Embed Size (px)
Citation preview

Heft 2 ] 1991 J 145
B Orn. 132, 1991: S. 145--163
Early Stages of Vocal Ontogeny in the Magpie (Pica pica)
Tomäs Redondo
Introduction Much of our knowledge about the ontogeny of bird vocalizations comes from
studies on the development of song in oscine birds (KROODSMA & Mm>EI< 1982). Such studies have revealed a general pattern of song ontogeny across many different species: juvenile birds have non-differentiated precursors of song elements which become gradually modified into their adult-typical configuration through a continuous process of change (MARLER & PETERS 1982). However, compared with the enormous amount of information available on song development in passerines, we know little about the ontogeny of non-song vocalizations (ZANN 1975; BEAVER 1978; COSENS 1981; SMITH 1983; HOWES-JONES 1984). As a consequence, emphasis on the former topic has pro- bably introduced a source of conceptual blas in our interpretation of the latter. Most studies dealing with the ontogeny of non-song calls in passerines and other avian orders have largely assumed that (i) development consists of a continuous phenomenon in which mature vocalizations arise after a graded process of change, starting from im- mature vocalizations (MARLEe, 1956; MESSMER & MESSMER 1956; LANYON 1960; MOYN~- HAN 1962; THOMPSON & RICE 1970; ZANN 1975; WILKINSON & HUXLEY 1978); and (ii) that structural variations in the calls at early, non-differentiated stages, make functional sense only in as rar as they serve as precursors for later vocalizations (MaRLER & MUNDINGER 1975; WILKINSON & HUXLEY 1978).
The question of how a behavioural trait develops is usually approached by identify- ing several stages of the same trait and ordering them in a continuous time sequence The criterion by which such stages are said to represent different performances of the same trait is two-fold: they share some characteristic structural elements and/or are evoked by the same set of causal factors (GROOTHUIS 1989). Many studies of call development in birds fail, however, to meet one, and often both, of these requirements: vocalizations arise from earlier ones with no transitional forms between them (MESSMER & MESSMER 1956; THOMPSON & RICE 1970; WlLKINSON & HUXLEY 1978; COSENS 1981; HOWES-JONES 1984); different developmental stages of the same call (as identified by structural similarities) are frequently associated with different motiva- tional stares (MAi<LER 1956; MESSMER & MESSMER 1956; LANYON 1957 in HOWES-JONES 1984; COSENS 1981; HOWES-JONES 1984); and calls which are included in separate developmental routes at a given age show motivational graduations between them (MESSMER &i; MESSMER 1956; WILKINSON & HUXLEY 1978; COSENS 1981; MHNERT & BERGMANN 1983; THOREN & BERGMANN 1987a, b).

T. REDO~,DO [ J" Orn. 146 L 132
Before understanding why modets of song development fall to explain the observed patterns of call ontogeny successfully, more information needs to be gathered on how calls develop and what are they used for by young birds. Studies on behavioural development have long suffered from a theoretical tradition which focused on the adult phenotype as the major target for natural selection, hence overlooked that im- mature traits may be functional in the short-term, adapted to particular ontogenetic niches (CHISZAR 1985; ALBERTS & CRAMER 1988; GOMENDIO 1988). No comprehensive account of developrnental patterns can be obtained by ignoring that animals taust be functional at all stages of their life cycle (OPPENHEIM 1980), SO an evolutionary perspective may help us to gain further insight into many problems concerning the ontogeny of behaviour (Bn;oFF 1985).
In this paper, I analyze the vocal development of Magpie (Picä pica) nestlings and fledglings from a functional standpoint. Magpies suffer from high mortality rates dur- ing the earliest part of their life cycle (R~DoNDO & Can~ANZA 1989) which, as in most altricial species, appear to be related mainly to the combined effects of predation and starvation (RICKLEFS 1969). Here, I will suggest rather than considering juvenile calls as simple precursors of aduk vocalizations, they seem to have evolved as an adap- tation to diminish both sources of mortality.
Material and Methods
Field recordings of magpie nest]ing calls were conducted at Espiel, Córdoba, during the 1985 and 1987 breeding seasons. Since hatching occurs asynchronously, nestling age was categorized into seven three-days age classes, thus covering the whole nestling period (about 24 days). Recordings from 92 nestlings (38 nests) were obtained in the absence of parents, by using an Uher 4200 Report Monitor tape-recorder through a condenser microphone AKG 568 EB, at 9.5 cm/s speed. The behavioural context in which nestling calls were emitted was classified into four categories (begging, resting, crouching and escaping frorn the nest). Family groups of fledglings were observed for up to eight days after leaving the nest. Sonagrams for a sample of each type of call/nest/day were obtained in a FFT digital sonagraph UNISCAN II 4600. For tonal calls, I measured duration and maximum and minimum frequency of the loudest fundamental band of energy. Frequency modulation in tonal calls appeared like series of V-like valleys. The slope (in Hz/ms) of the initial limb in the steepest valley was measured as an estimate of maximum frequency modulation.
During 1988, 28 nestlings were hand-reared from the time of hatching. In the laboratory, nestlings were kept in artificial (plaster) magpie nests, each containing three to seven nestlings of the same age, in wood boxes with a side entrance under controlled conditions of humidity and temperature. Boxes were placed at 1 m above ground, allowing nestlings to abandon them easily. I considered that a chick had left the nest when it was found out of the box for three consecutive days. Nestlings were red every hour from 08 00 to 20 00 when 1--14 days old and every two hours when older. These feeding rates resembled those observed in the wild (BuITRON 1988). Food supplied to nestlings was a modification of LANYON'S (1979) recipe (pro- tein 48.3 % of dry weight), finely homogeneized so as to flow through a 4 mm canula attached to a syringe. Feeding schedule consisted of serial feeding of nestlings in the same nest, but feeding order changed daily on a rotative basis. Feeding took place in isolation, inside a nest box identical to those in which nestlings were kept, which was equipped with recording ap-

Heft 2 ] 1991 ] Vocai ontogeny in Pica pica 147
paratus. I calculated begging rates of nestlings with a short (1 h) and a long (2.5 h) food depriva- tion time as an estimate of their hunger motivation. A positive correlation between fasting time and hunger motivation (measured as the amount of food intake) has been reported in rats (LI~ MA61VEN i983). Since hatching, nestlings had become conditioned to a human word as a begging-eliciting stimulus in order to obtain a sufficient number of calls during feeding ses- sions. Stimulation rate was controlled for as a covariate for comparing begging rates between short and long periods of food deprivation (analysis of covariance, Zat~ 1984).
