Embed Size (px)
Citation preview

Doxycycline Inhibition of Interleukin-1in the Corneal Epithelium
Abraham Solomon,1 Mark Rosenblatt,1 De-Quan Li,1 Zuguo Liu,1 Dagoberto Monroy,1
Zhonghua Ji,1 Balakrishna L. Lokeshwar,2 and Stephen C. Pflugfelder1
PURPOSE. To evaluate the effect of doxycycline on the regulation of interleukin (IL)-1 expression andactivity in human cultured corneal epithelium.
METHODS. Human corneal limbal epithelium (HLE) was cultured from explants prepared from limbalrings of donor corneas. Primary cultured limbal epithelial cells were treated with either 10 mg/mllipopolysaccharide (LPS), LPS with 10 mg/ml doxycycline, or LPS with 0.1 mg/ml methylpred-nisolone (MP) for 24 hours. The intracellular and supernatant protein amounts of IL-1a, theprecursor and mature forms of IL-1b, IL-1 receptor antagonist (IL-1 RA), and the intracellular levelof IL-1b–converting enzyme (ICE) were measured with enzyme-linked immunosorbent assays(ELISAs). Western blot analysis was performed to evaluate IL-1 RA protein. mRNA steady stateamounts were determined by RNase protection assay (RPA) for IL-1a, IL-1b, IL-1 RA, and ICE.
RESULTS. LPS increased the mRNA and protein amounts of intracellular and released IL-1a, matureIL-1b, and IL-1 RA. Doxycycline inhibited the LPS-induced IL-1b increase in the mRNA and proteinamounts in the corneal epithelium and upregulated the expression of the anti-inflammatory IL-1 RAprotein. In addition, doxycycline reduced the steady state level of the cellular ICE protein but didnot affect the level of ICE transcripts. IL-1b secreted to the conditioned media of HLE wasfunctionally active in inducing matrix metalloproteinase (MMP)-1 and MMP-3 in cultured cornealfibroblasts. Doxycycline significantly decreased IL-1b bioactivity in the supernatants from LPS-treated corneal epithelial cultures. These effects were comparable to those induced by thecorticosteroid, MP.
CONCLUSIONS. Doxycycline can suppress the steady state amounts of mRNA and protein of IL-b anddecrease the bioactivity of this major inflammatory cytokine. These data may partially explain theclinically observed anti-inflammatory properties of doxycycline. The observation that doxycyclinewas equally potent as a corticosteroid, combined with the relative absence of adverse effects,makes it a potent drug for a wide spectrum of ocular surface inflammatory diseases. (InvestOphthalmol Vis Sci. 2000;41:2544–2557)
Keratitis sicca, the corneal epithelial disease that devel-ops in dry eye, is among the most common and prob-lematic conditions faced by ophthalmologists. In mild
cases, it is associated with symptoms of irritation, redness, andblurred vision. In the more severe forms, sight-threateningcorneal problems may develop, such as filamentary keratitis,corneal epithelial erosions, corneal stromal vascularization,and ulceration. The exact mechanism by which keratitis siccadevelops has not been established. Our group has reportedthat inflammation may be the primary factor causing this con-
dition.1 The proinflammatory cytokine interleukin (IL)-1 hasbeen identified as a factor that may play a key role in theinitiation and perpetuation of this inflammation. We have ob-served that as tear clearance from the ocular surface decreases,the concentrations of both isoforms of the proinflammatorycytokine IL-1, IL-1a,2 and IL-1b (Solomon et al., unpublishedresults, 2000), increase in the tear fluid. The IL-1 gene family isa group of potent cytokines that function as major mediators ofinflammation and immune response.3 This family is composedof three forms: two proinflammatory forms, IL-1a and IL-1b,each having a precursor form, and an anti-inflammatory form,IL-1 receptor antagonist (IL-1 RA).
Recent data suggest that the IL-1 cytokines play an impor-tant role in the regulation of inflammation and wound healingon the ocular surface. IL-1b was found in the epithelium,stroma, and endothelium of the cornea, at the mRNA andprotein levels.4 Type 1 receptor for IL-1 is expressed in stromalfibroblasts.5 Both IL-1a and IL-1b have been found to modulatematrix metalloproteinase (MMP) expression by corneal stromalfibroblasts6 and their own synthesis in keratocytes,7 to regulateapoptosis of keratocytes in response to corneal epithelialwounding,8 and to upregulate hepatocyte growth factor andkeratocyte growth factor in corneal fibroblasts.4 These findings
From the 1Ocular Surface and Tear Center, Bascom Palmer EyeInstitute, Department of Ophthalmology; and 2Department of Urology,University of Miami School of Medicine, Florida.
Supported in part by Public Health Service Research GrantEY11915 (SCP); Department of Health and Human Services, NationalEye Institute Grant CA61038 (BLL); National Cancer Institute; an un-restricted Grant from Research to Prevent Blindness; and the Drs.David and Maureen Smith Ocular Surface and Tear Research Fund.
Submitted for publication November 23, 1999; revised February23, 2000; accepted March 17, 2000.
Commercial relationships policy: N.Corresponding author: Stephen C. Pflugfelder, Bascom Palmer Eye
Institute, 900 NW 17th Street, Miami, FL [email protected]
Investigative Ophthalmology & Visual Science, August 2000, Vol. 41, No. 92544 Copyright © Association for Research in Vision and Ophthalmology

make IL-1 a prime candidate for inducing ocular surface dis-ease, especially the chronic subclinical ocular surface inflam-mation of dry eye.
Traditional therapies for keratoconjunctivitis have con-sisted of artificial tears and aqueous-conserving therapies, suchas punctal occlusion. Although these therapies transiently im-prove irritation symptoms, they are often ineffective in treatingthe severe complications of dry eye, such as recurrent cornealepithelial erosion and corneal stromal ulceration. Therapiestargeting the underlying inflammatory environment of the oc-ular surface would represent a major improvement in themanagement of these conditions and would have a majorclinical impact. Consistent with the concept that inflammationis a key feature in the pathophysiology of keratitis sicca is thefinding that both aqueous tear deficiency and meibomian glanddisease are effectively treated with the corticosteroid methyl-prednisolone (MP).9,10 Unfortunately the long-term use of top-ical corticosteroids is limited by potential sight-threatening sideeffects, such as glaucoma and cataract. Therefore, there is aclinical need for nontoxic steroid-sparing anti-inflammatorytherapies that target IL-1 expression in the corneal epithelium.
Systemically administered tetracycline antibiotics havelong been recognized as effective therapies for ocular surfaceinflammatory diseases. The semisynthetic tetracycline, doxycy-cline, has been reported to successfully treat the common dryeye condition acne rosacea,11 as well as recurrent cornealerosions12 and phlyctenular keratoconjunctivitis.13 We hypoth-esized that one of the mechanisms of action of doxycycline indry eye is the downregulation of the IL-1–mediated inflamma-tory cascade in the corneal epithelium. Therefore, the purposeof this study was to evaluate the effect of doxycycline on theregulation of IL-1 expression and activity in the human cornealepithelium.
METHODS
Reagents
Dulbecco’s modified Eagle’s medium (DMEM), fetal bovineserum (FBS), HEPES buffer, F12 (Ham’s), were from Life Tech-nologies (Rockville, MD). Tissue culture plates were from Bec-ton Dickinson (Franklin Lakes, NJ). Cholera toxin subunit A,epidermal growth factor (EGF), hydrocortisone, LPS (derivedfrom Serratia marcescens), doxycycline, and MP were fromSigma (St. Louis, MO). IL-b precursor, IL-1b–converting en-zyme (ICE), and IL-b mature enzyme-linked immunosorbentassay (ELISA) kits were from Cistron (Pine Brook, NJ); IL-1 aand IL-1 RA ELISA kits were from R&D systems (Minneapolis,MN); and MMP-1 and MMP-3 ELISA kits were from OncogeneResearch Products of Calbiochem (Cambridge, MA). RNA lysisand RNase protection kits were from Ambion (Austin, TX). IL-1RA was from Genzyme (Cambridge, MA). The BCA proteinassay kit was from Pierce (Rockford, IL).
Culture of Human Corneal Limbal Epithelium
Human corneal limbal epithelium (HLE) was cultured fromexplants of human donor corneoscleral rims, provided by theFlorida Lions Eye Bank at the Bascom Palmer Eye Institute.Each corneoscleral rim was trimmed, the endothelial layer andiris remnants were removed, and the tissue was treated withdispase for 15 minutes. Each rim was dissected into 12 equal
parts, which were applied to six-well plastic dishes and cov-ered with a drop of FBS overnight. The explants were culturedin supplemented hormonal epithelial medium (SHEM) contain-ing equal amounts of DMEM and Ham’s F12 medium, supple-mented with 5% FBS, 0.5% dimethyl sulfoxide, 2 ng/ml EGF, 5mg/ml insulin, 5 mg/ml transferrin, 5 ng/ml selenium, 0.5 mg/mlhydrocortisone, 30 ng/ml cholera toxin A, 50 mg/ml gentami-cin, and 1.25 mg/ml amphotericin B. Cultures were incubatedat 37°C under 95% humidity and 5% CO2. The medium waschanged every 2 days. Cultures were maintained for 10 to 14days until confluence and then switched to the serum-freemedium described above, without FBS, for 24 hours before theadditions of treatments.
