Embed Size (px)
Citation preview

BIOLOGICALCONTROL
Habitat Diversification for Improving Biological ControlAbundance of Anagrus epos (Hymenoptera Mymaridae) in
Grape Vineyards
BROOK C MURPHY JAY A ROSENHEIM ANDJEFFREY GRANETT
Department of Entomology Universityof California Davis CA 95616
Environ Entomol 25(2) 495-504 (1996)ABSTRACT A frequently cited habitat diversificationtactic is the use of prune tree refugesthat support overwintering populations of Anagrus epos (Girault) a mymarid egg parasite ofthe western grape leafhopperErythroneura elegantula Osborn in vineyardsHere we test theeffect of prune trees on early-season abundance of adult A epos in vineyards A epos wasfound in vineyards downwind of prune trees at more than twice the densities of vineyardslackingpnllle trees despite significantvariation in A epos immigration from sources outsidethe pnme tree-vineyard system Densities of A epos overwintering within prune trees ex-plained a significantamount of the variation in A epos trap capture in vineyardsFurthermoreanother factor associatedwith prune trees was found to influence A epos abundance in vine-yards a windbreak effect created by the prune trees concentrated dispersing A epos on theleeward side of the prune trees tllereby further enhancing A epos numbers
KEY WORnS Anagrus epos Erythroneura elegantula habitat diversificationconservationbiologicalcontrol grape
HABITATDIVERSIFICATIONHASlong been promot-ed as a tactic to conserve natural enemies and en-hance biological control of insect pests and as aintegrated method of developing sustainable pestcontrol systems in production agriculture Both ap-proaches argue that habitat diversification can pro-vide essential resources for natural enemies suchas feeding sites alternate hosts or prey or over-wintering sites that can enhance their abundancein the surrounding environment and help preventpest outbreaks (Herzog and Funderburk 1985 Na-tional Research Council 1989 van Emden 1990Altieri 1992)
Whether viewed as a biological control tactic oras an integrated approach to sustainable pest con-trol habitat diversification in or around crop fieldscan have a profound effect on how insect herbivoreand nahlral enemy populations interact An under-standing of the ecological mechanisms involvedwill also provide a rich resource for the develop-lIlent of pest management tactics in production ag-riculture (Herzog and Funderburk 1985)
One of the key hypotheses underlying the con-cept of habitat diversification is that many pestproblems are the result of a loss of habitat criticalfor supporting natural enemy populations (Letour-neau 1987 Russell 1989 Andow 1991) Althoughthe ecological basis behind habitat diversificationand natural enemy effectiveness has been dis-cllssed for many years and many proposed systemshave been evaluated in a preliminary way (re-viewed by Flint and Roberts 1988 Russell 1989
Altieri 1992) most studies have been qualitativerather than quantitative in nature Furthermorethere are no current tactics that have been widelyadopted for pest control in production agricultureHere we test and examine the ecological mecha-nisms associated with prune tree habitats plantedadjacent to grape vineyards on a key natural enemyspecies
Anagrus epos-Erythroneura elegantula SystemA frequently cited example used to illustrate theeffect of habitat diversification on natural enemyeffectiveness is the planting of overwintering hab-itats for Anagrus epos (Girault) near grape vine-yards to enhance biological control of the westerngrape leafhopper Erythroneura elegantula Os-born
Erythroneura elegantula is a major pest ofgrapes in many regions of the western UnitedStates High leafhopper numbers result in eco-nomic losses caused by cosmetic damage to grapeberries from leafhopper frass reduced vine vigorfrom heavy leaf feeding and leaf loss and fruitdamage from sun exposure Furthermore highdensities of adult leafhoppers can disrupt harvestby flying into the eyes nose and mouth of fieldlaborers
A key natural enemy of the grape leafhopper isA epos an important egg parasite of E elegantulaas well as other leafhopper species (Gordh andDunbar 1977 Williams 1984) A epos has beenreported to parasitize a significant proportion of Eelegantula eggs within commercial vineyards
0046-225X1960495-0504$02000 copy 1996 Entomolngical Society of America
496 ENVIRONMENTAL ENTOMOLOCY Vol 25 no 2
(Doutt and Nakata 1973) A major h1ctor hypoth-esized to be associated with the abundance andeffectiveness of A epos is the presence of nearbyhabitat that supports alternate leafhopper hosts(Doutt and Nakata 1965 1973) A epos overwin-ters within leafhopper eggs E elegantula overwin-ters as an adult thus an alternate source of leaf-hopper eggs is required to support overwinteringA epos numbers
Doutt and Nakata (1965) first observed thatvineyards located downwind from riparian habitatshad higher levels of E elegantula egg parasitismand lower E elegantula numbers relative to vine-yards located distant from these habitats Theyfound that blackberry brambles Rubus spp abun-dant in riparian habitats support Dikrella eaZifor-niea (Lawson) a year-round host of A epos Theyobserved that early spring emergence of A eposfrom blackberry results in earlier vineyard coloni-zation by parasites which coincided with oviposi-tion of overwintered E elegantula They also spec-ulated that earlier vineyard colonization at higherdensities produced a stronger numerical responseand thereby improved biological control (Douttand Nakata 1973) Unfortunately field trials deter-mined blackberry brambles grown away from ri-parian habitats did not maintain abundant popu-lations of A epos and further attempts wereabandoned (Flal1erty et al 1985)
Kido et at (1984) demonstrated that prune treessupport the prune leafhopper Edwardsiana prun-ieola (Edwards) which overwinters in the eggstage and serves as an overwintering host for Aepos Emergence of overwintering A epos fromoverwintering E prunieola eggs laid beneath thebark of prune twigs also was found to coincide with1st-generation E elegantula oviposition in vine-yards suggesting that pnme trees also serve as anoverwintering habitat for A epos Studies in bothcommercial and experimental plantings of pnilletrees demonstrated that A epos populations couldbe supported year-round Williams (1984) andKido et al (1984) also showed that A epos rearedfrom E prunieola or D ealifomiea can parasitizeE elegant1l1aeggs successfully and vice versa Fi-nally it has been shown that elevated A epos pop-ulations were associated with higher E elegant1l1aegg parasitism in vineyards near pnme trees (Kidoet al 1984 Pickett et aI 1990) Although thesestudies established the basic framework of thegrape leafhopper-A epos interaction there havebeen no replicated field trials that assess the effi-cacy of prune tree habitats or that test the ecolog-ical mechanisms underlying the effect
Anagrus epos Abundance Doutt and Nakata(1973) and Kido et al (1984) hypothesized thatoverwintering refuges near vineyards increase ear-ly-season A epos vineyard colonization as a resultof overwintering A epos emerging from refugesand dispersing into nearby vineyards They testedthese predictions in field trials using yellow stickytraps to measure early-season abundance of adult
A epos in vineyards or by examining E elegantulaeggs and comparing rates of egg parasitism Col-onization appeared to be enhanced in vineyardsdownwind from prune orchards or riparian habi-tats containing blackberry brambles They con-cluded that the greater abundance seen in fieldtrials supported the overwintering refuge hypoth-esis However because these initial experimentsused unreplicated comparisons the results al-though consistent with the refuge hypothesis werepreliminary in nature
The interpretation presented by Doutt and Nak-ata (1973) and Kido et a1(1984) also rested on anuntested assumption namely that an increase inparasite captures or egg parasitism near refugeswas the direct result of A epos emergence fromoverwintering sites A 2nd potential mechanismthat could explain increased parasite capturesdownwind of prune trees or trees found in riparianhabitats is a windbreak effect generated by thephysical structure of these habitats A windbreakeffect occurs when a structural barrier creates hlr-bulence in the airflow and a sheltered zone on theleeward side of the barrier The turbulent airflowmay act to increase the probability that a dispers-ing insect will settle into these sheltered zonesthereby enhancing colonization Dispersing insectsfrom many families including wasps in the familyMymaridae have been shown to accumulate be-hind windbreaks (Lewis and Stephenson 1966)Wilson et a1 (1989) showed that A epos reachhigher densities in pnmes when artificial wind-breaks are positioned on the upwind side of prunetrees Corbett and Rosenheim (in press) have alsorecently demonstrated a pattern of A epos cap-tures in vineyards downwind of pnme trees that isconsistent with the operation of a windbreak Thusenhanced A epos abundance could be the resultof overwintering populations within the prunetrees a windbreak effect or some combination ofthose 2 factors
In the current shldy we tested 2 key hypotheseswith respect to the effect of overwintering sites onA epos abundance in vineyards as follows (1) thatprune trees adjacent to vineyards enhance early-season A epos numbers in vineyards and (2) thatdifferences in abundance are the result of differ-ences in the densities of overwintering A eposemerging from pnme trees In addition to those 2hypotheses we also analyzed capture data for ev-idence of a windbreak effect influencing A eposabundance downwind of pnme trees
Materials and Methods
Experimental Design A epos colonizes virtu-ally all grape vineyards at some point during thegrowing season the time and density of abundancecan however be highly variable E elegantulapopulations also can be variable among vineyardsites in a given year Because of the vmiation ofboth parasite and leafhopper numbers a rigorous
April 1996 MURPHY ET AL HABITAT DIVERSIFICATION FOR BIOLOGICAL CONTHOL 497
Fil( 1 Pairedvineyardplot locationsin northernandctntra CaliforniaThe explodedview showsthe orien-tationof pnme tree and controlvineyardplots
evaluation of prune tree-enhanced biological con-trol required a large number of replicates Surveyswere conducted throughout the major wine grapeviticultural regions of central and northern Cali-fornia to locate 26 paired commercia] vineyardsone with adjacent upwind pnme trees and the oth-er lacking pnme trees (Fig 1) Paired vineyardswere matched for grape cultivar trellising anclmanagement practices Control (nonprune tree)vinlyards were positioned O~O kilometers fromrefuge sites and either parallel to or upwind ofpnll1e tree vineyard sites to minimize movementof ]eafllOppersor parasites between treatment andcontrol vineyards The paired plots were groupedinto 2 viticultural growing regions defined by thecUllmlative degree days occurring between Apriland October (Winkler et al 1974) Comparisonswere made between cooler growing regionslaquo3000 DD) and warmer regions (gt3000 DD)Two of the pnme tree sites used in the study wereplanted specifically for leafhopper control the re-mainder were commercial or remnant prune or-chards growing adjacent to the vineyard plots
Given the nature of the pnme tree habitat beingtested we were unable to ensure tnle randomiza-tion within our experimental design True random-ization would have required us to assign prune treeplantings randomly to plots and would have re-quired a delay of 3-5 yr for the pnme trees tobecome established and support large populationsof the alternate leafhopper host As a result weused vineyard sites located near previously estab-lished prune trees Had we chosen to work pri-
marily in vineyards with prune trees planted spe-cifically for pest management purposes we mighthave obtained a seriously biased sample of vine-yards for at least the 2 following reasons (1) thesevineyards might be more likely to have experi-enced severe leafhopper problems motivatinggrowers to look for enhanced means of controland (2) growers using prune tree plantings mightbe a relatively progressive subset of all growersand might also differ in other aspects of vineyardmanagement Because 10 of the 12 prune tree siteswere commercial pnme orchards and were notplanted for pest control we feel that the lack oftrue randomization is unlikely to introduce a sys-tematic bias into the analysis
Vineyard A epos Abundance Vineyard plotswere monitored to estimate densities of immigrat-ing A epos and E elegantula during the early sea-son before parasites or leafllOpper adults beganemerging from the 1st generation of reproductionin vineyards For both prune tree and control vine-yard plots 2 yellow sticky traps (75 by 125 mmyellow plastic cards Hilcor Plastics Los Ange]esCAl coated with Tanglefoot were attached to eachof 6 wooden poles (12 traps total per site) Poleswere placed in a transect at 10-m intervals alongthe 3rd vine row from the upwind edge of eachvineyard block Traps were oriented perpendicularto the predominant wind direction and positioned06 and 12 m above the vine canopy to minimizeany influence of vine canopies on trap captureTraps were deployed beginning 1 April (7 pairedsites) 15 Apri] (3 additional pairs) and 1 May (2final pairs) and were replaced twice monthlythrough 15 June 1992 All traps were taken to thelaboratory and examined under a dissection micro-scope to count the number of adult A epos cap-tured
Four vineyard plots were monitored weekly be-ginning 15 April to estimate the beginning of leaf-hopper oviposition and the 1st-generation (Fl)
nymphal emergence The beginning of ovipositionwas determined by examining field-collected grapeleaves under a dissection microscope for the pres-ence of E elegantula eggs The presence ofnymphs was determined by examining grape leavesin the field
Prune Tree Effect The prune tree sites variedwith respect to tree number tree size and orchardmanagement practices These differences in turncould influence overwintering A epos densitiesand cause variation in the treatment effect withinour design Therefore to control for variation inthe pnme refuge effect and to test hypothesesabout the contribution of A epos overwintering inprunes to vineyard abundance we sampled pmnetwigs to estimate overwintering A epos densities(details below) We also recorded the nlllnber oftrees and orchard management practices and es-timated the average rate of tree growth to deter-mine if these factors influence densities of over-wintering A epos (detai]s below)
498 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
The primary difference in management practic-es found among sites was whether or not treeswere irrigated Seven of the 12 sites used in thestudy received little or no supplemental water dur-ing the growing season We classified prune treesites that had a permanent irrigation system as ir-rigated trees Prune trees that were occasionallyprovided supplemental irrigation using portablesystems or that were never irrigated were classi-fied as nonirrigated
Overwintering A epos Densities Anagrus eposemerge from overwintering prune leafhopper eggsoviposited beneath the bark of 1-6-yr-old branchwood (Mulla 1957) Because 2-6-yr-old branchwood can accumulate A epos emergence holesover several years we sampled only 1-yr-oldbranch wood to distinguish the current yearsemergence holes from those of previous yearsPrune twigs were collected in late June and earlyJuly after the completion of A epos emergence Aminimum of 4 pnme twigs (l-yr-old) sampled from10 randomly selected prune trees at each site wasexamined (see twig sampling details described be-low) The length and diameter of twigs were mea-sured to calculate the surface area of 1-yr-oldgrowth (area = 7T d I where d is 1-yr-old twigdiameter I is twig length) Prune twigs were thenexamined under dissection microscopes for thepresence of A epos emergence holes These datawere then used to estimate the average number ofemergence holes per square centimeter of twigsurface area for each refuge site
PrWle Tree Growth Average tree growth var-ied substantially among prune tree sites To eval-uate the possible influence of this factor on A eposdensities we recorded the following 3 parametersas measures of tree growth average length diam-eter and number per tree of 1-yr-old twigs Theaverage length and diameter of twigs were deter-mined by randomly selecting 10 trees within eachpnme tree site for sampling A main branch wasremoved from each compass direction of each treefor a total of 40 branches per site Branches weretaken to the laboratory and the length and diam-eter of each 1-yr-old twig were recorded
The total number of 1-yr-old twigs per tree wasestimated by randomly selecting 4 trees from eachprune tree site for sampling Each tree was dividedin half vertically and horizontally to establish 4quadrants One quadrant position was randomlychosen in each of the 4 trees to count hvigs The4 hvig counts 1 example from each quadrant po-sition were pooled to estimate the mean numberof hvigs per tree
Prwle Tree Site Traps YellowstiCh)traps alsowere used to monitor A epos and E prunicolaadult numbers within pnme trees For each pnmesite 1 trap was placed in 3 randomly selectedprune trees 20-25 m from the ground Trapswere replaced twice monthly as described above
Statistical Procedures The average number ofA epos caught on traps within the vineyard plots
was analyzed using a split-plot analysis of variance(ANOVA) model Growing region was the mainfactor within the model and presence or absenceof a prune tree site was the subfactor Each vine-yard pair was treated as a statistical block to controlfor spatial variability in A epos densities Becausethe number of vineyard pairs sampled increased asthe season progressed separate univariate analyseswere conducted for each sampling period Theanalyseswere conducted to test the null hypothesisthat A epos abundance rates in vineyards are in-dependent of the presence or absence of prunetrees
Prune Tree Effect To test if differences invineyard parasite abundance can be explained bythe emergence of overwintering A epos frompnme trees multiple regression analyses were per-formed using A epos emergence hole densitiesfrom prunes mId A epos capture in control vine-yards as independent variables and A epos capturein prune tree vineyards as the response variableWe interpret A epos trap caphlfe in a control vine-yard as an estimate of the local abundance of par-asites moving into vineyards in the absence ofprune trees (area-wide A epos numbers) We test-ed the null hypothesis that differences in A eposabundance are independent of variation in over-wintering A epos densities within prune trees
Multiplicative Effects We evaluated the poten-tial influence of a vindbreak effect on A eposabundance in vineyards using model II regressionanalyses (Sokal and Rohlf 1981) to examine therelationship between A epos captures in the con-trol vineyards (the independent variable) and Aepos caphlfes in prune tree vineyards (the depen-dent variable) (statistical model prune tree vine-yard captures = 13 control vineyard captures + c+ error) If overwintering A epos within prunetrees were the only factor affecting A epos trapcapture we would expect the slope of the regres-sion equation (13) to approximate 10 with an in-crease in the intercept (c) reflecting an additiveincrease in the number of A epos contributed bypmne trees If on the other hand the slope pa-rameter is significantly greater than 10 a multi-plicative effect in pnme tree vineyard trap captureis present indicating the presence of another fac-tor such as a windbreak effect increasing A eposcapture independent of any contribution fromoverwintering A epos emerging from pnme trees
This analysis was used becanse model I regres-sion produces a biased estimate of the slope forlow values of the correlation coefficient (Sokal andRohlf 1981 Pagel and Harvey 1988) Model IIanalysis is also the most appropriate method foranalyzing the functional relationship between 2variables when both variables are subject to exper-imental error (Sokal and Rohlf 1981) A principalaxis (major axis) model II regression estimates theslope parameter by estimating the principal axisbetween 2 normally distributed random variables(Pagel and Harvey 1988) We used this analysis to
April 1996 MURPHY ET AL HABITAT DIVERSIFICATION FOR BIOLOGICAL CONTHOL 499
Results
Seasonal Patterns Examination of the early-season pattern of parasite and leafhopper trap cap-hires revealed that during the 1st sample period inearly to mid-April 1 or more A epos were caughtin 7 of the 14 vineyard plots monitored (Fig 2)During the 2nd sample period A epos was de-
Fill 2 Seasonal patterns of A epos and E elegantulain vineyard plots (A) A epos adult trap captures (meanplusmn SE) in control and treatment plots (B) E elegantulaadult trap captures (mcan plusmn SE) in control and treatmentplots (e) Period of E elegantrtla oviposition in vineyardplots (D) Pelod of A epos F I emergence (E) Period ofFI nymphal emergence Hatched lines indicate early ap-pearance in some vineyard plots Solid lines indicate ap-pemanle in all plots
tected in 18 of 20 vineyard plots The increasingA epos capture over time indicated that parasitesbegan moving into vineyards in April severalweeks after overwintered E elegantula adults be-gin moving into vineyards Overwintered adultstypically begin moving into vineyards from theiroverwintering habitats during March (Doutt andNakata 1965 Williams 1984) E elegantula eggswere detected in vineyards beginning in late April(Fig 2C) demonstrating that the appearance of Aepos coincided with the beginning of the oviposi-tion period of overwintering leafllOpper adultsThe large increases in A epos capture during the4th sample period in late May coincided with the1st appearance of A epos emergence holes ingrape leaves marking the emergence of adults ofthe Fl parasite generation (Fig 2D) Also duringthe 4th sample period Fl leafhopper nymphs be-gan appearing on grape leaves in most vineyards(Fig2E)
Regression analyses were done to assess the re-lationship between the final density of A cposemergence holes in 1-yr-old prune twigs and trapcapture in pnme trees to estimate the time of Aepos emergence from pnme twigs During the 1stsample period (15-18 April) only a single prunetree site had an A epos capture Regression anal-yses for the 2nd (24 April-l May) and 3rd (8-15May) periods revealed that A epos emergenceholes explained a significant amount of the varia-tion in trap captures (R2 = 088 n = 9 P = 0002and R2 = 042 n = 12 P = 0022 respectively)No relationship was found between area-wide trapcaptures in control vineyards and trap capturewithin pnme trees (R2 = 005 n = 9 P = 056 forthe 2nd sample period R2 = 017 n = 12 P =018 for the 3rd sample period) The conclllsionfrom these analyses is that sticky traps in prunetrees reflect the A epos emergence pattern fromprune trees and are not influenced significantly byarea-wide A epos moving from outside sourcesThe high degree of correlation during the 2ndsample period suggests that peak overwinteringemergence of A epos from prune twigs occllrredbetween 18 April and 1 May
Vineyard A epos Abundance We confined ouranalyses of A epos abundance to the 1st 3 sampledates when A epos captured on traps were mostlikely to have originated from sources outside thevineyard Traps in prune tree vineyards capturedgt2 times the number of parasites as did traps incontrol vineyards during the 2nd and 3rd sampledates (Fig 2A Table 1) No significant differencesin capture were detected between the 2 growingregions but variation in A epos captures was sub-stantial across the paired vineyards as indicated bythe significant block effect
Prune Tree Effect Modell regression analysesrevealed that captures of A epos in pnme treevineyards were closely correlated with captures incontrol vineyards for the 2nd and 3rd sample dates(R2 = 053 n = 10 P = 0016 R2 = 065 n = 12
F Lealltopper eggs
FlA epos emergence11I _
S8-S1S 522-529 613middot610
Date
11I1111I11I1111I11I11I
r3 Control vineyards
o Treatment vineyards
c
D
E
415-18 424middot511
e 2S
B Bbulll 20bull~ ISoSt 10~~ 5-3bull-lt
45
~ 40 A3 35bullis 30bull~ 2S
bullbull 20~ IS
10
ibull-lt
test the null hypothesis that there is no multipli-cative increase in A epos trap capture in prunetree vineyards relative to control vineyard captures
For all statistical analyses except the model IIregressions trap captures were subjected to a log(x+ 1) transformation and A epos emergence holedata were square root-transformed to normalizethe distribution of means Mean comparisons testswere accomplished using single degree of freedomtests (orthogonal contrasts) and all statistical anal-yses except the model II regressions were per-formed using the JMP statistical program for theMacintosh (SAS institute 1989) A Bonferroni cor-rection for multiple comparisons was used to main-tain the total experimentwise a error rate at 005for the ANOVA and regression analyses (Sokal andRohlf 1981)
500 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
Table 1 Nested ANOVA of the number of A epos(101110 trw1sformed) captured on traps in incyards withresl)ect to prune refuges growing regions and pairedvineyard plots (block effcct)
Date Source of df F 1variation
15-18 Apr Treahnent 1 393 0094Blocks (region) 5 617 0023Region 1 013 0735
24 Apr-l May Treatment 1 647 0032Blocks (region) 8 514 0012Region 1 460 0061
8-15 May Treahnent 1 1132 0006Blocks (region) 10 572 0004Begion 1 356 0086
One-tailed tests for hypothesis testing l lt clitical valueafter Bonferroni correction (1 lt 0033)
P = 0001 respectively) but not for the 1st sampledate (R2 = 048 n = 7 P = 009) The significantregressions of A epos capture between the treat-ment and control plots underlines the importanceof blocking by vineyard pair a key feature of ourexperimental design This result also confirms thatarea-wide A epos numbers were a significantsource of parasites in vineyards whether pnmetrees were present or not
The multiple regression analyses indicated thatA epos emergence from prunes was a factor ex-plaining A epos trap captures in prune tree vine-yards after controlling for area-wide abundancefrom control plot trap captures during the 2ndsample period (Table 2) This coincided with theestimated time of A epos emergence from prunetwigs as determined by sticky traps in pnme treesThe partial regression coefficients indicated thatemergence from pnmes explained a substantialamount of the variation in A epos capture duringthe first 2 sample periods in April but by the 3rdsample period in early May area-wide A epos cap-tures explained most variation in trap captures
Variation in Overwintering A epos DensitiesPrune tree sites varied in terms of the number oftrees per site the rate of tree growth the numberof I-yr-old twigs per tree and the density of Aepos emergence holes (Table 3) Much of this vari-
ation was generated by variation in irrigation prac-tices (Table 4) Irrigated trees were associated withgreater tree growth (average surface area of I-yr-old twigs) and lower twig numbers than nonirri-gated trees Irrigation was also associated withhigher A epos overwintering densities as well ashigher trap captures of E prunicola and A eposadults in tree canopies ANCOVA revealed thatboth irrigation (F = 2306 df = 1 12 P = 0001)and prune tree number (F = 1121 df = 1 12 P= 0009) had significant influences on the densityof A epos emergence holes in prune trees Therewas a positive linear relationship between thenumber of prune trees and the density of A eposemergence holes in trees (intercept = -00496slope = 00773) Nevertheless neither pnme treenumber nor irrigation had detectable effects onparasite catch in associated vineyards during the 3sample periods (P gt 0047)
We also estimated the average number of Aepos emergence holes per tree and the total num-ber of emergence holes per prune tree site to testif these parameters could explain captures in ad-jacent vineyards The number of emergence holesper tree was nonsignificant during the 1st sampleperiod (F = 89 df = 1 6 P = 0041) significantduring the 2nd period (F = 721 df = 1 9 P =0031) and non-significant during the 3rd period(F = 026 df = 1 11 P = 0625) The numberper prune tree site was however nonsignificantduring all 3 sample periods (P ~ 0096)
Multiplicative Effects The principal axis anal-yses (model II regression) of A epos trap capturesin control and prune tree vineyards suggest that amultiplicative effect was present during the 2ndand 3rd sample periods when both slope param-eters were significantly gt10 (Fig 3 B and C)During the 1st sample period the estimated slopewas similar to the other sample periods but notsignificantlydifferent from 10 (Fig 3A) These re-sults suggest that independent of any contributionfrom A epos overwintering in pmne trees prunetrees were associated with a multiplicative effectaugmenting A epos capture on the downwind sideof prune trees
Table 2 Multiple regression analysis of the relationship bctwcen the dependent variable 10fllO (A epos trap bullatch)ill prune trec plots and the oulcpendent variable sluarc root (A epos emergence density) from prlUlC sites and10gIO (A epos trap catch) 01 control plots
Sample date Source of variation df Slope (1 SE) F R2 pb
15--18 Apr Whole model test 2 7 119 086 0026Emergence density 17 106 (~ 0325) 106 075 0031Control captu res 17 067 (10237) 80 069 0047
