Embed Size (px)
Citation preview

14. CALCAREOUS NANNOPLANKTON BIOGEOGRAPHY ANDITS PALEOCLIMATE IMPLICATIONS: CENOZOIC OF THE FALKLAND PLATEAU
(DSDP LEG 36) AND MIOCENE OF THE ATLANTIC OCEAN
Bilal U. Haq and G.P. Lohmann, Woods Hole Oceanographic Institution, Woods Hole, Massachusettsand
Sherwood W. Wise, Jr., Florida State University, Tallahassee, Florida
INTRODUCTION
Latitudinal differentiation in calcareous nan-noplankton assemblages is known to have existedthroughout the Cenozoic and probably through mostof the Mesozoic. In the early Cenozoic such lati-tudinally differentiated assemblages have shown dis-tinct migrations across latitudes with time. In a study ofthe nannoplankton biogeography of the Atlantic basin,Haq and Lohmann (1976) have documented five suchmajor migrationary events in the early Cenozoic: (1) in-vasion of low latitudes by Paleocene high latitudeassemblages between 59 and 57 m.y.B.P., (2) invasionof high latitudes by late Paleocene-early Eocene low tomid latitude assemblages between 54 and 52 m.y.B.P.,(3) invasion of middle latitudes by Eocene high latitudeassemblages between 48 and 43 m.y.B.P., (4) invasionof low latitudes by late Eocene-early Oligocene highlatitude assemblages between 37 and 36 m.y.B.P., and(5) invasion of middle latitudes by Oligocene highlatitude assemblages between 32 and 27 m.y.B.P. Theseevents were interpreted as responses of the nannoflorato major fluctuations in the climate of the earlyCenozoic (see Haq and Lohmann, 1976, fig. 18).
The above study was based on DSDP material fromthe North and Central Atlantic only. Due to lack ofdrill sites in the southern high latitudes at the time ofthe study, no biogeographic data from South Atlantichigher than 30°S (Leg 3 sites) could be considered.During DSDP Leg 36, two open-ocean sites weredrilled on the eastern end of the Falkland Plateau(Figure 1), both of which are above 50°S latitude (Site327: 50°42.38'S and Site 329: 50°39.31'S) in waterdepths shallow enough for the preservation of earlyCenozoic and Miocene calcareous plankton. Due to itsposition, this area is critical to the delineation ofbiogeographic patterns and the paleooceanography ofthe Southern Ocean. The objective of this paper is toextend Atlantic nannoplankton biogeography into thesouthern high latitudes and to compare the results withthe distributional patterns in the northern highlatitudes and the low to middle latitudes of the Atlantic.
TIME INTERVALS CONSIDEREDAll the available Cenozoic material from DSDP Hole
327A and Site 329 was examined for the preservation ofcalcareous nannoplankton. All samples from Hole327A, Core 9 through Core 2, showed poor preserva-tion and excessive fragmentation of coccoliths, and a
meaningful census of Coccolith taxa could not be ob-tained. Samples from Site 329 showed varying degreesof Coccolith dissolution, with moderately well pre-served coccoliths at some levels in Cores 33 through 3.Upper Paleocene, lower Oligocene, and Miocenehorizons in particular contained coccoliths well enoughpreserved that the relative abundances of variousgroups could be determined. The horizon containingearly Eocene {Tribrachiatus orthostylus Zone—see Wiseand Wind, this volume) showed poor preservation andfragmentation of more susceptible taxa and is not con-sidered here.
On the basis of preservation, 12 time intervals (two inthe late Paleocene, two in the early Oligocene, and eightin the Miocene) were chosen for Coccolith census andintrabasinal comparison:
80° 60° 40°
Figure 1. Location of DSDP Leg 36 Sites.
745

B. U. HAQ, G. P. LOHMANN, S. W. WISE, JR.
Time Interval 1: Lower Part of Discoaster multiradiatus(NP9) Zone (56 to 55 m.y.B.P.)
This horizon is characterized by late Paleocene Coc-colith taxa, but only a single discoaster species, D. mul-tiradiatus. Sample 329-33-3, 118-120 cm is considered arepresentative sample for this interval.
Time Interval 2: Upper Part of Discoaster multiradiatus(NP9) Zone (55.5 to 53 m.y.B.P.)
At Site 329, Core 32, species of discoasters, such asD. falcatus, D. lenticularis, and (in the upper section ofthe core) D. barbadiensis, were recorded in addition tothe flora recorded in Core 33. These additional dis-coaster taxa are more typical of the upper part of theNP9 Zone. Sample 329-32-4, 5-7 cm is chosen to repre-sent this time interval.
Time Interval 3: Ericsonia subdisticha/Helicopontosphaera reticulata (NP 21/22) Zones (37.5to 35 m.y.B.P.)
This horizon contains a characteristic late Eocene-early Oligocene nannoflora; however, the zonediagnostic taxa, such as helicopontosphaerids andsphenoliths are absent. Due to the presence of Ericsoniasubdisticha and rare occurrence of Reticulofenestra um-bilica and Cyclococcolithus formosus, this time slice canbe assigned to a mixed NP 21/22 Zone. Sample 329-30-3, 78-80 cm is considered a representative sample forthis interval.
Time Interval 4: Upper Part of Helicopontosphaerareticulata to Sphenolithus predistentus (NP 22/23) Zones(36 to 32 m.y.B.P.)
This level covers a rather large period of 4 m.y. NoCyclococcolithus formosus was seen, but very rarespecimens of Reticulofenestra umbilica are still present.On the basis of foraminifera (see Tjalsma, this volume),this level has been assigned to the later part of the earlyOligocene. Due to these considerations we assign thishorizon to the NP 22/23 nanno zones and an age of 36to 32 m.y.B.P. Sample 329-28-1, 15-17 cm is consideredrepresentative for this interval.
Time Interval 5: Early Miocene (22.5 to 21 m.y.B.P)In Core 27 rare to few specimens of Reticulofenestra
bisecta and Dictyococcites abisectus were observed.These forms could not be reworked since we found noother reworked taxa. Other Coccolith taxa observed,though not zone diagnostic, are common in the earliestMiocene. On the basis of these observations, we placethis time interval between 22.5 to 21 m.y.B.P. Sample329-27-1, 120-124 cm is considered representative.
Time Intervals 6-12: Middle to Late Miocene (16-14,12-10, 11-9, 10.5-8.5, 10-8, 9-7, and 8-6 m.y.B.P.)
Eight time intervals are chosen from Cores 23 to 3representing sediments from middle to late Miocene onthe basis of the availability of well-preserved nan-noflora. In the absence of age-diagnostic species, agesassigned to these time intervals are based on sedimenta-tion rates of these cores. For representative samples seeTable 1.
CENSUS DATA AND PALEOBIOGEOGRAPHYIn smear slides made from raw samples representing
each time interval 100 to 300 specimens (depending onthe floral diversity) were counted. Table 2 lists the cen-sus data expressed as percent.