To demonstrate the effect of doxycycline on the cornealepithelium and to compare it with that of a corticosteroid,primary cultures of HLE were treated with 10 mg/ml bacterialLPS alone or in combination with either 0.1 mg/ml MP or 10mg/ml doxycycline. These treatments were maintained for 24hours.
After a 24-hour treatment, the culture supernatant wascollected from each well, centrifuged, and stored in 280°Cuntil assayed by ELISA. Cell lysis solution, containing 50 mMTris-HCl (pH 7.6), 300 mM NaCl, and 0.5% Triton X-100, wasadded to the cells for 3 hours, and the cellular protein wascollected, centrifuged, and stored in 280°C until assayed byELISA. In parallel cultures, the cells were subjected to lysisbuffer (Direct Protect; Ambion), and total RNA was isolated forfurther assessment by RNase protection assay (RPA). The ELISAand RPA were targeted at determining the protein and mRNAlevels, respectively, of IL-1a, IL-1b, IL-1 RA, and ICE.
RPA Template Construction
Partial cDNAs for human IL-1a, IL-1b, IL-1 RA, and glyceralde-hyde-3-phosphate dehydrogenase (GAPDH) were prepared byreverse transcription–polymerase chain reaction (RT-PCR).PolyA1 RNA was isolated from cultured human corneal epi-thelial cells using oligo-dT–coated beads (Oligotex DirectmRNA Isolation System; Qiagen, Valencia, CA), according tothe manufacturer’s instructions. RT was performed using 200ng mRNA as template and gene-specific primers (see Table 1)were prepared to human IL-1a (gene accession, X02531), IL-1b(gene accession, X02532), IL-1 RA (gene accession, M63099),and GAPDH (gene accession, NM 002046), according to themanufacturer’s instructions (Superscript II Reverse Transcrip-tion Kit; Life Technologies). The resultant first-strand cDNAwas used for PCR (PCR Kit, Life Technologies) using a gene-specific upstream primer and the same downstream primer-2used for RT. An aliquot of the initial PCR reaction (except forthe GAPDH probe that required only a single round of PCR)was reamplified using the same upstream primer and a thirdgene-specific primer, downstream primer-1. The primers usedin this second amplification were designed with 12 nucleotideadditions (four copies of a trinucleotide repeat containing asingle deoxyuracil in each repeat) at their 59 ends, to facilitaterapid cloning of the amplimers (pAMP1 System; Life Technol-ogies).
RPA Probes
RPA requires hybridization of sense mRNA to complementaryradiolabeled antisense RNA. Subsequently, double-stranded
IOVS, August 2000, Vol. 41, No. 9 Doxycycline Inhibition of IL-1 in Corneal Epithelium 2545

mRNA-radiolabeled antisense RNA hybrids are treated withRNase specific for single-stranded RNA. Protected hybrids canthen be resolved and quantified using gel electrophoresis andautoradiography. Radiolabeled antisense RNA was transcribedusing the a kit (Maxiscript T7; Ambion) and labeling with[a-32P]CTP (800 Ci/mmol). Plasmids were digested at a uniqueBamHI site upstream of the cloned cDNAs. RNA probes weregenerated for IL-1a, IL-1b, and IL-1 RA, and GAPDH. TheGAPDH message is a housekeeping gene that is found to beexpressed 10 to 20 times more than the messages for thecytokines measured in the RPA. The GAPDH probe was there-fore transcribed to yield a specific activity 10 times less thanthat of the cytokine probes, to allow simultaneous detection ofprotected cytokine probe fragments as well as GAPDH probefragments, given the range of sensitivity provided by the x-rayfilm. After transcription, probes were DNase treated to removetemplate DNA, and unincorporated nucleotides were removedusing RNA Quick Spin columns (Roche Molecular Biochemi-cals, Indianapolis, IN). A template set containing DNA tem-plates for ICE and GAPDH RNA probes, was purchased fromPharMingen (San Diego, CA).
RNase Protection Assay for IL-1a, IL-1b, IL-1 RA,and ICE
RNase protection assays were performed (Direct Protect Sys-tem; Ambion) as described by the manufacturer. Briefly, cul-tured human corneal epithelial cells were resuspended in lysisbuffer at approximately 107 cells/ml. The cell lysis buffer in-cluded concentrated guanidine thiocyanate, which rapidlysolubilizes cells and also rapidly inactivates ribonucleases. As-says were performed using 50 ml of cell lysate, 105 cpm of eachcytokine probe (specific activity, 5 3 10 5 cpm/mg) and 4 3104 cpm of the GAPDH probe (specific activity, 5 3 10 4
cpm/mg) for each sample. Samples were allowed to hybridizeovernight at 37°C. They were then treated for 30 minutes at37°C with RNase solution, after which the RNase was inacti-vated with proteinase K. Protected RNA fragments were pre-cipitated and separated on a 6% polyacrylamide urea-Tris-base,boric acid, EDTA (TBE) sequencing gel.
RPAs for IL-1a, IL-1b, and IL-1 RA were repeated fourtimes on primary cultures derived from four different donorcorneas. RPAs for ICE were repeated three times on primarycultures from three donor corneas. Autoradiographs fromthese gels were scanned and then analyzed using image anal-ysis software (Gel-Pro; Media Cybernetics, Silver Spring, MD).The digitized data for each band was plotted, and the area
under the curve for each peak was calculated with statisticalsoftware (GraphPad Prism; GraphPad Software, San Diego,CA). The value for each cytokine band was divided by thecorresponding value of the GAPDH band in the same lane tocalculate the relative mRNA amount for each gene. Results areshown as means 6 SEM of relative mRNA amounts from threeor four experiments.
Immunodetection of IL-1b Precursor, IL-1bMature, IL-1a, IL-1 RA, and ICE
The conditioned media and cell lysates of corneal limbal epi-thelial cells from four independent primary cultures, derivedfrom four different donor corneas were collected, centrifuged,and stored at 280°C until assayed. The concentrations of IL-1bprecursor and IL-1b mature in the cell lysates and in thesupernatants and of ICE in cell lysates were measured byELISAs according to the respective manufacturer’s protocol.The cellular protein concentration in cell lysates was measuredwith the BCA protein assay kit.
Western Blot Analysis for IL-1 RA
To evaluate the expression of IL-1 RA protein in the condi-tioned medium and in the cells, we further incubated primarylimbal epithelium with LPS, LPS and MP, LPS and doxycycline,or LPS with MP and doxycycline, using the same concentra-tions as described earlier.
Cell lysates and conditioned media containing equal quan-tities of protein were subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) using 4% to15%, 0.75-mm thick polyacrylamide gel (Mini-ready; Bio-Rad,Hercules, CA) at a constant 200 V for 45 minutes, in a mini-protean electrophoresis apparatus (Bio-Rad). A positive control(human recombinant IL-1 RA; R&D) and prestained (7.5–203kDa) molecular weight protein markers (Bio-Rad) were runsimultaneously with the samples. Resolved proteins were trans-ferred to nitrocellulose membranes (BioTrance NT, Ann Arbor,MI) using a minitank blot apparatus (Bio-Rad). Membraneswere blocked in 3% fat-free milk for 45 minutes After a 1-hourincubation with polyclonal rabbit anti-human IL-1 RA antibodydiluted in 1% bovine serum albumin, Tris-buffered saline, and0.5% Tween 20, the membranes were incubated with IgG-horseradish peroxidase–conjugated goat anti-rabbit (Sigma) di-luted 1:80,000 in 1% bovine serum albumin, Tris-buffered sa-line, and 0.5% Tween 20. Signals were detected with animmunodetection kit (Renaissance Enhanced Chemilumines-cence [ECL]; NEN Life Science Products, Boston, MA), and
TABLE 1. Primers Used in RNAse Protection Assay
cDNA Size (bp) Upstream Primer (5*–3*) Downstream Primer-1 (5*–3*) Downstream Primer-2 (5*–3*)
IL-1a 407 CAA GGA GAG CAT GGT GGTAGT AGC AAC CAA CG
GCA CTG GTT GGT CTT CATCTT GGG C
TAG TGC CGT GAG TTT CCCAGA AGA AGA GGA GG
IL-1b 348 GCT ACG AAT CTC CGA CCACCA CTA CAG C
CCT TGT ACA AAG GAC ATGGAG AAC ACC
CTT ATC ATC TTT CAA CACGCA GGA CAG G
IL-1 RA 308 CCA TTC AGA GAC GAT CTGCCG ACC
GCT TGT CCT GCT TTC TGTTCT CGC
CTG TCT GAG CGG ATG AAGGCG AAG C
GAPDH 188 GAC ATC AAG AAG GTG GTGAAG CAG GC
CCA AAT TCG TTG TCA TACCAG GAA ATG AGC
2546 Solomon et al. IOVS, August 2000, Vol. 41, No. 9
then exposed to x-ray film (Eastman Kodak, Rochester, NY)from 30 seconds to 3 minutes.
IL-1b Activity Assay
To evaluate the functional activity of IL-1b secreted by culturedHLE, we sought to develop a bioassay that is based on theinduction of MMP-1 and MMP-3 in corneal fibroblasts by IL-1b.6
IL-1b has been previously demonstrated to induce MMP-1 and
MMP-3 secretion in human synovial fibroblasts,14 endometrialstromal cells,15 and fibrochondrocytes.16 We therefore cul-tured early-passaged corneal fibroblasts in conditioned mediathat were collected from the HLE cultures. The resultantMMP-1 and -3 supernatant concentrations were measured.