24 Apr-l May Whole model test 210 168 083 0002Emergence density 110 237(1 0717) 109 073 0013Control captures 110 084 (10242) 119 075 OoIl
8-]5 May Whole model test 2 12 1015 069 0005E mergence density 1 19- 091 (10825) 121 035 0299Control captures 112 148 (~ 0363) 1674 081 0003
H2 for individual factors are partial correlation coefficients (r2)b l lt clitical value after Bonferroni correction (1 lt 0017)
April 1996 MURPHY ET AL HABITAT DIVERSIFICATION FOR BIOLOGICAL CONTROL 501
Table 3 Descrilltive larmnelers of pnnle lrees mnong experimenlalllols
Variable
No pnllle trees per siteNo 1-yr-old twigs per treel-yr-old twig surface area cm2
A (--pOS tlllfgtrgencp densitycm2
Mean
422511217
2030021
SE
11191774
4000056
Range
27-1216435-223837-1225
00023-00640
Discllssion
The goal of this research was to assess the influ-ence of pnme trees on early-season A epos abun-dance in vineyards and to test the hypothesizedecological mechanisms underlying this systemPomp trees were associated with significant in-creases in the density of A epos captured in nearbygrape vineyards relative to vineyards lacking prunetrees Pome tree vineyards were found on aver-agp to receive twice the number of early-seasonparasite captures Overwintering A epos emer-gpnce from prune trees was a significant predictorof A cpos trap capture in nearby vineyards duringthe 2nd sample period which coincides with ourestimated period of A epos emergence from prunetwigs Examination of the functional relationshipbetween control and pnme tree vineyard parasitecaptures also revealed a multiplicative response intrap capture on the downwind side of pnme treevineyard plots This suggests that another factorassociated with pnme trees other than overwin-tering A epos populations enhanced parasitenumhtrs in refuge vineyards
Seasonal Patterns Based on limited field dataDoutt and Nakata (1973) and Kido et al (1984)speculated that the effectiveness of overwinteringrefuges near vineyards was at least in part causedby the time of A epos emergence from overwin-tering hosts They based this hypothesis on the ob-senation that parasite emergence from prune treesor blackberries occurs during mid-April at the ap-proximate time overwintered E elegantula beginoviposition in vimmiddotyards They further speculatedthat enhancing early-season A epos colonizerscoupled with the high numerical response of theparasite (A epos completes 3 generations for eachE d(~antllia generation) improves biological con-trol of leafhoppers in vineyards The seasonal pat-tern of leafllOppers and parasites seen in the cur-rent study concurs with these previousobservations Across the replicated vineyard pairsour data show a consistent pattern of both area-
wide A epos captures and A epos emergence fromprune trees coinciding with the onset of oviposi-tion by overwintered E elegantula in vineyardsThus parasite abundance appears to be synchro-nized with the appearance of host eggs in grapevineyards
Vineyard A epos Abundance The replicatedcomparisons of A epos abundance in vineyardsshowed that the presence of overwintering habitatsproduced a substantial early-season increase in thedensity of A epos trapped in grape vineyards aresult consistent with the observations reported byDoutt and Nakata (1973) and Kido et a (1984)The elevated capture rates were seen despite dif-ferences in geographical regions and variation inthe number of area-wide A epos captures whichindicates that the prune tree effect operated undera wide variety of vineyard management and envi-ronmental conditions Based on these results weconcluded that the presence of prune trees wasassociated with elevated A epos numbers and asa result parasites may colonize these vineyards athigher rates than vineyards lacking nearby pnmetrees
Ecological Mechanisms Doutt and Nakata(1973) and Kido et a (1984) assumed that anyincrease in parasite abundance in vineyards (in-ferred using trap data or egg parasitism rates) wasthe direct result of overwintering emergence fromnearby habitats In the current studywe tested thisassumption by comparing the density of overwin-tering A epos emergence holes with A epos trapcapture in pnme tree vineyards after controllingfor area-wide colonizers The significant regressionduring the 2nd sample period indicated prunetrees were a significant source of parasite capturesduring the period of A epos emergence from over-wintered E prunicola eggs These results reveal 2important points regarding the influence of pnmetrees on parasite abundance in vineyards The 1stis that overwintering emergence is a significantecological factor determining early-season parasite
Table 4 Effeet of irrigalion on prIme lree growlh mld populalions of E prunicola mId A epos
Growth rate No twigs A epos A epos tmppecI Elnmicolacm2n (SE) emergence holescm2 trapped
Irrigated 304 498 642 176 00325 00063 950 209 2216+ 5411Nonirrigated 131 420 1475 163 Om05 0058 110 193 059t 507
a Surface area of 1-yr-old pnmc twigs (mean t SE) Numbers of adults caught on traps within prune tree canopies
Signifkantly different 001 lt P lt 005 SigniRcantl) different P lt 001
NoA epos captures in control plots
8060
Vol 25 no 2
402000
00
ENVIRONMENTAL ENTOMOLOGY
0008 00 10 20 30 40 50
B c50 80
bull 40 bull bull 6030
4020
10 20
06
502
lQ-Q~ A~
08bull~=E 06Q= 04tIJ~ 02aQto~l~~~0Z
Fig 3 Scatterplots of the functional relationship between A epos capture in pnme tree refuge vineyardsagainstA epos capture in control vineyards (A) 15-18 April (B = 1705 975 CL L = 058 U = nonestimatable) (8) 24April-l May (B = 238 975 CL L = 205 U = 279) (C) 8--15 May (B = 310 975 CL L = 221 U = 485)Confidence limits were established for 975 as a correction for multiple comparisons
numbers The 2nd is that the effect of prune treeson A epos abundance was dependent not only onthe presence of prune trees themselves but on thedensity of overwintering parasites residing withinthe trees This latter point may have important im-plications for using prune trees as a commercialpest control tactic (discussed below)
Lewis and Stephenson (1966) showed that flyinginsects dispersing in the surrounding environmentaccumulate in sheltered regions near physical bar-riers (natural or artificial windbreaks) They alsofound that insects aggregate behind windbreaks asa multiplicative function of the number of insectsdispersing in the surrounding environment Thiseffect was particularly pronounced for mymaridwasps Using elemental labeling techniques Cor-bett and Rosenheim (in press) found a pattern ofA epos capture suggesting that area-wide A eposnumbers accumulate at a rate 4 times greater im-mediately downwind of prune trees than found up-wind of trees Based on these results they hypoth-esized that a windbreak effect may also be amechanism enhancing abundance within the prunetree-vineyard system We tested this hypothesisacross our replicated study by examining the func-tional relationship between control vineyard trapcaptures (area-wide colonizers in the surroundingenvironment) and prune tree vineyard captures(number of colonizers behind windbreaks) Our re-sults revealed that during each of the 3 sampleperiods the slope of the relationship was 217Thus a multiplicative increase in capture was as-sociated with the presence of prune trees but in-dependent of any contribution from overwinteringA epos densities This result supported the wind-break hypothesis and demonstrated the effect may
be consistent for vineyards with a windbreak pres-ent Furthermore this result identifies an impor-tant additional mechanism that may be affectingA epos vineyard colonization in this system
These results underscore the importance of in-vestigating the ecological mechanisms underlyingsystems of habitat diversification in addition totests of overall effectiveness of diversification Un-derstanding the mechanisms involved will enhanceour ability to manipulate these systems to maxi-mize their effectiveness and provide a usefulframework to begin analyses of new systems
Variation in the Prunc Tree Effect Becauseoverwintering A epos densities varied among theprune tree sites we were interested in how themanagement of pnme trees affects overwinteringA epos populations and their subsequent abun-dance in vineyards Irrigation practices were foundto be a significant factor cplaining much of thevariation in overwintering A epos densities ob-served in pnme trees (Table 4) and somc of thevariation in trap catches in pnme tree vineyardsThe number of prune trees within a site also af-fected overwintering A epos density How treenumber exerts an influence on the density of over-wintering A epos (per square centimeter of pnmebark) is not known but we speculate that greaternumbers of trees may create a more favorable mi-crohabitat increasing the number of A epos thatsuccessfully emerge from their overwinteringhosts
Although tree number had a strong positive in-fluence on the density of A epos emergence holesper square centimeter of prune tree bark it wasnot a predictor of A epos captures in pnme treevineyards This is an enigmatic result At the outset
April IHH6 MUHllIY ET 1 HABITAT DIVEHSIFICATlON FOH BIOLOGICAL CONTHOL 503
of till study we expected that pnllle tree sites con-taining morl trelS would producl a larger pool ofmigrants for thl downwind vineyards The in-enased density (pcr squangt centimeter of bark) ofovcrwintcring A epos in prune trce sites with larg-cr tne 1lI11l11wrsonly strengthened onr expcctationthat trcc numbpr would lw a major determinant ofcady-season abundance in prune trep vineyardsIndclltl wc expccted that we would bl able to es-timatp t]1( minimum numbPI of trees needed togenlrate the gtelof augmentation of A cpos nec-pssary to achieve pest management Instead whatwe found is that trpe number is less a predictor ofalmmlancp in vinpyards than is the number of AlJOS produced ppr centinIPter of bark We also ca]-cu]ahd thc total number of A epos emerged fromovclvintlring sitps vithin an cntire prune tree site(a composite cstimate obtained as the product ofA e]1os ltknsity pcr squarc ccntimetcr of bark thetotal arca plr tngtp and the number of trpes persitc) as t]IP estimatp for A pos contributions fromprunl trpc sites This estimatc was however awcakcr prcdictor of early-season abundance invincyards than Pitill l thc numbcr of A cpos persquarc ccntimctcr of bark or pcr prune tree Howcan we cxplain this resn]t One possibility is thattrees within the largcr prune trcc sites Illay havecontributed uncIlm]]y to the number of vineyardmigrants vith thc rows of trccs directly adjacentto the vincyard producing the bulk of the waspstrapped in vincyards Additional experimentationwill be needed to test this and other potential ex-plauations for the lack of a trec number effect
These results have 2 implications for establish-ing and managing pnnll tree rcfuges for pest con-trol The 1st is that cnhanccd A cpos abundancemay he attainable with a relatively small numberof tnps TIIP 2nd is that irrigation of pnllle treesvill bp of primary importance for supporting highdpnsitics of ovprwintcring i cpos populationsMon detail(gtd rpcommcndations for establishingpnme trcc rcfugps will be presented in future pa-pers
Questious about thc pffect of vegetational diver-sity on tIll diversity and abundance of insect pop-ulations ngtcenty have rcceived considerable atteu-tion from ecologists and pest control specialistsThe rpsu]ts of thp cnrrent study not only supportedtl( hypothdical effect of pnme tree refuges on Acpos abundance but in a broader context providea test of the effect of wgctational diversity on nat-ural encmy abundance As a result this study hassatisfied tIl( 1st of 3 hypothesized natural enemyresponses which pr((licts greater natura] enemyabundance in diversified systems (Bussell 1989Andow 1991) In addition 011study identified thefollOving 2 mpchanisms underlying the influenceof prune trees the presence of an ove1vinteringhost and a windbreak effpct
Future papers vill examine the effect of over-wintering refuges on the remaining 2 hypotheses-the effect of pnnw trees on E elegalltula egg mor-
taity and E elcgalltula population densities withingrape vineyards
Acknowledgments
We extend special thanks to Nerissa Fretman MaddySamant John Tlnnter Jason May and Paul Higgs (De-partment of Entomology University of Califol1lia Davis)for field collections and sample processing Vc also ac-Irnowledge thc assistancc of Hobert Hoy Stan Grant andBrad Coleman (E amp J Gallo Winery Livingston CAl forlogistical and material support This research was fuudedby the Westem Hegional Integrated Pest Managementprogram the California Department of Ptsticidt Hegu-lation and the Locli-Woodbridge vVinegrapt Commis-sion This research was part of BGMs doctoral disser-tation at the University of California at Davis
References Cited
Altieri M A 1992 Agroecological foundatious of al-temative agliculture in Califomia Agric Ecosyst En-viron 39 23-53
Andow D A 1991 Vegetational diversity and artll1o-pod population response Annu Hev Entomol 36561-586
Corbett A und J A Rosenheim 1996 Impact of anatural enemy overwintering refuge and its iuterac-tion vith the surrounding landscape Ecol Entomol(in press)
Doutt R L and J Nakat bullbull 1965 Overwintering ref-uge of Anagrus epos (Hymenoptera Mymaridae) JEcon Entomol 58 586
1973 The Ruhus leafllOpper and its egg parasitoid Anendemic biotic system useful in grape-pest manage-ment Environ Entomol 2 381~186
Flaherty D L L T Wilson V M Stern and IIKido 1985 Biologicalcontrol in San Joaquin Valleyvineyards pp 501-520 Tn D G Herzog and M AHoy [eds] Biological control of agricnltural IPM sys-tems Viley New York
Flint M L and P A Roberts 1988 Using cropdiversity to manage pest problems some Californiaexamples Am J Altern Agric1 163-167
Gorclh G lIml D M Dunlmr 1977 A new Allagmsimportant in the biological control of Stephanitis tak-eljai and a key to the N01ih American species FlaEntomol 60 85-95
Herzo~ D c and J E Funderburk 1985 Plantresistance and cultural practiee interactious with hio-logical control pp 67-88 Til D G Ilerzog and MA Hoy [eds] Biological control of agliclllInral IPMsystems Wiley New York
Kiclo H D L Flaherty D F 80sh Dlul K A Val-ero 1984 French pmne trees as ovclwiuteringsites for the grape leafllOpper egg parasite Am JEnol Vitie 35 156-160
Letourneau D K 1987 The enemies hypothesis Tri-trophic interactions and vegetational diversity in trop-ical agroecosystel11sEcology 68 1616-1622
Lewis T bullbulllid J W Stephenson 1966 The penne-ability of artificial windbreaks and the distribution offlying insects in the leeward shelttred Will AnnAppl BioI 58 355-363
Mulla M S 1957 The biology of Typhlo(fhapnmi-cola (Edwards) and 1 quercus (Fabricius) (Cicadelli-dae lIomoptera) Ann Entomol SOlAm 50 76-87
504 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
National Research Council 1989 Alternative agri-culture National AcademyWashington DC
PaAel M D and P H Harvey 1988 Recent de-velopments in the analysis of comparative data QRev BioI 63 418-440
Pickett C H L T Wilson and D L Flaherty1990 The role of refuges in crop protection withreference to plantings of French prune trees in agrape agroecosystem pp 151-165 In N J BostanianL T Wilson and T J Dennehy [eds] Monitoringand integrated management of arthropod pests ofsmall fruit crops Intercept Andover
RusseD E P 1989 Enemies hypothesis A review ofthe effect of vegetational diversity on predatory in-sects and parasitoids Environ Entomol 18 590-599
Sokal R R and F J Rohlf 1981 Biometry 2nded Freeman San Francisco
SAS Institute 1989 JMP statistical program SAS In-stitute Cary NC
van Emden H F 1990 Plant diversity and naturalenemy efficiency in agroecosystems pp 63-80In MMackauer L E Ehler and J Roland [eds] Criticalissues in biological control Intercept Andover UK
Williams D W 1984 Ecology of the blackberry-leaf-hopper-parasite system and its relevance to Californiagrape agroecosystems Hilgardia 52 1--33
Wilson L T C H Pickett and D L Flaherty1989 French prune trees refuge for grape leafhop-per parasite Calif Agric 43(2) 7-8
Winkler A J J A Cook W M Kliewer and L ALider 1974 Climate and soils pp 58-75 In Gen-eral viticulture University of California Press Berke-ley
Received for publication 5 December 1994 accepted 6November 1995

496 ENVIRONMENTAL ENTOMOLOCY Vol 25 no 2
(Doutt and Nakata 1973) A major h1ctor hypoth-esized to be associated with the abundance andeffectiveness of A epos is the presence of nearbyhabitat that supports alternate leafhopper hosts(Doutt and Nakata 1965 1973) A epos overwin-ters within leafhopper eggs E elegantula overwin-ters as an adult thus an alternate source of leaf-hopper eggs is required to support overwinteringA epos numbers
Doutt and Nakata (1965) first observed thatvineyards located downwind from riparian habitatshad higher levels of E elegantula egg parasitismand lower E elegantula numbers relative to vine-yards located distant from these habitats Theyfound that blackberry brambles Rubus spp abun-dant in riparian habitats support Dikrella eaZifor-niea (Lawson) a year-round host of A epos Theyobserved that early spring emergence of A eposfrom blackberry results in earlier vineyard coloni-zation by parasites which coincided with oviposi-tion of overwintered E elegantula They also spec-ulated that earlier vineyard colonization at higherdensities produced a stronger numerical responseand thereby improved biological control (Douttand Nakata 1973) Unfortunately field trials deter-mined blackberry brambles grown away from ri-parian habitats did not maintain abundant popu-lations of A epos and further attempts wereabandoned (Flal1erty et al 1985)
Kido et at (1984) demonstrated that prune treessupport the prune leafhopper Edwardsiana prun-ieola (Edwards) which overwinters in the eggstage and serves as an overwintering host for Aepos Emergence of overwintering A epos fromoverwintering E prunieola eggs laid beneath thebark of prune twigs also was found to coincide with1st-generation E elegantula oviposition in vine-yards suggesting that pnme trees also serve as anoverwintering habitat for A epos Studies in bothcommercial and experimental plantings of pnilletrees demonstrated that A epos populations couldbe supported year-round Williams (1984) andKido et al (1984) also showed that A epos rearedfrom E prunieola or D ealifomiea can parasitizeE elegant1l1aeggs successfully and vice versa Fi-nally it has been shown that elevated A epos pop-ulations were associated with higher E elegant1l1aegg parasitism in vineyards near pnme trees (Kidoet al 1984 Pickett et aI 1990) Although thesestudies established the basic framework of thegrape leafhopper-A epos interaction there havebeen no replicated field trials that assess the effi-cacy of prune tree habitats or that test the ecolog-ical mechanisms underlying the effect
Anagrus epos Abundance Doutt and Nakata(1973) and Kido et al (1984) hypothesized thatoverwintering refuges near vineyards increase ear-ly-season A epos vineyard colonization as a resultof overwintering A epos emerging from refugesand dispersing into nearby vineyards They testedthese predictions in field trials using yellow stickytraps to measure early-season abundance of adult
A epos in vineyards or by examining E elegantulaeggs and comparing rates of egg parasitism Col-onization appeared to be enhanced in vineyardsdownwind from prune orchards or riparian habi-tats containing blackberry brambles They con-cluded that the greater abundance seen in fieldtrials supported the overwintering refuge hypoth-esis However because these initial experimentsused unreplicated comparisons the results al-though consistent with the refuge hypothesis werepreliminary in nature
The interpretation presented by Doutt and Nak-ata (1973) and Kido et a1(1984) also rested on anuntested assumption namely that an increase inparasite captures or egg parasitism near refugeswas the direct result of A epos emergence fromoverwintering sites A 2nd potential mechanismthat could explain increased parasite capturesdownwind of prune trees or trees found in riparianhabitats is a windbreak effect generated by thephysical structure of these habitats A windbreakeffect occurs when a structural barrier creates hlr-bulence in the airflow and a sheltered zone on theleeward side of the barrier The turbulent airflowmay act to increase the probability that a dispers-ing insect will settle into these sheltered zonesthereby enhancing colonization Dispersing insectsfrom many families including wasps in the familyMymaridae have been shown to accumulate be-hind windbreaks (Lewis and Stephenson 1966)Wilson et a1 (1989) showed that A epos reachhigher densities in pnmes when artificial wind-breaks are positioned on the upwind side of prunetrees Corbett and Rosenheim (in press) have alsorecently demonstrated a pattern of A epos cap-tures in vineyards downwind of pnme trees that isconsistent with the operation of a windbreak Thusenhanced A epos abundance could be the resultof overwintering populations within the prunetrees a windbreak effect or some combination ofthose 2 factors
In the current shldy we tested 2 key hypotheseswith respect to the effect of overwintering sites onA epos abundance in vineyards as follows (1) thatprune trees adjacent to vineyards enhance early-season A epos numbers in vineyards and (2) thatdifferences in abundance are the result of differ-ences in the densities of overwintering A eposemerging from pnme trees In addition to those 2hypotheses we also analyzed capture data for ev-idence of a windbreak effect influencing A eposabundance downwind of pnme trees
Materials and Methods
Experimental Design A epos colonizes virtu-ally all grape vineyards at some point during thegrowing season the time and density of abundancecan however be highly variable E elegantulapopulations also can be variable among vineyardsites in a given year Because of the vmiation ofboth parasite and leafhopper numbers a rigorous
April 1996 MURPHY ET AL HABITAT DIVERSIFICATION FOR BIOLOGICAL CONTHOL 497
Fil( 1 Pairedvineyardplot locationsin northernandctntra CaliforniaThe explodedview showsthe orien-tationof pnme tree and controlvineyardplots
evaluation of prune tree-enhanced biological con-trol required a large number of replicates Surveyswere conducted throughout the major wine grapeviticultural regions of central and northern Cali-fornia to locate 26 paired commercia] vineyardsone with adjacent upwind pnme trees and the oth-er lacking pnme trees (Fig 1) Paired vineyardswere matched for grape cultivar trellising anclmanagement practices Control (nonprune tree)vinlyards were positioned O~O kilometers fromrefuge sites and either parallel to or upwind ofpnll1e tree vineyard sites to minimize movementof ]eafllOppersor parasites between treatment andcontrol vineyards The paired plots were groupedinto 2 viticultural growing regions defined by thecUllmlative degree days occurring between Apriland October (Winkler et al 1974) Comparisonswere made between cooler growing regionslaquo3000 DD) and warmer regions (gt3000 DD)Two of the pnme tree sites used in the study wereplanted specifically for leafhopper control the re-mainder were commercial or remnant prune or-chards growing adjacent to the vineyard plots
Given the nature of the pnme tree habitat beingtested we were unable to ensure tnle randomiza-tion within our experimental design True random-ization would have required us to assign prune treeplantings randomly to plots and would have re-quired a delay of 3-5 yr for the pnme trees tobecome established and support large populationsof the alternate leafhopper host As a result weused vineyard sites located near previously estab-lished prune trees Had we chosen to work pri-
marily in vineyards with prune trees planted spe-cifically for pest management purposes we mighthave obtained a seriously biased sample of vine-yards for at least the 2 following reasons (1) thesevineyards might be more likely to have experi-enced severe leafhopper problems motivatinggrowers to look for enhanced means of controland (2) growers using prune tree plantings mightbe a relatively progressive subset of all growersand might also differ in other aspects of vineyardmanagement Because 10 of the 12 prune tree siteswere commercial pnme orchards and were notplanted for pest control we feel that the lack oftrue randomization is unlikely to introduce a sys-tematic bias into the analysis
Vineyard A epos Abundance Vineyard plotswere monitored to estimate densities of immigrat-ing A epos and E elegantula during the early sea-son before parasites or leafllOpper adults beganemerging from the 1st generation of reproductionin vineyards For both prune tree and control vine-yard plots 2 yellow sticky traps (75 by 125 mmyellow plastic cards Hilcor Plastics Los Ange]esCAl coated with Tanglefoot were attached to eachof 6 wooden poles (12 traps total per site) Poleswere placed in a transect at 10-m intervals alongthe 3rd vine row from the upwind edge of eachvineyard block Traps were oriented perpendicularto the predominant wind direction and positioned06 and 12 m above the vine canopy to minimizeany influence of vine canopies on trap captureTraps were deployed beginning 1 April (7 pairedsites) 15 Apri] (3 additional pairs) and 1 May (2final pairs) and were replaced twice monthlythrough 15 June 1992 All traps were taken to thelaboratory and examined under a dissection micro-scope to count the number of adult A epos cap-tured
Four vineyard plots were monitored weekly be-ginning 15 April to estimate the beginning of leaf-hopper oviposition and the 1st-generation (Fl)
nymphal emergence The beginning of ovipositionwas determined by examining field-collected grapeleaves under a dissection microscope for the pres-ence of E elegantula eggs The presence ofnymphs was determined by examining grape leavesin the field
Prune Tree Effect The prune tree sites variedwith respect to tree number tree size and orchardmanagement practices These differences in turncould influence overwintering A epos densitiesand cause variation in the treatment effect withinour design Therefore to control for variation inthe pnme refuge effect and to test hypothesesabout the contribution of A epos overwintering inprunes to vineyard abundance we sampled pmnetwigs to estimate overwintering A epos densities(details below) We also recorded the nlllnber oftrees and orchard management practices and es-timated the average rate of tree growth to deter-mine if these factors influence densities of over-wintering A epos (detai]s below)
498 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
The primary difference in management practic-es found among sites was whether or not treeswere irrigated Seven of the 12 sites used in thestudy received little or no supplemental water dur-ing the growing season We classified prune treesites that had a permanent irrigation system as ir-rigated trees Prune trees that were occasionallyprovided supplemental irrigation using portablesystems or that were never irrigated were classi-fied as nonirrigated
Overwintering A epos Densities Anagrus eposemerge from overwintering prune leafhopper eggsoviposited beneath the bark of 1-6-yr-old branchwood (Mulla 1957) Because 2-6-yr-old branchwood can accumulate A epos emergence holesover several years we sampled only 1-yr-oldbranch wood to distinguish the current yearsemergence holes from those of previous yearsPrune twigs were collected in late June and earlyJuly after the completion of A epos emergence Aminimum of 4 pnme twigs (l-yr-old) sampled from10 randomly selected prune trees at each site wasexamined (see twig sampling details described be-low) The length and diameter of twigs were mea-sured to calculate the surface area of 1-yr-oldgrowth (area = 7T d I where d is 1-yr-old twigdiameter I is twig length) Prune twigs were thenexamined under dissection microscopes for thepresence of A epos emergence holes These datawere then used to estimate the average number ofemergence holes per square centimeter of twigsurface area for each refuge site
PrWle Tree Growth Average tree growth var-ied substantially among prune tree sites To eval-uate the possible influence of this factor on A eposdensities we recorded the following 3 parametersas measures of tree growth average length diam-eter and number per tree of 1-yr-old twigs Theaverage length and diameter of twigs were deter-mined by randomly selecting 10 trees within eachpnme tree site for sampling A main branch wasremoved from each compass direction of each treefor a total of 40 branches per site Branches weretaken to the laboratory and the length and diam-eter of each 1-yr-old twig were recorded
The total number of 1-yr-old twigs per tree wasestimated by randomly selecting 4 trees from eachprune tree site for sampling Each tree was dividedin half vertically and horizontally to establish 4quadrants One quadrant position was randomlychosen in each of the 4 trees to count hvigs The4 hvig counts 1 example from each quadrant po-sition were pooled to estimate the mean numberof hvigs per tree
Prwle Tree Site Traps YellowstiCh)traps alsowere used to monitor A epos and E prunicolaadult numbers within pnme trees For each pnmesite 1 trap was placed in 3 randomly selectedprune trees 20-25 m from the ground Trapswere replaced twice monthly as described above
Statistical Procedures The average number ofA epos caught on traps within the vineyard plots