Nineteen nestlings completed their development successfully and were kept in the labora- tory for their first week after fledging. They were released once they had managed to feed by themselves in an outdoor aviary.
Nestling growth was determined by fitting fasting body weight (measured prior to the first feeding session) to a logistic model (RIcK~ErS 1983) by a non-linear regression procedure (SvATCRaPH, Statistical Graphics Co., Rockville, Maryland, 1986). The development of the> moregulatory ability was determined according to O ' C o ~ o R (1984). Nestlings were pur on clothed Petri capsules and exposed to an ambient temperature of 25 °C for 30 min. Measurements of cloacal temperature before and after cooling were taken by a cloacal ther- mometer (Miller & Weber, Inc.), from which a Thermoregulatory Index can be computed. Adult cloacal temperature was arbitrarily established at 39 °C. Finally, I determined modal ages of appearance of some relevant features related to sensory-motor development: grasping a pencil; balancing on a stick; and opto-motor responses (tracking a circling finger with head movements).
I thank E. CuRio, M. FicxeN and M. GoM~aeio for helpful criticism of earlier drafts of this paper; L. ARIAS DE REYNA and coworkers for assistance with field work; A. PACHE and B. PASCU*L for solving lots of problems at the laboratory, and J. CaRRANZA for many facilities during chick reari~g and tape analysis. Research funds from Catedra de Biolog~a y Etologia, Facultad de Veterinaria, Cäceres, provided partial financial support. During the writing of the paper, the author enjoyed a grant from the Programa de Perfeccionamiento de Doctores y Tecnólogos en Espafia, MEC.
Results 1. G e n e r a l D e v e l o p m e n t
In the wild, newly hatched nestlings usually gaped silently, but vocalizations become frequent when 1 - 2 days old. Auditory channels appear fully open at 4 - 6 days. Eyes open between 7-- 10 days. At first, begging is directed upwards and becomes orientated from 12 days onwards. Neossoptiles emerge between 7 - 1 0 days, nestlings appearing fully feathered on dorsal view by day 14. Initial preening movements can be observed as early as 9 days. Neck extension and wing elevation appear at 13 and 19 days, respec- tively. Flying exercises were first observed at 17 days of age Figure 1 shows the increase in body weight and thermoregulatory ability, as weil as the timing of appearance of some patterns of locomotory coordination in laboratory-reared nesttings. All in- dividual growth curves fitted closely with a logistic model (minimum F value obtain- ed, F = 2886, df = 2, 28, p <0.001). Mean values of asymptotic weight and intrinsic growth rate were~ respectively, 158.4 g +7.87 95 % CI and 0.262 days -1 +0.011 95 % CI. Maximum weight gain per day occurs at 10.9 days + 1.4 SD. Figure 1 also shows that endothermy is poor ly developed before 11 days and that coordination of motor patterns improves rapidly between 14 and 20 days. Fledging &tes observed in the wild

[ j . Orn. 148 T. REI)ONDO 132
Percentage of Adult I
20 ~ i O ~ t I I I I I I I I I t I ! k I I I I I I I i I 1 ~ -
2 5 8 11 14 17 20 23 26
N e s t l i n g A g e (days)
Fig. 1. The development o{ thermoregulatory ability as a percentage of adult thermoregu]atory index and growth curve expressed as percentage of asymptotic weight. Vertical bars around mean values of ther- moregulatory index show ranges. Arrows indicate modal ages of appearance of some patterns of locomotory coordination: Optomotor response (OM), Grasping (GR) and Balancing on a stick (BA).
(24 days 4-2.6 SD, N = 25 nests) were earlier than those obtained in the laboratory (26.1 4-2.98 SD, N = 19), the difference being probably explained by premature fledg- ing of wild broods in response to disturbance
I I I
[F I I I
BT
SW
BCN I L I I I E
AC J I I I ] [ I I I I
DC I I E I l l l I l l
BS
,?
I I I I i l > i
0 5 10 15 20 25 30" 35 Nestling Age (days)
Fig. 2. Ages of occurrence of nestling and fledgling calls. Tonal and harsh vocalizations are denoted by smooth and ticked lines, respectively. SW: Soft Whistle; BT: Begging Trill; BS: Begging Scream; DC: Distress Call; AL: Alarm Calt; BCN: Brief Contact Note The arrow points to mean age of fledging.

Heft 2] Voca] ontogeny in Pica pica 149 1991 /
2. V o c a l i z a t i o n s
The vocal repertoire of wild nestlings consisted of four cal]s (Fig. 2): Begging Trill, Soft Whistle, Distress Cal] and Alarm Call. Three of these ca]ls (Begging, Distress and Alarm Calls) remained in the repertoire of fledglings, although Begging Trills changed into Begging Screams at just about the time of leaving the nest. In addition, fledgling» also gare Brief Contact Notes. Laboratory-reared nestlings were never observed to give Distress Calls hut they uttered all the remaining vocalizations in similar contexts and following similar developmental courses as nestlings observed in the fiel&
10
8
6
4
2
0 0
Frequency (kHz) : , ~ :::~» *%,
• -~:. "-,t.',--'~-, :.
0.5 Time (8)
Frequency (kHz) 10
81 .. I ~ ~ " « ' : . ~ : ~ , :~ ' ~ ~ : : ' i - .
0 0.5 1 Time (s)
Fig. 3. Upper: Begging Trills of nestlings (left to right) 5, 8 and 11 days old, respectively. Lower: Begging Tri]l of a nestling 17 days o]d.

150 T. REDONDO [ J" Orn. k 132
Begging Trill
These calls had a tonal structure~ composed of up to four bands. The first begging calls, recorded when nestlings were 1 - 2 days old, appeared in the sonagram as inverted U structures. This simple structure becomes highly complex as nestlings grow older, showing a dramatic increase in duration and frequency-modulation (Fig. 3; Table). To my ear, calls also increased in amplitude with age hut no intensity measurement was done As a result of frequency modulation (FM), energy is widely dispersed along a wide frequency fange This leads to calls becoming increasingly noisier with age, but cioser examination of the sonagrams at a lower speed revealed that they still had a tonal quality. This process of increasing duration and FM ends when nestlings are about to leave the nest.