Late Paleocene AssemblagesThe late Paleocene assemblages of the Falkland
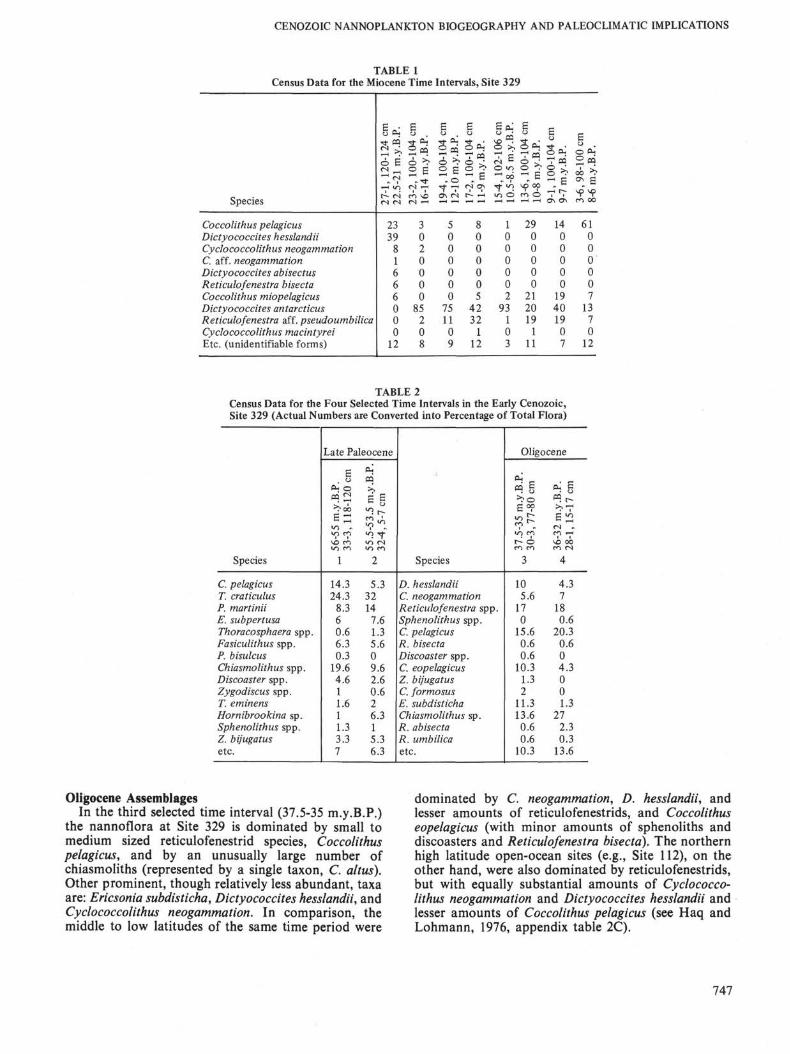
Plateau are characterized by the same taxa thatdominate the mid to low latitudes of this time.However, there are distinct differences in the relativeabundance of these taxa, and additional taxa also occurat this site (Figure 2).
During the first time slice (56-55 m.y.B.P.), a Toweiuscraticulus-Coccolithus pelagicus assemblage dominates,much like the mid to low latitudes of the same time (seeHaq and Lohmann, 1976, fig. 6B). This is a relativelylow-latitude assemblage (Haq and Lohmann, 1976,table 9) which migrates into the higher latitudes duringthe late Paleocene-early Eocene interval. Like the midto low latitudes, minor amounts of Ericsonia subper-tusa, fasciculiths, and discoasters are also present.
This nannoflora, however, differs in some significantrespects from the northern high latitude floras of thesame time. In the northern high latitudes (e.g., DSDPHole 117A) Prinsius martinii is the dominantassemblage from 63 to 54 m.y.B.P., and the Toweiuscraticulus-Coccolithus pelagicus assemblage is lessprominent, becoming dominant only in the latestPaleocene-early Eocene. On the Falkland Plateau,however, the latter assemblage is more prominent thanthe former which comprises only about 8.3% of thetotal flora (as compared to 55.3% at northern Site 117).
Another striking difference between the northern andsouthern high latitudes is the prominence ofchiasmoliths on the Falkland Plateau (up to 19.6% ofthe total flora). Chiasmoliths are found in much smallernumbers (up to 4.3%) at Site 117. Another difference isthe presence of Hornibrookina austral is at Site 329,which is absent in the northern high latitudes of theAtlantic and, in fact, has been recorded only once fromthe northern hemisphere (Crimea, Perch-Nielsen, per-sonal communication). Genus Hornibrookina has beenreported from Danian to early Eocene of such southernhigh latitude areas as New Zealand, Tasman Sea(Edwards and Perch-Nielsen, 1975), BellingshausenAbyssal Plain (Haq, 1976), and may prove to be aprovincial taxon peculiar to the southern latitudes.
In the latest Paleocene (55.5-53.5 m.y.B.P.) the domi-nant assemblages remain the same as earlier, however,chiasmoliths are reduced in prominence (from 19.6% to9.6%). Hornibrookina australis, on the other hand, in-creases in abundance (from 1% in time interval 1, to6.3% in time interval 2).
These comparisons indicate a "mixed" nature of theassemblages of Falkland Plateau during the latePaleocene. The dominance by warm low latitudeassemblages, the prominence of chiasmoliths (cool-water), and the continued presence of Prinsius martinii(cool water) in time-interval 2 (as compared to northernhigh latitudes where this taxon is absent during thistime), all suggest dual influences of cool and warmwater nannofloras.
746
CENOZOIC NANNOPLANKTON BIOGEOGRAPHY AND PALEOCLIMATIC IMPLICATIONS
TABLE 1Census Data for the Miocene Time Intervals, Site 329
Species
Coccolithus pelagicusDictyococcites hesslandüCyclococcolithus neogammationC. aff. neogammationDictyococcites abisectusReticulofenestra bisectaCoccolithus miopelagicusDictyococcites antarcticusReticulofenestra aff. pseudoumbilicaCyclococcolithus macintyreiEtc. (unidentifiable forms)
εo P-j
^* pa
1 ! >>
6 ε<N ^
r-H IT)
t~- <N<N <N
2339
81666000
12
UIO
\
2© >?
2 ε< N ~ ~CO VO<N •
3020000
85208
o
2 «ó >>2 ε
„©
σs <N
5000000
7511
09
εo
1 M-l
© •%•
1 ε<N~σ•s
8000005
4232
112
go;
© >.
<N ε© in
"~1 °?
IO ©
1000002
93103
uio \
T1 pa2 ><© .
vo opCO ©
2900000
212019
111
o
O 0HT pa
§ >>
"* ε
1400000
194019
07
oo Pi
2oó >>as g
CO 00
61000007
1370
12
TABLE 2Census Data for the Four Selected Time Intervals in the Early Cenozoic,Site 329 (Actual Numbers are Converted into Percentage of Total Flora)
Species
C. pelagicusT. craticulusP. martinüE. subpertusaThoracosphaera spp.Fasiculithus spp.P. bisulcusChiasmolithus spp.Discoaster spp.Zygodiscus spp.r. eminensHornibrookina sp.Sphenolithus spp.Z. bijugatusetc.
Late Paleocene
oPi opa ^>>oò
wo>O CO
VD CO>O CO
1
14.324.3
8.360.66.30.3
19.64.611.611.33.37
B.P
.
>>
?^U-> <Nio ro
2
5.332147.61.35.609.62.60.626.315.36.3
Species
D. hesslandüC. neogammationReticulofenestra spp.Sphenolithus spp.C. pelagicusR. bisectaDiscoaster spp.C. eopelagicusZ. bijugatusC. formosusE. subdistichaChiasmolithus sp.R. abisectaR. umbilicaetc.