was analyzed using a split-plot analysis of variance(ANOVA) model Growing region was the mainfactor within the model and presence or absenceof a prune tree site was the subfactor Each vine-yard pair was treated as a statistical block to controlfor spatial variability in A epos densities Becausethe number of vineyard pairs sampled increased asthe season progressed separate univariate analyseswere conducted for each sampling period Theanalyseswere conducted to test the null hypothesisthat A epos abundance rates in vineyards are in-dependent of the presence or absence of prunetrees
Prune Tree Effect To test if differences invineyard parasite abundance can be explained bythe emergence of overwintering A epos frompnme trees multiple regression analyses were per-formed using A epos emergence hole densitiesfrom prunes mId A epos capture in control vine-yards as independent variables and A epos capturein prune tree vineyards as the response variableWe interpret A epos trap caphlfe in a control vine-yard as an estimate of the local abundance of par-asites moving into vineyards in the absence ofprune trees (area-wide A epos numbers) We test-ed the null hypothesis that differences in A eposabundance are independent of variation in over-wintering A epos densities within prune trees
Multiplicative Effects We evaluated the poten-tial influence of a vindbreak effect on A eposabundance in vineyards using model II regressionanalyses (Sokal and Rohlf 1981) to examine therelationship between A epos captures in the con-trol vineyards (the independent variable) and Aepos caphlfes in prune tree vineyards (the depen-dent variable) (statistical model prune tree vine-yard captures = 13 control vineyard captures + c+ error) If overwintering A epos within prunetrees were the only factor affecting A epos trapcapture we would expect the slope of the regres-sion equation (13) to approximate 10 with an in-crease in the intercept (c) reflecting an additiveincrease in the number of A epos contributed bypmne trees If on the other hand the slope pa-rameter is significantly greater than 10 a multi-plicative effect in pnme tree vineyard trap captureis present indicating the presence of another fac-tor such as a windbreak effect increasing A eposcapture independent of any contribution fromoverwintering A epos emerging from pnme trees
This analysis was used becanse model I regres-sion produces a biased estimate of the slope forlow values of the correlation coefficient (Sokal andRohlf 1981 Pagel and Harvey 1988) Model IIanalysis is also the most appropriate method foranalyzing the functional relationship between 2variables when both variables are subject to exper-imental error (Sokal and Rohlf 1981) A principalaxis (major axis) model II regression estimates theslope parameter by estimating the principal axisbetween 2 normally distributed random variables(Pagel and Harvey 1988) We used this analysis to
April 1996 MURPHY ET AL HABITAT DIVERSIFICATION FOR BIOLOGICAL CONTHOL 499
Results
Seasonal Patterns Examination of the early-season pattern of parasite and leafhopper trap cap-hires revealed that during the 1st sample period inearly to mid-April 1 or more A epos were caughtin 7 of the 14 vineyard plots monitored (Fig 2)During the 2nd sample period A epos was de-
Fill 2 Seasonal patterns of A epos and E elegantulain vineyard plots (A) A epos adult trap captures (meanplusmn SE) in control and treatment plots (B) E elegantulaadult trap captures (mcan plusmn SE) in control and treatmentplots (e) Period of E elegantrtla oviposition in vineyardplots (D) Pelod of A epos F I emergence (E) Period ofFI nymphal emergence Hatched lines indicate early ap-pearance in some vineyard plots Solid lines indicate ap-pemanle in all plots
tected in 18 of 20 vineyard plots The increasingA epos capture over time indicated that parasitesbegan moving into vineyards in April severalweeks after overwintered E elegantula adults be-gin moving into vineyards Overwintered adultstypically begin moving into vineyards from theiroverwintering habitats during March (Doutt andNakata 1965 Williams 1984) E elegantula eggswere detected in vineyards beginning in late April(Fig 2C) demonstrating that the appearance of Aepos coincided with the beginning of the oviposi-tion period of overwintering leafllOpper adultsThe large increases in A epos capture during the4th sample period in late May coincided with the1st appearance of A epos emergence holes ingrape leaves marking the emergence of adults ofthe Fl parasite generation (Fig 2D) Also duringthe 4th sample period Fl leafhopper nymphs be-gan appearing on grape leaves in most vineyards(Fig2E)
Regression analyses were done to assess the re-lationship between the final density of A cposemergence holes in 1-yr-old prune twigs and trapcapture in pnme trees to estimate the time of Aepos emergence from pnme twigs During the 1stsample period (15-18 April) only a single prunetree site had an A epos capture Regression anal-yses for the 2nd (24 April-l May) and 3rd (8-15May) periods revealed that A epos emergenceholes explained a significant amount of the varia-tion in trap captures (R2 = 088 n = 9 P = 0002and R2 = 042 n = 12 P = 0022 respectively)No relationship was found between area-wide trapcaptures in control vineyards and trap capturewithin pnme trees (R2 = 005 n = 9 P = 056 forthe 2nd sample period R2 = 017 n = 12 P =018 for the 3rd sample period) The conclllsionfrom these analyses is that sticky traps in prunetrees reflect the A epos emergence pattern fromprune trees and are not influenced significantly byarea-wide A epos moving from outside sourcesThe high degree of correlation during the 2ndsample period suggests that peak overwinteringemergence of A epos from prune twigs occllrredbetween 18 April and 1 May
Vineyard A epos Abundance We confined ouranalyses of A epos abundance to the 1st 3 sampledates when A epos captured on traps were mostlikely to have originated from sources outside thevineyard Traps in prune tree vineyards capturedgt2 times the number of parasites as did traps incontrol vineyards during the 2nd and 3rd sampledates (Fig 2A Table 1) No significant differencesin capture were detected between the 2 growingregions but variation in A epos captures was sub-stantial across the paired vineyards as indicated bythe significant block effect
Prune Tree Effect Modell regression analysesrevealed that captures of A epos in pnme treevineyards were closely correlated with captures incontrol vineyards for the 2nd and 3rd sample dates(R2 = 053 n = 10 P = 0016 R2 = 065 n = 12
F Lealltopper eggs
FlA epos emergence11I _
S8-S1S 522-529 613middot610
Date
11I1111I11I1111I11I11I
r3 Control vineyards
o Treatment vineyards
c
D
E
415-18 424middot511
e 2S
B Bbulll 20bull~ ISoSt 10~~ 5-3bull-lt
45
~ 40 A3 35bullis 30bull~ 2S
bullbull 20~ IS
10
ibull-lt
test the null hypothesis that there is no multipli-cative increase in A epos trap capture in prunetree vineyards relative to control vineyard captures
For all statistical analyses except the model IIregressions trap captures were subjected to a log(x+ 1) transformation and A epos emergence holedata were square root-transformed to normalizethe distribution of means Mean comparisons testswere accomplished using single degree of freedomtests (orthogonal contrasts) and all statistical anal-yses except the model II regressions were per-formed using the JMP statistical program for theMacintosh (SAS institute 1989) A Bonferroni cor-rection for multiple comparisons was used to main-tain the total experimentwise a error rate at 005for the ANOVA and regression analyses (Sokal andRohlf 1981)
500 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
Table 1 Nested ANOVA of the number of A epos(101110 trw1sformed) captured on traps in incyards withresl)ect to prune refuges growing regions and pairedvineyard plots (block effcct)
Date Source of df F 1variation
15-18 Apr Treahnent 1 393 0094Blocks (region) 5 617 0023Region 1 013 0735
24 Apr-l May Treatment 1 647 0032Blocks (region) 8 514 0012Region 1 460 0061
8-15 May Treahnent 1 1132 0006Blocks (region) 10 572 0004Begion 1 356 0086
One-tailed tests for hypothesis testing l lt clitical valueafter Bonferroni correction (1 lt 0033)
P = 0001 respectively) but not for the 1st sampledate (R2 = 048 n = 7 P = 009) The significantregressions of A epos capture between the treat-ment and control plots underlines the importanceof blocking by vineyard pair a key feature of ourexperimental design This result also confirms thatarea-wide A epos numbers were a significantsource of parasites in vineyards whether pnmetrees were present or not
The multiple regression analyses indicated thatA epos emergence from prunes was a factor ex-plaining A epos trap captures in prune tree vine-yards after controlling for area-wide abundancefrom control plot trap captures during the 2ndsample period (Table 2) This coincided with theestimated time of A epos emergence from prunetwigs as determined by sticky traps in pnme treesThe partial regression coefficients indicated thatemergence from pnmes explained a substantialamount of the variation in A epos capture duringthe first 2 sample periods in April but by the 3rdsample period in early May area-wide A epos cap-tures explained most variation in trap captures
Variation in Overwintering A epos DensitiesPrune tree sites varied in terms of the number oftrees per site the rate of tree growth the numberof I-yr-old twigs per tree and the density of Aepos emergence holes (Table 3) Much of this vari-
ation was generated by variation in irrigation prac-tices (Table 4) Irrigated trees were associated withgreater tree growth (average surface area of I-yr-old twigs) and lower twig numbers than nonirri-gated trees Irrigation was also associated withhigher A epos overwintering densities as well ashigher trap captures of E prunicola and A eposadults in tree canopies ANCOVA revealed thatboth irrigation (F = 2306 df = 1 12 P = 0001)and prune tree number (F = 1121 df = 1 12 P= 0009) had significant influences on the densityof A epos emergence holes in prune trees Therewas a positive linear relationship between thenumber of prune trees and the density of A eposemergence holes in trees (intercept = -00496slope = 00773) Nevertheless neither pnme treenumber nor irrigation had detectable effects onparasite catch in associated vineyards during the 3sample periods (P gt 0047)
We also estimated the average number of Aepos emergence holes per tree and the total num-ber of emergence holes per prune tree site to testif these parameters could explain captures in ad-jacent vineyards The number of emergence holesper tree was nonsignificant during the 1st sampleperiod (F = 89 df = 1 6 P = 0041) significantduring the 2nd period (F = 721 df = 1 9 P =0031) and non-significant during the 3rd period(F = 026 df = 1 11 P = 0625) The numberper prune tree site was however nonsignificantduring all 3 sample periods (P ~ 0096)
Multiplicative Effects The principal axis anal-yses (model II regression) of A epos trap capturesin control and prune tree vineyards suggest that amultiplicative effect was present during the 2ndand 3rd sample periods when both slope param-eters were significantly gt10 (Fig 3 B and C)During the 1st sample period the estimated slopewas similar to the other sample periods but notsignificantlydifferent from 10 (Fig 3A) These re-sults suggest that independent of any contributionfrom A epos overwintering in pmne trees prunetrees were associated with a multiplicative effectaugmenting A epos capture on the downwind sideof prune trees
Table 2 Multiple regression analysis of the relationship bctwcen the dependent variable 10fllO (A epos trap bullatch)ill prune trec plots and the oulcpendent variable sluarc root (A epos emergence density) from prlUlC sites and10gIO (A epos trap catch) 01 control plots
Sample date Source of variation df Slope (1 SE) F R2 pb
15--18 Apr Whole model test 2 7 119 086 0026Emergence density 17 106 (~ 0325) 106 075 0031Control captu res 17 067 (10237) 80 069 0047
24 Apr-l May Whole model test 210 168 083 0002Emergence density 110 237(1 0717) 109 073 0013Control captures 110 084 (10242) 119 075 OoIl
8-]5 May Whole model test 2 12 1015 069 0005E mergence density 1 19- 091 (10825) 121 035 0299Control captures 112 148 (~ 0363) 1674 081 0003
H2 for individual factors are partial correlation coefficients (r2)b l lt clitical value after Bonferroni correction (1 lt 0017)
April 1996 MURPHY ET AL HABITAT DIVERSIFICATION FOR BIOLOGICAL CONTROL 501
Table 3 Descrilltive larmnelers of pnnle lrees mnong experimenlalllols
Variable
No pnllle trees per siteNo 1-yr-old twigs per treel-yr-old twig surface area cm2
A (--pOS tlllfgtrgencp densitycm2
Mean
422511217
2030021
SE
11191774
4000056
Range
27-1216435-223837-1225
00023-00640
Discllssion
The goal of this research was to assess the influ-ence of pnme trees on early-season A epos abun-dance in vineyards and to test the hypothesizedecological mechanisms underlying this systemPomp trees were associated with significant in-creases in the density of A epos captured in nearbygrape vineyards relative to vineyards lacking prunetrees Pome tree vineyards were found on aver-agp to receive twice the number of early-seasonparasite captures Overwintering A epos emer-gpnce from prune trees was a significant predictorof A cpos trap capture in nearby vineyards duringthe 2nd sample period which coincides with ourestimated period of A epos emergence from prunetwigs Examination of the functional relationshipbetween control and pnme tree vineyard parasitecaptures also revealed a multiplicative response intrap capture on the downwind side of pnme treevineyard plots This suggests that another factorassociated with pnme trees other than overwin-tering A epos populations enhanced parasitenumhtrs in refuge vineyards
Seasonal Patterns Based on limited field dataDoutt and Nakata (1973) and Kido et al (1984)speculated that the effectiveness of overwinteringrefuges near vineyards was at least in part causedby the time of A epos emergence from overwin-tering hosts They based this hypothesis on the ob-senation that parasite emergence from prune treesor blackberries occurs during mid-April at the ap-proximate time overwintered E elegantula beginoviposition in vimmiddotyards They further speculatedthat enhancing early-season A epos colonizerscoupled with the high numerical response of theparasite (A epos completes 3 generations for eachE d(~antllia generation) improves biological con-trol of leafhoppers in vineyards The seasonal pat-tern of leafllOppers and parasites seen in the cur-rent study concurs with these previousobservations Across the replicated vineyard pairsour data show a consistent pattern of both area-
wide A epos captures and A epos emergence fromprune trees coinciding with the onset of oviposi-tion by overwintered E elegantula in vineyardsThus parasite abundance appears to be synchro-nized with the appearance of host eggs in grapevineyards
Vineyard A epos Abundance The replicatedcomparisons of A epos abundance in vineyardsshowed that the presence of overwintering habitatsproduced a substantial early-season increase in thedensity of A epos trapped in grape vineyards aresult consistent with the observations reported byDoutt and Nakata (1973) and Kido et a (1984)The elevated capture rates were seen despite dif-ferences in geographical regions and variation inthe number of area-wide A epos captures whichindicates that the prune tree effect operated undera wide variety of vineyard management and envi-ronmental conditions Based on these results weconcluded that the presence of prune trees wasassociated with elevated A epos numbers and asa result parasites may colonize these vineyards athigher rates than vineyards lacking nearby pnmetrees
Ecological Mechanisms Doutt and Nakata(1973) and Kido et a (1984) assumed that anyincrease in parasite abundance in vineyards (in-ferred using trap data or egg parasitism rates) wasthe direct result of overwintering emergence fromnearby habitats In the current studywe tested thisassumption by comparing the density of overwin-tering A epos emergence holes with A epos trapcapture in pnme tree vineyards after controllingfor area-wide colonizers The significant regressionduring the 2nd sample period indicated prunetrees were a significant source of parasite capturesduring the period of A epos emergence from over-wintered E prunicola eggs These results reveal 2important points regarding the influence of pnmetrees on parasite abundance in vineyards The 1stis that overwintering emergence is a significantecological factor determining early-season parasite
Table 4 Effeet of irrigalion on prIme lree growlh mld populalions of E prunicola mId A epos
Growth rate No twigs A epos A epos tmppecI Elnmicolacm2n (SE) emergence holescm2 trapped
Irrigated 304 498 642 176 00325 00063 950 209 2216+ 5411Nonirrigated 131 420 1475 163 Om05 0058 110 193 059t 507
a Surface area of 1-yr-old pnmc twigs (mean t SE) Numbers of adults caught on traps within prune tree canopies
Signifkantly different 001 lt P lt 005 SigniRcantl) different P lt 001
NoA epos captures in control plots
8060
Vol 25 no 2
402000
00
ENVIRONMENTAL ENTOMOLOGY
0008 00 10 20 30 40 50
B c50 80
bull 40 bull bull 6030
4020
10 20
06
502
lQ-Q~ A~
08bull~=E 06Q= 04tIJ~ 02aQto~l~~~0Z
Fig 3 Scatterplots of the functional relationship between A epos capture in pnme tree refuge vineyardsagainstA epos capture in control vineyards (A) 15-18 April (B = 1705 975 CL L = 058 U = nonestimatable) (8) 24April-l May (B = 238 975 CL L = 205 U = 279) (C) 8--15 May (B = 310 975 CL L = 221 U = 485)Confidence limits were established for 975 as a correction for multiple comparisons
numbers The 2nd is that the effect of prune treeson A epos abundance was dependent not only onthe presence of prune trees themselves but on thedensity of overwintering parasites residing withinthe trees This latter point may have important im-plications for using prune trees as a commercialpest control tactic (discussed below)
Lewis and Stephenson (1966) showed that flyinginsects dispersing in the surrounding environmentaccumulate in sheltered regions near physical bar-riers (natural or artificial windbreaks) They alsofound that insects aggregate behind windbreaks asa multiplicative function of the number of insectsdispersing in the surrounding environment Thiseffect was particularly pronounced for mymaridwasps Using elemental labeling techniques Cor-bett and Rosenheim (in press) found a pattern ofA epos capture suggesting that area-wide A eposnumbers accumulate at a rate 4 times greater im-mediately downwind of prune trees than found up-wind of trees Based on these results they hypoth-esized that a windbreak effect may also be amechanism enhancing abundance within the prunetree-vineyard system We tested this hypothesisacross our replicated study by examining the func-tional relationship between control vineyard trapcaptures (area-wide colonizers in the surroundingenvironment) and prune tree vineyard captures(number of colonizers behind windbreaks) Our re-sults revealed that during each of the 3 sampleperiods the slope of the relationship was 217Thus a multiplicative increase in capture was as-sociated with the presence of prune trees but in-dependent of any contribution from overwinteringA epos densities This result supported the wind-break hypothesis and demonstrated the effect may
be consistent for vineyards with a windbreak pres-ent Furthermore this result identifies an impor-tant additional mechanism that may be affectingA epos vineyard colonization in this system
These results underscore the importance of in-vestigating the ecological mechanisms underlyingsystems of habitat diversification in addition totests of overall effectiveness of diversification Un-derstanding the mechanisms involved will enhanceour ability to manipulate these systems to maxi-mize their effectiveness and provide a usefulframework to begin analyses of new systems
Variation in the Prunc Tree Effect Becauseoverwintering A epos densities varied among theprune tree sites we were interested in how themanagement of pnme trees affects overwinteringA epos populations and their subsequent abun-dance in vineyards Irrigation practices were foundto be a significant factor cplaining much of thevariation in overwintering A epos densities ob-served in pnme trees (Table 4) and somc of thevariation in trap catches in pnme tree vineyardsThe number of prune trees within a site also af-fected overwintering A epos density How treenumber exerts an influence on the density of over-wintering A epos (per square centimeter of pnmebark) is not known but we speculate that greaternumbers of trees may create a more favorable mi-crohabitat increasing the number of A epos thatsuccessfully emerge from their overwinteringhosts
Although tree number had a strong positive in-fluence on the density of A epos emergence holesper square centimeter of prune tree bark it wasnot a predictor of A epos captures in pnme treevineyards This is an enigmatic result At the outset
April IHH6 MUHllIY ET 1 HABITAT DIVEHSIFICATlON FOH BIOLOGICAL CONTHOL 503
of till study we expected that pnllle tree sites con-taining morl trelS would producl a larger pool ofmigrants for thl downwind vineyards The in-enased density (pcr squangt centimeter of bark) ofovcrwintcring A epos in prune trce sites with larg-cr tne 1lI11l11wrsonly strengthened onr expcctationthat trcc numbpr would lw a major determinant ofcady-season abundance in prune trep vineyardsIndclltl wc expccted that we would bl able to es-timatp t]1( minimum numbPI of trees needed togenlrate the gtelof augmentation of A cpos nec-pssary to achieve pest management Instead whatwe found is that trpe number is less a predictor ofalmmlancp in vinpyards than is the number of AlJOS produced ppr centinIPter of bark We also ca]-cu]ahd thc total number of A epos emerged fromovclvintlring sitps vithin an cntire prune tree site(a composite cstimate obtained as the product ofA e]1os ltknsity pcr squarc ccntimetcr of bark thetotal arca plr tngtp and the number of trpes persitc) as t]IP estimatp for A pos contributions fromprunl trpc sites This estimatc was however awcakcr prcdictor of early-season abundance invincyards than Pitill l thc numbcr of A cpos persquarc ccntimctcr of bark or pcr prune tree Howcan we cxplain this resn]t One possibility is thattrees within the largcr prune trcc sites Illay havecontributed uncIlm]]y to the number of vineyardmigrants vith thc rows of trccs directly adjacentto the vincyard producing the bulk of the waspstrapped in vincyards Additional experimentationwill be needed to test this and other potential ex-plauations for the lack of a trec number effect
These results have 2 implications for establish-ing and managing pnnll tree rcfuges for pest con-trol The 1st is that cnhanccd A cpos abundancemay he attainable with a relatively small numberof tnps TIIP 2nd is that irrigation of pnllle treesvill bp of primary importance for supporting highdpnsitics of ovprwintcring i cpos populationsMon detail(gtd rpcommcndations for establishingpnme trcc rcfugps will be presented in future pa-pers
Questious about thc pffect of vegetational diver-sity on tIll diversity and abundance of insect pop-ulations ngtcenty have rcceived considerable atteu-tion from ecologists and pest control specialistsThe rpsu]ts of thp cnrrent study not only supportedtl( hypothdical effect of pnme tree refuges on Acpos abundance but in a broader context providea test of the effect of wgctational diversity on nat-ural encmy abundance As a result this study hassatisfied tIl( 1st of 3 hypothesized natural enemyresponses which pr((licts greater natura] enemyabundance in diversified systems (Bussell 1989Andow 1991) In addition 011study identified thefollOving 2 mpchanisms underlying the influenceof prune trees the presence of an ove1vinteringhost and a windbreak effpct
Future papers vill examine the effect of over-wintering refuges on the remaining 2 hypotheses-the effect of pnnw trees on E elegalltula egg mor-
taity and E elcgalltula population densities withingrape vineyards
Acknowledgments
We extend special thanks to Nerissa Fretman MaddySamant John Tlnnter Jason May and Paul Higgs (De-partment of Entomology University of Califol1lia Davis)for field collections and sample processing Vc also ac-Irnowledge thc assistancc of Hobert Hoy Stan Grant andBrad Coleman (E amp J Gallo Winery Livingston CAl forlogistical and material support This research was fuudedby the Westem Hegional Integrated Pest Managementprogram the California Department of Ptsticidt Hegu-lation and the Locli-Woodbridge vVinegrapt Commis-sion This research was part of BGMs doctoral disser-tation at the University of California at Davis
References Cited
Altieri M A 1992 Agroecological foundatious of al-temative agliculture in Califomia Agric Ecosyst En-viron 39 23-53
Andow D A 1991 Vegetational diversity and artll1o-pod population response Annu Hev Entomol 36561-586
Corbett A und J A Rosenheim 1996 Impact of anatural enemy overwintering refuge and its iuterac-tion vith the surrounding landscape Ecol Entomol(in press)
Doutt R L and J Nakat bullbull 1965 Overwintering ref-uge of Anagrus epos (Hymenoptera Mymaridae) JEcon Entomol 58 586
1973 The Ruhus leafllOpper and its egg parasitoid Anendemic biotic system useful in grape-pest manage-ment Environ Entomol 2 381~186
Flaherty D L L T Wilson V M Stern and IIKido 1985 Biologicalcontrol in San Joaquin Valleyvineyards pp 501-520 Tn D G Herzog and M AHoy [eds] Biological control of agricnltural IPM sys-tems Viley New York
Flint M L and P A Roberts 1988 Using cropdiversity to manage pest problems some Californiaexamples Am J Altern Agric1 163-167
Gorclh G lIml D M Dunlmr 1977 A new Allagmsimportant in the biological control of Stephanitis tak-eljai and a key to the N01ih American species FlaEntomol 60 85-95
Herzo~ D c and J E Funderburk 1985 Plantresistance and cultural practiee interactious with hio-logical control pp 67-88 Til D G Ilerzog and MA Hoy [eds] Biological control of agliclllInral IPMsystems Wiley New York
Kiclo H D L Flaherty D F 80sh Dlul K A Val-ero 1984 French pmne trees as ovclwiuteringsites for the grape leafllOpper egg parasite Am JEnol Vitie 35 156-160
Letourneau D K 1987 The enemies hypothesis Tri-trophic interactions and vegetational diversity in trop-ical agroecosystel11sEcology 68 1616-1622
Lewis T bullbulllid J W Stephenson 1966 The penne-ability of artificial windbreaks and the distribution offlying insects in the leeward shelttred Will AnnAppl BioI 58 355-363
Mulla M S 1957 The biology of Typhlo(fhapnmi-cola (Edwards) and 1 quercus (Fabricius) (Cicadelli-dae lIomoptera) Ann Entomol SOlAm 50 76-87
504 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
National Research Council 1989 Alternative agri-culture National AcademyWashington DC
PaAel M D and P H Harvey 1988 Recent de-velopments in the analysis of comparative data QRev BioI 63 418-440
Pickett C H L T Wilson and D L Flaherty1990 The role of refuges in crop protection withreference to plantings of French prune trees in agrape agroecosystem pp 151-165 In N J BostanianL T Wilson and T J Dennehy [eds] Monitoringand integrated management of arthropod pests ofsmall fruit crops Intercept Andover
RusseD E P 1989 Enemies hypothesis A review ofthe effect of vegetational diversity on predatory in-sects and parasitoids Environ Entomol 18 590-599
Sokal R R and F J Rohlf 1981 Biometry 2nded Freeman San Francisco
SAS Institute 1989 JMP statistical program SAS In-stitute Cary NC
van Emden H F 1990 Plant diversity and naturalenemy efficiency in agroecosystems pp 63-80In MMackauer L E Ehler and J Roland [eds] Criticalissues in biological control Intercept Andover UK
Williams D W 1984 Ecology of the blackberry-leaf-hopper-parasite system and its relevance to Californiagrape agroecosystems Hilgardia 52 1--33
Wilson L T C H Pickett and D L Flaherty1989 French prune trees refuge for grape leafhop-per parasite Calif Agric 43(2) 7-8
Winkler A J J A Cook W M Kliewer and L ALider 1974 Climate and soils pp 58-75 In Gen-eral viticulture University of California Press Berke-ley
Received for publication 5 December 1994 accepted 6November 1995

April 1996 MURPHY ET AL HABITAT DIVERSIFICATION FOR BIOLOGICAL CONTHOL 497
Fil( 1 Pairedvineyardplot locationsin northernandctntra CaliforniaThe explodedview showsthe orien-tationof pnme tree and controlvineyardplots
evaluation of prune tree-enhanced biological con-trol required a large number of replicates Surveyswere conducted throughout the major wine grapeviticultural regions of central and northern Cali-fornia to locate 26 paired commercia] vineyardsone with adjacent upwind pnme trees and the oth-er lacking pnme trees (Fig 1) Paired vineyardswere matched for grape cultivar trellising anclmanagement practices Control (nonprune tree)vinlyards were positioned O~O kilometers fromrefuge sites and either parallel to or upwind ofpnll1e tree vineyard sites to minimize movementof ]eafllOppersor parasites between treatment andcontrol vineyards The paired plots were groupedinto 2 viticultural growing regions defined by thecUllmlative degree days occurring between Apriland October (Winkler et al 1974) Comparisonswere made between cooler growing regionslaquo3000 DD) and warmer regions (gt3000 DD)Two of the pnme tree sites used in the study wereplanted specifically for leafhopper control the re-mainder were commercial or remnant prune or-chards growing adjacent to the vineyard plots
