Embed Size (px)
Citation preview

Michael Kaden-Hoffmann
Isolation and Identification of an Unknown Bacteria Found in Ski Gloves
Introduction
Bacteria are unparalleled in their abilities to exist in almost every location on
earth (Dykhuizen, 1998). This is due to bacteria’s ability to use a variety of energy
sources and the enormous diversity in structure in regards to there cells and
cellular components. While most bacteria are harmless to human life, some can be a
nuisance, and others can be deadly.
Our hands are home to a huge and diverse microbiome that is necessary for
life. However due to the objects we touch our hands are also host pathogenic
bacteria that are suppressed by our microbiome (Edmonds-Wilson et al., 2015).
When we touch things we spread not just our microbiome, but also these pathogenic
bacteria around. To combat the spread of bacteria on a day-to-day bases, we often
wash our hands and clothing. However, one thing we often neglect to wash enough
is the gloves that we wear in the winter. For that reason gloves are home to a large
number of bacteria, which can lead to bad smells and skin problems (Vargas-Díez,
1996). The goal of this study was to isolate and identify a single bacterium found in
ski gloves to get a glimpse of the microbiome I found there.
To identify my isolated bacterium I exposed it to a multitude of tests, both
phenotypic and genotypic, to learn how it survives in its environment and fulfills its
energy requirements. I looked at my isolate under a microscope and employed
various staining techniques to identify physical characteristics. In addition to those
tests I extracted DNA from my isolate, which was sequenced. The resulting partial

Michael Kaden-Hoffmann
genome was then used to identify taxonomy and specific metabolic traits of my
isolate. Due to being found in gloves and being cultured on a tryptic soy agar plate I
expected my bacterium to an aerobic strain that uses some from of sugar as an
energy source.
Methods
Sample Collection and Isolation
To start this experiment I used a moist sterile cotton swab to swab a ski
glove. This swab was then streaked over a tryptic soy agar (TSA) plate in a zigzag
pattern. This plate was then left to grow for a week a room temperature. Then a lone
orange colony was chosen from the TSA plate and transferred to a new TSA plate
using the quadrant streak method as detailed in Lab 2. This quadrant streak was
grown in a 37°C incubator for two days. Then a single colony was selected and
transferred to a new TSA plate via the same quadrant streak technique used before.
Over the following two weeks this was repeated three additional times to create a
culture containing a colony isolated species of bacterium. After the fourth quadrant
streak was grown, a single colony was selected and transferred to a tube containing
a sterile TSA slant.
Physiological Testing
To begin the identification of the isolated bacterium I performed a series of
tests to identify the physiological traits of the bacterium. Colony color, shape,
margins, elevation, and motility were recorded from the previously grown TSA
plates. A new TSA plate was cultured using the pure culture streak as detailed in Lab
2. From this plate a sample was taken and a Gram stain performed using the

Michael Kaden-Hoffmann
protocol in Lab 4. The bacterium was then examined under the microscope for color,
shape, and grouping. Next an API 20E test strip was inoculated with the bacterium
and left to grow in a 37°C incubator for two days as detailed in Lab 6. The API 20E
test strip was used to determine if the bacterium was positive or negative for 20
different metabolic reactions. In addition to the API 20E test strip, a fluid
thioglycollate test, an oxidase test, and a catalase test were also performed using the
protocols in Lab 6. The fluid thioglycollate test was used to determine the oxygen
class of the bacterium, the oxidase test was used to determine if the bacterium
contained cytochrome c oxidase, and the catalase test was performed to test for the
presence of the enzyme catalase. Lastly a disk diffusion test adapted from the
standard Kirby-Bauer method was preformed as detailing in Lab 9 to test for
antibiotic resistance traits in the bacterium.
Genetic Sequencing and Analysis
In addition to the physiological tests I also performed a DNA test on the
bacterium to help with identification. This was done in two steps; DNA extraction
and sequencing followed by DNA sequence data analysis. To isolate the DNA a
PowerSoil DNA Isolation Kit was used following the protocols in Lab 5. The isolated
DNA was then sent to the Arctic Biology DNA Core Lab, where a specialized
technician did the sequencing. The Core Lab sent back a file, which contained the
unanalyzed DNA sequence fragments. The first step was to upload the files to
BaseSpace where I could assemble and analyze the DNA. Once the files were in
BaseSpace I used the program SPAdes Genome Assembler to assemble the DNA
fragments into contigs. These contigs were then uploaded to the program Kraken

