Embed Size (px)
Citation preview

1
Title: Relative diversity and community structure of ciliates in stream biofilms 1
according to molecular and microscopy methods 2
Running title: Diversity of ciliates in stream biofilms 3
Andrew Dopheide1, Gavin Lear1, Rebecca Stott2 and Gillian Lewis1* 4
School of Biological Sciences, University of Auckland, 3a Symonds Street, Auckland, New 5
Zealand1, and National Institute for Water and Atmospheric Research, P.O. Box 11-115, 6
Hamilton, New Zealand2 7
*Corresponding author. Mailing address: School of Biological Sciences, University of Auckland, 8
Private Bag 92019, Auckland, New Zealand. Phone: 64 (9) 373 7599. Fax 64 (9) 373 7416. E-9
mail: [email protected] 10
11
ABSTRACT 12
Ciliates are an important component of aquatic ecosystems, acting as predators of bacteria and 13
protozoa and providing nutrition for organisms at higher trophic levels. Understanding of the 14
diversity and ecological role of ciliates in stream biofilms is limited, however. Ciliate diversity in 15
biofilm samples from four differently impacted streams was assessed using microscopy and T-16
RFLP analysis of 18S rRNA sequences. Analysis of both 3' and 5' terminal fragments yielded very 17
similar estimates of ciliate diversity. The diversity detected using microscopy was consistently 18
lower than that suggested by T-RFLP analysis, indicating the existence of genetic diversity not 19
apparent to morphological examination. Both microscopy and T-RFLP analyses provided similar 20
relative trends in diversity between different streams, with the lowest level of biofilm-associated 21
ciliate diversity in samples from the least-impacted stream and the highest diversity in samples 22
from moderately to highly impacted streams. Multivariate analysis provided evidence of 23
significantly different ciliate communities in biofilm samples from different streams and seasons, 24
particularly between a highly degraded urban stream and less impacted streams. Microscopy and 25
T-RFLP data both suggested the existence of widely distributed, resilient biofilm-associated ciliates 26
as well as ciliate taxa restricted to sites with particular environmental conditions, with 27
cosmopolitan taxa being more abundant than those with restricted distributions. Differences 28
between ciliate assemblages were associated with water quality characteristics typical of urban 29
Copyright © 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.00412-09 AEM Accepts, published online ahead of print on 26 June 2009
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

2
stream degradation, and may be related to factors including nutrient availability and 30
macroinvertebrate communities. Microscopic and molecular techniques were considered to be 31
useful complementary approaches for investigation of biofilm ciliate communities. 32
33
INTRODUCTION 34
Heterotrophic micro-eukaryotes such as ciliates are thought to be of considerable importance in 35
aquatic ecosystems, as they are major predators of bacteria and constitute a nutritional resource 36
for other protozoa, invertebrates and probably fish larvae (9, 22, 36, 52, 62, 63, 71). In addition, 37
protozoan bacterivory contributes to enhanced decomposition of leaf detritus—a vital nutrient 38
resource in streams—by increasing turnover of bacterial populations through predation (57). It is 39
not well understood, however, how ciliate diversity and community structure in streams are 40
affected by changing environmental conditions, or how ciliate communities affect other stream 41
biota and processes. The effects of various physical, chemical and biological factors on freshwater 42
protozoan communities have been considered by a number of studies, but most of these have 43
focused upon planktonic organisms in lentic habitats (for example, 2, 11, 44). However, the 44
complex microbial communities in biofilms have been recognised as important contributors to 45
critical ecological processes such as primary production, nitrogen fixation and nutrient cycling, and 46
may underpin the function of stream food webs (31, 45, 61). The few studies which have 47
investigated benthic habitats in lotic systems have found evidence of the existence of diverse 48
communities of abundant ciliates (3, 20, 56) and shifts in community structure in response to 49
ecophysiological parameters (30, 42, 43). With one exception, however, these investigations were 50
based on aquatic sediments, and the organisms within epilithic biofilms have continued to receive 51
little attention. 52
Most studies of ciliate diversity and ecology have utilised microscopy-based methods of 53
identification (for example, 3, 56), as ciliate cells are relatively large and morphologically diverse. 54
Such methods demand a high level of taxonomic expertise, however, and are difficult and time-55
consuming—for example, many ciliates are fragile and fast moving, and often require difficult 56
fixing and staining protocols for reliable identification. Molecular biological tools offer the 57
possibility of more accurate and efficient methods for protozoan study, and may provide a useful 58
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

3
complement to traditional approaches (12, 18, 28, 65), yet we know of only a few molecular studies 59
of environmental ciliate diversity (18, 20, 37). A series of recent investigations have used culture-60
independent analysis of 18S rRNA gene sequences to reveal the existence of diverse micro-61
eukaryote communities in assorted marine, anoxic and extreme environments (40, 48, 66, 69, 70, 62
72). Furthermore, a growing body of evidence suggests the existence of significant genetic diversity 63
among various ciliate taxa which has escaped detection by microscopy (14, 18, 23, 34, 60, 64, 78), 64
pointing to the potential for molecular techniques to generate new insights into ciliate diversity and 65
ecology, and suggesting a need for comparison of the effectiveness of these different techniques in 66
environmental samples. 67
Terminal restriction fragment length polymorphism (T-RFLP) analysis provides an efficient, 68
inexpensive and semi-quantitative means for comparing microbial molecular diversity between 69
different samples, and has been widely used to investigate bacterial communities, although only a 70
handful of studies have applied T-RFLP methods to the analysis of micro-eukaryote diversity (6, 16, 71
17). In this study, ciliate diversity and community structure was investigated in biofilm samples 72
from streams representing a range of levels of anthropogenic degradation, with the objective of 73
testing the null hypothesis that human impacts have no effect upon this important heterotrophic 74
component of stream ecosystems. To achieve this, ciliate-targeted PCR primers were used in 75
conjunction with T-RFLP and multivariate statistical analyses. Additionally, ciliate diversity 76
measures obtained using molecular techniques were compared with those derived from 77
microscopy-based methods in order to assess the relative effectiveness of these approaches. 78
79
MATERIALS AND METHODS 80
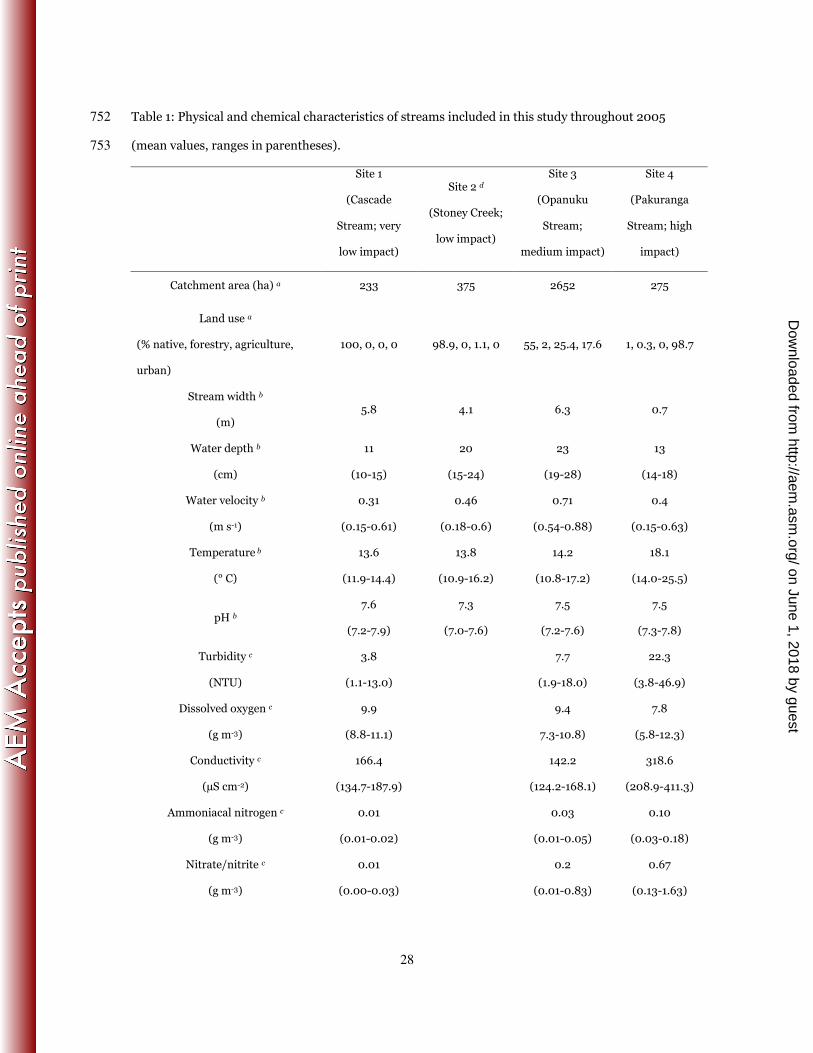
Sampling sites. Biofilm samples were collected from each of four differently impacted streams in 81
Auckland, New Zealand. Site 1 (Cascade Stream) is a largely unimpacted stream, located in an 82
undeveloped native forest catchment (36°53'32"S, 174°31'07"E). Site 2 (Stoney Creek) is mildly 83
impacted, located in a partially developed native forest catchment with nearby houses and roads 84
(36°54'24"S, 174°34'06"E). Site 2 is a lower order tributary of Site 3 (Opanuku Stream) which is 85
proximate to rural agricultural development and is moderately impacted (36°53'42"S, 174°35'44"E). 86
Site 4 (Pakuranga Stream) is located in a highly developed urban catchment (36°53'50"S, 87
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