Oligocene
« 1> o
$ £<A cçTt> ©
3
105.6
170
15.60.60.6
10.31.32
11.313.60.60.6
10.3
pi ε" ^
p i o<N -CO y-i
vó oòCO <N
4
4.37
180.6
20.30.604.3001.3
272.30.3
13.6
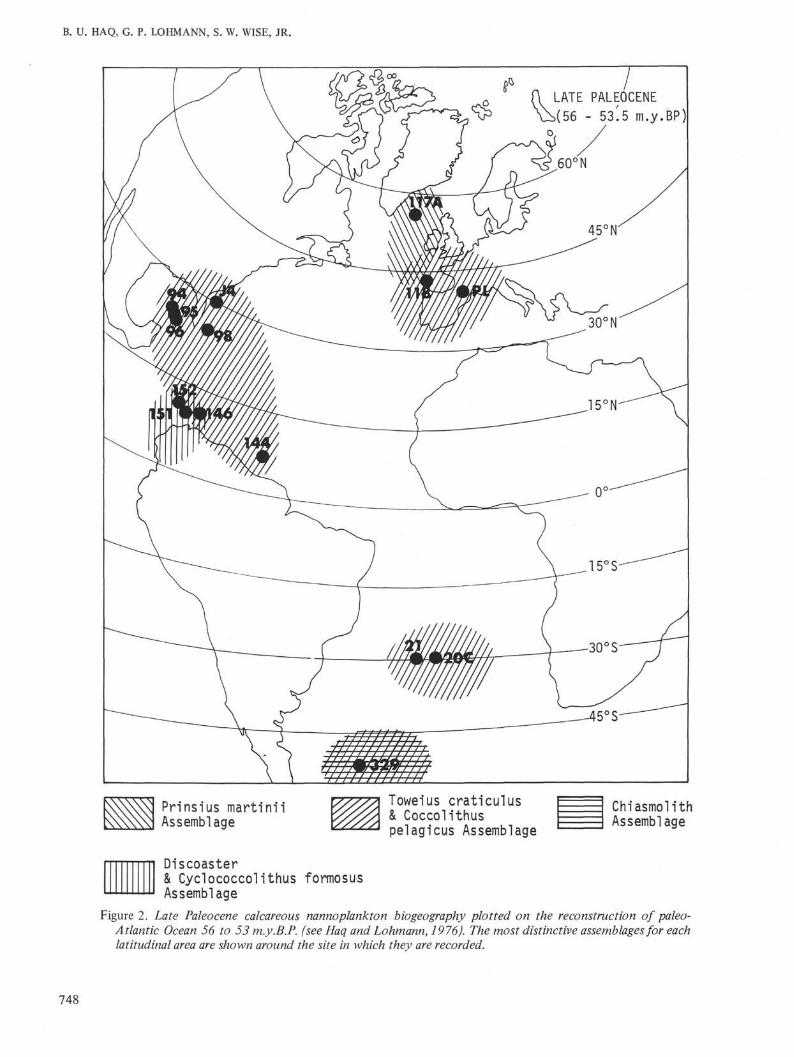
Oligocene AssemblagesIn the third selected time interval (37.5-35 m.y.B.P.)
the nannoflora at Site 329 is dominated by small tomedium sized reticulofenestrid species, Coccolithuspelagicus, and by an unusually large number ofchiasmoliths (represented by a single taxon, C. altus).Other prominent, though relatively less abundant, taxaare: Ericsonia subdisticha, Dictyococcites hesslandii, andCyclococcolithus neogammation. In comparison, themiddle to low latitudes of the same time period were
dominated by C. neogammation, D. hesslandii, andlesser amounts of reticulofenestrids, and Coccolithuseopelagicus (with minor amounts of sphenoliths anddiscoasters and Reticulofenestra bisecta). The northernhigh latitude open-ocean sites (e.g., Site 112), on theother hand, were also dominated by reticulofenestrids,but with equally substantial amounts of Cyclococco-lithus neogammation and Dictyococcites hesslandii andlesser amounts of Coccolithus pelagicus (see Haq andLohmann, 1976, appendix table 2C).
747
B. U. HAQ, G. P. LOHMANN, S. W. WISE, JR.
A LATE PALEOCENE\^>(56 - 53!5 m.y.BP)
Prinsius martiniiAssemblage
Toweius craticulus& Coccolithuspelagicus Assemblage
Discoaster& Cyclococcolithus formosusAssemblage
ChiasmolithAssemblage
Figure 2. Late Paleocene calcareous nannoplankton biogeography plotted on the reconstruction of paleo-Atlantic Ocean 56 to 53 m.y.B.P. (see Haq and Lohmann, 1976). The most distinctive assemblages for eachlatitudinal area are shown around the site in which they are recorded.
748

CENOZOIC NANNOPLANKTON BIOGEOGRAPHY AND PALEOCLIMATE IMPLICATIONS
The most striking difference between northern andsouthern high latitudes of this time, much like the latePaleocene, is the occurrence of a large number ofchiasmoliths which are either very rare or absent inmost other Oligocene sites of the Atlantic (see Figure3).
In time interval 4 (36-32 m.y.B.P.), the biogeographicrelationships remain essentially similar to time interval3, except that the chiasmoliths become more dominantthan reticulofenestrids and Coccolithus pelagicus (up to27% of the total flora). There is a noticeable decrease inthe abundance of Dictyococcites hesslandii and Coc-colithus eopelagicus, and Ericsonia subdisticha is reduceddrastically. In the northern high latitudes at this timeDictyococcites hesslandii, Cyclococcolithus neogamma-tion, and reticulofenestrids were all equally prominentand chiasmoliths were absent. Thus, there is a greaterbiogeographic distinction between the northern andsouthern high latitudes of this time as compared to thelast time interval.