Corneal fibroblasts were cultured as previously de-scribed.17 Briefly, the central corneas of donor eye bank eyeswere isolated with a 6-mm trephine after removal of the epi-
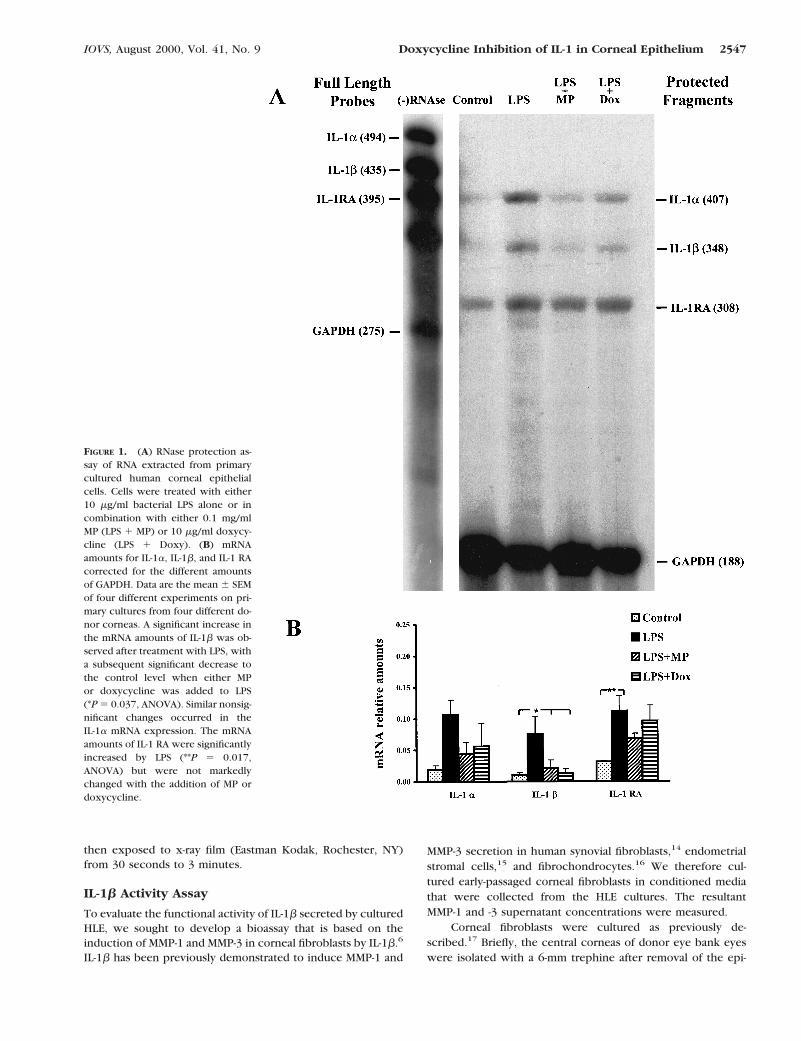
FIGURE 1. (A) RNase protection as-say of RNA extracted from primarycultured human corneal epithelialcells. Cells were treated with either10 mg/ml bacterial LPS alone or incombination with either 0.1 mg/mlMP (LPS 1 MP) or 10 mg/ml doxycy-cline (LPS 1 Doxy). (B) mRNAamounts for IL-1a, IL-1b, and IL-1 RAcorrected for the different amountsof GAPDH. Data are the mean 6 SEMof four different experiments on pri-mary cultures from four different do-nor corneas. A significant increase inthe mRNA amounts of IL-1b was ob-served after treatment with LPS, witha subsequent significant decrease tothe control level when either MPor doxycycline was added to LPS(*P 5 0.037, ANOVA). Similar nonsig-nificant changes occurred in theIL-1a mRNA expression. The mRNAamounts of IL-1 RA were significantlyincreased by LPS (**P 5 0.017,ANOVA) but were not markedlychanged with the addition of MP ordoxycycline.
IOVS, August 2000, Vol. 41, No. 9 Doxycycline Inhibition of IL-1 in Corneal Epithelium 2547
thelium and the endothelium with a cell scraper. Explantcultures were prepared in the same manner as described ear-lier for limbal epithelial culture, except that DMEM contain-ing 10% FBS (D-FBS) was used. Cultures were incubated at37°C under 95% humidity and 5% CO2, and the mediumwas changed twice a week. Fibroblasts were subculturedwith 0.05% trypsin and 0.85 mM EDTA in a calcium-free MEMat 80% to 90% confluence with 1:3 to 1:4 split for threepassages.
Third-passage fibroblasts were seeded in six-well tissueculture plates at a density of 2 3 105 cells per well. After 5 daysin culture, on confluence, cultures were switched to a serum-free medium containing DMEM supplemented with 5 mg/mlinsulin, 5 mg/ml transferrin, 5 ng/ml selenium, 50 mg/ml gen-tamicin, and 1.25 mg/ml amphotericin B (D-ITS). Some cultureswere maintained in D-FBS. After a 24-hour incubation in D-ITS,cultures were divided into two groups.
The first group of cultures served as a positive control andwere treated directly with human recombinant IL-1b with one
of the following: D-ITS alone, recombinant human (rh)-matureIL-1b (10 ng/ml), rh-pre-IL-1b (10 ng/ml), and rh-pre-IL-1b (10ng/ml) with matrix MMP-9 (1 mg/ml).
The second group of cultures were treated with condi-tioned media (CM) derived from HLE cultures that had beentreated as described earlier. These treatments included CMfrom HLE culture treated with medium alone (CM-SHEM), CMfrom HLE culture treated with LPS (CM-LPS), CM from HLEculture treated with LPS and doxycycline (CM-LPS 1 doxy),and CM from HLE culture treated with doxycycline (CM-doxy).To exclude the possibility that the drugs contained in theconditioned media (LPS and doxycycline) altered MMP secre-tion in corneal fibroblasts, two additional treatments wereadded: SHEM with LPS (10 mg/ml; SHEM-LPS) and SHEM withLPS and doxycycline (10 mg/ml each; SHEM-LPS 1 doxy).
Cultures were incubated with one of these treatments for24 hours, and thereafter their supernatants were collectedfor measurement of MMP-1 and MMP-3 concentrations byELISAs.
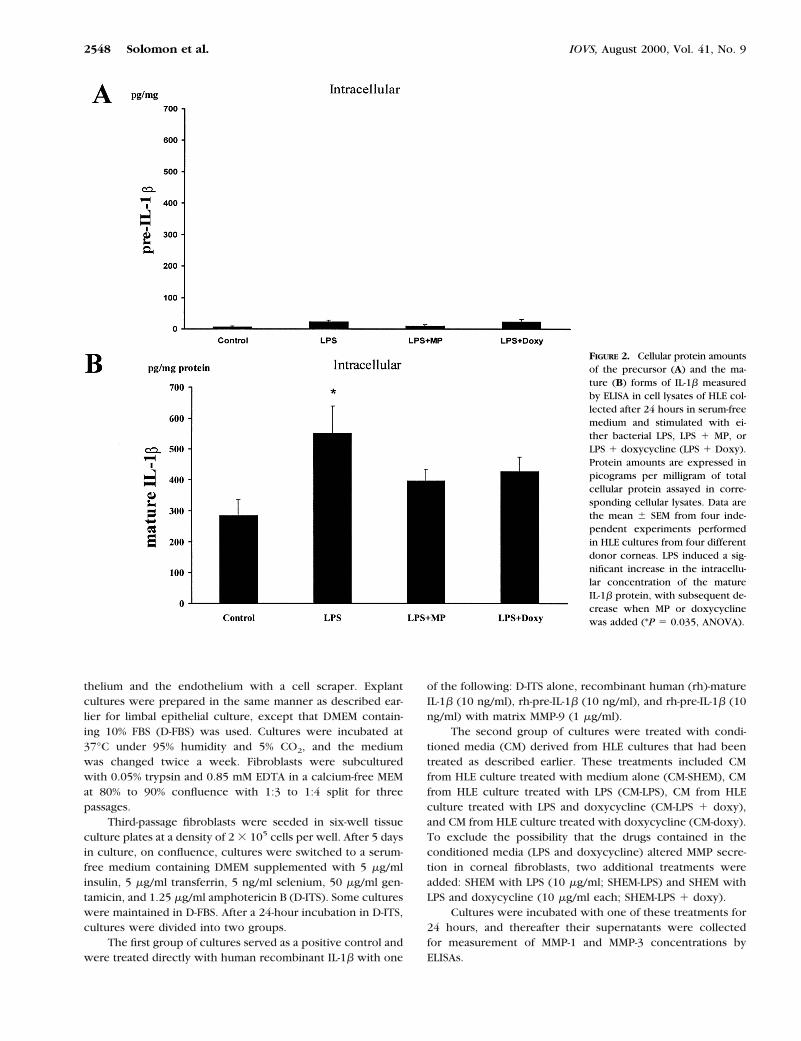
FIGURE 2. Cellular protein amountsof the precursor (A) and the ma-ture (B) forms of IL-1b measuredby ELISA in cell lysates of HLE col-lected after 24 hours in serum-freemedium and stimulated with ei-ther bacterial LPS, LPS 1 MP, orLPS 1 doxycycline (LPS 1 Doxy).Protein amounts are expressed inpicograms per milligram of totalcellular protein assayed in corre-sponding cellular lysates. Data arethe mean 6 SEM from four inde-pendent experiments performedin HLE cultures from four differentdonor corneas. LPS induced a sig-nificant increase in the intracellu-lar concentration of the matureIL-1b protein, with subsequent de-crease when MP or doxycyclinewas added (*P 5 0.035, ANOVA).