4
174°54'21"E) and is highly impacted. Sites 1, 2 and 3 all have natural stony substrates whilst Site 4 88
consists of a concrete channel at the sampling location. 89
Sites 1 and 3 are respectively ranked as having the best and 5th-best water quality of 25 streams 90
throughout the Auckland region based on monthly monitoring between 1995 and 2005; three 91
locations in the Site 4 stream catchment are ranked in the worst five (4, 5). Physical and chemical 92
attributes of the streams are presented in Table 1. 93
Sample collection. Quantitative methods for sampling biofilm material and associated protozoa 94
from submerged surfaces were developed. There are no clearly established protocols for sampling 95
protozoa associated with epilithic biofilms in lotic systems, and for this reason two methods were 96
tested in this study. For both methods stream biofilm was collected from substrate surfaces while 97
submerged, to avoid the potential loss of material upon removal of stones from the water column 98
(29). The first method involved the use of sterile Speci-Sponges® (Nasco, Fort Atkinson, WI, USA) 99
to thoroughly swab submerged surfaces (rocks or concrete channel) within a 55 cm2 area defined 100
by a circular neoprene template. Dislodged biofilm material was then squeezed from the collecting 101
sponges into sterile Whirl-Pak® bags (Nasco). 102
The second biofilm collection method involved a brush/syringe sampler based on devices 103
recommended for subsurface sampling of epilithic periphyton (1, 39, 54, 67). The brush/syringe 104
sampler, illustrated in figure S1 in the supplemental material, consisted of a 60 ml syringe with its 105
end removed to create a wide opening, and a toothbrush head glued to the end of the syringe 106
plunger. A rubber ring was attached to the end of the syringe to seal the sampler against the rock 107
surface and to minimise the loss of dislodged material due to water currents. Biofilm material was 108
removed from a 4.91 cm2 area by pressing down and rotating the syringe plunger. Loosened 109
material was drawn up into a 10 ml collection syringe attached to the base of the larger syringe with 110
plastic tubing. Samples were then decanted into sterile Whirl-Pak® bags (Nasco). 111
Biofilm sampling was carried out during January (summer), May (autumn), August (winter) and 112
November (spring) of 2005. On each sampling occasion, biofilm material was collected from two 113
20 m reaches of each stream. In general, the exposed surfaces of 4-10 randomly selected rocks 114
(220-550 cm2 in total) were sampled using the sponge method, and the surfaces of 10 rocks (about 115
50 cm2 in total) were sampled using the syringe method, from within each 20 m reach of each 116
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

5
stream. Similarly, ten samples were collected from each of two 20 m reaches along the concrete 117
channel at Site 4, using each sampling method. The ten samples obtained using each method at 118
each sample point were combined, giving a total of four composite samples per stream (one sponge 119
sample and one syringe sample from each of two sampling points in each stream). Samples were 120
chilled on ice for transport. 121
Assessment of ciliate diversity by microscopy-based analysis. Samples were stored at 4 122
°C and analysed within four to ten hours of collection. For the enumeration of ciliates, sub-samples 123
of 1 ml were transferred to a Sedgewick Rafter cell and scanned at 25 x magnification to generate 124
preliminary lists of taxa. Sub-samples were then examined at 200 x to 630 x magnification. Due to 125
low density of biofilm material and associated ciliates, concentration of samples from Site 1 prior to 126
examination was typically required, as follows: 25-100 ml samples were concentrated by filtering 127
through 25 µm nylon mesh and backwashing the retentate into a graduated 15 ml tube using 128
filtered water (typical final volume ~3 ml). Aliquots (1 ml) were then transferred to a Sedgewick 129
Rafter cell and examined at 200 x to 630 x magnification. Ciliate species were identified to at least 130
genus level, where possible, using criteria described in taxonomic keys (25, 53). Photographs were 131
used to ensure that identifications were consistent. The relative abundance of different taxa was 132
scored on a scale of one to eight, with each value respectively corresponding to an approximate 133
abundance of 1-5, 5-10, 10-15, 15-20, 20-50, 50-100, 100-200, and over 200 cells per ml. 134
DNA extraction and PCR amplification. Sub-samples (30 ml) of each combined biofilm 135
sample were transferred to pre-weighed sterile 35 ml centrifuge tubes and centrifuged at 6000 x g 136
for 10 min at 4 °C. Supernatants were removed and pellets resuspended in 15 % glycerol to achieve 137
final concentrations of 100 to 200 mg biofilm ml-1. Samples were then frozen at -80 °C until 138
required. 139
DNA was extracted from biofilm samples as previously described (20). Following extraction, the 140
concentration of DNA in each extract was assessed using a Quant-iT PicoGreen dsDNA kit 141
(Invitrogen, Auckland, New Zealand) according to manufacturer’s directions, in combination with 142
absorbance measurements at 260 nm using a Nanodrop ND-1000 spectrophotometer (Thermo 143
Fisher Scientific, MA, USA), and electrophoresis on 1 % agarose gels stained with Sybr Safe 144
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

6
(Invitrogen). Based on combined results of these procedures, the concentration of DNA in extracts 145
was standardised to approximately 20 ng µl-1 before use as template in PCR reactions. 146
PCR primers 384F (5'-YTB GAT GGT AGT GTA TTG GA-3'; 20) and 1147R (5'-GAC GGT ATC TRA 147
TCG TCT TT-3'; 20), targeting a ~700 bp fragment of the ciliate 18S rRNA gene, were respectively 148
labelled at their 5' termini with HEX and FAM fluorophores (Invitrogen). DNA was amplified from 149
biofilm extracts using these primers in 50 µL PCR reaction mixes (25 µL GoTaq Green master mix 150
(Promega, In Vitro Technologies, Auckland, New Zealand), 0.5 µM forward and reverse primers, 151
0.4 % BSA (Invitrogen) and 2 µL of template DNA). The following PCR protocol was used: initial 152
incubation for 5 min at 94 °C, then 30 amplification cycles of 45 s at 94 °C, 60 s at 55 °C and 90 s at 153
72 °C, followed by a final extension step of 7 min at 72 °C. PCR products were purified using a 154
Purelink PCR Purification Kit (Invitrogen) according to manufacturer’s instructions. The 155
concentration of fluorescently-labelled PCR products was determined by absorbance 156
measurements at 260 nm using a Nanodrop ND-1000 spectrophotometer. 157
Terminal restriction fragment length polymorphism analysis. Terminal restriction 158
fragment polymorphism (T-RFLP) analysis is a semi-quantitative molecular fingerprinting 159
technique which provides an efficient method of comparing populations. Fluorescently-labelled 160
PCR products are digested with one or more restriction enzymes, resulting in the production of 161
fluorescently-labelled terminal fragments, the length (bp) and abundance of which can be 162
automatically detected. This results in the generation of profiles in which the number of peaks 163
indicates the number of different terminal fragments present whilst the height and area of peaks 164
indicate their relative abundance. As terminal fragment length (bp) varies across different taxa, 165
this data can provide a profile of community structure within each sample. 166
For T-RFLP analysis in this study, the DNA concentration in each purified PCR product was 167
adjusted to 20 ng µl-1. PCR products were digested with the restriction endonucleases HaeIII and 168
RsaI (Invitrogen) in 10 µl reactions, incubated overnight at 37 ° C. Each digestion reaction 169
contained 1 U of each enzyme, 1 µl reaction buffer (Invitrogen) and approximately 175 ng of 170
purified amplicon. Digested samples were electrophoresed alongside a size standard with markers 171
at 20 bp intervals up to 1200 bp (LIZ1200; Applied Biosystems, Melbourne, Australia). Terminal 172
restriction fragments were detected using a 3130XL Genetic Analyzer (Applied Biosystems). This 173
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

