Embed Size (px)
Citation preview

© Department of Chemistry, The University of Western Ontario
Chem 2223b Intersession 2008: Amino Acids, Polypeptides, and Proteins
• This chapter discusses the chemistry of amino acids and polypeptides (peptides). Specifically, we will examine the properties and reactions of amino acids, some common structures of proteins, the laboratory synthesis of peptides, how enzymes accelerate reactions, and how proteins are synthesized biochemically.
• Background material, from Chem 2213a or otherwise, that is important includes:
o Properties of weak acids, buffers, and titration curves
o Amines, alcohols, thiols, and carboxylic acids and their derivatives
Trends in the reactivity of acid derivatives
Reactions such as nucleophlic addition and nucleophilic acyl substitution
o Stereochemistry
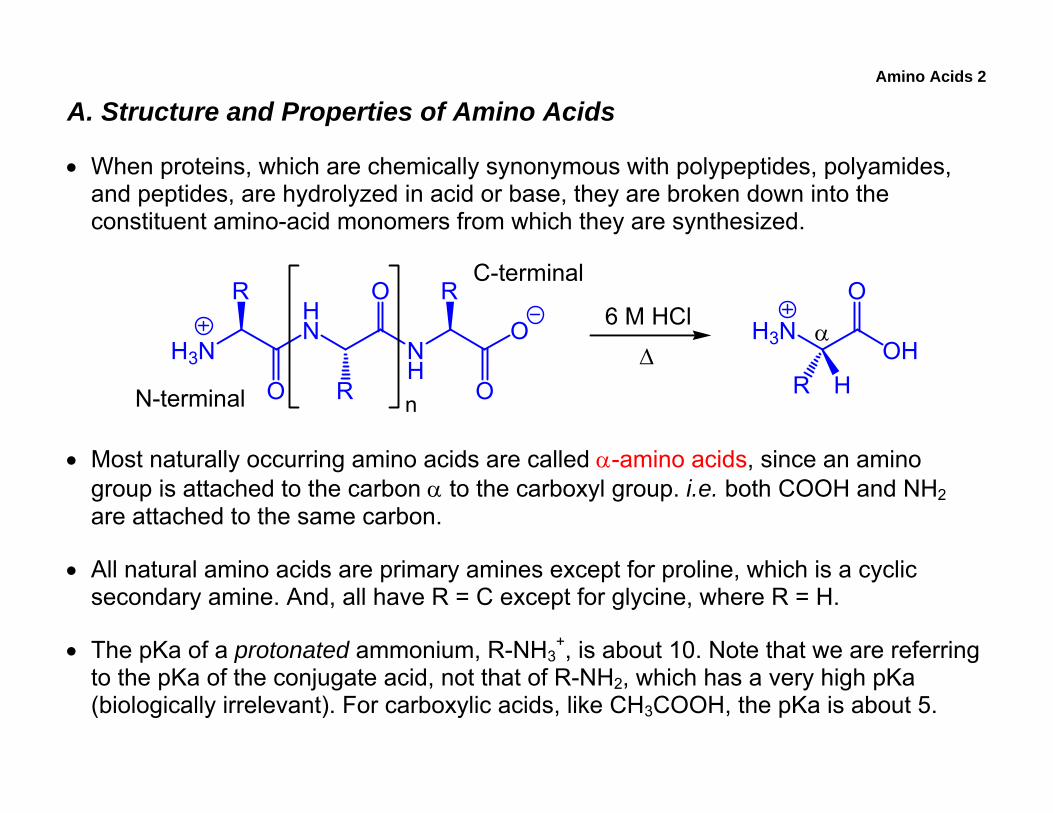
Amino Acids 2
A. Structure and Properties of Amino Acids
• When proteins, which are chemically synonymous with polypeptides, polyamides, and peptides, are hydrolyzed in acid or base, they are broken down into the constituent amino-acid monomers from which they are synthesized.
H3N
HN
NH
R
O
R
O
O
OR n
H3NOH
O
R H
6 M HCl
Δα
N-terminal
C-terminal
• Most naturally occurring amino acids are called α-amino acids, since an amino group is attached to the carbon α to the carboxyl group. i.e. both COOH and NH2 are attached to the same carbon.
• All natural amino acids are primary amines except for proline, which is a cyclic secondary amine. And, all have R = C except for glycine, where R = H.
• The pKa of a protonated ammonium, R-NH3+, is about 10. Note that we are referring
to the pKa of the conjugate acid, not that of R-NH2, which has a very high pKa (biologically irrelevant). For carboxylic acids, like CH3COOH, the pKa is about 5.