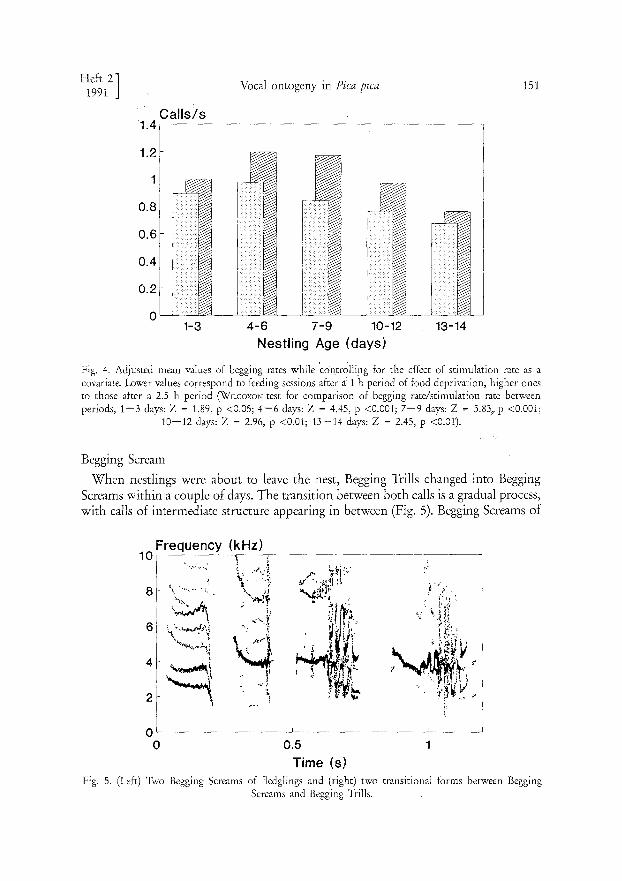
Rates of calling were dependent upon the nestlings' level of hunger motivation. As Fig. 4 shows, begging rates were higher after a 2.5 h long period of food deprivation than after a 1 h long period.
Mean values (+SE) of maximum and minimum frequency, duration, frequency fange and maximum rate of frequency modulation of the dominant fundamental band in Begging Trills
(top) and Soft whistles (bottom).
Age FRMIN FRMAX DURATION RANGE FM RATE (days) (Hz) (Hz) (ms) (Hz) (Hz/ms)
Mean SE Mean SE Mean SE Mean SE Mean SE
i-- 3 4 737 120 6 206 125 126 6.9 7 674 179 21.0 1.7 4-- 6 4 689 121 6 691 139 180 10.0 8 693 203 22.9 1.3 7-- 9 3 222 63 5 510 106 307 19.4 7 798 207 32.5 2.3
10--12 2 281 58 4 938 234 559 50.3 7 594 459 33.0 3.4 13--15 2 331 74 5 226 114 511 34.3 8 120 224 46.6 4.4 16--18 1 700 50 4 499 87 701 41.6 7 299 167 69.9 3.9 F [1,25] 1) 115.1*** 42.5*** 37.2*** 2.4 35.6 »**
1-- 3 3 383 402 5 625 497 120 15.0 7 867 752 17.6 4.1 4-- 6 3 529 210 4 627 297 126 13.1 5 923 440 t2.0 1.1 7-- 9 3 222 150 4 380 181 129 11.5 5 537 262 14.4 1.4
10--12 2 633 164 3 067 217 117 43.2 3 500 275 9.6 5.5 13--15 2 194 177 2 969 186 237 48.2 3 744 323 8.7 2.3 16--18 1 931 102 2 617 134 156 21.6 3 303 226 29.7 7.2 F [1,14] 1) 55.5*** 99.3*** 4.8* 56.5*** 0.2
Comparison between regression lines (analysis of covariance):
Slopes: F [1,39] 8.5** 3.9 16.9 **'~ Elevations: F [1,40] -- 90.9*** --
m
1) Linear regression between mean values per nest per day and ne~tling ag~ » p <0 .05 ; *«'~ p <0.01; '«*«~ p <0 .001 .
Heft 2] 1991 j
Calls/s 1.4
1 . 2 !
o.8- ~~~~~~!!~!i~i!i!iii~
0 . 6 -
0.4 ~~~i!ii~i:~iiii~;~~
0 . 2 - i;i;i;;iiiiiiil;~
0 1-3
Vocai ontogeny in Pica •ica 151
~i~ii;iiiiiiiiii 4 - 6
i:% ::)
i : : 4 / :
li:i:i:?~:i:;;i ~
15 ~: ::
ii!il, iii,lii~ii~
13-14 7-9 10-12
Nestling Age (days)
Fig. 4. Adjusted mean values of begging rates while 'contr0lling for the effect of stimulation rate as a covariate Lower values correspond to feeding sessions after ä 1 h period of food deprivation, higher ones to those after a 2.5 h period (WiLcoxo> ~ test for comparison of begging rate/stimulation rate between periods, 1 - 3 days: Z = 1.89, p <0.05; 4 - 6 days: Z = 4.45, p <0.001; 7 - 9 days: Z = 5.83, p <0.001;
10--12 days: Z = 2.96, p <0.01; 13-14 days: Z = 2.45, p <0.01).
Begging Scream
W h e n nestl ings were abou t to leave the nest, Begging Trills changed into Begging Screams w i th in a couple of days. The t rans i t ion between b o t h calls is a gradual process, w i th calls of in termediate s t ructure appear ing in between (Fig. 5). Begging Screams of
10
8
6
Frequency (kHz)
~t ù " ' - . - . . : .
' I
i
2
: . & .~ * I " ~ r
; ",,4} h
I' . « ¢
I ,
it0 ~
} " / / ' i :
' I
0 I J
0 0.5 1
Tirne (s) Fig. 5. (Ldt) Two Begging Screams of fledglings and (right) two transitiona] forms between Begging
Screams and Begging Trills.

152 T. REDONDO [ J" Orn. [ 132
fledglings were of a higher amplitude and shorter duration than nestling Trilts and lacked their characteristic FM and energy dispersion. In the laboratory, the transition between calls was a rather sudden event. Typically, a nestling, which previously had not given any signs of change, started to produce Screams prior to the first feeding of the day or after a 2.5 h period of food deprivation. These calls contained a large amount of intermediate transitional forms. Dates of first appearance of Begging Screams were closely associated with fledging &tes (Fig. 6).
Soft Whistle
This vocalization was first recorded when nesttings were 1--2 days old but became much more frequent between 5 and 11 days. Soft Whistles were given by nestlings while in resting postures and were especially common just after bouts of begging. They could be easily elicited by täctile or auditory stimuli coming from myself or nestmates. Like Begging Calls, Soft Whistles had a tonal structure and sounded like brief, musical notes of very low amplitude Throughout the nestling period, intermediate gradual calls between Soft Whistles and Begging Trills could be observed during a recording session but such calls were not included in the analyzed samples. Soft Whistles had a lower pitch and were shorter than Begging Trills, an d typically lacked their intense FM and broad frequency-range (Fig. 7; Table). In the wild, Soft Whistles were not recorded after 17 days of ag« Hand-reared nestlings continued to give brief, soft and low-pitched calls after begging bouts well after this age but these calls readily graded into Begging Calls similar to those given by nestlings with a low hunger motivation. Consequently, I concluded that Soft Whistles represent a characteristic vocalization in the early nestling period.