2548 Solomon et al. IOVS, August 2000, Vol. 41, No. 9
Measurement of ICE Activity
The specific activity of ICE in cell lysates was measured, aspreviously described,18 by incubating lysates with the flu-orogenic substrate AcTyr-Val-Ala-Asp-AMC (YVAD-AMC; Up-
state Biotechnology, Lake Placid, NY) in buffer containing100 mM HEPES, 10% sucrose, 0.1% NP40, and 10 mM dithio-threitol (pH 7.5) for 1 hour at 37°C. Fluorescence wasdetermined with a fluorometer at 360 nm (excitation) and530 nm (emission).
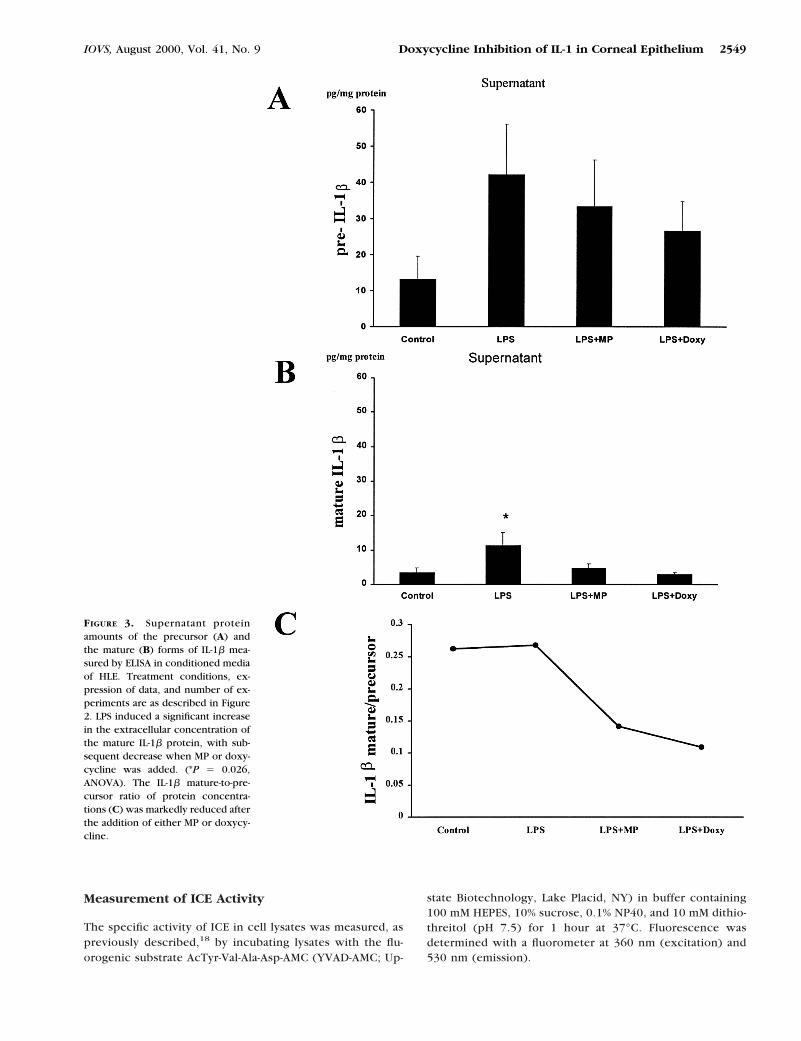
FIGURE 3. Supernatant proteinamounts of the precursor (A) andthe mature (B) forms of IL-1b mea-sured by ELISA in conditioned mediaof HLE. Treatment conditions, ex-pression of data, and number of ex-periments are as described in Figure2. LPS induced a significant increasein the extracellular concentration ofthe mature IL-1b protein, with sub-sequent decrease when MP or doxy-cycline was added. (*P 5 0.026,ANOVA). The IL-1b mature-to-pre-cursor ratio of protein concentra-tions (C) was markedly reduced afterthe addition of either MP or doxycy-cline.
IOVS, August 2000, Vol. 41, No. 9 Doxycycline Inhibition of IL-1 in Corneal Epithelium 2549
Statistical Analysis
Results are expressed as mean 6 SEM of at least three experi-ments performed on cultures derived from at least three differentdonor corneas, respectively. Statistical analysis was performedusing Student’s t-test or one-way analysis of variance (ANOVA)where appropriate. P , 0.05 was considered significant.
RESULTS
Expression of the IL-1 Genes in the HumanCorneal Epithelium
All three genes of the IL-1 family: IL-1a, IL-1b, and IL-1 RA weredemonstrated at the RNA and protein levels in primary cultures
of human corneal limbal epithelium (Fig. 1A, first lane; Figs. 2and 3, 5 and 6). Both precursor and mature forms of IL-1b werefound in these cultured epithelial cells and their supernatants.Considerably higher concentrations of the mature IL-1b werefound within the cells (Figs. 2A, 2B), whereas higher precursorIL-1b concentrations were found in the extracellular environ-ment (Figs. 3A, 3B).
Regulation of IL-1b by Endotoxin, Doxycycline,and MP
The IL-1b mRNA was expressed at very low levels in thenonactivated corneal epithelium. However, when stimulatedwith LPS, it was significantly upregulated (sevenfold; P 50.037) when compared with its baseline level (Figs. 1A, 1B).
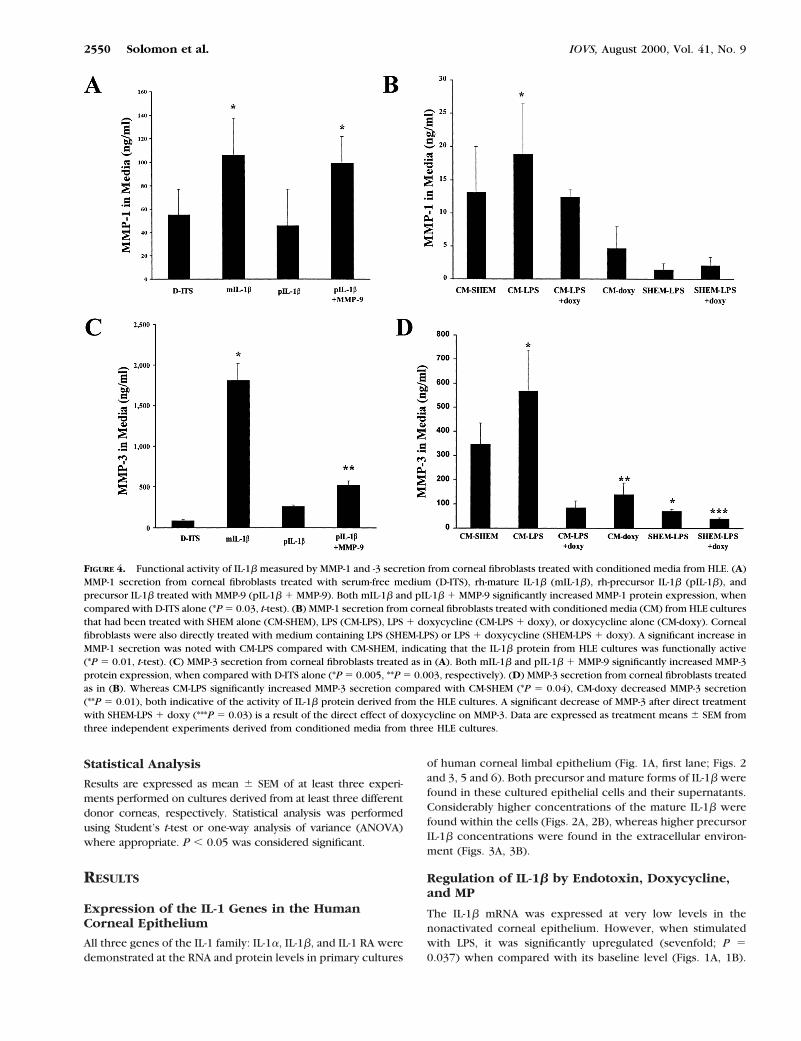
FIGURE 4. Functional activity of IL-1b measured by MMP-1 and -3 secretion from corneal fibroblasts treated with conditioned media from HLE. (A)MMP-1 secretion from corneal fibroblasts treated with serum-free medium (D-ITS), rh-mature IL-1b (mIL-1b), rh-precursor IL-1b (pIL-1b), andprecursor IL-1b treated with MMP-9 (pIL-1b 1 MMP-9). Both mIL-1b and pIL-1b 1 MMP-9 significantly increased MMP-1 protein expression, whencompared with D-ITS alone (*P 5 0.03, t-test). (B) MMP-1 secretion from corneal fibroblasts treated with conditioned media (CM) from HLE culturesthat had been treated with SHEM alone (CM-SHEM), LPS (CM-LPS), LPS 1 doxycycline (CM-LPS 1 doxy), or doxycycline alone (CM-doxy). Cornealfibroblasts were also directly treated with medium containing LPS (SHEM-LPS) or LPS 1 doxycycline (SHEM-LPS 1 doxy). A significant increase inMMP-1 secretion was noted with CM-LPS compared with CM-SHEM, indicating that the IL-1b protein from HLE cultures was functionally active(*P 5 0.01, t-test). (C) MMP-3 secretion from corneal fibroblasts treated as in (A). Both mIL-1b and pIL-1b 1 MMP-9 significantly increased MMP-3protein expression, when compared with D-ITS alone (*P 5 0.005, **P 5 0.003, respectively). (D) MMP-3 secretion from corneal fibroblasts treatedas in (B). Whereas CM-LPS significantly increased MMP-3 secretion compared with CM-SHEM (*P 5 0.04), CM-doxy decreased MMP-3 secretion(**P 5 0.01), both indicative of the activity of IL-1b protein derived from the HLE cultures. A significant decrease of MMP-3 after direct treatmentwith SHEM-LPS 1 doxy (***P 5 0.03) is a result of the direct effect of doxycycline on MMP-3. Data are expressed as treatment means 6 SEM fromthree independent experiments derived from conditioned media from three HLE cultures.