Miocene Assemblages and Paleobiogeography of theAtlantic Ocean
In this section together with Leg 36 results, we in-clude the preliminary results of the study on Miocenebiogeography of the entire Atlantic Ocean. Theseresults have not been published earlier and are includedhere so that the Miocene assemblage of the FalklandPlateau can be effectively compared with the remainderof the Atlantic Ocean. Figures 4 to 11 show thetime/latitudinal distributions of various nannoplank-ton assemblages during the Miocene.
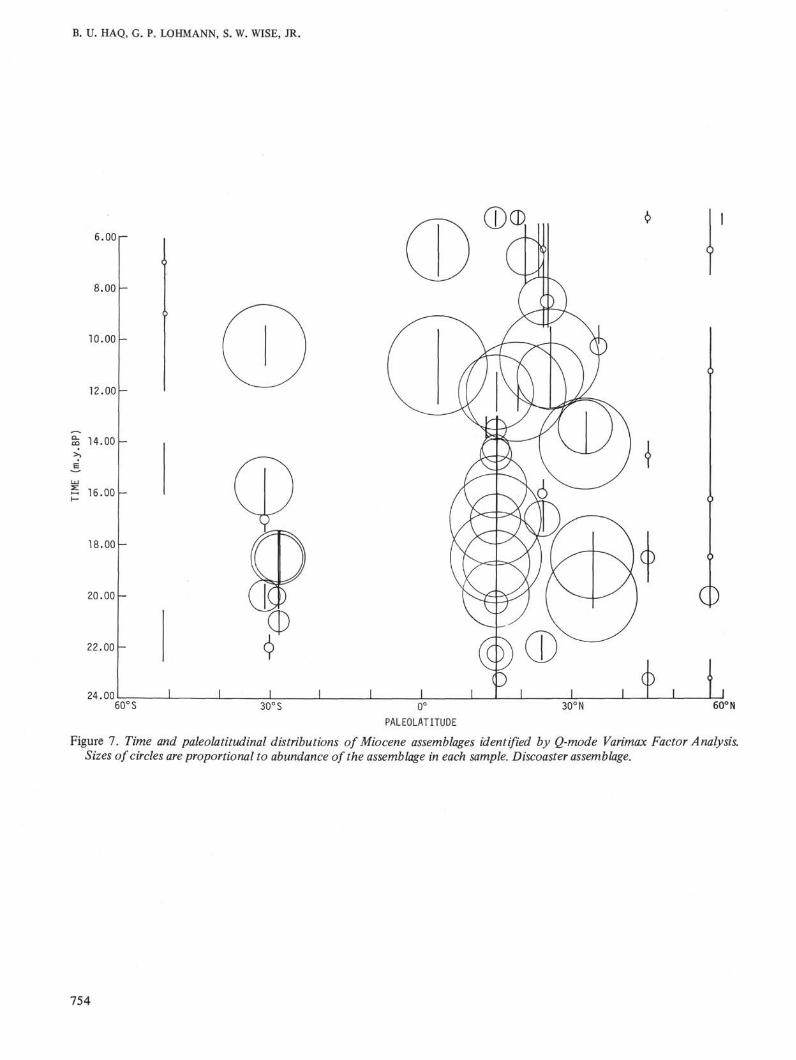
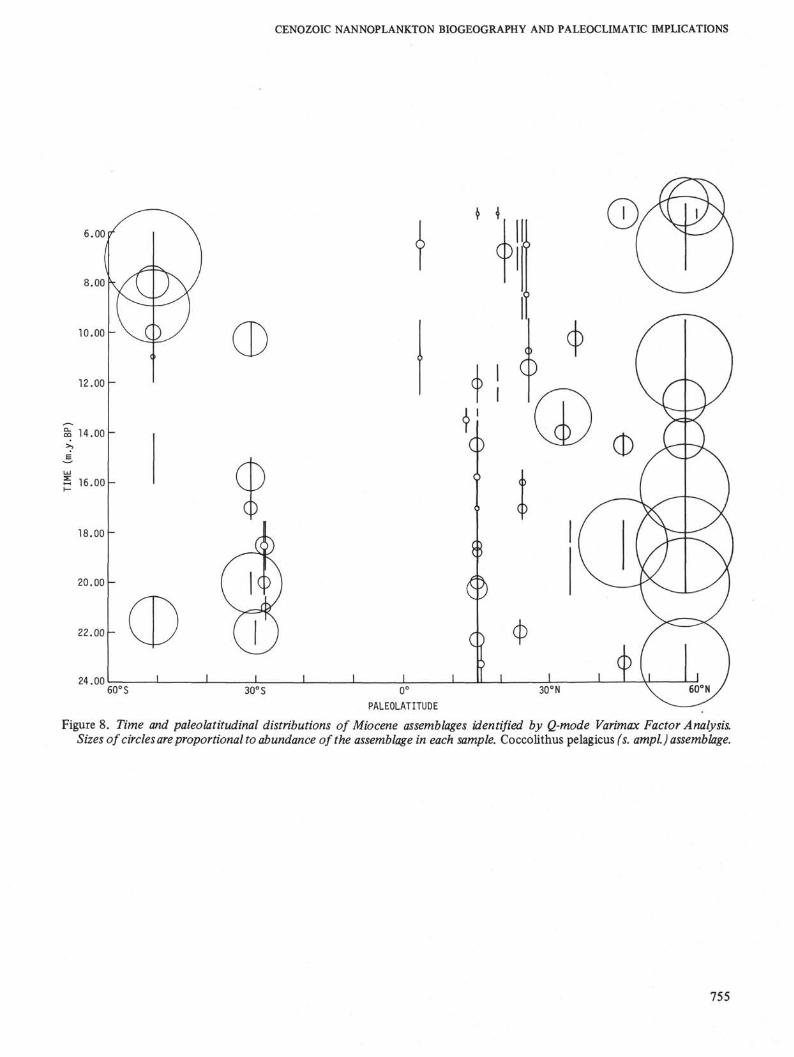
Like the early Tertiary, Miocene assemblages showdistinct latitudinal differentiations. In the earliestMiocene (NN1 Zone) the late Oligocene biogeographicpatterns continue. Cyclococcolithus neogammation andDictyococcites hesslandii continue to dominate the lowto mid latitudes, but in the northern high latitudes thesetaxa are less prominent, Coccolithus pelagicus being themost dominant taxon. This essentially Oligocenepattern changes in the NN2 Zone when a discoasterassemblage achieves dominance in the low to midlatitudes and Cyclococcolithus neogammation continuesto be abundant only in the low latitudes. During theremainder of the early Miocene (NN2 to NN4 zones),these patterns remain relatively stable, and there is littlevariation in the makeup of the latitudinal assemblageswith the exception of a general reduction in the abun-dance of Dictyococcites hesslandii. The northern highlatitudes contain a stable Coccolithus pelagicusassemblage.
During the early Miocene the Falkland Plateau areawas dominated by the cosmopolitan taxon, Dictyococ-cites hesslandii, and the cool Coccolithus pelagicus.Cyclococcolithus neogammation occurs in less abundantnumbers (see Table 1).
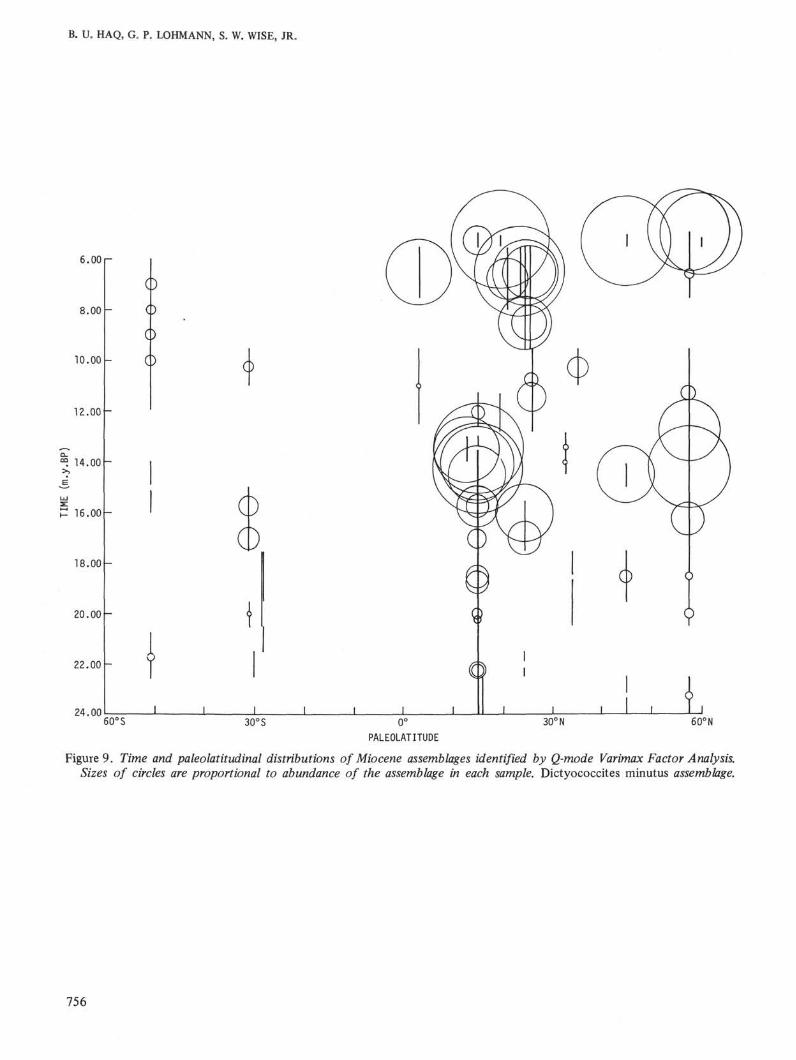
In the lower part of the middle Miocene (NN5 Zone)the patterns change again. A tropical assemblagedominated by two small Coccolith taxa, Dictyococcitesminutus (Haq) n. comb., and Cyclococcolithus aff.stradneri Jafar, appears in the low latitudes and spreadsquickly to the mid latitudes. These taxa, with lesser
amounts of Reticulofenestra pseudoumbilica and dis-coasters, remain dominant in the lower latitudes up tothe NN8 Zone of the upper mid Miocene. The earlierdiscoaster dominated low-latitude assemblage is dis-placed towards the mid latitudes at this time (DSDPSite 10). Minor amounts of Dictyococcites minutus alsostart appearing in the northern high latitudes in theNN5 Zone and become dominant in the NN6 Zone,remaining dominant until the NN8 Zone.