Michael Kaden-Hoffmann
Metagenomics, where they were compared to a database for identification of the
bacterium. Also the contigs were uploaded to the program Prokka Genome
Annotation, which identified genes found within the contigs. Due to poor results
from the Kraken Metagenomics program the contigs were also uploaded to the
BLAST database where they were compared to known DNA sequences to identify
the bacterium.
Results
The results of the identification tests were mixed with only the physiological
traits of the bacterium, the Gram stain, the catalase test, and the oxidase test
producing conclusive results (Table 1). Inspection of the colonies of the isolate
showed that it had a deep orange cool (Figure 1), round shape, and no visible
motility.
Test Name Result
Gram Stain Gram positive
Catalase Test Catalase positive
Oxidase Test Cytochrome c oxidase positive
Table 1. Tests performed and the results.

Michael Kaden-Hoffmann
Figure 1. Above is photo of the last quadrant streak of my isolate after two days of growth in a 37°C incubator.
The API 20e test strip and the API Staph test strips both yielded inconclusive results
as no changes were observed after they were inoculated with the bacterium. Also,
the fluid thioglycollate test was inconclusive as the bacteria grew immediately after
inoculation along the inoculation site, but then never continued to grow anywhere
in the tube, even when put into a 37°C incubator.
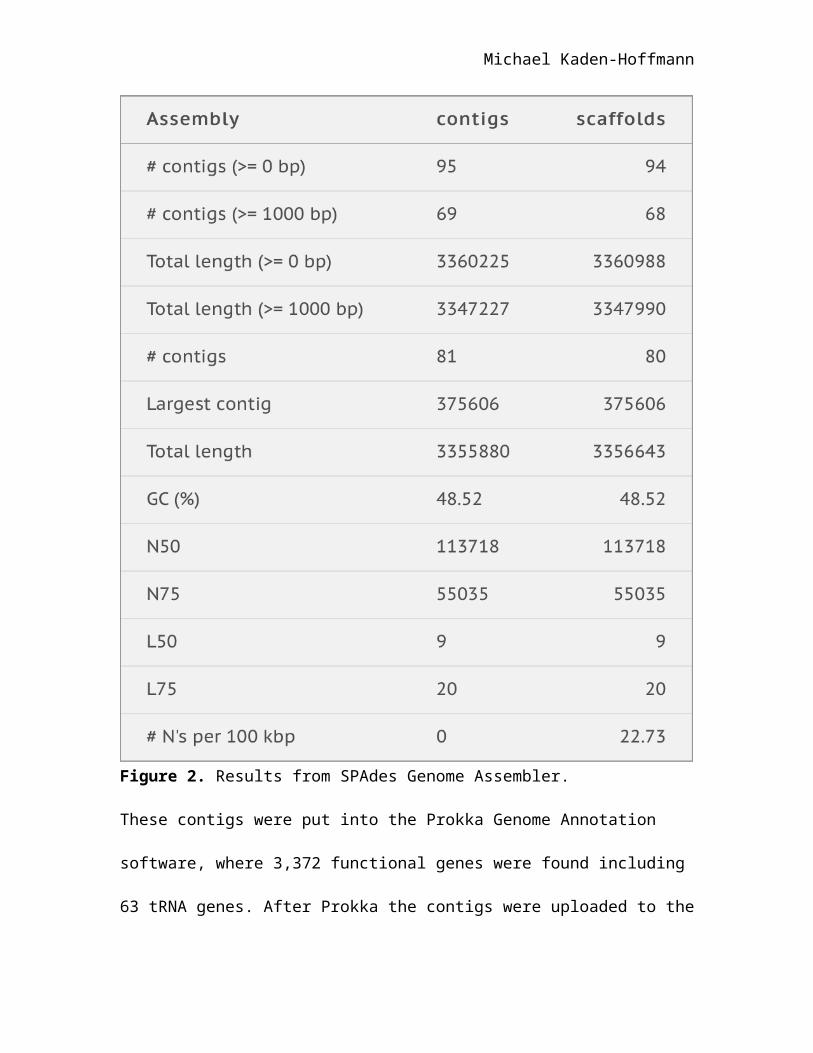
The most useful information in identification of the bacterium came from the
DNA sequencing data. The DNA sequencing data assembled into 95 contigs made of