7
resulted in generation of peak profiles representing the abundance of HEX- and FAM-labelled 174
terminal fragments, which were analysed using GeneMapper 4.0 (Applied Biosystems), which 175
automatically calculates the number, height and area of peaks and their corresponding fragment 176
lengths (bp). Profiles from two runs of each sample were compared to check for consistency, and 177
any inconsistent results were discarded. T-RFLP peaks with a height of ≤50 rfu (relative 178
fluorescence units) and of less than 10 base pairs in length were excluded from analysis to 179
eliminate background interference. Each remaining peak was presumed to represent a different 180
terminal restriction fragment and hence a different ciliate-derived 18S rRNA gene sequence. Peaks 181
representing terminal fragments in excess of about 650 base pairs in length were assumed to 182
represent PCR products which were not cut during restriction digestion. 183
The length (bp) and area of HEX- and FAM-labelled peaks in each T-RFLP profile was imported 184
into Microsoft Excel. Peak positions were rounded to the nearest whole number, and the overall 185
area of each profile was standardised to 1, to ensure comparability between samples. 186
Statistical analyses. ANOVA was used to test for significant differences among numbers of 187
ciliate taxa detected in samples from different streams in each month. Significant ANOVA results 188
were followed with post-hoc Tukey-Kramer HSD multiple comparison tests. Differences between 189
numbers of taxa in samples obtained using the two different sampling methods, and according to 190
the two different analysis methods, were investigated using t-tests. These analyses were carried out 191
in JMP 7.0 (SAS Institute Inc., USA). As each different PCR product can be expected to produce 192
both a HEX-labelled fragment (primer 384F) and a FAM-labelled fragment (primer 1147R), the 193
numbers of HEX-labelled and FAM-labelled T-RFLP peaks were averaged to provide a composite 194
diversity estimate for each sample. 195
For multivariate analysis, data from both HEX-labelled and FAM-labelled T-RFLP peaks were 196
combined into a single data set for each sample, and subjected to a square-root transformation to 197
moderate the influence of large peaks in subsequent analyses. Relative abundance data obtained by 198
microscopy were not transformed. Multivariate analyses were carried out in Primer v6.1.6 (Primer-199
E Ltd., UK). Bray-Curtis similarity between all pairs of biofilm samples was calculated. Results 200
were ordinated by non-metric multidimensional scaling (MDS), which clusters samples with higher 201
levels of pairwise similarity more closely than samples with lower pairwise similarity, thereby 202
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

8
allowing the visualisation of similarities and differences among sets of samples. ANOSIM was used 203
to test the null hypotheses of no significant differences between different streams, seasons, and 204
sampling methods. ANOSIM compares within-group similarity and between-group similarity; R 205
values of around zero indicate within-group and between-group similarity are the same, whilst R 206
values approaching 1 indicate samples within groups are more similar to each other than to 207
samples from different groups, allowing the null hypothesis to be rejected (15). 208
209
RESULTS 210
Sampling methods. 211
The effectiveness of sponge- and syringe-based biofilm sampling methods for detecting ciliates was 212
compared, but there was no clear evidence of greater efficacy of either method in terms of ciliate 213
diversity detected, according to either microscopy or T-RFLP. Little evidence for significant 214
differences in ciliate community structure was found between sponge-derived and syringe-derived 215
samples according to ANOSIM (Table 2), and furthermore, sponge- and syringe-derived samples 216
from the same stream and sampling date were typically grouped very closely in MDS plots, 217
suggesting very similar composition of the communities sampled by each method. Sponge and 218
syringe-derived samples from each site and date were therefore pooled for subsequent analyses. 219
Ciliate diversity in stream biofilm samples (microscopy and T-RFLP methods). Our 220
methodology targeted a standard area of substrate for all samples, and our diversity results 221
represent the number of taxa detected per area sampled. However, biofilm biomass was 222
particularly sparse at Site 1, especially in winter (G. Lewis and S. Tsai, unpublished data). Similarly, 223
density of ciliates at Site 1 was generally very low, necessitating concentration of these samples for 224
analysis. Even after concentration, the number of ciliate cells detected and identified in Site 1 225
samples was low, and this may have affected the level of diversity detected. 226
The number of ciliate taxa detected in stream biofilm samples using microscopy-based methods 227
ranged from zero for samples from Site 1 in May and August, to 17 for a Site 4 sample from May 228
(Figure 1). Site 1 typically had the fewest biofilm-associated ciliate taxa, whilst samples from Site 3 229
and Site 4 contained the highest numbers of ciliate taxa. Significant differences were detected 230
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

9
among microscopy results from May and November (ANOVA, p < 0.05), but not from January or 231
August. 232
The number of different peaks present in a T-RFLP profile is assumed to reflect the number of 233
different ciliate 18S rRNA gene sequences—and therefore the number of ciliate taxa—present in the 234
stream biofilm sample. The number of T-RFLP peaks detected ranged from five (Site 1, November) 235
to 61 (Site 3, January) (Figure 1). Significant differences were detected in all months (ANOVA, p < 236
0.005). The pattern of ciliate diversity across different streams according to T-RFLP analysis was 237
broadly similar to that derived from microscopic investigations, with the lowest ciliate diversity 238
detected in the most pristine stream and higher diversity at the more impacted sites. 239
Overall, in each stream the average number of peaks detected in T-RFLP profiles exceeded the 240
average number of ciliate taxa detected by microscopy (t-test, p < 0.0001) (Figure 2). In total, 183 241
different HEX-labelled terminal fragments and 191 different FAM-labelled terminal fragments 242
were detected among all samples, compared with 68 different ciliate taxa identified by microscopy. 243
Differences in ciliate diversity and community structure between streams and 244
seasons according to microscopy and T-RFLP. Microscopy-based analysis of biofilm 245
samples found evidence of ciliate taxa common to multiple stream environments as well as many 246
taxa restricted to individual sites (Figure 3). Taxa common to all four streams were detected only in 247
January and November, although these taxa were always present in at least one stream throughout 248
the year. In January a greater number of unique taxa were found in Site 2 than in Site 3. In all 249
other months, taxa unique to Site 3 and Site 4 together accounted for the majority of taxa detected, 250
although these two streams also had generally higher overall levels of diversity than the other sites. 251
Taxa unique to Site 4 were particularly frequent in August. In November, taxa unique to Site 2 were 252
not detected, and taxa unique to Site 1 were evident only in May. Fewer than one in five of the taxa 253
unique to particular streams were detected on more than one sampling date. 254
Overall, only 7 % of ciliate taxa identified throughout the year using microscopy were common to 255
all four streams (Figure 3). Forty-four percent of the detected taxa were each found in only one 256
stream, most commonly Site 3 or Site 4, and no taxa were unique to the least impacted stream, Site 257
1. 258
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

10
According to microscopy, ciliates common to all four streams were generally small-sized species 259
from Oligohymenophorea or Phyllopharyngea, such as Glaucoma sp., Trochilia sp. or Cyclidium sp. 260
These ubiquitous species were generally more abundant in the more impacted streams. Several 261
unidentified hypotrichs were typically characteristic of Site 2 biofilm samples, whilst taxa unique to 262
Site 3 included Actinobolina sp., Aspidisca lynceus and sessile peritrich species such as Epistylis sp. 263
and Vorticella sp. A large number of taxa were found only at Site 4, including Spirostomum sp., 264
Stylonychia sp., Euplotes sp., Strombilidium sp., and predatory taxa including Monodinium sp. 265
and several species of Litonotus. 266
Visual inspection of T-RFLP profiles shows that some peaks are present in profiles from multiple 267
streams (although these peaks are often markedly different in magnitude) whilst other peaks 268
appear to be unique to particular stream biofilms (Figure 4). Overall, 17 % of the different T-RFLP 269
peaks detected throughout the year were found in all four streams. Forty five percent of the T-270
RFLP peaks occurred only in one stream (Figure 3), consistent with the microscopy data. The 271
proportion unique to each stream ranged from 5 % for Site 1 to 16 % for Site 3. 272
The proportions of T-RFLP peaks found in different streams showed a higher degree of consistency 273
between months than the microscopy data (Figure 3). Peaks unique to each of the four streams 274
were detected in all months except August, when biofilm biomass at Site 1 was insufficient for 275
molecular analysis. Compared with the microscopy data, the proportion of T-RFLP peaks unique to 276
Site 3 and Site 4 together accounted for less of the total diversity detected, except for in January 277
samples. Peaks unique to Site 1 were more frequently detected, however, as were peaks unique to 278
Site 2 in November samples. As for the microscopy data, T-RFLP peaks unique to Site 4 were most 279
frequent in August samples, although the proportion of peaks unique to Site 3 was highest in 280
January. 281
The T-RFLP peaks found in all four streams throughout the year (17 % of all peaks) together 282
accounted for 75 % of total profile area, indicating that the corresponding terminal fragments were 283
relatively abundant (Figure 5). Conversely, the peaks that were each detected in only one stream 284
(45 % of all peaks) accounted for less than 6 % of the total T-RFLP profile area, and as for the 285
microscopy data, few of these unique T-RFLP peaks were detected on more than one sampling date. 286
These findings suggest the existence of populations of abundant and cosmopolitan ciliate taxa in 287
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

