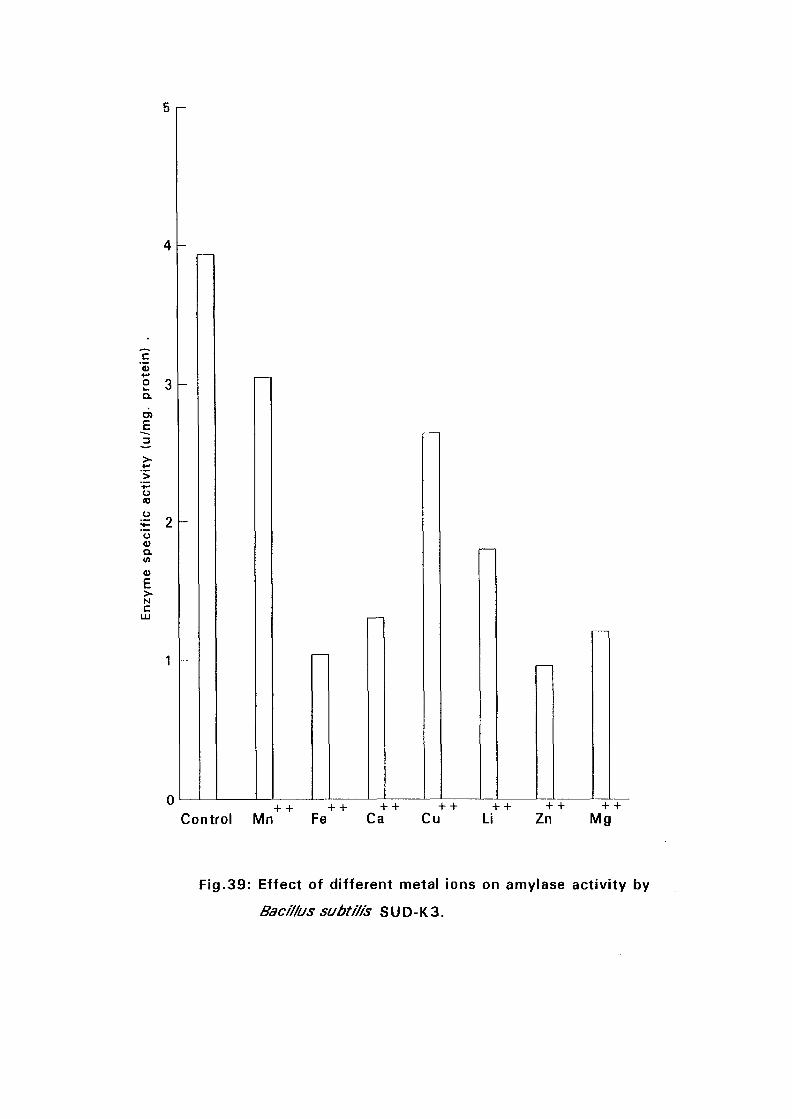
+
S 3Q.
E
E 2
o
Eua)o.V)
4)
E>~Nc
Control Mn Fe Ca Cu Li Zn Mg
Fig.39: Effect of different metal ions on amylase activity by
Bacillus subti/is S U D - K 3.
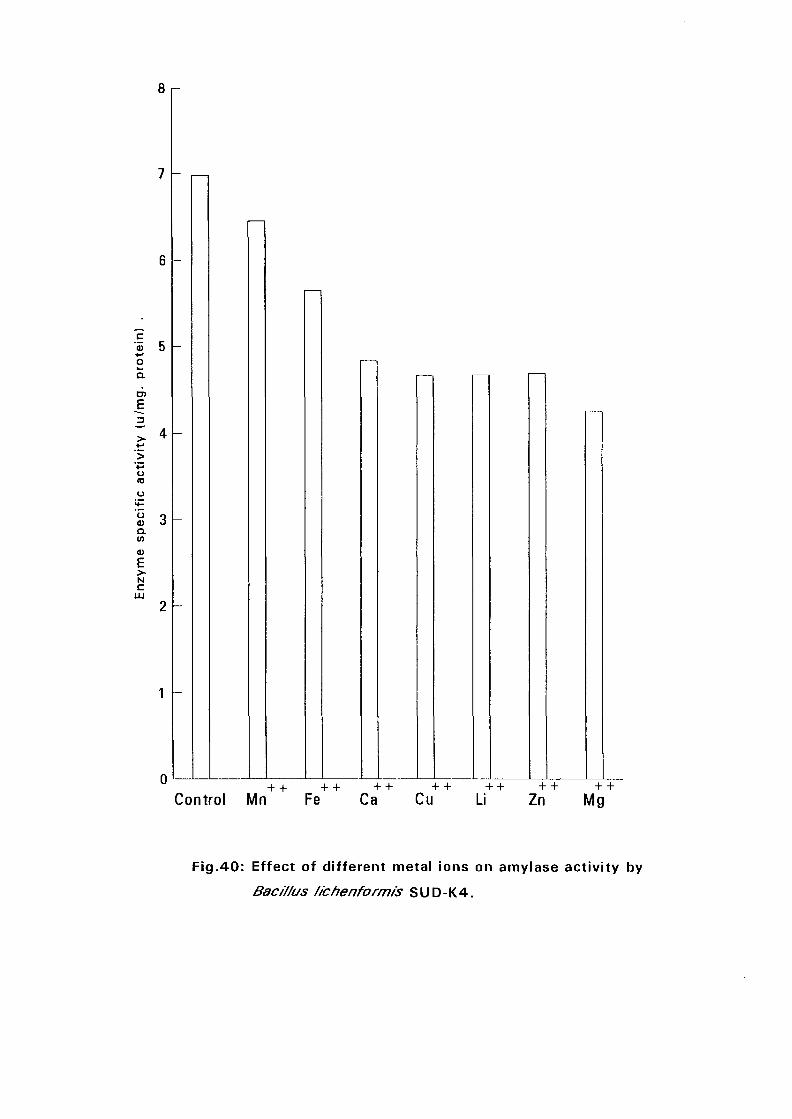
c• 5+-•oQ.
D)
uto
ud>Q .(A
93
E>.NC
0+ + ++ ++ ++ ++ ++ +
Control Mn Fe Ca Cu Li Zn Mg
Fig.40: Effect of different metal ions on amylase activity by
Bac/7/us //chenformis SUD-K4.
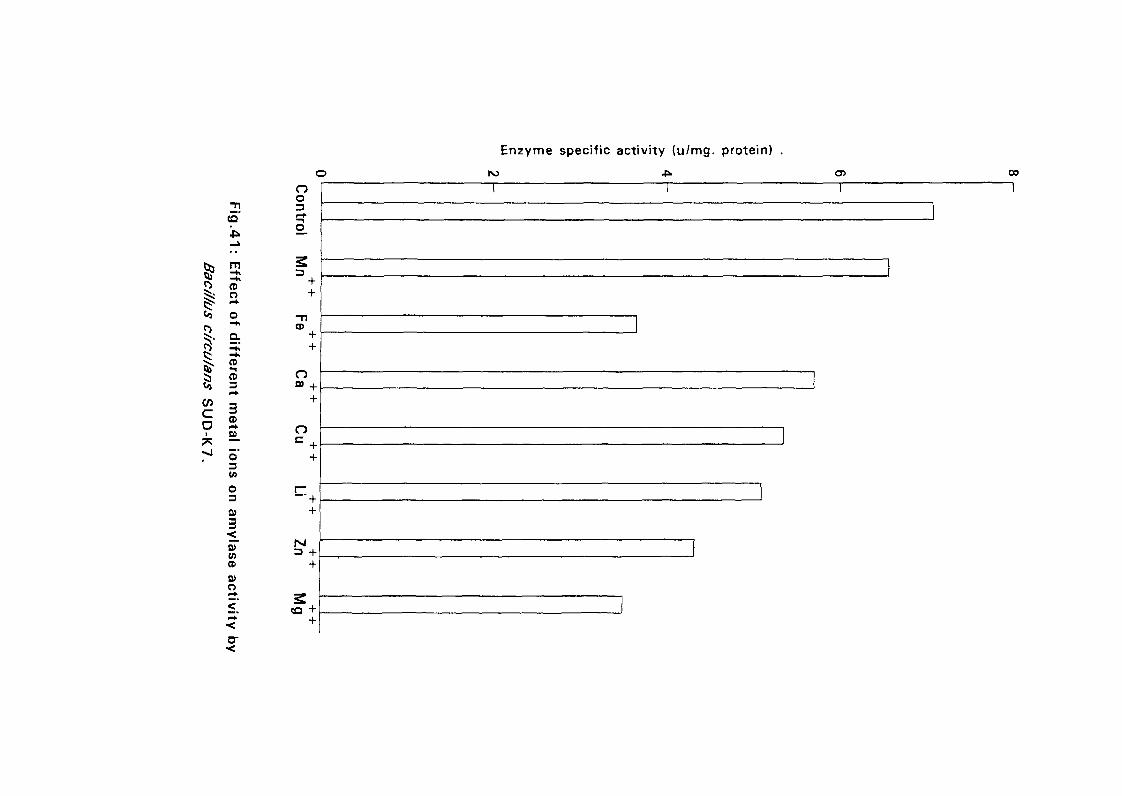
to
oo3
Enzyme specific activity (u/mg. protein)
r>o oo
mCD
o++
CD
rco
CD
§ CD« 3
3CD»•+
o3V>
o3
++
o
o++
:
Q>CACD
0)O
rsi
:
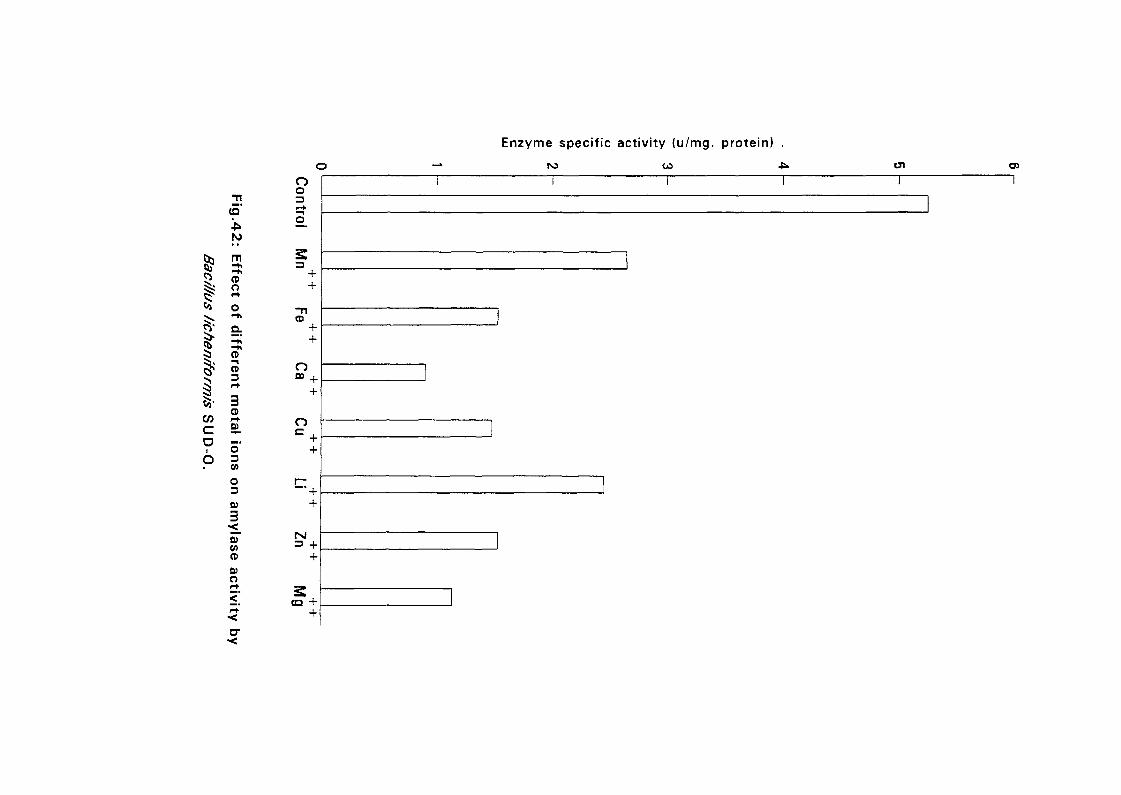
Enzyme specific activity (u/mg. protein) .
•VL
CD*
OJ
oo
N)
m-*CDO
CD
65'CO
c
CD
CD3
3CD
ao3coo3fi)
3COCD
0)O
Oas +
o
+
CD +