Michael Kaden-Hoffmann
3,355,880 base pairs in SPAdes Genome Assembler, with the longest contig being
375,606 base pairs long (Figure 2).
Figure 2. Results from SPAdes Genome Assembler.
These contigs were put into the Prokka Genome Annotation software, where 3,372
functional genes were found including 63 tRNA genes. After Prokka the contigs were

Michael Kaden-Hoffmann
uploaded to the Kraken Metagenomics program for species identification. However,
Kraken Metagenomics could not identify the bacterium, as the DNA sequences didn’t
match any in the database. Thus, I uploaded the contigs to BLAST and compared my
sequencing data with that database (Altschul et al., 1990). The isolate’s DNA
sequences matched most closely with DNA sequences of the genera Planococcus and
Planomicobium with each having a query cover values of 84%.
The last test performed on the bacterium was the antibiotic susceptibility
test. In this test the isolate was grown in eight environments where each
environment had a different antibiotic tablet at the center. Of the eight antibiotics
tested, the bacterium was only resistant to Oxacillin (Table 2).
Antibacterial Compound Susceptibility
Cefoperazone Susceptible
Vancomycin Susceptible
Amikacin Susceptible
Trimethopim Susceptible
Gentamicin Susceptible
Oxacillin Resistant
Tobramycin Susceptible
Cenfazolin Susceptible
Table 2. Antibiotic compounds and the susceptibility of the isolate to them.
Discussion
Since the API 20E test strip and API Staph strip both yielded no results, the
DNA sequencing is the primary source of insight into the identification of the

Michael Kaden-Hoffmann
bacterium. When compared to the BLAST database the DNA sequences of the isolate
matched most closely with the genera Planococcus and Planomicobium (Altschul et
al., 1990; Margolles et. al., 2012). Both genera are very closely related, with the
genus Planomicobium only being separated from Planococcus in 2001 (Yoon et. al.,
2001). The main differences cited as the reason for the taxonomic separation of the
genera in the Yoon et. al. paper, was the shape of the cells and some biochemical
differences (Yoon et. al., 2001). Planomicobium are cited as being coccobacillus with
a shape change to bacillus over time, while Planococcus are said to be remain cocci
shaped (Yoon et. al., 2001). Since the isolate remained cocci shaped over the course
of the study, I identified it as Planococcus.
Looking at studies done on the genus Planococcus, the colonies are orange
and circularly shaped about 1-2mm in diameter when grown on TSA plates
(Mayilraj, 2005). This matches exactly what was observed in with the isolate in my
study. Mayilraj’s research also showed that some Planococcus grow on mannitol and
can grow on plates with high salt content, but didn’t produce acidic products, which
was something that was also observed with the isolate in this study (Mayilraj,
2005). However, evidence suggests that certain species of Planococcus, do not grow
on mannitol, but can similarly tolerate high salt content (Getty et. al., 2002).
Planococcus species are all Gram-positive or Gram-variable, catalase positive, and
strict aerobes, which matches up with the isolate from this study (Getty et. al., 2002;
Mayilraj, 2005). However, both the Getty and Mayilraj studies found Planococcus to
be oxidase negative, which was not the case with this bacterium, possibly indicating
a new or unknown species. With few DNA sequences for Planococcus existing in the
Michael Kaden-Hoffmann
NCBI database used in BLAST, the question of whether or not this was a new species
of Planococcus could not be answered.
Planococcus have so far only been isolated from cold places like in
permafrost in Antarctica, under glaciers in the Himalayas, and in lakes in Antarctica,
however all the studies done have shown that they grow the best at temperatures
around 25°C in the lab (Getty et. al., 2002; Mayilraj, 2005; Margolles et. al., 2012).
This optimal growth at was 25°C also observed in this study, as well as growth at
around 3°C but at a much slower rate. The sampling location for this study (a ski
glove) is not in a cold place like the locations in the other studies, however it is
exposed to cold places where the bacteria normally grow, thus hinting at a possible
source of origin. If I were going to conduct additional research to further confirm
the identity of my isolate, I would conduct the appropriate tests (see source) to
assess its motility (Bassiri). This insight is supported by multiple studies, which
found that both the Planococcus and Planomicobium genera exhibit bacteria that are
motile, having a single flagella to move around with (Getty et al., 2002;Mayilraj,
2005;Margolles et. al., 2012;Yoon et. al., 2001).