Amino Acids 3
1. Acid-Base Properties
• For the dissociation of any weak acid, we can write
HA + H2O A + H3O Keq =[A ] [H3O ]
[HA] [H2O]
Let Ka = Keq × [H2O] then Ka =[A ] [H3O ]
[HA]
• Taking the negative log and rearranging, we have…
pH = pKa + log[Base form]
[Acid form]
• Thus, when pH = pKa, the [A−] = [HA], and this is the midpoint of a titration curve. When pH > pKa, then [A−] > [HA]. Similarly, at pH < pKa, then [A−] < [HA].
• The equation can be used to estimate state of an amino acid at a given pH.
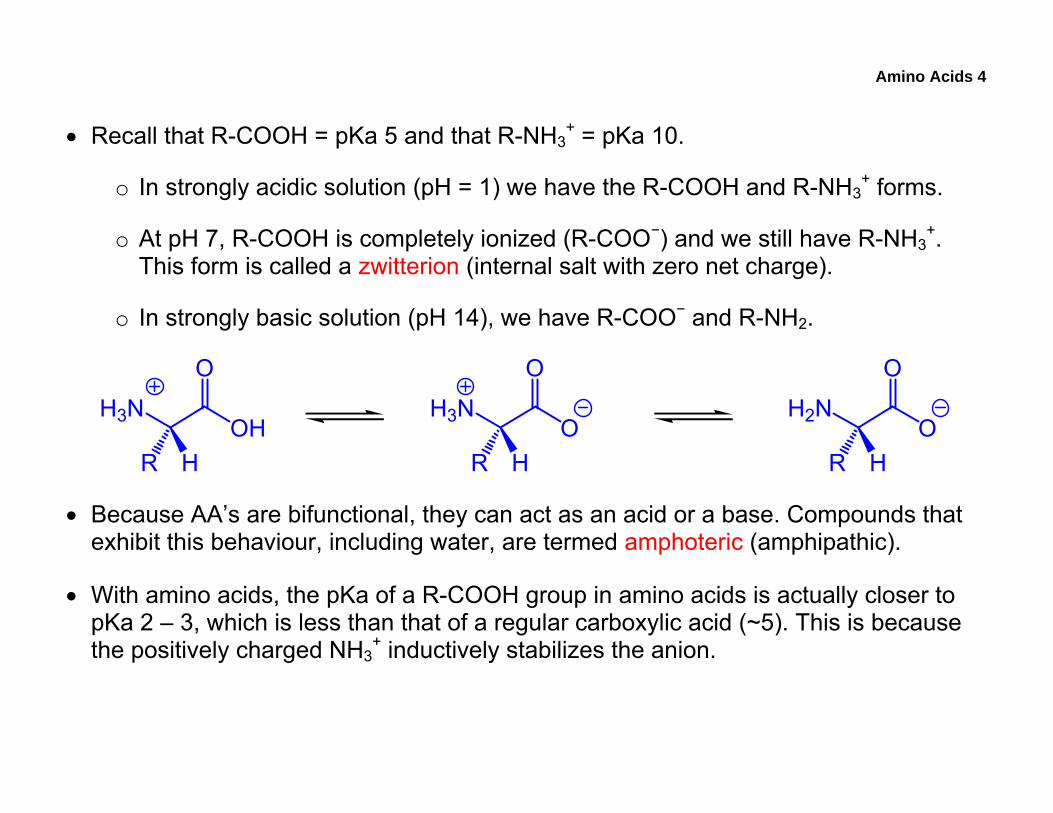
Amino Acids 4
• Recall that R-COOH = pKa 5 and that R-NH3+ = pKa 10.
o In strongly acidic solution (pH = 1) we have the R-COOH and R-NH3+ forms.
o At pH 7, R-COOH is completely ionized (R-COO−) and we still have R-NH3+.
This form is called a zwitterion (internal salt with zero net charge).
o In strongly basic solution (pH 14), we have R-COO− and R-NH2.
H3NOH
O
R H
H3NO
O
R H
H2NO
O
R H
• Because AA’s are bifunctional, they can act as an acid or a base. Compounds that exhibit this behaviour, including water, are termed amphoteric (amphipathic).
• With amino acids, the pKa of a R-COOH group in amino acids is actually closer to pKa 2 – 3, which is less than that of a regular carboxylic acid (~5). This is because the positively charged NH3
+ inductively stabilizes the anion.
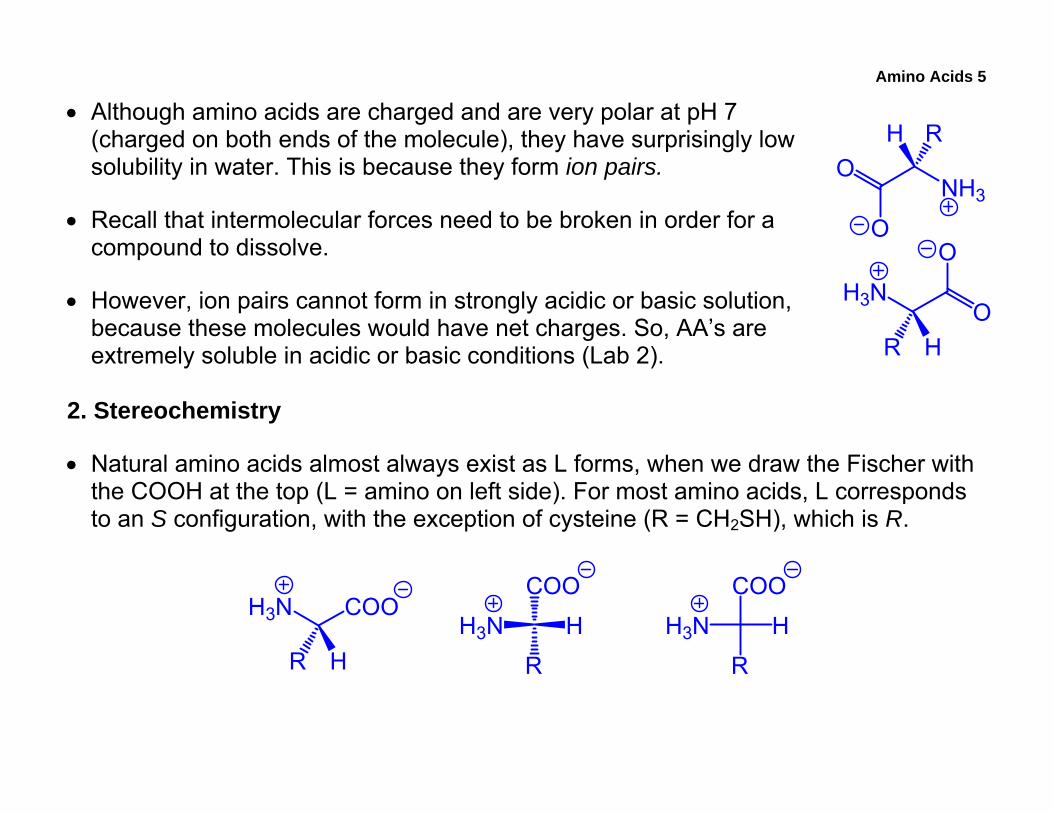
Amino Acids 5
• Although amino acids are charged and are very polar at pH 7 (charged on both ends of the molecule), they have surprisingly low solubility in water. This is because they form ion pairs.
• Recall that intermolecular forces need to be broken in order for a compound to dissolve.
• However, ion pairs cannot form in strongly acidic or basic solution, because these molecules would have net charges. So, AA’s are extremely soluble in acidic or basic conditions (Lab 2).
2. Stereochemistry
• Natural amino acids almost always exist as L forms, when we draw the Fischer with the COOH at the top (L = amino on left side). For most amino acids, L corresponds to an S configuration, with the exception of cysteine (R = CH2SH), which is R.
H3N COO
R HH3N
COO
H
R
H3N
COO
H
R
H3NO
O
R H
NH3O
O
RH
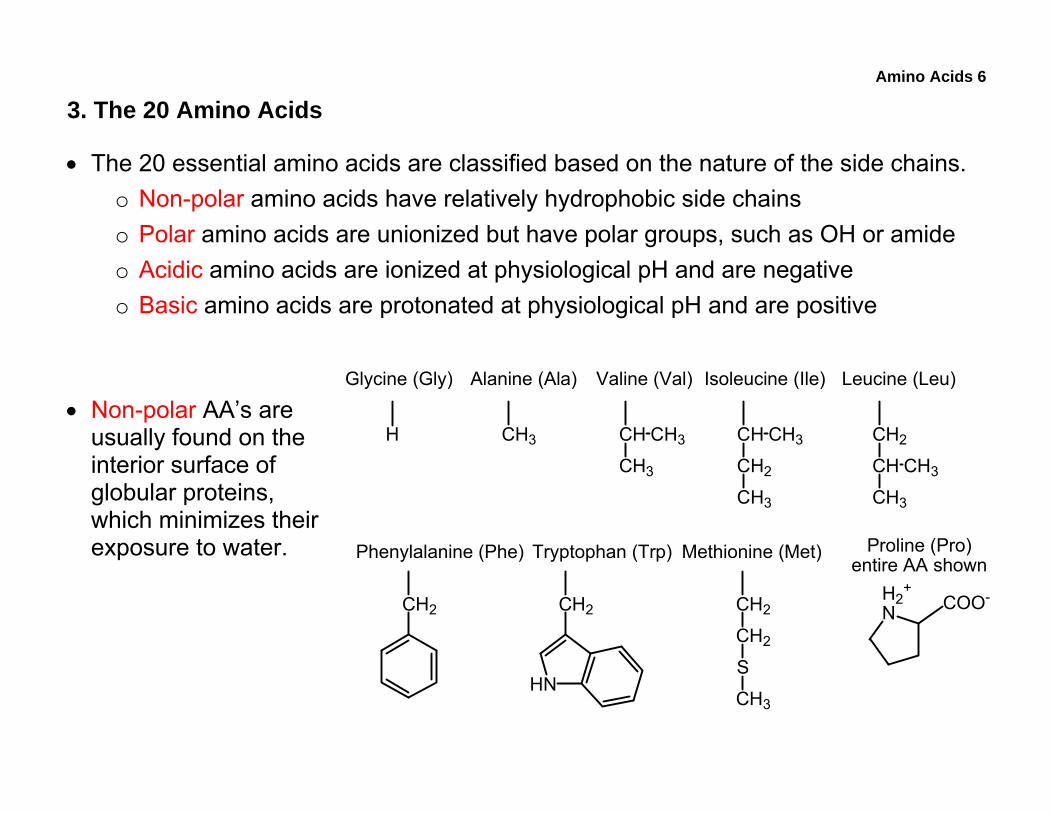
Amino Acids 6
3. The 20 Amino Acids
• The 20 essential amino acids are classified based on the nature of the side chains. o Non-polar amino acids have relatively hydrophobic side chains o Polar amino acids are unionized but have polar groups, such as OH or amide o Acidic amino acids are ionized at physiological pH and are negative o Basic amino acids are protonated at physiological pH and are positive
• Non-polar AA’s are usually found on the interior surface of globular proteins, which minimizes their exposure to water.
CH3H CH CH3
CH3
CH CH3
CH2
CH3
CH2
CH CH3
CH3
Glycine (Gly) Alanine (Ala) Valine (Val) Isoleucine (Ile) Leucine (Leu)
CH2 CH2
HN
CH2
CH2
S
CH3
Phenylalanine (Phe) Tryptophan (Trp) Methionine (Met)
H2+
N
Proline (Pro)entire AA shown
COO-
Amino Acids 7
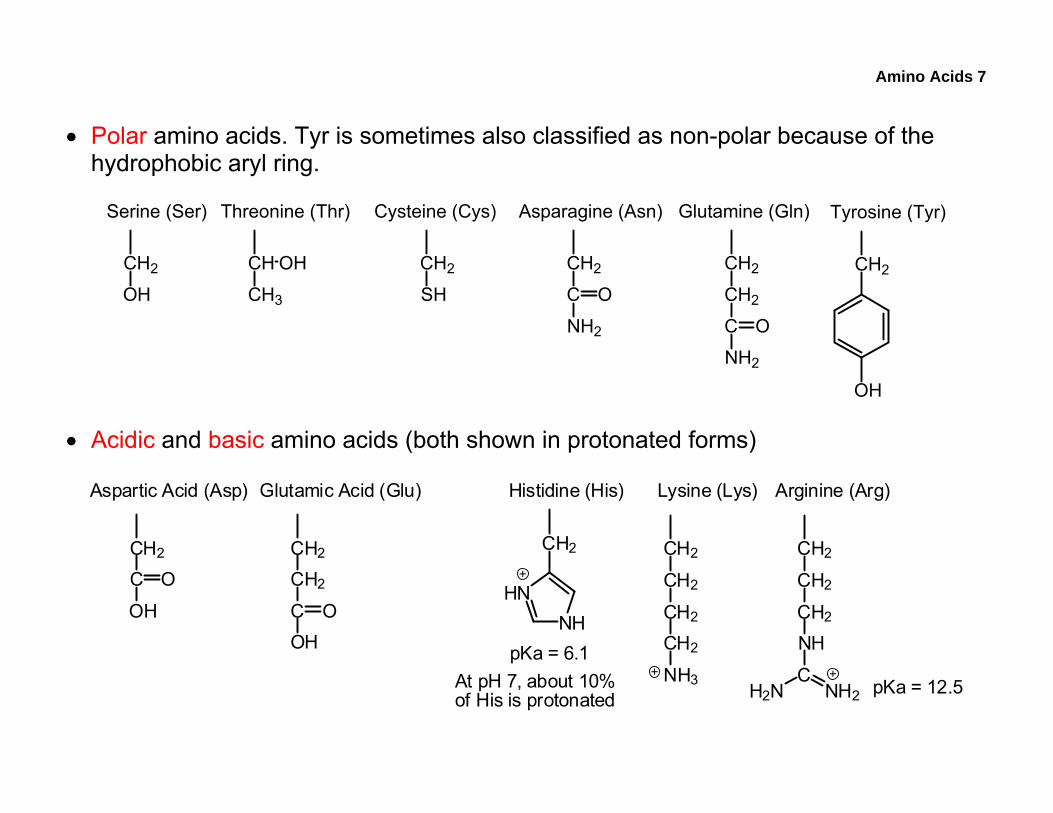
• Polar amino acids. Tyr is sometimes also classified as non-polar because of the hydrophobic aryl ring.
CH2
OH
CH2
OH
CH OH
CH3
Tyrosine (Tyr)Serine (Ser) Threonine (Thr)
CH2
C
NH2
O
CH2
CH2
C
NH2
O
CH2
SH
Cysteine (Cys) Asparagine (Asn) Glutamine (Gln)