36
34
32
30
28
26
24
22
20
18 2O
Transi t ion (days) [3
[3
_ _ I i I I _ I
22 24 26 28 30 32 34
Fledging age (days)
Fig. 6. Age of first appearance of transitional forms between Begging Trills and Begging Screams plotted against fledging age for 19 laboratory-reared nestlings.

Heft 2 ] 1991 j Vocal ontogeny in Pica pica
Frequency (kHz) 10,
6~
4 ! "
I 0 L
ù
ù :~" " - " V
0 0.5
Time (s)
153
Frequency (kHz) lOt
8 ) . "
6 i :.-''"~ :
41 "".": .«, !
7
ù
0 0.5 1 Time (s)
• : : ! .
.. : ' . , . . . : , ~
Fig. 7. Upper: Soft Whistles of nestlings (left to right) 2, 5, 5, 8 and 11 days oid. Lower: Soft Whistles of nesdings 11, 14, 14 and I7 days old.
I considered three hypotheses for the functional significance of this call:
A) Soft Whistles act as a brooding solicitation signal, i.e they advertise to parents about the cooling of nestlings which are still unable to thermoregulate This hypothesis was supported by the fact that Soß Whistles were triggered by tactile stimuli. In addition, their frequency of occurrence in the wild dropped around the age endothermy develops (11 days) (Fig. 8).
B) Soft Whistles are a satiation signal, as was suggested by the fact that they were fre- quently uttered after, and graded with, begging bouts.

154
100
T. REDONDO
Percentage of Nests
[ J. Orn. 132
80
6O
40
20 i
0 L
/ TR
-->
2 5 8 11 14 17 20 23 26 Nestling Age (days)
Fig. 8. Percentage of nests of a given age in which nestlings uttered Soft Whistles (N = 24). The arrow points to the age at which nestlings show the highest daily gain in thermoregulatory ability.
C) Soft Whistles advertise to parents that nestlings are alive and in good condition. Physical activity of nestlings may provide clues about their condition, but its {requen- cy of occurrence is probably limited due to energetic constraints. This hypothesis was supported by the fact that Soft Whistles were elicited by stimuli coming from the caretaker, mainly &ring periods of nestling inactivky. In addition, those nestlings which died ffom pathological processes in the laboratory, completely ceased vocaliza- tions &ring the period they showed symptoms of illness.
These hypotheses predict that emission rates of Soff Whistles shonld increase with (i) a decrease in nestlings' body temperature (hypothesis A); (il) a decrease in hunger motivation at a constant temperature (hypothesis B); and (iii) offspring's awareness of the presence of a caretaker at the nest when both temperature and hunger motivation are held constant (hypothesis C).
Hypothesis A was rejected after I observed that &ring the cooling sessions for &ter- mining thermoregulation, the utterance of Soft Whistles was negligible, i.a cooling in h ib i t ed vocalizations. In order to test hypothesis B, I calculated calling rotes of nestlings 5 - 9 days old at nest temperature during 2 min of observation at 20, 40, and 60 min after they received the first feeding of the day. Fig. 9 shows that, contrary to the prediction, an increase in hunger motivation actually resulted in an increase in call- ing rates. Finally, I also calculated calling rates of nestlings 6 - 7 days old at nest temperature, 40 min after they had received the second feeding of the day, &ring two 2 min periods before and after 10 s of acoustic stimu]ation (softly talking to nestlings). Stimulation caused calling rates to increase from 0.70 -+-0.43 SE to 6.98 4-1.05 SE calls per nestling per minute (WmcoxoN test, T = 101.2, p < 0.01, N = 14). Hypothesis C also accounts for the fact that Soft Whistles are given mainly between begging bouts

Hef t 2 ] 1991 J Vocal on togeny in Pica pica 155
and at early ages, for they should be prevalent precisely when motor activity is low or poorly developed.
Distress and Alarm Calls
The first truly harsh calls which appeared in the repertoire consisted of Distress Catls given by rlestlings at the age of 17 däys. These and subsequent calls were uttered when nestlings were seized or handled by us. Fig. 10 shows that energy is distributed through a wide frequency fange but it concentrates around 3 kHz.
The occurrence of Distress Calls reaches a maximum around 20 days and declines thereafter. This is a consequence of a sudden shift in the anti-predator response of nest- lings which occurs at this age Chicks younger than 20 days typically respond to an intrusion at the nest by crouching and freezing, a passive response which appears when nestlings are 14 days old (R~DONDO & CARI~ANZA 1989). At about 20 days, nestiings develop an active response to intruders which consists of jumping out of the nest while giving Alarm Calls (Fig. 11). This response rnay result in prernature fiedging if nestlings fall down frorn the nest tree. Alarm Calls are also genuine harsh vocalizations given in series of 1 - 4 brief syllables (Fig. 10). Compared with Distress Calls, they show a rnore uniform pattern of energy distribution, with arnplitude peaks around 3, 6 and 9 kHz.
Distress and Alarm calls appear de novo during development. They show no transi- tional forms with any other call in the repertoire, not with each other.
Calls/nestling/s 5,
4 L
Fig. 9. Mean (SE) rates of emission I of Soft Whistles by nestlings 5--9 1 [ days old at 20, 40 and 60 min after they had received the first feeding in the day (KRusKA>WALHS test, H d = 5.5, df = 2, p = 0.06; SeEAR- 01 ~IAN'S correlation coeficient, r =
0.3i, p <0.01, N = 105).
2 0 :~~!i:!i ii{i!i!~ 4 O
!iiiiii i ::: : ::i
¸¸:<5¸1
!~!iii!!! iiiii~il ....
5::q:!
iiii!ili i i i i i i i l l
6 0
Fasting Tirne (min)

156 T. R E D O N D O
20
1 5
10
Frequency (kHz) ù
{.- , ù.~
ù,
' : " : ~F- " "
~f~~
.; ,. ,
, . ' . t " . ' , l , ' . "
, : 2~i.~~ '~"
I
0 0.5
Frequency (kHz) 2O
i ~':
15 L i ~ , !t B .% « . i~!
~, :7 ;i
Or ~
20
1 5
~- , , : ' . . ; " .