Michael Kaden-Hoffmann
Citations
Altschul, S.F., Gish, W., Miller, W., Myers, E.W. & Lipman, D.J. (1990) "Basic local alignment search tool." J. Mol. Biol. 215:403-410.
Dykhuizen, Daniel. (1998) “Santa Rosalia revisited: Why are there so many species of bacteria?”. Antonie van Leeuwenhoek 73.1: 25-33.
Edmonds-Wilson S., Nurinova N., Zapka C., Fierer N., and Wilson M. (2015) "Review Of Human Hand Microbiome Research". Journal of Dermatological Science 80.1: 3-12.
Margolles, A., Gueimonde, M., & Sanchez, B. (2012). Genome Sequence of the Antarctic Psychrophile Bacterium Planococcus antarcticus DSM 14505. Journal Of Bacteriology, 194(16), 4465-4465. http://dx.doi.org/10.1128/jb.00888-12
Mayilraj, S. (2005). Planococcus stackebrandtii sp. nov., isolated from a cold desert of the Himalayas, India. INTERNATIONAL JOURNAL OF SYSTEMATIC AND EVOLUTIONARY MICROBIOLOGY, 55(1), 91-94. http://dx.doi.org/10.1099/ijs.0.63290-0
Getty, R., Prakash, J., Vairamani, M., Prabhakar, S., Matsumoto, G., & Shivaji, S. (2002). Planococcus antarcticus and Planococcus psychrophilus spp. nov. isolated from cyanobacterial mat samples collected from ponds in Antarctica. Extremophiles, 6(3), 253-26, http://dx.doi.org/10.1007/s00792-001-0250-7
Vargas-Díez E., Buezo G., Aragües M., Daudén E., and Ory, F. (1996) “Papular-Purpuric Gloves-and-Socks Syndrome". International Journal of Dermatology 35.9: 626-632
Yoon, J., Lee, K., Kang, S., Kho, Y., Lee, E., Park, Y., & Kang, K. (2001). Planomicrobium koreense gen. nov., sp. nov., a bacterium isolated from the Korean traditional fermented seafood jeotgal, and transfer of Planococcus okeanokoites (Nakagawa et al. 1996) and Planococcus mcmeekinii (Junge et al. 1998) to the genus Planomicrobium. International Journal Of Systematic And Evolutionary Microbiology, 51(4), 1511-1520. http://dx.doi.org/10.1099/00207713-51-4-1511
Bassiri, E., http://www.sas.upenn.edu/LabManuals/biol275/Table_of_Contents_files/8-Motility.pdf





