Given the nature of the pnme tree habitat beingtested we were unable to ensure tnle randomiza-tion within our experimental design True random-ization would have required us to assign prune treeplantings randomly to plots and would have re-quired a delay of 3-5 yr for the pnme trees tobecome established and support large populationsof the alternate leafhopper host As a result weused vineyard sites located near previously estab-lished prune trees Had we chosen to work pri-
marily in vineyards with prune trees planted spe-cifically for pest management purposes we mighthave obtained a seriously biased sample of vine-yards for at least the 2 following reasons (1) thesevineyards might be more likely to have experi-enced severe leafhopper problems motivatinggrowers to look for enhanced means of controland (2) growers using prune tree plantings mightbe a relatively progressive subset of all growersand might also differ in other aspects of vineyardmanagement Because 10 of the 12 prune tree siteswere commercial pnme orchards and were notplanted for pest control we feel that the lack oftrue randomization is unlikely to introduce a sys-tematic bias into the analysis
Vineyard A epos Abundance Vineyard plotswere monitored to estimate densities of immigrat-ing A epos and E elegantula during the early sea-son before parasites or leafllOpper adults beganemerging from the 1st generation of reproductionin vineyards For both prune tree and control vine-yard plots 2 yellow sticky traps (75 by 125 mmyellow plastic cards Hilcor Plastics Los Ange]esCAl coated with Tanglefoot were attached to eachof 6 wooden poles (12 traps total per site) Poleswere placed in a transect at 10-m intervals alongthe 3rd vine row from the upwind edge of eachvineyard block Traps were oriented perpendicularto the predominant wind direction and positioned06 and 12 m above the vine canopy to minimizeany influence of vine canopies on trap captureTraps were deployed beginning 1 April (7 pairedsites) 15 Apri] (3 additional pairs) and 1 May (2final pairs) and were replaced twice monthlythrough 15 June 1992 All traps were taken to thelaboratory and examined under a dissection micro-scope to count the number of adult A epos cap-tured
Four vineyard plots were monitored weekly be-ginning 15 April to estimate the beginning of leaf-hopper oviposition and the 1st-generation (Fl)
nymphal emergence The beginning of ovipositionwas determined by examining field-collected grapeleaves under a dissection microscope for the pres-ence of E elegantula eggs The presence ofnymphs was determined by examining grape leavesin the field
Prune Tree Effect The prune tree sites variedwith respect to tree number tree size and orchardmanagement practices These differences in turncould influence overwintering A epos densitiesand cause variation in the treatment effect withinour design Therefore to control for variation inthe pnme refuge effect and to test hypothesesabout the contribution of A epos overwintering inprunes to vineyard abundance we sampled pmnetwigs to estimate overwintering A epos densities(details below) We also recorded the nlllnber oftrees and orchard management practices and es-timated the average rate of tree growth to deter-mine if these factors influence densities of over-wintering A epos (detai]s below)
498 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
The primary difference in management practic-es found among sites was whether or not treeswere irrigated Seven of the 12 sites used in thestudy received little or no supplemental water dur-ing the growing season We classified prune treesites that had a permanent irrigation system as ir-rigated trees Prune trees that were occasionallyprovided supplemental irrigation using portablesystems or that were never irrigated were classi-fied as nonirrigated
Overwintering A epos Densities Anagrus eposemerge from overwintering prune leafhopper eggsoviposited beneath the bark of 1-6-yr-old branchwood (Mulla 1957) Because 2-6-yr-old branchwood can accumulate A epos emergence holesover several years we sampled only 1-yr-oldbranch wood to distinguish the current yearsemergence holes from those of previous yearsPrune twigs were collected in late June and earlyJuly after the completion of A epos emergence Aminimum of 4 pnme twigs (l-yr-old) sampled from10 randomly selected prune trees at each site wasexamined (see twig sampling details described be-low) The length and diameter of twigs were mea-sured to calculate the surface area of 1-yr-oldgrowth (area = 7T d I where d is 1-yr-old twigdiameter I is twig length) Prune twigs were thenexamined under dissection microscopes for thepresence of A epos emergence holes These datawere then used to estimate the average number ofemergence holes per square centimeter of twigsurface area for each refuge site
PrWle Tree Growth Average tree growth var-ied substantially among prune tree sites To eval-uate the possible influence of this factor on A eposdensities we recorded the following 3 parametersas measures of tree growth average length diam-eter and number per tree of 1-yr-old twigs Theaverage length and diameter of twigs were deter-mined by randomly selecting 10 trees within eachpnme tree site for sampling A main branch wasremoved from each compass direction of each treefor a total of 40 branches per site Branches weretaken to the laboratory and the length and diam-eter of each 1-yr-old twig were recorded
The total number of 1-yr-old twigs per tree wasestimated by randomly selecting 4 trees from eachprune tree site for sampling Each tree was dividedin half vertically and horizontally to establish 4quadrants One quadrant position was randomlychosen in each of the 4 trees to count hvigs The4 hvig counts 1 example from each quadrant po-sition were pooled to estimate the mean numberof hvigs per tree
Prwle Tree Site Traps YellowstiCh)traps alsowere used to monitor A epos and E prunicolaadult numbers within pnme trees For each pnmesite 1 trap was placed in 3 randomly selectedprune trees 20-25 m from the ground Trapswere replaced twice monthly as described above
Statistical Procedures The average number ofA epos caught on traps within the vineyard plots
was analyzed using a split-plot analysis of variance(ANOVA) model Growing region was the mainfactor within the model and presence or absenceof a prune tree site was the subfactor Each vine-yard pair was treated as a statistical block to controlfor spatial variability in A epos densities Becausethe number of vineyard pairs sampled increased asthe season progressed separate univariate analyseswere conducted for each sampling period Theanalyseswere conducted to test the null hypothesisthat A epos abundance rates in vineyards are in-dependent of the presence or absence of prunetrees
Prune Tree Effect To test if differences invineyard parasite abundance can be explained bythe emergence of overwintering A epos frompnme trees multiple regression analyses were per-formed using A epos emergence hole densitiesfrom prunes mId A epos capture in control vine-yards as independent variables and A epos capturein prune tree vineyards as the response variableWe interpret A epos trap caphlfe in a control vine-yard as an estimate of the local abundance of par-asites moving into vineyards in the absence ofprune trees (area-wide A epos numbers) We test-ed the null hypothesis that differences in A eposabundance are independent of variation in over-wintering A epos densities within prune trees
Multiplicative Effects We evaluated the poten-tial influence of a vindbreak effect on A eposabundance in vineyards using model II regressionanalyses (Sokal and Rohlf 1981) to examine therelationship between A epos captures in the con-trol vineyards (the independent variable) and Aepos caphlfes in prune tree vineyards (the depen-dent variable) (statistical model prune tree vine-yard captures = 13 control vineyard captures + c+ error) If overwintering A epos within prunetrees were the only factor affecting A epos trapcapture we would expect the slope of the regres-sion equation (13) to approximate 10 with an in-crease in the intercept (c) reflecting an additiveincrease in the number of A epos contributed bypmne trees If on the other hand the slope pa-rameter is significantly greater than 10 a multi-plicative effect in pnme tree vineyard trap captureis present indicating the presence of another fac-tor such as a windbreak effect increasing A eposcapture independent of any contribution fromoverwintering A epos emerging from pnme trees
This analysis was used becanse model I regres-sion produces a biased estimate of the slope forlow values of the correlation coefficient (Sokal andRohlf 1981 Pagel and Harvey 1988) Model IIanalysis is also the most appropriate method foranalyzing the functional relationship between 2variables when both variables are subject to exper-imental error (Sokal and Rohlf 1981) A principalaxis (major axis) model II regression estimates theslope parameter by estimating the principal axisbetween 2 normally distributed random variables(Pagel and Harvey 1988) We used this analysis to
April 1996 MURPHY ET AL HABITAT DIVERSIFICATION FOR BIOLOGICAL CONTHOL 499
Results
Seasonal Patterns Examination of the early-season pattern of parasite and leafhopper trap cap-hires revealed that during the 1st sample period inearly to mid-April 1 or more A epos were caughtin 7 of the 14 vineyard plots monitored (Fig 2)During the 2nd sample period A epos was de-
Fill 2 Seasonal patterns of A epos and E elegantulain vineyard plots (A) A epos adult trap captures (meanplusmn SE) in control and treatment plots (B) E elegantulaadult trap captures (mcan plusmn SE) in control and treatmentplots (e) Period of E elegantrtla oviposition in vineyardplots (D) Pelod of A epos F I emergence (E) Period ofFI nymphal emergence Hatched lines indicate early ap-pearance in some vineyard plots Solid lines indicate ap-pemanle in all plots
tected in 18 of 20 vineyard plots The increasingA epos capture over time indicated that parasitesbegan moving into vineyards in April severalweeks after overwintered E elegantula adults be-gin moving into vineyards Overwintered adultstypically begin moving into vineyards from theiroverwintering habitats during March (Doutt andNakata 1965 Williams 1984) E elegantula eggswere detected in vineyards beginning in late April(Fig 2C) demonstrating that the appearance of Aepos coincided with the beginning of the oviposi-tion period of overwintering leafllOpper adultsThe large increases in A epos capture during the4th sample period in late May coincided with the1st appearance of A epos emergence holes ingrape leaves marking the emergence of adults ofthe Fl parasite generation (Fig 2D) Also duringthe 4th sample period Fl leafhopper nymphs be-gan appearing on grape leaves in most vineyards(Fig2E)
Regression analyses were done to assess the re-lationship between the final density of A cposemergence holes in 1-yr-old prune twigs and trapcapture in pnme trees to estimate the time of Aepos emergence from pnme twigs During the 1stsample period (15-18 April) only a single prunetree site had an A epos capture Regression anal-yses for the 2nd (24 April-l May) and 3rd (8-15May) periods revealed that A epos emergenceholes explained a significant amount of the varia-tion in trap captures (R2 = 088 n = 9 P = 0002and R2 = 042 n = 12 P = 0022 respectively)No relationship was found between area-wide trapcaptures in control vineyards and trap capturewithin pnme trees (R2 = 005 n = 9 P = 056 forthe 2nd sample period R2 = 017 n = 12 P =018 for the 3rd sample period) The conclllsionfrom these analyses is that sticky traps in prunetrees reflect the A epos emergence pattern fromprune trees and are not influenced significantly byarea-wide A epos moving from outside sourcesThe high degree of correlation during the 2ndsample period suggests that peak overwinteringemergence of A epos from prune twigs occllrredbetween 18 April and 1 May
Vineyard A epos Abundance We confined ouranalyses of A epos abundance to the 1st 3 sampledates when A epos captured on traps were mostlikely to have originated from sources outside thevineyard Traps in prune tree vineyards capturedgt2 times the number of parasites as did traps incontrol vineyards during the 2nd and 3rd sampledates (Fig 2A Table 1) No significant differencesin capture were detected between the 2 growingregions but variation in A epos captures was sub-stantial across the paired vineyards as indicated bythe significant block effect
Prune Tree Effect Modell regression analysesrevealed that captures of A epos in pnme treevineyards were closely correlated with captures incontrol vineyards for the 2nd and 3rd sample dates(R2 = 053 n = 10 P = 0016 R2 = 065 n = 12
F Lealltopper eggs
FlA epos emergence11I _
S8-S1S 522-529 613middot610
Date
11I1111I11I1111I11I11I
r3 Control vineyards
o Treatment vineyards
c
D
E
415-18 424middot511
e 2S
B Bbulll 20bull~ ISoSt 10~~ 5-3bull-lt
45
~ 40 A3 35bullis 30bull~ 2S
bullbull 20~ IS
10
ibull-lt
test the null hypothesis that there is no multipli-cative increase in A epos trap capture in prunetree vineyards relative to control vineyard captures
For all statistical analyses except the model IIregressions trap captures were subjected to a log(x+ 1) transformation and A epos emergence holedata were square root-transformed to normalizethe distribution of means Mean comparisons testswere accomplished using single degree of freedomtests (orthogonal contrasts) and all statistical anal-yses except the model II regressions were per-formed using the JMP statistical program for theMacintosh (SAS institute 1989) A Bonferroni cor-rection for multiple comparisons was used to main-tain the total experimentwise a error rate at 005for the ANOVA and regression analyses (Sokal andRohlf 1981)
500 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
Table 1 Nested ANOVA of the number of A epos(101110 trw1sformed) captured on traps in incyards withresl)ect to prune refuges growing regions and pairedvineyard plots (block effcct)
Date Source of df F 1variation
15-18 Apr Treahnent 1 393 0094Blocks (region) 5 617 0023Region 1 013 0735
24 Apr-l May Treatment 1 647 0032Blocks (region) 8 514 0012Region 1 460 0061
8-15 May Treahnent 1 1132 0006Blocks (region) 10 572 0004Begion 1 356 0086
One-tailed tests for hypothesis testing l lt clitical valueafter Bonferroni correction (1 lt 0033)
P = 0001 respectively) but not for the 1st sampledate (R2 = 048 n = 7 P = 009) The significantregressions of A epos capture between the treat-ment and control plots underlines the importanceof blocking by vineyard pair a key feature of ourexperimental design This result also confirms thatarea-wide A epos numbers were a significantsource of parasites in vineyards whether pnmetrees were present or not
The multiple regression analyses indicated thatA epos emergence from prunes was a factor ex-plaining A epos trap captures in prune tree vine-yards after controlling for area-wide abundancefrom control plot trap captures during the 2ndsample period (Table 2) This coincided with theestimated time of A epos emergence from prunetwigs as determined by sticky traps in pnme treesThe partial regression coefficients indicated thatemergence from pnmes explained a substantialamount of the variation in A epos capture duringthe first 2 sample periods in April but by the 3rdsample period in early May area-wide A epos cap-tures explained most variation in trap captures
Variation in Overwintering A epos DensitiesPrune tree sites varied in terms of the number oftrees per site the rate of tree growth the numberof I-yr-old twigs per tree and the density of Aepos emergence holes (Table 3) Much of this vari-
ation was generated by variation in irrigation prac-tices (Table 4) Irrigated trees were associated withgreater tree growth (average surface area of I-yr-old twigs) and lower twig numbers than nonirri-gated trees Irrigation was also associated withhigher A epos overwintering densities as well ashigher trap captures of E prunicola and A eposadults in tree canopies ANCOVA revealed thatboth irrigation (F = 2306 df = 1 12 P = 0001)and prune tree number (F = 1121 df = 1 12 P= 0009) had significant influences on the densityof A epos emergence holes in prune trees Therewas a positive linear relationship between thenumber of prune trees and the density of A eposemergence holes in trees (intercept = -00496slope = 00773) Nevertheless neither pnme treenumber nor irrigation had detectable effects onparasite catch in associated vineyards during the 3sample periods (P gt 0047)
We also estimated the average number of Aepos emergence holes per tree and the total num-ber of emergence holes per prune tree site to testif these parameters could explain captures in ad-jacent vineyards The number of emergence holesper tree was nonsignificant during the 1st sampleperiod (F = 89 df = 1 6 P = 0041) significantduring the 2nd period (F = 721 df = 1 9 P =0031) and non-significant during the 3rd period(F = 026 df = 1 11 P = 0625) The numberper prune tree site was however nonsignificantduring all 3 sample periods (P ~ 0096)
Multiplicative Effects The principal axis anal-yses (model II regression) of A epos trap capturesin control and prune tree vineyards suggest that amultiplicative effect was present during the 2ndand 3rd sample periods when both slope param-eters were significantly gt10 (Fig 3 B and C)During the 1st sample period the estimated slopewas similar to the other sample periods but notsignificantlydifferent from 10 (Fig 3A) These re-sults suggest that independent of any contributionfrom A epos overwintering in pmne trees prunetrees were associated with a multiplicative effectaugmenting A epos capture on the downwind sideof prune trees
Table 2 Multiple regression analysis of the relationship bctwcen the dependent variable 10fllO (A epos trap bullatch)ill prune trec plots and the oulcpendent variable sluarc root (A epos emergence density) from prlUlC sites and10gIO (A epos trap catch) 01 control plots
Sample date Source of variation df Slope (1 SE) F R2 pb
15--18 Apr Whole model test 2 7 119 086 0026Emergence density 17 106 (~ 0325) 106 075 0031Control captu res 17 067 (10237) 80 069 0047
24 Apr-l May Whole model test 210 168 083 0002Emergence density 110 237(1 0717) 109 073 0013Control captures 110 084 (10242) 119 075 OoIl
8-]5 May Whole model test 2 12 1015 069 0005E mergence density 1 19- 091 (10825) 121 035 0299Control captures 112 148 (~ 0363) 1674 081 0003
H2 for individual factors are partial correlation coefficients (r2)b l lt clitical value after Bonferroni correction (1 lt 0017)
April 1996 MURPHY ET AL HABITAT DIVERSIFICATION FOR BIOLOGICAL CONTROL 501
Table 3 Descrilltive larmnelers of pnnle lrees mnong experimenlalllols
Variable
No pnllle trees per siteNo 1-yr-old twigs per treel-yr-old twig surface area cm2
A (--pOS tlllfgtrgencp densitycm2
Mean
422511217
2030021
SE
11191774
4000056
Range
27-1216435-223837-1225
00023-00640
Discllssion
The goal of this research was to assess the influ-ence of pnme trees on early-season A epos abun-dance in vineyards and to test the hypothesizedecological mechanisms underlying this systemPomp trees were associated with significant in-creases in the density of A epos captured in nearbygrape vineyards relative to vineyards lacking prunetrees Pome tree vineyards were found on aver-agp to receive twice the number of early-seasonparasite captures Overwintering A epos emer-gpnce from prune trees was a significant predictorof A cpos trap capture in nearby vineyards duringthe 2nd sample period which coincides with ourestimated period of A epos emergence from prunetwigs Examination of the functional relationshipbetween control and pnme tree vineyard parasitecaptures also revealed a multiplicative response intrap capture on the downwind side of pnme treevineyard plots This suggests that another factorassociated with pnme trees other than overwin-tering A epos populations enhanced parasitenumhtrs in refuge vineyards
Seasonal Patterns Based on limited field dataDoutt and Nakata (1973) and Kido et al (1984)speculated that the effectiveness of overwinteringrefuges near vineyards was at least in part causedby the time of A epos emergence from overwin-tering hosts They based this hypothesis on the ob-senation that parasite emergence from prune treesor blackberries occurs during mid-April at the ap-proximate time overwintered E elegantula beginoviposition in vimmiddotyards They further speculatedthat enhancing early-season A epos colonizerscoupled with the high numerical response of theparasite (A epos completes 3 generations for eachE d(~antllia generation) improves biological con-trol of leafhoppers in vineyards The seasonal pat-tern of leafllOppers and parasites seen in the cur-rent study concurs with these previousobservations Across the replicated vineyard pairsour data show a consistent pattern of both area-
wide A epos captures and A epos emergence fromprune trees coinciding with the onset of oviposi-tion by overwintered E elegantula in vineyardsThus parasite abundance appears to be synchro-nized with the appearance of host eggs in grapevineyards
Vineyard A epos Abundance The replicatedcomparisons of A epos abundance in vineyardsshowed that the presence of overwintering habitatsproduced a substantial early-season increase in thedensity of A epos trapped in grape vineyards aresult consistent with the observations reported byDoutt and Nakata (1973) and Kido et a (1984)The elevated capture rates were seen despite dif-ferences in geographical regions and variation inthe number of area-wide A epos captures whichindicates that the prune tree effect operated undera wide variety of vineyard management and envi-ronmental conditions Based on these results weconcluded that the presence of prune trees wasassociated with elevated A epos numbers and asa result parasites may colonize these vineyards athigher rates than vineyards lacking nearby pnmetrees
Ecological Mechanisms Doutt and Nakata(1973) and Kido et a (1984) assumed that anyincrease in parasite abundance in vineyards (in-ferred using trap data or egg parasitism rates) wasthe direct result of overwintering emergence fromnearby habitats In the current studywe tested thisassumption by comparing the density of overwin-tering A epos emergence holes with A epos trapcapture in pnme tree vineyards after controllingfor area-wide colonizers The significant regressionduring the 2nd sample period indicated prunetrees were a significant source of parasite capturesduring the period of A epos emergence from over-wintered E prunicola eggs These results reveal 2important points regarding the influence of pnmetrees on parasite abundance in vineyards The 1stis that overwintering emergence is a significantecological factor determining early-season parasite
Table 4 Effeet of irrigalion on prIme lree growlh mld populalions of E prunicola mId A epos
Growth rate No twigs A epos A epos tmppecI Elnmicolacm2n (SE) emergence holescm2 trapped
Irrigated 304 498 642 176 00325 00063 950 209 2216+ 5411Nonirrigated 131 420 1475 163 Om05 0058 110 193 059t 507
a Surface area of 1-yr-old pnmc twigs (mean t SE) Numbers of adults caught on traps within prune tree canopies
Signifkantly different 001 lt P lt 005 SigniRcantl) different P lt 001
NoA epos captures in control plots
8060
Vol 25 no 2
402000
00
ENVIRONMENTAL ENTOMOLOGY
0008 00 10 20 30 40 50
B c50 80
bull 40 bull bull 6030
4020
10 20
06
502
lQ-Q~ A~
08bull~=E 06Q= 04tIJ~ 02aQto~l~~~0Z
Fig 3 Scatterplots of the functional relationship between A epos capture in pnme tree refuge vineyardsagainstA epos capture in control vineyards (A) 15-18 April (B = 1705 975 CL L = 058 U = nonestimatable) (8) 24April-l May (B = 238 975 CL L = 205 U = 279) (C) 8--15 May (B = 310 975 CL L = 221 U = 485)Confidence limits were established for 975 as a correction for multiple comparisons
numbers The 2nd is that the effect of prune treeson A epos abundance was dependent not only onthe presence of prune trees themselves but on thedensity of overwintering parasites residing withinthe trees This latter point may have important im-plications for using prune trees as a commercialpest control tactic (discussed below)
Lewis and Stephenson (1966) showed that flyinginsects dispersing in the surrounding environmentaccumulate in sheltered regions near physical bar-riers (natural or artificial windbreaks) They alsofound that insects aggregate behind windbreaks asa multiplicative function of the number of insectsdispersing in the surrounding environment Thiseffect was particularly pronounced for mymaridwasps Using elemental labeling techniques Cor-bett and Rosenheim (in press) found a pattern ofA epos capture suggesting that area-wide A eposnumbers accumulate at a rate 4 times greater im-mediately downwind of prune trees than found up-wind of trees Based on these results they hypoth-esized that a windbreak effect may also be amechanism enhancing abundance within the prunetree-vineyard system We tested this hypothesisacross our replicated study by examining the func-tional relationship between control vineyard trapcaptures (area-wide colonizers in the surroundingenvironment) and prune tree vineyard captures(number of colonizers behind windbreaks) Our re-sults revealed that during each of the 3 sampleperiods the slope of the relationship was 217Thus a multiplicative increase in capture was as-sociated with the presence of prune trees but in-dependent of any contribution from overwinteringA epos densities This result supported the wind-break hypothesis and demonstrated the effect may
be consistent for vineyards with a windbreak pres-ent Furthermore this result identifies an impor-tant additional mechanism that may be affectingA epos vineyard colonization in this system
These results underscore the importance of in-vestigating the ecological mechanisms underlyingsystems of habitat diversification in addition totests of overall effectiveness of diversification Un-derstanding the mechanisms involved will enhanceour ability to manipulate these systems to maxi-mize their effectiveness and provide a usefulframework to begin analyses of new systems
Variation in the Prunc Tree Effect Becauseoverwintering A epos densities varied among theprune tree sites we were interested in how themanagement of pnme trees affects overwinteringA epos populations and their subsequent abun-dance in vineyards Irrigation practices were foundto be a significant factor cplaining much of thevariation in overwintering A epos densities ob-served in pnme trees (Table 4) and somc of thevariation in trap catches in pnme tree vineyardsThe number of prune trees within a site also af-fected overwintering A epos density How treenumber exerts an influence on the density of over-wintering A epos (per square centimeter of pnmebark) is not known but we speculate that greaternumbers of trees may create a more favorable mi-crohabitat increasing the number of A epos thatsuccessfully emerge from their overwinteringhosts
Although tree number had a strong positive in-fluence on the density of A epos emergence holesper square centimeter of prune tree bark it wasnot a predictor of A epos captures in pnme treevineyards This is an enigmatic result At the outset
April IHH6 MUHllIY ET 1 HABITAT DIVEHSIFICATlON FOH BIOLOGICAL CONTHOL 503
of till study we expected that pnllle tree sites con-taining morl trelS would producl a larger pool ofmigrants for thl downwind vineyards The in-enased density (pcr squangt centimeter of bark) ofovcrwintcring A epos in prune trce sites with larg-cr tne 1lI11l11wrsonly strengthened onr expcctationthat trcc numbpr would lw a major determinant ofcady-season abundance in prune trep vineyardsIndclltl wc expccted that we would bl able to es-timatp t]1( minimum numbPI of trees needed togenlrate the gtelof augmentation of A cpos nec-pssary to achieve pest management Instead whatwe found is that trpe number is less a predictor ofalmmlancp in vinpyards than is the number of AlJOS produced ppr centinIPter of bark We also ca]-cu]ahd thc total number of A epos emerged fromovclvintlring sitps vithin an cntire prune tree site(a composite cstimate obtained as the product ofA e]1os ltknsity pcr squarc ccntimetcr of bark thetotal arca plr tngtp and the number of trpes persitc) as t]IP estimatp for A pos contributions fromprunl trpc sites This estimatc was however awcakcr prcdictor of early-season abundance invincyards than Pitill l thc numbcr of A cpos persquarc ccntimctcr of bark or pcr prune tree Howcan we cxplain this resn]t One possibility is thattrees within the largcr prune trcc sites Illay havecontributed uncIlm]]y to the number of vineyardmigrants vith thc rows of trccs directly adjacentto the vincyard producing the bulk of the waspstrapped in vincyards Additional experimentationwill be needed to test this and other potential ex-plauations for the lack of a trec number effect
These results have 2 implications for establish-ing and managing pnnll tree rcfuges for pest con-trol The 1st is that cnhanccd A cpos abundancemay he attainable with a relatively small numberof tnps TIIP 2nd is that irrigation of pnllle treesvill bp of primary importance for supporting highdpnsitics of ovprwintcring i cpos populationsMon detail(gtd rpcommcndations for establishingpnme trcc rcfugps will be presented in future pa-pers