different isolate does not show any dependence on metal ion for their
activity.
Yamaski and Kanno (1991) tested various metal ions on glucoamylase
activity of Mucor javanicus and reported that Ca**, Cu**, Mg** and Mn**
had no effect on amylase activity. At the same time Jin et al (1992) using
Bacillus sp. JF strain showed that Ca1 + and Mg++ ions didn't affect enzyme
activity, but addition of Fe++, Cu++, Zn++ and Ag** ions resulted in inhibition
of activity. Khire and Pant (1993) tested the effect of Zn^, Mg"", Mil**
Co*"1", Cu^, Fe**, Ca"", Li"" and Sr^ions on the amylase of Bacillus sp. 64
and reported that all these metal ions had no effect except Fe** and Co**
which had a stimulatory effect and Cu** which had an inhibitory effect. Ilori
et al (1995), showed that Ca**, Mg**, Na+ and K+increased the amylase
activity from Lactobacillns brevis when applied at a concentration of 50, 30,
20 and 5 mM in order.
4.4.6. Effect of NaCl on Amylase Activity:
Results of the effect of NaCl on amylase activity from the different isolates
is shown in Fig. 43. It is seen that there was a reduction in enzymes
activities at all concentrations of NaCl, although this inhibition varies from
one enzyme to other.
Moseley and Keay(1970) using Bacillus subtilis NRRL B3411 found similar
result. However Khire and Pant (1993) reported that the amylase from
Bacillus sp. 64 was stable in NaCl with 80% of the original activity being
retained at 5 M concentration of NaCl.
4.5. Identification of Enzyme Product:
The products of starch hydrolysis by amylase enzyme of the six bacilli were
identified by thin layer chromatography (TLC). The results are shown in Fig.
55
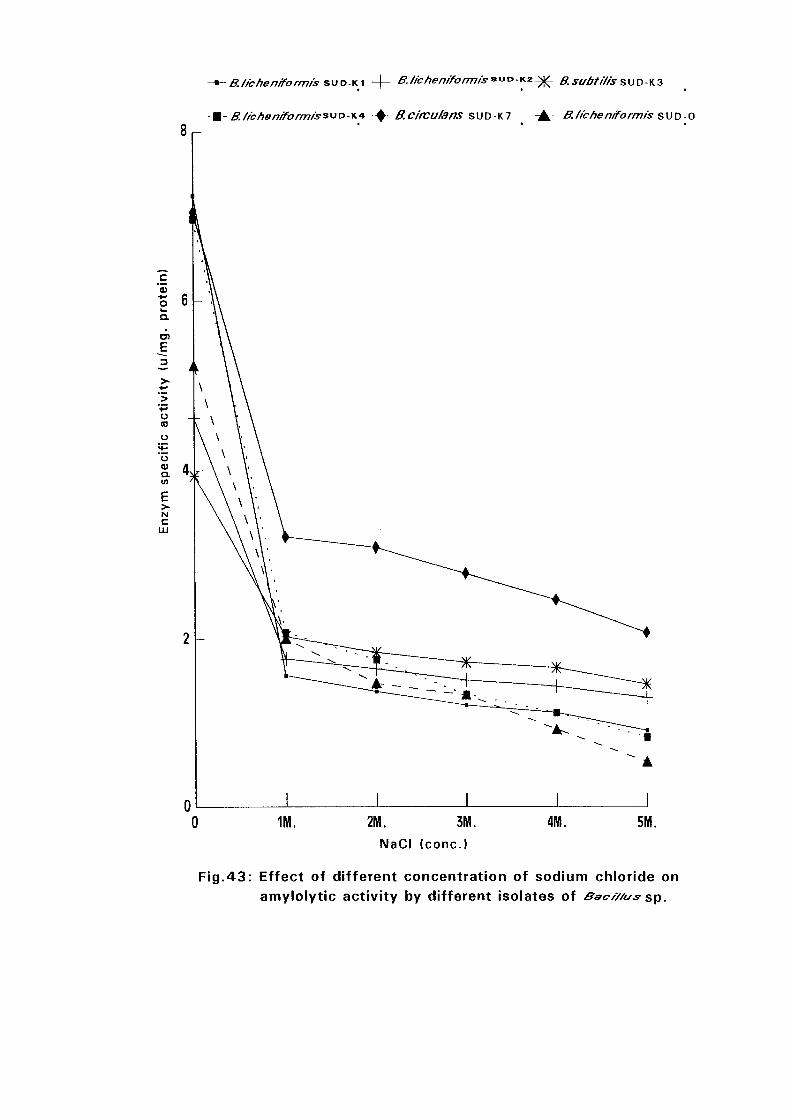
<- B./fchenfformts S U D K I -\
I- B./ichenfformis*uo-*** -+ - B.Circu/ans SUD-K7
B.subti/is SUD-K3
B./ichenfformis suD-0
S 6 -
.E
uCO
o£ 4vU
Nc
UJ
2M. 3M.NaCI (cone.)
4M. 5NI.
Fig.43: Effect of different concentration of sodium chloride on
amyloiytic activity by different isolates of Bac///us s\>.