• Acidic and basic amino acids (both shown in protonated forms)
CH2
HNNH
Histidine (His)
CH2
C
OH
OCH2
CH2
COH
O
CH2
CH2
CH2
CH2
NH3
CH2
CH2
CH2
NH
C
Aspartic Acid (Asp) Glutamic Acid (Glu) Lysine (Lys) Arginine (Arg)
H2N NH2
pKa = 6.1
pKa = 12.5At pH 7, about 10%of His is protonated
Amino Acids 8
B. Primary Structure
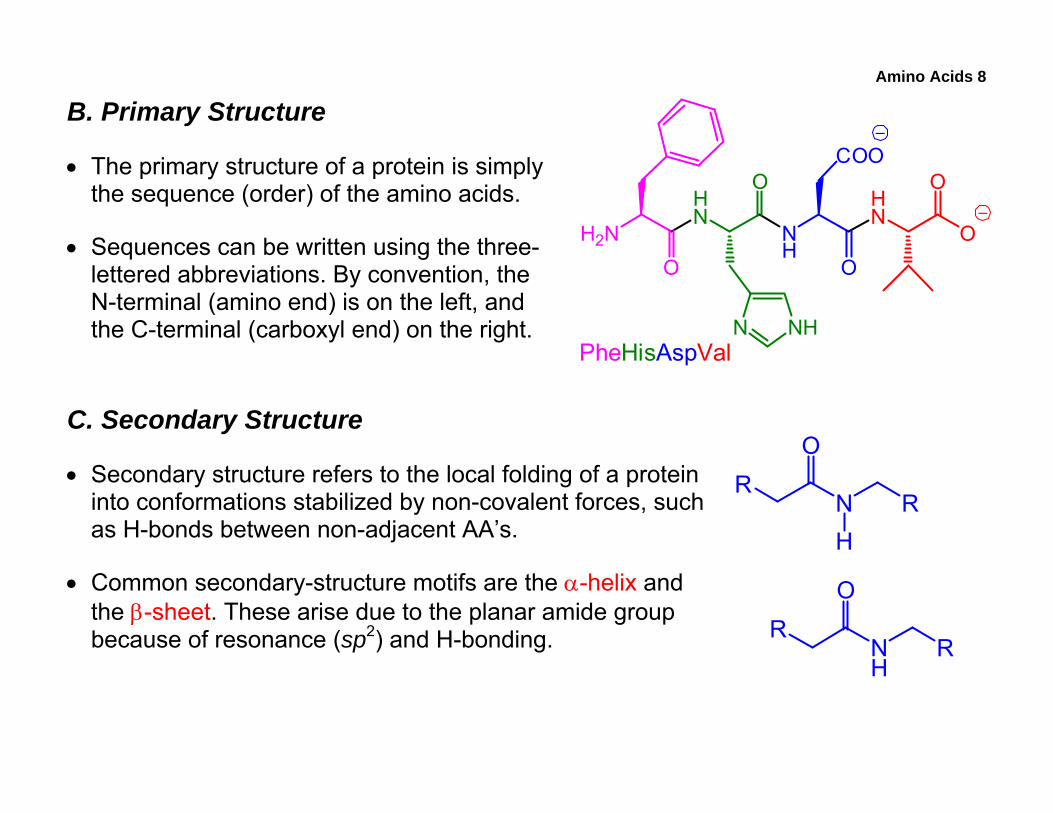
• The primary structure of a protein is simply the sequence (order) of the amino acids.
• Sequences can be written using the three-lettered abbreviations. By convention, the N-terminal (amino end) is on the left, and the C-terminal (carboxyl end) on the right.
C. Secondary Structure
• Secondary structure refers to the local folding of a protein into conformations stabilized by non-covalent forces, such as H-bonds between non-adjacent AA’s.
• Common secondary-structure motifs are the α-helix and the β-sheet. These arise due to the planar amide group because of resonance (sp2) and H-bonding.
R
NH
R
O
RN R
O
H
H2N
HN
NH
HN
O
O
OO
OCOO
N NHPheHisAspVal
Amino Acids 9
1. α-Helix
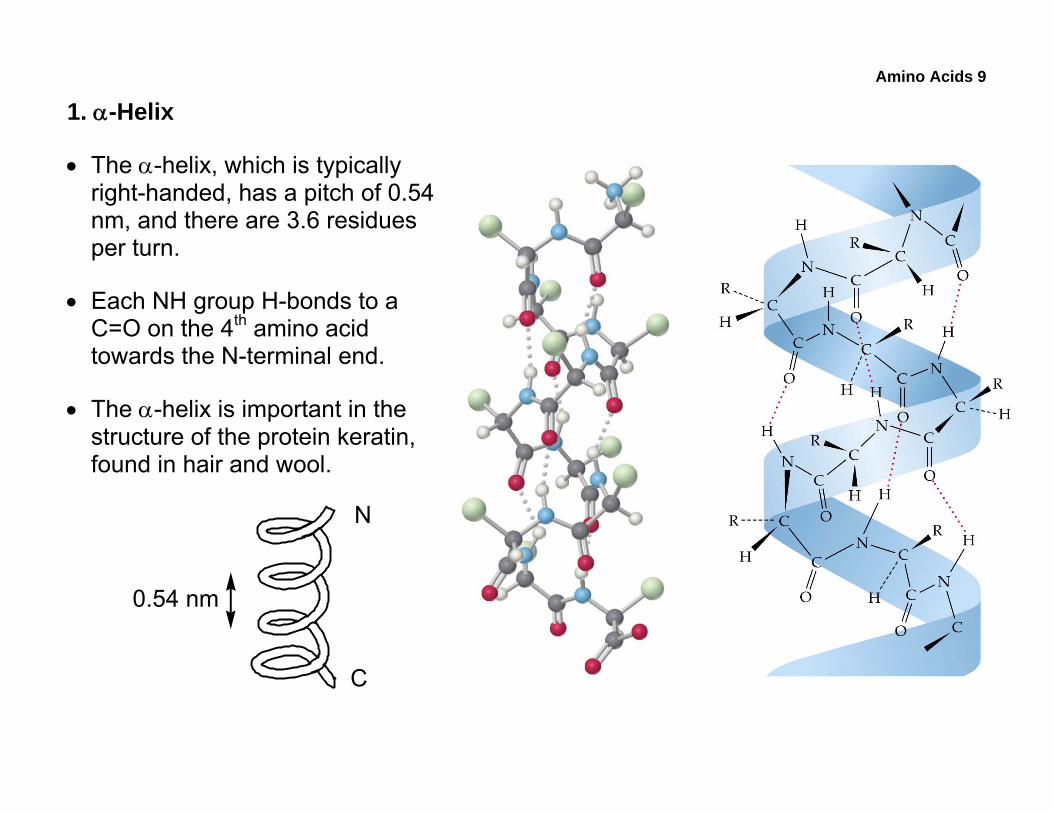
• The α-helix, which is typically right-handed, has a pitch of 0.54 nm, and there are 3.6 residues per turn.
• Each NH group H-bonds to a C=O on the 4th amino acid towards the N-terminal end.
• The α-helix is important in the structure of the protein keratin, found in hair and wool.
0.54 nm
N
C
Amino Acids 10
2. β-Sheet
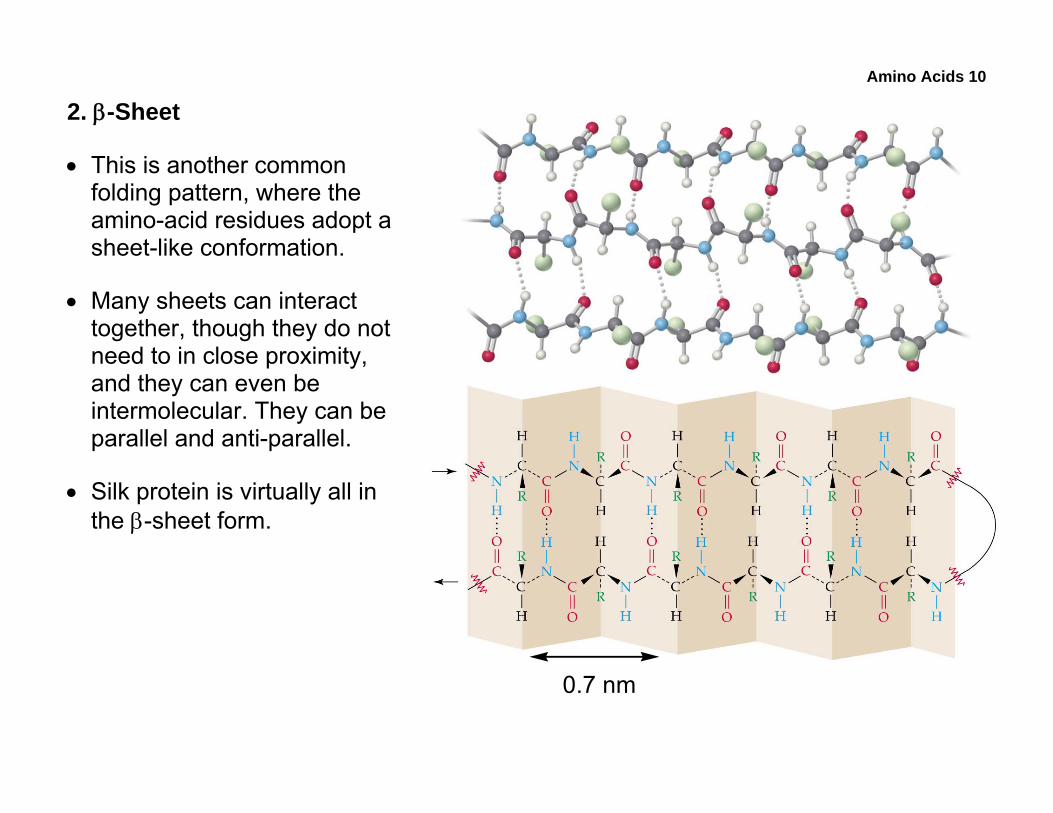
• This is another common folding pattern, where the amino-acid residues adopt a sheet-like conformation.
• Many sheets can interact together, though they do not need to in close proximity, and they can even be intermolecular. They can be parallel and anti-parallel.
• Silk protein is virtually all in the β-sheet form.
0.7 nm
Amino Acids 11
D. Tertiary Structure
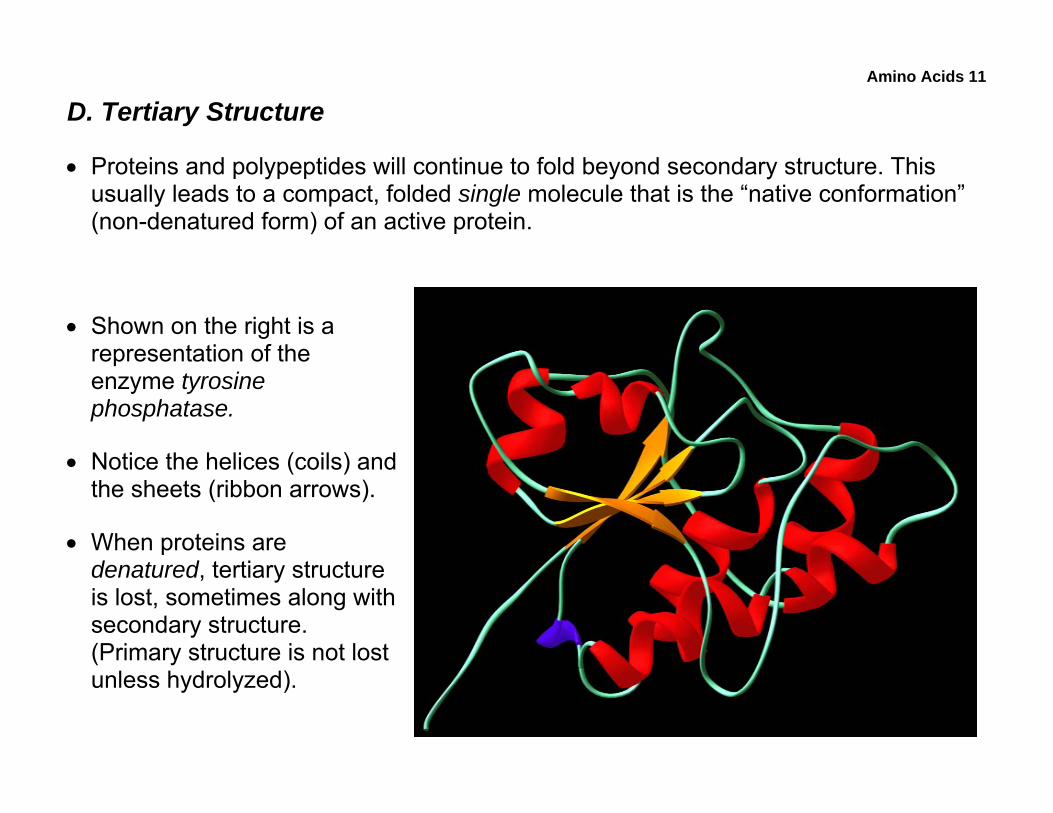
• Proteins and polypeptides will continue to fold beyond secondary structure. This usually leads to a compact, folded single molecule that is the “native conformation” (non-denatured form) of an active protein.
• Shown on the right is a representation of the enzyme tyrosine phosphatase.
• Notice the helices (coils) and the sheets (ribbon arrows).
• When proteins are denatured, tertiary structure is lost, sometimes along with secondary structure. (Primary structure is not lost unless hydrolyzed).
Amino Acids 12
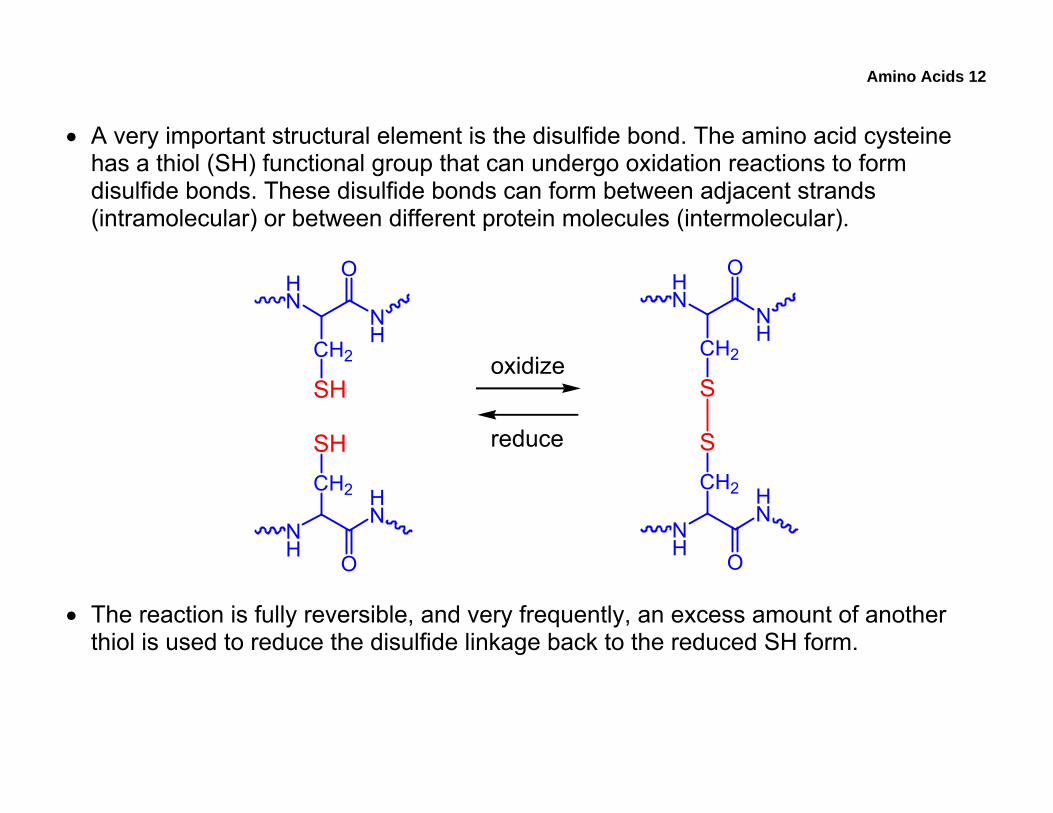
• A very important structural element is the disulfide bond. The amino acid cysteine has a thiol (SH) functional group that can undergo oxidation reactions to form disulfide bonds. These disulfide bonds can form between adjacent strands (intramolecular) or between different protein molecules (intermolecular).
HN
NH
O
CH2
SH
NH
HN
O
CH2
SH
HN
NH
O
CH2
S
NH
HN
O
CH2
S
oxidize
reduce
• The reaction is fully reversible, and very frequently, an excess amount of another thiol is used to reduce the disulfide linkage back to the reduced SH form.
Amino Acids 13
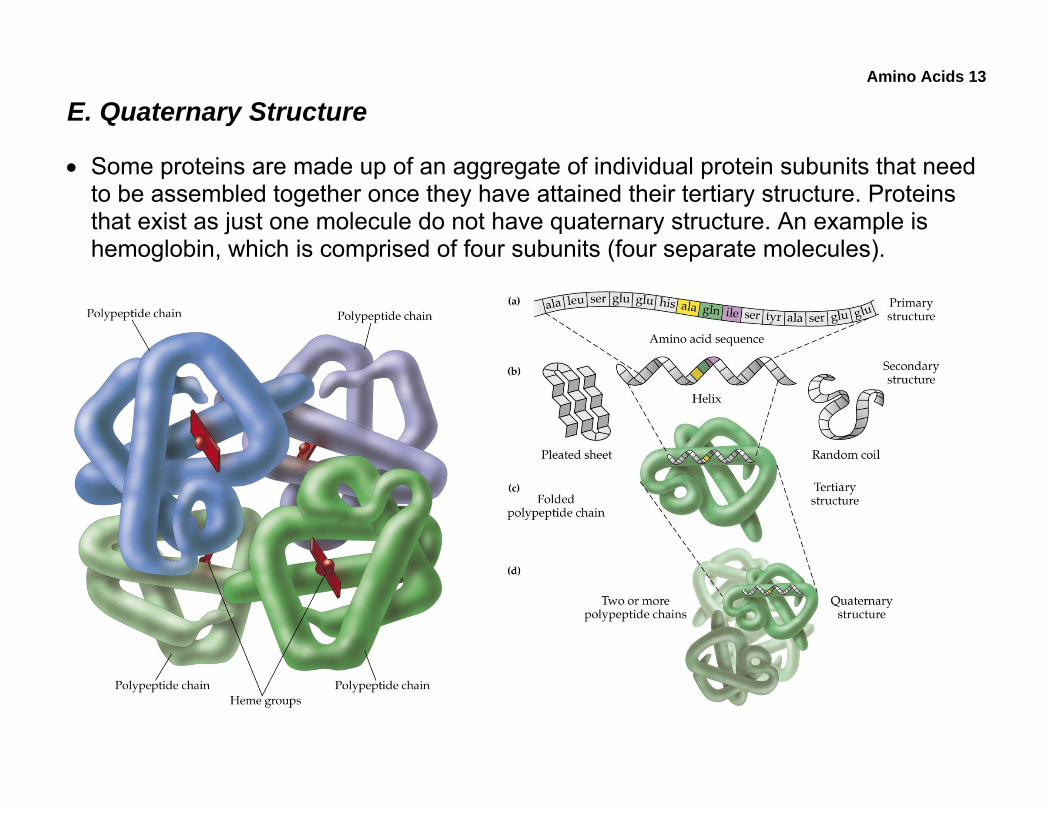
E. Quaternary Structure
• Some proteins are made up of an aggregate of individual protein subunits that need to be assembled together once they have attained their tertiary structure. Proteins that exist as just one molecule do not have quaternary structure. An example is hemoglobin, which is comprised of four subunits (four separate molecules).