% " a : d ' . " ?~ "" :'~ "
• « ' Ü"!: 77:.~ ~y..;ù ;.;,~?~. ~ .~~'~~
10
1 1.5
,:»,t. -»'
i~i;" .:~«: ': ù.'[i
• :.,. '.~ :,;
';.::. "_.;,
( N { : ' ;
t
I I
0 0.5 1 1.5
Frequency (kHz)
v..
~;}»ù-~. O - -
~-~,, 1' !i
. «:~!..,
*«Z»~
I
ù
i - .
:;fr,Z.
~~~,
~ i ~ : " I
.[
: N ? : : a~
Time (s)
Time (8)
0 0.5 1 1.5 Time (s)
[ ]-. O r n .
132
Fig. 10. l-larsh vocalizations of fledglings. Upper: four Distress Calls; middie: four Alarm Catls; lower: five Brief Contact Notes.

Heft 2 ] 1991 J Vocal ontoge W in Hca pica
Percentage of Nests ! ,00!
8 0 ! •
00l ! 40
157
20
2 5 8 11 14 17 20 23 26 29
Nestling Age (days)
Fig. 11. Percentage of nests of a given age in which nestlings uttered Distress Calls (lines) or Alarm Calis (bars) (N = 51).
Both Distress and Alarm calls were of high amplitude and triggered intense defensive responses in adult birds, especially Alarm Calls. In 4 out of 9 cases, patents increased their scolding rates and approached closer to the nest after nestlings gave only Distress Calls. The comparative figure for Alarm Calls was an increase in defence intensity in 15 out of 18 cases (FIsHER's test, p = 0.052).
Brief Contact Note
This is a harsh, low-pitched call of very low amplitude given by fledglings. Fig. 10 shows that the energy is distributed rather uniformly throughout a frequency fange of 0.5-- 10 kHz. The onset of the call is abrupt, but energy vanishes smoothly at the end of the call. Like anti-predator calls, Brief Contact Notes appear de novo, without evident transitional forms with preceding calls.
This call was rarely observed in the wild, but laboratory-reared fledglings gare it very frequently, although its occurrence was restricted to calling bouts associated with two behavioural contexts: exploration and play. Exploration and walking was the most fre- quent context of calling by solitary fledglings. Social play episodes (including hopping, wing fluttering, chasing and pecking others' tail) also included high rates of vocaliza- tion.
Discuss ion
Five out of the six calls which compose the vocal repertoire are linked either to a context related to feeding (two begging calls and Soft Whistles) or anti-predator defence (Alarm and Distress Calls). These are precisely the two contexts in which patents allo- cate most of their effort. Compared with other parental activities (like brooding and

158 T. REDONDO [ J' 132Orn"
nest sanitation), feeding nestlings and defending them against predators monopolize most of the risk, time and energy budgets of patents (O'CoNNOk 1984), their relative contribution to parental duties increasing as the offspring grow older (BuITRON 1988; REDONDO & CARRANZA 1989). Greater allocation of parental effort towards feeding and protecting nestlings has probably evolved to compensate for the two major eco- logical sources of nestting mortality: starvation and predation (RIcKLE~S 1969). In my study population, about 26 % of the hatched nestlings died from starvation (within- brood losses) and 34 % were killed by predators (N = 266).
The above considerations suggest that nestling vocalizations may have evolved in response to direct ecological pressures, hence their structural design is expected to be molded by forces related to the ecology of sound propagation. The structure of Begg- ing Trills seems to be a compromise between the neccessities for nestlings to make themselves conspicuous to patents and cryptic to non-intended receivers, like predators (REI3ONI)O & AI~IAs ~E REYNa 1988). A wide frequency fange may help begging calls to be easily located (ROOKE & KNIGHT 1977). In addittion, such calls show a high degree of spectral complexity, caused by energy dispersion and FM, which facilitates their degradation into the environment (R*cHaP, DS & WiteY 1980; COSeNS & FALLS 1984). Age-related changes in the structure of Begging Trills involve an increase in both energy dispersion and FM which is likely to compensate for the more detectable (louder, longer and lower-pitched, MaRTEN & MAI~LER 1977) calls uttered by older nestlings (REDONDO & EXPOSlTO 1990).
When nestlings leave the nest, a sudden (albeit continuous) change occurs in the structure of begging calls. Fledgling Begging Screams are rauch shorter and louder than nestling Begging Trills and lack their characteristic dispersion of energy. The higher loudness and tonal quality of Begging Screams make them less prone to environmental degradation; hence they retain their structure at longer distances than Begging Trills (RICHARDS ~X~ WILEY 1980). Such a design makes sense in the light of parent-fledgling relationships. Shortly after fledging, chicks await concealed in vegetation for patents to come with food (BuITRON 1988) but the location of the group of fledglings may change due to spontaneous mobility or as a consequence of disturbances. Fledgling magpies beg for food from a W other adult magpie coming nearby but parents recognize and selectively feed their own offspring (LiNsDaLE 1937), presumably on the basis of an acoustic signature system.
A sudden change in the structure of begging calls at fledging time is a widespread phenomenon. Seven studies (MARLER 1956; THOMPSON & RtCE 1970; ZaNN 1975; WINKLEI~ & SHORT 1978; WILKINSON 1980; BEECHER et al. 1981; HOWES-JONES 1984) make explicit that fledgling begging calls gain in tonal quality with respect to preceding nestling calls. The opposite trend is found in two studies (McLaREN 1976; THOMPSON 1976) while others do not specify the structural changes accompanying the transition between calls (MESSMER & MESSMER 1956; HORWlCH 1969; MOREHOUSE & BREWER 1968; MARL~R & MUNDINGER 1975; RUSSELL & BALDA 1978; ELSACKER et al. 1986; more references in HAt<PER 1986). It is also interesting to note that in species in which acoustic recognition of the offspring is well developed, signature calls are precisely

Heft 2] Vocal ontogeny in Pica pica 159 1991 /
fledgling begging calls (RusSEL> & BALDA 1978; BEECHER et al. 1981; ELSACKER et al.. 1986).