44. The degradation products of amylase produced by Bacillus licheniformis
SUD K,, SUD K4, SUD O and Bacillus subfilis SUD K3 were mainly
oligosaccharides (identified by comparison with published data) and maltose
(identified by comparison with authentic marker), Bacillus licheniformis
SUD K2 and Bacillus circulans SUD K7 produced oligosaccharides, maltose
and glucose. It has been shown that a-amylases products are dextrins only
or dextrin and maltose and sometimes, dextrine maltose and glucose ( Bird
et al, 1954). Accordingly, all the amylases isolated in this study could be
identified as a-amylases. In this respect these results conform with the data
published earlier (Mastuzaki et al 1974; Yamane and Maruo, 1974 and
Michelena and Castillo. 1984). However it differs from the results published
by Me Wealthy and Hartman (1977) who reported that a-amylases from
Bacteriodes amylophilus yielded oligosaccharides only and accordingly was
called liquefying a-amylases. It could be concluded that these enzymes are
saccharogenic. a-amylases due to the production of maltose and glucose.
4.6.Partial Purification of oc-amylase Enzymes:
Results of the column chromatography of the crude extract of the six bacilli
is shown in Table (4). The chromatography profile is shown in Fig. 45-50. It
is seen that the specific activity of amylases from all bacilli was increased
15-19 folds over the crude extract, while the protein content decreased to
about 4-5% of it's original value. Also it is seen that all the isolates gave a
single peak of a—amylase. At the same time the yield of all enzymes varied
in the range of 62-80% , a fact which when considered with increase in
specific activity would indicate that DEAE-Sephadex is an excellent
exchanger for these enzymes. The results obtained from this investigation of
the starch-degrading system showed that the a-amylases of all isolates were
56
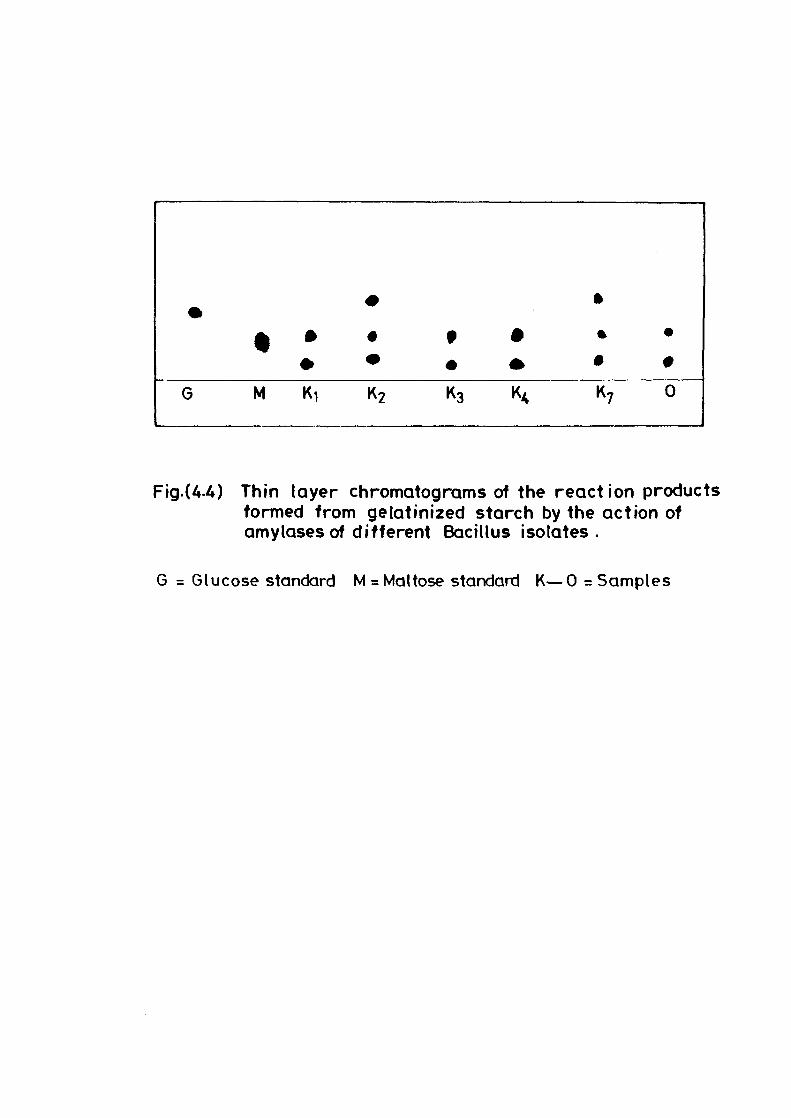
G M
ft
•
K,
#
•
K2
• •
K3 K,
• #
K7 0
Fig.(A4) Thin layer chromatograms of the reaction productsformed from gelatinized starch by the action ofamylases of different Bacillus isolates.
G = Glucose standard M = Mattose standard K—0 = Samples
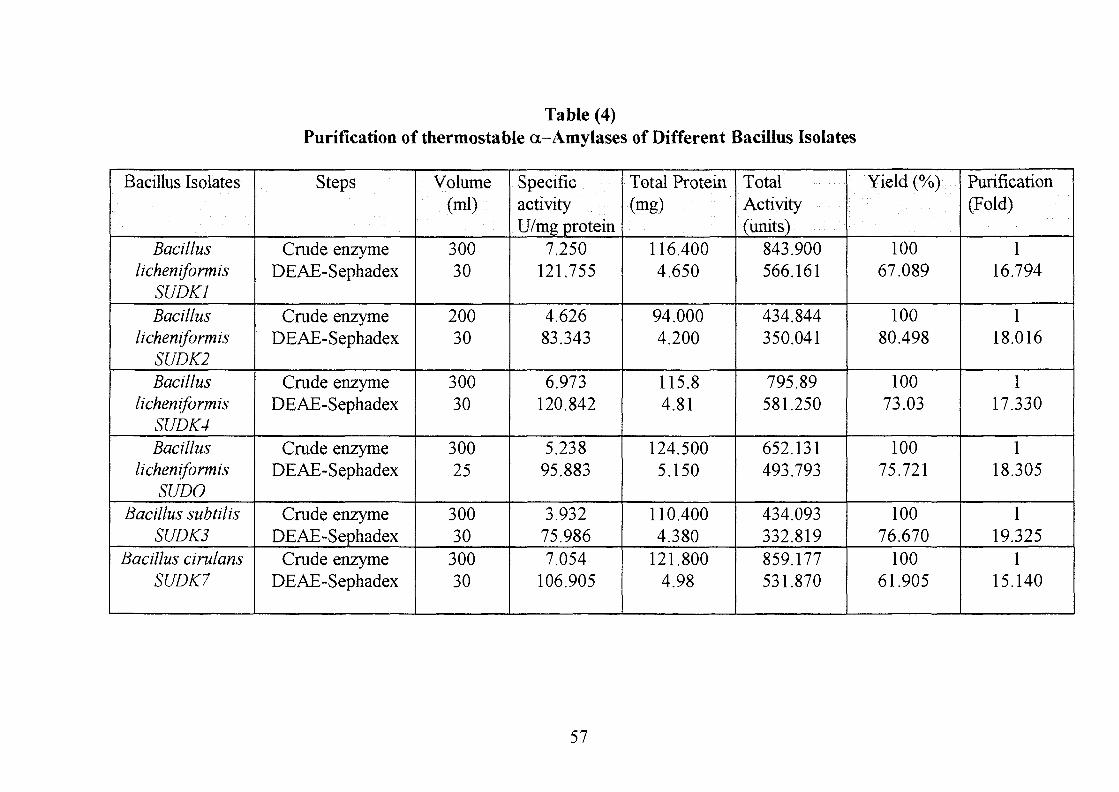
Table (4)Purification of thermostable a-Amylases of Different Bacillus Isolates
Bacillus Isolates
Bacilluslicheniformis
SUDK1Bacillus
licheniformisSUDK2Bacillus
licheniformisSUDK4Bacillus
licheniformisSUDO
Bacillus subtilisSUDK3
Bacillus cirulansSUDK7
Steps
Crude enzymeDEAE-Sephadex
Crude enzymeDEAE-Sephadex
Crude enzymeDEAE-Sephadex
Crude enzymeDEAE-Sephadex
Crude enzymeDEAE-Sephadex
Crude enzymeDEAE-Sephadex
Volume(ml)
30030
20030
30030
30025
30030
30030
SpecificactivityU/mg protein
7.250121.755
4.62683.343
6.973120.842
5.23895.883
3.93275.9867.054
106.905
Total Protein(mg)
116.4004.650
94.0004.200
115.84.81
124.5005.150
110.4004.380
121.8004.98
TotalActivity(units)
843.900566.161
434.844350.041
795.89581.250
652.131493.793
434.093332.819859.177531.870
Yield (%>..•
10067.089
10080.498
10073.03
10075.721
10076.670
10061.905
Purification(Fold)
116.794
118.016
117.330
118.305
119.325
115.140
57
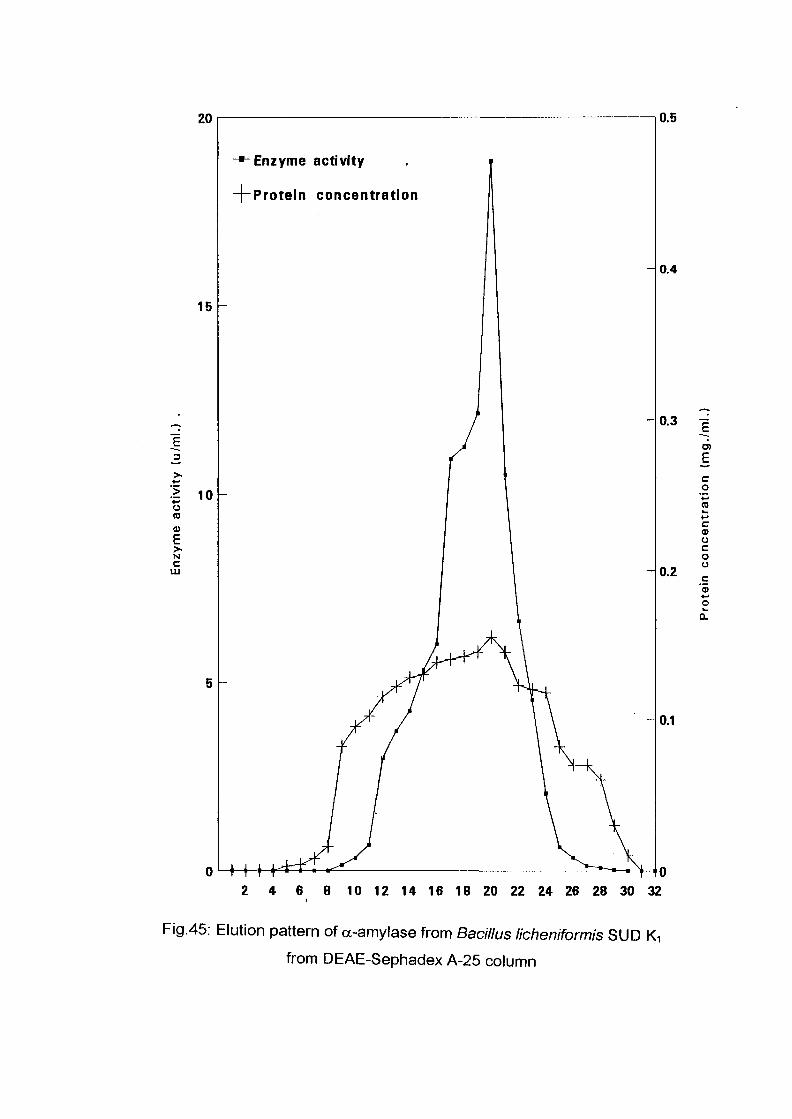
20
15
1 10uCO®ENC
UJ
-*• Enzyme activity
-|-Protein concentration
0.5
0.4
0.3 "i
0.2
co
c0)ocoo
O
a.
0.1
2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
Fig.45: Elution pattern of a-amylase from Bacillus licheniformis SUD Ki
from DEAE-Sephadex A-25 column
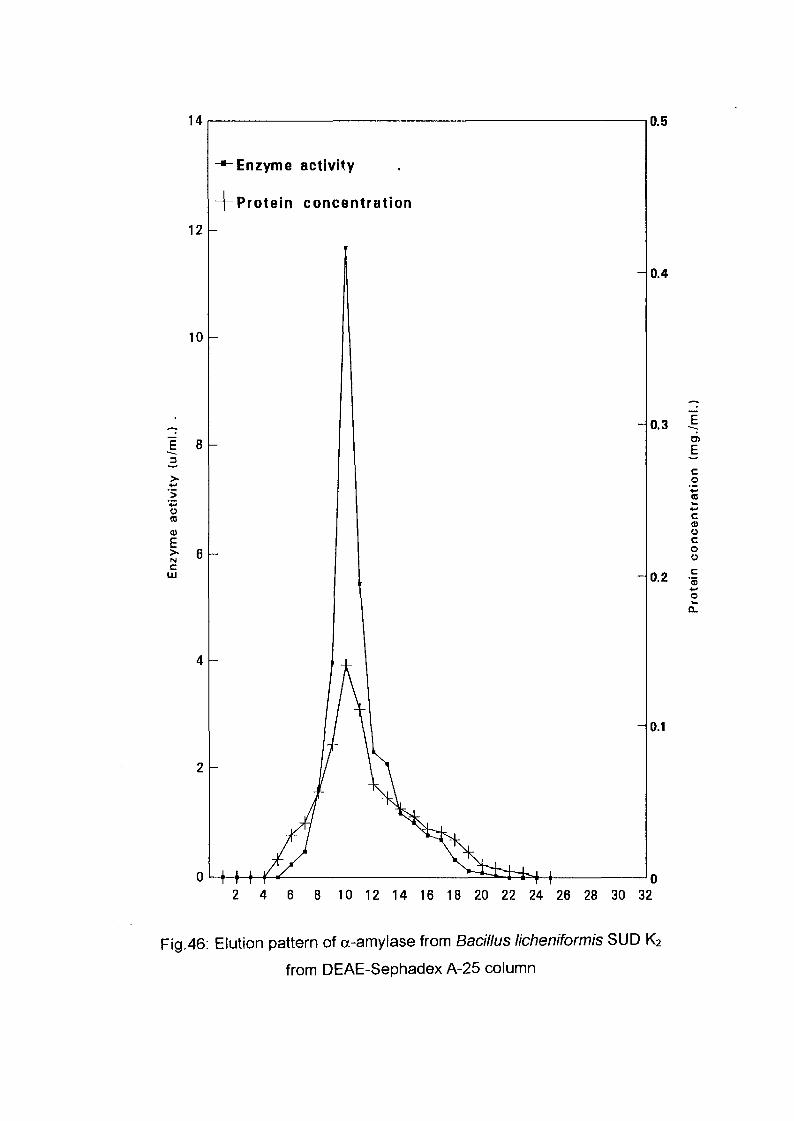
14
12 -
10
E 8
+oto
E>-N
c
Enzyme activity
Protein concentration
nL+++
0.5
0.4
0.3
0.2
0.1
2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
Fig.46: Elution pattern of a-amylase from Bacillus licheniformis SUD K2
from DEAE-Sephadex A-25 column
.E
co
ca>ocoo
c' 3
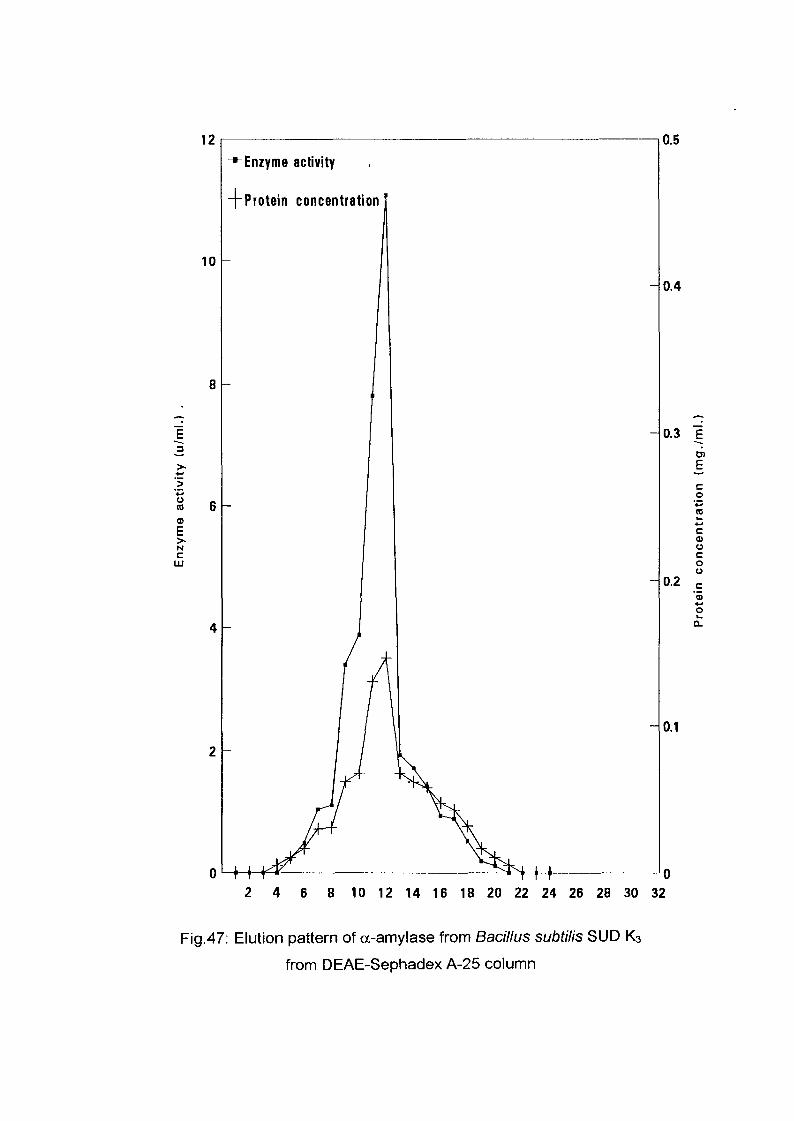
12
10
"5
uCO
0)ENc
LU
0.5
-"-Enzyme activity
-J-Protein concentration
0.4
0.3
0.2
Eco
cCDOCoo
oQ.
0.1
-H-2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32
Fig.47: Elution pattern of a-amylase from Bacillus subtilis SUD K3
from DEAE-Sephadex A-25 column
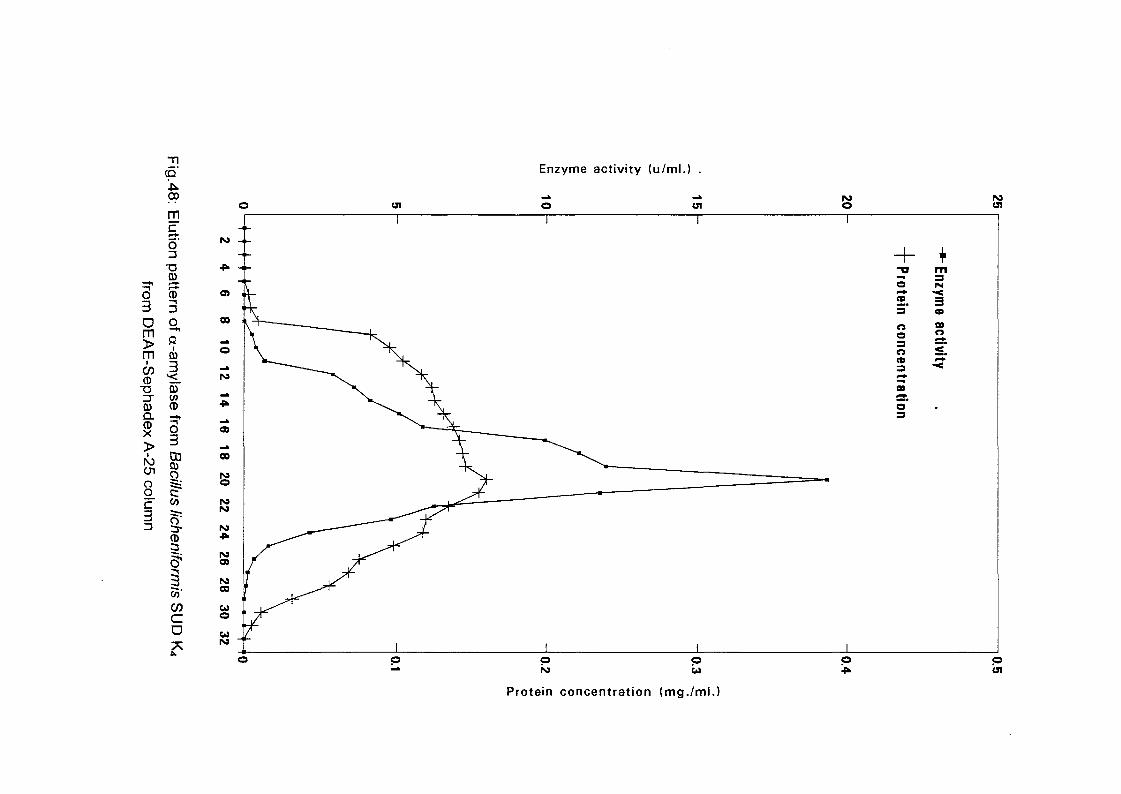
O
mcoCD
T3
0)Q.CDX
(p00
m
5'T303
CD3
Enzyme activity (u/ml.) .
CD
o3
I ICD3
CO-COca
uiO Ol
PO -
OS i
1
00
o
•p.
a)
00
PO©
ro
35
ro00
w ,o 'u _po •"
1
—
( ^
ti
o1
— ^
Ti —
± —.—-——
1
CJ1
1
— — - _ __ —
1
1
+-arotein c
oi
nCDltra
i
o"
r •
1
u
+mtizym
e actiivity
-
pW UI
Protein concentration (mg./ml.)
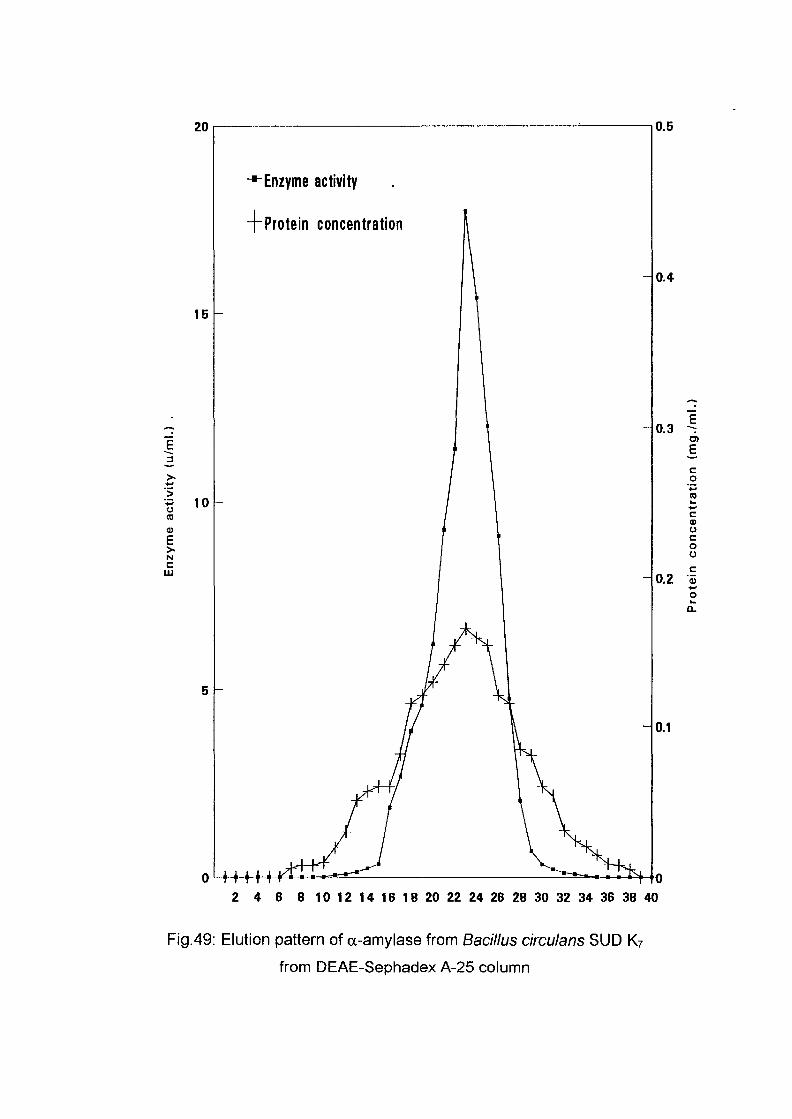
20
15
E3
ICOtoE>•
Nc
111
-•-Enzyme activity
T Protein concentration
0.5
0.4
0.3
E
co
c<uocoo
o.z
0.1
o ' l H H2 4 6 8 10 12 14 16 IB 20 22 24 26 28 30 32 34 36 38 40
Fig.49: Elution pattern of a-amylase from Bacillus circulans SUD K7
from DEAE-Sephadex A-25 column
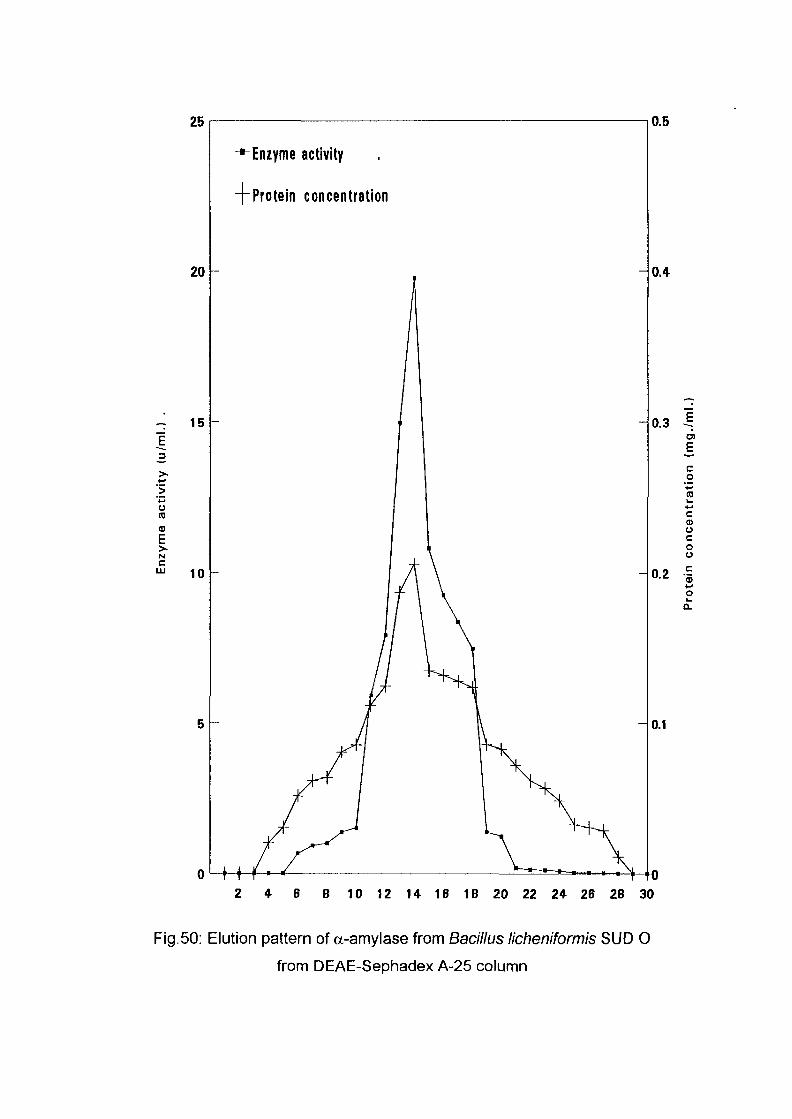
25
20
~ 15
_3
>•+-*'>'+*O(0<D
EN
10
-^Enzyme activity
-("Protein concentration
oM-t
0.5
0.4
0.3
0.2 -5
co
cCDOcooc5
0.1
-TO2 4 6 8 10 12 14 16 18 20 22 24 26 28 30
Fig.50: Elution pattern of a-amylase from Bacillus licheniformis SUD 0
from DEAE-Sephadex A-25 column