11
stream biofilms from different environments, together with rarer taxa of low abundance and with 288
restricted spatial and temporal distributions. 289
Microscopy relative abundance data and T-RFLP peak area data were used to generate non-metric 290
multidimensional scaling (MDS) plots in which each data point represents the assemblage of ciliate 291
taxa or T-RFLP peaks detected in one sample. The proximity of the data points to each other 292
reflects the relative similarity of their ciliate assemblages. MDS plots based on microscopy data 293
show separation of samples from highly impacted Site 4 and moderately impacted Site 3 from each 294
other, and from samples from less impacted Site 2 and Site 1 (Figure 6). Similarly, MDS plots based 295
on T-RFLP data show Site 4 samples forming a clearly separated group, while Site 3 and Site 2 296
samples form overlapping clusters, and Site 1 samples are scattered widely. Temporal patterns are 297
less clear, with little discernable grouping of data points by sampling month according to 298
microscopy data. According to T-RFLP data, November samples and May samples from Site 1 are 299
more dispersed than January and August samples. 300
ANOSIM analysis of microscopy and T-RFLP data provided evidence of significant differences 301
between ciliate communities in different streams, and between samples from different seasons 302
(Table 3). For the microscopy data, significant and generally large differences were found between 303
ciliate assemblages from all streams, and moderate differences between assemblages from different 304
sampling dates. Similarly, significant differences were found between ciliate assemblages from all 305
streams according to T-RFLP analysis, with the exception of Site 1 and Site 2. The largest 306
differences according to T-RFLP were found between Site 4 and both Site 2 and Site 3, while the 307
largest differences according to microscopy were found between Site 1 and Site 3, and Site 3 and 308
Site 4. The largest difference between sampling months according to T-RFLP analysis was between 309
January and August, but these months showed relatively little difference according to microscopy. 310
Significant differences were not detected between January and May, and January and November T-311
RFLP results. 312
Links between environmental data and ciliate assemblage MDS data. Links between 313
ciliate community assemblage data and environmental trends can be visualised as bubbles overlaid 314
on MDS ordination plots, with the size of bubbles representing the magnitude of environmental 315
parameters at sites and dates corresponding to biofilm sampling occasions. The resulting figures 316
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

12
suggest observed MDS ordination patterns are associated with a combination of environmental 317
factors (Figure 7). Separation of Site 4 samples from others appears to be associated with factors 318
typically associated with urban stream degradation, such as higher levels of nitrogenous 319
compounds and lower levels of dissolved oxygen, in addition to very low levels of forest cover. 320
Turbidity, temperature and phosphorus levels are similarly elevated at Site 4 on certain sampling 321
occasions, while water depth, water velocity and pH do not follow this trend. Conversely, Site 1 322
samples are associated with high native forest cover, low levels of nitrogenous compounds, total 323
phosphorus and turbidity, and elevated oxygen. Grouping of Site 3 samples in a cluster adjacent to 324
Site 4 samples appears to be related to intermediate levels of nitrogenous compounds and total 325
phosphorus, although pH and levels of dissolved reactive phosphorus are generally lower than for 326
Site 1 samples. 327
328
DISCUSSION 329
Ciliate diversity according to microscopy and T-RFLP analysis. Ciliates have been 330
considered very amenable to microscopic study due to their high level of morphological diversity 331
and relatively large size. However, it has been suggested that a current list of described ciliate 332
morphospecies may contain five to ten times as many biological species (13, 24, 26). Furthermore, 333
ciliate morphospecies have been shown to include organisms with clearly different ecophysiological 334
characteristics (75). This suggests that the morphospecies concept may substantially underestimate 335
ciliate species diversity and ecosystem complexity (26, 75). Morphologically identical but 336
genetically, physiologically or biochemically divergent ciliates are likely to occupy separate 337
ecological niches, and measurement of this functional ciliate diversity is relevant to ecological 338
studies. While molecular techniques have recently contributed to great insights into protistan 339
diversity in various inaccessible and extreme environments, few studies have applied these 340
techniques to specific phyla such as Ciliophora. 341
This investigation found that the number of ciliate taxa suggested by T-RFLP analysis was more 342
than double that indicated by microscopy. This suggests the existence of a significant component of 343
genetic ciliate diversity in stream biofilms which is not evident to microscopic examination of 344
morphology. A similar finding was made in a recent study of oligotrich ciliate diversity in seawater, 345
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

13
with diversity according to molecular analysis about 10 fold higher than according to 346
morphological observations (18). In the present study, ciliates were identified using simple light 347
microscopy methods, and although most species can be detected at relatively low magnification, it 348
is possible that some organisms may have been overlooked due to small size or inconspicuousness, 349
or due to being present in encysted form. Silver staining procedures and electron microscopy can 350
improve taxonomic discrimination of ciliate taxa based on morphological and morphometrical 351
analysis. However, even when these more complex methods have been used, ciliate diversity based 352
on molecular analysis has still been found to exceed diversity according to morphology (34). A 353
growing number of studies provide evidence of cryptic molecular diversity exceeding apparent 354
morphological diversity in various ciliate taxa, including Carchesium (78), Cyclidium (23), 355
Halteria (34), Oxytricha (60), Strombidium (34), Tetrahymena (41, 49, 64) and Zoothamnium 356
(14). The consistency of these findings suggests little reason to expect any difference in the great 357
majority of ciliate taxa which have not yet been subjected to genetic analysis. Cyclidium and 358
ribosomal RNA gene sequences closely matching those of Oxytricha, Tetrahymena and 359
Zoothamnium have all previously been detected in these Auckland streams (20), and therefore may 360
have contributed to the cryptic genetic diversity detected using T-RFLP in this study. 361
Although our molecular analysis indicates a high level of genetic diversity underlying ciliate 362
morphospecies, the PCR primers used in this study, while highly ciliate-specific, are not perfectly 363
so (20). It is possible that a limited number of non-ciliate sequences and resulting terminal 364
restriction fragments may be represented in our results. However, this effect may be outweighed by 365
the tendency of T-RFLP analysis to underestimate diversity of closely related taxa, due to 366
conservation of restriction sites and consequent generation of terminal fragments of identical 367
length (16). Additionally, ciliate taxa with indistinguishable 18S rRNA gene sequences may be 368
discriminated by examination of other genes (41), suggesting that the sequences targeted in this 369
study may not provide complete resolution of different species. It thus seems likely that the 370
assessments of ciliate diversity provided by T-RFLP analyses in this study are conservative. 371
Limitations and complementarity of methods. Although T-RFLP analysis is an efficient 372
method of obtaining and comparing microbial genetic diversity data, interpretation of T-RFLP 373
information, in isolation, remains challenging. T-RFLP analysis lacks a straightforward means of 374
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

14
reliably assigning taxonomic identities to observed peaks in complex samples, particularly for 375
groups of organisms for which availability of DNA sequence data is limited, such as ciliates. The 376
value of diversity measures derived from T-RFLP analyses has been questioned, on the bases that 377
different organisms may contribute to single T-RFLP peaks, different restriction enzymes will 378
produce different results, and the use of thresholds to eliminate background “noise” from T-RFLP 379
profiles means terminal fragments (and organisms) of low abundance will be excluded from the 380
resulting analysis (10). Multivariate statistical analysis of T-RFLP data is considered reliable 381
however, with conclusions little affected by the exclusion of minor T-RFLP peaks or the choice of 382
restriction enzyme (8, 77). T-RFLP is thus a useful method for comparing complex microbial 383
community structures. 384
Microscopy-based analysis of morphology does permit identification of ciliates, subject to sufficient 385
taxonomic expertise being available, and can allow the classification of ciliates into functional 386
categories such as feeding groups which can be used to examine the ecological role of protozoa (55). 387
The level of taxonomic resolution used can affect whether significant differences between 388
protozoan communities will be detected, with identification of protozoa to taxonomic levels higher 389
than genus being less effective in discriminating surface-associated protozoan communities (35). 390
Microscopy does have the advantage of quantitative power–cells can be counted, albeit tediously—391
which may be lacking in PCR-based assays. Clearly, morphological identification is only possible 392
within the constraints of the morphospecies concept, which has recognised limitations (26). 393
In this study, both T-RFLP and microscopy-based analyses showed broadly similar overall trends 394
of diversity in the different streams, and both methods produced evidence of significant differences 395
between ciliate communities in differently impacted stream biofilms. It seems, therefore, that T-396
RFLP and microscopic analyses may be considered as complementary methods, the former 397
providing a robust and efficient method for comparing ciliate community structure, and the latter 398
allowing attribution of differences between microbial assemblages and systems to particular ciliate 399
taxa or functional groups. Of course, group-targeted PCR primers such as those used in this study 400
can be used for cloning and sequencing which—if sequence and morphological data have been 401
reconciled—does allow reliable identification of microbes in environmental samples, thus avoiding 402
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