2550 Solomon et al. IOVS, August 2000, Vol. 41, No. 9
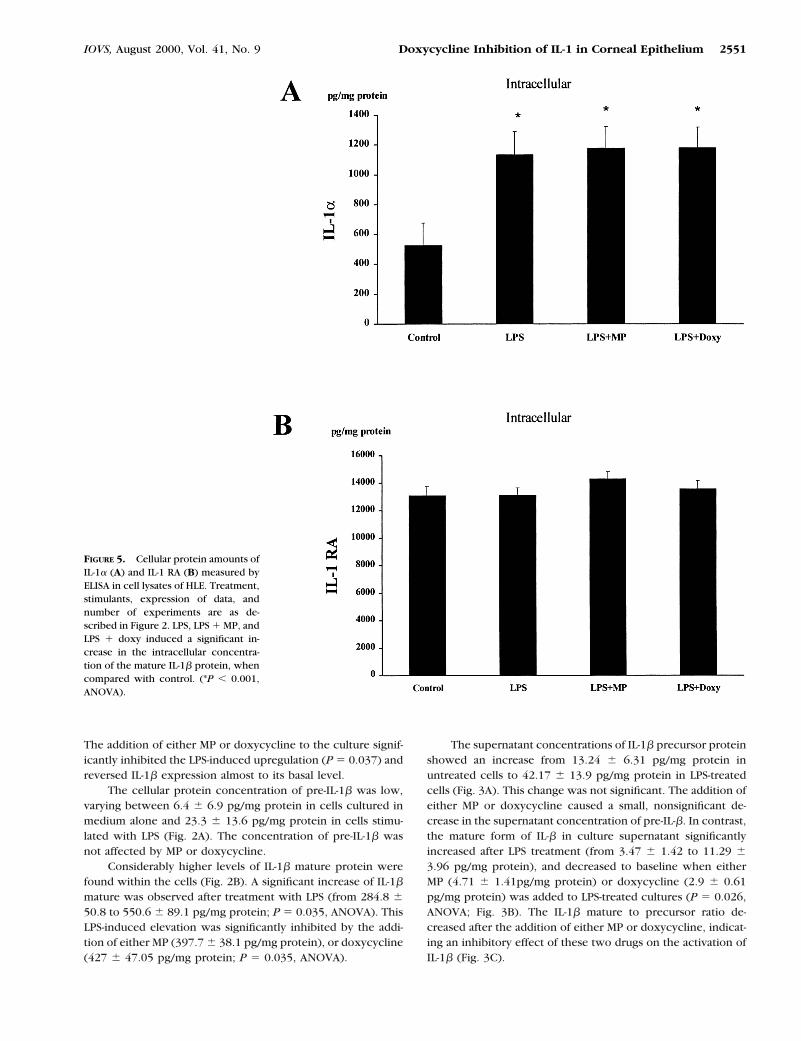
The addition of either MP or doxycycline to the culture signif-icantly inhibited the LPS-induced upregulation (P 5 0.037) andreversed IL-1b expression almost to its basal level.
The cellular protein concentration of pre-IL-1b was low,varying between 6.4 6 6.9 pg/mg protein in cells cultured inmedium alone and 23.3 6 13.6 pg/mg protein in cells stimu-lated with LPS (Fig. 2A). The concentration of pre-IL-1b wasnot affected by MP or doxycycline.
Considerably higher levels of IL-1b mature protein werefound within the cells (Fig. 2B). A significant increase of IL-1b
mature was observed after treatment with LPS (from 284.8 650.8 to 550.6 6 89.1 pg/mg protein; P 5 0.035, ANOVA). ThisLPS-induced elevation was significantly inhibited by the addi-tion of either MP (397.7 6 38.1 pg/mg protein), or doxycycline(427 6 47.05 pg/mg protein; P 5 0.035, ANOVA).
The supernatant concentrations of IL-1b precursor proteinshowed an increase from 13.24 6 6.31 pg/mg protein inuntreated cells to 42.17 6 13.9 pg/mg protein in LPS-treatedcells (Fig. 3A). This change was not significant. The addition ofeither MP or doxycycline caused a small, nonsignificant de-crease in the supernatant concentration of pre-IL-b. In contrast,the mature form of IL-b in culture supernatant significantlyincreased after LPS treatment (from 3.47 6 1.42 to 11.29 63.96 pg/mg protein), and decreased to baseline when eitherMP (4.71 6 1.41pg/mg protein) or doxycycline (2.9 6 0.61pg/mg protein) was added to LPS-treated cultures (P 5 0.026,ANOVA; Fig. 3B). The IL-1b mature to precursor ratio de-creased after the addition of either MP or doxycycline, indicat-ing an inhibitory effect of these two drugs on the activation ofIL-1b (Fig. 3C).
FIGURE 5. Cellular protein amounts ofIL-1a (A) and IL-1 RA (B) measured byELISA in cell lysates of HLE. Treatment,stimulants, expression of data, andnumber of experiments are as de-scribed in Figure 2. LPS, LPS 1 MP, andLPS 1 doxy induced a significant in-crease in the intracellular concentra-tion of the mature IL-1b protein, whencompared with control. (*P , 0.001,ANOVA).
IOVS, August 2000, Vol. 41, No. 9 Doxycycline Inhibition of IL-1 in Corneal Epithelium 2551
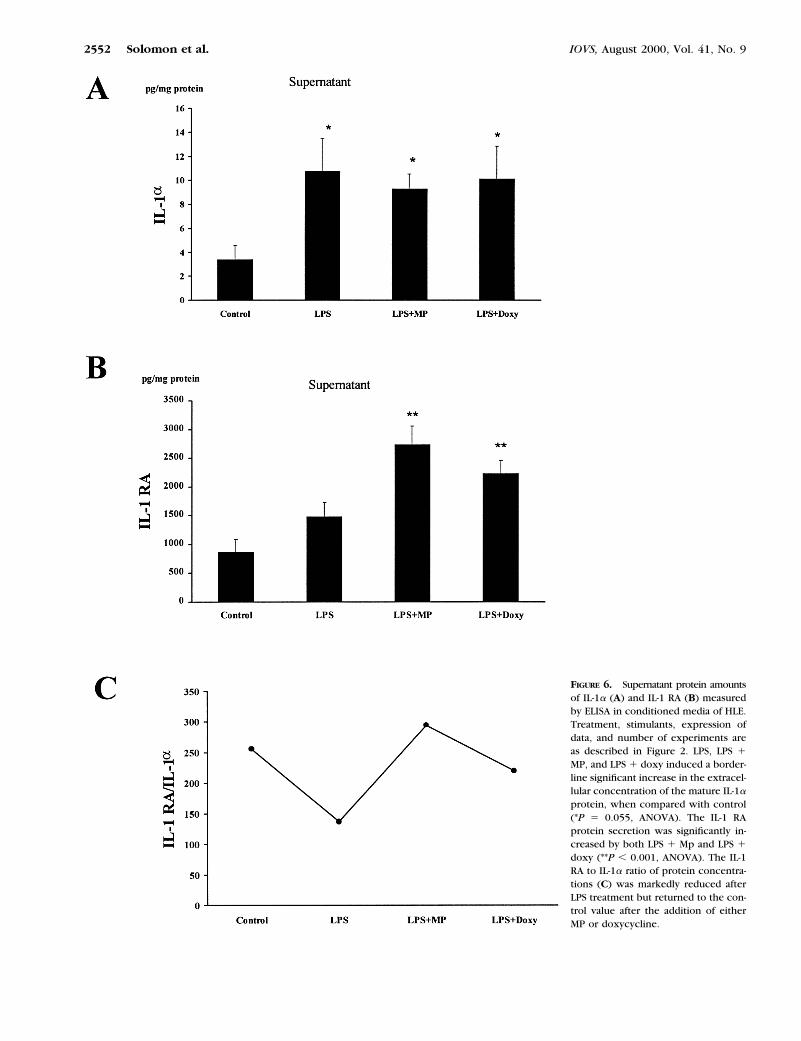
FIGURE 6. Supernatant protein amountsof IL-1a (A) and IL-1 RA (B) measuredby ELISA in conditioned media of HLE.Treatment, stimulants, expression ofdata, and number of experiments areas described in Figure 2. LPS, LPS 1MP, and LPS 1 doxy induced a border-line significant increase in the extracel-lular concentration of the mature IL-1aprotein, when compared with control(*P 5 0.055, ANOVA). The IL-1 RAprotein secretion was significantly in-creased by both LPS 1 Mp and LPS 1doxy (**P , 0.001, ANOVA). The IL-1RA to IL-1a ratio of protein concentra-tions (C) was markedly reduced afterLPS treatment but returned to the con-trol value after the addition of eitherMP or doxycycline.