one enzyme system. Kanno (1986) and Jin el al (1992) purified a-amylase
of Bacillus acidocaldarius and Bacillus sp. J-F strain and found one activity
peak while Griffin and Fogarty (1973) using DEAE-Sephadex obtained two
peaks from the extract of Bacillus polymyxa.
4.7. Effect of Storage Temperature on Amylase Activity:
The partially purified oc-amylases of the different isolates were tested for
storage stability. Two storage temperatures, namely 4°C and -20°C were
evaluated with regard to their effect on enzymic stability. The results are
presented in Fig. 51-56
It is seen that storing the enzymes at -20°C did not result in any significant
loss of activity up to six months. All the enzymes retained 90% or more of
their original activity during this period. At 4°C all the enzymes showed
gradual loss in their activity and by the eighth week 60% or more of their
activity was lost. This finding is very important. If these enzymes are to be
used industrially they must be stable for appreciable periods of time to meet
the requirements for handling, shipping and storage. The inactivation of
enzyme preparation at the latter temperature (4°C) may be due to the action
of contaminating proteases or may be due to the presence of the
microorganism. This result is generally similar to the finding of Michelena
and Castillo (1984) who worked on the a-amylase of Aspergillus foeiidus.
4.8. Determination of Km and Vmax values of a-amylase:
Kinetic parameter of substrate hydrolysis of the a-amylase of the different
Bacillus isolates were determined using different concentrations of soluble
starch. From the double reciprocal plot shown in Fig. 57 (Linweaver and
58
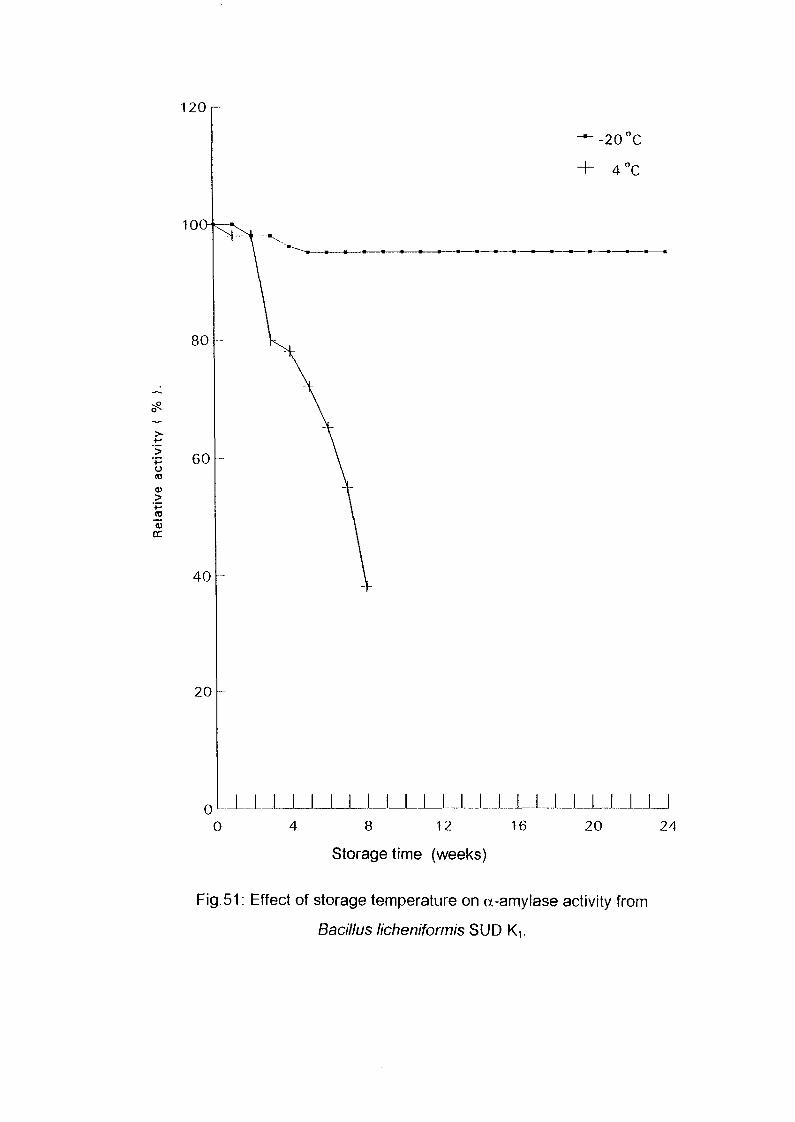
120 r-
(0
>
QC
-20°C
16
JLJL_LJL_L_U20 24
Storage time (weeks)
Fig.51: Effect of storage temperature on a-amylase activity from
Bacillus licheniformis SUD Ki.
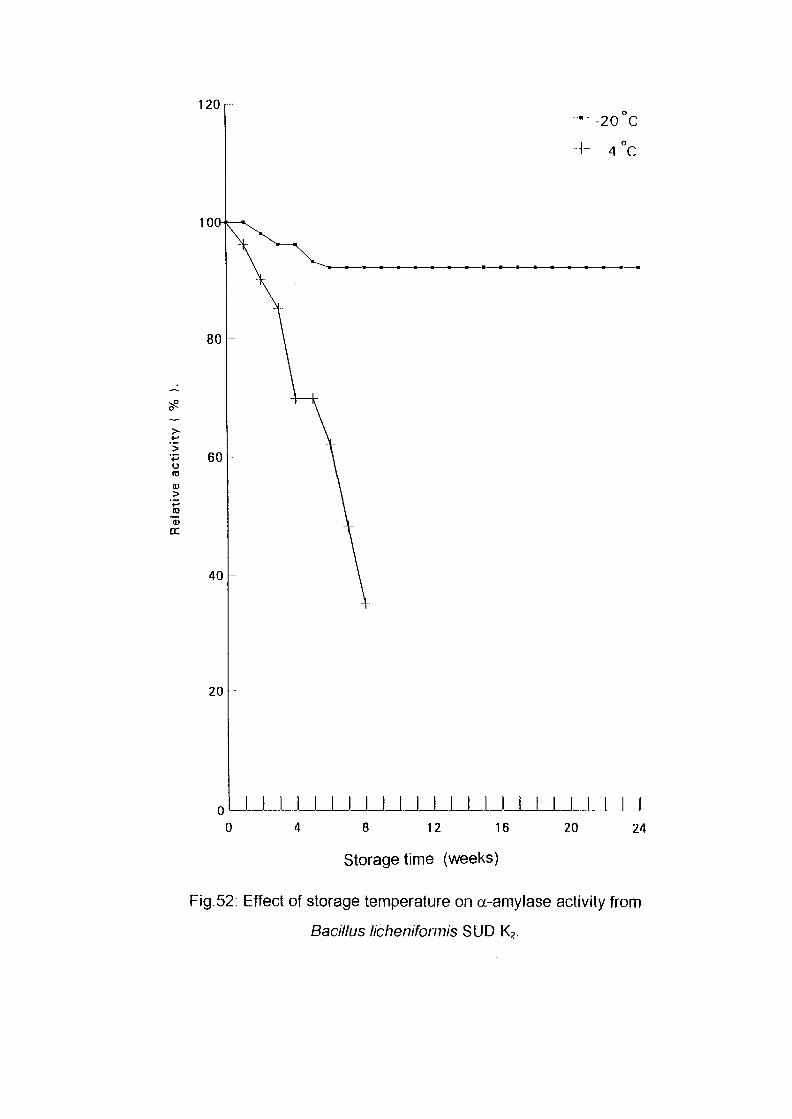
120
oto
— -20 C
+ 4°C
L_UJLi_LJLI_L8 12 16
Storage time (weeks)
20 24
Fig.52: Effect of storage temperature on a-amylase activity from
Bacillus licheniformis SUD K2.
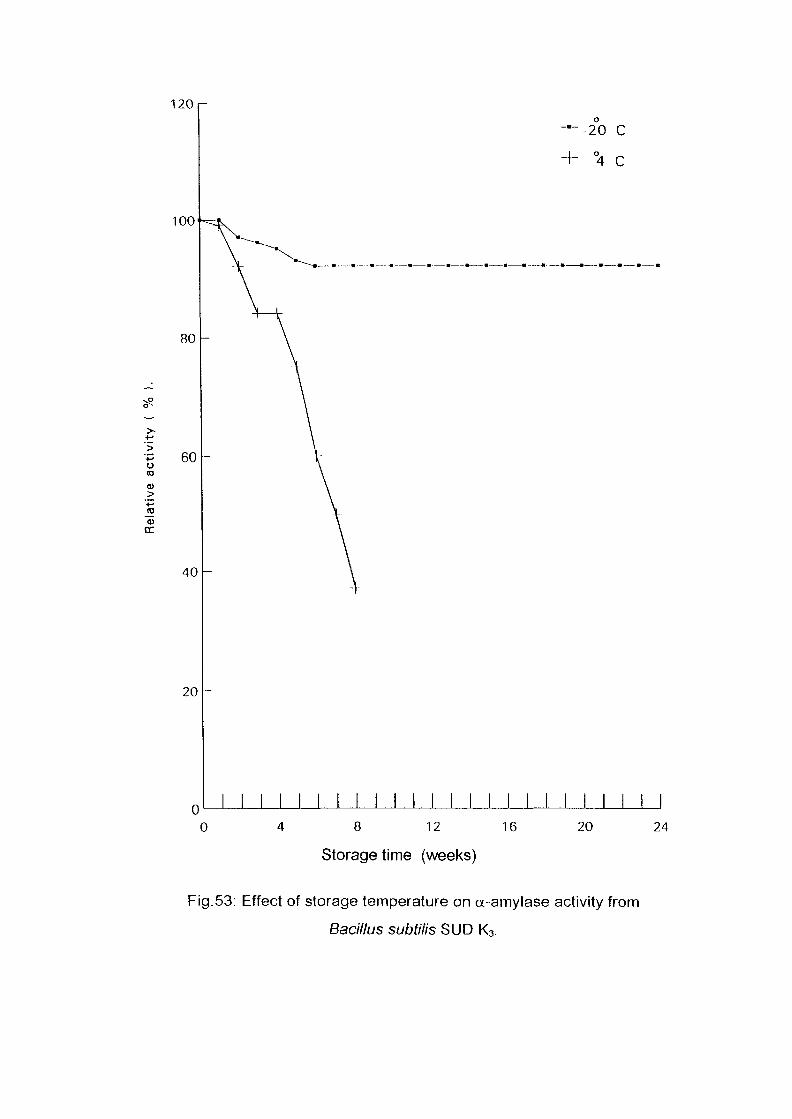
>-
oto
JO
120r-
100-^1
o
-20 C
°4 C
I I I I I I I I 1 I I 1 I 1 18 12 16
Storage time (weeks)
20 24
Fig.53: Effect of storage temperature on u-amylase activity from
Bacillus subtilis SUD K3.
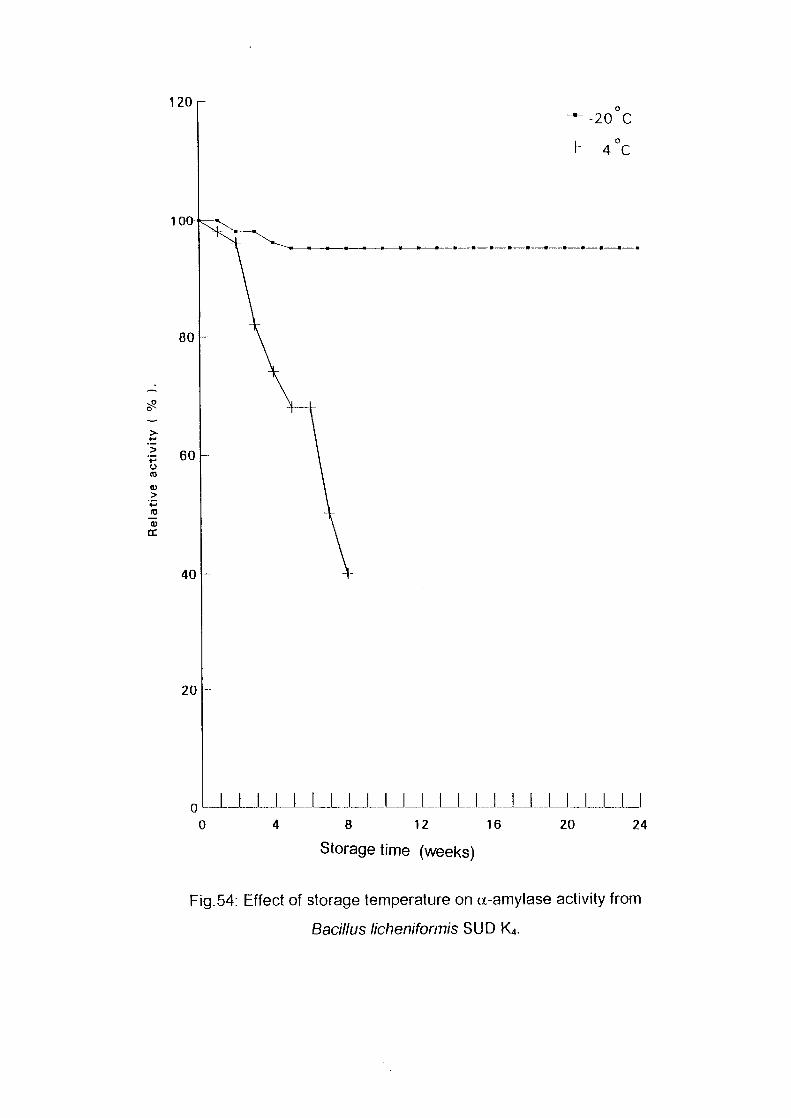
120
o03
>
-20°Co
4 C
JLJUJ_J_JJ_J_J_JJ_UJJ_JJLJU8 12 16 20 24
Storage time (weeks)
Fig.54: Effect of storage temperature on a-amylase activity from
Bacillus licheniformis SUD K4.
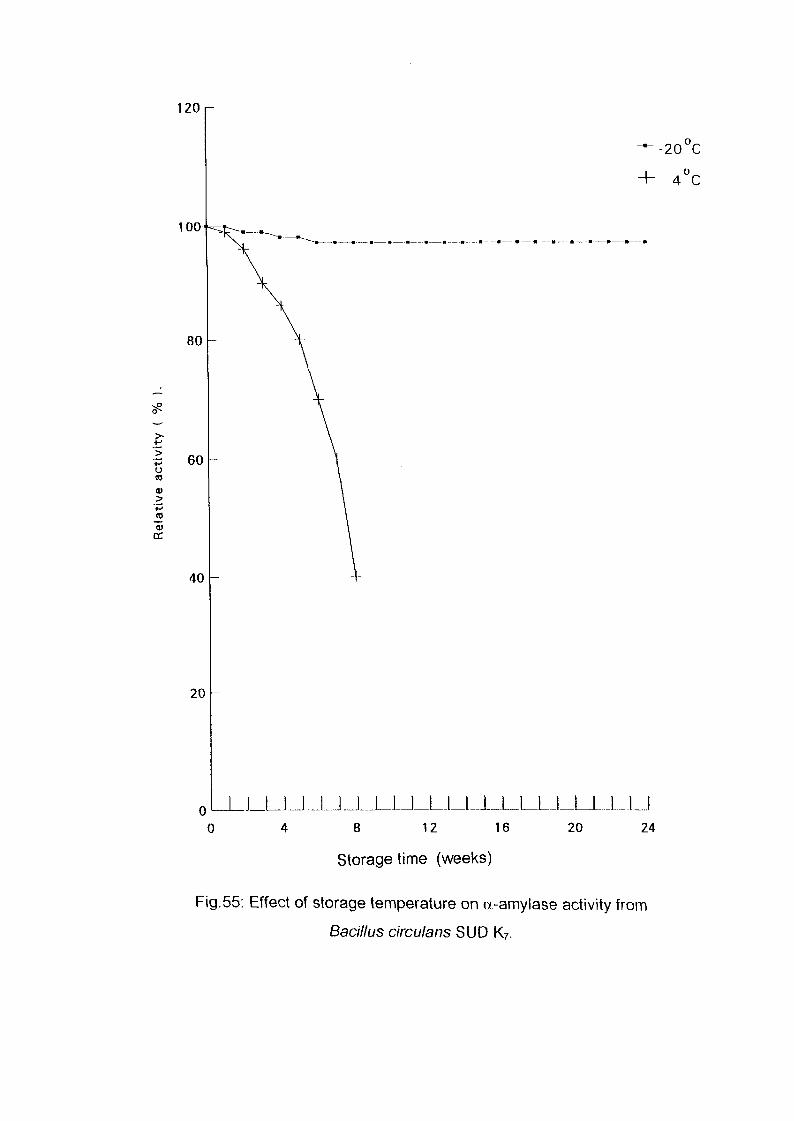
oto
0)DC
120
100-
-20°C
4°C
8 12 16
Storage time (weeks)
20 24
Fig.55: Effect of storage temperature on a-amylase activity from
Bacillus circulans SUD K7.
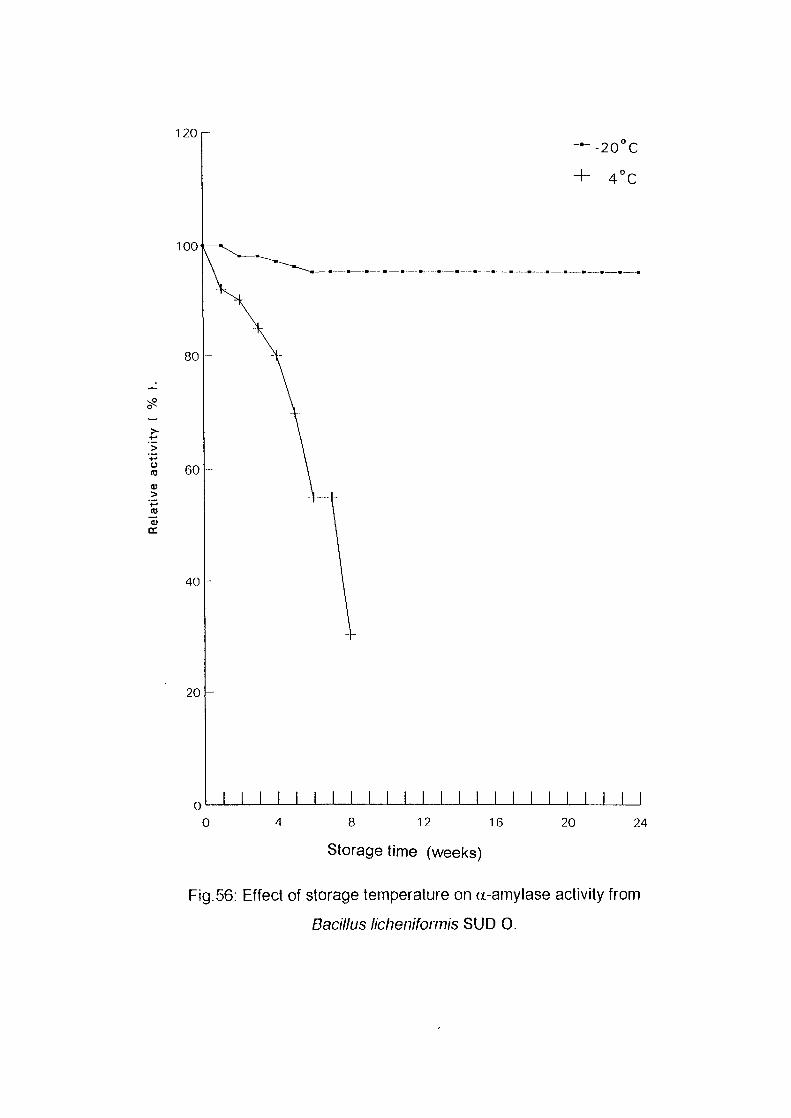
120r
100
<U>
80
60
40
20
—-20°C
+ 4°C
I I I I I I I I I I8 12 16
Storage time (weeks)
20 24
Fig.56: Effect of storage temperature on a-amylase activity from
Bacillus licheniformis SUD 0.
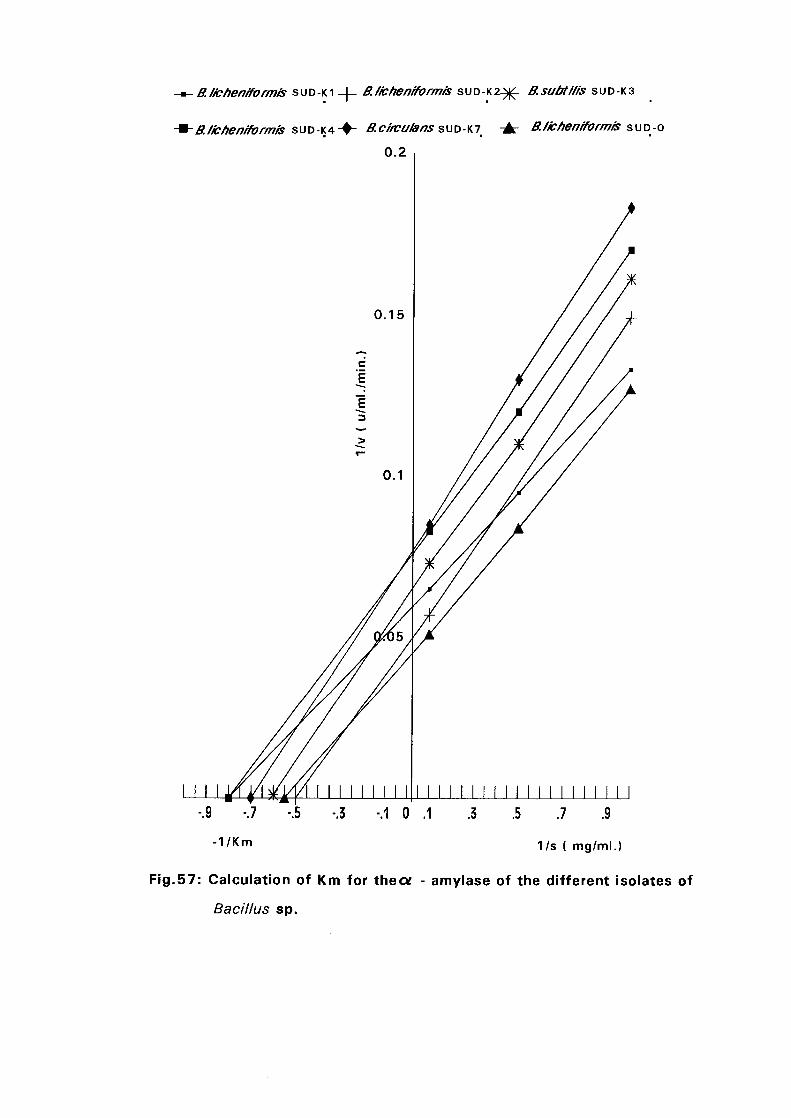
B. f/cheniformis SUD-KI - { - &/tcheniformis B.subti//s SUD-K3
Rfichenrformis SUD-K4+ B.CinculansSUD-K7
0.2
BJicheniforfnis SUD-O
.5 .7 .9
1/s { mg/ml.)
Fig.57: Calculation of Km for the« - amylase of the different isolates of
Bacillus sp.