Amino Acids 14
F. Determination of Amino Acid Composition
• Amino Acid Analysis (AAA) is a technique used to determine the composition and relative amounts of amino acids present in a peptide. AAA does not determine the primary structure (sequence) of the peptide.
• There are three steps to this technique:
1. Complete acid hydrolysis of a peptide into its constituent amino acids.
2. Separation of this amino-acid mixture by ion-exchange HPLC.
3. Quantification of the separated amino acids. This is done by converting them into a chromophore, then measuring their abundance by UV-vis spectroscopy. The intensity of the absorption is proportional to amount (Beer-Lambert Law).

Amino Acids 15
• The mechanism of the acid-catalyzed hydrolysis of acid derivatives, such as amides, esters, and anhydrides, is a nucleophilic acyl substitution, the characteristic reaction of acid derivatives. In acid hydrolysis, water is the nucleophile.

Amino Acids 16
• Hydrolysis can also be performed under basic conditions, where hydroxide is the nucleophile in the nucleophilic acyl substitution reaction.

Amino Acids 17
• The overall process for HPLC is rather automated, as shown in this scheme.
Amino Acids 18
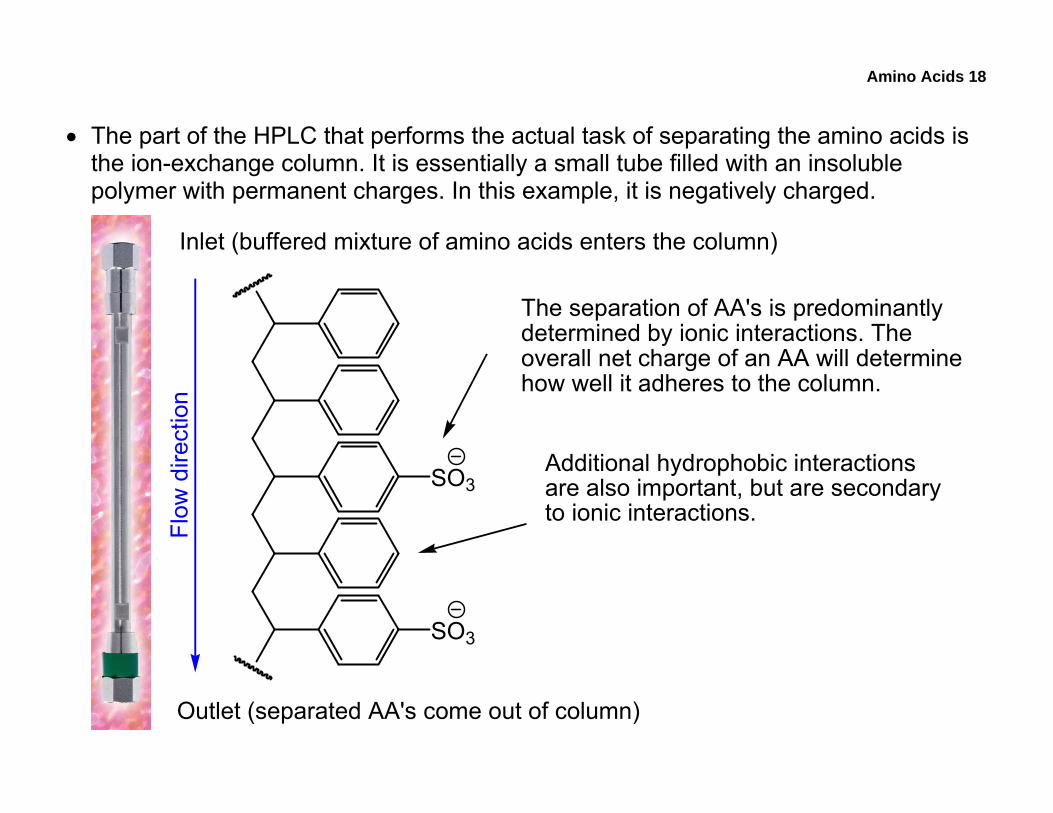
• The part of the HPLC that performs the actual task of separating the amino acids is the ion-exchange column. It is essentially a small tube filled with an insoluble polymer with permanent charges. In this example, it is negatively charged.
SO3
SO3
The separation of AA's is predominantlydetermined by ionic interactions. Theoverall net charge of an AA will determinehow well it adheres to the column.
Additional hydrophobic interactionsare also important, but are secondaryto ionic interactions.
Inlet (buffered mixture of amino acids enters the column)
Outlet (separated AA's come out of column)
Flow
dire
ctio
n
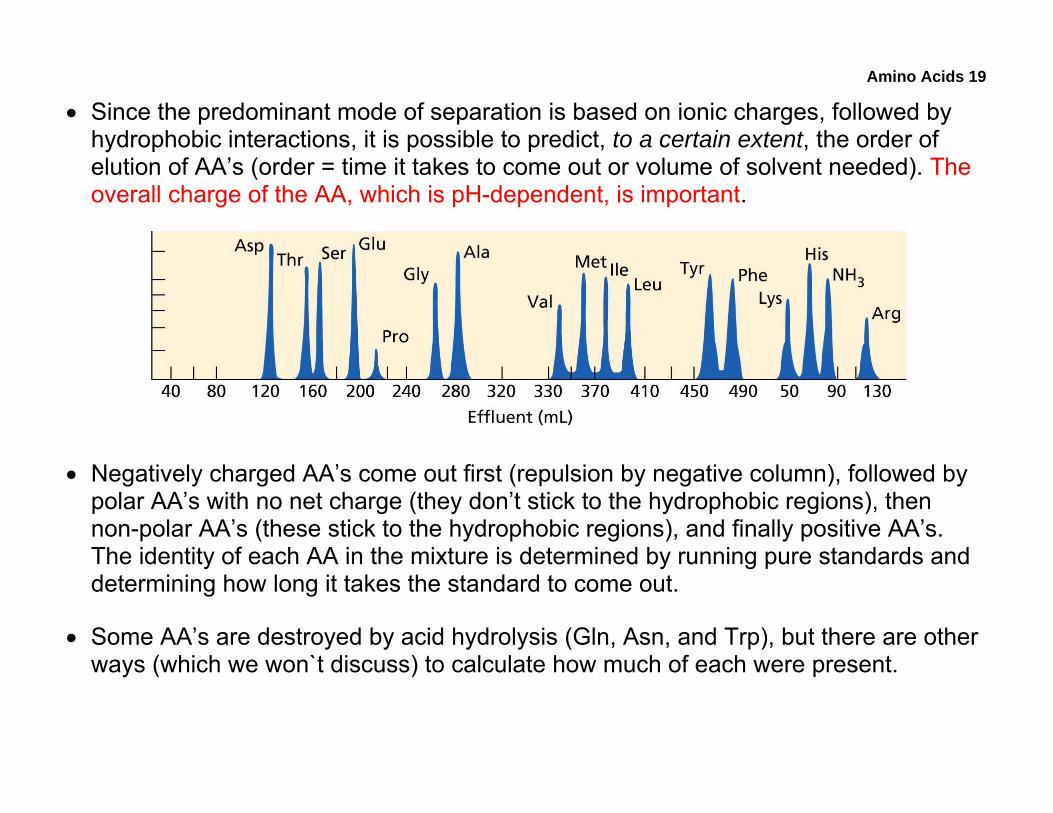
Amino Acids 19
• Since the predominant mode of separation is based on ionic charges, followed by hydrophobic interactions, it is possible to predict, to a certain extent, the order of elution of AA’s (order = time it takes to come out or volume of solvent needed). The overall charge of the AA, which is pH-dependent, is important.
• Negatively charged AA’s come out first (repulsion by negative column), followed by polar AA’s with no net charge (they don’t stick to the hydrophobic regions), then non-polar AA’s (these stick to the hydrophobic regions), and finally positive AA’s. The identity of each AA in the mixture is determined by running pure standards and determining how long it takes the standard to come out.
• Some AA’s are destroyed by acid hydrolysis (Gln, Asn, and Trp), but there are other ways (which we won`t discuss) to calculate how much of each were present.
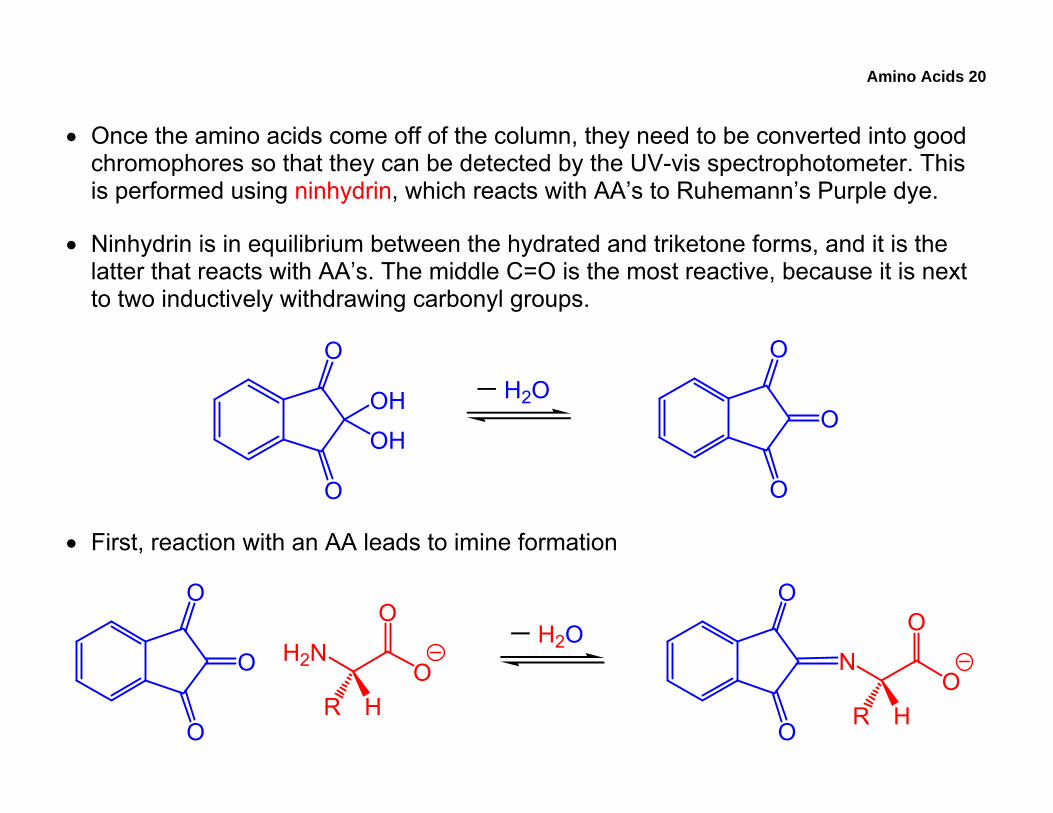
Amino Acids 20
• Once the amino acids come off of the column, they need to be converted into good chromophores so that they can be detected by the UV-vis spectrophotometer. This is performed using ninhydrin, which reacts with AA’s to Ruhemann’s Purple dye.
• Ninhydrin is in equilibrium between the hydrated and triketone forms, and it is the latter that reacts with AA’s. The middle C=O is the most reactive, because it is next to two inductively withdrawing carbonyl groups.
O
O
O
O
OH
OH
H2OO
• First, reaction with an AA leads to imine formation
H2NO
O
R H
O
O
OH2O
NO
O
R H
O
O

Amino Acids 21
• The mechanism of imine formation between an aldehyde/ketone and a primary amine is a nucleophilic addition reaction, characteristic of carbonyl compounds. The reaction is reversible, and in the reverse direction, the steps are the opposite.
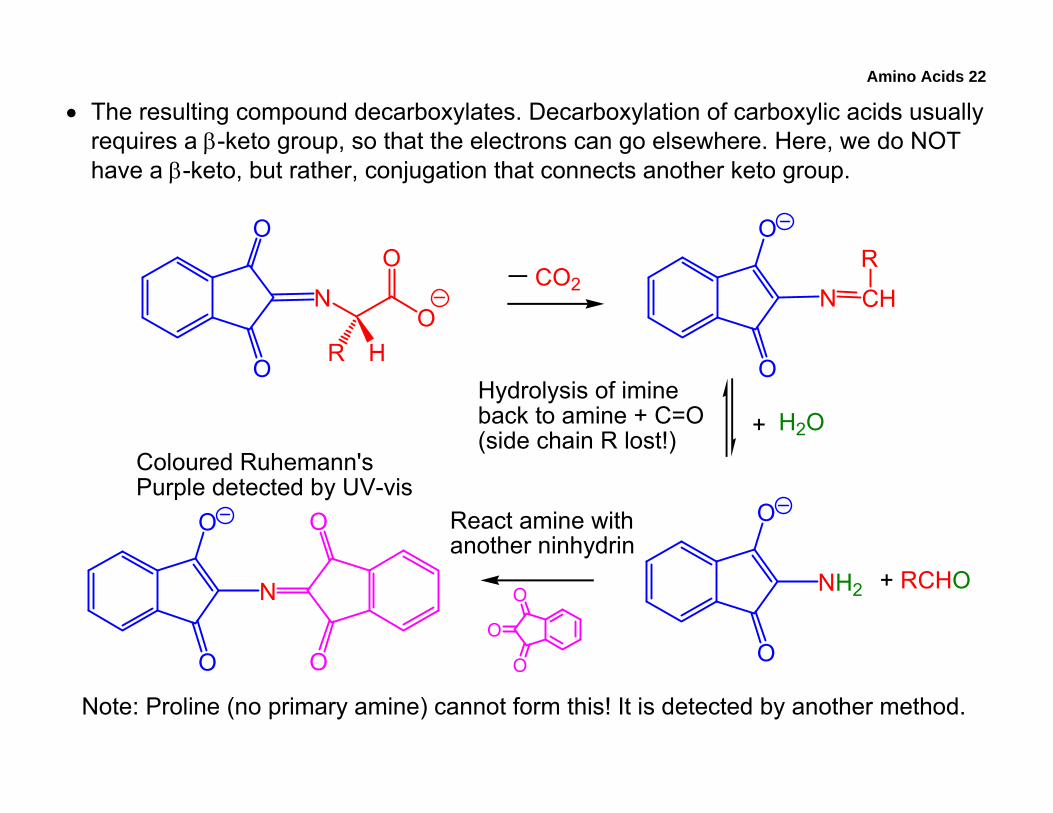
Amino Acids 22
• The resulting compound decarboxylates. Decarboxylation of carboxylic acids usually requires a β-keto group, so that the electrons can go elsewhere. Here, we do NOT have a β-keto, but rather, conjugation that connects another keto group.
NO
O
R H
O
O
CO2N CH
RO
O
H2O+Hydrolysis of imineback to amine + C=O(side chain R lost!)
NH2
O
O
+ RCHOO
O
O
React amine withanother ninhydrin
N
O
O
O
O
Coloured Ruhemann'sPurple detected by UV-vis
Note: Proline (no primary amine) cannot form this! It is detected by another method.

Amino Acids 23
• Summary of amino acid analysis:
o The technique determines the composition of the protein or polypeptide, not the sequence of the amino acids.
o In HPLC:
The identity of the AA is determined by its retention time on the column.
Amino acids react with ninhydrin after they have been separated (after they come out of the column). This is a post-column derivatization technique.
Amino acids are destroyed when they react with ninhydrin. The side chain is lost as an aldehyde.
Regardless of the amino acid that reacts with ninhydrin, the same chromophore is produced. Because each AA produces the same chromophore, we can compare the relative amounts of each AA by measuring the amount of chromophore produced (Beer-Lambert law).

Amino Acids 24
G. Sequence Analysis (Primary Structure)
• The goal of sequence analysis is to determine what order the amino acids are connected in. Several methods are often used in conjunction with each other.
1. Partial Hydrolysis
• The large polypeptide is first partially hydrolyzed into many smaller fragments. This can be achieved chemically (acid) or enzymatically.
o One reaction that is examined in an upper-level course offered by Biochem is the treatment of the polypeptide using CNBr (cyanogen bromide), which cleaves after a Met residue when the polypeptide is written N C.
• Realize that we are always working with many, many molecules of the same peptide, not just one molecule. Thus, after partial hydrolysis, we will have many small peptides (oligopeptides) that can overlap in sequence.
• Each of these oligopeptides is then subjected to N-terminal analysis, N-terminal sequencing, and C-terminal sequencing.