There is little doubt that anti-predator nestling calls have evolved as a trait which ensures parental protection (KLuMP & SHALTER 1984). Theory predicts that parental care should be allocated in direct proportion to the relative reproductive value of the offspring (MI>INSKI 1978). Reproductive value of nestlings increases during the nesting cycle (ANDERSSON et al. 1980) and especially shortly before fledging, when chicks develop the ability to escape by themselves from predators (REDONDO 1989). The sen- sory and motor coordination neccessary to perform a successful escaping response im- proves greatly between 14 and 20 days, just before the appearance of Alarm Calls. By means of Alarm Calls, nestlings might advertise to their parerlts about their novel gain in reproductive value (REDONDO & CARRANZA 1989). In accordance with their func- tion of attracting patents to the nest, anti-predator calls seem to be designed for facilitating detection and location of the source. Both a high amplitude and a concen- tration of energy in low frequencies improve signal detection (MARTEN & M•RLER 1977). Distress Calls show a clear energy peak around 3 kHz, which corresponds to the point of maximal acoustic sensitivity in adults (RoBERTS et al. 1983) and they are efficient over long distances (BREMONI) & AUt~~N 1990). Alarm Calls have a high fre- quency fange, an abrupt onset and a repetitive structure, all features that make them easily located (ROOKE & KNIGHT 1977). Since Alarm Calls are given in connection with evasive behaviour, this feature may facilitate a rapid location of endangered nestl- ings by adults.
The structure of Brief Contact Notes seems to provide clues for allowing source location (abrupt onset and wide frequency fange) at short distances (low amplitude and dispersive structure). Brief Contact Notes probably help to maintain the cohesion between siblings during episodes of locomotory activity (like exploration and play), since visual contact is limited within vegetation.
Finally, Soft Whistles function as a kind of very short-range communication. Unlike begging calls, they are not the result of sibling competition for the attention of parents (REDoNDO & AI~IAS DE REYNA 1988). Consequently, they seem to be barely locatable (narrow frequency fange) and detectable (low amplitude). Calls probably analogous to magpie Soft Whistles are described by THOMPSON & RICE (1970) and W*NKLER & SHORT (1978).
Developmental variations in the vocal repertoire can be summarized as follows. Ear- iy stages are characterized by tonal calls, while late (ca. fledging) phases include noisy calls similar to those of adult birds. The four fledgling calls remain in the aduk reper- toire with minor modifications (REDONDO & EXPOSITO 1990). Noisy calls become fre- quent around 18 däys. Before this age, the most frequent vocalizations (Begging Trilts) undergo structural changes which make them increasingly no{sier as a consequence of intense FM. This sequence of events could be interpreted as if structural changes (FM) in Begging Trilts provide nestlings with a mechanism by which they acquire the skill for producing adult noisy calls. However, a detailed analysis of energy dispersion in Begging Trills caused by FM showed that such a process is not probably involved in

r j. Orn. 160 T. REDONDO !
' 132
the production of noisy adult calls but probably reduces chances of nestiings being detected by potential predators (REDONDO & EXPOSITO 1990). In addition, begging calls change their structure at fledging time in a way that can hardly be interpreted in developmental terms (they lack the noisy features of preceding nestling calls) but which is consistent with the novel ecological requirements of fledglings.
The hypothesis that nestling calls act as precursors for late vocalizations is also faced with the problem of discontinuities in the developmental sequence Both Distress and Alarm Calls and Brief Contact Notes appear de novo, without transitional forms with preceding calls. Also, they are probably controlled by a different set of motivational factors. This is especially clear in the case of begging and anti-predator calls, since those stimuli which dick fear (crouching) responses actually supress begging (RYDEN 1978; BISCHOF & LASSEK 1985).
The sudden appearance of vocalizations during development has been widely reported, both in altricial (MARLER 1956; MESSMER & MESSMER 1956; THOMPSON ~x~ R~CE 1970; HOWES-JONES 1984) and precocial species (WILKINSON & HUXLEY 1978; COSENS 1981). A continuous pattern of late call development starting from begging calls is orten invoked (MARLER & MUNDINGER 1975; ZANN 1975; GEBAUER 1984; HOW~S-JONES 1984; ]~ARLE 1986) but, with a single exception, none of these studies provides supporting evidence Transitional forms between begging and distress calls have been described in the grassfinch Poephila cincta (ZANN 1975). Note, however, that this single case leaves open the question of the nature of the motivational change underlying the transition.
In fact, the transition between begging calls at fledging is the only instance of call ontogeny which fulfills the double criterion of structural and rnotivational continuity. With respect to the remaining fledgling calls, the rule is a sudden appearance of several call types once the nestlings have left the nest. Such a 'vocal diversification at fledging' is also widely reported (MEsSMER & MESSMER 1956; THOMPSON & RICE 1970; BALPH 1975; ZANN 1975; SMITH 1977; HOWES-JONES 1984). But, contrary to what is observed at earlier stages of development, there is good evidence for the existence of a con- tinuous pattern of development of adult calls starting from fledgling calls (MARLER 1956; THOMPSON & ~CE 1970; ZANN 1975 and refs. therein; SMITH 1977; HOWES- JONES 1984). Indeed, adult calls in their definkive form may already be present in the repertoire of fledglings (MEssMER & MESSMER 1956; THOMPSON & RICE 1970; ZANN 1975; THOMPSON 1976; HOWES-JONES 1984). This suggests that neural mechanisms of some displays may be functional at an early point in ontogeny (SCHLEIDT & SHALT~R 1973; GROOTHUtS 1989), as in the well-known study of ANDREW (1966) about testo- sterone-induced crowing in young chicks. What is relevant for our discussion is that such findings question whether birds require precursors which are functional at nestl- ing stages for developing adult calls at all.
S u m m a r y
The vocal repertoire of magpie (Pica pica) chicks consists of six calls: Begging Trill (BT), Soft Whistle (SW), Begging Scream (BS), Alarm Call (AC), Distress Call (DC) and Brief Contact

Heft 2 ] 1991 J Vocai ontogeny in Pica pica 161
Note (BCN). Both BT and SW have a tonal structure and their occurrence is restricted to the nestling period. At fledging, there is a gradual change from BT into BS and a sudden ap- pearance of harsh calls similar to those of adult birds (AC, DC, BCN), without evident transi- tional forms with preceding tonal calls. Both the existence and the structural design of calls seem to be adapted for providing nestlings with immediate benefits linked to the two major chapters of allocation of parental care. Emission rates of BT increase with hunger motivation under laboratory conditions. Their structure suggests that they are easily located but liable to suffer from environmental degradation. BS of fledglings may be more resistant to degradation, a trait which may facilitate the identification by parents of their own offspring. Both AC and DC attract patents to defend the nest against potential predators, and their structure make them to be easily located and detectable at long distances. BCN are given by fledglings during bouts of locomotory activity (exploration and play) and they probably help in maintaining the cohesion of the group under conditions of poor visibility. In accordance, this call may be fairly located at short distances. The function of SW was unclear. It is given during periods of nestling inactivity between begging bouts, and could be easily elicited by tactile and auditory stimuli. After laboratory experiments, it is concluded that SW serve to indicate parents that nestlings are in good condition, hence to benefit from the parental willingness to invest in a brood with high prospects of survival. Since (i) there is a widespread lack of con- tinuity in the development of adult vocalizations starting from nestling calls, and (ii) nestling calls seem to have evolved to provide birds with benefits in the short-term, these facts argue against the prevailing idea that the main function of calls early in ontogeny is to act as precur- sors of adult vocalizations.