Questious about thc pffect of vegetational diver-sity on tIll diversity and abundance of insect pop-ulations ngtcenty have rcceived considerable atteu-tion from ecologists and pest control specialistsThe rpsu]ts of thp cnrrent study not only supportedtl( hypothdical effect of pnme tree refuges on Acpos abundance but in a broader context providea test of the effect of wgctational diversity on nat-ural encmy abundance As a result this study hassatisfied tIl( 1st of 3 hypothesized natural enemyresponses which pr((licts greater natura] enemyabundance in diversified systems (Bussell 1989Andow 1991) In addition 011study identified thefollOving 2 mpchanisms underlying the influenceof prune trees the presence of an ove1vinteringhost and a windbreak effpct
Future papers vill examine the effect of over-wintering refuges on the remaining 2 hypotheses-the effect of pnnw trees on E elegalltula egg mor-
taity and E elcgalltula population densities withingrape vineyards
Acknowledgments
We extend special thanks to Nerissa Fretman MaddySamant John Tlnnter Jason May and Paul Higgs (De-partment of Entomology University of Califol1lia Davis)for field collections and sample processing Vc also ac-Irnowledge thc assistancc of Hobert Hoy Stan Grant andBrad Coleman (E amp J Gallo Winery Livingston CAl forlogistical and material support This research was fuudedby the Westem Hegional Integrated Pest Managementprogram the California Department of Ptsticidt Hegu-lation and the Locli-Woodbridge vVinegrapt Commis-sion This research was part of BGMs doctoral disser-tation at the University of California at Davis
References Cited
Altieri M A 1992 Agroecological foundatious of al-temative agliculture in Califomia Agric Ecosyst En-viron 39 23-53
Andow D A 1991 Vegetational diversity and artll1o-pod population response Annu Hev Entomol 36561-586
Corbett A und J A Rosenheim 1996 Impact of anatural enemy overwintering refuge and its iuterac-tion vith the surrounding landscape Ecol Entomol(in press)
Doutt R L and J Nakat bullbull 1965 Overwintering ref-uge of Anagrus epos (Hymenoptera Mymaridae) JEcon Entomol 58 586
1973 The Ruhus leafllOpper and its egg parasitoid Anendemic biotic system useful in grape-pest manage-ment Environ Entomol 2 381~186
Flaherty D L L T Wilson V M Stern and IIKido 1985 Biologicalcontrol in San Joaquin Valleyvineyards pp 501-520 Tn D G Herzog and M AHoy [eds] Biological control of agricnltural IPM sys-tems Viley New York
Flint M L and P A Roberts 1988 Using cropdiversity to manage pest problems some Californiaexamples Am J Altern Agric1 163-167
Gorclh G lIml D M Dunlmr 1977 A new Allagmsimportant in the biological control of Stephanitis tak-eljai and a key to the N01ih American species FlaEntomol 60 85-95
Herzo~ D c and J E Funderburk 1985 Plantresistance and cultural practiee interactious with hio-logical control pp 67-88 Til D G Ilerzog and MA Hoy [eds] Biological control of agliclllInral IPMsystems Wiley New York
Kiclo H D L Flaherty D F 80sh Dlul K A Val-ero 1984 French pmne trees as ovclwiuteringsites for the grape leafllOpper egg parasite Am JEnol Vitie 35 156-160
Letourneau D K 1987 The enemies hypothesis Tri-trophic interactions and vegetational diversity in trop-ical agroecosystel11sEcology 68 1616-1622
Lewis T bullbulllid J W Stephenson 1966 The penne-ability of artificial windbreaks and the distribution offlying insects in the leeward shelttred Will AnnAppl BioI 58 355-363
Mulla M S 1957 The biology of Typhlo(fhapnmi-cola (Edwards) and 1 quercus (Fabricius) (Cicadelli-dae lIomoptera) Ann Entomol SOlAm 50 76-87
504 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
National Research Council 1989 Alternative agri-culture National AcademyWashington DC
PaAel M D and P H Harvey 1988 Recent de-velopments in the analysis of comparative data QRev BioI 63 418-440
Pickett C H L T Wilson and D L Flaherty1990 The role of refuges in crop protection withreference to plantings of French prune trees in agrape agroecosystem pp 151-165 In N J BostanianL T Wilson and T J Dennehy [eds] Monitoringand integrated management of arthropod pests ofsmall fruit crops Intercept Andover
RusseD E P 1989 Enemies hypothesis A review ofthe effect of vegetational diversity on predatory in-sects and parasitoids Environ Entomol 18 590-599
Sokal R R and F J Rohlf 1981 Biometry 2nded Freeman San Francisco
SAS Institute 1989 JMP statistical program SAS In-stitute Cary NC
van Emden H F 1990 Plant diversity and naturalenemy efficiency in agroecosystems pp 63-80In MMackauer L E Ehler and J Roland [eds] Criticalissues in biological control Intercept Andover UK
Williams D W 1984 Ecology of the blackberry-leaf-hopper-parasite system and its relevance to Californiagrape agroecosystems Hilgardia 52 1--33
Wilson L T C H Pickett and D L Flaherty1989 French prune trees refuge for grape leafhop-per parasite Calif Agric 43(2) 7-8
Winkler A J J A Cook W M Kliewer and L ALider 1974 Climate and soils pp 58-75 In Gen-eral viticulture University of California Press Berke-ley
Received for publication 5 December 1994 accepted 6November 1995

498 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
The primary difference in management practic-es found among sites was whether or not treeswere irrigated Seven of the 12 sites used in thestudy received little or no supplemental water dur-ing the growing season We classified prune treesites that had a permanent irrigation system as ir-rigated trees Prune trees that were occasionallyprovided supplemental irrigation using portablesystems or that were never irrigated were classi-fied as nonirrigated
Overwintering A epos Densities Anagrus eposemerge from overwintering prune leafhopper eggsoviposited beneath the bark of 1-6-yr-old branchwood (Mulla 1957) Because 2-6-yr-old branchwood can accumulate A epos emergence holesover several years we sampled only 1-yr-oldbranch wood to distinguish the current yearsemergence holes from those of previous yearsPrune twigs were collected in late June and earlyJuly after the completion of A epos emergence Aminimum of 4 pnme twigs (l-yr-old) sampled from10 randomly selected prune trees at each site wasexamined (see twig sampling details described be-low) The length and diameter of twigs were mea-sured to calculate the surface area of 1-yr-oldgrowth (area = 7T d I where d is 1-yr-old twigdiameter I is twig length) Prune twigs were thenexamined under dissection microscopes for thepresence of A epos emergence holes These datawere then used to estimate the average number ofemergence holes per square centimeter of twigsurface area for each refuge site
PrWle Tree Growth Average tree growth var-ied substantially among prune tree sites To eval-uate the possible influence of this factor on A eposdensities we recorded the following 3 parametersas measures of tree growth average length diam-eter and number per tree of 1-yr-old twigs Theaverage length and diameter of twigs were deter-mined by randomly selecting 10 trees within eachpnme tree site for sampling A main branch wasremoved from each compass direction of each treefor a total of 40 branches per site Branches weretaken to the laboratory and the length and diam-eter of each 1-yr-old twig were recorded
The total number of 1-yr-old twigs per tree wasestimated by randomly selecting 4 trees from eachprune tree site for sampling Each tree was dividedin half vertically and horizontally to establish 4quadrants One quadrant position was randomlychosen in each of the 4 trees to count hvigs The4 hvig counts 1 example from each quadrant po-sition were pooled to estimate the mean numberof hvigs per tree
Prwle Tree Site Traps YellowstiCh)traps alsowere used to monitor A epos and E prunicolaadult numbers within pnme trees For each pnmesite 1 trap was placed in 3 randomly selectedprune trees 20-25 m from the ground Trapswere replaced twice monthly as described above
Statistical Procedures The average number ofA epos caught on traps within the vineyard plots
was analyzed using a split-plot analysis of variance(ANOVA) model Growing region was the mainfactor within the model and presence or absenceof a prune tree site was the subfactor Each vine-yard pair was treated as a statistical block to controlfor spatial variability in A epos densities Becausethe number of vineyard pairs sampled increased asthe season progressed separate univariate analyseswere conducted for each sampling period Theanalyseswere conducted to test the null hypothesisthat A epos abundance rates in vineyards are in-dependent of the presence or absence of prunetrees
Prune Tree Effect To test if differences invineyard parasite abundance can be explained bythe emergence of overwintering A epos frompnme trees multiple regression analyses were per-formed using A epos emergence hole densitiesfrom prunes mId A epos capture in control vine-yards as independent variables and A epos capturein prune tree vineyards as the response variableWe interpret A epos trap caphlfe in a control vine-yard as an estimate of the local abundance of par-asites moving into vineyards in the absence ofprune trees (area-wide A epos numbers) We test-ed the null hypothesis that differences in A eposabundance are independent of variation in over-wintering A epos densities within prune trees
Multiplicative Effects We evaluated the poten-tial influence of a vindbreak effect on A eposabundance in vineyards using model II regressionanalyses (Sokal and Rohlf 1981) to examine therelationship between A epos captures in the con-trol vineyards (the independent variable) and Aepos caphlfes in prune tree vineyards (the depen-dent variable) (statistical model prune tree vine-yard captures = 13 control vineyard captures + c+ error) If overwintering A epos within prunetrees were the only factor affecting A epos trapcapture we would expect the slope of the regres-sion equation (13) to approximate 10 with an in-crease in the intercept (c) reflecting an additiveincrease in the number of A epos contributed bypmne trees If on the other hand the slope pa-rameter is significantly greater than 10 a multi-plicative effect in pnme tree vineyard trap captureis present indicating the presence of another fac-tor such as a windbreak effect increasing A eposcapture independent of any contribution fromoverwintering A epos emerging from pnme trees
This analysis was used becanse model I regres-sion produces a biased estimate of the slope forlow values of the correlation coefficient (Sokal andRohlf 1981 Pagel and Harvey 1988) Model IIanalysis is also the most appropriate method foranalyzing the functional relationship between 2variables when both variables are subject to exper-imental error (Sokal and Rohlf 1981) A principalaxis (major axis) model II regression estimates theslope parameter by estimating the principal axisbetween 2 normally distributed random variables(Pagel and Harvey 1988) We used this analysis to
April 1996 MURPHY ET AL HABITAT DIVERSIFICATION FOR BIOLOGICAL CONTHOL 499
Results
Seasonal Patterns Examination of the early-season pattern of parasite and leafhopper trap cap-hires revealed that during the 1st sample period inearly to mid-April 1 or more A epos were caughtin 7 of the 14 vineyard plots monitored (Fig 2)During the 2nd sample period A epos was de-
Fill 2 Seasonal patterns of A epos and E elegantulain vineyard plots (A) A epos adult trap captures (meanplusmn SE) in control and treatment plots (B) E elegantulaadult trap captures (mcan plusmn SE) in control and treatmentplots (e) Period of E elegantrtla oviposition in vineyardplots (D) Pelod of A epos F I emergence (E) Period ofFI nymphal emergence Hatched lines indicate early ap-pearance in some vineyard plots Solid lines indicate ap-pemanle in all plots
tected in 18 of 20 vineyard plots The increasingA epos capture over time indicated that parasitesbegan moving into vineyards in April severalweeks after overwintered E elegantula adults be-gin moving into vineyards Overwintered adultstypically begin moving into vineyards from theiroverwintering habitats during March (Doutt andNakata 1965 Williams 1984) E elegantula eggswere detected in vineyards beginning in late April(Fig 2C) demonstrating that the appearance of Aepos coincided with the beginning of the oviposi-tion period of overwintering leafllOpper adultsThe large increases in A epos capture during the4th sample period in late May coincided with the1st appearance of A epos emergence holes ingrape leaves marking the emergence of adults ofthe Fl parasite generation (Fig 2D) Also duringthe 4th sample period Fl leafhopper nymphs be-gan appearing on grape leaves in most vineyards(Fig2E)
Regression analyses were done to assess the re-lationship between the final density of A cposemergence holes in 1-yr-old prune twigs and trapcapture in pnme trees to estimate the time of Aepos emergence from pnme twigs During the 1stsample period (15-18 April) only a single prunetree site had an A epos capture Regression anal-yses for the 2nd (24 April-l May) and 3rd (8-15May) periods revealed that A epos emergenceholes explained a significant amount of the varia-tion in trap captures (R2 = 088 n = 9 P = 0002and R2 = 042 n = 12 P = 0022 respectively)No relationship was found between area-wide trapcaptures in control vineyards and trap capturewithin pnme trees (R2 = 005 n = 9 P = 056 forthe 2nd sample period R2 = 017 n = 12 P =018 for the 3rd sample period) The conclllsionfrom these analyses is that sticky traps in prunetrees reflect the A epos emergence pattern fromprune trees and are not influenced significantly byarea-wide A epos moving from outside sourcesThe high degree of correlation during the 2ndsample period suggests that peak overwinteringemergence of A epos from prune twigs occllrredbetween 18 April and 1 May
Vineyard A epos Abundance We confined ouranalyses of A epos abundance to the 1st 3 sampledates when A epos captured on traps were mostlikely to have originated from sources outside thevineyard Traps in prune tree vineyards capturedgt2 times the number of parasites as did traps incontrol vineyards during the 2nd and 3rd sampledates (Fig 2A Table 1) No significant differencesin capture were detected between the 2 growingregions but variation in A epos captures was sub-stantial across the paired vineyards as indicated bythe significant block effect
Prune Tree Effect Modell regression analysesrevealed that captures of A epos in pnme treevineyards were closely correlated with captures incontrol vineyards for the 2nd and 3rd sample dates(R2 = 053 n = 10 P = 0016 R2 = 065 n = 12
F Lealltopper eggs
FlA epos emergence11I _
S8-S1S 522-529 613middot610
Date
11I1111I11I1111I11I11I
r3 Control vineyards
o Treatment vineyards
c
D
E
415-18 424middot511
e 2S
B Bbulll 20bull~ ISoSt 10~~ 5-3bull-lt
45
~ 40 A3 35bullis 30bull~ 2S
bullbull 20~ IS
10
ibull-lt
test the null hypothesis that there is no multipli-cative increase in A epos trap capture in prunetree vineyards relative to control vineyard captures
For all statistical analyses except the model IIregressions trap captures were subjected to a log(x+ 1) transformation and A epos emergence holedata were square root-transformed to normalizethe distribution of means Mean comparisons testswere accomplished using single degree of freedomtests (orthogonal contrasts) and all statistical anal-yses except the model II regressions were per-formed using the JMP statistical program for theMacintosh (SAS institute 1989) A Bonferroni cor-rection for multiple comparisons was used to main-tain the total experimentwise a error rate at 005for the ANOVA and regression analyses (Sokal andRohlf 1981)
500 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
Table 1 Nested ANOVA of the number of A epos(101110 trw1sformed) captured on traps in incyards withresl)ect to prune refuges growing regions and pairedvineyard plots (block effcct)
Date Source of df F 1variation
15-18 Apr Treahnent 1 393 0094Blocks (region) 5 617 0023Region 1 013 0735
24 Apr-l May Treatment 1 647 0032Blocks (region) 8 514 0012Region 1 460 0061
8-15 May Treahnent 1 1132 0006Blocks (region) 10 572 0004Begion 1 356 0086
One-tailed tests for hypothesis testing l lt clitical valueafter Bonferroni correction (1 lt 0033)
P = 0001 respectively) but not for the 1st sampledate (R2 = 048 n = 7 P = 009) The significantregressions of A epos capture between the treat-ment and control plots underlines the importanceof blocking by vineyard pair a key feature of ourexperimental design This result also confirms thatarea-wide A epos numbers were a significantsource of parasites in vineyards whether pnmetrees were present or not
The multiple regression analyses indicated thatA epos emergence from prunes was a factor ex-plaining A epos trap captures in prune tree vine-yards after controlling for area-wide abundancefrom control plot trap captures during the 2ndsample period (Table 2) This coincided with theestimated time of A epos emergence from prunetwigs as determined by sticky traps in pnme treesThe partial regression coefficients indicated thatemergence from pnmes explained a substantialamount of the variation in A epos capture duringthe first 2 sample periods in April but by the 3rdsample period in early May area-wide A epos cap-tures explained most variation in trap captures
Variation in Overwintering A epos DensitiesPrune tree sites varied in terms of the number oftrees per site the rate of tree growth the numberof I-yr-old twigs per tree and the density of Aepos emergence holes (Table 3) Much of this vari-
ation was generated by variation in irrigation prac-tices (Table 4) Irrigated trees were associated withgreater tree growth (average surface area of I-yr-old twigs) and lower twig numbers than nonirri-gated trees Irrigation was also associated withhigher A epos overwintering densities as well ashigher trap captures of E prunicola and A eposadults in tree canopies ANCOVA revealed thatboth irrigation (F = 2306 df = 1 12 P = 0001)and prune tree number (F = 1121 df = 1 12 P= 0009) had significant influences on the densityof A epos emergence holes in prune trees Therewas a positive linear relationship between thenumber of prune trees and the density of A eposemergence holes in trees (intercept = -00496slope = 00773) Nevertheless neither pnme treenumber nor irrigation had detectable effects onparasite catch in associated vineyards during the 3sample periods (P gt 0047)
We also estimated the average number of Aepos emergence holes per tree and the total num-ber of emergence holes per prune tree site to testif these parameters could explain captures in ad-jacent vineyards The number of emergence holesper tree was nonsignificant during the 1st sampleperiod (F = 89 df = 1 6 P = 0041) significantduring the 2nd period (F = 721 df = 1 9 P =0031) and non-significant during the 3rd period(F = 026 df = 1 11 P = 0625) The numberper prune tree site was however nonsignificantduring all 3 sample periods (P ~ 0096)
Multiplicative Effects The principal axis anal-yses (model II regression) of A epos trap capturesin control and prune tree vineyards suggest that amultiplicative effect was present during the 2ndand 3rd sample periods when both slope param-eters were significantly gt10 (Fig 3 B and C)During the 1st sample period the estimated slopewas similar to the other sample periods but notsignificantlydifferent from 10 (Fig 3A) These re-sults suggest that independent of any contributionfrom A epos overwintering in pmne trees prunetrees were associated with a multiplicative effectaugmenting A epos capture on the downwind sideof prune trees
Table 2 Multiple regression analysis of the relationship bctwcen the dependent variable 10fllO (A epos trap bullatch)ill prune trec plots and the oulcpendent variable sluarc root (A epos emergence density) from prlUlC sites and10gIO (A epos trap catch) 01 control plots
Sample date Source of variation df Slope (1 SE) F R2 pb
15--18 Apr Whole model test 2 7 119 086 0026Emergence density 17 106 (~ 0325) 106 075 0031Control captu res 17 067 (10237) 80 069 0047
24 Apr-l May Whole model test 210 168 083 0002Emergence density 110 237(1 0717) 109 073 0013Control captures 110 084 (10242) 119 075 OoIl
8-]5 May Whole model test 2 12 1015 069 0005E mergence density 1 19- 091 (10825) 121 035 0299Control captures 112 148 (~ 0363) 1674 081 0003
H2 for individual factors are partial correlation coefficients (r2)b l lt clitical value after Bonferroni correction (1 lt 0017)
April 1996 MURPHY ET AL HABITAT DIVERSIFICATION FOR BIOLOGICAL CONTROL 501
Table 3 Descrilltive larmnelers of pnnle lrees mnong experimenlalllols
Variable
No pnllle trees per siteNo 1-yr-old twigs per treel-yr-old twig surface area cm2
A (--pOS tlllfgtrgencp densitycm2
Mean
422511217
2030021
SE
11191774
4000056
Range
27-1216435-223837-1225
00023-00640
Discllssion
The goal of this research was to assess the influ-ence of pnme trees on early-season A epos abun-dance in vineyards and to test the hypothesizedecological mechanisms underlying this systemPomp trees were associated with significant in-creases in the density of A epos captured in nearbygrape vineyards relative to vineyards lacking prunetrees Pome tree vineyards were found on aver-agp to receive twice the number of early-seasonparasite captures Overwintering A epos emer-gpnce from prune trees was a significant predictorof A cpos trap capture in nearby vineyards duringthe 2nd sample period which coincides with ourestimated period of A epos emergence from prunetwigs Examination of the functional relationshipbetween control and pnme tree vineyard parasitecaptures also revealed a multiplicative response intrap capture on the downwind side of pnme treevineyard plots This suggests that another factorassociated with pnme trees other than overwin-tering A epos populations enhanced parasitenumhtrs in refuge vineyards
Seasonal Patterns Based on limited field dataDoutt and Nakata (1973) and Kido et al (1984)speculated that the effectiveness of overwinteringrefuges near vineyards was at least in part causedby the time of A epos emergence from overwin-tering hosts They based this hypothesis on the ob-senation that parasite emergence from prune treesor blackberries occurs during mid-April at the ap-proximate time overwintered E elegantula beginoviposition in vimmiddotyards They further speculatedthat enhancing early-season A epos colonizerscoupled with the high numerical response of theparasite (A epos completes 3 generations for eachE d(~antllia generation) improves biological con-trol of leafhoppers in vineyards The seasonal pat-tern of leafllOppers and parasites seen in the cur-rent study concurs with these previousobservations Across the replicated vineyard pairsour data show a consistent pattern of both area-
wide A epos captures and A epos emergence fromprune trees coinciding with the onset of oviposi-tion by overwintered E elegantula in vineyardsThus parasite abundance appears to be synchro-nized with the appearance of host eggs in grapevineyards
Vineyard A epos Abundance The replicatedcomparisons of A epos abundance in vineyardsshowed that the presence of overwintering habitatsproduced a substantial early-season increase in thedensity of A epos trapped in grape vineyards aresult consistent with the observations reported byDoutt and Nakata (1973) and Kido et a (1984)The elevated capture rates were seen despite dif-ferences in geographical regions and variation inthe number of area-wide A epos captures whichindicates that the prune tree effect operated undera wide variety of vineyard management and envi-ronmental conditions Based on these results weconcluded that the presence of prune trees wasassociated with elevated A epos numbers and asa result parasites may colonize these vineyards athigher rates than vineyards lacking nearby pnmetrees
Ecological Mechanisms Doutt and Nakata(1973) and Kido et a (1984) assumed that anyincrease in parasite abundance in vineyards (in-ferred using trap data or egg parasitism rates) wasthe direct result of overwintering emergence fromnearby habitats In the current studywe tested thisassumption by comparing the density of overwin-tering A epos emergence holes with A epos trapcapture in pnme tree vineyards after controllingfor area-wide colonizers The significant regressionduring the 2nd sample period indicated prunetrees were a significant source of parasite capturesduring the period of A epos emergence from over-wintered E prunicola eggs These results reveal 2important points regarding the influence of pnmetrees on parasite abundance in vineyards The 1stis that overwintering emergence is a significantecological factor determining early-season parasite
Table 4 Effeet of irrigalion on prIme lree growlh mld populalions of E prunicola mId A epos
Growth rate No twigs A epos A epos tmppecI Elnmicolacm2n (SE) emergence holescm2 trapped
Irrigated 304 498 642 176 00325 00063 950 209 2216+ 5411Nonirrigated 131 420 1475 163 Om05 0058 110 193 059t 507
a Surface area of 1-yr-old pnmc twigs (mean t SE) Numbers of adults caught on traps within prune tree canopies
Signifkantly different 001 lt P lt 005 SigniRcantl) different P lt 001
NoA epos captures in control plots
8060
Vol 25 no 2
402000
00
ENVIRONMENTAL ENTOMOLOGY
0008 00 10 20 30 40 50
B c50 80
bull 40 bull bull 6030
4020
10 20
06
502
lQ-Q~ A~
08bull~=E 06Q= 04tIJ~ 02aQto~l~~~0Z
Fig 3 Scatterplots of the functional relationship between A epos capture in pnme tree refuge vineyardsagainstA epos capture in control vineyards (A) 15-18 April (B = 1705 975 CL L = 058 U = nonestimatable) (8) 24April-l May (B = 238 975 CL L = 205 U = 279) (C) 8--15 May (B = 310 975 CL L = 221 U = 485)Confidence limits were established for 975 as a correction for multiple comparisons
numbers The 2nd is that the effect of prune treeson A epos abundance was dependent not only onthe presence of prune trees themselves but on thedensity of overwintering parasites residing withinthe trees This latter point may have important im-plications for using prune trees as a commercialpest control tactic (discussed below)
Lewis and Stephenson (1966) showed that flyinginsects dispersing in the surrounding environmentaccumulate in sheltered regions near physical bar-riers (natural or artificial windbreaks) They alsofound that insects aggregate behind windbreaks asa multiplicative function of the number of insectsdispersing in the surrounding environment Thiseffect was particularly pronounced for mymaridwasps Using elemental labeling techniques Cor-bett and Rosenheim (in press) found a pattern ofA epos capture suggesting that area-wide A eposnumbers accumulate at a rate 4 times greater im-mediately downwind of prune trees than found up-wind of trees Based on these results they hypoth-esized that a windbreak effect may also be amechanism enhancing abundance within the prunetree-vineyard system We tested this hypothesisacross our replicated study by examining the func-tional relationship between control vineyard trapcaptures (area-wide colonizers in the surroundingenvironment) and prune tree vineyard captures(number of colonizers behind windbreaks) Our re-sults revealed that during each of the 3 sampleperiods the slope of the relationship was 217Thus a multiplicative increase in capture was as-sociated with the presence of prune trees but in-dependent of any contribution from overwinteringA epos densities This result supported the wind-break hypothesis and demonstrated the effect may
be consistent for vineyards with a windbreak pres-ent Furthermore this result identifies an impor-tant additional mechanism that may be affectingA epos vineyard colonization in this system
These results underscore the importance of in-vestigating the ecological mechanisms underlyingsystems of habitat diversification in addition totests of overall effectiveness of diversification Un-derstanding the mechanisms involved will enhanceour ability to manipulate these systems to maxi-mize their effectiveness and provide a usefulframework to begin analyses of new systems
Variation in the Prunc Tree Effect Becauseoverwintering A epos densities varied among theprune tree sites we were interested in how themanagement of pnme trees affects overwinteringA epos populations and their subsequent abun-dance in vineyards Irrigation practices were foundto be a significant factor cplaining much of thevariation in overwintering A epos densities ob-served in pnme trees (Table 4) and somc of thevariation in trap catches in pnme tree vineyardsThe number of prune trees within a site also af-fected overwintering A epos density How treenumber exerts an influence on the density of over-wintering A epos (per square centimeter of pnmebark) is not known but we speculate that greaternumbers of trees may create a more favorable mi-crohabitat increasing the number of A epos thatsuccessfully emerge from their overwinteringhosts
Although tree number had a strong positive in-fluence on the density of A epos emergence holesper square centimeter of prune tree bark it wasnot a predictor of A epos captures in pnme treevineyards This is an enigmatic result At the outset
April IHH6 MUHllIY ET 1 HABITAT DIVEHSIFICATlON FOH BIOLOGICAL CONTHOL 503