2552 Solomon et al. IOVS, August 2000, Vol. 41, No. 9

Functional Activity of IL-1b Secreted fromCultured Corneal Epithelium
Mature IL-1b, but not precursor IL-1b, significantly increasedMMP-1 and MMP-3 secretion by corneal fibroblasts comparedwith the media control (D-ITS), showing that this is a feasiblemethod for determining IL-1b bioactivity (Figs. 4A, 4C). Prein-cubation of precursor IL-1b with MMP-9, an MMP that isknown to process precursor IL-1b to the mature biologicallyactive form,19 resulted in MMP-1 and MMP-3 levels comparableto those observed with mature IL-b. MMP-1 and -3 were de-tected in corneal fibroblast cultures treated with HLE-condi-tioned media (Figs. 4B, 4D). Significantly increased productionof these MMPs was observed when the LPS-treated HLE-condi-tioned media were added. Treatment of the fibroblast cultureswith conditioned media from LPS 1 doxycycline–treated HLEabrogated this LPS-induced increase (Figs. 4B, 4D). In contrast,direct treatment of the corneal fibroblasts with LPS resulted innegligible protein concentrations, excluding a direct effect ofdrug that remained in the HLE-conditioned media.
Regulation of IL-1a by Endotoxin and MP
The mRNA of IL-1a showed upregulation after stimulation withLPS. This increase was partially reversed when either MP ordoxycycline was added to LPS (Fig. 1B). The changes in themRNA amounts for IL-1a were not statistically significant, al-though they followed a trend similar to that of IL-1b.
The intracellular IL-1a protein concentration significantlyincreased from 525 6 151 to 1134 6 155 pg/mg protein (P ,0.001, ANOVA) in LPS-treated cultures and remained highwhen either MP or doxycycline was added to the culture (Fig.5A). A similar trend was observed in the culture supernatants(Fig. 6A). LPS increased IL-1a concentration from 3.4 6 1.2 to10.7 6 2.7 pg/mg protein (P 5 0.055), but neither MP nordoxycycline affected the concentration of this cytokine. Thecellular concentration of IL-1 a protein in corneal epithelialcells was 10-fold higher than the concentration released intothe culture medium.
Effect of Doxycycline on IL-1 RA
IL-1 receptor antagonist was upregulated by LPS alone or witheither MP or doxycycline at the mRNA and protein levels. LPSinduced a 3.5-fold increase of mRNA expression (P 5 0.017),which was partially inhibited when MP was added but wasunchanged when doxycycline was added to LPS (Fig. 1B).
No differences were noted between the four culturegroups in the concentrations of the intracellular IL-1 RAprotein (Fig. 5B). However, the concentrations of IL-1 RA inthe cultures supernatants demonstrated a significant in-crease when either MP or doxycycline was added to LPS(862 6 222, 2730 6 333, and 2230 6 229 pg/mg protein forcontrol, LPS 1 MP, and LPS 1 doxycycline, respectively(P , 0.001; Fig. 6B).
Western blot analysis for IL-1 RA demonstrated a markedincrease in the bands for culture supernatants when doxycy-cline was added to LPS. The combination of doxycycline andMP added to LPS demonstrated the highest IL-1 RA proteinexpression (Fig. 7, top). The expression of the intracellular IL-1RA was not changed by any of the treatments, when comparedwith medium alone (Fig. 7, bottom). These data correspondwith the ELISA data, showing significant changes in secretedIL-1 RA, whereas the intracellular expression was stable (Figs.5B, 6B).
At the supernatant level, the IL-1 RA-to-IL-1a ratio mark-edly decreased after the addition of LPS alone, but addition ofeither of these two drugs to LPS returned the ratio to thatobserved in the control cultures (Fig. 6C).
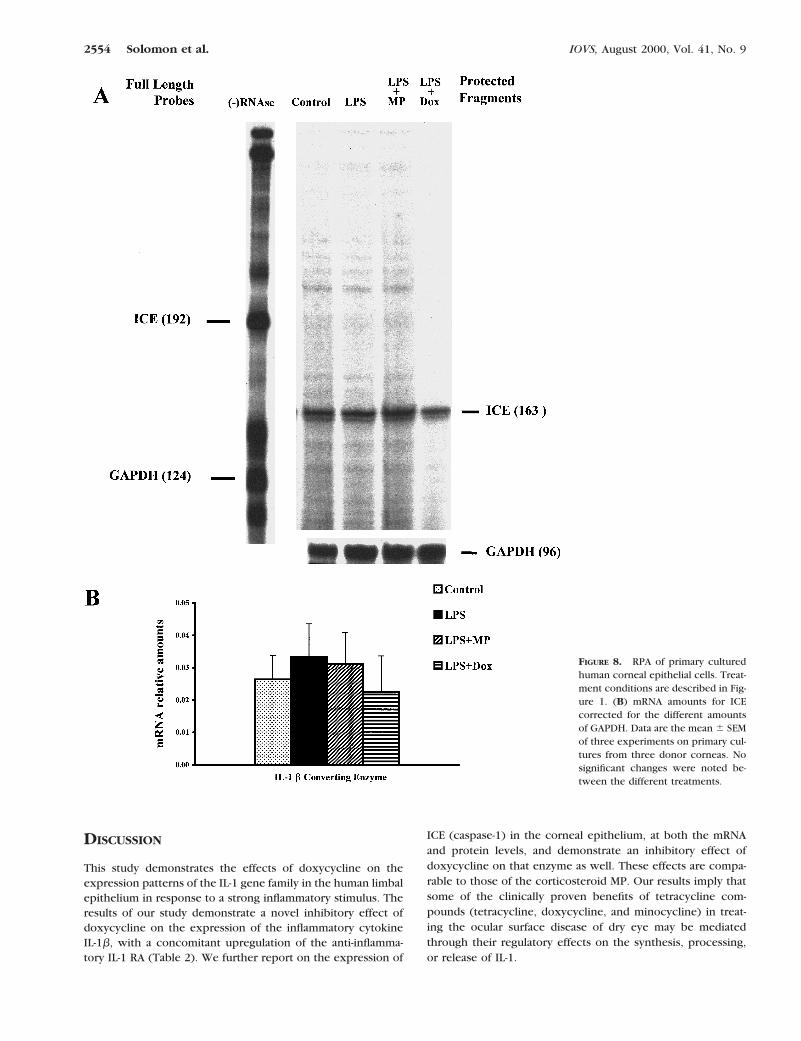
Effect of Doxycycline on ICE
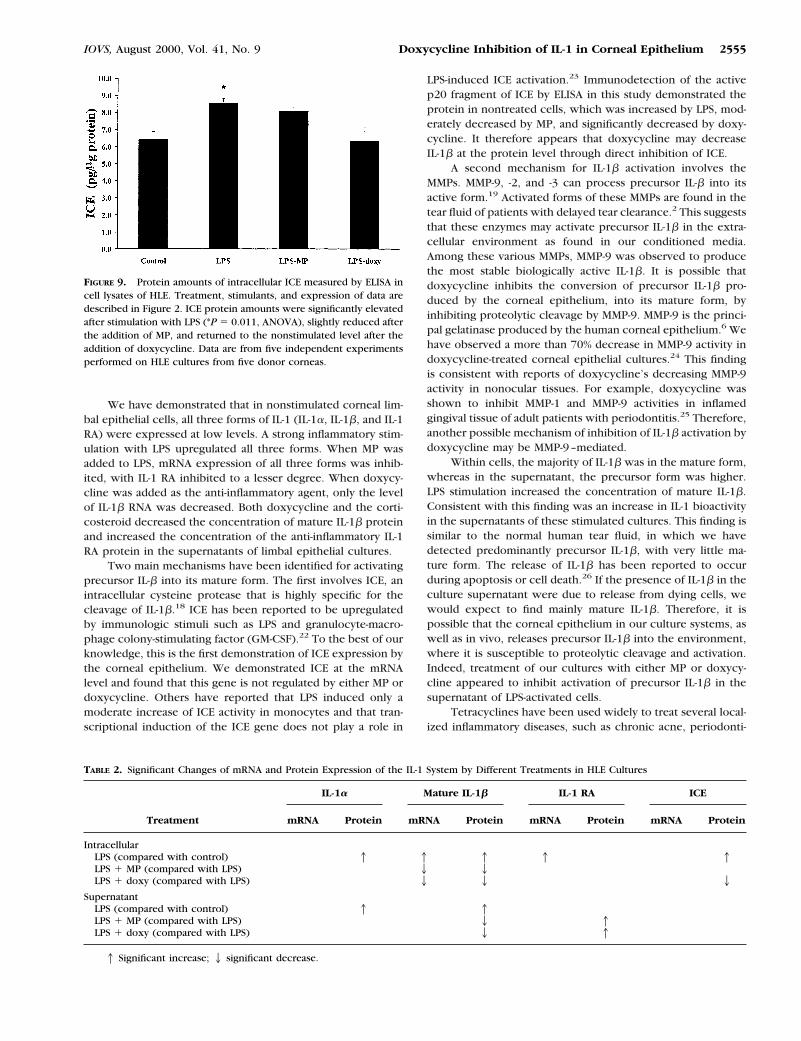
The mRNA transcripts of ICE were demonstrated in nontreatedlimbal epithelial cells (Fig. 8), but no changes were demon-strated among the different treatments. The active p20 subunitof ICE protein was demonstrated by ELISA in limbal epitheliumcell lysates (Fig. 9). Its concentration increased significantlyafter stimulation with LPS (from 6.4 6 1.1 to 8.5 6 0.6 pg/mgprotein; P 5 0.011, ANOVA). There was no significant changein the cellular concentration of ICE protein in the MP-treatedgroup. Doxycycline, however, significantly reduced the intra-cellular protein concentration when compared with LPS alone(6.3 6 1.8 pg/mg protein, P 5 0.011, ANOVA).