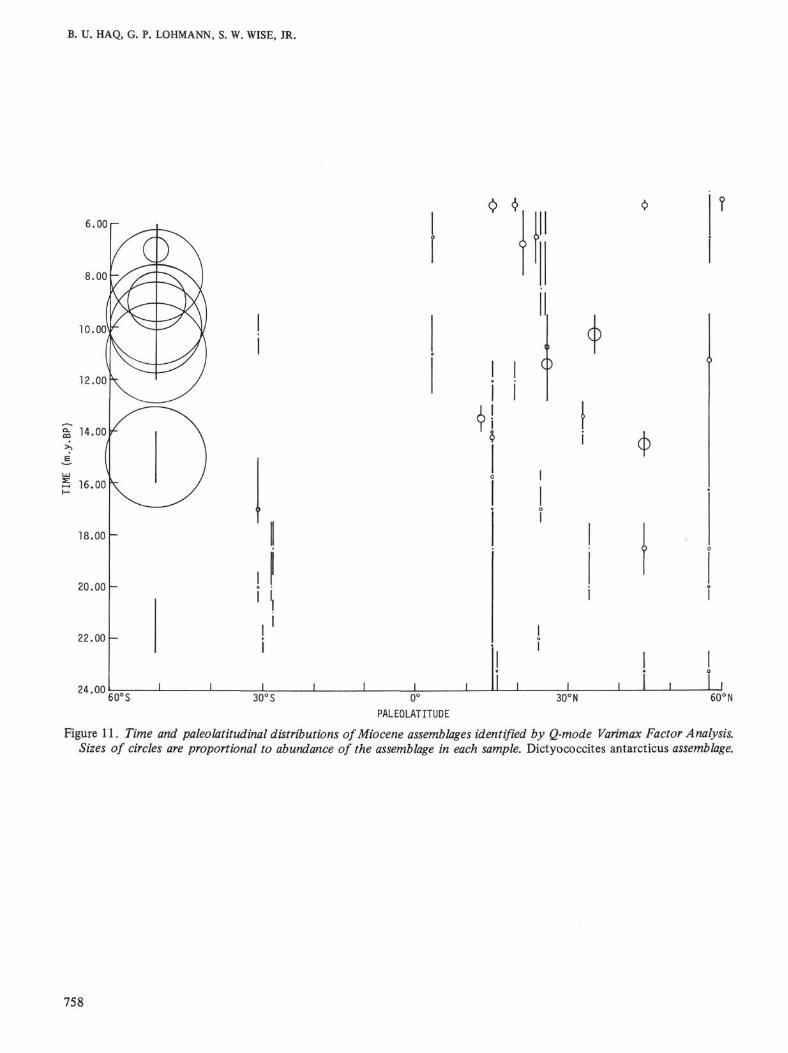
At the Falkland Plateau (Site 329) a new assemblagedominated by Dictyococcites antarcticus Haq, appearsin the middle Miocene. D. antarcticus seems restrictedto southern high latitudes and true D. antarcticus hasnot been recorded from other areas. This assemblage isessentially the same recorded in the Miocene of theBellingshausen Abyssal Plain during DSDP Leg 35 (seeHaq, 1976).
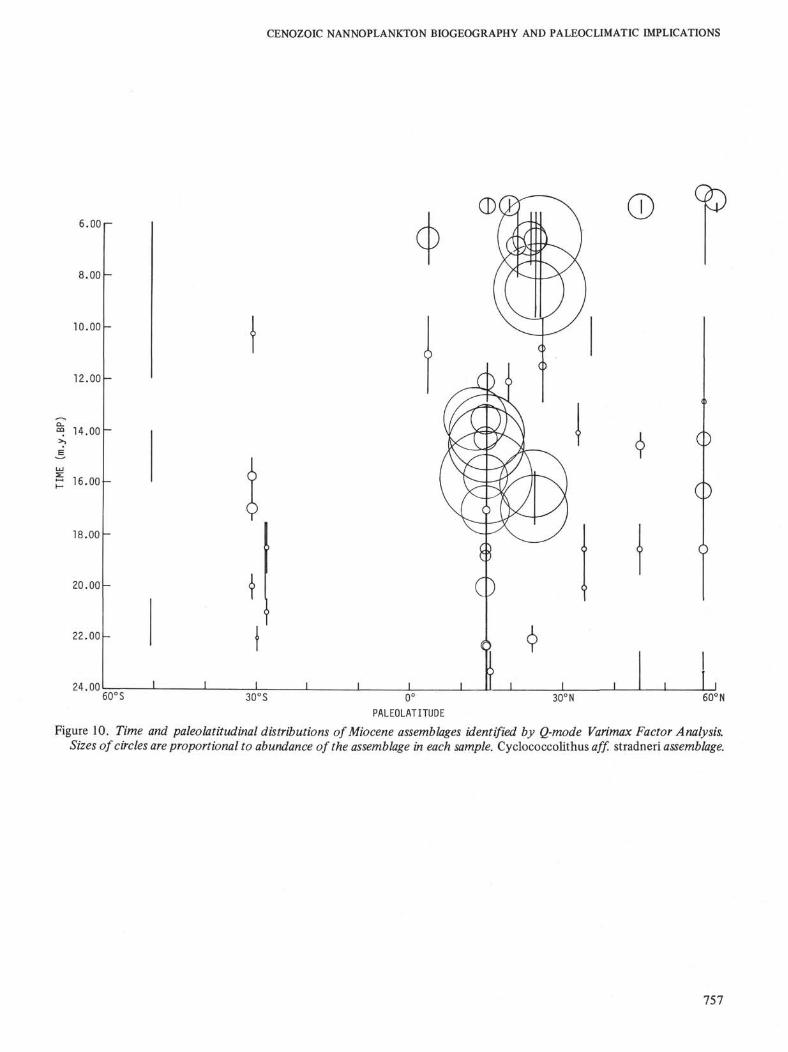
In the upper middle Miocene the discoaster-dom-inated assemblage again returns to the low latitudesand Dictyococcites minutus and Cyclococcolithus aff.stradneri are sharply reduced in both the low and midlatitudes. Northern high latitudes are also once againdominated by Coccolithus pelagicus in the NN9-NN10zones. This general pattern continues into the lowerpart of the late Miocene (lower NN11 Zone).
In the upper part of the late Miocene (upper NN11and NN12 zones) and earliest Pliocene (NN13 Zone) asecond expansion of Dictyococcites minutus andCyclococcolithus aff. stradneri takes place in the lowlatitudes. Once again these tropical elements spreadinto the mid and high latitudes—the northern highlatitudes are dominated by Dictyococcites minutus andCoccolithus pelagicus through the NN12/NN13 zones.
Throughout the middle and late Miocene the Falk-land Plateau assemblages are essentially dominated byDictyococcites antarcticus, with some variations in thelate Miocene when Coccolithus pelagicus becomes im-portant in one interval and Reticulofenestra pseudoum-bilica becomes prominent in another interval (see Table1).
Miocene Paleoclimatology of the Atlantic OceanThe Miocene migrations of nannoplankton assem-
blages through latitudes can be interpreted as re-sponses of the latitudinally restricted floras to majorclimatic changes. The Miocene climatic history can besummarized as follows: (1) The relatively warm lateOligocene climatic trend continues into the earliestMiocene (23 to 21.5 m.y.B.P.). This is indicated by thepresence of late Oligocene Dictyococcites hesslandii andCyclococcolithus neogammation assemblages in the lowto high latitudes. Elements of these assemblages arealso common on the Falkland Plateau during this time;(2) A cooling event between 19 and 17 m.y.B.P. is in-dicated by the decrease in the Dictyococcites hesslandiiand Cyclococcolithus neogammation assemblages in lowlatitudes and invasion of the high latitude Coccolithuspelagicus assemblage into mid latitudes; (3) A majorwarming trend between 15 and 13 m.y.B.P. is evidencedby the emergence and spread of the tropical Dictyococ-cites minutus and Cyclococcolithus aff. stradneri assem-blages in all latitudes. An austral assemblage
749
B. U. HAQ, G_ P. LOHMANN, S. W. WISE, JR.
EARLY OLIGOCENE(37.5 - 32 m.y.BP)
/60°N
45°N
Cyclococcolithusneogammation-Dictyococciteshesslandii Assemblage
RetiCulofenetrid-R. bisectaAssemblage
Reticulofenestrid•ChiasmolithAssemblage
Figure 3. Early Oligocene calcareous nannoplankton biogeography.