of till study we expected that pnllle tree sites con-taining morl trelS would producl a larger pool ofmigrants for thl downwind vineyards The in-enased density (pcr squangt centimeter of bark) ofovcrwintcring A epos in prune trce sites with larg-cr tne 1lI11l11wrsonly strengthened onr expcctationthat trcc numbpr would lw a major determinant ofcady-season abundance in prune trep vineyardsIndclltl wc expccted that we would bl able to es-timatp t]1( minimum numbPI of trees needed togenlrate the gtelof augmentation of A cpos nec-pssary to achieve pest management Instead whatwe found is that trpe number is less a predictor ofalmmlancp in vinpyards than is the number of AlJOS produced ppr centinIPter of bark We also ca]-cu]ahd thc total number of A epos emerged fromovclvintlring sitps vithin an cntire prune tree site(a composite cstimate obtained as the product ofA e]1os ltknsity pcr squarc ccntimetcr of bark thetotal arca plr tngtp and the number of trpes persitc) as t]IP estimatp for A pos contributions fromprunl trpc sites This estimatc was however awcakcr prcdictor of early-season abundance invincyards than Pitill l thc numbcr of A cpos persquarc ccntimctcr of bark or pcr prune tree Howcan we cxplain this resn]t One possibility is thattrees within the largcr prune trcc sites Illay havecontributed uncIlm]]y to the number of vineyardmigrants vith thc rows of trccs directly adjacentto the vincyard producing the bulk of the waspstrapped in vincyards Additional experimentationwill be needed to test this and other potential ex-plauations for the lack of a trec number effect
These results have 2 implications for establish-ing and managing pnnll tree rcfuges for pest con-trol The 1st is that cnhanccd A cpos abundancemay he attainable with a relatively small numberof tnps TIIP 2nd is that irrigation of pnllle treesvill bp of primary importance for supporting highdpnsitics of ovprwintcring i cpos populationsMon detail(gtd rpcommcndations for establishingpnme trcc rcfugps will be presented in future pa-pers
Questious about thc pffect of vegetational diver-sity on tIll diversity and abundance of insect pop-ulations ngtcenty have rcceived considerable atteu-tion from ecologists and pest control specialistsThe rpsu]ts of thp cnrrent study not only supportedtl( hypothdical effect of pnme tree refuges on Acpos abundance but in a broader context providea test of the effect of wgctational diversity on nat-ural encmy abundance As a result this study hassatisfied tIl( 1st of 3 hypothesized natural enemyresponses which pr((licts greater natura] enemyabundance in diversified systems (Bussell 1989Andow 1991) In addition 011study identified thefollOving 2 mpchanisms underlying the influenceof prune trees the presence of an ove1vinteringhost and a windbreak effpct
Future papers vill examine the effect of over-wintering refuges on the remaining 2 hypotheses-the effect of pnnw trees on E elegalltula egg mor-
taity and E elcgalltula population densities withingrape vineyards
Acknowledgments
We extend special thanks to Nerissa Fretman MaddySamant John Tlnnter Jason May and Paul Higgs (De-partment of Entomology University of Califol1lia Davis)for field collections and sample processing Vc also ac-Irnowledge thc assistancc of Hobert Hoy Stan Grant andBrad Coleman (E amp J Gallo Winery Livingston CAl forlogistical and material support This research was fuudedby the Westem Hegional Integrated Pest Managementprogram the California Department of Ptsticidt Hegu-lation and the Locli-Woodbridge vVinegrapt Commis-sion This research was part of BGMs doctoral disser-tation at the University of California at Davis
References Cited
Altieri M A 1992 Agroecological foundatious of al-temative agliculture in Califomia Agric Ecosyst En-viron 39 23-53
Andow D A 1991 Vegetational diversity and artll1o-pod population response Annu Hev Entomol 36561-586
Corbett A und J A Rosenheim 1996 Impact of anatural enemy overwintering refuge and its iuterac-tion vith the surrounding landscape Ecol Entomol(in press)
Doutt R L and J Nakat bullbull 1965 Overwintering ref-uge of Anagrus epos (Hymenoptera Mymaridae) JEcon Entomol 58 586
1973 The Ruhus leafllOpper and its egg parasitoid Anendemic biotic system useful in grape-pest manage-ment Environ Entomol 2 381~186
Flaherty D L L T Wilson V M Stern and IIKido 1985 Biologicalcontrol in San Joaquin Valleyvineyards pp 501-520 Tn D G Herzog and M AHoy [eds] Biological control of agricnltural IPM sys-tems Viley New York
Flint M L and P A Roberts 1988 Using cropdiversity to manage pest problems some Californiaexamples Am J Altern Agric1 163-167
Gorclh G lIml D M Dunlmr 1977 A new Allagmsimportant in the biological control of Stephanitis tak-eljai and a key to the N01ih American species FlaEntomol 60 85-95
Herzo~ D c and J E Funderburk 1985 Plantresistance and cultural practiee interactious with hio-logical control pp 67-88 Til D G Ilerzog and MA Hoy [eds] Biological control of agliclllInral IPMsystems Wiley New York
Kiclo H D L Flaherty D F 80sh Dlul K A Val-ero 1984 French pmne trees as ovclwiuteringsites for the grape leafllOpper egg parasite Am JEnol Vitie 35 156-160
Letourneau D K 1987 The enemies hypothesis Tri-trophic interactions and vegetational diversity in trop-ical agroecosystel11sEcology 68 1616-1622
Lewis T bullbulllid J W Stephenson 1966 The penne-ability of artificial windbreaks and the distribution offlying insects in the leeward shelttred Will AnnAppl BioI 58 355-363
Mulla M S 1957 The biology of Typhlo(fhapnmi-cola (Edwards) and 1 quercus (Fabricius) (Cicadelli-dae lIomoptera) Ann Entomol SOlAm 50 76-87
504 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
National Research Council 1989 Alternative agri-culture National AcademyWashington DC
PaAel M D and P H Harvey 1988 Recent de-velopments in the analysis of comparative data QRev BioI 63 418-440
Pickett C H L T Wilson and D L Flaherty1990 The role of refuges in crop protection withreference to plantings of French prune trees in agrape agroecosystem pp 151-165 In N J BostanianL T Wilson and T J Dennehy [eds] Monitoringand integrated management of arthropod pests ofsmall fruit crops Intercept Andover
RusseD E P 1989 Enemies hypothesis A review ofthe effect of vegetational diversity on predatory in-sects and parasitoids Environ Entomol 18 590-599
Sokal R R and F J Rohlf 1981 Biometry 2nded Freeman San Francisco
SAS Institute 1989 JMP statistical program SAS In-stitute Cary NC
van Emden H F 1990 Plant diversity and naturalenemy efficiency in agroecosystems pp 63-80In MMackauer L E Ehler and J Roland [eds] Criticalissues in biological control Intercept Andover UK
Williams D W 1984 Ecology of the blackberry-leaf-hopper-parasite system and its relevance to Californiagrape agroecosystems Hilgardia 52 1--33
Wilson L T C H Pickett and D L Flaherty1989 French prune trees refuge for grape leafhop-per parasite Calif Agric 43(2) 7-8
Winkler A J J A Cook W M Kliewer and L ALider 1974 Climate and soils pp 58-75 In Gen-eral viticulture University of California Press Berke-ley
Received for publication 5 December 1994 accepted 6November 1995

April 1996 MURPHY ET AL HABITAT DIVERSIFICATION FOR BIOLOGICAL CONTHOL 499
Results
Seasonal Patterns Examination of the early-season pattern of parasite and leafhopper trap cap-hires revealed that during the 1st sample period inearly to mid-April 1 or more A epos were caughtin 7 of the 14 vineyard plots monitored (Fig 2)During the 2nd sample period A epos was de-
Fill 2 Seasonal patterns of A epos and E elegantulain vineyard plots (A) A epos adult trap captures (meanplusmn SE) in control and treatment plots (B) E elegantulaadult trap captures (mcan plusmn SE) in control and treatmentplots (e) Period of E elegantrtla oviposition in vineyardplots (D) Pelod of A epos F I emergence (E) Period ofFI nymphal emergence Hatched lines indicate early ap-pearance in some vineyard plots Solid lines indicate ap-pemanle in all plots
tected in 18 of 20 vineyard plots The increasingA epos capture over time indicated that parasitesbegan moving into vineyards in April severalweeks after overwintered E elegantula adults be-gin moving into vineyards Overwintered adultstypically begin moving into vineyards from theiroverwintering habitats during March (Doutt andNakata 1965 Williams 1984) E elegantula eggswere detected in vineyards beginning in late April(Fig 2C) demonstrating that the appearance of Aepos coincided with the beginning of the oviposi-tion period of overwintering leafllOpper adultsThe large increases in A epos capture during the4th sample period in late May coincided with the1st appearance of A epos emergence holes ingrape leaves marking the emergence of adults ofthe Fl parasite generation (Fig 2D) Also duringthe 4th sample period Fl leafhopper nymphs be-gan appearing on grape leaves in most vineyards(Fig2E)
Regression analyses were done to assess the re-lationship between the final density of A cposemergence holes in 1-yr-old prune twigs and trapcapture in pnme trees to estimate the time of Aepos emergence from pnme twigs During the 1stsample period (15-18 April) only a single prunetree site had an A epos capture Regression anal-yses for the 2nd (24 April-l May) and 3rd (8-15May) periods revealed that A epos emergenceholes explained a significant amount of the varia-tion in trap captures (R2 = 088 n = 9 P = 0002and R2 = 042 n = 12 P = 0022 respectively)No relationship was found between area-wide trapcaptures in control vineyards and trap capturewithin pnme trees (R2 = 005 n = 9 P = 056 forthe 2nd sample period R2 = 017 n = 12 P =018 for the 3rd sample period) The conclllsionfrom these analyses is that sticky traps in prunetrees reflect the A epos emergence pattern fromprune trees and are not influenced significantly byarea-wide A epos moving from outside sourcesThe high degree of correlation during the 2ndsample period suggests that peak overwinteringemergence of A epos from prune twigs occllrredbetween 18 April and 1 May
Vineyard A epos Abundance We confined ouranalyses of A epos abundance to the 1st 3 sampledates when A epos captured on traps were mostlikely to have originated from sources outside thevineyard Traps in prune tree vineyards capturedgt2 times the number of parasites as did traps incontrol vineyards during the 2nd and 3rd sampledates (Fig 2A Table 1) No significant differencesin capture were detected between the 2 growingregions but variation in A epos captures was sub-stantial across the paired vineyards as indicated bythe significant block effect
Prune Tree Effect Modell regression analysesrevealed that captures of A epos in pnme treevineyards were closely correlated with captures incontrol vineyards for the 2nd and 3rd sample dates(R2 = 053 n = 10 P = 0016 R2 = 065 n = 12
F Lealltopper eggs
FlA epos emergence11I _
S8-S1S 522-529 613middot610
Date
11I1111I11I1111I11I11I
r3 Control vineyards
o Treatment vineyards
c
D
E
415-18 424middot511
e 2S
B Bbulll 20bull~ ISoSt 10~~ 5-3bull-lt
45
~ 40 A3 35bullis 30bull~ 2S
bullbull 20~ IS
10
ibull-lt
test the null hypothesis that there is no multipli-cative increase in A epos trap capture in prunetree vineyards relative to control vineyard captures
For all statistical analyses except the model IIregressions trap captures were subjected to a log(x+ 1) transformation and A epos emergence holedata were square root-transformed to normalizethe distribution of means Mean comparisons testswere accomplished using single degree of freedomtests (orthogonal contrasts) and all statistical anal-yses except the model II regressions were per-formed using the JMP statistical program for theMacintosh (SAS institute 1989) A Bonferroni cor-rection for multiple comparisons was used to main-tain the total experimentwise a error rate at 005for the ANOVA and regression analyses (Sokal andRohlf 1981)
500 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
Table 1 Nested ANOVA of the number of A epos(101110 trw1sformed) captured on traps in incyards withresl)ect to prune refuges growing regions and pairedvineyard plots (block effcct)
Date Source of df F 1variation
15-18 Apr Treahnent 1 393 0094Blocks (region) 5 617 0023Region 1 013 0735
24 Apr-l May Treatment 1 647 0032Blocks (region) 8 514 0012Region 1 460 0061
8-15 May Treahnent 1 1132 0006Blocks (region) 10 572 0004Begion 1 356 0086
One-tailed tests for hypothesis testing l lt clitical valueafter Bonferroni correction (1 lt 0033)
P = 0001 respectively) but not for the 1st sampledate (R2 = 048 n = 7 P = 009) The significantregressions of A epos capture between the treat-ment and control plots underlines the importanceof blocking by vineyard pair a key feature of ourexperimental design This result also confirms thatarea-wide A epos numbers were a significantsource of parasites in vineyards whether pnmetrees were present or not
The multiple regression analyses indicated thatA epos emergence from prunes was a factor ex-plaining A epos trap captures in prune tree vine-yards after controlling for area-wide abundancefrom control plot trap captures during the 2ndsample period (Table 2) This coincided with theestimated time of A epos emergence from prunetwigs as determined by sticky traps in pnme treesThe partial regression coefficients indicated thatemergence from pnmes explained a substantialamount of the variation in A epos capture duringthe first 2 sample periods in April but by the 3rdsample period in early May area-wide A epos cap-tures explained most variation in trap captures
Variation in Overwintering A epos DensitiesPrune tree sites varied in terms of the number oftrees per site the rate of tree growth the numberof I-yr-old twigs per tree and the density of Aepos emergence holes (Table 3) Much of this vari-
ation was generated by variation in irrigation prac-tices (Table 4) Irrigated trees were associated withgreater tree growth (average surface area of I-yr-old twigs) and lower twig numbers than nonirri-gated trees Irrigation was also associated withhigher A epos overwintering densities as well ashigher trap captures of E prunicola and A eposadults in tree canopies ANCOVA revealed thatboth irrigation (F = 2306 df = 1 12 P = 0001)and prune tree number (F = 1121 df = 1 12 P= 0009) had significant influences on the densityof A epos emergence holes in prune trees Therewas a positive linear relationship between thenumber of prune trees and the density of A eposemergence holes in trees (intercept = -00496slope = 00773) Nevertheless neither pnme treenumber nor irrigation had detectable effects onparasite catch in associated vineyards during the 3sample periods (P gt 0047)
We also estimated the average number of Aepos emergence holes per tree and the total num-ber of emergence holes per prune tree site to testif these parameters could explain captures in ad-jacent vineyards The number of emergence holesper tree was nonsignificant during the 1st sampleperiod (F = 89 df = 1 6 P = 0041) significantduring the 2nd period (F = 721 df = 1 9 P =0031) and non-significant during the 3rd period(F = 026 df = 1 11 P = 0625) The numberper prune tree site was however nonsignificantduring all 3 sample periods (P ~ 0096)
Multiplicative Effects The principal axis anal-yses (model II regression) of A epos trap capturesin control and prune tree vineyards suggest that amultiplicative effect was present during the 2ndand 3rd sample periods when both slope param-eters were significantly gt10 (Fig 3 B and C)During the 1st sample period the estimated slopewas similar to the other sample periods but notsignificantlydifferent from 10 (Fig 3A) These re-sults suggest that independent of any contributionfrom A epos overwintering in pmne trees prunetrees were associated with a multiplicative effectaugmenting A epos capture on the downwind sideof prune trees
Table 2 Multiple regression analysis of the relationship bctwcen the dependent variable 10fllO (A epos trap bullatch)ill prune trec plots and the oulcpendent variable sluarc root (A epos emergence density) from prlUlC sites and10gIO (A epos trap catch) 01 control plots
Sample date Source of variation df Slope (1 SE) F R2 pb
15--18 Apr Whole model test 2 7 119 086 0026Emergence density 17 106 (~ 0325) 106 075 0031Control captu res 17 067 (10237) 80 069 0047
24 Apr-l May Whole model test 210 168 083 0002Emergence density 110 237(1 0717) 109 073 0013Control captures 110 084 (10242) 119 075 OoIl
8-]5 May Whole model test 2 12 1015 069 0005E mergence density 1 19- 091 (10825) 121 035 0299Control captures 112 148 (~ 0363) 1674 081 0003
H2 for individual factors are partial correlation coefficients (r2)b l lt clitical value after Bonferroni correction (1 lt 0017)
April 1996 MURPHY ET AL HABITAT DIVERSIFICATION FOR BIOLOGICAL CONTROL 501
Table 3 Descrilltive larmnelers of pnnle lrees mnong experimenlalllols
Variable
No pnllle trees per siteNo 1-yr-old twigs per treel-yr-old twig surface area cm2
A (--pOS tlllfgtrgencp densitycm2
Mean
422511217
2030021
SE
11191774
4000056
Range
27-1216435-223837-1225
00023-00640
Discllssion
The goal of this research was to assess the influ-ence of pnme trees on early-season A epos abun-dance in vineyards and to test the hypothesizedecological mechanisms underlying this systemPomp trees were associated with significant in-creases in the density of A epos captured in nearbygrape vineyards relative to vineyards lacking prunetrees Pome tree vineyards were found on aver-agp to receive twice the number of early-seasonparasite captures Overwintering A epos emer-gpnce from prune trees was a significant predictorof A cpos trap capture in nearby vineyards duringthe 2nd sample period which coincides with ourestimated period of A epos emergence from prunetwigs Examination of the functional relationshipbetween control and pnme tree vineyard parasitecaptures also revealed a multiplicative response intrap capture on the downwind side of pnme treevineyard plots This suggests that another factorassociated with pnme trees other than overwin-tering A epos populations enhanced parasitenumhtrs in refuge vineyards
Seasonal Patterns Based on limited field dataDoutt and Nakata (1973) and Kido et al (1984)speculated that the effectiveness of overwinteringrefuges near vineyards was at least in part causedby the time of A epos emergence from overwin-tering hosts They based this hypothesis on the ob-senation that parasite emergence from prune treesor blackberries occurs during mid-April at the ap-proximate time overwintered E elegantula beginoviposition in vimmiddotyards They further speculatedthat enhancing early-season A epos colonizerscoupled with the high numerical response of theparasite (A epos completes 3 generations for eachE d(~antllia generation) improves biological con-trol of leafhoppers in vineyards The seasonal pat-tern of leafllOppers and parasites seen in the cur-rent study concurs with these previousobservations Across the replicated vineyard pairsour data show a consistent pattern of both area-
wide A epos captures and A epos emergence fromprune trees coinciding with the onset of oviposi-tion by overwintered E elegantula in vineyardsThus parasite abundance appears to be synchro-nized with the appearance of host eggs in grapevineyards
Vineyard A epos Abundance The replicatedcomparisons of A epos abundance in vineyardsshowed that the presence of overwintering habitatsproduced a substantial early-season increase in thedensity of A epos trapped in grape vineyards aresult consistent with the observations reported byDoutt and Nakata (1973) and Kido et a (1984)The elevated capture rates were seen despite dif-ferences in geographical regions and variation inthe number of area-wide A epos captures whichindicates that the prune tree effect operated undera wide variety of vineyard management and envi-ronmental conditions Based on these results weconcluded that the presence of prune trees wasassociated with elevated A epos numbers and asa result parasites may colonize these vineyards athigher rates than vineyards lacking nearby pnmetrees
Ecological Mechanisms Doutt and Nakata(1973) and Kido et a (1984) assumed that anyincrease in parasite abundance in vineyards (in-ferred using trap data or egg parasitism rates) wasthe direct result of overwintering emergence fromnearby habitats In the current studywe tested thisassumption by comparing the density of overwin-tering A epos emergence holes with A epos trapcapture in pnme tree vineyards after controllingfor area-wide colonizers The significant regressionduring the 2nd sample period indicated prunetrees were a significant source of parasite capturesduring the period of A epos emergence from over-wintered E prunicola eggs These results reveal 2important points regarding the influence of pnmetrees on parasite abundance in vineyards The 1stis that overwintering emergence is a significantecological factor determining early-season parasite
Table 4 Effeet of irrigalion on prIme lree growlh mld populalions of E prunicola mId A epos
Growth rate No twigs A epos A epos tmppecI Elnmicolacm2n (SE) emergence holescm2 trapped
Irrigated 304 498 642 176 00325 00063 950 209 2216+ 5411Nonirrigated 131 420 1475 163 Om05 0058 110 193 059t 507
a Surface area of 1-yr-old pnmc twigs (mean t SE) Numbers of adults caught on traps within prune tree canopies
Signifkantly different 001 lt P lt 005 SigniRcantl) different P lt 001
NoA epos captures in control plots
8060
Vol 25 no 2
402000
00
ENVIRONMENTAL ENTOMOLOGY
0008 00 10 20 30 40 50
B c50 80
bull 40 bull bull 6030
4020
10 20
06
502
lQ-Q~ A~
08bull~=E 06Q= 04tIJ~ 02aQto~l~~~0Z
Fig 3 Scatterplots of the functional relationship between A epos capture in pnme tree refuge vineyardsagainstA epos capture in control vineyards (A) 15-18 April (B = 1705 975 CL L = 058 U = nonestimatable) (8) 24April-l May (B = 238 975 CL L = 205 U = 279) (C) 8--15 May (B = 310 975 CL L = 221 U = 485)Confidence limits were established for 975 as a correction for multiple comparisons
numbers The 2nd is that the effect of prune treeson A epos abundance was dependent not only onthe presence of prune trees themselves but on thedensity of overwintering parasites residing withinthe trees This latter point may have important im-plications for using prune trees as a commercialpest control tactic (discussed below)
Lewis and Stephenson (1966) showed that flyinginsects dispersing in the surrounding environmentaccumulate in sheltered regions near physical bar-riers (natural or artificial windbreaks) They alsofound that insects aggregate behind windbreaks asa multiplicative function of the number of insectsdispersing in the surrounding environment Thiseffect was particularly pronounced for mymaridwasps Using elemental labeling techniques Cor-bett and Rosenheim (in press) found a pattern ofA epos capture suggesting that area-wide A eposnumbers accumulate at a rate 4 times greater im-mediately downwind of prune trees than found up-wind of trees Based on these results they hypoth-esized that a windbreak effect may also be amechanism enhancing abundance within the prunetree-vineyard system We tested this hypothesisacross our replicated study by examining the func-tional relationship between control vineyard trapcaptures (area-wide colonizers in the surroundingenvironment) and prune tree vineyard captures(number of colonizers behind windbreaks) Our re-sults revealed that during each of the 3 sampleperiods the slope of the relationship was 217Thus a multiplicative increase in capture was as-sociated with the presence of prune trees but in-dependent of any contribution from overwinteringA epos densities This result supported the wind-break hypothesis and demonstrated the effect may
be consistent for vineyards with a windbreak pres-ent Furthermore this result identifies an impor-tant additional mechanism that may be affectingA epos vineyard colonization in this system
These results underscore the importance of in-vestigating the ecological mechanisms underlyingsystems of habitat diversification in addition totests of overall effectiveness of diversification Un-derstanding the mechanisms involved will enhanceour ability to manipulate these systems to maxi-mize their effectiveness and provide a usefulframework to begin analyses of new systems
Variation in the Prunc Tree Effect Becauseoverwintering A epos densities varied among theprune tree sites we were interested in how themanagement of pnme trees affects overwinteringA epos populations and their subsequent abun-dance in vineyards Irrigation practices were foundto be a significant factor cplaining much of thevariation in overwintering A epos densities ob-served in pnme trees (Table 4) and somc of thevariation in trap catches in pnme tree vineyardsThe number of prune trees within a site also af-fected overwintering A epos density How treenumber exerts an influence on the density of over-wintering A epos (per square centimeter of pnmebark) is not known but we speculate that greaternumbers of trees may create a more favorable mi-crohabitat increasing the number of A epos thatsuccessfully emerge from their overwinteringhosts
Although tree number had a strong positive in-fluence on the density of A epos emergence holesper square centimeter of prune tree bark it wasnot a predictor of A epos captures in pnme treevineyards This is an enigmatic result At the outset
April IHH6 MUHllIY ET 1 HABITAT DIVEHSIFICATlON FOH BIOLOGICAL CONTHOL 503
of till study we expected that pnllle tree sites con-taining morl trelS would producl a larger pool ofmigrants for thl downwind vineyards The in-enased density (pcr squangt centimeter of bark) ofovcrwintcring A epos in prune trce sites with larg-cr tne 1lI11l11wrsonly strengthened onr expcctationthat trcc numbpr would lw a major determinant ofcady-season abundance in prune trep vineyardsIndclltl wc expccted that we would bl able to es-timatp t]1( minimum numbPI of trees needed togenlrate the gtelof augmentation of A cpos nec-pssary to achieve pest management Instead whatwe found is that trpe number is less a predictor ofalmmlancp in vinpyards than is the number of AlJOS produced ppr centinIPter of bark We also ca]-cu]ahd thc total number of A epos emerged fromovclvintlring sitps vithin an cntire prune tree site(a composite cstimate obtained as the product ofA e]1os ltknsity pcr squarc ccntimetcr of bark thetotal arca plr tngtp and the number of trpes persitc) as t]IP estimatp for A pos contributions fromprunl trpc sites This estimatc was however awcakcr prcdictor of early-season abundance invincyards than Pitill l thc numbcr of A cpos persquarc ccntimctcr of bark or pcr prune tree Howcan we cxplain this resn]t One possibility is thattrees within the largcr prune trcc sites Illay havecontributed uncIlm]]y to the number of vineyardmigrants vith thc rows of trccs directly adjacentto the vincyard producing the bulk of the waspstrapped in vincyards Additional experimentationwill be needed to test this and other potential ex-plauations for the lack of a trec number effect
These results have 2 implications for establish-ing and managing pnnll tree rcfuges for pest con-trol The 1st is that cnhanccd A cpos abundancemay he attainable with a relatively small numberof tnps TIIP 2nd is that irrigation of pnllle treesvill bp of primary importance for supporting highdpnsitics of ovprwintcring i cpos populationsMon detail(gtd rpcommcndations for establishingpnme trcc rcfugps will be presented in future pa-pers
Questious about thc pffect of vegetational diver-sity on tIll diversity and abundance of insect pop-ulations ngtcenty have rcceived considerable atteu-tion from ecologists and pest control specialistsThe rpsu]ts of thp cnrrent study not only supportedtl( hypothdical effect of pnme tree refuges on Acpos abundance but in a broader context providea test of the effect of wgctational diversity on nat-ural encmy abundance As a result this study hassatisfied tIl( 1st of 3 hypothesized natural enemyresponses which pr((licts greater natura] enemyabundance in diversified systems (Bussell 1989Andow 1991) In addition 011study identified thefollOving 2 mpchanisms underlying the influenceof prune trees the presence of an ove1vinteringhost and a windbreak effpct
Future papers vill examine the effect of over-wintering refuges on the remaining 2 hypotheses-the effect of pnnw trees on E elegalltula egg mor-
taity and E elcgalltula population densities withingrape vineyards
Acknowledgments
We extend special thanks to Nerissa Fretman MaddySamant John Tlnnter Jason May and Paul Higgs (De-partment of Entomology University of Califol1lia Davis)for field collections and sample processing Vc also ac-Irnowledge thc assistancc of Hobert Hoy Stan Grant andBrad Coleman (E amp J Gallo Winery Livingston CAl forlogistical and material support This research was fuudedby the Westem Hegional Integrated Pest Managementprogram the California Department of Ptsticidt Hegu-lation and the Locli-Woodbridge vVinegrapt Commis-sion This research was part of BGMs doctoral disser-tation at the University of California at Davis
References Cited
Altieri M A 1992 Agroecological foundatious of al-temative agliculture in Califomia Agric Ecosyst En-viron 39 23-53
Andow D A 1991 Vegetational diversity and artll1o-pod population response Annu Hev Entomol 36561-586
Corbett A und J A Rosenheim 1996 Impact of anatural enemy overwintering refuge and its iuterac-tion vith the surrounding landscape Ecol Entomol(in press)
Doutt R L and J Nakat bullbull 1965 Overwintering ref-uge of Anagrus epos (Hymenoptera Mymaridae) JEcon Entomol 58 586
1973 The Ruhus leafllOpper and its egg parasitoid Anendemic biotic system useful in grape-pest manage-ment Environ Entomol 2 381~186
Flaherty D L L T Wilson V M Stern and IIKido 1985 Biologicalcontrol in San Joaquin Valleyvineyards pp 501-520 Tn D G Herzog and M AHoy [eds] Biological control of agricnltural IPM sys-tems Viley New York
Flint M L and P A Roberts 1988 Using cropdiversity to manage pest problems some Californiaexamples Am J Altern Agric1 163-167