Burk, 1934) the Vmax and Km were determined (Table 5). It is seen that the
Km values for all enzyme species lie in the range 1.25-2.00 mg./ml while the
Vmax. varied in the range of 13-23 U/ml/min.
The affinity of a-amylase for soluble starch of these thermophilic Bacillus
isolates as judged by the Km value were generally similar to the a-amylases
of thennophilic Bacillus acidiocaldarius Km = 0.8-1.6 mg/ml (Buonocore et
al, 1976 and Kanno, 1986). The affinity of these amylases are considerably
higher 1.25 to 2.0 fold than that of Kelbsiellae pneumoniae Km =2.5 mg/ml
and 2.34 to 3.7 fold than that of Bacillus caldovelox; km = 4.68 mg/ml.
(Monma et al, 1983 and Fogarty el al, 1991) respectively.
However, it is lower than that of Bacillus s1 ear othermophilus Km = 0.77
mg/ml by 1.6-2.6 fold and 1.9-3.1 fold lower than that of maltohexaose
producing a-amylases of Bacillus circulans Km = 0.65 mg/ml (Yutani et
al, 1973 and Takasaki, 1982).
59
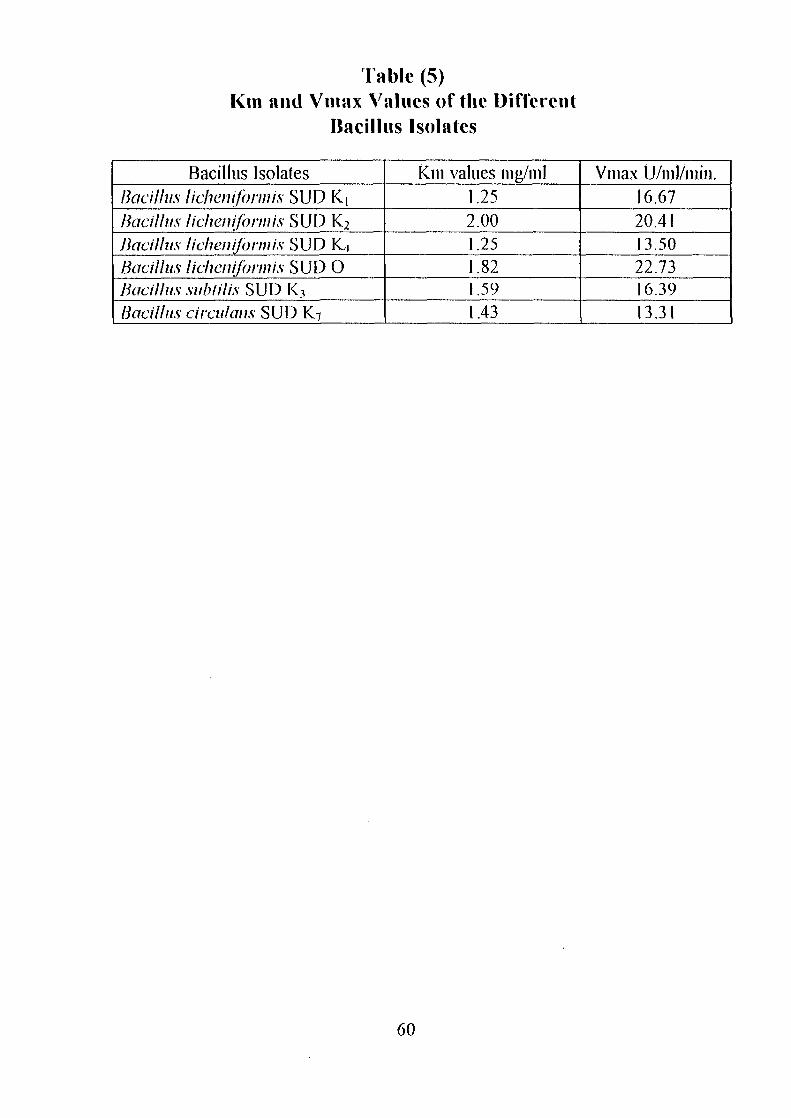
Fable (5)Km and Vinax Values of the Different
Bacillus Isolates
Bacillus IsolatesBacillus lichenifonnis SUD K|Bacillus lichenifonnis SUD K2
Bacillus lichenifonnis SUD K4Bacillus lichenifonnis SUD 0Bacillus subtil is SUD K3
Bacillus circulans SUD K7
Km values mg/ml1.252.001.251.821.591.43
Vinax U/nil/min.16.6720.4113.5022.7316.3913.31
60