Although the active part of ICE (the p20 isoform) wasreadily measured by ELISA, we could not demonstrate ICEactivity in our samples using the ICE-specific fluorogenic sub-strate (YVAD-AMC). A possible explanation is that this assay,which was previously described for detecting ICE in mono-cytes and macrophages20 and THP-1 myelomonocytic cells, 21
did not have sufficient sensitivity to detect this enzyme’s activ-ity in HLE.
FIGURE 7. Western blot analysis ofIL-1 RA in conditioned media (top)and in cell lysates (bottom) of HLEcells cultured in medium alone (Con-trol), or treated with LPS, LPS 1 MP,LPS 1 doxycycline (LPS 1 doxy), orthe combination of LPS with MP anddoxycycline (LPS 1 MP 1 doxy).The positive control is rh-IL-1 RA.Samples were taken from the samecultures used for the ELISA analysesshown in Figures 5B and 6B. The dataare representative of four experi-ments.
IOVS, August 2000, Vol. 41, No. 9 Doxycycline Inhibition of IL-1 in Corneal Epithelium 2553
DISCUSSION
This study demonstrates the effects of doxycycline on theexpression patterns of the IL-1 gene family in the human limbalepithelium in response to a strong inflammatory stimulus. Theresults of our study demonstrate a novel inhibitory effect ofdoxycycline on the expression of the inflammatory cytokineIL-1b, with a concomitant upregulation of the anti-inflamma-tory IL-1 RA (Table 2). We further report on the expression of
ICE (caspase-1) in the corneal epithelium, at both the mRNAand protein levels, and demonstrate an inhibitory effect ofdoxycycline on that enzyme as well. These effects are compa-rable to those of the corticosteroid MP. Our results imply thatsome of the clinically proven benefits of tetracycline com-pounds (tetracycline, doxycycline, and minocycline) in treat-ing the ocular surface disease of dry eye may be mediatedthrough their regulatory effects on the synthesis, processing,or release of IL-1.
FIGURE 8. RPA of primary culturedhuman corneal epithelial cells. Treat-ment conditions are described in Fig-ure 1. (B) mRNA amounts for ICEcorrected for the different amountsof GAPDH. Data are the mean 6 SEMof three experiments on primary cul-tures from three donor corneas. Nosignificant changes were noted be-tween the different treatments.
2554 Solomon et al. IOVS, August 2000, Vol. 41, No. 9
We have demonstrated that in nonstimulated corneal lim-bal epithelial cells, all three forms of IL-1 (IL-1a, IL-1b, and IL-1RA) were expressed at low levels. A strong inflammatory stim-ulation with LPS upregulated all three forms. When MP wasadded to LPS, mRNA expression of all three forms was inhib-ited, with IL-1 RA inhibited to a lesser degree. When doxycy-cline was added as the anti-inflammatory agent, only the levelof IL-1b RNA was decreased. Both doxycycline and the corti-costeroid decreased the concentration of mature IL-1b proteinand increased the concentration of the anti-inflammatory IL-1RA protein in the supernatants of limbal epithelial cultures.
Two main mechanisms have been identified for activatingprecursor IL-b into its mature form. The first involves ICE, anintracellular cysteine protease that is highly specific for thecleavage of IL-1b.18 ICE has been reported to be upregulatedby immunologic stimuli such as LPS and granulocyte-macro-phage colony-stimulating factor (GM-CSF).22 To the best of ourknowledge, this is the first demonstration of ICE expression bythe corneal epithelium. We demonstrated ICE at the mRNAlevel and found that this gene is not regulated by either MP ordoxycycline. Others have reported that LPS induced only amoderate increase of ICE activity in monocytes and that tran-scriptional induction of the ICE gene does not play a role in
LPS-induced ICE activation.23 Immunodetection of the activep20 fragment of ICE by ELISA in this study demonstrated theprotein in nontreated cells, which was increased by LPS, mod-erately decreased by MP, and significantly decreased by doxy-cycline. It therefore appears that doxycycline may decreaseIL-1b at the protein level through direct inhibition of ICE.
A second mechanism for IL-1b activation involves theMMPs. MMP-9, -2, and -3 can process precursor IL-b into itsactive form.19 Activated forms of these MMPs are found in thetear fluid of patients with delayed tear clearance.2 This suggeststhat these enzymes may activate precursor IL-1b in the extra-cellular environment as found in our conditioned media.Among these various MMPs, MMP-9 was observed to producethe most stable biologically active IL-1b. It is possible thatdoxycycline inhibits the conversion of precursor IL-1b pro-duced by the corneal epithelium, into its mature form, byinhibiting proteolytic cleavage by MMP-9. MMP-9 is the princi-pal gelatinase produced by the human corneal epithelium.6 Wehave observed a more than 70% decrease in MMP-9 activity indoxycycline-treated corneal epithelial cultures.24 This findingis consistent with reports of doxycycline’s decreasing MMP-9activity in nonocular tissues. For example, doxycycline wasshown to inhibit MMP-1 and MMP-9 activities in inflamedgingival tissue of adult patients with periodontitis.25 Therefore,another possible mechanism of inhibition of IL-1b activation bydoxycycline may be MMP-9–mediated.
Within cells, the majority of IL-1b was in the mature form,whereas in the supernatant, the precursor form was higher.LPS stimulation increased the concentration of mature IL-1b.Consistent with this finding was an increase in IL-1 bioactivityin the supernatants of these stimulated cultures. This finding issimilar to the normal human tear fluid, in which we havedetected predominantly precursor IL-1b, with very little ma-ture form. The release of IL-1b has been reported to occurduring apoptosis or cell death.26 If the presence of IL-1b in theculture supernatant were due to release from dying cells, wewould expect to find mainly mature IL-1b. Therefore, it ispossible that the corneal epithelium in our culture systems, aswell as in vivo, releases precursor IL-1b into the environment,where it is susceptible to proteolytic cleavage and activation.Indeed, treatment of our cultures with either MP or doxycy-cline appeared to inhibit activation of precursor IL-1b in thesupernatant of LPS-activated cells.
Tetracyclines have been used widely to treat several local-ized inflammatory diseases, such as chronic acne, periodonti-
FIGURE 9. Protein amounts of intracellular ICE measured by ELISA incell lysates of HLE. Treatment, stimulants, and expression of data aredescribed in Figure 2. ICE protein amounts were significantly elevatedafter stimulation with LPS (*P 5 0.011, ANOVA), slightly reduced afterthe addition of MP, and returned to the nonstimulated level after theaddition of doxycycline. Data are from five independent experimentsperformed on HLE cultures from five donor corneas.
TABLE 2. Significant Changes of mRNA and Protein Expression of the IL-1 System by Different Treatments in HLE Cultures
Treatment
IL-1a Mature IL-1b IL-1 RA ICE
mRNA Protein mRNA Protein mRNA Protein mRNA Protein
IntracellularLPS (compared with control) 1 1 1 1 1LPS 1 MP (compared with LPS) 2 2LPS 1 doxy (compared with LPS) 2 2 2
SupernatantLPS (compared with control) 1 1LPS 1 MP (compared with LPS) 2 1LPS 1 doxy (compared with LPS) 2 1
1 Significant increase; 2 significant decrease.
IOVS, August 2000, Vol. 41, No. 9 Doxycycline Inhibition of IL-1 in Corneal Epithelium 2555

tis, and the dermatologic and ocular manifestations of rosa-cea.11,27–29 One study27 reported dramatic improvement in thesymptoms and signs of ocular rosacea in 111 patients treatedwith oral tetracycline or doxycycline. The exact mechanismsof doxycycline-induced suppression of the corneal epitheliumIL-1b system are not clear. Tetracycline was found to inhibitthe LPS-induced IL-1b secretion, but not the mRNA accumula-tion in human monocytes, suggesting a posttranscriptionalblock in cytokine production.30 In addition, tetracyclineblocked LPS-stimulated tumor necrosis factor (TNF)-a secre-tion, by inducing retention of membrane-associated TNF-a inmonocyte cell membranes, thus preventing this cytokine’srelease into the culture media.31 Because no transport mecha-nism has been identified for the IL-1b protein, this mechanismof inhibition is unlikely.
The functional activity of IL-1b generated from our limbalepithelial cultures was tested on corneal fibroblasts, measuringtheir MMP-1 and -3 secretion. This bioassay provides an in vitrosystem that closely resembles the in vivo situation, whereinepithelial cells of the ocular surface produce inflammatorycytokines that regulate MMP secretion by corneal and conjunc-tival fibroblasts.6 Using MMP-1 and -3 secretion as a measure ofIL-1b activity in conditioned media from HLE cultures, wefound significantly lower concentrations of these matrix-de-grading enzymes when conditioned media from LPS 1 doxy-cycline-treated cultures were added. This result may be eitherIL-1b–mediated or a result of a direct effect of doxycycline onthese enzymes.