Gorclh G lIml D M Dunlmr 1977 A new Allagmsimportant in the biological control of Stephanitis tak-eljai and a key to the N01ih American species FlaEntomol 60 85-95
Herzo~ D c and J E Funderburk 1985 Plantresistance and cultural practiee interactious with hio-logical control pp 67-88 Til D G Ilerzog and MA Hoy [eds] Biological control of agliclllInral IPMsystems Wiley New York
Kiclo H D L Flaherty D F 80sh Dlul K A Val-ero 1984 French pmne trees as ovclwiuteringsites for the grape leafllOpper egg parasite Am JEnol Vitie 35 156-160
Letourneau D K 1987 The enemies hypothesis Tri-trophic interactions and vegetational diversity in trop-ical agroecosystel11sEcology 68 1616-1622
Lewis T bullbulllid J W Stephenson 1966 The penne-ability of artificial windbreaks and the distribution offlying insects in the leeward shelttred Will AnnAppl BioI 58 355-363
Mulla M S 1957 The biology of Typhlo(fhapnmi-cola (Edwards) and 1 quercus (Fabricius) (Cicadelli-dae lIomoptera) Ann Entomol SOlAm 50 76-87
504 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
National Research Council 1989 Alternative agri-culture National AcademyWashington DC
PaAel M D and P H Harvey 1988 Recent de-velopments in the analysis of comparative data QRev BioI 63 418-440
Pickett C H L T Wilson and D L Flaherty1990 The role of refuges in crop protection withreference to plantings of French prune trees in agrape agroecosystem pp 151-165 In N J BostanianL T Wilson and T J Dennehy [eds] Monitoringand integrated management of arthropod pests ofsmall fruit crops Intercept Andover
RusseD E P 1989 Enemies hypothesis A review ofthe effect of vegetational diversity on predatory in-sects and parasitoids Environ Entomol 18 590-599
Sokal R R and F J Rohlf 1981 Biometry 2nded Freeman San Francisco
SAS Institute 1989 JMP statistical program SAS In-stitute Cary NC
van Emden H F 1990 Plant diversity and naturalenemy efficiency in agroecosystems pp 63-80In MMackauer L E Ehler and J Roland [eds] Criticalissues in biological control Intercept Andover UK
Williams D W 1984 Ecology of the blackberry-leaf-hopper-parasite system and its relevance to Californiagrape agroecosystems Hilgardia 52 1--33
Wilson L T C H Pickett and D L Flaherty1989 French prune trees refuge for grape leafhop-per parasite Calif Agric 43(2) 7-8
Winkler A J J A Cook W M Kliewer and L ALider 1974 Climate and soils pp 58-75 In Gen-eral viticulture University of California Press Berke-ley
Received for publication 5 December 1994 accepted 6November 1995

500 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
Table 1 Nested ANOVA of the number of A epos(101110 trw1sformed) captured on traps in incyards withresl)ect to prune refuges growing regions and pairedvineyard plots (block effcct)
Date Source of df F 1variation
15-18 Apr Treahnent 1 393 0094Blocks (region) 5 617 0023Region 1 013 0735
24 Apr-l May Treatment 1 647 0032Blocks (region) 8 514 0012Region 1 460 0061
8-15 May Treahnent 1 1132 0006Blocks (region) 10 572 0004Begion 1 356 0086
One-tailed tests for hypothesis testing l lt clitical valueafter Bonferroni correction (1 lt 0033)
P = 0001 respectively) but not for the 1st sampledate (R2 = 048 n = 7 P = 009) The significantregressions of A epos capture between the treat-ment and control plots underlines the importanceof blocking by vineyard pair a key feature of ourexperimental design This result also confirms thatarea-wide A epos numbers were a significantsource of parasites in vineyards whether pnmetrees were present or not
The multiple regression analyses indicated thatA epos emergence from prunes was a factor ex-plaining A epos trap captures in prune tree vine-yards after controlling for area-wide abundancefrom control plot trap captures during the 2ndsample period (Table 2) This coincided with theestimated time of A epos emergence from prunetwigs as determined by sticky traps in pnme treesThe partial regression coefficients indicated thatemergence from pnmes explained a substantialamount of the variation in A epos capture duringthe first 2 sample periods in April but by the 3rdsample period in early May area-wide A epos cap-tures explained most variation in trap captures
Variation in Overwintering A epos DensitiesPrune tree sites varied in terms of the number oftrees per site the rate of tree growth the numberof I-yr-old twigs per tree and the density of Aepos emergence holes (Table 3) Much of this vari-
ation was generated by variation in irrigation prac-tices (Table 4) Irrigated trees were associated withgreater tree growth (average surface area of I-yr-old twigs) and lower twig numbers than nonirri-gated trees Irrigation was also associated withhigher A epos overwintering densities as well ashigher trap captures of E prunicola and A eposadults in tree canopies ANCOVA revealed thatboth irrigation (F = 2306 df = 1 12 P = 0001)and prune tree number (F = 1121 df = 1 12 P= 0009) had significant influences on the densityof A epos emergence holes in prune trees Therewas a positive linear relationship between thenumber of prune trees and the density of A eposemergence holes in trees (intercept = -00496slope = 00773) Nevertheless neither pnme treenumber nor irrigation had detectable effects onparasite catch in associated vineyards during the 3sample periods (P gt 0047)
We also estimated the average number of Aepos emergence holes per tree and the total num-ber of emergence holes per prune tree site to testif these parameters could explain captures in ad-jacent vineyards The number of emergence holesper tree was nonsignificant during the 1st sampleperiod (F = 89 df = 1 6 P = 0041) significantduring the 2nd period (F = 721 df = 1 9 P =0031) and non-significant during the 3rd period(F = 026 df = 1 11 P = 0625) The numberper prune tree site was however nonsignificantduring all 3 sample periods (P ~ 0096)
Multiplicative Effects The principal axis anal-yses (model II regression) of A epos trap capturesin control and prune tree vineyards suggest that amultiplicative effect was present during the 2ndand 3rd sample periods when both slope param-eters were significantly gt10 (Fig 3 B and C)During the 1st sample period the estimated slopewas similar to the other sample periods but notsignificantlydifferent from 10 (Fig 3A) These re-sults suggest that independent of any contributionfrom A epos overwintering in pmne trees prunetrees were associated with a multiplicative effectaugmenting A epos capture on the downwind sideof prune trees
Table 2 Multiple regression analysis of the relationship bctwcen the dependent variable 10fllO (A epos trap bullatch)ill prune trec plots and the oulcpendent variable sluarc root (A epos emergence density) from prlUlC sites and10gIO (A epos trap catch) 01 control plots
Sample date Source of variation df Slope (1 SE) F R2 pb
15--18 Apr Whole model test 2 7 119 086 0026Emergence density 17 106 (~ 0325) 106 075 0031Control captu res 17 067 (10237) 80 069 0047
24 Apr-l May Whole model test 210 168 083 0002Emergence density 110 237(1 0717) 109 073 0013Control captures 110 084 (10242) 119 075 OoIl
8-]5 May Whole model test 2 12 1015 069 0005E mergence density 1 19- 091 (10825) 121 035 0299Control captures 112 148 (~ 0363) 1674 081 0003
H2 for individual factors are partial correlation coefficients (r2)b l lt clitical value after Bonferroni correction (1 lt 0017)
April 1996 MURPHY ET AL HABITAT DIVERSIFICATION FOR BIOLOGICAL CONTROL 501
Table 3 Descrilltive larmnelers of pnnle lrees mnong experimenlalllols
Variable
No pnllle trees per siteNo 1-yr-old twigs per treel-yr-old twig surface area cm2
A (--pOS tlllfgtrgencp densitycm2
Mean
422511217
2030021
SE
11191774
4000056
Range
27-1216435-223837-1225
00023-00640
Discllssion
The goal of this research was to assess the influ-ence of pnme trees on early-season A epos abun-dance in vineyards and to test the hypothesizedecological mechanisms underlying this systemPomp trees were associated with significant in-creases in the density of A epos captured in nearbygrape vineyards relative to vineyards lacking prunetrees Pome tree vineyards were found on aver-agp to receive twice the number of early-seasonparasite captures Overwintering A epos emer-gpnce from prune trees was a significant predictorof A cpos trap capture in nearby vineyards duringthe 2nd sample period which coincides with ourestimated period of A epos emergence from prunetwigs Examination of the functional relationshipbetween control and pnme tree vineyard parasitecaptures also revealed a multiplicative response intrap capture on the downwind side of pnme treevineyard plots This suggests that another factorassociated with pnme trees other than overwin-tering A epos populations enhanced parasitenumhtrs in refuge vineyards
Seasonal Patterns Based on limited field dataDoutt and Nakata (1973) and Kido et al (1984)speculated that the effectiveness of overwinteringrefuges near vineyards was at least in part causedby the time of A epos emergence from overwin-tering hosts They based this hypothesis on the ob-senation that parasite emergence from prune treesor blackberries occurs during mid-April at the ap-proximate time overwintered E elegantula beginoviposition in vimmiddotyards They further speculatedthat enhancing early-season A epos colonizerscoupled with the high numerical response of theparasite (A epos completes 3 generations for eachE d(~antllia generation) improves biological con-trol of leafhoppers in vineyards The seasonal pat-tern of leafllOppers and parasites seen in the cur-rent study concurs with these previousobservations Across the replicated vineyard pairsour data show a consistent pattern of both area-
wide A epos captures and A epos emergence fromprune trees coinciding with the onset of oviposi-tion by overwintered E elegantula in vineyardsThus parasite abundance appears to be synchro-nized with the appearance of host eggs in grapevineyards
Vineyard A epos Abundance The replicatedcomparisons of A epos abundance in vineyardsshowed that the presence of overwintering habitatsproduced a substantial early-season increase in thedensity of A epos trapped in grape vineyards aresult consistent with the observations reported byDoutt and Nakata (1973) and Kido et a (1984)The elevated capture rates were seen despite dif-ferences in geographical regions and variation inthe number of area-wide A epos captures whichindicates that the prune tree effect operated undera wide variety of vineyard management and envi-ronmental conditions Based on these results weconcluded that the presence of prune trees wasassociated with elevated A epos numbers and asa result parasites may colonize these vineyards athigher rates than vineyards lacking nearby pnmetrees
Ecological Mechanisms Doutt and Nakata(1973) and Kido et a (1984) assumed that anyincrease in parasite abundance in vineyards (in-ferred using trap data or egg parasitism rates) wasthe direct result of overwintering emergence fromnearby habitats In the current studywe tested thisassumption by comparing the density of overwin-tering A epos emergence holes with A epos trapcapture in pnme tree vineyards after controllingfor area-wide colonizers The significant regressionduring the 2nd sample period indicated prunetrees were a significant source of parasite capturesduring the period of A epos emergence from over-wintered E prunicola eggs These results reveal 2important points regarding the influence of pnmetrees on parasite abundance in vineyards The 1stis that overwintering emergence is a significantecological factor determining early-season parasite
Table 4 Effeet of irrigalion on prIme lree growlh mld populalions of E prunicola mId A epos
Growth rate No twigs A epos A epos tmppecI Elnmicolacm2n (SE) emergence holescm2 trapped
Irrigated 304 498 642 176 00325 00063 950 209 2216+ 5411Nonirrigated 131 420 1475 163 Om05 0058 110 193 059t 507
a Surface area of 1-yr-old pnmc twigs (mean t SE) Numbers of adults caught on traps within prune tree canopies
Signifkantly different 001 lt P lt 005 SigniRcantl) different P lt 001
NoA epos captures in control plots
8060
Vol 25 no 2
402000
00
ENVIRONMENTAL ENTOMOLOGY
0008 00 10 20 30 40 50
B c50 80
bull 40 bull bull 6030
4020
10 20
06
502
lQ-Q~ A~
08bull~=E 06Q= 04tIJ~ 02aQto~l~~~0Z
Fig 3 Scatterplots of the functional relationship between A epos capture in pnme tree refuge vineyardsagainstA epos capture in control vineyards (A) 15-18 April (B = 1705 975 CL L = 058 U = nonestimatable) (8) 24April-l May (B = 238 975 CL L = 205 U = 279) (C) 8--15 May (B = 310 975 CL L = 221 U = 485)Confidence limits were established for 975 as a correction for multiple comparisons
numbers The 2nd is that the effect of prune treeson A epos abundance was dependent not only onthe presence of prune trees themselves but on thedensity of overwintering parasites residing withinthe trees This latter point may have important im-plications for using prune trees as a commercialpest control tactic (discussed below)
Lewis and Stephenson (1966) showed that flyinginsects dispersing in the surrounding environmentaccumulate in sheltered regions near physical bar-riers (natural or artificial windbreaks) They alsofound that insects aggregate behind windbreaks asa multiplicative function of the number of insectsdispersing in the surrounding environment Thiseffect was particularly pronounced for mymaridwasps Using elemental labeling techniques Cor-bett and Rosenheim (in press) found a pattern ofA epos capture suggesting that area-wide A eposnumbers accumulate at a rate 4 times greater im-mediately downwind of prune trees than found up-wind of trees Based on these results they hypoth-esized that a windbreak effect may also be amechanism enhancing abundance within the prunetree-vineyard system We tested this hypothesisacross our replicated study by examining the func-tional relationship between control vineyard trapcaptures (area-wide colonizers in the surroundingenvironment) and prune tree vineyard captures(number of colonizers behind windbreaks) Our re-sults revealed that during each of the 3 sampleperiods the slope of the relationship was 217Thus a multiplicative increase in capture was as-sociated with the presence of prune trees but in-dependent of any contribution from overwinteringA epos densities This result supported the wind-break hypothesis and demonstrated the effect may
be consistent for vineyards with a windbreak pres-ent Furthermore this result identifies an impor-tant additional mechanism that may be affectingA epos vineyard colonization in this system
These results underscore the importance of in-vestigating the ecological mechanisms underlyingsystems of habitat diversification in addition totests of overall effectiveness of diversification Un-derstanding the mechanisms involved will enhanceour ability to manipulate these systems to maxi-mize their effectiveness and provide a usefulframework to begin analyses of new systems
Variation in the Prunc Tree Effect Becauseoverwintering A epos densities varied among theprune tree sites we were interested in how themanagement of pnme trees affects overwinteringA epos populations and their subsequent abun-dance in vineyards Irrigation practices were foundto be a significant factor cplaining much of thevariation in overwintering A epos densities ob-served in pnme trees (Table 4) and somc of thevariation in trap catches in pnme tree vineyardsThe number of prune trees within a site also af-fected overwintering A epos density How treenumber exerts an influence on the density of over-wintering A epos (per square centimeter of pnmebark) is not known but we speculate that greaternumbers of trees may create a more favorable mi-crohabitat increasing the number of A epos thatsuccessfully emerge from their overwinteringhosts
Although tree number had a strong positive in-fluence on the density of A epos emergence holesper square centimeter of prune tree bark it wasnot a predictor of A epos captures in pnme treevineyards This is an enigmatic result At the outset
April IHH6 MUHllIY ET 1 HABITAT DIVEHSIFICATlON FOH BIOLOGICAL CONTHOL 503
of till study we expected that pnllle tree sites con-taining morl trelS would producl a larger pool ofmigrants for thl downwind vineyards The in-enased density (pcr squangt centimeter of bark) ofovcrwintcring A epos in prune trce sites with larg-cr tne 1lI11l11wrsonly strengthened onr expcctationthat trcc numbpr would lw a major determinant ofcady-season abundance in prune trep vineyardsIndclltl wc expccted that we would bl able to es-timatp t]1( minimum numbPI of trees needed togenlrate the gtelof augmentation of A cpos nec-pssary to achieve pest management Instead whatwe found is that trpe number is less a predictor ofalmmlancp in vinpyards than is the number of AlJOS produced ppr centinIPter of bark We also ca]-cu]ahd thc total number of A epos emerged fromovclvintlring sitps vithin an cntire prune tree site(a composite cstimate obtained as the product ofA e]1os ltknsity pcr squarc ccntimetcr of bark thetotal arca plr tngtp and the number of trpes persitc) as t]IP estimatp for A pos contributions fromprunl trpc sites This estimatc was however awcakcr prcdictor of early-season abundance invincyards than Pitill l thc numbcr of A cpos persquarc ccntimctcr of bark or pcr prune tree Howcan we cxplain this resn]t One possibility is thattrees within the largcr prune trcc sites Illay havecontributed uncIlm]]y to the number of vineyardmigrants vith thc rows of trccs directly adjacentto the vincyard producing the bulk of the waspstrapped in vincyards Additional experimentationwill be needed to test this and other potential ex-plauations for the lack of a trec number effect
These results have 2 implications for establish-ing and managing pnnll tree rcfuges for pest con-trol The 1st is that cnhanccd A cpos abundancemay he attainable with a relatively small numberof tnps TIIP 2nd is that irrigation of pnllle treesvill bp of primary importance for supporting highdpnsitics of ovprwintcring i cpos populationsMon detail(gtd rpcommcndations for establishingpnme trcc rcfugps will be presented in future pa-pers
Questious about thc pffect of vegetational diver-sity on tIll diversity and abundance of insect pop-ulations ngtcenty have rcceived considerable atteu-tion from ecologists and pest control specialistsThe rpsu]ts of thp cnrrent study not only supportedtl( hypothdical effect of pnme tree refuges on Acpos abundance but in a broader context providea test of the effect of wgctational diversity on nat-ural encmy abundance As a result this study hassatisfied tIl( 1st of 3 hypothesized natural enemyresponses which pr((licts greater natura] enemyabundance in diversified systems (Bussell 1989Andow 1991) In addition 011study identified thefollOving 2 mpchanisms underlying the influenceof prune trees the presence of an ove1vinteringhost and a windbreak effpct
Future papers vill examine the effect of over-wintering refuges on the remaining 2 hypotheses-the effect of pnnw trees on E elegalltula egg mor-
taity and E elcgalltula population densities withingrape vineyards
Acknowledgments
We extend special thanks to Nerissa Fretman MaddySamant John Tlnnter Jason May and Paul Higgs (De-partment of Entomology University of Califol1lia Davis)for field collections and sample processing Vc also ac-Irnowledge thc assistancc of Hobert Hoy Stan Grant andBrad Coleman (E amp J Gallo Winery Livingston CAl forlogistical and material support This research was fuudedby the Westem Hegional Integrated Pest Managementprogram the California Department of Ptsticidt Hegu-lation and the Locli-Woodbridge vVinegrapt Commis-sion This research was part of BGMs doctoral disser-tation at the University of California at Davis
References Cited
Altieri M A 1992 Agroecological foundatious of al-temative agliculture in Califomia Agric Ecosyst En-viron 39 23-53
Andow D A 1991 Vegetational diversity and artll1o-pod population response Annu Hev Entomol 36561-586
Corbett A und J A Rosenheim 1996 Impact of anatural enemy overwintering refuge and its iuterac-tion vith the surrounding landscape Ecol Entomol(in press)
Doutt R L and J Nakat bullbull 1965 Overwintering ref-uge of Anagrus epos (Hymenoptera Mymaridae) JEcon Entomol 58 586
1973 The Ruhus leafllOpper and its egg parasitoid Anendemic biotic system useful in grape-pest manage-ment Environ Entomol 2 381~186
Flaherty D L L T Wilson V M Stern and IIKido 1985 Biologicalcontrol in San Joaquin Valleyvineyards pp 501-520 Tn D G Herzog and M AHoy [eds] Biological control of agricnltural IPM sys-tems Viley New York
Flint M L and P A Roberts 1988 Using cropdiversity to manage pest problems some Californiaexamples Am J Altern Agric1 163-167
Gorclh G lIml D M Dunlmr 1977 A new Allagmsimportant in the biological control of Stephanitis tak-eljai and a key to the N01ih American species FlaEntomol 60 85-95
Herzo~ D c and J E Funderburk 1985 Plantresistance and cultural practiee interactious with hio-logical control pp 67-88 Til D G Ilerzog and MA Hoy [eds] Biological control of agliclllInral IPMsystems Wiley New York
Kiclo H D L Flaherty D F 80sh Dlul K A Val-ero 1984 French pmne trees as ovclwiuteringsites for the grape leafllOpper egg parasite Am JEnol Vitie 35 156-160
Letourneau D K 1987 The enemies hypothesis Tri-trophic interactions and vegetational diversity in trop-ical agroecosystel11sEcology 68 1616-1622
Lewis T bullbulllid J W Stephenson 1966 The penne-ability of artificial windbreaks and the distribution offlying insects in the leeward shelttred Will AnnAppl BioI 58 355-363
Mulla M S 1957 The biology of Typhlo(fhapnmi-cola (Edwards) and 1 quercus (Fabricius) (Cicadelli-dae lIomoptera) Ann Entomol SOlAm 50 76-87
504 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
National Research Council 1989 Alternative agri-culture National AcademyWashington DC
PaAel M D and P H Harvey 1988 Recent de-velopments in the analysis of comparative data QRev BioI 63 418-440
Pickett C H L T Wilson and D L Flaherty1990 The role of refuges in crop protection withreference to plantings of French prune trees in agrape agroecosystem pp 151-165 In N J BostanianL T Wilson and T J Dennehy [eds] Monitoringand integrated management of arthropod pests ofsmall fruit crops Intercept Andover
RusseD E P 1989 Enemies hypothesis A review ofthe effect of vegetational diversity on predatory in-sects and parasitoids Environ Entomol 18 590-599
Sokal R R and F J Rohlf 1981 Biometry 2nded Freeman San Francisco
SAS Institute 1989 JMP statistical program SAS In-stitute Cary NC
van Emden H F 1990 Plant diversity and naturalenemy efficiency in agroecosystems pp 63-80In MMackauer L E Ehler and J Roland [eds] Criticalissues in biological control Intercept Andover UK
Williams D W 1984 Ecology of the blackberry-leaf-hopper-parasite system and its relevance to Californiagrape agroecosystems Hilgardia 52 1--33
Wilson L T C H Pickett and D L Flaherty1989 French prune trees refuge for grape leafhop-per parasite Calif Agric 43(2) 7-8
Winkler A J J A Cook W M Kliewer and L ALider 1974 Climate and soils pp 58-75 In Gen-eral viticulture University of California Press Berke-ley
Received for publication 5 December 1994 accepted 6November 1995

April 1996 MURPHY ET AL HABITAT DIVERSIFICATION FOR BIOLOGICAL CONTROL 501
Table 3 Descrilltive larmnelers of pnnle lrees mnong experimenlalllols
Variable
No pnllle trees per siteNo 1-yr-old twigs per treel-yr-old twig surface area cm2
A (--pOS tlllfgtrgencp densitycm2
Mean
422511217
2030021
SE
11191774
4000056
Range
27-1216435-223837-1225
00023-00640
Discllssion
The goal of this research was to assess the influ-ence of pnme trees on early-season A epos abun-dance in vineyards and to test the hypothesizedecological mechanisms underlying this systemPomp trees were associated with significant in-creases in the density of A epos captured in nearbygrape vineyards relative to vineyards lacking prunetrees Pome tree vineyards were found on aver-agp to receive twice the number of early-seasonparasite captures Overwintering A epos emer-gpnce from prune trees was a significant predictorof A cpos trap capture in nearby vineyards duringthe 2nd sample period which coincides with ourestimated period of A epos emergence from prunetwigs Examination of the functional relationshipbetween control and pnme tree vineyard parasitecaptures also revealed a multiplicative response intrap capture on the downwind side of pnme treevineyard plots This suggests that another factorassociated with pnme trees other than overwin-tering A epos populations enhanced parasitenumhtrs in refuge vineyards
Seasonal Patterns Based on limited field dataDoutt and Nakata (1973) and Kido et al (1984)speculated that the effectiveness of overwinteringrefuges near vineyards was at least in part causedby the time of A epos emergence from overwin-tering hosts They based this hypothesis on the ob-senation that parasite emergence from prune treesor blackberries occurs during mid-April at the ap-proximate time overwintered E elegantula beginoviposition in vimmiddotyards They further speculatedthat enhancing early-season A epos colonizerscoupled with the high numerical response of theparasite (A epos completes 3 generations for eachE d(~antllia generation) improves biological con-trol of leafhoppers in vineyards The seasonal pat-tern of leafllOppers and parasites seen in the cur-rent study concurs with these previousobservations Across the replicated vineyard pairsour data show a consistent pattern of both area-
wide A epos captures and A epos emergence fromprune trees coinciding with the onset of oviposi-tion by overwintered E elegantula in vineyardsThus parasite abundance appears to be synchro-nized with the appearance of host eggs in grapevineyards
Vineyard A epos Abundance The replicatedcomparisons of A epos abundance in vineyardsshowed that the presence of overwintering habitatsproduced a substantial early-season increase in thedensity of A epos trapped in grape vineyards aresult consistent with the observations reported byDoutt and Nakata (1973) and Kido et a (1984)The elevated capture rates were seen despite dif-ferences in geographical regions and variation inthe number of area-wide A epos captures whichindicates that the prune tree effect operated undera wide variety of vineyard management and envi-ronmental conditions Based on these results weconcluded that the presence of prune trees wasassociated with elevated A epos numbers and asa result parasites may colonize these vineyards athigher rates than vineyards lacking nearby pnmetrees
Ecological Mechanisms Doutt and Nakata(1973) and Kido et a (1984) assumed that anyincrease in parasite abundance in vineyards (in-ferred using trap data or egg parasitism rates) wasthe direct result of overwintering emergence fromnearby habitats In the current studywe tested thisassumption by comparing the density of overwin-tering A epos emergence holes with A epos trapcapture in pnme tree vineyards after controllingfor area-wide colonizers The significant regressionduring the 2nd sample period indicated prunetrees were a significant source of parasite capturesduring the period of A epos emergence from over-wintered E prunicola eggs These results reveal 2important points regarding the influence of pnmetrees on parasite abundance in vineyards The 1stis that overwintering emergence is a significantecological factor determining early-season parasite
Table 4 Effeet of irrigalion on prIme lree growlh mld populalions of E prunicola mId A epos
Growth rate No twigs A epos A epos tmppecI Elnmicolacm2n (SE) emergence holescm2 trapped
Irrigated 304 498 642 176 00325 00063 950 209 2216+ 5411Nonirrigated 131 420 1475 163 Om05 0058 110 193 059t 507
a Surface area of 1-yr-old pnmc twigs (mean t SE) Numbers of adults caught on traps within prune tree canopies
Signifkantly different 001 lt P lt 005 SigniRcantl) different P lt 001
NoA epos captures in control plots
8060
Vol 25 no 2
402000
00
ENVIRONMENTAL ENTOMOLOGY
0008 00 10 20 30 40 50
B c50 80
bull 40 bull bull 6030
4020
10 20
06
502
lQ-Q~ A~
08bull~=E 06Q= 04tIJ~ 02aQto~l~~~0Z
Fig 3 Scatterplots of the functional relationship between A epos capture in pnme tree refuge vineyardsagainstA epos capture in control vineyards (A) 15-18 April (B = 1705 975 CL L = 058 U = nonestimatable) (8) 24April-l May (B = 238 975 CL L = 205 U = 279) (C) 8--15 May (B = 310 975 CL L = 221 U = 485)Confidence limits were established for 975 as a correction for multiple comparisons
numbers The 2nd is that the effect of prune treeson A epos abundance was dependent not only onthe presence of prune trees themselves but on thedensity of overwintering parasites residing withinthe trees This latter point may have important im-plications for using prune trees as a commercialpest control tactic (discussed below)
Lewis and Stephenson (1966) showed that flyinginsects dispersing in the surrounding environmentaccumulate in sheltered regions near physical bar-riers (natural or artificial windbreaks) They alsofound that insects aggregate behind windbreaks asa multiplicative function of the number of insectsdispersing in the surrounding environment Thiseffect was particularly pronounced for mymaridwasps Using elemental labeling techniques Cor-bett and Rosenheim (in press) found a pattern ofA epos capture suggesting that area-wide A eposnumbers accumulate at a rate 4 times greater im-mediately downwind of prune trees than found up-wind of trees Based on these results they hypoth-esized that a windbreak effect may also be amechanism enhancing abundance within the prunetree-vineyard system We tested this hypothesisacross our replicated study by examining the func-tional relationship between control vineyard trapcaptures (area-wide colonizers in the surroundingenvironment) and prune tree vineyard captures(number of colonizers behind windbreaks) Our re-sults revealed that during each of the 3 sampleperiods the slope of the relationship was 217Thus a multiplicative increase in capture was as-sociated with the presence of prune trees but in-dependent of any contribution from overwinteringA epos densities This result supported the wind-break hypothesis and demonstrated the effect may