15
one of the limitations of T-RFLP. FISH-based techniques offer a useful means of linking molecular 403
sequence data with microscopy-derived morphological information (68). 404
Further studies combining molecular and microscopic methods are necessary for the expansion of 405
currently limited sequence database coverage of micro-eukaryotes (21). Assuming availability of 406
micro-eukaryote DNA sequence information improves, it seems probable that in future molecular 407
identification methods may prove more straightforward and accurate than morphology-based 408
methods. It is nonetheless likely that combined approaches may prove more informative than 409
either molecular or microscopy-based techniques alone, however (68, 73). Molecular profiling 410
methods allow efficient and robust comparisons of community structure, while identification and 411
description of taxa using sequencing, microscopy, and FISH-based techniques can provide 412
additional insights and links to ecological, phenotypic and physiological information, and may 413
allow pinpointing of ecologically important organisms. 414
Differences between biofilm ciliate communities in different streams. Both microscopy 415
and T-RFLP analysis methods have provided clear evidence of differences in ciliate assemblages in 416
biofilms from differently impacted streams. These differences can be associated with 417
environmental parameters typical of urban stream degradation, suggesting that our null hypothesis, 418
that human impacts have no effect upon biofilm-associated ciliate communities, can be rejected. 419
The fewest ciliate taxa were found at Site 1, a relatively pristine stream, while the most ciliate taxa 420
were detected in samples from moderately impacted Site 3 and highly impacted Site 4. This trend 421
of greater diversity at the more impacted sites seems contrary to the generally accepted tendency 422
for ecological perturbation to lead to a simplification of community structure. A number of 423
contrasting biotic and abiotic factors suggest possible reasons for this finding. Site 1 and Site 2 are 424
characterised by little exposure to sunlight or anthropogenic pollutants. Site 3 and Site 4, in 425
contrast, are exposed to elevated nitrogen loads respectively derived from nearby areas of 426
agricultural and urban land use. Reduced density and height of riparian vegetation expose Site 2 427
and Site 3 to more sunlight than Site 1, and Site 4 receives virtually no shade whatsoever. Nutrient 428
enrichment and sunlight have been shown to promote periphyton growth in lotic systems (27, 32, 429
74), suggesting that the more impacted sites are likely to have communities of more abundant 430
phototrophic organisms compared to the less impacted streams. This is consistent with 431
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

16
observations of increased biofilm biomass and more diverse bacterial and algal (especially diatom) 432
communities at sites 3 and 4 (G. Lewis and S. Tsai, unpublished data). The three less impacted 433
sites in this study also receive significant amounts of allochthonous debris from surrounding 434
vegetation whilst Site 4 does not, suggesting a difference in the relative importance of detritus-435
derived nutrients in these streams. Elevated nutrient availability can increase benthic ciliate 436
abundance in rivers and streams (19, 50, 58), and may affect ciliate abundance, biomass and 437
community composition in lentic habitats (2, 33, 51, 76). It seems possible that the abundant 438
biofilms in the more impacted streams in this study may provide more resources and a wider 439
variety of feeding niches for heterotrophic protozoan organisms. The greater variety and 440
abundance of bacterivorous, algivorous and predatory ciliates detected in samples from the two 441
more impacted sites is consistent with this suggestion. 442
Site 1 is home to diverse and abundant benthic macroinvertebrates, whilst Site 4 has a 443
macrobenthic invertebrate fauna of very low diversity, consisting almost entirely of chironomid 444
larvae (38). Biofilms at Site 1 may therefore be subjected to very different grazing pressures than 445
biofilms at Site 4, which is likely to further affect the nutrient resources available in these streams. 446
Macroinvertebrates may also negatively affect protozoa through predation (51, 71). In addition, 447
macroinvertebrates consume meiofauna such as rotifers (59), which may also predate upon 448
protozoa (47). Studies of effects of top-down predation pressures on ciliates in lakes and ponds 449
have had mixed results (2, 51, 76). There is very little information available on the nature of trophic 450
interactions between ciliates and invertebrates in stream biofilms, although one study found 451
evidence of invertebrate predation and/or competition negatively affecting biofilm-associated 452
ciliates (46). Nevertheless, it can be speculated that the homogeneity of the invertebrate 453
community at Site 4 may mean that ciliates of only certain types and sizes are subjected to 454
predation pressures, resulting in the selective proliferation of non-target taxa. In contrast, the 455
diverse invertebrates at Site 1 may represent a broad competitive and predatory factor, perhaps 456
contributing to the lower abundance and diversity of ciliates at this site. 457
Different catchment land uses may cause development of different biofilm-associated ciliate 458
assemblages by favouring tolerant taxa while eliminating sensitive organisms. Being surrounded by 459
extensive urban development, Site 4 is probably exposed to various pollutants in addition to 460
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

17
increased levels of nitrogenous compounds, higher temperatures and lower levels of dissolved 461
oxygen. Furthermore, the artificial substrate in Site 4 may lack refugia for flow-sensitive or light-462
sensitive organisms. A previous investigation suggested Site 4 may be home to fewer sessile 463
peritrich taxa and a higher frequency of predatory ciliates such as Litonotus sp. and Loxophyllum 464
sp. than the other streams investigated in this study (20). Similarly, in this study, predatory ciliates 465
such as Monodinium sp. and Litonotus sp. were detected by microscopy-based analysis only in Site 466
4 biofilm. These, and other characteristic taxa identified in Site 4, are typically β-α mesosaprobic or 467
polysaprobic tolerant species, indicating that they can tolerate reasonably high organic loads and 468
hence more heavily polluted environments (25). This suggests that physico-chemical conditions in 469
Site 4 influence the development of a very different ciliate community compared to the less 470
impacted streams included in this study. How this different ciliate community affects the ecological 471
processes and interactions occurring in this stream awaits further investigation. 472
Conclusion. Ciliates and other protozoa are major predators of bacteria, and provide an 473
important trophic link in aquatic habitats such as stream biofilms (52). Understanding protozoan 474
community diversity and abundance is therefore important for gaining insights into the function of 475
these “hot spots” of microbial activity, which contribute substantially to ecosystem processes in 476
streams (7). Both molecular and microscopy-based analyses provided evidence of diverse biofilm-477
associated ciliate communities, with greater diversity in the more impacted streams and significant 478
differences between ciliate assemblages in streams in different states of degradation. These 479
observations may be related to a variety of differences in environmental parameters characteristic 480
of urban stream degradation, such as elevated nutrient and sunlight availability, as well as different 481
assemblages of autotrophic biofilm organisms and communities of benthic macroinvertebrates. 482
The discrepancy between numbers of taxa suggested by T-RFLP analysis compared to microscopy-483
based analysis in this study adds to evidence that ciliate diversity has been underestimated by 484
traditional microscopic approaches. Microscopic analysis allowed identification of ciliate taxa 485
characteristic of different stream environments, however. Future application of these 486
complementary techniques will improve our understanding of the causes and effects of stream 487
degradation at the microbial level, leading to development of more effective stream monitoring and 488
remediation strategies. 489
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