4.9. Conclusions:
As conclusions the important findings could be summarised in the
followings:
1-A11 the enzymes have no specific requirements for metal ions for their
activity, this fact is of dual importance. From one side the poisonous
effects of heavy metals could be avoided since these enzymes are used in
the food industry. From the other side the absence of metal ions as
essentials for enzymic activity.
2-It is seen that these enzymes operate maximally at relatively low substrate
concentration. This would indicate that these enzymes are best used in an
immobilized form, to achieve the highest activity and thus the best
economic situation.
3-Six isolates were found to be thermophilic with their amylases operating
maximally at 60-70°C.
4-Of the six isolates two could be the same strain. These are Bacillus
licheniformis SUDK] and SUDK4. When the effect of temperature and
time interval, pH, substrate concentration, metal ions, organic nitrogen
and carbon source were studied in relation to amylase production and
when effects of temperature, pH, time interval and substrate concentration
were studied in relation to enzymic activity, the above two isolates were
found to be very similar. On this basis they could be the same strain.
5-Of special importance among the six strains are Bacillus circulans
SUDK7 and Bacillus licheniformis SUDK2 since they produce glucose.
4.10. Suggestions for further work:
As regards for future work it is recommended to purify and characterise,
physically and biochemically, all the six enzymes. Secondly, it is
recommended to immobilise the six enzymes both as free enzymes and
61

attached to the bacterial cells and study their behavior and characteristics as
amylolytic enzymes. Thirdly, it is important to study the hydrolytic
characters of these enzyme towards raw starch (unpiirified grain flour).
62

Achi, O.K. and Njoku-obi, A.N.U.(1992).Production of a raw-starch saccharifing
amylase by Bacillus alvei grown on different agricultural substrates.
World Journal of Microbiology and Biotechnology 8, 206-207.
Ahmed, S.B.(1988). Taninns contents, amylase activity and cyanide content of
germinated sorghum grain. MSc. Thesis U.of K.
Ali, S., Mohamood, S., Alam, R. and Hossein, Z. (1989). Culture conditions
for production of glucoamylase from rice bran by Aspergillus terms.
MICRCEN Journal 5, 525-532.
Ali, S., Hossein, Z. , Mahmood , S. and Alam, R.(1990). Induction of
glucoamylase production by non-starch carbohydrates in Aspergillus
terms. World Journal of Microbiology and Biotechnology 6, 19-22.
Allen, J. D., Thoma, J. A. (1978). Model for carbohydrase action. Aspergillus
oryzae a-amylase degradation of maltoriose. Biochemical Journal 17,
2345-2350.
APHA (1955). Standard method for examination of water sewage and industrial
wastes 10th ed.American public Health Association. New York.
Bajpai, P., Rajesh, K.G. and Baipai, P. K. (1992).Optimization studies for the
production of a-amylase using cheese whey medium . Enzyme Microbial
Technology 14, 679-683.
63

Balls, A. K., Walden, M. K. and Thompson, R. R. I. (1948). In: "Enzymes".
Malcolm, D. and Edwin, C.W. (eds.) (1964). 2nd.ed.pp. 802-808.
Academic Press NewYork.
Baifoed, H. L. (1981) In: "Essay in applied Microbiology" Norris J. R. and
Richmond M. W. (eds.) J. Wiley .London.
Bartholomew J. W. and Mittwer's, T. (1950). A simplified bacterial spore stain.
Stain Technology 25, 153.
Bealin-Kelly, F, Kelly, C.T. and Fogarty, W.M. (1990).The cc-amylase of the
caldo-active bacterium Bacillus caidoveiox. Biochemical sociely
transect ion 18, 310-311.
Bergmann, F. W., Abe, J. and Hizukuri, S.(1988). selection of microorganisms
which produce raw- starch degrading wnylases Applied
Microbiology and Biotechnology 27, 443-446.
Bemfeld, P. (1955). Amylases a and b. Methods in Enzymology 1, 149-158.
Bezbaruch, R. L., Gogoi, B. K., Pilai, R. R. and Nigam, J. N. (1991).
Amylase production by three Bacillus strains active at alkaline pH.
Journal of Basic Microbiology 31, 13-20.
Bhumibhamon, O. (1983). Production of glucoamylase by submerged culture.
Thai Journal of Agricultural Science 16, 173-184.
Bhumibhamon, O. (1986).Glucoamylase of a fungus isolated from a rotting
cassava tuber MIRCEN Journal 2, 473-482.
64

Bird, R. and Hopkin, R. H. (1954). In: "General Biochemistry" Fruton, J.S. and
Simmods, S. (eds.) (1958). 2nd ed. pp. 434-438. J. Wiley London.
Boel, E. , Brady, L., Brzozowski, A.M., Derewenda, Z., Dodson, G. G.,
Jensen, V. J., Peterson, S.B., Swift, H., Thim, L. and Woldlike, H.F.
(1990). Calcuim binding in a-amylases: An x-ray diffraction study at
2.1A° resolution of two enzymes from Aspergilhis niger. Biochemistry 29,
6244-6249.
Botes, D. P. , Joubert, F. J. and Novellie, L. (1967). Kaffircorn malting and
brewing studies. Purification and properties of sorghum P-amylases.
Journal of Science, Food and Agriculture 18, 409-414.
Boyer, E.W. and Ingle, M. B. (1972). Extracellular amylase from a Bacillus spp.
Journal of Bacteriology 9, 992-1000.
Boyer, E.W. and Ingle, M.B. (1977).In: "Hand book of Enzyme
Biotechnology". Wiseman, A. (ed.) 2nd.ed. (1985). pp. 330-334. Ellis
Horwood Publishers New York.
Bradford, M. M. (1976) A rapid and sensitive method for the quantition of
microgram quantities of protein -dye binding. Analytical Biochemistry 72,
248-254.
Budair, A.A. (1977). Chemical studies on sorghum grain and their products.
M.Sc. Thesis U. of K.
Buisson,G., Duee, E. , Haser, R.and Payan, F. (1987). Three dimentional
structure of porcine pancreatic a-amylase at 2.9A°resolution. Role of
calcium in structure and activity. EMBO. Journal 6, 3909-3916.
65

Buonocore, N. , Caporale, C , Derosa, M.and Cambocoria, A.(1976). Stable
inducible thennophilic a-amylase from Bacillus acidocaldarius. Journal
of Bacteriology 128, 515-512.
Burbidge, E. and Collier, B. (1968). Production of bacterial, amylases.
Process Biochemistry 3, 53-56.
Burgess-Cassler, A. and Imam, S. (1991). Partial purification and comparative
characterization of a-amylase secreted by Lactobacillus amylovorus.
Current Microbiology 23, 207-213.
Campbell, L. L. (1954). Ciystallization of a-amylase from a thennophilic
bacterium. Journal of the American Chemical Society 76, 5256-5257.
Campbell, L. L. (1955). Purification and some properties of an a-amylase from a
facultative thermophilic bacterium. Archives of biochemistry and
Biophysics 54, 154-161.
Campbell, L. L. and Manning, G.B. (1961). Thermostable a-amylase of
Bacillus stearothermophilus. III. Amino acid composition. Journal of
Biological Chemistry 236, 2962-2965.
Chiang, J.P., Alter, J.E., and Steinberg, E.M. (1979). Purification and
characterization of a thermostable a-amylase from Bacillus
Ucheniformis. StarkS(ark3\, 86- 92.
Chu, H.P. (1949). The lectithinase of Bacillus cereus and it's comparison with
Clostridium welchii a toxin'. Journal of General Microbiology 312, 255
-273.
66

Claus, D. and Berkeley, P.C.W. (1986). Genus Bacillus In: "Bergey's Manual
of Systematic Bacteriology". Vol.2: Sneath, P.H.A., Bair, N.S., Shatpe,
M.E. and Holt, J.G. (eds.). pp. 1115-1123. Williams and Wilkins,
Baltimore.
Clayton, D.W., Jurasek, L.,Paice, M.G. and Leary, OS. (1984). In: "Handbook
of Enzyme Biotechnology". Wiseman, A. (ed.)2nd. ed. pp. 366-367. Ellis
Horwood Publisher, New York.
Coleman, G. and Elliot ,W. (1962). Characterization of a-amylase formation
by Bacillus sublifis. Journal of General Microbiology 83, 256-263.
Cori,G.T. and Laner, J.(1951). Action of amylo 1,6-glucosidase and
phosphoiylase on glycogen and amylopectin. Journal of Biological
Chemistry 188, 17-29.
Crawford, R. J.M.(1962). Citrate utilizing activity of certain starter bacteria.
16th ed. International Dairy Congress B, pp. 322
Cruickshank, R. (1960). In: "Handbook of Bacteriology". Mackie and McCatney
(eds.) 10lh ed., pp. 320-323. Livingstone, London.
Daiber, K. H. and Novellie, L. (1968). Kaffircom malting and brewing studies.
Gibberellic acid and amylase formation in kaffircorn. Journal of Science,
Food and Agriculture 19, 87-90.
Danielson, C.E. (1948). Molecular weight of malt amylase. Nature 162, 525.
Danielson, C.E.and Sandegien, E.(1947). Investigation of bailey and malt
amylase. A eta Chemica Scandinavica 1, 917-928.
67

Davis, J.G.(1959). Milk testing. 2nd.ed. Dairy Industries Ltd pp. 175-187.
London.
De Mot, R., Van Oudenijek. E., Hougoerts, P. and Verachtert, H. (1984)
Effect of medium composition on amylase production by some starch
degrading yeasts. FEMS. Microbiology Letter 25, 169-173.
Dixon, M. (1953). The determination of enzyme inhibitor constants. Biochewica]
Journal 55, 170-171.
Ellis, J. J., Wang, J. L. and Hesseltine. C.W. (1974). Rhizopus and
Chlamydomucor strains surveyed for milk-clotting, amyloltic and
antibiotic activities. Mycologia 66, 593-599.
Flemingi, I.D. and Stone, B. A. (1965). Fractionation of Aspergillus niger
amyloglucosidase. Biochemistry Journal 97, 13p.
Fogarty, W.M. (1983). Microbial amylases In: " Microbial Enzymes and
Biotechnology". Fogarty, W.M. (ed.)pp. 1-92. Applied Science Publishers,
London.
Fogarty, W.M. and Kelly, C.T. (1980). Amylases, amyloglucosidase and
related glucanases. In:" Microbial Enzymes and Byconversions".
(Economic Microbiology). Rose, AH. (ed.) Vol. 5, pp 115-170. Academic
Press, London.
Fogaity, W.M. and Kelly, C.T. (1990). recent advances in microbial amylases
In: " Microbial Enzymes and Biotechnology". Fogarty, W.M. and Kelly,
C.T. (eds.). 2nd . ed. pp. 71-132. Elsevier Applied Science Publishers,
68