750

Samples Palolatitude Age (m.y.BP)
123456789
101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960616263646566676869
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
DSDP
18-3, 2-14422-1 , 6-8127-3, 2-12495-2, 6-99116-19, 1-91119-11, 1-24149-26, 1-64151-7, 1-5149-23, 3-10215-9, 6-120136-3, 4-10116-14, CC151-4, 2-12514-1 A, 3-65151-4, 1-120116-13, CC14-1A, 3-5218-2, 2-144119-7, 3-125136-3, 2-10149-21, 1-91329-27, 1-12094-9, 6-14415-7, 1-27149-20, 5-125151-4, 1-68116-10, 6-12294-9, 2-7415-6, 6-144149-17, 5-141141-5, 1-5329-23, 2-100119-5, 3-122149-15, 4-10116-9, 1-2910-2, 3-145151-3, 6-145151-3, 2-115149-15, 3-12430-12, 1-7.5116-8, 3-13910-2, 3-10116-6, 1-29141-7, 2-82149-14, 1-110142-8, 1-2797-4, 2-897-3, 3-124329-19, 4-100135-3, 1-6015-5, 6-144329-17, 2-100329-15, 4-10294-8, 5-12498-4, 6-139329-13, 6-100329-9, 1-100329-3, 6-9889-6, 5-74142-7, 6-12094-7, 1-113116-5, 5-11998-3, 3-49139-2, 4-145151-2, 3-140141-6, 4-145118-5, 2-44116-3, 6-117114-6, 6-134
-28C
-30c
16°24°58°45°15°15°15°
-31°34°58°15°
-28°15°58°
-28°-28°
45°34°15°
-51°25°
-31°15°15°58°25°
-31°15°15°
-51°45°15°58°33°15°15°15°13°58°33°58°19°15°
3°26°26°
-51°35°
-31°-51°-51°
25°25°
-51°-51°-51°
21°3°
25°58°25°24°15°19°45°58°60°
SSNNNNNNNSNNNSNNSSNNNSNSNNNNSNNSNNNNNNNNNNNNNNNNSNS
ssNNSSsNNNNNNNNNNN
21.522.524.022.524.024.024.024.021.020.520.520.520.520.520.019.519.519.519.519.519.522.517.517.517.517.517.516.516.516.516.516.015.015.015.014.515.014.514.014.014.014.013.012.812.812.512.812.012.011.011.011.010.5
9.59.5
10.09.08.08.07.57.57.57.57.55.55.55.55.55.5
20.521.522.521.522.522.520.520.519.519.519.519.519.519.517.517.517.517.517.517.517.521.016.516.516.516.515.015.515.015.015.014.014.013.513.513.514.013.513.013.011.512.8
9.511.311.3
9.510.0
9.510.0
9.59.59.08.57.57.58.07.06.05.55.55.55.55.55.55.05.05.04.04.5
6.00
8.00
10.00
12.00
14.00
16.00
18.00
20.00
22.00
?d.nn
—
-
_
-
58
57
56
53 T
I51
1*9 1
32 i
29
2k
_
17
i :T
IB
1 ,
1
22 T2
1 1 1 1 1
]
8059
8
f;i.
5 4
k&
> r
7
I
" T
1
. 139
3 8
37
30
26
211 5
10
8r •1-
1 |
61 I"/
63
55
T
T50k8
1kl
•
1 r28
23
20
11
19
4
T I
II i l l
.1
6 2
41
3 5
2 7
1G
12
r
60°S 30°S 0°
PALEOLATITUDE
30°N 60° N
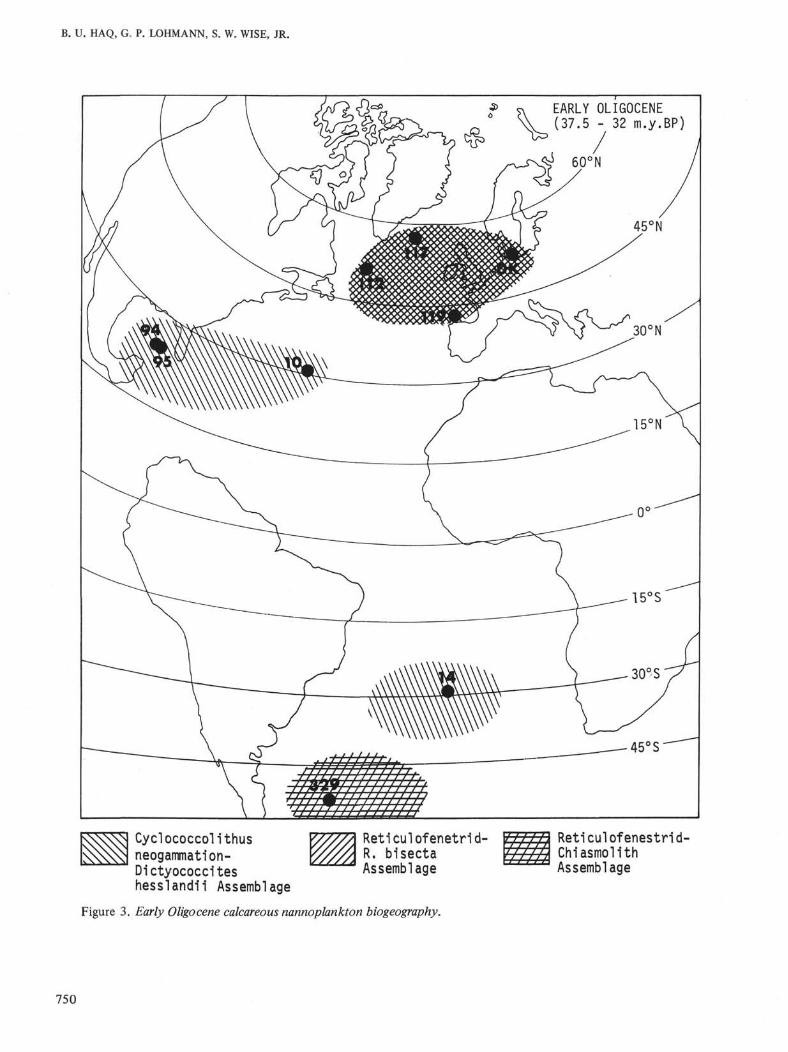
Figure 4. Time and paleolatitudinal distribution of Miocene samples. Numbers refer to sample list on the left. Length of line segment beside each num-ber reflects uncertainty of sampled estimated age. Some samples are used in illustrating the distribution patterns of assemblages in Figures 5-11.