18
490
ACKNOWLEDGEMENTS 491
We wish to thank anonymous reviewers for their helpful contributions to the manuscript. Funding 492
for this research was provided by the New Zealand Foundation for Research, Science and 493
Technology Public Good Science Fund UOA306. 494
495
REFERENCES 496
1. Aloi, J. E. 1990. A critical review of recent freshwater periphyton field methods. Canadian 497
Journal of Fisheries & Aquatic Sciences 47:656-670. 498
2. Andrushchyshyn, O. P., A. K. Magnusson, and D. D. Williams. 2006. Responses of 499
intermittent pond ciliate populations and communities to in situ bottom-up and top-down 500
manipulations. Aquatic Microbial Ecology 42:293-310. 501
3. Andrushchyshyn, O. P., K. P. Wilson, and D. D. Williams. 2007. Ciliate 502
communities in shallow groundwater: Seasonal and spatial characteristics. Freshwater 503
Biology 52:1745-1761. 504
4. ARC. 2007. River Water Quality: State and Trends in the Auckland Region. Auckland 505
Regional Council Technical Publication 336. 506
5. ARC. 2007. State of the Environment Monitoring: Rivers & Streams Water Quality Data 507
Report 2005. Auckland Regional Council Technical Publication 327. 508
6. Baldwin, A. J., J. A. Moss, J. D. Pakulski, P. Catala, F. Joux, and W. H. Jeffrey. 509
2005. Microbial diversity in a Pacific Ocean transect from the Arctic to Antarctic circles. 510
Aquatic Microbial Ecology 41:91-102. 511
7. Battin, T. J., L. A. Kaplan, J. D. Newbold, and C. M. E. Hansen. 2003. 512
Contributions of microbial biofilms to ecosystem processes in stream mesocosms. Nature 513
426:439-442. 514
8. Bennett, L. T., S. Kasel, and J. Tibbits. 2008. Non-parametric multivariate 515
comparisons of soil fungal composition: Sensitivity to thresholds and indications of 516
structural redundancy in T-RFLP data. Soil Biology and Biochemistry 40:1601-1611. 517
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

19
9. Berninger, U. G., B. J. Finlay, and P. Kuuppo-Leinikki. 1991. Protozoan control of 518
bacterial abundances in freshwater. Limnology & Oceanography 36:139-147. 519
10. Blackwood, C. B., D. Hudleston, D. R. Zak, and J. S. Buyer. 2007. Interpreting 520
ecological diversity indices applied to terminal restriction fragment length polymorphism 521
data: Insights from simulated microbial communities. Applied and Environmental 522
Microbiology 73:5276-5283. 523
11. Burns, C. W., and L. M. Galbraith. 2007. Relating planktonic microbial food web 524
structure in lentic freshwater ecosystems to water quality and land use. Journal of Plankton 525
Research 29:127-139. 526
12. Caron, D. A., P. D. Countway, and M. V. Brown. 2004. The growing contribution of 527
molecular biology and immunology to protistan ecology: molecular signatures as ecological 528
tools. Journal of Eukaryotic Microbiology 51:38-48. 529
13. Chao, A., P. C. Li, S. Agatha, and W. Foissner. 2006. A statistical approach to 530
estimate soil ciliate diversity and distribution based on data from five continents. Oikos 531
114:479-493. 532
14. Clamp, J. C., and D. Williams. 2006. A molecular phylogenetic investigation of 533
Zoothamnium (Ciliophora, Peritrichia, Sessilida). Journal of Eukaryotic Microbiology 534
53:494-498. 535
15. Clarke, K. R. 1993. Non-parametric multivariate analyses of changes in community 536
structure. Australian Journal of Ecology 18:117-143. 537
16. Countway, P. D., R. J. Gast, P. Savai, and D. A. Caron. 2005. Protistan diversity 538
estimates based on 18S rDNA from seawater incubations in the Western North Atlantic. 539
Journal of Eukaryotic Microbiology 52:95-106. 540
17. Díez, B., C. Pedrós-Alió, T. L. Marsh, and R. Massana. 2001. Application of 541
denaturing gradient gel electrophoresis (DGGE) to study the diversity of marine 542
picoeukaryotic assemblages and comparison of DGGE with other molecular techniques. 543
Applied and Environmental Microbiology 67:2942-2951. 544
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

20
18. Doherty, M., B. A. Costas, G. B. McManus, and L. A. Katz. 2007. Culture-545
independent assessment of planktonic ciliate diversity in coastal northwest Atlantic waters. 546
Aquatic Microbial Ecology 48:141-154. 547
19. Domènech, R., A. Gaudes, J. C. López-Doval, H. Salvadó, and I. Muñoz. 2006. 548
Effects of short-term nutrient addition on microfauna density in a Mediterranean stream. 549
Hydrobiologia 568:207-215. 550
20. Dopheide, A., G. Lear, R. Stott, and G. Lewis. 2008. Molecular characterization of 551
ciliate diversity in stream biofilms. Applied and Environmental Microbiology 74:1740-1747. 552
21. Duff, R. J., H. Ball, and P. J. Lavrentyev. 2008. Application of combined 553
morphological-molecular approaches to the identification of planktonic protists from 554
environmental samples. Journal of Eukaryotic Microbiology 55:306-312. 555
22. Finlay, B. J., and G. F. Esteban. 1998. Freshwater protozoa: Biodiversity and ecological 556
function. Biodiversity and Conservation 7:1163-1186. 557
23. Finlay, B. J., G. F. Esteban, S. Brown, T. Fenchell, and K. Hoef-Emden. 2006. 558
Multiple cosmopolitan ecotypes within a microbial eukaryote morphospecies. Protist 559
157:377-390. 560
24. Foissner, W. 2006. Biogeography and dispersal of micro-organisms: A review 561
emphasizing protists. Acta Protozoologica 45:111-136. 562
25. Foissner, W., and H. Berger. 1996. A user-friendly guide to the ciliates (Protozoa, 563
Ciliophora) commonly used by hydrobiologists as bioindicators in rivers, lakes, and waste 564
waters, with notes on their ecology. Freshwater Biology 35:375-482. 565
26. Foissner, W., A. Chao, and L. A. Katz. 2008. Diversity and geographic distribution of 566
ciliates (Protista: Ciliophora). Biodiversity and Conservation 17:345-363. 567
27. Francoeur, S. N. 2001. Meta-analysis of lotic nutrient amendment experiments: 568
Detecting and quantifying subtle responses. Journal of the North American Benthological 569
Society 20:358-368. 570
28. Fried, J., R. Psenner, W. Ludwig, and K. H. Schleifer. 2002. Improvement of ciliate 571
identification and quantification: A new protocol for fluorescence in situ hybridization 572
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

21
(FISH) in combination with silver stain techniques. Systematic and Applied Microbiology 573
25:555-571. 574
29. Fukuda, M., A. Ashida, Y. Tomaru, and S. I. Nakano. 2004. An improved method 575
for collecting heterotrophic microorganisms living on pebbles in streams. Limnology 5:41-576
46. 577
30. Gu�cker, B., and H. Fischer. 2003. Flagellate and ciliate distribution in sediments of a 578
lowland river: Relationships with environmental gradients and bacteria. Aquatic Microbial 579
Ecology 31:67-76. 580
31. Hall Jr., R. O., and J. L. Meyer. 1998. The trophic significance of bacteria in a detritus-581
based stream food web. Ecology Letters 79:1995-2012. 582
32. Hillebrand, H. 2002. Top-down versus bottom-up control of autotrophic biomass - A 583
meta-analysis on experiments with periphyton. Journal of the North American 584
Benthological Society 21:349-369. 585
33. Hillebrand, H., M. Kahlert, A. L. Haglund, U. G. Berninger, S. Nagel, and S. 586
Wickham. 2002. Control of microbenthic communities by grazing and nutrient supply. 587
Ecology 83:2205-2219. 588
34. Katz, L. A., G. B. McManus, O. L. O. Snoeyenbos-West, A. Griffin, K. Pirog, B. 589
Costas, and W. Foissner. 2005. Reframing the 'everything is everywhere' debate: 590
Evidence for high gene flow and diversity in ciliate morphospecies. Aquatic Microbial 591
Ecology 41:55-65. 592
35. Kepner Jr., R. L., and J. R. Pratt. 1996. Characterization of surface-associated 593
protozoan communities in a Lake Erie coastal wetland (Old Woman Creek, Ohio). Journal 594
of Great Lakes Research 22:63-76. 595
36. Kneitel, J. M., and J. M. Chase. 2004. Disturbance, predator, and resource interactions 596
alter container community composition. Ecology 85:2088-2093. 597
37. Lara, E., C. Berney, H. Harms, and A. Chatzinotas. 2007. Cultivation-independent 598
analysis reveals a shift in ciliate 18S rRNA gene diversity in a polycyclic aromatic 599
hydrocarbon-polluted soil. FEMS Microbiology Ecology 62:365-373. 600
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