Amino Acids 25
2. N-Terminal Identification
• The purpose of this technique is to tag the amino acid present at the N-terminal, which has a primary amino group. Once this amino acid is tagged, the peptide can be hydrolyzed, followed by identification of the tagged AA via HPLC by comparing its elution time with those of known, tagged standards.
• Tagging is done with dinitrofluorobenzene (DNFB, Sanger’s Reagent), which forms a coloured product (no ninhydrin needed) via a nucleophilic aromatic substitution reaction. Most aromatic compounds do not substitute, but this works because the multiple electron-withdrawing groups make the sp2 carbon sufficiently δ+.
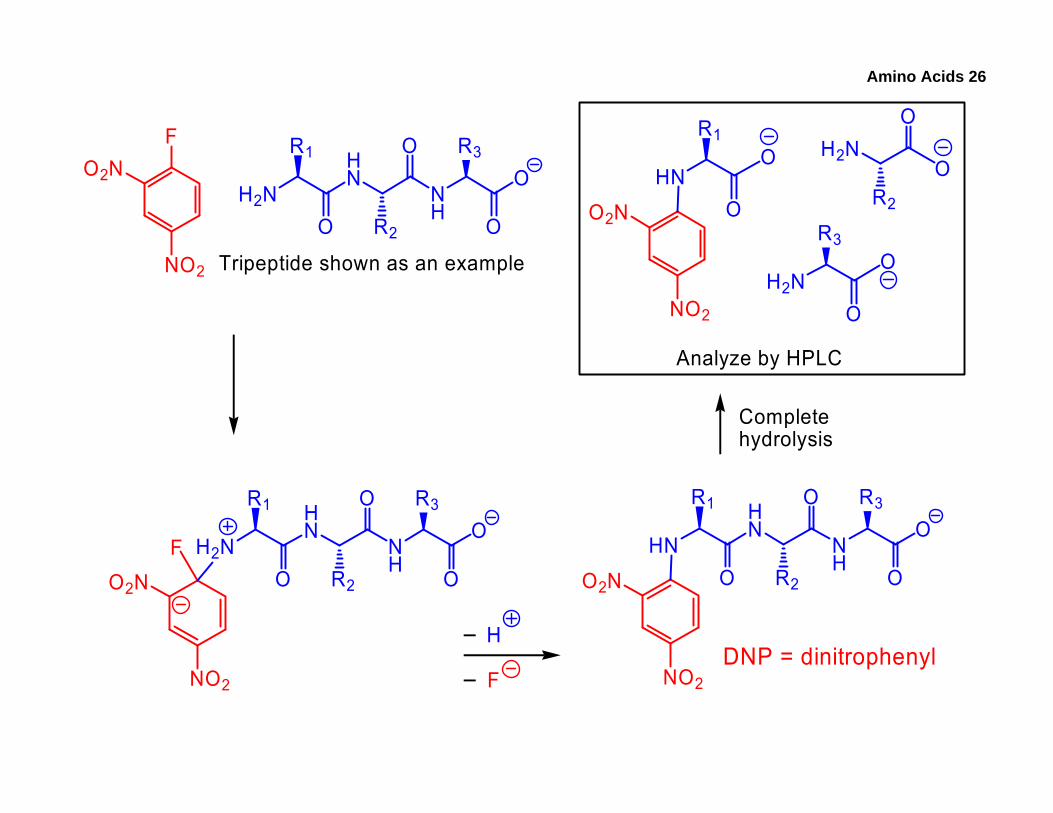
Amino Acids 26
H2NHN
NH
R1
OR3
O
O
OR2
Tripeptide shown as an example
FO2N
NO2
H2NHN
NH
R1
OR3
O
O
OR2
FO2N
NO2
HNHN
NH
R1
OR3
O
O
OR2O2N
NO2
H
F
Completehydrolysis
HN
R1
OO2N
NO2
O H2NO
R2
O
H2NO
R3O
Analyze by HPLC
DNP = dinitrophenyl
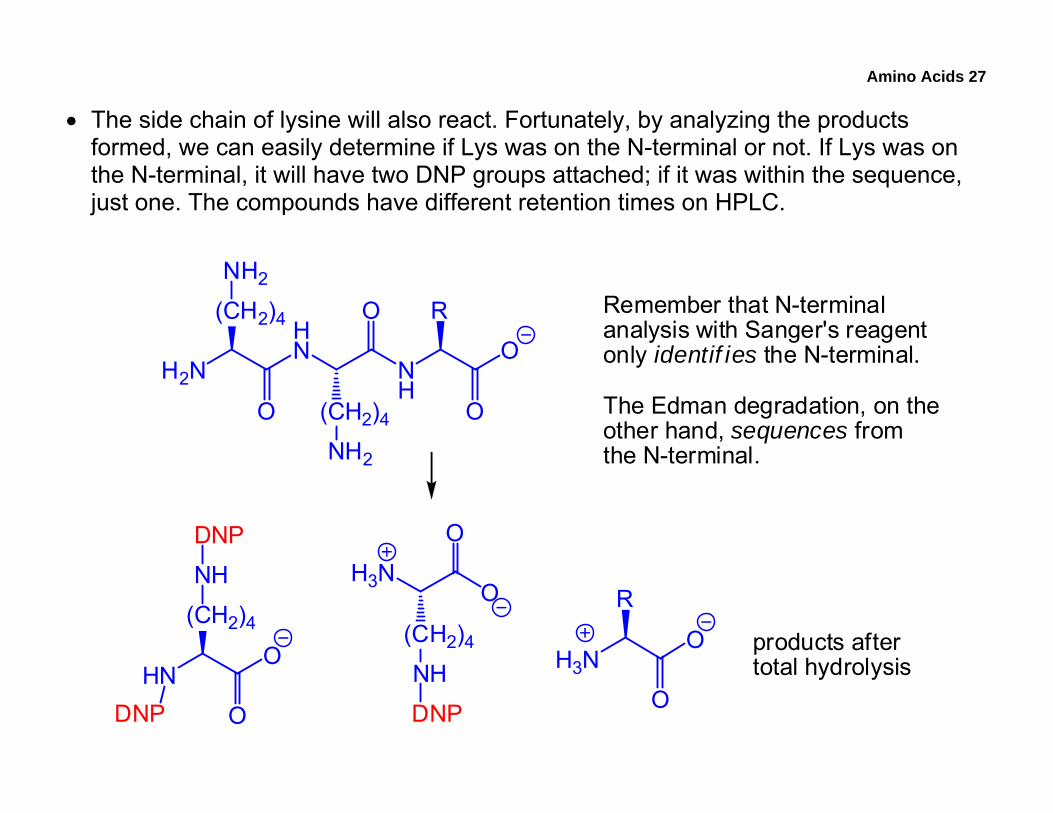
Amino Acids 27
• The side chain of lysine will also react. Fortunately, by analyzing the products formed, we can easily determine if Lys was on the N-terminal or not. If Lys was on the N-terminal, it will have two DNP groups attached; if it was within the sequence, just one. The compounds have different retention times on HPLC.
H2N
HN
NH
(CH2)4
O
R
O
O
O(CH2)4
NH2
NH2
HNO
(CH2)4
O
NH
DNP
DNP
H3NO
(CH2)4
O
NH
DNP
H3NO
R
O
Remember that N-terminalanalysis with Sanger's reagentonly identif ies the N-terminal.
The Edman degradation, on theother hand, sequences fromthe N-terminal.
products aftertotal hydrolysis
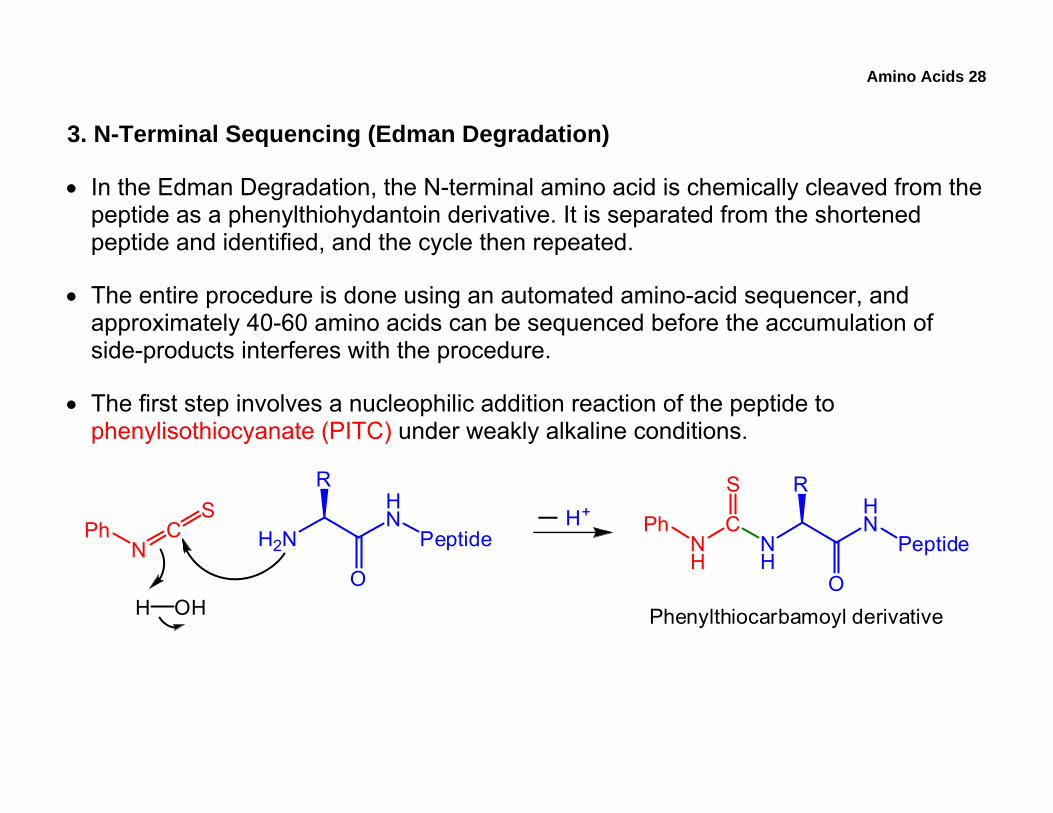
Amino Acids 28
3. N-Terminal Sequencing (Edman Degradation)
• In the Edman Degradation, the N-terminal amino acid is chemically cleaved from the peptide as a phenylthiohydantoin derivative. It is separated from the shortened peptide and identified, and the cycle then repeated.
• The entire procedure is done using an automated amino-acid sequencer, and approximately 40-60 amino acids can be sequenced before the accumulation of side-products interferes with the procedure.
• The first step involves a nucleophilic addition reaction of the peptide to phenylisothiocyanate (PITC) under weakly alkaline conditions.
PhN
CS
H2N
HN
Peptide
R
OH OH
H+
NH
HN
Peptide
R
O
CNH
S
Ph
Phenylthiocarbamoyl derivative
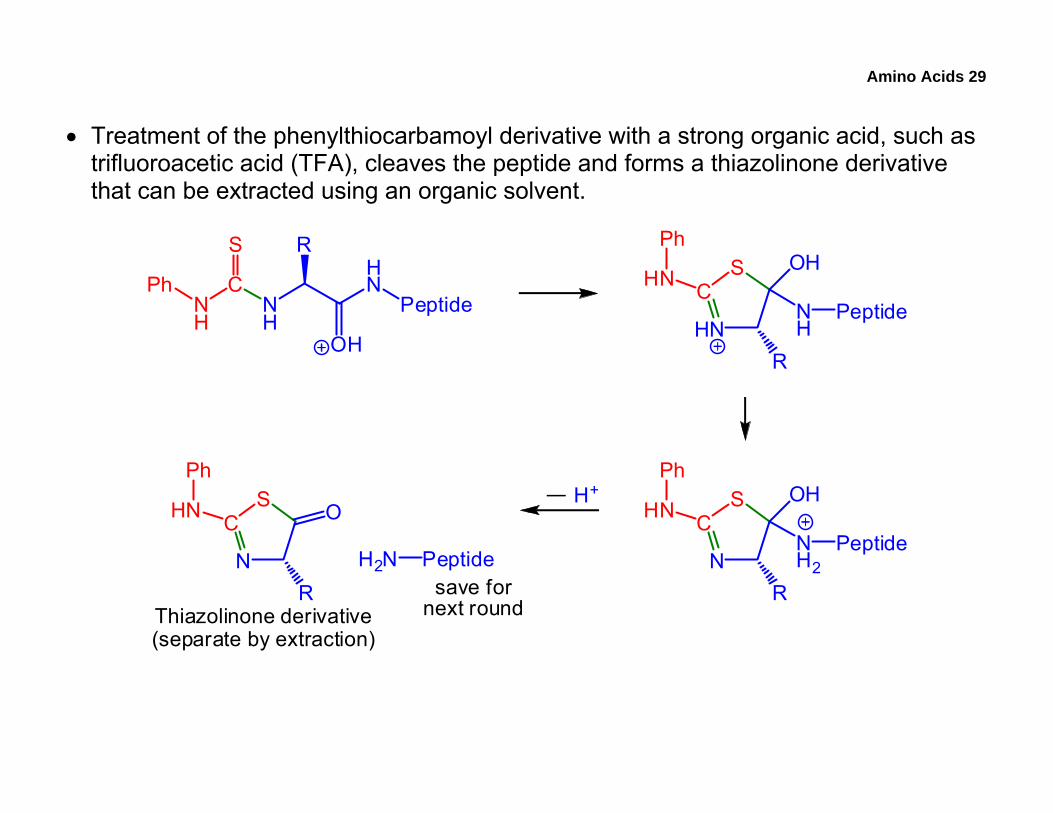
Amino Acids 29
• Treatment of the phenylthiocarbamoyl derivative with a strong organic acid, such as trifluoroacetic acid (TFA), cleaves the peptide and forms a thiazolinone derivative that can be extracted using an organic solvent.
NH
HN
Peptide
R
OH
CNH
S
Ph
HN
SC
OH
NH
Peptide
R
HN
Ph
N
SC
OH
NH2
Peptide
R
HN
Ph
N
SC O
R
HN
PhH+
H2N Peptide
Thiazolinone derivative(separate by extraction)
save fornext round
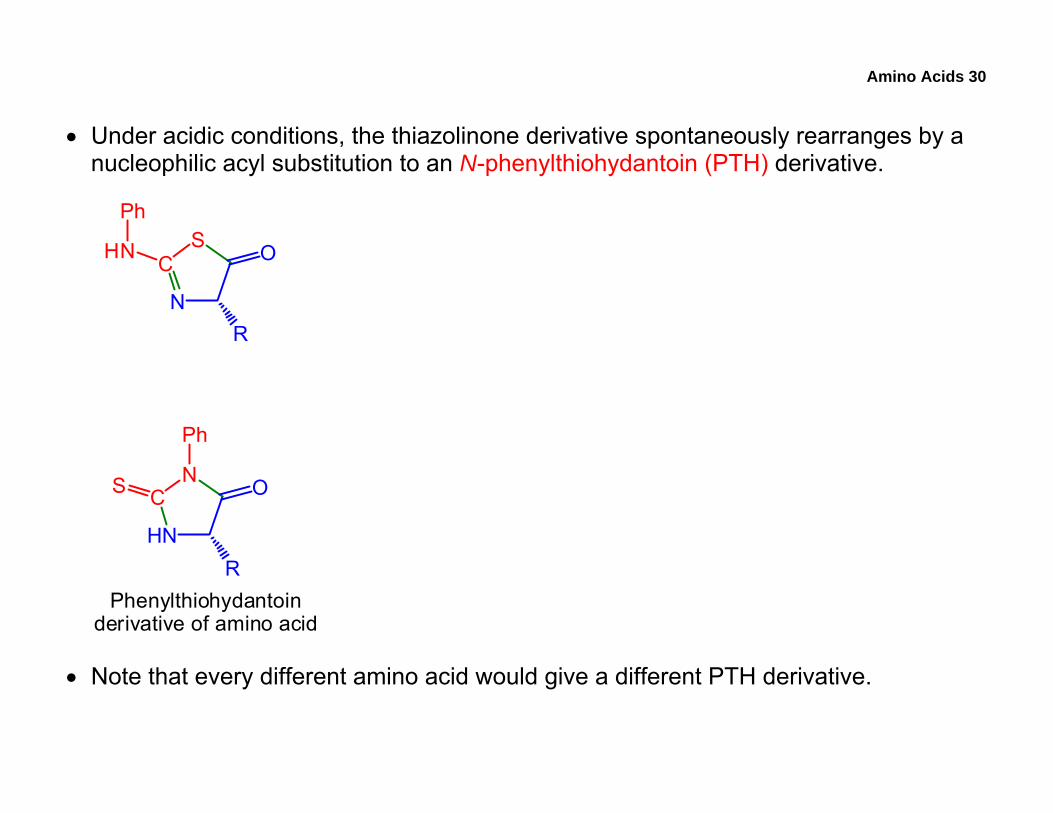
Amino Acids 30
• Under acidic conditions, the thiazolinone derivative spontaneously rearranges by a nucleophilic acyl substitution to an N-phenylthiohydantoin (PTH) derivative.
N
SC O
R
HN
Ph
Phenylthiohydantoinderivative of amino acid
HN
NC O
R
S
Ph
• Note that every different amino acid would give a different PTH derivative.