Amsterdam.
Fogarty, W.M. , Bealin, F., Kelly, C.T.and Doyle, E.M. (1991). A novel
maltohexaose-forming a-amylase from Bacillus caldovelox. Pattern and
mechanisms of action. Applied Microbiology and Biofechnologv 36, 184-
189.
French, D. and Knapp, D. W. (1950). The maltase ofC/ostridiuin
acetobutylictim, it's specificity range and mode of action. Journal of
Biological Chemistry 187, 463-495.
Fukumoto, J. and Tsujsaka, Y. (1956). In: " Biochemistry of Industrial
Microorganisms". Rain Bow, C. and Rose, A.H. (eds.)(1963). pp. 106-
107. Academic Press, London.
Fukumolo, J., Yamamoto, T.and Tsui a, D.(I957). Effect of carbon sources and
base analogues of nucleic acid on llie formation of bacterial aniylase.
Nature (London). 180, 438-439.
Glymph, J. L. and stutzenberger, F. (1977). Production, Purification and
characterization of a-amylase from Thermonospora curvata. Applied and
Environmental Microbiology 34, 391-397. Godfrey, T. and Reichelt, J.
(1983). Textiles In: "Industrial Enzymology".
Godfrey, T. and and Reichelt, J. (eds.) pp. 397-409. The NaturePress, New York.
Gordon, R. E., Haynes, W. C. and Hor-Nay Pang, C. (1973). "The Genus
Bacillus", Agriculture Research Service, U.S. Department of Agriculture,
Washington D.C.
69

Goyal, N. , Sidu, G. S., Chakrabort, T. and Gupta, J. K. (1995).
Thennostabiliry of a-amylase produced by Bacillus sp. E2. a thennophilic
mutant. Journal of Microbiology and Biotechnology 1 1, 593-594.
Griffin, P. J. and Fogarty, W. M. (1973). Pelriminary observations on the
starch degrading system elaborated by Bacillus polymyxa. Biochemical
Society Transactions 1, 397-398.
Guilbot, A. and Mercier ,C. (1985). Starch. In: "The Polysaccharides "Aspinall,
G.O. (ed.) pp. 209-282. Academic Press, New York.
Gupta, A. K. and Gautan, S. P. (1995). Improved production of extracelluar a-
amylase by the thermophilic fungus Malhranchea sulfurea following
protoplast fusion. World Journal of Microbiology and Biotechnology 11,
193-195.
Haas, G. J. (1976). Alcohol beverages and fermented foods. In: "Industrial
Microbiology". Miller, B.M., Litsky, W. (eds.) pp. 165-191. McGraw-
Hill, New York.
Hang, Y.D.(1989). Direct fennentation of com to L(+)-lactic acid by Rhizopus
oryzae. Biotechnology Letters 11, 299-300.
Hankin, L. and Anagnostakis, S. L. (1975). The use of solid media for
detection of enzyme production by fungi. Mycologia 67, 597-600.
Harrigan, W.F.and Margaret, E. McCance, (1966). "Laboratory Methods in
Microbiology", pp. 3-316. Academic Press, London.
Harris, G.(1968). In: "Barley and Malt". Cook, A.H. (ed.) pp. 624-640.
70

Academic Press, London.
Hartman, P.A., Wilerson, R. JR. and Tetraut, P.A. (1955). Bacillus siear-
thermophiltis. 1-Thermal and pH stability of the amylase. Applied
Microbiology 3, 7-10.
Hassid, W. Z. and Newfeld, E. F. (1955). Hydrolysis of amylase by (3-
amylase and Z-enzyme. Archives of Biochemistry and Biophysics 159,
405-419.
Hayashida, S., Teramoto, Y. and Inoue, T. (1988). Production and
characteriation of raw-potato starch digesting a-amylase from Bacillus
subtilis 65. Applied and Environmental Microbiology 54, 1516-1522.
Hayashi, T., Akiba, T. and Horikoshik, K. (1988). Properties of new alkaline
maltohexaose forming amylases. Applied Microbiology and Biotechnology
28,281-285.
Heinen, U. J. and Heinen, W. (1972). Characterization and properties of
a caldo-active bacterium producing extracellular enzyme and two related
strains. Archives of Microbiology 82, 1-23.
Hesseltine, C.W., Smith, M., Bradle, B. and KO, S.D. (1963). Investigation
of tempeh, an Indonesian food. Development in Industrial Microbiology 4,
275-287.
Higashihara, M. and Okada, S.(1974). Studies on p-amylase of Bacillus
megaterium strain No. 32. Agriculture and Biological Chemist ty 38(5),
1023-1028.
Ilori, M.O.Amund, O. O. and Omidiji, O.(1995). Purification and properties
71

Ilori, M.O.,Amund. O. O. and Oinidiji, 0.(1995). Purification and properties
of a glucose forming amylase by LacfobacH/u.s brevis. World Journal of
Microbilogy andBiotechnology 11, 595-596.
Imam, Sll., Burgess-Cassler,A., Core, G.L., Gordon, S.ll. and Baker, F. L. C.
(1991). A study of corn starch granule digestion by an unusually high
molecular weight a-amylase secreted by Lactobacillus amylovorus:
identification of an unusally high molecular weight. Current
Microbiology 22, 365-370.
Ingle, M. B. and Boyer, E.W. (1976). Production of industrial enzymes In:
"Microbiology".Schlessinger, D. (ed.) pp. 420-426. American Society for
Microbiology, Washington D.C.
Jaeyong, K., Nanmori, T. and Shinka, R. (1989). Thermostable- raw starch
digesting amylase from Bacillus stearothermophilus. Applied and
Enivronmental Microbiology 55, 1636-1639.
Jin, F., Li, X.,Zhang, C., Shu, O., Wu, II., Liu, Z., Zhang, X. ,Wu, X. and
Cheng, Y. (1992). Purification and characterization of a thermostable a-
amylase from Bacillus sp.JF strain Journal oj General and Applied
Microbiology 38, 293-302.
Ju-Nichi, A., Fredcrica, B.W., Kazuoki, 0. and Susmu, II. (1985). Raw
starch digesting enzyme of Aspergillus spp. K-27- Denpum Kogoku 32,
128-135.
Kainuma, K. , Katsuo, W., Shoichi, K. , Akiko, N. and Shigeo, S. (1975).
Purification and some properties of a novel maltohexaose- producing
exoamylase from Aerobactor aerogenes. Biochimica e( Biophysica Acta
72

410,333-345.
Kanlayakrit, W., Ishimatsu, K. , Nakao, M.and Hayashida, S. (1987).
Characterization of raw-starch digesting glucoamylase from thermophilic
Rhizonwcor pusiUns. Journal of Fermentation Technology. 65, 379-385.
Kanno, M. (1986). A Bacillus acidocaldarius a-amylase that is highly stable
lo heat under acidic conditions. Agriculture and Biological Chemistry
50(1), 23-31.
Karlson, P. (1974). " Introduction to Modem Biochemistry". 4th. ed. pp. 343-
346 Academic Press, New York.
Khire, J. M. and Pant, A. (1992). Thermostable salt tolerant amylase from
Bacillus sp. 64. World Journal of Microbiology and Biotechnology 8, 167
-170.
Kirochhoff, M. J. (1816). In: " Chemistry and Industry of Starch". Ken,
P.W.(ed.) (1949). 2nd.ed. pp. 407-438. Academic Press, London.
Kita, Y., Sakaguchi, S., Nitta, Y. and Watanabe,T. (1982). Kinetics study on
chemical modification of Taka amylase A.I 1 Ethoxy-Carbonylation of
histidine residues. Journal of Biochemistry 92, 1499-1504.
Kneen, E. and Beckord, L.D. (1946). In: "Chemistry and Industry of starch".
Ken, W. (ed.) (1949). 2nd. ed. pp. 407-438. Academic Press, London.
Kneen, E. (1949). In: "Chemistry and Industry of starch". Kerr, W. (ed.) (1949).
2nd. ed. pp. 407-438. Academic Press, London.
73

nutrition and physiology of mesophilic species in the genus Bacillus.
Journal of General Microbiology 4(3), 508-538.
Koaze, Y. , Nakajima, Y., Hidaka, H., Niwa, T., Adachi, K., Niid, T.,
Shomura, T.and Uede, M. (1975). In:" Handbook of Enzyme
Biotechnology." Wiseman, A.(ed.) (1985). 2nd. ed. pp. 329-330. Ellis
Horwood Publishers, NewYork.
Komaki, T. (1956). Industrial application of amylase Denpum Kogyo
Gakkaishi 4, 33-39.
Kuo, M. J. and Hartman, P. A. (1966). Isolation of amylolytic strains of
Thermoactinomyces vulgar is and production of thermophilic
actinomycetes amylases. Journal of Bacteriology 92, 723-726.
Kwan, H.S., So, K.H., Chan, K. Y. and Cheng, S. C. (1993). production of
thennotolerant p-amylase by Bacillus circulans. World Journal of
Microbiology and Biotechnology 9, 50-52.
Laner, J. and Nickle, C. M. (1955). Gastrointestinal digestion of starch. 1- The
action of oligio 1,6- glucosidase or branched saccharides. Journal of
Biological Chemistry 215, 723-736.
Lazer, O. (1938). In: "Chemistry and industry of starch". Ken, R.W. (ed.)
(1949). 2nd ed. pp. 407-438. Academic Press, London.
Levin, M. (1916). The Voges Proskauer Reaction. Journal of Bacteriology 1,
153-164.
Lim, G. ,Tan, T.R. and Rahim, N. A. (1987). Variation in atnylase and
74

protease activities among Rhizopus isolates. M1RCEN Journal 3, 319-322.
Linardi, V. R. and Machado, K. M. (1990). Production of amylases by yeasts.
Canadian Journal of Microbiology 36, 751 -753.
Lineback, D. R., Russell, I. J. and Rosamassen, C. (1969). Two forms of
glucoamylase ofAspergillus niger. Archives of Biochemistry and
Biophysics 134, 539-553.
Lineback, D. Rand Aira, L.A. (1972). Structure and Characterization of the
two forms of glucoamylase from Aspergillus niger. Cereal Chemistry 49,
283-298.
Lineweaver, H. and Bulk, D. (1934). The determination of enzyme dissociation
constant. Journal of the American Chemical Society 56, 658-666.
MacAllister, R.V. (1979). Nutritive sweeteners made from starch. In: "Advances
in Carbohydrate Chemistry and Biochemistry". Tipson, R.J. and Horton
D. (eds.) Vol. 36, 15-56.
Madsen, G.B., Norman, B.E. and Slaff, S. (1973). A new heat stable bacterial
amylase and it's use in high temperature starch liquefaction. Die Stark 25,
304-308.
Maeda, S. and Tsao, G.T. (1979). In:" Handbook of Enzyme Biotechnology".
Wiseman, A.(ed.) (1985). 2nd. ed. pp. 328-330. Ellis Horwood Publishers,
NewYork.
Mai, N.T., Giang, D. T., Minh, N.T.N. and Thao, V.T. (1992). Thennophilic
75

amylase producing bacteria from Vietnamese soils. World.Journal of
Microbiology and Biotechnology 8. 505-508.
Manjunath, P. , Shenoy, B.C. and Roglmvendra Rao, M.R. (1983). Fungal gluco-
amylases. Journal of Applied Biochemistry 5, 235-260.
Manning, G.B. and Campbell, L. L. (1961). Thermostable a-amylase of Bacillus
stearofhermophilus. 1-Crystallization and some chemical properties.
Journal of Biological Chemistry 236, 2925-2957.
Matsuura, Y., Kusumoki, M. , Harada, W. and Kakuko, W. (1984). Structure
and possible catalytic residues of Taka-amylase A. Journal of
Biochemistry. 95, 697-702.
Matsuzaki, H. , Yamane, K. Yamaguchi, K., Nagata,Y. and Maruo, B. (1974).
Hybrid a-amylase produced by Iransfot ments of Bacillus suhdlis.
1-Purification and characterization of extracellular a-amylase produced by
parental straifis and transfoi ments. Biochimica ef Biophysica Ada 365,
248-258.
McWealthy, S. J. and Hattman, P.A. (1977). Purification and some properties of
an extracellular a-amylase from Bacteroids amylophilus. Journal of
Bacteriology 129, 1537-1544.
Medda, S. and Chandra, A. K. (1980). New strains of Bacillus lichenifarmis
and Bacillus coagulans producing thermostable a-amylase active at
alkaline pH. Journal of Applied Bacteriology. 48, 47-58
Meers, J.L. (1972). The regulation of a-amylase production in Bacillus
licheniformis. Atonic Vanleeuwhoek. Journal of Microbiology and
76

Serology 38, 585-590.
Meyer, K.H., Fisher, E.H. and Bernfeld, P. (1951). In: " General
Biochemistry". Fritton, S. S. and Simmods, S. (eds.) (1951). 2nd. ed. pp.
434-450. J. Wiley, New York.
Michelena, V. N. and Castillo, F. J. (1984). Production of amylase by
Aspergillus foelidus on rice flour medium and characterization of the
enzyme. Journal of Applied Bacteriology 56, 395-407.
Miller, G. L. (1959). Use of dinitrosalicyltc acid reagent for determination of
reducing sugar. Analytical Chemistry 31, 426-428.
Mitsue, T., Saha,B.C. and Ueda, S. (1979). Glucoamylase of Aspergillus
oryzae cultured on steamed rice. Journal of Applied Biochemistry 1, 410-
422.
Monma, N., Nakakuki, T. and Kainuma, K. (1983). Formation and hydrolysis
of maltohexaose. Agriculture and Biological Chemistry 47, 1769-1774.
Morgan, F.J. and Priest, F. G. (1981). Characterization of a thermostable a -
atnylase from Bacillus lichenifbrmis NCIB 6346. Journal of Applied
Bacteriology 50, 107-114.
Moseley, M. II. and Keay, L. (1970). Purification and characterization of the
amylase of Bacillus suhtilis NRRLB 3411. Bacteriology and
Bioengineering 12, 251-271.
Mundy, J. (1982). Isolation and characterization of two immunologically
distinct forms of [3-amylase from seeds of germinated Sorghum
hicolor (L.)Monech. Carlsberg Research Communications 47, 263-274.
77