oNon
>δzin
B. U. HAQ, G. P. LOHMANN, S W. WISE, JR,
β.OOr
8.00
12.00
14.00
16.00
r lδ.oo
19.00
20.00
22.00
24.0060° S 30°S 0°
PALEOLATITUDE
30° N 60°N
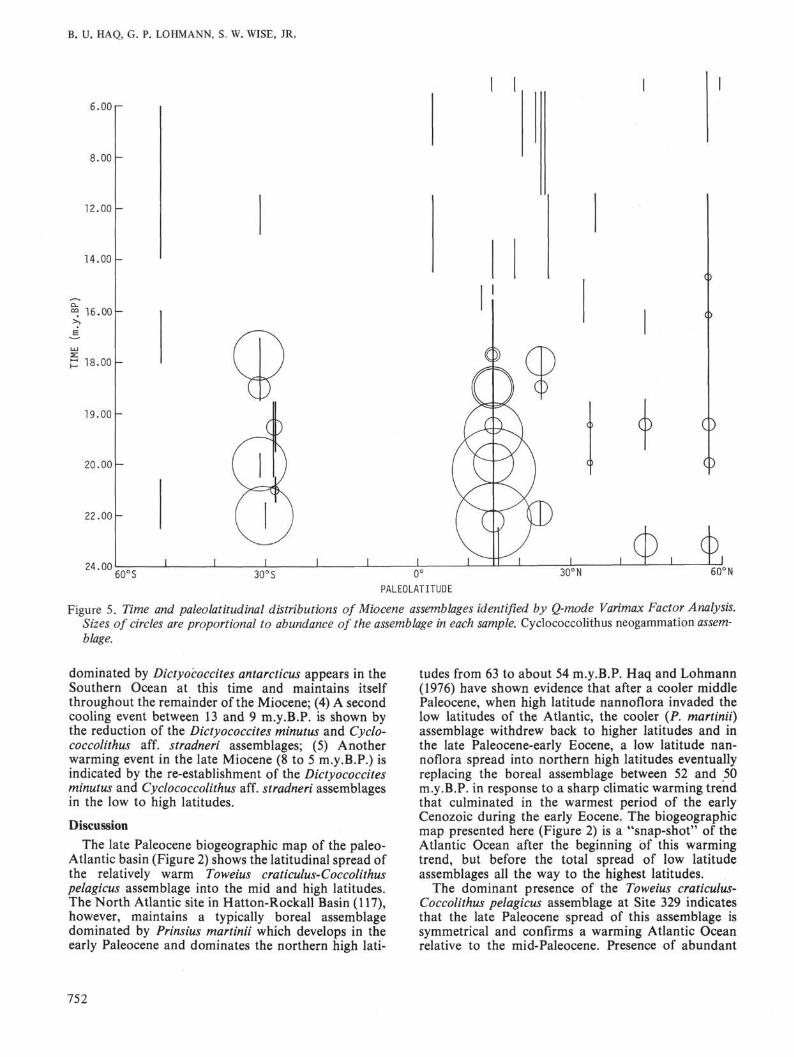
Figure 5. Time and paleolatitudinal distributions of Miocene assemblages identified by Q-mode Varimax Factor Analysis.Sizes of circles are proportional to abundance of the assemblage in each sample. Cyclococcolithus neogammation assem-blage.
dominated by Dictyococcites antarcticus appears in theSouthern Ocean at this time and maintains itselfthroughout the remainder of the Miocene; (4) A secondcooling event between 13 and 9 m.y.B.P. is shown bythe reduction of the Dictyococcites minutus and Cyclo-coccolithus aff. stradneri assemblages; (5) Anotherwarming event in the late Miocene (8 to 5 m.y.B.P.) isindicated by the re-establishment of the Dictyococcitesminutus and Cyclococcolithus aff. stradneri assemblagesin the low to high latitudes.
Discussion
The late Paleocene biogeographic map of the paleo-Atlantic basin (Figure 2) shows the latitudinal spread ofthe relatively warm Toweius craticulus-Coccolithuspelagicus assemblage into the mid and high latitudes.The North Atlantic site in Hatton-Rockall Basin (117),however, maintains a typically boreal assemblagedominated by Prinsius martinii which develops in theearly Paleocene and dominates the northern high lati-
tudes from 63 to about 54 m.y.B.P. Haq and Lohmann(1976) have shown evidence that after a cooler middlePaleocene, when high latitude nannoflora invaded thelow latitudes of the Atlantic, the cooler (P. martinii)assemblage withdrew back to higher latitudes and inthe late Paleocene-early Eocene, a low latitude nan-noflora spread into northern high latitudes eventuallyreplacing the boreal assemblage between 52 and 50m.y.B.P. in response to a sharp climatic warming trendthat culminated in the warmest period of the earlyCenozoic during the early Eocene. The biogeographicmap presented here (Figure 2) is a "snap-shot" of theAtlantic Ocean after the beginning of this warmingtrend, but before the total spread of low latitudeassemblages all the way to the highest latitudes.
The dominant presence of the Toweius craticulus-Coccolithus pelagicus assemblage at Site 329 indicatesthat the late Paleocene spread of this assemblage issymmetrical and confirms a warming Atlantic Oceanrelative to the mid-Paleocene. Presence of abundant
752
CENOZOIC NANNOPLANKTON BIOGEOGRAPHY AND PALEOCLIMATIC IMPLICATIONS
6.00
8.00
10.00
12.00
14.00
16.00
18.00
20.00
22.00
24.00
C)
60°S 30° S 0°PALEOLATITUDE
60°N
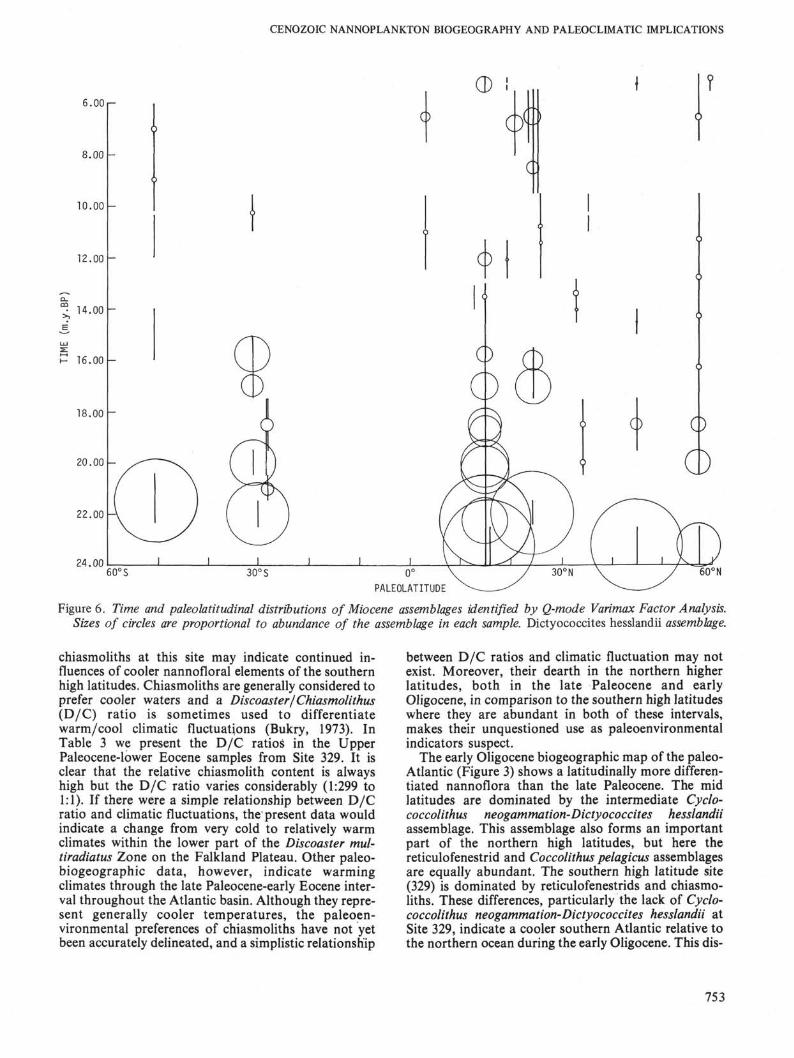
Figure 6. Time and paleolatitudinal distributions of Miocene assemblages identified by Q-mode Varimax Factor Analysis.Sizes of circles are proportional to abundance of the assemblage in each sample. Dictyococcites hesslandii assemblage.
chiasmoliths at this site may indicate continued in-fluences of cooler nannofloral elements of the southernhigh latitudes. Chiasmoliths are generally considered toprefer cooler waters and a Discoaster/Chiasmolithus(D/C) ratio is sometimes used to differentiatewarm/cool climatic fluctuations (Bukry, 1973). InTable 3 we present the D/C ratios in the UpperPaleocene-lower Eocene samples from Site 329. It isclear that the relative chiasmolith content is alwayshigh but the D/C ratio varies considerably (1:299 to1:1). If there were a simple relationship between D/Cratio and climatic fluctuations, the present data wouldindicate a change from very cold to relatively warmclimates within the lower part of the Discoaster mul-tiradiatus Zone on the Falkland Plateau. Other paleo-biogeographic data, however, indicate warmingclimates through the late Paleocene-early Eocene inter-val throughout the Atlantic basin. Although they repre-sent generally cooler temperatures, the paleoen-vironmental preferences of chiasmoliths have not yetbeen accurately delineated, and a simplistic relationship
between D/C ratios and climatic fluctuation may notexist. Moreover, their dearth in the northern higherlatitudes, both in the late Paleocene and earlyOligocene, in comparison to the southern high latitudeswhere they are abundant in both of these intervals,makes their unquestioned use as paleoenvironmentalindicators suspect.