Zusammenfassung Das Lautrepertoire von Elsternestlingen besteht aus 6 Lautäußerungen: Betteltrillern (BT),
Sanftes Pfeifen (SP), Bettelkreischen (BK), Alarmruf (AR), Angstschreien (AS) und kurzer Kontaktruf (KP). BT und SP sind tonal und treten nur während der Nestlingsperiode auf. Zum Zeitpunkt des Ausfliegens geht BT graduell in BK ~iber und plötzlich treten auch ohne vorausgehende tonale Übergangsformen rauhe Rufe ähnlich denen der Altvögel auf (AR, AS, KR). Sowohl die Existenz als auch die Struktur der Lautäußerungen scheinen als Anpassungen im Zusammenhang mit einer Optimierung der Brutpflege zu interpretieren zu sein. Die Emis- sionsrate von BT steigt unter Laborbedingungen mit der Hungermotivation. Die Struktur legt nahe, daß BT leicht geortet werden kann, aber auch leicht von der Umgebung verschluckt wird. BS der flüggen Jungen scheint davon weniger betroffen zu sein, so daß die Eltern leichter ihre Jungen identifizieren können. AR und AS veranlassen die Altvögel, ihr Nest gegenüber potentiellen Prädatoren zu verteidigen; die Struktur der Laute begünstigt ihre Ortung und Wahrnehmung über größere Entfernungen. KR werden von den flüggen Jungen bei lebhafter lokomotorischer Aktivität (Exploration, Spiel) geäußert; sie tragen vielleicht dazu bei, die Gruppe auch unter erschwerten optischen Kontaktmöglichkeiten zusammenzuhalten. Die Funktion von SP blieb unklar. SP war während inaktiver Perioden zwischen Bettelverha]ten zu hören und konnte leicht durch taktile und akustische Reize ausgelöst werden. Nach Labor- experimenten ist zu schließen, daß SP den Eltern ~Wohlbefinden" der Jungen anzeigt und so Bereitschaft zur Investition in eine Brut mit hoher Überlebenswahrscheinlichkeit fördert. Daß (1) zwischen den Rufen der Altvögel und dem Repertoire der Nestlingen weitgehend keine kontinuierlichen Übergänge festzustellen und (2) Nestlingsrufe offensichtlich im Hinblick auf kuzfristige Gewinne zu interpretieren sind, spricht gegen die allgemein vorherrschende An- sicht, das Lautrepertoire in frühen Stadien der Ontogenie sei hauptsächlich ein Vorläufer der Adultlaute.

162 T. REDONDO [ J" Orn. k 132
Literature
ALBERTS, J. R., & C. P. CRaME~ (1988): Ecology and experience. Sources of means and mean- ings of developmental change. In: E. M. BLass, Handbook of Behavioral Neurobiology 9: 1--39. N. Y. * ANDERSSON, M., C. G. WIKLUND, ~X~ H. RUNI)CREN(1980): Parental defence of offspring: a model and an example. Anim. Behav. 28: 536--542. • ANDREW, R. J. (1966): Precocious adult behaviour in the young chick. Anim, Behav. 14: 485--500. • BALPH, M. H. (1975): Development of young Brewer's blackbirds. Wilson Bull. 87: 207--230. * BEECI~ER, M. D., I. M. BEECHEa, & S. HAHN (1981): Parent-offspring recognition in bank swallows (Riparia riparia): II, Development and acoustic basis. Anim. Behav. 29: 95-101. ® BEKOFF, M. (1985): Evolutionary perspectives of behavioral development. Z. Tierpsychol. 69: 166-- 167. * BiscHoF, H. J., & R. LASSEK (1985): The gaping reaction and the development of fear in young zebra finches (Taeniopygia guttata castanotis). Z. Tierpsychol, 69: 55--65. « BP, EMONI), J. C., & T. AUBIN (1990): Responses to distress calls by black-Eeaded gulls, Larus ridibundus: the role of non-degraded features. Anim. Behav. 39: 503--511. • BUITRON, D. (1988): Female and male specialization in parental care and its consequences in black-billed magpies. Condor 90: 29-39. t CHiSZaR, D. (1985): Ontogeny of communicative behaviors. In: E. S, GOLLIN, The Comparative Development of Adaptive Skills: Evolutionary Implica- tions: 207--238. Hillsdale • CoseNs, S. E. (1981): Development of vocalizations in the American Coot. Can. J. Zool. 59: 1921-1928. • Ditto & J. B. FALLS (1984): A comparison of sound propagation and song frequency in temperate marsh and grassland habitats. Behav. Ecol. Sociobiol. 15: 161--t70. • ELsacr~a, L. VaN, R. PINXTEN, & R. VERHEYEN (1986): Parent-offspring recognition in starlings (Stur*zus v. vulgaris). In: L. PASSERA, & J. P. LaCHAUD, The Individual and Society: 73-78. Toulouse. • EaaLE, R. A. (1986): Vocalizations of the South Affican Cliff Swallow Hirundo spilodera. S. Afr. J. Zool. 21: 229-232. * GEBaUER, A. (1984): Die Lautentwicklung beim Buntspecht, Dendrocopos major. Ann. Orn. 8: 107--127. • GOMeNDIO, M. (1988): The development of different types of play in gazelles: implications for the nature and functions of play. Anim. Behav. 36: 825--836. • GROOTHUIS, T. (1989): On the ontogeny of display behaviour in the black-headed gull: I. The gradual emergence of the aduk forms. Behaviour 109: 76--124. • Ha~ptG A. B. (1986): The evolution of begging: sibling competirion and parent-offspring conflict. Am. Nat. 128: 99-114. • HORWlCH, R. H. (1969): Behavioral ontogew of the mockingbird. Wilson Bull. 91: 434---440. • Howes-JoNEs, D. (1984): The vocal behaviour of young warbling vireos. Can. J. Zool. 62: 205-230. • KLUMP, G. M., & M. D. SHa5~reR (1984): Acoustic behaviour of birds and mammals in the predator context: I. Factors governing the structure of alarm dgnals. II. The functional significance andevolution of alarm calls. Z. Tierpsychol. 