22
38. Lear, G., I. K. G. Boothroyd, S. J. Turner, K. Roberts, and G. D. Lewis. 2009. A 601
comparison of bacteria and benthic invertebrates as indicators of ecological health in 602
streams. Freshwater Biology 54:1532-1543. 603
39. Loeb, S. L. 1981. An in situ method for measuring the primary productivity and standing 604
crop of the epilithic periphyton community in lentic systems. Limnology and Oceanography 605
26:394-399. 606
40. López-García, P., F. Rodríguez-Valera, C. Pedrós-Alió, and D. Moreira. 2001. 607
Unexpected diversity of small eukaryotes in deep-sea Antarctic plankton. Nature 409:603-608
607. 609
41. Lynn, D. H., and M. C. Stru�der-Kypke. 2006. Species of Tetrahymena identical by 610
small subunit rRNA gene sequences are discriminated by mitochondrial cytochrome c 611
oxidase I gene sequences. Journal of Eukaryotic Microbiology 53:385-387. 612
42. Madoni, P., and N. Bassanini. 1999. Longitudinal changes in the ciliated protozoa 613
communities along a fluvial system polluted by organic matter. European Journal of 614
Protistology 35:391-402. 615
43. Madoni, P., and S. Braghiroli. 2007. Changes in the ciliate assemblage along a fluvial 616
system related to physical, chemical and geomorphological characteristics. European 617
Journal of Protistology 43:67-75. 618
44. Madoni, P., and F. Sartore. 2003. Long-term changes in the structure of ciliate 619
communities in a small isolated pond. Italian Journal of Zoology 70:313-320. 620
45. Marxsen, J. 2006. Bacterial production in the carbon flow of a central European stream, 621
the Breitenbach. Freshwater Biology 51:1838-1861. 622
46. McCormick, P. V. 1991. Lotic protistan herbivore selectivity and its potential impact on 623
benthic algal assemblages. Journal of the North American Benthological Society 10:238-624
250. 625
47. McCormick, P. V., and J. Cairns Jr. 1991. Effects of micrometazoa on the protistan 626
assemblage of a littoral food web. Freshwater Biology 26:111-119. 627
48. Moon-van der Staay, S. Y., R. De Wachter, and D. Vaulot. 2001. Oceanic 18S rDNA 628
sequences from picoplankton reveal unsuspected eukaryotic diversity. Nature 409:607-610. 629
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

23
49. Nanney, D. L., C. Park, R. Preparata, and E. M. Simon. 1998. Comparison of 630
sequence differences in a variable 23S rRNA domain among sets of cryptic species of 631
ciliated protozoa. Journal of Eukaryotic Microbiology 45:91-100. 632
50. Norf, H., H. Arndt, and M. Weitere. 2009. Responses of biofilm-dwelling ciliate 633
communities to planktonic and benthic resource enrichment. Microbial Ecology 57:687-634
700. 635
51. Pace, M. L., and E. Funke. 1991. Regulation of planktonic microbial communities by 636
nutrients and herbivores. Ecology 72:904-914. 637
52. Parry, J. D. 2004. Protozoan grazing of freshwater biofilms, p. 167-196, Advances in 638
Applied Microbiology, vol. 54. 639
53. Patterson, D. J. 1998. Free-living Freshwater Protozoa. Manson Publishing Ltd., London. 640
54. Persson, J., H. Hansson, and I. Renberg. 2006. The Stone Brusher, a new sampler for 641
submerged epilithic material in shallow streams and lakes. Hydrobiologia 560:385-392. 642
55. Pratt, J. R., and J. Cairns Jr. 1985. Functional groups in the protozoa: Roles in 643
differing ecosystems. Journal of Protozoology 32:415-423. 644
56. Reiss, J., and J. M. Schmid-Araya. 2008. Existing in plenty: Abundance, biomass and 645
diversity of ciliates and meiofauna in small streams. Freshwater Biology 53:652-668. 646
57. Ribblett, S. G., M. A. Palmer, and D. W. Coats. 2005. The importance of 647
bacterivorous protists in the decomposition of stream leaf litter. Freshwater Biology 648
50:516-526. 649
58. Rublee, P. A., and A. Partusch-Talley. 1995. Microfaunal response to fertilization of an 650
Arctic tundra stream. Freshwater Biology 34:81-90. 651
59. Schmid-Araya, J. M., and P. E. Schmid. 2000. Trophic relationships: Integrating 652
meiofauna into a realistic benthic food web. Freshwater Biology 44:149-163. 653
60. Schmidt, S. L., D. Bernhard, M. Schlegel, and W. Foissner. 2007. Phylogeny of the 654
Stichotrichia (Ciliophora; Spirotrichea) reconstructed with nuclear small subunit rRNA 655
gene sequences: Discrepancies and accordances with morphological data. Journal of 656
Eukaryotic Microbiology 54:201-209. 657
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

24
61. Sheldon, F., and K. F. Walker. 1997. Changes in biofilms induced by flow regulation 658
could explain extinctions of aquatic snails in the lower River Murray, Australia. 659
Hydrobiologia 347:97-108. 660
62. Sherr, E. B., and B. F. Sherr. 1994. Bacterivory and herbivory: key roles of 661
phagotrophic protists in pelagic food webs. Microbial Ecology 28:223-235. 662
63. Sherr, E. B., and B. F. Sherr. 2002. Significance of predation by protists in aquatic 663
microbial food webs. Antonie van Leeuwenhoek, International Journal of General and 664
Molecular Microbiology 81:293-308. 665
64. Simon, E. M., D. L. Nanney, and F. P. Doerder. 2008. The "Tetrahymena 666
pyriformis" complex of cryptic species. Biodiversity and Conservation 17:365-380. 667
65. Sims, G. P., R. Aitken, and A. Rogerson. 2002. Identification and phylogenetic 668
analysis of morphologically similar naked amoebae using small subunit ribosomal RNA. 669
Journal of Eukaryotic Microbiology 49:478-484. 670
66. Slapeta, J., D. Moreira, and P. Lopez-Garcia. 2005. The extent of protist diversity: 671
Insights from molecular ecology of freshwater eukaryotes. Proceedings of the Royal Society 672
- Biological Sciences (Series B) 272:2073-2081. 673
67. Stockner, J. G., and F. A. J. Armstrong. 1971. Periphyton of the experimental lakes 674
area, northwestern Ontario. Journal of the Fisheries Reseach Board of Canada 28:215-229. 675
68. Stoeck, T., S. S. Epstein, and W. H. Fowle. 2003. Methodology of Protistan Discovery: 676
From rRNA Detection to Quality Scanning Electron Microscope Images. Applied and 677
Environmental Microbiology 69:6856-6863. 678
69. Stoeck, T., B. Hayward, S. S. Epstein, G. T. Taylor, and R. Varela. 2006. A 679
multiple PCR-primer approach to access the microeukaryotic diversity in environmental 680
samples. Protist 157:31-43. 681
70. Stoeck, T., G. T. Taylor, and S. S. Epstein. 2003. Novel eukaryotes from the 682
permanently anoxic Cariaco Basin (Caribbean Sea). Applied and Environmental 683
Microbiology 69:5656-5663. 684
71. Stoecker, D. K., and J. McDowell Capuzzo. 1990. Predation on protozoa: its 685
importance to zooplankton. Journal of Plankton Research 12:891-908. 686
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

25
72. Takishita, K., M. Tsuchiya, M. Kawato, K. Oguri, H. Kitazato, and T. Maruyama. 687
2007. Genetic diversity of microbial eukaryotes in anoxic sediment of the saline meromictic 688
Lake Namako-ike (Japan): On the detection of anaerobic or anoxic-tolerant lineages of 689
eukaryotes. Protist 158:51-64. 690
73. Vaerewijck, M. J. M., K. Sabbe, J. Bare, and K. Houf. 2008. Microscopic and 691
molecular studies of the diversity of free-living protozoa in meat-cutting plants. Applied 692
and Environmental Microbiology 74:5741-5749. 693
74. Von Schiller, D., E. Martí, J. L. Riera, and F. Sabater. 2007. Effects of nutrients and 694
light on periphyton biomass and nitrogen uptake in Mediterranean streams with 695
contrasting land uses. Freshwater Biology 52:891-906. 696
75. Weisse, T., and S. Rammer. 2006. Pronounced ecophysiological clonal differences of 697
two common freshwater ciliates, Coleps spetai (Prostomatida) and Rimostrombidium 698
lacustris (Oligotrichida), challenge the morphospecies concept. Journal of Plankton 699
Research 28:55-63. 700
76. Wickham, S. A., S. Nagel, and H. Hillebrand. 2004. Control of epibenthic ciliate 701
communities by grazers and nutrients. Aquatic Microbial Ecology 35:153-162. 702
77. Zhang, R., V. Thiyagarajan, and P. Y. Qian. 2008. Evaluation of terminal-restriction 703
fragment length polymorphism analysis in contrasting marine environments. FEMS 704
Microbiology Ecology 65:169-178. 705
78. Zhang, W. J., J. Yang, Y. H. Yu, S. W. Shu, and Y. F. Shen. 2006. Population 706
genetic structure of Carchesium polypinum (Ciliophora: Peritrichia) in four Chinese lakes 707
inferred from ISSR fingerprinting: High diversity but low differentiation. Journal of 708
Eukaryotic Microbiology 53:358-363. 709
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