The early Oligocene biogeographic map of the paleo-Atlantic (Figure 3) shows a latitudinally more differen-tiated nannoflora than the late Paleocene. The midlatitudes are dominated by the intermediate Cyclo-coccolithus neogammation-Dictyococcites hesslandiiassemblage. This assemblage also forms an importantpart of the northern high latitudes, but here thereticulofenestrid and Coccolithus pelagicus assemblagesare equally abundant. The southern high latitude site(329) is dominated by reticulofenestrids and chiasmo-liths. These differences, particularly the lack of Cyclo-coccolithus neogammation-Dictyococcites hesslandii atSite 329, indicate a cooler southern Atlantic relative tothe northern ocean during the early Oligocene. This dis-
753
B. U. HAQ, G. P. LOHMANN, S. W. WISE, JR.
6.00
8.00
10.00
12.00
14.00
~ 16.00
18.00
20.00
22.00
24.0060°S 30°S 0°
PALEOLATITUDE
30°N 60°N
Figure 7. Time and paleolatitudinal distributions of Miocene assemblages identified by Q-mode Varimax Factor Analysis.Sizes of circles are proportional to abundance of the assemblage in each sample. Discoaster assemblage.
754
CENOZOIC NANNOPLANKTON BIOGEOGRAPHY AND PALEOCLIMATIC IMPLICATIONS
6.00
8.00
10.00
12.00
14.00
16.00
18.00
20.00
22.00
24.0060° S 30° S 0°
PALEOLATITUDE
Figure 8. Time and paleolatitudinal distributions of Miocene assemblages identified by Q-mode Varimax Factor Analysis.Sizes of circles are proportional to abundance of the assemblage in each sample. Coccolithus pelagicus (s. ampl.) assemblage.
755
B. U. HAQ, G. P. LOHMANN, S. W. WISE, JR.
6.001-
8.00 -
22.00 -
24.0060°S 30° S 30° N
PALEOLATITUDE60°N
Figure 9. Time and paleolatitudinal distributions of Miocene assemblages identified by Q-mode Varimax Factor Analysis.Sizes of circles are proportional to abundance of the assemblage in each sample. Dictyococcites minutus assemblage.
756
CENOZOIC NANNOPLANKTON BIOGEOGRAPHY AND PALEOCLIMATIC IMPLICATIONS
6.00
8.00
10.00
12.00
14.00>>
E 16.001-
18.00
20.00 -
22.00
24.00
ó
60°S 30°S 0°PALEOLATITUDE
30° N 60° N
Figure 10. Time and paleolatitudinal distributions of Miocene assemblages identified by Q-mode Varimax Factor Analysis.Sizes of circles are proportional to abundance of the assemblage in each sample. Cyclococcolithus aff. stradneri assemblage.
757
B. U. HAQ, G. P. LOHMANN, S. W. WISE, JR.
6 . O O 1 —
8.00 -
Φ Φ
Φ
60°S 30°S 0°
PALEOLATITUDE
30° N 60°N
Figure 11. Time and paleolatitudinal distributions of Miocene assemblages identified by Q-mode Varimax Factor Analysis.Sizes of circles are proportional to abundance of the assemblage in each sample. Dictyococcites antarcticus assemblage.
758

CENOZOIC NANNOPLANKTON BIOGEOGRAPHY AND PALEOCLMATIC IMPLICATIONS
TABLE 3Discoaster: Chiasmolithus Ratios, Site 329
Sample(Interval in cm)
32-1,86-8832-1, 127-12932-1, 143-14432-4, 5-732-4, 33-3532-4, 67-6932-4, 107-10932-4, 127-12933-1, 94-9533-1, 12533-1, 139-14133-2,9-1133-2, 70-7233-2, 125-126145-14633-3, 10-1233-3, 70-7233-3,118-12033-3, 143-14533-4, 77-8033-4, 100-10233-4, 134-135
(Discoaster:Chiasmolithus)
26:27424:27612:28859:24142:25823:27750-25078:22248:25225:27562:238
146:15430:27012:28811:28911:28926:27464:23692:2084:2961:2993:297
Ratio
1:111:111:241:41:61:121:51:31:51:111:41:11:91:241:261:261:111:41:21:741:2991:99
crepancy could be explained by invoking a paleo-GulfStream and its warming influence; Sites 112 and 117would lie directly in its path. Alternatively, the general-ly cooler southern high latitudes during this period maybe due to the same reason that eventually caused theglobal climatic deterioration in the early Oligocene, i.e.,the onset of Antarctic glaciation near theEocene/Oligocene boundary and increased bottomwater production and circulation (Kennett et al., 1975).This general trend of cooler southern high latitudes ex-tends into the Miocene as evidenced by the dominanceof the cool Dictyococcites antarcticus assemblage at Site329.
TAXONOMIC NOTES
Cyclococcolithus aff. stradneri Jafar, Verh. Kon. Ned. Akad. Wet.Afd. Natuurk., v. 28, no. 1, p. 75, 76, pi. 9, fig. 22-25.
Remarks: The small circular placoliths observed in thetropical/subtropical Atlantic DSDP sites have dimensions similar toC. stradneri. However, the present species does not show a prominentcentral pore. The placoliths have relatively large thickness comparedto the diameter of the shields, and the two shields are almost alwaysseparated from each other, leaving a knob on one shield and a"pseudo-pore" on the other.
Dictyococcites minutus (Haq) n. comb. Basionym: Prinsius minutusHaq, Stockholm Contrib. Geol., v. 25, no. 2, p. 78, pi. 7, fig. 4,5.
ACKNOWLEDGMENTS
This paper was reviewed by W.A. Berggren and R.C.Tjalsma. Research was supported by Grant GA 21983 of theSubmarine Geology and Geophysics program, OceanographySection of the National Science Foundation. This is WoodsHole Oceanographic Institution Contribution No. 3748.
REFERENCES
Bukry, D., 1973. Coccolith and silicoflagellate stratigraphy,Tasman Sea and Southwestern Pacific Ocean, Deep SeaDrilling Project Leg 21. In Burns, R.E., Andrews, J.E., etal., Initial Reports of the Deep Sea Drilling Project,Volume 21: Washington (U.S. Government Printing Of-fice), p. 885-893.
Edwards, A.R. and Perch-Nielsen, K., 1975. Calcareous nan-nofossils from the southern SW Pacific, DSDP Leg 29. InKennett, J.P., Houtz, R.E., et al., Initial Reports of theDeep Sea Drilling Project, Volume 29: Washington (U.S.Government Printing Office), p. 469-539.
Haq, B.U., 1976. Coccoliths in cores from BellingshausenAbyssal Plain and Antarctic continental Rise (DSDP Leg35). In Craddock, C, Hollister, CD., et al., Initial Reportsof the Deep Sea Drilling Project, Volume 35: Washington(U.S. Government Printing Office), p. 557.
Haq, B.U. and Lohmann, G.P., 1976. Early Cenozoiccalcareous nannoplankton biogeography of the AtlanticOcean: Marine Micropaleo., v. 1, no. 2.
Kennett, J.P., Houtz, R.E., et al., 1975. Cenozoic paleo-oceanography in the Southwest Pacific Ocean, Antarcticglaciation, and the development of the Circum-AntarcticCurrent. In Kennett, J.P., Houtz, R.E., et al., InitialReports of the Deep Sea Drilling Project, Volume 29:Washington (U.S. Government Printing Office), p. 1155-1169.
759





