66: 189--226. • LaNYON, W. E. (1960): The ontoge W of vocalizations in birds. In: W. E. LaNYON, & N. N. TaVOLCA, Animal Sounds and Communication: 321-347. Publ. No. 7. Am. Inst. Biol. Sci. Washington. • Dit- to (1979): Development of song in the wood thrush (Hylocicbla mustelina), with notes on a technique for hand-rearing passerines ffom the egg. Am. Mus. Novit. 2666: 1--27. • LE MACNEN, J. (1983): Body energy balance and food intake: a neuroendocrine regulatory mechanism. Physiol. Rev. 63: 314--386. • LII'VSDALE, J. M. (1937): The natural history of magpies. Pac. Coast Avif. 25: 1--234. • McLa~EN, M. A. 1976. Vocalizations of the boreal chickadee. Auk 93: 451--463. • MARLER, R (1956): The voice of the chaffinch and its func- tion as a language. Ibis 98: 231--261. • Ditto & R C. MUNDINCER (1975): Vocalizations, social organization and breeding biology of the twite (Acanthis flavirõstris). Ibis 117: 1--17. • Ditto & S. PET~aS (1982): Subsong and plastic song: their role in the vocal learning process. In: D. E. KaooI)sva, & E.H. MI>>ER, Acoustic communication in birds Vol 2, 25--50. N. Y. • MaRTEN, K., & R MaRLEP, (1977): Sound transmission and its significance for animal

Heft 2 ] 1991 ] Vocal ontogeny in Pica pica 163
vocaiizations. I. Temperate habitats. Behav. Ecol. Sociobiol. 2: 271--290. ® MEJNeRT, U., & H.-H. BERCMANN (1983): Zur Jugendentwicklung der Lautäußerungen beim Birkhuhn (Tetrao tetrix). Behaviour 85: 242--259. • M~SSMER, E, & E. M~SSMeR (1956): Die Entwicklung der Lautäußerungen und einiger Verhaltensweisen der Amsel (Turdus rner~¢la merula L.) unter natürlichen Bedingungen und nach Einzelaufzucht in schalldichten Räumen. Z. Tierpsychol. 13: 341--441. • MILINSKI, M. (1978): Kin selection and reproductive value. Z. Tierpsychol. 47: 328--329. • MORe~OUSE, E. L., & R. BREWER, (1968): Feeding of nestling and fledgling eastern kingbirds. Auk 85: 44--54. • MO'CMHaN, M. (1962): Hostile and sexual behavior pat- terns of South American and Pacific Laridae. Behaviour Suppl. 8: 1--365. • O'Co?,NOR, R. J. (1984): The growth and development of birds. Chichester. • OPeENHHM, R. W. (1980): Metamorphosis and adaptation in the behaviour of developing organisms. Devi. Psychobiol. I3: 353--356. • REDONDO, T. (1989): Avian nest defence: theoretical models and evidence. Behaviour 111: 161--195. • Ditto & L. ARIAS De REYNa (1988): Locatability of begging calls in nestling altricial birds. Anim. Behav. 36: 653--661. • Ditto & J. CARRa>aza (1989): Oft- spring reproductive value and nest defense in the magpie. Behav. Ecol. Sociobiol. 25: 369--378. • Ditto & E ExPOSlTO (1990): Structural variations in the begging calls of nestling magpies (Pica pica) and their role in the development of adult voice. Ethology 84: 307--318. • RICHAI~DS, D. G., & R. H. WILEY (1980): Reverberations and amplitude fluctuations in the propagation of sound in a lotest: implications for animal communication. Am. Nat. 115: 381--399. • R~CKLEFS, R: E. (1969): An analysis of nesting mortality in birds. Smithson. Contr. Zooh 9: 1-48. • Ditto (1983): Avian posmatai development. In: D. S. FARNES, J. R. KINo, & K.C. PAaK~S, Avian Biology 7: 1--83. N. Y. • ROBERTS, J., MOReNO, A., KaCELNIK, A., & M. HUNTeR (1983): Physical considerations in the frequency limits of birdsong. Acoustic Letters 6: 100--105. • Roo~E, I. J., & T. A. K>~ICHT, (1977): Alarm calls of honeyeaters with reference to locating sources of sound. Emu 77: 193-- 198. • RUSSELL, P., & J. H. BaLI)A, (1978): The care of young pifion jays and their integration into the flock. J. Orn. 119: 1 4 6 - 1 7 1 . . RYI)EN, O. O. (1978): Differential responsiveness of great tit nestlings, Parus major, to natural auditory stimuli. Z. Tierpsychol. 47: 236--253. ® SCHLHDT, W. M., & M. D. SHALTE~(1973): Stereotypy of a fixed action pattern during ontogeny in Coturnix cotur- nix coturnix. Z. Tierpsychol. 33: 35--37. • SVITH, S. M. (1977): The behavior and vocaliza- tions of young turquoise-browed motmots. Biotropica 9: 127-- 130. • Ditto (1983): The on- togeny of avian behaviour. In: D. S. FAt<N~R, KING, J. R., & K. C. PaBKES, Avian Biology 7: 85--160. N. Y. * THo_~tPso~, W. L. (1976): Vocalizations of the lazuli bunting. Condor 78: 195-207. • Ditto & J. O'H. R,c~ (1970): Calls of the indigo bunting, Passerina eyanea. Z. Tierpsychoh 27: 35--46. • THOt<~N, B. ten, & H.-H. BeaCMaNN (1987a): Veränderung und Konstanz von Merkmalen in der jugendlichen Stimmentwicklung der Nonnengans (Branta leucopsis). Behaviour 100: 61--91. • Ditto (1987b): Die Entwicklung der Lautäußerungen bei der Graugans (Anser anser). J. Orn. 128: 181-207. • W~>K*NSON, R. (1980): Calls of nestling chaffinches Fringilla coelebs: the use of two sound sources. Z. Tierpsychol. 54: 346-356. • Ditto & C. R. HUXL~Y (1978): Vocalizations of chicks and juveniles and the development of calls in the Aldabra white-throated rail Dryolimnas cuvieri aldabranus (Aves: Rallidae). J. Zool. Lond. 186: 487--505. • W~NK~~~, H., & L. L. SHOaT (1978): A com- parative analysis of acoustical signals in pied woodpeckers (Aves, Picoides). Bull. Am. Mus. Nat. Hist. 160: 1--110. • ZaNN, R. (1975): Inter- and intraspecific variation in the calls of three species of grassfinches of the subgenus Poephila (Gould) (Estrildidae). Z. Tierpsychol. 39: 85--125. • Zat<, J. H. (1984): Biostatistical Analysis. Prentice Hall, New Jersey.
Author's address: Estación Biológica de Dofiana, C.S.I.C., Apartado 1056, E-41080, Sevilla, Spain





