be consistent for vineyards with a windbreak pres-ent Furthermore this result identifies an impor-tant additional mechanism that may be affectingA epos vineyard colonization in this system
These results underscore the importance of in-vestigating the ecological mechanisms underlyingsystems of habitat diversification in addition totests of overall effectiveness of diversification Un-derstanding the mechanisms involved will enhanceour ability to manipulate these systems to maxi-mize their effectiveness and provide a usefulframework to begin analyses of new systems
Variation in the Prunc Tree Effect Becauseoverwintering A epos densities varied among theprune tree sites we were interested in how themanagement of pnme trees affects overwinteringA epos populations and their subsequent abun-dance in vineyards Irrigation practices were foundto be a significant factor cplaining much of thevariation in overwintering A epos densities ob-served in pnme trees (Table 4) and somc of thevariation in trap catches in pnme tree vineyardsThe number of prune trees within a site also af-fected overwintering A epos density How treenumber exerts an influence on the density of over-wintering A epos (per square centimeter of pnmebark) is not known but we speculate that greaternumbers of trees may create a more favorable mi-crohabitat increasing the number of A epos thatsuccessfully emerge from their overwinteringhosts
Although tree number had a strong positive in-fluence on the density of A epos emergence holesper square centimeter of prune tree bark it wasnot a predictor of A epos captures in pnme treevineyards This is an enigmatic result At the outset
April IHH6 MUHllIY ET 1 HABITAT DIVEHSIFICATlON FOH BIOLOGICAL CONTHOL 503
of till study we expected that pnllle tree sites con-taining morl trelS would producl a larger pool ofmigrants for thl downwind vineyards The in-enased density (pcr squangt centimeter of bark) ofovcrwintcring A epos in prune trce sites with larg-cr tne 1lI11l11wrsonly strengthened onr expcctationthat trcc numbpr would lw a major determinant ofcady-season abundance in prune trep vineyardsIndclltl wc expccted that we would bl able to es-timatp t]1( minimum numbPI of trees needed togenlrate the gtelof augmentation of A cpos nec-pssary to achieve pest management Instead whatwe found is that trpe number is less a predictor ofalmmlancp in vinpyards than is the number of AlJOS produced ppr centinIPter of bark We also ca]-cu]ahd thc total number of A epos emerged fromovclvintlring sitps vithin an cntire prune tree site(a composite cstimate obtained as the product ofA e]1os ltknsity pcr squarc ccntimetcr of bark thetotal arca plr tngtp and the number of trpes persitc) as t]IP estimatp for A pos contributions fromprunl trpc sites This estimatc was however awcakcr prcdictor of early-season abundance invincyards than Pitill l thc numbcr of A cpos persquarc ccntimctcr of bark or pcr prune tree Howcan we cxplain this resn]t One possibility is thattrees within the largcr prune trcc sites Illay havecontributed uncIlm]]y to the number of vineyardmigrants vith thc rows of trccs directly adjacentto the vincyard producing the bulk of the waspstrapped in vincyards Additional experimentationwill be needed to test this and other potential ex-plauations for the lack of a trec number effect
These results have 2 implications for establish-ing and managing pnnll tree rcfuges for pest con-trol The 1st is that cnhanccd A cpos abundancemay he attainable with a relatively small numberof tnps TIIP 2nd is that irrigation of pnllle treesvill bp of primary importance for supporting highdpnsitics of ovprwintcring i cpos populationsMon detail(gtd rpcommcndations for establishingpnme trcc rcfugps will be presented in future pa-pers
Questious about thc pffect of vegetational diver-sity on tIll diversity and abundance of insect pop-ulations ngtcenty have rcceived considerable atteu-tion from ecologists and pest control specialistsThe rpsu]ts of thp cnrrent study not only supportedtl( hypothdical effect of pnme tree refuges on Acpos abundance but in a broader context providea test of the effect of wgctational diversity on nat-ural encmy abundance As a result this study hassatisfied tIl( 1st of 3 hypothesized natural enemyresponses which pr((licts greater natura] enemyabundance in diversified systems (Bussell 1989Andow 1991) In addition 011study identified thefollOving 2 mpchanisms underlying the influenceof prune trees the presence of an ove1vinteringhost and a windbreak effpct
Future papers vill examine the effect of over-wintering refuges on the remaining 2 hypotheses-the effect of pnnw trees on E elegalltula egg mor-
taity and E elcgalltula population densities withingrape vineyards
Acknowledgments
We extend special thanks to Nerissa Fretman MaddySamant John Tlnnter Jason May and Paul Higgs (De-partment of Entomology University of Califol1lia Davis)for field collections and sample processing Vc also ac-Irnowledge thc assistancc of Hobert Hoy Stan Grant andBrad Coleman (E amp J Gallo Winery Livingston CAl forlogistical and material support This research was fuudedby the Westem Hegional Integrated Pest Managementprogram the California Department of Ptsticidt Hegu-lation and the Locli-Woodbridge vVinegrapt Commis-sion This research was part of BGMs doctoral disser-tation at the University of California at Davis
References Cited
Altieri M A 1992 Agroecological foundatious of al-temative agliculture in Califomia Agric Ecosyst En-viron 39 23-53
Andow D A 1991 Vegetational diversity and artll1o-pod population response Annu Hev Entomol 36561-586
Corbett A und J A Rosenheim 1996 Impact of anatural enemy overwintering refuge and its iuterac-tion vith the surrounding landscape Ecol Entomol(in press)
Doutt R L and J Nakat bullbull 1965 Overwintering ref-uge of Anagrus epos (Hymenoptera Mymaridae) JEcon Entomol 58 586
1973 The Ruhus leafllOpper and its egg parasitoid Anendemic biotic system useful in grape-pest manage-ment Environ Entomol 2 381~186
Flaherty D L L T Wilson V M Stern and IIKido 1985 Biologicalcontrol in San Joaquin Valleyvineyards pp 501-520 Tn D G Herzog and M AHoy [eds] Biological control of agricnltural IPM sys-tems Viley New York
Flint M L and P A Roberts 1988 Using cropdiversity to manage pest problems some Californiaexamples Am J Altern Agric1 163-167
Gorclh G lIml D M Dunlmr 1977 A new Allagmsimportant in the biological control of Stephanitis tak-eljai and a key to the N01ih American species FlaEntomol 60 85-95
Herzo~ D c and J E Funderburk 1985 Plantresistance and cultural practiee interactious with hio-logical control pp 67-88 Til D G Ilerzog and MA Hoy [eds] Biological control of agliclllInral IPMsystems Wiley New York
Kiclo H D L Flaherty D F 80sh Dlul K A Val-ero 1984 French pmne trees as ovclwiuteringsites for the grape leafllOpper egg parasite Am JEnol Vitie 35 156-160
Letourneau D K 1987 The enemies hypothesis Tri-trophic interactions and vegetational diversity in trop-ical agroecosystel11sEcology 68 1616-1622
Lewis T bullbulllid J W Stephenson 1966 The penne-ability of artificial windbreaks and the distribution offlying insects in the leeward shelttred Will AnnAppl BioI 58 355-363
Mulla M S 1957 The biology of Typhlo(fhapnmi-cola (Edwards) and 1 quercus (Fabricius) (Cicadelli-dae lIomoptera) Ann Entomol SOlAm 50 76-87
504 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
National Research Council 1989 Alternative agri-culture National AcademyWashington DC
PaAel M D and P H Harvey 1988 Recent de-velopments in the analysis of comparative data QRev BioI 63 418-440
Pickett C H L T Wilson and D L Flaherty1990 The role of refuges in crop protection withreference to plantings of French prune trees in agrape agroecosystem pp 151-165 In N J BostanianL T Wilson and T J Dennehy [eds] Monitoringand integrated management of arthropod pests ofsmall fruit crops Intercept Andover
RusseD E P 1989 Enemies hypothesis A review ofthe effect of vegetational diversity on predatory in-sects and parasitoids Environ Entomol 18 590-599
Sokal R R and F J Rohlf 1981 Biometry 2nded Freeman San Francisco
SAS Institute 1989 JMP statistical program SAS In-stitute Cary NC
van Emden H F 1990 Plant diversity and naturalenemy efficiency in agroecosystems pp 63-80In MMackauer L E Ehler and J Roland [eds] Criticalissues in biological control Intercept Andover UK
Williams D W 1984 Ecology of the blackberry-leaf-hopper-parasite system and its relevance to Californiagrape agroecosystems Hilgardia 52 1--33
Wilson L T C H Pickett and D L Flaherty1989 French prune trees refuge for grape leafhop-per parasite Calif Agric 43(2) 7-8
Winkler A J J A Cook W M Kliewer and L ALider 1974 Climate and soils pp 58-75 In Gen-eral viticulture University of California Press Berke-ley
Received for publication 5 December 1994 accepted 6November 1995

NoA epos captures in control plots
8060
Vol 25 no 2
402000
00
ENVIRONMENTAL ENTOMOLOGY
0008 00 10 20 30 40 50
B c50 80
bull 40 bull bull 6030
4020
10 20
06
502
lQ-Q~ A~
08bull~=E 06Q= 04tIJ~ 02aQto~l~~~0Z
Fig 3 Scatterplots of the functional relationship between A epos capture in pnme tree refuge vineyardsagainstA epos capture in control vineyards (A) 15-18 April (B = 1705 975 CL L = 058 U = nonestimatable) (8) 24April-l May (B = 238 975 CL L = 205 U = 279) (C) 8--15 May (B = 310 975 CL L = 221 U = 485)Confidence limits were established for 975 as a correction for multiple comparisons
numbers The 2nd is that the effect of prune treeson A epos abundance was dependent not only onthe presence of prune trees themselves but on thedensity of overwintering parasites residing withinthe trees This latter point may have important im-plications for using prune trees as a commercialpest control tactic (discussed below)
Lewis and Stephenson (1966) showed that flyinginsects dispersing in the surrounding environmentaccumulate in sheltered regions near physical bar-riers (natural or artificial windbreaks) They alsofound that insects aggregate behind windbreaks asa multiplicative function of the number of insectsdispersing in the surrounding environment Thiseffect was particularly pronounced for mymaridwasps Using elemental labeling techniques Cor-bett and Rosenheim (in press) found a pattern ofA epos capture suggesting that area-wide A eposnumbers accumulate at a rate 4 times greater im-mediately downwind of prune trees than found up-wind of trees Based on these results they hypoth-esized that a windbreak effect may also be amechanism enhancing abundance within the prunetree-vineyard system We tested this hypothesisacross our replicated study by examining the func-tional relationship between control vineyard trapcaptures (area-wide colonizers in the surroundingenvironment) and prune tree vineyard captures(number of colonizers behind windbreaks) Our re-sults revealed that during each of the 3 sampleperiods the slope of the relationship was 217Thus a multiplicative increase in capture was as-sociated with the presence of prune trees but in-dependent of any contribution from overwinteringA epos densities This result supported the wind-break hypothesis and demonstrated the effect may
be consistent for vineyards with a windbreak pres-ent Furthermore this result identifies an impor-tant additional mechanism that may be affectingA epos vineyard colonization in this system
These results underscore the importance of in-vestigating the ecological mechanisms underlyingsystems of habitat diversification in addition totests of overall effectiveness of diversification Un-derstanding the mechanisms involved will enhanceour ability to manipulate these systems to maxi-mize their effectiveness and provide a usefulframework to begin analyses of new systems
Variation in the Prunc Tree Effect Becauseoverwintering A epos densities varied among theprune tree sites we were interested in how themanagement of pnme trees affects overwinteringA epos populations and their subsequent abun-dance in vineyards Irrigation practices were foundto be a significant factor cplaining much of thevariation in overwintering A epos densities ob-served in pnme trees (Table 4) and somc of thevariation in trap catches in pnme tree vineyardsThe number of prune trees within a site also af-fected overwintering A epos density How treenumber exerts an influence on the density of over-wintering A epos (per square centimeter of pnmebark) is not known but we speculate that greaternumbers of trees may create a more favorable mi-crohabitat increasing the number of A epos thatsuccessfully emerge from their overwinteringhosts
Although tree number had a strong positive in-fluence on the density of A epos emergence holesper square centimeter of prune tree bark it wasnot a predictor of A epos captures in pnme treevineyards This is an enigmatic result At the outset
April IHH6 MUHllIY ET 1 HABITAT DIVEHSIFICATlON FOH BIOLOGICAL CONTHOL 503
of till study we expected that pnllle tree sites con-taining morl trelS would producl a larger pool ofmigrants for thl downwind vineyards The in-enased density (pcr squangt centimeter of bark) ofovcrwintcring A epos in prune trce sites with larg-cr tne 1lI11l11wrsonly strengthened onr expcctationthat trcc numbpr would lw a major determinant ofcady-season abundance in prune trep vineyardsIndclltl wc expccted that we would bl able to es-timatp t]1( minimum numbPI of trees needed togenlrate the gtelof augmentation of A cpos nec-pssary to achieve pest management Instead whatwe found is that trpe number is less a predictor ofalmmlancp in vinpyards than is the number of AlJOS produced ppr centinIPter of bark We also ca]-cu]ahd thc total number of A epos emerged fromovclvintlring sitps vithin an cntire prune tree site(a composite cstimate obtained as the product ofA e]1os ltknsity pcr squarc ccntimetcr of bark thetotal arca plr tngtp and the number of trpes persitc) as t]IP estimatp for A pos contributions fromprunl trpc sites This estimatc was however awcakcr prcdictor of early-season abundance invincyards than Pitill l thc numbcr of A cpos persquarc ccntimctcr of bark or pcr prune tree Howcan we cxplain this resn]t One possibility is thattrees within the largcr prune trcc sites Illay havecontributed uncIlm]]y to the number of vineyardmigrants vith thc rows of trccs directly adjacentto the vincyard producing the bulk of the waspstrapped in vincyards Additional experimentationwill be needed to test this and other potential ex-plauations for the lack of a trec number effect
These results have 2 implications for establish-ing and managing pnnll tree rcfuges for pest con-trol The 1st is that cnhanccd A cpos abundancemay he attainable with a relatively small numberof tnps TIIP 2nd is that irrigation of pnllle treesvill bp of primary importance for supporting highdpnsitics of ovprwintcring i cpos populationsMon detail(gtd rpcommcndations for establishingpnme trcc rcfugps will be presented in future pa-pers
Questious about thc pffect of vegetational diver-sity on tIll diversity and abundance of insect pop-ulations ngtcenty have rcceived considerable atteu-tion from ecologists and pest control specialistsThe rpsu]ts of thp cnrrent study not only supportedtl( hypothdical effect of pnme tree refuges on Acpos abundance but in a broader context providea test of the effect of wgctational diversity on nat-ural encmy abundance As a result this study hassatisfied tIl( 1st of 3 hypothesized natural enemyresponses which pr((licts greater natura] enemyabundance in diversified systems (Bussell 1989Andow 1991) In addition 011study identified thefollOving 2 mpchanisms underlying the influenceof prune trees the presence of an ove1vinteringhost and a windbreak effpct
Future papers vill examine the effect of over-wintering refuges on the remaining 2 hypotheses-the effect of pnnw trees on E elegalltula egg mor-
taity and E elcgalltula population densities withingrape vineyards
Acknowledgments
We extend special thanks to Nerissa Fretman MaddySamant John Tlnnter Jason May and Paul Higgs (De-partment of Entomology University of Califol1lia Davis)for field collections and sample processing Vc also ac-Irnowledge thc assistancc of Hobert Hoy Stan Grant andBrad Coleman (E amp J Gallo Winery Livingston CAl forlogistical and material support This research was fuudedby the Westem Hegional Integrated Pest Managementprogram the California Department of Ptsticidt Hegu-lation and the Locli-Woodbridge vVinegrapt Commis-sion This research was part of BGMs doctoral disser-tation at the University of California at Davis
References Cited
Altieri M A 1992 Agroecological foundatious of al-temative agliculture in Califomia Agric Ecosyst En-viron 39 23-53
Andow D A 1991 Vegetational diversity and artll1o-pod population response Annu Hev Entomol 36561-586
Corbett A und J A Rosenheim 1996 Impact of anatural enemy overwintering refuge and its iuterac-tion vith the surrounding landscape Ecol Entomol(in press)
Doutt R L and J Nakat bullbull 1965 Overwintering ref-uge of Anagrus epos (Hymenoptera Mymaridae) JEcon Entomol 58 586
1973 The Ruhus leafllOpper and its egg parasitoid Anendemic biotic system useful in grape-pest manage-ment Environ Entomol 2 381~186
Flaherty D L L T Wilson V M Stern and IIKido 1985 Biologicalcontrol in San Joaquin Valleyvineyards pp 501-520 Tn D G Herzog and M AHoy [eds] Biological control of agricnltural IPM sys-tems Viley New York
Flint M L and P A Roberts 1988 Using cropdiversity to manage pest problems some Californiaexamples Am J Altern Agric1 163-167
Gorclh G lIml D M Dunlmr 1977 A new Allagmsimportant in the biological control of Stephanitis tak-eljai and a key to the N01ih American species FlaEntomol 60 85-95
Herzo~ D c and J E Funderburk 1985 Plantresistance and cultural practiee interactious with hio-logical control pp 67-88 Til D G Ilerzog and MA Hoy [eds] Biological control of agliclllInral IPMsystems Wiley New York
Kiclo H D L Flaherty D F 80sh Dlul K A Val-ero 1984 French pmne trees as ovclwiuteringsites for the grape leafllOpper egg parasite Am JEnol Vitie 35 156-160
Letourneau D K 1987 The enemies hypothesis Tri-trophic interactions and vegetational diversity in trop-ical agroecosystel11sEcology 68 1616-1622
Lewis T bullbulllid J W Stephenson 1966 The penne-ability of artificial windbreaks and the distribution offlying insects in the leeward shelttred Will AnnAppl BioI 58 355-363
Mulla M S 1957 The biology of Typhlo(fhapnmi-cola (Edwards) and 1 quercus (Fabricius) (Cicadelli-dae lIomoptera) Ann Entomol SOlAm 50 76-87
504 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
National Research Council 1989 Alternative agri-culture National AcademyWashington DC
PaAel M D and P H Harvey 1988 Recent de-velopments in the analysis of comparative data QRev BioI 63 418-440
Pickett C H L T Wilson and D L Flaherty1990 The role of refuges in crop protection withreference to plantings of French prune trees in agrape agroecosystem pp 151-165 In N J BostanianL T Wilson and T J Dennehy [eds] Monitoringand integrated management of arthropod pests ofsmall fruit crops Intercept Andover
RusseD E P 1989 Enemies hypothesis A review ofthe effect of vegetational diversity on predatory in-sects and parasitoids Environ Entomol 18 590-599
Sokal R R and F J Rohlf 1981 Biometry 2nded Freeman San Francisco
SAS Institute 1989 JMP statistical program SAS In-stitute Cary NC
van Emden H F 1990 Plant diversity and naturalenemy efficiency in agroecosystems pp 63-80In MMackauer L E Ehler and J Roland [eds] Criticalissues in biological control Intercept Andover UK
Williams D W 1984 Ecology of the blackberry-leaf-hopper-parasite system and its relevance to Californiagrape agroecosystems Hilgardia 52 1--33
Wilson L T C H Pickett and D L Flaherty1989 French prune trees refuge for grape leafhop-per parasite Calif Agric 43(2) 7-8
Winkler A J J A Cook W M Kliewer and L ALider 1974 Climate and soils pp 58-75 In Gen-eral viticulture University of California Press Berke-ley
Received for publication 5 December 1994 accepted 6November 1995

April IHH6 MUHllIY ET 1 HABITAT DIVEHSIFICATlON FOH BIOLOGICAL CONTHOL 503
of till study we expected that pnllle tree sites con-taining morl trelS would producl a larger pool ofmigrants for thl downwind vineyards The in-enased density (pcr squangt centimeter of bark) ofovcrwintcring A epos in prune trce sites with larg-cr tne 1lI11l11wrsonly strengthened onr expcctationthat trcc numbpr would lw a major determinant ofcady-season abundance in prune trep vineyardsIndclltl wc expccted that we would bl able to es-timatp t]1( minimum numbPI of trees needed togenlrate the gtelof augmentation of A cpos nec-pssary to achieve pest management Instead whatwe found is that trpe number is less a predictor ofalmmlancp in vinpyards than is the number of AlJOS produced ppr centinIPter of bark We also ca]-cu]ahd thc total number of A epos emerged fromovclvintlring sitps vithin an cntire prune tree site(a composite cstimate obtained as the product ofA e]1os ltknsity pcr squarc ccntimetcr of bark thetotal arca plr tngtp and the number of trpes persitc) as t]IP estimatp for A pos contributions fromprunl trpc sites This estimatc was however awcakcr prcdictor of early-season abundance invincyards than Pitill l thc numbcr of A cpos persquarc ccntimctcr of bark or pcr prune tree Howcan we cxplain this resn]t One possibility is thattrees within the largcr prune trcc sites Illay havecontributed uncIlm]]y to the number of vineyardmigrants vith thc rows of trccs directly adjacentto the vincyard producing the bulk of the waspstrapped in vincyards Additional experimentationwill be needed to test this and other potential ex-plauations for the lack of a trec number effect
These results have 2 implications for establish-ing and managing pnnll tree rcfuges for pest con-trol The 1st is that cnhanccd A cpos abundancemay he attainable with a relatively small numberof tnps TIIP 2nd is that irrigation of pnllle treesvill bp of primary importance for supporting highdpnsitics of ovprwintcring i cpos populationsMon detail(gtd rpcommcndations for establishingpnme trcc rcfugps will be presented in future pa-pers
Questious about thc pffect of vegetational diver-sity on tIll diversity and abundance of insect pop-ulations ngtcenty have rcceived considerable atteu-tion from ecologists and pest control specialistsThe rpsu]ts of thp cnrrent study not only supportedtl( hypothdical effect of pnme tree refuges on Acpos abundance but in a broader context providea test of the effect of wgctational diversity on nat-ural encmy abundance As a result this study hassatisfied tIl( 1st of 3 hypothesized natural enemyresponses which pr((licts greater natura] enemyabundance in diversified systems (Bussell 1989Andow 1991) In addition 011study identified thefollOving 2 mpchanisms underlying the influenceof prune trees the presence of an ove1vinteringhost and a windbreak effpct
Future papers vill examine the effect of over-wintering refuges on the remaining 2 hypotheses-the effect of pnnw trees on E elegalltula egg mor-
taity and E elcgalltula population densities withingrape vineyards
Acknowledgments
We extend special thanks to Nerissa Fretman MaddySamant John Tlnnter Jason May and Paul Higgs (De-partment of Entomology University of Califol1lia Davis)for field collections and sample processing Vc also ac-Irnowledge thc assistancc of Hobert Hoy Stan Grant andBrad Coleman (E amp J Gallo Winery Livingston CAl forlogistical and material support This research was fuudedby the Westem Hegional Integrated Pest Managementprogram the California Department of Ptsticidt Hegu-lation and the Locli-Woodbridge vVinegrapt Commis-sion This research was part of BGMs doctoral disser-tation at the University of California at Davis
References Cited
Altieri M A 1992 Agroecological foundatious of al-temative agliculture in Califomia Agric Ecosyst En-viron 39 23-53
Andow D A 1991 Vegetational diversity and artll1o-pod population response Annu Hev Entomol 36561-586
Corbett A und J A Rosenheim 1996 Impact of anatural enemy overwintering refuge and its iuterac-tion vith the surrounding landscape Ecol Entomol(in press)
Doutt R L and J Nakat bullbull 1965 Overwintering ref-uge of Anagrus epos (Hymenoptera Mymaridae) JEcon Entomol 58 586
1973 The Ruhus leafllOpper and its egg parasitoid Anendemic biotic system useful in grape-pest manage-ment Environ Entomol 2 381~186
Flaherty D L L T Wilson V M Stern and IIKido 1985 Biologicalcontrol in San Joaquin Valleyvineyards pp 501-520 Tn D G Herzog and M AHoy [eds] Biological control of agricnltural IPM sys-tems Viley New York
Flint M L and P A Roberts 1988 Using cropdiversity to manage pest problems some Californiaexamples Am J Altern Agric1 163-167
Gorclh G lIml D M Dunlmr 1977 A new Allagmsimportant in the biological control of Stephanitis tak-eljai and a key to the N01ih American species FlaEntomol 60 85-95
Herzo~ D c and J E Funderburk 1985 Plantresistance and cultural practiee interactious with hio-logical control pp 67-88 Til D G Ilerzog and MA Hoy [eds] Biological control of agliclllInral IPMsystems Wiley New York
Kiclo H D L Flaherty D F 80sh Dlul K A Val-ero 1984 French pmne trees as ovclwiuteringsites for the grape leafllOpper egg parasite Am JEnol Vitie 35 156-160
Letourneau D K 1987 The enemies hypothesis Tri-trophic interactions and vegetational diversity in trop-ical agroecosystel11sEcology 68 1616-1622
Lewis T bullbulllid J W Stephenson 1966 The penne-ability of artificial windbreaks and the distribution offlying insects in the leeward shelttred Will AnnAppl BioI 58 355-363
Mulla M S 1957 The biology of Typhlo(fhapnmi-cola (Edwards) and 1 quercus (Fabricius) (Cicadelli-dae lIomoptera) Ann Entomol SOlAm 50 76-87
504 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
National Research Council 1989 Alternative agri-culture National AcademyWashington DC
PaAel M D and P H Harvey 1988 Recent de-velopments in the analysis of comparative data QRev BioI 63 418-440
Pickett C H L T Wilson and D L Flaherty1990 The role of refuges in crop protection withreference to plantings of French prune trees in agrape agroecosystem pp 151-165 In N J BostanianL T Wilson and T J Dennehy [eds] Monitoringand integrated management of arthropod pests ofsmall fruit crops Intercept Andover
RusseD E P 1989 Enemies hypothesis A review ofthe effect of vegetational diversity on predatory in-sects and parasitoids Environ Entomol 18 590-599
Sokal R R and F J Rohlf 1981 Biometry 2nded Freeman San Francisco
SAS Institute 1989 JMP statistical program SAS In-stitute Cary NC
van Emden H F 1990 Plant diversity and naturalenemy efficiency in agroecosystems pp 63-80In MMackauer L E Ehler and J Roland [eds] Criticalissues in biological control Intercept Andover UK
Williams D W 1984 Ecology of the blackberry-leaf-hopper-parasite system and its relevance to Californiagrape agroecosystems Hilgardia 52 1--33
Wilson L T C H Pickett and D L Flaherty1989 French prune trees refuge for grape leafhop-per parasite Calif Agric 43(2) 7-8
Winkler A J J A Cook W M Kliewer and L ALider 1974 Climate and soils pp 58-75 In Gen-eral viticulture University of California Press Berke-ley
Received for publication 5 December 1994 accepted 6November 1995

504 ENVIRONMENTAL ENTOMOLOGY Vol 25 no 2
National Research Council 1989 Alternative agri-culture National AcademyWashington DC
PaAel M D and P H Harvey 1988 Recent de-velopments in the analysis of comparative data QRev BioI 63 418-440
Pickett C H L T Wilson and D L Flaherty1990 The role of refuges in crop protection withreference to plantings of French prune trees in agrape agroecosystem pp 151-165 In N J BostanianL T Wilson and T J Dennehy [eds] Monitoringand integrated management of arthropod pests ofsmall fruit crops Intercept Andover
RusseD E P 1989 Enemies hypothesis A review ofthe effect of vegetational diversity on predatory in-sects and parasitoids Environ Entomol 18 590-599
Sokal R R and F J Rohlf 1981 Biometry 2nded Freeman San Francisco
SAS Institute 1989 JMP statistical program SAS In-stitute Cary NC
van Emden H F 1990 Plant diversity and naturalenemy efficiency in agroecosystems pp 63-80In MMackauer L E Ehler and J Roland [eds] Criticalissues in biological control Intercept Andover UK
Williams D W 1984 Ecology of the blackberry-leaf-hopper-parasite system and its relevance to Californiagrape agroecosystems Hilgardia 52 1--33
Wilson L T C H Pickett and D L Flaherty1989 French prune trees refuge for grape leafhop-per parasite Calif Agric 43(2) 7-8
Winkler A J J A Cook W M Kliewer and L ALider 1974 Climate and soils pp 58-75 In Gen-eral viticulture University of California Press Berke-ley
Received for publication 5 December 1994 accepted 6November 1995







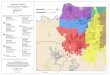


















![913[1]enlistment rule](https://img.pdfslide.us/doc/110x75/55cf9d4f550346d033ad11ce/9131enlistment-rule.jpg)