26
Figure 1: Numbers of ciliate taxa detected by microscopy and numbers of ciliate 18S rRNA gene 710
sequences indicated by T-RFLP analysis of stream biofilm samples. Numbers 1 and 2 denote 711
samples from two different points within each stream. Each bar is the mean of counts from two 712
samples (microscopy data), or the mean of HEX-labelled and FAM-labelled terminal fragment 713
counts from two samples (T-RFLP data). Error bars are +/- one standard deviation. Significant 714
differences were found between streams during each month according to T-RFLP, and in May and 715
November according to microscopy data (ANOVA, p < 0.05). Within each month, samples not 716
linked by the same letter (A - C) are significantly different (Tukey-Kramer HSD, p = 0.05). 717
* Samples from Site 1 were typically concentrated due to low biomass and ciliate abundance; 718
insufficient biofilm biomass in August for T-RFLP analysis. 719
720
Figure 2: Average numbers of ciliate taxa detected by microscopy (white), and average numbers of 721
ciliate 18S rRNA gene sequences indicated by T-RFLP analysis (grey), in stream biofilm samples 722
throughout one year. Each bar is the mean of counts from eight samples (microscopy data), or the 723
mean of HEX-labelled and FAM-labelled terminal fragment counts from eight samples (T-RFLP 724
data). Error bars are +/- one standard deviation. The average number of T-RFLP peaks detected 725
significantly exceeded the average number of taxa identified by microscopy in all cases (t-test, p < 726
0.0001). 727
728
Figure 3: Comparison of richness of ciliate taxa occurring in biofilms from different stream 729
environments, according to microscopy and T-RFLP analysis. 730
* No T-RFLP data for Site 1 in August due to insufficient biofilm biomass for analysis. 731
732
Figure 4: T-RFLP profiles derived from stream biofilm samples in November 2005, using ciliate-733
specific PCR primers. Sizes of grey and black peaks respectively indicate abundance of HEX-734
labelled and FAM-labelled terminal restriction fragments. 735
736
Figure 5: Abundance and diversity of ciliates in stream biofilms according to T-RFLP profiles. Bars 737
represent the combined area of T-RFLP peaks found in profiles from individual streams only, and 738
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

27
peaks found in profiles from multiple streams, as a proportion of total profile area for each 739
sampling date. Numbers of peaks contributing to each bar are indicated. 740
* No T-RFLP data from Site 1 for August. 741
742
Figure 6: MDS ordination of ciliate assemblages in stream biofilm samples based on microscopy 743
data (left; 2D stress = 0.15) and T-RFLP profiles (right; 2D stress = 0.19). Data are labelled by 744
sample location (top) and date (bottom). 745
746
Figure 7: MDS ordination of ciliate assemblages in stream biofilm samples, based on T-RFLP 747
analysis, showing magnitude of various environmental parameters associated with samples (2D 748
stress = 0.19). Bubble sizes are scaled to reflect the ranges of values indicated in table 1. Samples 749
for which environmental data is unavailable are omitted. 750
751
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
28
Table 1: Physical and chemical characteristics of streams included in this study throughout 2005 752
(mean values, ranges in parentheses). 753
Site 1
(Cascade
Stream; very
low impact)
Site 2 d
(Stoney Creek;
low impact)
Site 3
(Opanuku
Stream;
medium impact)
Site 4
(Pakuranga
Stream; high
impact)
Catchment area (ha) a 233 375 2652 275
Land use a
(% native, forestry, agriculture,
urban)
100, 0, 0, 0 98.9, 0, 1.1, 0 55, 2, 25.4, 17.6 1, 0.3, 0, 98.7
Stream width b
(m) 5.8 4.1 6.3 0.7
Water depth b
(cm)
11
(10-15)
20
(15-24)
23
(19-28)
13
(14-18)
Water velocity b
(m s-1)
0.31
(0.15-0.61)
0.46
(0.18-0.6)
0.71
(0.54-0.88)
0.4
(0.15-0.63)
Temperature b
(° C)
13.6
(11.9-14.4)
13.8
(10.9-16.2)
14.2
(10.8-17.2)
18.1
(14.0-25.5)
pH b 7.6
(7.2-7.9)
7.3
(7.0-7.6)
7.5
(7.2-7.6)
7.5
(7.3-7.8)
Turbidity c
(NTU)
3.8
(1.1-13.0)
7.7
(1.9-18.0)
22.3
(3.8-46.9)
Dissolved oxygen c
(g m-3)
9.9
(8.8-11.1)
9.4
7.3-10.8)
7.8
(5.8-12.3)
Conductivity c
(µS cm-2)
166.4
(134.7-187.9)
142.2
(124.2-168.1)
318.6
(208.9-411.3)
Ammoniacal nitrogen c
(g m-3)
0.01
(0.01-0.02)
0.03
(0.01-0.05)
0.10
(0.03-0.18)
Nitrate/nitrite c
(g m-3)
0.01
(0.00-0.03)
0.2
(0.01-0.83)
0.67
(0.13-1.63)
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

29
Total Kjeldahl nitrogen c
(g m-3)
0.31
(0.21-0.93)
0.33
(0.21-1.14)
0.75
(0.20-1.80)
Total nitrogen c
(g m-3)
0.22
(0.20-0.92)
0.41
(0.20-0.84)
0.96
(0.42-2.24)
Dissolved reactive phosphorus c
(g m-3)
0.021
(0.013-0.031)
0.016
(0.010-0.024)
0.026
(0.017-0.045)
Total phosphorus c
(g m-3)
0.032
(0.023-0.046)
0.045
(0.024-0.064)
0.178
(0.044-0.336)
a Data generated using the Land Cover database 2 (Terralink International Ltd, NZ). 754
b Data recorded by authors. 755
c Data from reference 5, unavailable for site 2; NTU denotes nephelometric turbidity units. 756
d Site 2 is an upstream tributary of site 3. It is presumed that water quality at site 2 is the same or 757
better than at site 3. 758
759
Table 2: Comparison of ciliate diversity in sponge and syringe samples according to microscopy 760
and T-RFLP analysis (ANOSIM). 761
Differences according to microscopy Differences according to T-RFLP
Sample comparison Global R statistic p Global R statistic p
January sponge-January syringe a 0.438 0.07
May sponge-May syringe 0.031 0.48 -0.106 0.67
August sponge-August syringe 0.167 0.44 -0.086 0.59
November sponge-November syringe a -0.313 1.00
a Molecular analysis of January sponge and November syringe samples was unsuccessful. 762
763
Table 3: ANOSIM comparison of ciliate assemblages in samples from different streams and 764
sampling dates according to microscopy and T-RFLP data. Statistically significant results (p < 0.05) 765
are indicated *. 766
Stream comparison Sampling date comparison
R statistic p R statistic p
Microscopy dataa
on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

30
Global comparison 0.719 0.001 * Global comparison 0.383 0.001 *
Site 1-Site 2 0.414 0.001 * January-May 0.487 0.002 *
Site 1-Site 3 0.988 0.001 * January-August 0.284 0.011 *
Site 1-Site 4 0.798 0.001 * January-November 0.469 0.001 *
Site 2-Site 3 0.748 0.001 * May-August 0.257 0.011 *
Site 2-Site 4 0.621 0.001 * May-November 0.428 0.001 *
Site 3-Site 4 0.842 0.001 * August-November 0.418 0.001 *
T-RFLP datab
Global comparison 0.49 0.001 * Global comparison 0.395 0.001 *
Site 1-Site 2 0.269 0.102 January-May 0.201 0.105
Site 1-Site 3 0.497 0.01 * January-August 0.722 0.002 *
Site 1-Site 4 0.508 0.006 * January-November 0.125 0.222
Site 2-Site 3 0.332 0.037 * May-August 0.502 0.001 *
Site 2-Site 4 0.769 0.002 * May-November 0.433 0.006 *
Site 3-Site 4 0.678 0.001 * August-November 0.376 0.020 *
a
N (microscopy data) = 13-16 samples per stream or month. 767
bN (T-RFLP data) = 8-12 samples per stream, 8-15 samples per month. 768 on June 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from



















![[OPS] One Piece 659 Fr (fairynopiece.shonenblog.com)](https://img.pdfslide.us/doc/110x75/568c39051a28ab0235a0d231/ops-one-piece-659-fr-fairynopieceshonenblogcom.jpg)












![[Shinobi] One Piece 659](https://img.pdfslide.us/doc/110x75/568bdcbd1a28ab2034b348d0/shinobi-one-piece-659.jpg)