Murao, S., Ohyama, K. and Ogura, S.(I977). Isolation of amylase inhibitor-
producing microorganism. Agriculture and Biological Chemistry 41(6),
919-924.
Murao, S., Ohyama, K. and Arai, M. (1979). |3— amylases from Bacillus
Polymyxa No. 72. Agriculture and Biological Chemistry 43(4), 719-726.
Muss, J., Brockett, F.P. and Connely, C. C. (1956). The effect of various ions on
the stability of crystalline salivary amylase. Archives of Biochemistry and
Biophysics 65, 261-268.
Myrback, K. and Neumiiler, G. (1951). In: "The Enzymes". Summer, J. B.and
Myrback, K.(eds.) 1st. ed. pp. 323-331 Academic Press, New York.
Niwa, T. ,Inouye, S., Tsuruoka,T. ,Koaze, Y. and Nida, T. (1970). "Nojirimycin"
as a potent inhibitor of glucosidase. Agricuiure and Biological Chemistry
34(6), 966-968.
Nomura, M. Maruo, B. and Akabori, J. (1956). Studies on amylase formation by
Bacillus subtilis-\- Effect of high concentration of polyethylene glycol on
amylase formation by Bacillus subtilis. Journal of Biochemistry (Tokoyo)
45, 143-152.
Norman, BE. (1979). The application of polysaccharides degrading enzyme
in the starch industry. ln:"Microbial Polysaccharides and Polysaccharase"
Berkeley, R.C.W., Goodways, G.W.and Ellwood, D.C. (eds.)pp. 339-376.
Academic Press, London.
Norman, B.E.(1981) In: "Enzyme and Food Processing". Birch, G.G.,
Blakebotougli, N. and Parker, K.J.(eds.) pp. 15-50. Applied Science
78

Publishers. London.
Novellie, L. (1960). Kaffircorn malting and brewing studies. 14. The extraction
and nature of insoluble amylases ofkaffitcorn malts. Journal of Science,
Food and Agriculture 11, 406-421.
Novellie, L. (1982a). Fermented beverages. In: "Proceeding of the international
Symposium Sorghum and Grain Quality". ICR1SAT (1981).pp 28-31.
Pantachera. India pp. 121-128.
Nyiri, L. (1971). The prepration of enzyme by fermentation
Chemical Engineering 11, 447-457.
Ogashara, K., Imanishi, A. and Isemura, T. (1970). Studies on thermophilic oc-
amylase from Bacillus stearothermophilm I- Some general and
pliysiochemical properties of thermophilic oc-amylase. Journal of
Biochemistry (Tokoyo) 67, 65-75.
Okolo, B. N., Ezeogu, L. 1. and Ebisike, CO. (1996) Raw starch digesting
amylase from Thermoactinomyces ihalpophilus F13. World Journal of
Micro-biology and Biotechnology 12, 637-638.
Okon, E.U. and Uvvaifo, A.0.(1984). Partial purification and properties of p-
amylase isolated from Sorghum hicolor. Journal of 'FoodChemistry 32,
11-14.
Olukayode, 0., Amund, O. O. and Olusola, A.O. (1987). Extracellular amylase
production by cassava- fermentation bacteria . Journal of Industrial
Microbiology 2,123-127.
79

Oten-Gyong, K., Moulin, G. and Glazy, P. (1980). Effect of medium
composition on biosynthesis and excretion of the amylase of
Sclwanniomyces castellii. European Journal of Applied Microbiology
and Biotechnology 1, 225-237.
Patel, S., Jain, N. and Madamwar, D. (1993).Production of a-amylase from
Halebaclerium haiobium. World Journal of Microbiology and
- Biotechnology 9, 25-28.
Payen, A. and Persoz, J. (1833). In:"Enzymes". Malcolm, D.and Edwin,
C.W.(eds) (1964). 2nd. ed. pp. 1-9. Academic Press, NewYork.
Pazur, ) . H., Knull, H. R. and Cepure, A. (1971). Glycoenzymes: Structure
and properties of the two forms of glucoamylase from Aspergillus niger.
Carbohydrate Research 20, 83-96.
Peat, S. , Gwen, J. T. and Whelan, W. J. (1952). The enzymic synthesis and
degradation of starch-Part XVII* Z. Enzyme. Journal of the Chemical
Society \, 722-739.
Pestana, F. and Castillo, .1.(1985). Glucoamylase production by Aspergiilus
awamori on rice flour media and partial characterization of the enzyme.
MIRCEN Journal 1, 225-237.
Peter, H. A. S., Nicholas, S. M. and Elisabeth, S. E. (1986) In: "Bergey's
Manual of Systematic Bacteriology" Vol.2, Sneath, P.M. A., Bair. N. S.,
Sharpe, M. E. and Molt, J.G. pp. 1104-1139. Williams and Wilkins,
Baltimore.
Priest, F. G.(1984). Commercial Enzymes. In:" Exlraceliuar Enzymes".
80

Aspect of Microbiology. Cole, A., Knowels, C. J. and Schelesinger, D.
eds.) Vol.9, pp. 32-50. American Society for Microbiology, Washington
DC.
Rani, R. R., Jana, S.C. and Nada, S.C. (1994). Saccharification of indigenous
starches (3-atnylase of Bacillus megaterium. World Journal of Micro-
biology and Biotechnology 10, 691-693.
Razzaque, A. and Ueda, S. (1978). Glucoamylase of Aspergillus oryzae.
Journal of Fermentation Technology 56, 296-302.
Reichelt, J. (1983). Starch.In:"Industrial Enzymology." Godfrey, T. and
Reichelt, J. (eds.) pp. 375-396. The Nature Press, New York.
Report (1958). Report of the Entrobacteriaceae Subcommittee of the
Nomenclature Committee of the International Association of Micro-
biological Society. International Bulltein of Bacteriological
Nomenclature Taxonomy 8, 25.
Robyt, J.F. and French, D.(1963). Action pattern and specificity of an amylase
from Bacillus subtil is. Archives of Biochemistry and Biophysics 100, 451-
467.
Robyt, J.F. and French, D.(1964). Purification and action pattern of an amylase
from Bacillus polymyxa. Archives of Biochemistry and Biophysics 104,
338-345.
Robyt, J.F. and Whelan, W.J. (1968). The a-amylase In: "Starch and it's
derivatives"Radely, J.A.(ed.). 4th. ed. pp 430-476 Chapman and Hall,
London.
81

Robyt, J.F. and Ackennan, R.J. (1971). Isolation, purification and
characterization of a maltotetrose producing amylase from Pseudomonas
sUitzeri. Archives of Biochemistry and Biophysics 145, 105-114.
Roch-Chui Yu and Hang, Y.D.(1990). Ainylolytic enzyme production by
Rhizopus oryzac grown on agricultural commodities. World Journal of
Microbiology and Biotechnology 6( 1), 15-18.
Satio, N. (1973). A thennophilic extracellular a-amylase from Bacillus
licheniformis. Archives of Biochemistry and Biophysics 155, 290-298.
Satio, N and Yamamoto, K. (1975). Regulatory factors affecting a-amylase
production in Bacillus licheniformis. Journal of Bacteriology 121, 848-
856.
Schwimtner, S. and Balls, A. K. (1949). In: "Enzymes". Malcolm, D. and Edwin,
C.W. (eds). (1964) 2nd. ed. pp. 802-811. Academic Press, New York.
Skinke, R. , Kunimi, Y. and Nishira, H. (1975). Production and properties of P-
amylases of Bacillus sp. BQ10. Journal of Fermentation Technology) 53,
693-697.
Solomon, B. and Levin, Y. (1974). Studies on the binding of amylogucosidase to
inert-protein. Biotechnology and Bioengineering 16, 1393-1398.
Srivastava, R. A. and Baruah, J.N. (1986). Culture conditions for the production
of thermostable amylase by Bacillus stearothermophilus. Applied and
I'jiivronmentai Microbiology 15, 179-184.
82

Stark, E. and Tetraut, P. (1951). Isolation of bacterial cell free starch
saccharifing enzyme from the medium at 70°C. Journal of Bacteriology
62, 247-249.
Steverson, E. M. Korus, R.C. Admassu, W. and Heimsuch, R.C. (1984).
Kinetics of the ainylase system of Saccharomyces Jibulegere. Itnzyme and
Microbial Technology 6, 549-554.
Svensson, B. , Moller, H. and Clark, A. J. (1988). Chemical modification of
carboxyl groups in glucoamylase from Aspergillus niger. Carlsherg
Research Communication 53, 331-342.
Takahasahi, T., Tscuchida, Y. and hie, M. (1978). Purification and some
properties of three forms of glucoamylase from a Rhizopus sp. Journal of
Biochemistry 82, 1183-1194.
Takasaki, Y (1982). Production of inaltohexaose by a-amylase from Bacillus
cirailan G6*. Agriculture and Biological Chemistry 46, 1121 -1129.
Takasaki, Y. and Takahara, Y. (1976). In:" Handbook of Enzyme
Biotechnology". Wiseman, A. (ed.)(1985). pp. 330-338. Ellis Horvvood
Publishers, New York.
Takasaki, Y., Shinohara, II., Tsuruhisa, M. Yayesh, S. and Imada,K. (1991).
Maltotetrose-producing amylase from Bacillus sp. MG-4*. Agriculture
and Biological Chemistry 55 (7), 1715-1720.
Tan?, Y. , Voungsu Vanlert, V.and Kumumanta, .1. (1986). Raw cassava starch
digestive glucoamylase of Aspergillus spp. N-2. isolated from cassava
starch. Journal of Fermentation Technology 64, 405-413.
83

Taniguchi, H., Odashima, F. , Igarashi, M. , Maruyama, Y. and Nakamura, M.
(1982). Characterization of potato starch digesting bacterium and it's
production of amylase . Agriciihiire and Biological Chemistry 46 (8),
2107-2115.
Taylor, P.M., Napier, E.J. and Fleming, I.D. (1978). Some properties of
glucoamylase produced by the thermophilic fungus Humicola lanuginosa
Carbohydrate Research 61, 301-308.
Tipples, R. H. and Tkachuk, R. (1965). Wheat-beta amylase. I-Isolation. Cereal
Chemistry 42, 111-124.
Tkachuk, R. and Tipples, R. H. (1966). Wheat beta-amylase 11. characterization.
Cereal Chemistry 43, 62-79.
Toda, H. and Narita, K. (1968). Correlation of the sulfhydral group with essential
calcium in Bacillus subtilis saccharifing a-amylase. The journal of
Biochemistry 63(3), 302-307.
Trevelayan, W.E., Procter, D. P. and Harrison, J. S. (1950). Detection of sugar
on paper chromatgrams. Nature (London) 166, 444-445.
Uchino, F. (1982). A thermophilic and unusually acidiophilic amylase produced
by a thennophilic acidiophilic Bacillus sp. Agriculture and Biological
chemistry 128, 515-521.
Underkofler, L. A. and Roy, D. K. (1951). In: " Enzymes". Malcolm, D. and
Edwin, C.W. (eds.) 2nd. ed. (1964). pp 803-811. Academic Press,
NewYork.
84

Upton, M. E. and Fogarly, W. M. (1977). Production and purification of
the thermostable amylase and protease of Thermomonospora viriciis.
Applied and Environmental Microbiology 33, 59-64.
Welker, N.E. and Campbell, L. L. (1963). Effect of carbon sources on formation
of a-amylase by Bacillus stearothermophilus. Journal of Bacteriology 86,
687-691.
Welker, N.E. and Campbell, L. L. (1967). Comparison of the a-amylase of
Bacillus subtilis and Bacillus amyloliquefacience. Journal of Bacteriology
24,1131-1135.
Whittenburry, R. (1964). Hydrogen peroxide formation and catalase activity in
the lactic acid bacteria. Journal of General Microbiology 35, 13.
Windish, M. W. and Mhatre, N. S. (1965). Microbial amylases . In:"Advances in
Applied Microbiology". Umbreit, W. W. (ed.) Vol. 7, pp 273-298.
Academic Press, NewYork.
Wiseman, A.(1985). " Handbook of Enzyme Biotechnology"2nd. ed. pp. 323-
373. Eliss Horwood Publishers, NewYork.
Yamane, K. and Maruo, B. (1974). Properties of thennosensitive extracelluar a-
amylase of Bacillus subtilis. Journal of Bacteriology 120, 792-798.
Yamasaki, Y., Suzuki, Y. and Ozawa, J. (1977a). Properties of two forms of
glucoamylase from PeniciIlium oxalicum. Agriculture and Biological
Chemistry 41(8), 1443-1449.
Yamasaki, Y., Suzuki, Y. and Ozawa, J. (1977b).Three forms of oc-glucosidase
and glucoamylase from Aspehgllus awamori Agriculture and Biological
85

Chemistry 41(11). 2149-2161.
Yamasaki,Y. and Kanno, H. (1991). Purification and properties of glucoamylase
from Mucor javanicus. Agriculture and Biological chemistry 55(9), 2429-
2430.
Yoshiyuki, T. (1982). Production of maltohexaose by a-amylase from Bacillus
circulcms G6* . Agriculture and Biological chemistry 46 (6), 1539-1547.
Yutani, K., Sasaki, I., Ogasahara, K. (1973). Comparison of thermostable a-
amylase from Bacillus stearothermophilus grown at different
temperatures. Journal of Biochemistry 73, 573-579.
86





