Amino Acids 31
4. C-Terminal Sequencing
• C-terminal sequencing identifies the amino acid at, and also provides sequence information from, the C-terminal. Carboxypeptidase sequentially removes C-terminal AA’s. With some luck, we can sequence several residues from the C-terminal (the digestion isn’t completely efficient, which is why we need to use small peptides… enzyme eventually quits).
• This is done by sampling the mixture during a carboxypeptidase digestion and analyzing the sample by HPLC to identify and quantify the released amino acids.
Am
ount
of A
A
Hours of digestion
IleSer
ThrMet
Peptide sequence:
N-term COOH

Amino Acids 32
• Up to now, some of the concepts we’ve covered include:
o Fundamental properties of amino acids, peptides, and proteins
o AA analysis and HPLC
o N-terminal identification, N-terminal sequencing, and C-terminal sequencing
o General mechanisms: nucleophilic addition, nucleophilic acyl substitution, nucleophilic aromatic substitution, and decarboxylation
• Using amino-acid structures provided in the lab manual, attempt:
o Practice problems: 1 – 22
o 2008 Midterm: 8 – 16
• For exam-studying purposes, review all the notes and redo the practice problems in the lab manual. Understand and be able to apply the concepts.
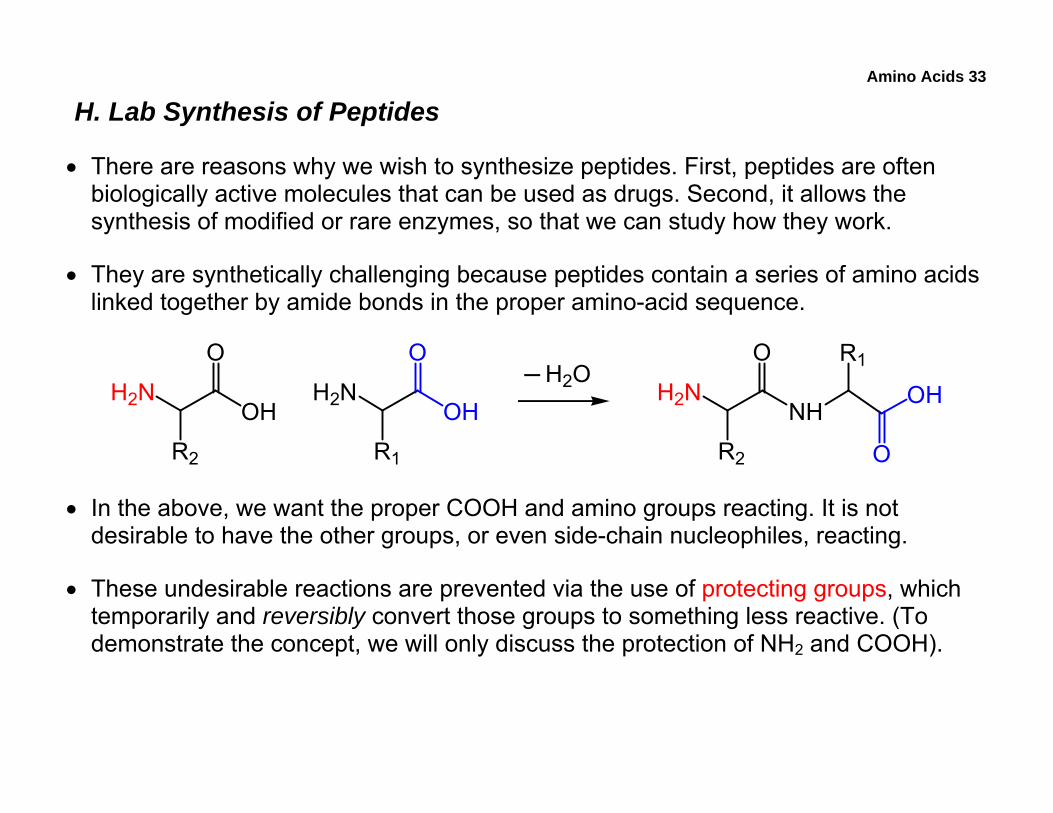
Amino Acids 33
H. Lab Synthesis of Peptides
• There are reasons why we wish to synthesize peptides. First, peptides are often biologically active molecules that can be used as drugs. Second, it allows the synthesis of modified or rare enzymes, so that we can study how they work.
• They are synthetically challenging because peptides contain a series of amino acids linked together by amide bonds in the proper amino-acid sequence.
H2NOH
O
R2
H2NOH
O
R1
H2NNH
O
R2
R1
O
OHH2O
• In the above, we want the proper COOH and amino groups reacting. It is not desirable to have the other groups, or even side-chain nucleophiles, reacting.
• These undesirable reactions are prevented via the use of protecting groups, which temporarily and reversibly convert those groups to something less reactive. (To demonstrate the concept, we will only discuss the protection of NH2 and COOH).