Intervention in one of the IL-1b regulatory mechanismsmay provide important strategies for the treatment of variousinflammatory processes on the ocular surface. The increasedtear fluid concentrations of IL-1b in dry eye conditions mayresult from release of proinflammatory cytokines, especiallyIL-1b, from stressed or damaged ocular surface epithelium intothe tear film. A low volume of tear fluid and its delayedclearance from the ocular surface may further increase theconcentration of IL-1b. This in turn may induce recruitmentand activation of ocular surface inflammatory cells. Increasedactivity of proteolytic enzymes (such as MMP-9) in the tearfluid of patients with delayed tear clearance may also processprecursor-IL-1b to its mature form, further increasing the con-centration of IL-1b in the tear fluid. This inflammatory cascadecould be responsible for the signs and symptoms of ocularirritation so commonly encountered in dry eye. Pharmacologicintervention may be capable of breaking this vicious inflamma-tory cycle. We have demonstrated that doxycycline has thepotential to reduce the protein levels of cellular and extracel-lular mature IL-1b at a level comparable with that of a potentcorticosteroid. We therefore conclude that based on our invitro model, the efficacy of doxycycline for treatment of kera-titis sicca may be due to inhibition of the IL-1b system.
References
1. Pflugfelder SC, Jones D, Ji Z, Afonso A, Monroy D. Altered cytokinebalance in the tear fluid and conjunctiva of patients with Sjogren’ssyndrome keratoconjunctivitis sicca. Curr Eye Res. 1999;19:201–211.
2. Afonso A, Sobrin L, Monroy DC, Selzer M, Lokeshwar B, PflugfelderSC. Tear fluid Gelatinase B activity correlates with IL-1a concen-tration and fluorescein clearance in ocular rosacea. Invest Oph-thalmol Vis Sci. 1999;40:2506–2512.
3. Dinarello CA. The interleukin-1 family: 10 years of discovery.FASEB J. 1994;8:1314–1325.
4. Weng J, Mohan RR, Li Q, Wilson SE. IL-1 upregulates keratinocytegrowth factor and hepatocyte growth factor mRNA and proteinproduction by cultured stromal fibroblast cells: Interleukin-1b ex-pression in the cornea. Cornea. 1997;16:465–471.
5. Wilson SE, Lloyd SA, He YG. Glucocorticoid receptor and interleu-kin-1 receptor messenger RNA expression in corneal cells. Cornea.1994;13:4–8.
6. Fini ME, Girard MT. Expression of collagenolytic/gelatinolytic met-alloproteinases by normal cornea. Invest Ophthalmol Vis Sci.1990;31:1779–1788.
7. West–Mays JA, Sadow PM, Strissel KJ, Cintron C, Fini ME. Inter-leukin-1 alpha mediates collagenase synthesis in passaged cornealfibroblasts and in fibroblasts isolated from penetrating keratectomywounds. Invest Ophthalmol Vis Sci. 1995;36(4):S866. Abstract nr3967.
8. Wilson SE, He YG, Weng J, Li Q, Vital M, Chwang EL. Epithelialinjury induces keratocyte apoptosis: hypothesized role for inter-leukin-1 system in modulation of corneal tissue organization andwound healing. Exp Eye Res. 1996;62:325–338.
9. Marsh P, Pflugfelder SC. Topical non-preserved methylpred-nisolone therapy of keratoconjunctivitis sicca. Ophthalmology.1999;106:811–816.
10. Prabhasawat P, Tseng SCG. Frequent association of delayed tearclearance in ocular irritation. Br J Ophthalmol. 1998;82:666–675.
11. Frucht–Pery J, Sagi E, Hemo I, Ever–Hadani P. Efficacy of doxycy-cline and tetracycline in ocular rosacea. Am J Ophthalmol. 1993;116:88–92.
12. Hope–Ross MW, Chell PB, Kervick GN, McDonnell PJ, Jones HS.Oral tetracycline in the treatment of recurrent corneal erosions.Eye. 1994;8:384–388.
13. Culbertson WW, Huang AJ, Mandelbaum SH, Pflugfelder SC,Boozalis GT, Miller D. Effective treatment of phlyctenular kerato-conjunctivitis with oral tetracycline (see comments). Ophthalmol-ogy. 1993;100:1358–1366.
14. MacNaul KL, Chartrain N, Lark M, Tocci MJ, Hutchinson NI. Dis-coordinate expression of stromelysin, collagenase, and tissue in-hibitor of metalloproteinases-1 in rheumatoid human synovialfibroblasts: synergistic effects of interleukin-1 and tumor necrosisfactor-alpha on stromelysin expression. J Biol Chem. 1990;265:17238–17245.
15. Rawdanowicz TJ, Hampton AL, Nagase H, Woolley DE, Salamon-sen LA. Matrix metalloproteinase production by cultured humanendometrial stromal cells: identification of interstitial collagenase,gelatinase- A, gelatinase-B, and stromelysin-1 and their differentialregulation by interleukin-1 alpha and tumor necrosis factor-alpha.J Clin Endocrinol Metab. 1994;79:530–536.
16. Jasser MZ, Mitchell PG, Cheung HS. Induction of stromelysin-1 andcollagenase synthesis in fibrochondrocytes by tumor necrosis fac-tor-alpha. Matrix Biol. 1994;14:241–249.
17. Li D-Q, Tseng SCG. Three patterns of cytokine expression poten-tially involved in epithelial-fibroblast interactions of human ocularsurface. J Cell Physiol. 1995;163:61–79.
18. Thornberry NA, Bull HG, Calaycay JR, Chapman K, Howard A,Kostura M. A novel heterodimeric cysteine protease is required forinterleukin-1 beta processing in monocytes. Nature. 1992;356:768.
19. Schonbeck U, Mach F, Libby P. Generation of biologically activeIL-1 beta by matrix metalloproteinases: a novel caspase-1 indepen-dent pathway of IL-1 beta processing. J Immunol. 1998;161:3340–3346.
20. Wewers MD, Dare HA, Winnard AV, Parker JM, Miller DK. IL-1b-converting enzyme (ICE) is present and functional in humanalveolar macrophages: macrophage IL-1b release limitation is ICEindependent. J Immunol. 1997;159:5964–5972.
21. Ayala JM, Yamin T-T, Egger LA, Chin J, Kostura M, Miller DK. IL-1b-converting enzyme is present in monocytic cells as an inactive45-kDa precursor. J Immunol. 1994;153:2592–2599.
22. Zepter K, Haffner A, Soohoo LF, et al. Induction of biologically
2556 Solomon et al. IOVS, August 2000, Vol. 41, No. 9

active IL-1 beta converting enzyme and mature IL-1 beta in humankeratinocytes by inflammatory and immunologic stimuli. J Immu-nol. 1997;159:6203–6208.
23. Schumann RR, Belka C, Reuter D, et al. Lipopolysaccharide acti-vates caspase-1 (interleukin-1-converting enzyme) in culturedmonocytic and endothelial cells. Blood. 1998;91:577–584.
24. Sobrin L, Liu Z, Monroy DC, Solomon A, Lokeshwar B, PflugfelderSC. Activators and inhibitors of matrix metalloproteinase-9 in hu-man tear fluid and corneal epithelial culture supernatant. InvestOphthalmol Vis Sci. 2000; 41:1703–1709.
25. Golub LM, Sorsa T, Lee HM, et al. Doxycyline inhibits neutrophil(PMN)-type matrix metalloproteinases in human adult periodonti-tis. J Clin Periodontol. 1995;22:100–109.
26. Hogquist KA, Nett MA, Unanue ER, Chaplin DD. Interleukin 1 isprocessed and released during apoptosis. Immunology. 1991;88:8485–8489.
27. Akpek EK, Merchant A, Pinar V, Foster CS. Ocular rosacea: patientcharacteristics and follow-up. Ophthalmology. 1997;104:1863–1867.
28. Bartholomew RS, Reid BJ, Cheesbrough MJ, Macdonald M, Gallo-way NR. Oxytetracycline in the treatment of ocular rosacea: adouble-blind trial. Br J Ophthalmol. 1982;66:386–388.
29. Zengin N, Tol H, Gunduz K, Okudan S, Balevi S, Endogru H.Meibomian gland dysfunction and tear film abnormalities in rosa-cea. Cornea. 1995;14:144–146.
30. Shapira L, Soskolne WA, Houri Y, Barak V, Halabi A, Stabholz A.Protection against endotoxic shock and lipopolysaccharide-in-duced local inflammation by tetracycline: correlation with inhibi-tion of cytokine secretion. Infect Immun. 1996;64:825–828.
31. Shapira L, Houri Y, Barak V, Soskolne WA, Halabi A, Stabholz A.Tetracycline inhibits porphyromonas gingivalis lipopolysaccharidelesions in vivo and TNF alpha processing in vitro. J PeriodontalRes. 1997;32:183–188.
IOVS, August 2000, Vol. 41, No. 9 Doxycycline Inhibition of IL-1 in Corneal Epithelium 2557














![2560 - 2561 282 35 f] 2 22-27 2540-2544 29-32 2546-2549 35 ... · 2560 - 2561 282 35 f] 2 22-27 2540-2544 29-32 2546-2549 35-36 2552-2553 1, 2. 3. 4. 5, 6, 8, 9, 10, 2560](https://img.pdfslide.us/doc/110x75/60f8dd67ae7a7704e365c0ca/2560-2561-282-35-f-2-22-27-2540-2544-29-32-2546-2549-35-2560-2561-282-35.jpg)