Amino Acids 34
1. Solution-Phase Synthesis
• We will examine two synthetic approaches, the solution-phase and the solid-phase.
• The choice of protecting group depends on many factors, including the type of functional group that we wish to protect, the conditions that the group must be stable under, and the conditions required to remove it. Of course, the conditions used to remove the protecting group must not destroy the product.
a. Protection of COOH
• The carboxylic acid group can be protected as an ester, and the benzyl (Bz) ester was experimentally found to be a good choice. It can be introduced by Fischer esterification and then later removed, when desired, using a strong acid. Yet, the acid is not strong enough to hydrolyze the amide (concept of graduated acid lability).
H2NO
O
R1
H2NOH
O
R1
Br
+HBr / HOAc
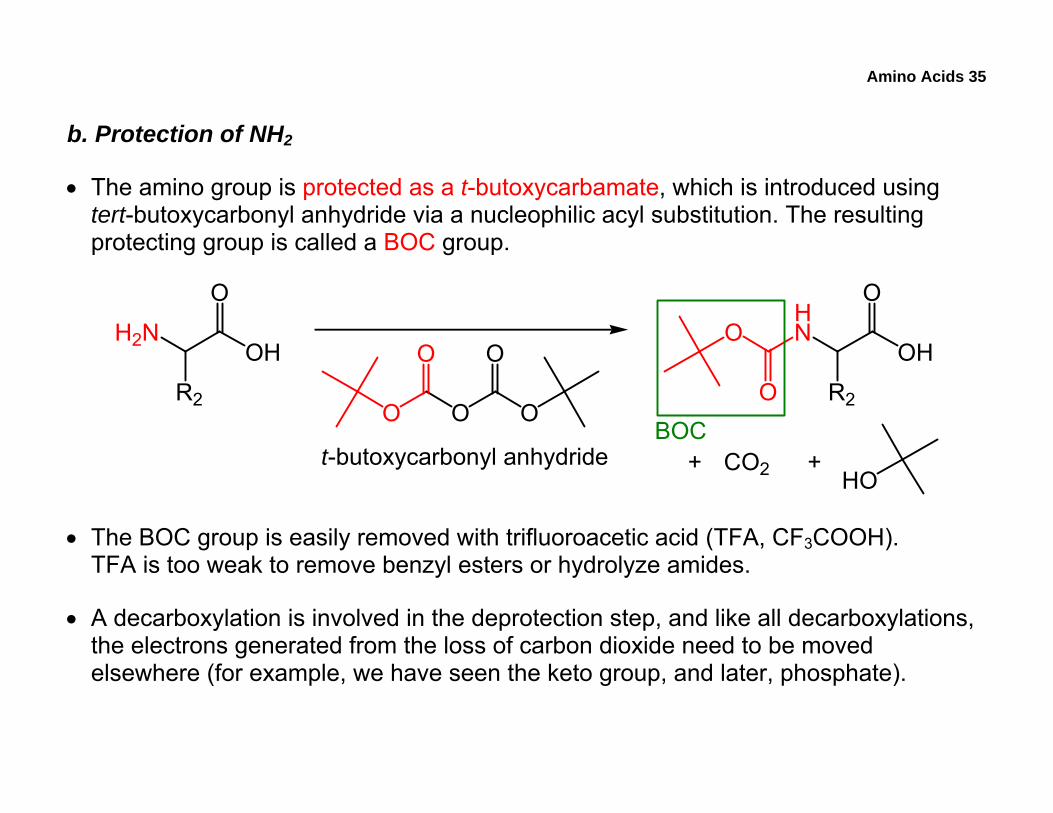
Amino Acids 35
b. Protection of NH2
• The amino group is protected as a t-butoxycarbamate, which is introduced using tert-butoxycarbonyl anhydride via a nucleophilic acyl substitution. The resulting protecting group is called a BOC group.
H2NOH
O
R2 O O O
O O
HN
OH
O
R2O
O
CO2 HO+ +t-butoxycarbonyl anhydride
BOC
• The BOC group is easily removed with trifluoroacetic acid (TFA, CF3COOH). TFA is too weak to remove benzyl esters or hydrolyze amides.
• A decarboxylation is involved in the deprotection step, and like all decarboxylations, the electrons generated from the loss of carbon dioxide need to be moved elsewhere (for example, we have seen the keto group, and later, phosphate).
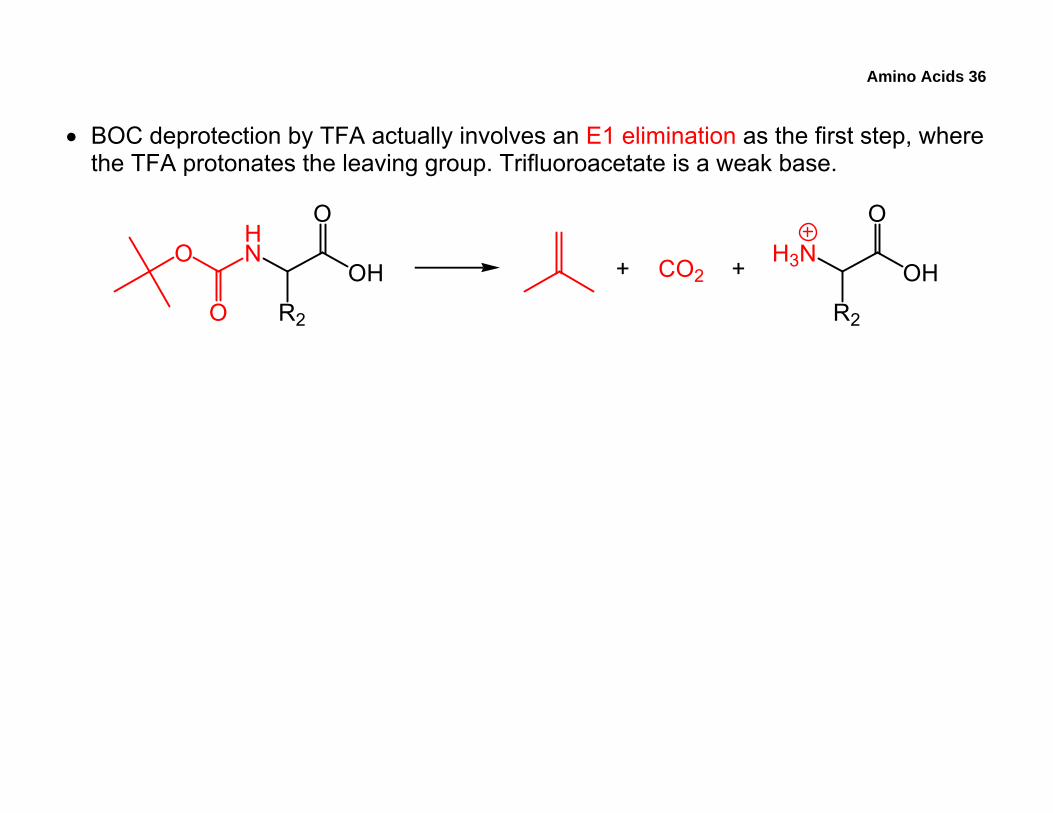
Amino Acids 36
• BOC deprotection by TFA actually involves an E1 elimination as the first step, where the TFA protonates the leaving group. Trifluoroacetate is a weak base.
HN
OH
O
R2O
O CO2+ + H3NOH
O
R2
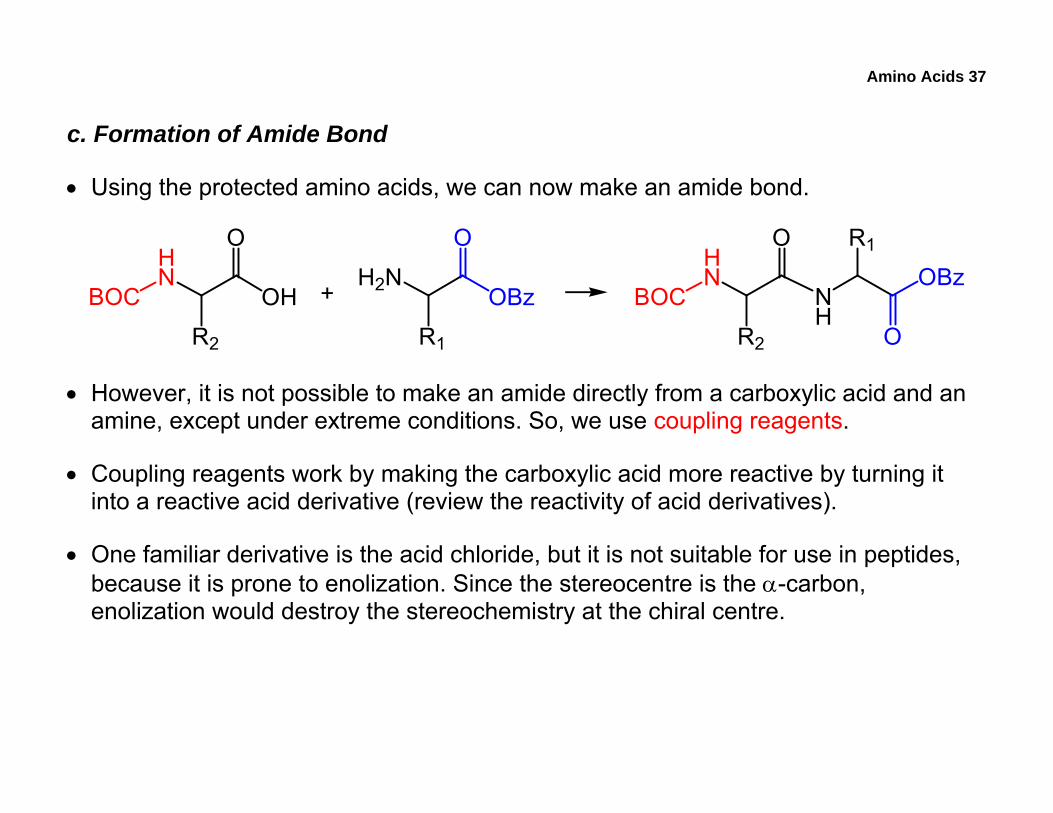
Amino Acids 37
c. Formation of Amide Bond
• Using the protected amino acids, we can now make an amide bond.
HN
OH
O
R2
BOCH2N
OBz
O
R1
+HN
O
R2
BOC NH
OBz
O
R1
• However, it is not possible to make an amide directly from a carboxylic acid and an amine, except under extreme conditions. So, we use coupling reagents.
• Coupling reagents work by making the carboxylic acid more reactive by turning it into a reactive acid derivative (review the reactivity of acid derivatives).
• One familiar derivative is the acid chloride, but it is not suitable for use in peptides, because it is prone to enolization. Since the stereocentre is the α-carbon, enolization would destroy the stereochemistry at the chiral centre.
Amino Acids 38
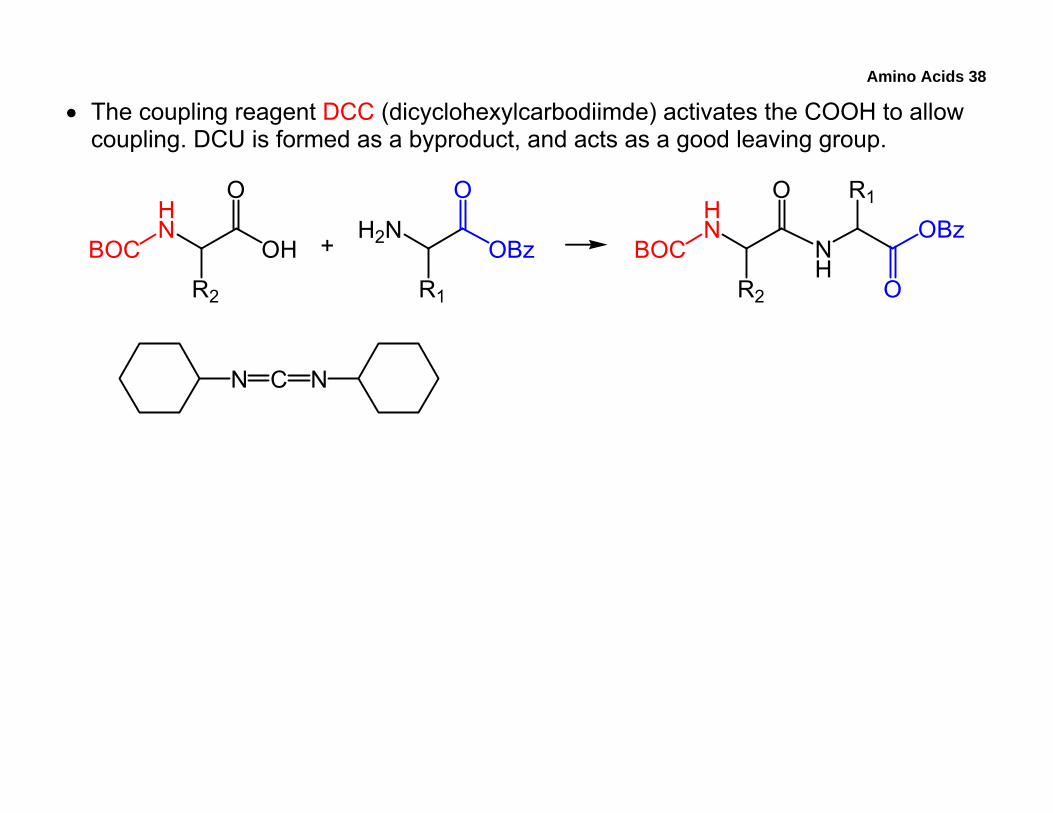
• The coupling reagent DCC (dicyclohexylcarbodiimde) activates the COOH to allow coupling. DCU is formed as a byproduct, and acts as a good leaving group.
HN
OH
O
R2
BOCH2N
OBz
O
R1
+HN
O
R2
BOC NH
OBz
O
R1
N C N
Amino Acids 39
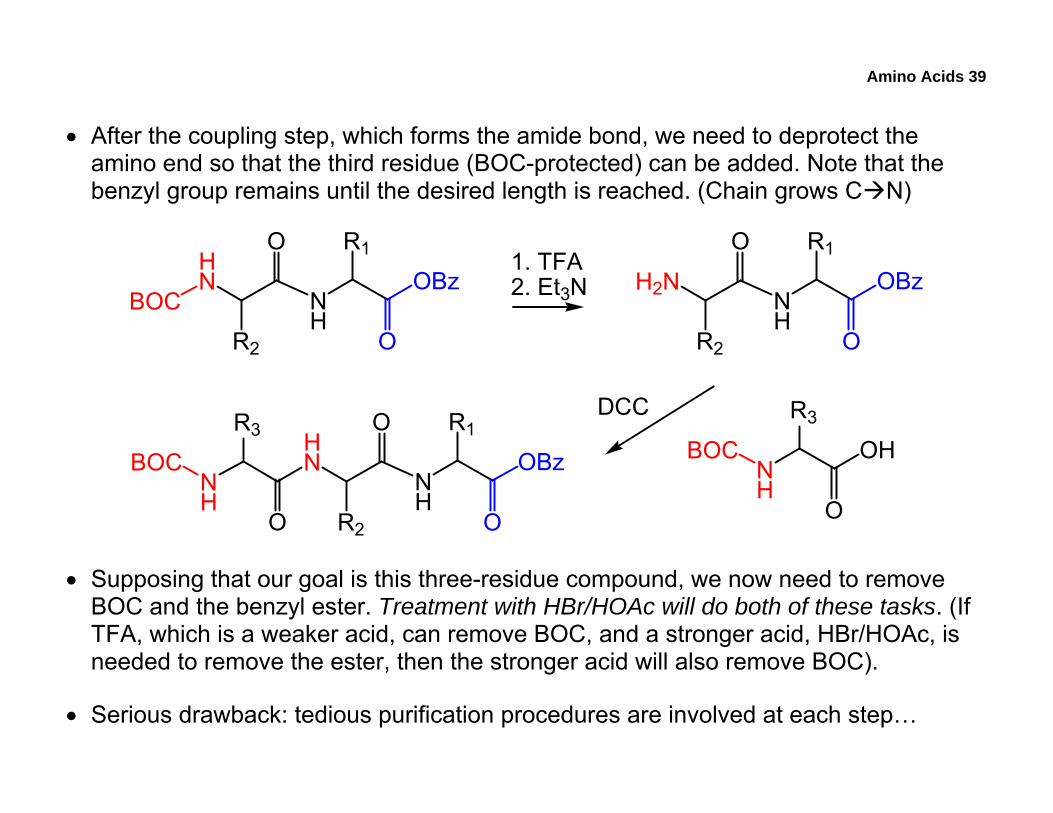
• After the coupling step, which forms the amide bond, we need to deprotect the amino end so that the third residue (BOC-protected) can be added. Note that the benzyl group remains until the desired length is reached. (Chain grows C N)
HN
O
R2
BOC NH
OBz
O
R1 1. TFA2. Et3N H2N
O
R2
NH
OBz
O
R1
NH
OH
O
R3
BOC
DCCHN
O
R2
NH
OBz
O
R1
O
R3
NH
BOC
• Supposing that our goal is this three-residue compound, we now need to remove BOC and the benzyl ester. Treatment with HBr/HOAc will do both of these tasks. (If TFA, which is a weaker acid, can remove BOC, and a stronger acid, HBr/HOAc, is needed to remove the ester, then the stronger acid will also remove BOC).
• Serious drawback: tedious purification procedures are involved at each step…
Amino Acids 40
2. Solid-Phase Synthesis
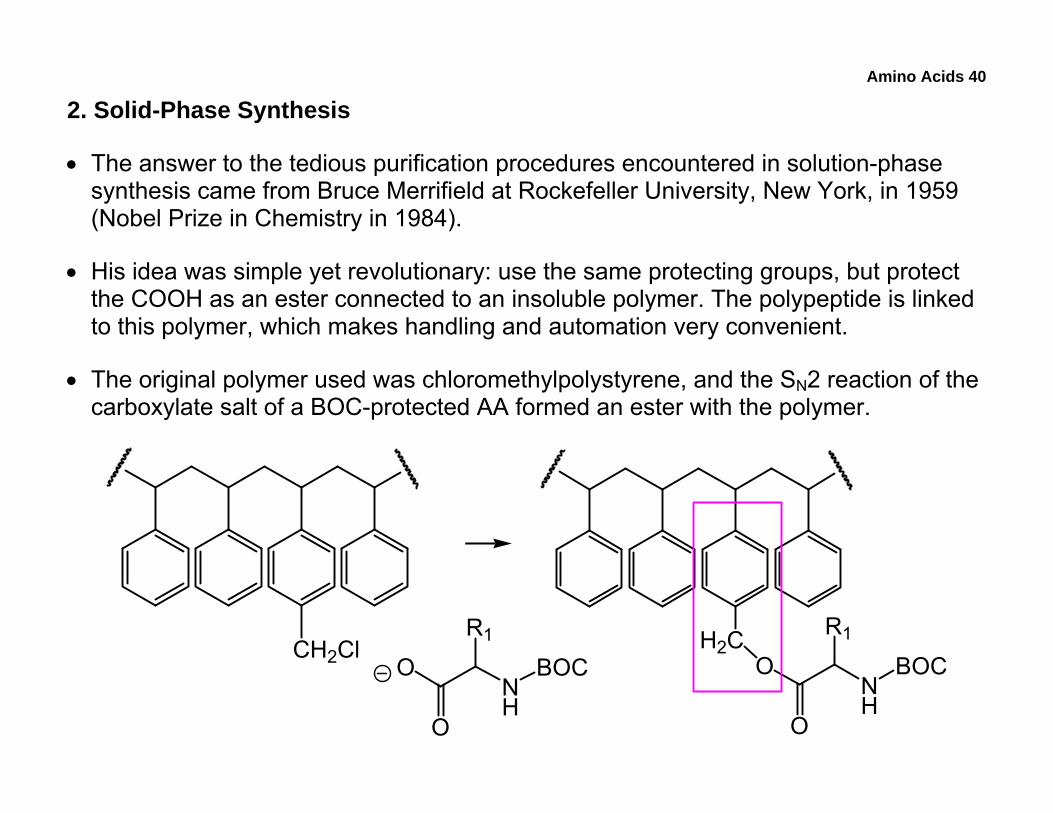
• The answer to the tedious purification procedures encountered in solution-phase synthesis came from Bruce Merrifield at Rockefeller University, New York, in 1959 (Nobel Prize in Chemistry in 1984).
• His idea was simple yet revolutionary: use the same protecting groups, but protect the COOH as an ester connected to an insoluble polymer. The polypeptide is linked to this polymer, which makes handling and automation very convenient.
• The original polymer used was chloromethylpolystyrene, and the SN2 reaction of the carboxylate salt of a BOC-protected AA formed an ester with the polymer.
CH2Cl H2C
NH
O
O
R1
BOCNH
O
O
R1
BOC
Amino Acids 41
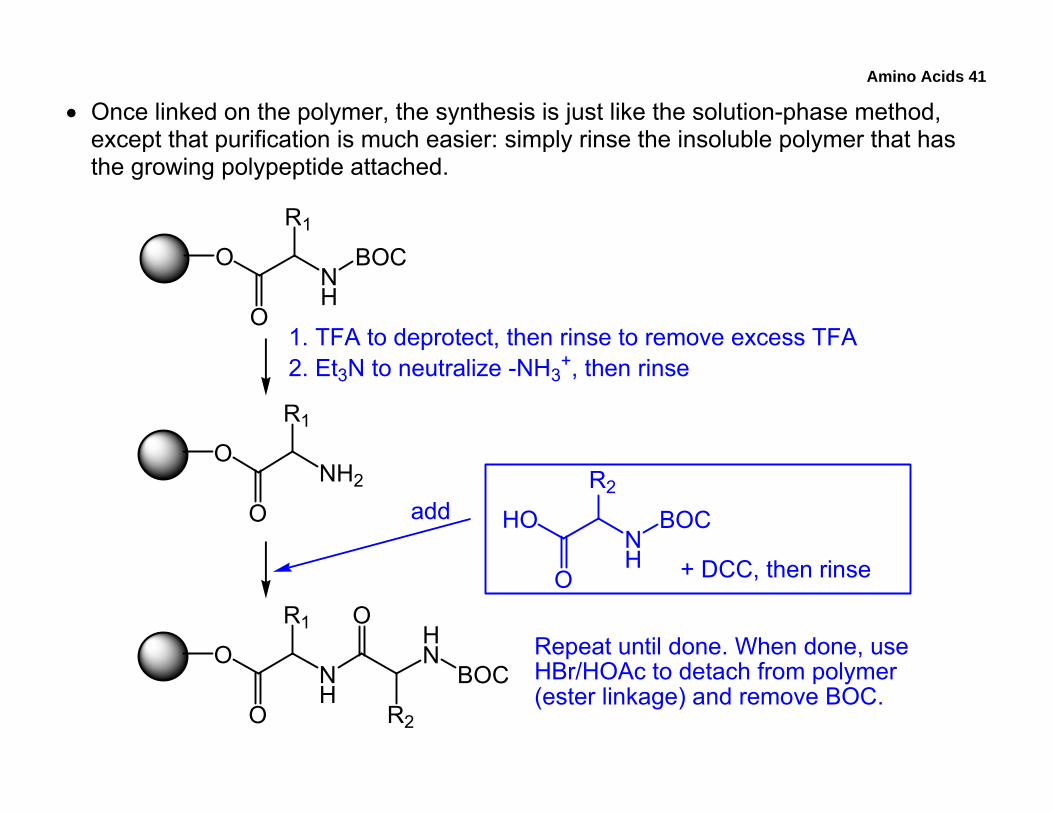
• Once linked on the polymer, the synthesis is just like the solution-phase method, except that purification is much easier: simply rinse the insoluble polymer that has the growing polypeptide attached.
NH
O
O
R1
BOC
NH2O
O
R1
1. TFA to deprotect, then rinse to remove excess TFA2. Et3N to neutralize -NH3
+, then rinse
NH
O
O
R1
NH
HO
O
R2
BOC
+ DCC, then rinse
add
O
R2
HN
BOCRepeat until done. When done, useHBr/HOAc to detach from polymer(ester linkage) and remove BOC.

Amino Acids 42
• Realize that in polypeptide synthesis, each one of the steps involved needs to proceed with a high efficiency. Relatively small changes in the yield of each step can lead to tremendous impact on the overall yield.
Number of steps: 100
Fractional yield per step Overall fractional yield 0.90 (0.90)100 = 0.00003 0.95 (0.95)100 = 0.005 0.99 (0.99)100 = 0.36 0.999 (0.999)100 = 0.90
• High efficiency is especially important at the coupling step. If there are 1000 growing polypeptides, and the coupling step is only 95% efficient, then only 950 will have the new residue attached, and hence the proper amino acid sequence. After the next coupling, only 903 will have the proper sequence. After 15 couplings, less than half.

Amino Acids 43
I. Enzymes and Catalysis
• Enzymes, which are a special class of proteins, are fascinating biological catalysts. They can increase the rates chemical of reactions up to a sextillion fold over their uncatalyzed counterparts.
1. Interactions in Substrate-Protein Binding
• For an enzyme to function as a catalyst, the enzyme needs to recognize and bind a substrate. These processes usually involve interactions between the substrate and the protein amide backbone and/or amino-acid side chains. In addition, the substrate must also be able to physically fit into the protein’s binding site.
• As seen in sulfanilamide, drugs often mimick natural substrates and competitively bind to the active site, thus preventing the natural substrate from binding.
• Three main interactions contribute to the initial binding of a substrate:
o Hydrogen bonding
o Hydrophobic forces
o Electrostatic forces

Amino Acids 44
a. Hydrogen bonding
• H-bonding arises from the interaction between a δ+ H covalently bonded to an electronegative atom and the lone pair of another electronegative atom (δ−). Hydrogen bonds are stronger when the H is more acidic and the lone pair is more basic. In practice, only H-bonding involving O and N are biologically significant.
• The typical strength of an H-bond is about 5 – 10 kcal/mol. However, H-bonding usually does not contribute to the substrate-protein binding energy, because any group that can H-bond will already be H-bonded to water. The binding of the substrate to the protein simply replaces these existing H-bonds.
AH OH2
enzyme
AH OH2
enzyme
BHOH+
BHOH
incorrect
+
AH B
enzyme
+ OH2HOH
AHB
enzyme
+ OH2HOH ΔG =
ΔG =
• If the substrate is incorrect or is oriented improperly, the interaction is unfavourable.
Amino Acids 45
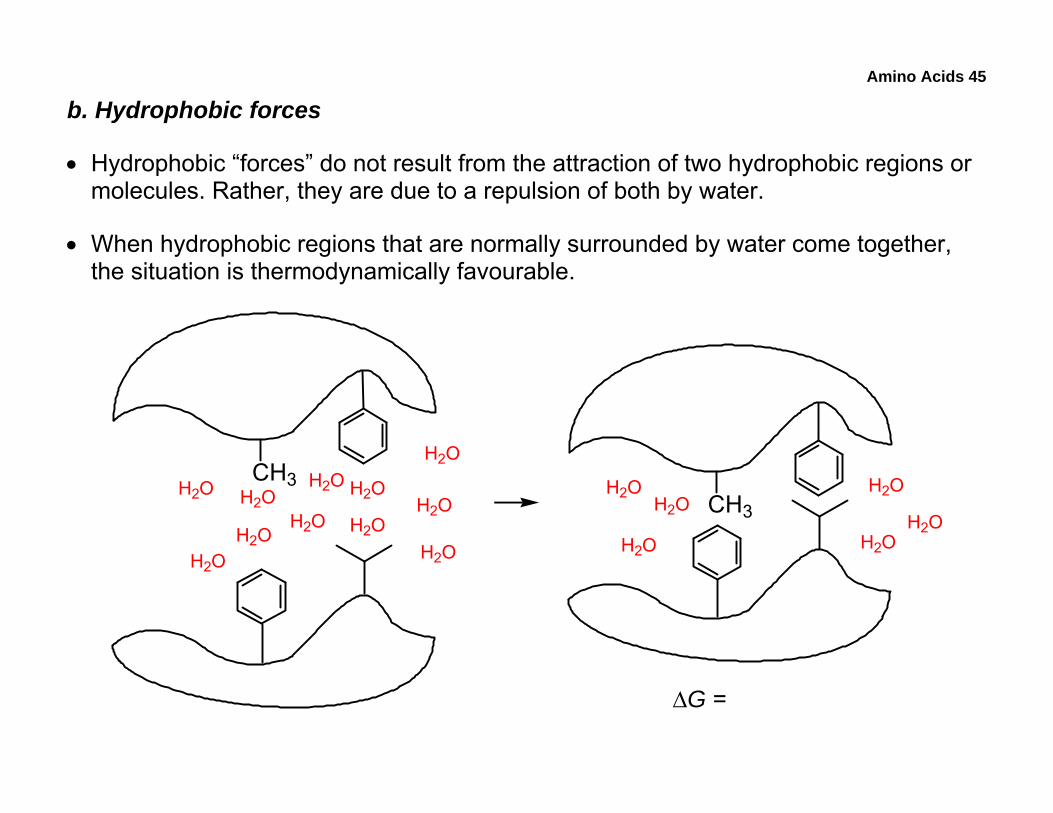
b. Hydrophobic forces
• Hydrophobic “forces” do not result from the attraction of two hydrophobic regions or molecules. Rather, they are due to a repulsion of both by water.
• When hydrophobic regions that are normally surrounded by water come together, the situation is thermodynamically favourable.
CH3CH3
H2OH2O
H2O
H2O
H2O
H2O
H2OH2O
H2OH2O
H2OH2O H2O
H2O
H2OH2O
H2O
ΔG =
Amino Acids 46
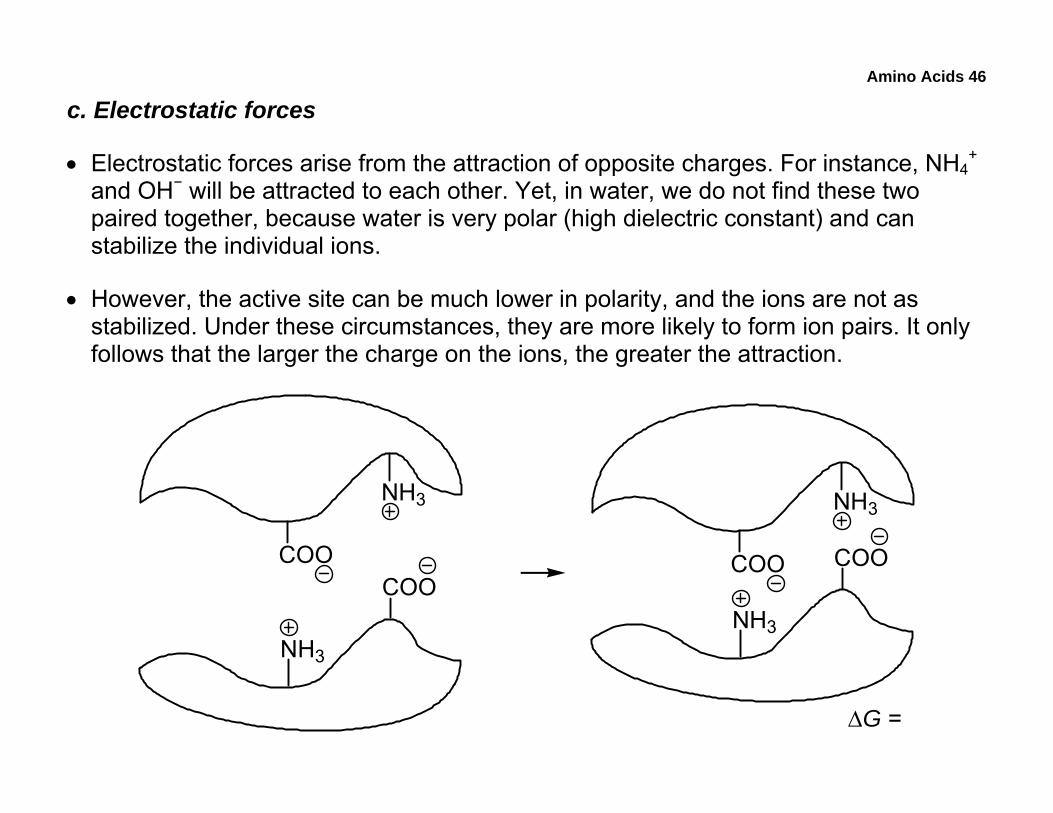
c. Electrostatic forces
• Electrostatic forces arise from the attraction of opposite charges. For instance, NH4+
and OH− will be attracted to each other. Yet, in water, we do not find these two paired together, because water is very polar (high dielectric constant) and can stabilize the individual ions.
• However, the active site can be much lower in polarity, and the ions are not as stabilized. Under these circumstances, they are more likely to form ion pairs. It only follows that the larger the charge on the ions, the greater the attraction.
NH3
COOCOO
NH3
ΔG =
NH3
COO COO
NH3
Amino Acids 47
2. Chiral Environment of Enzymes
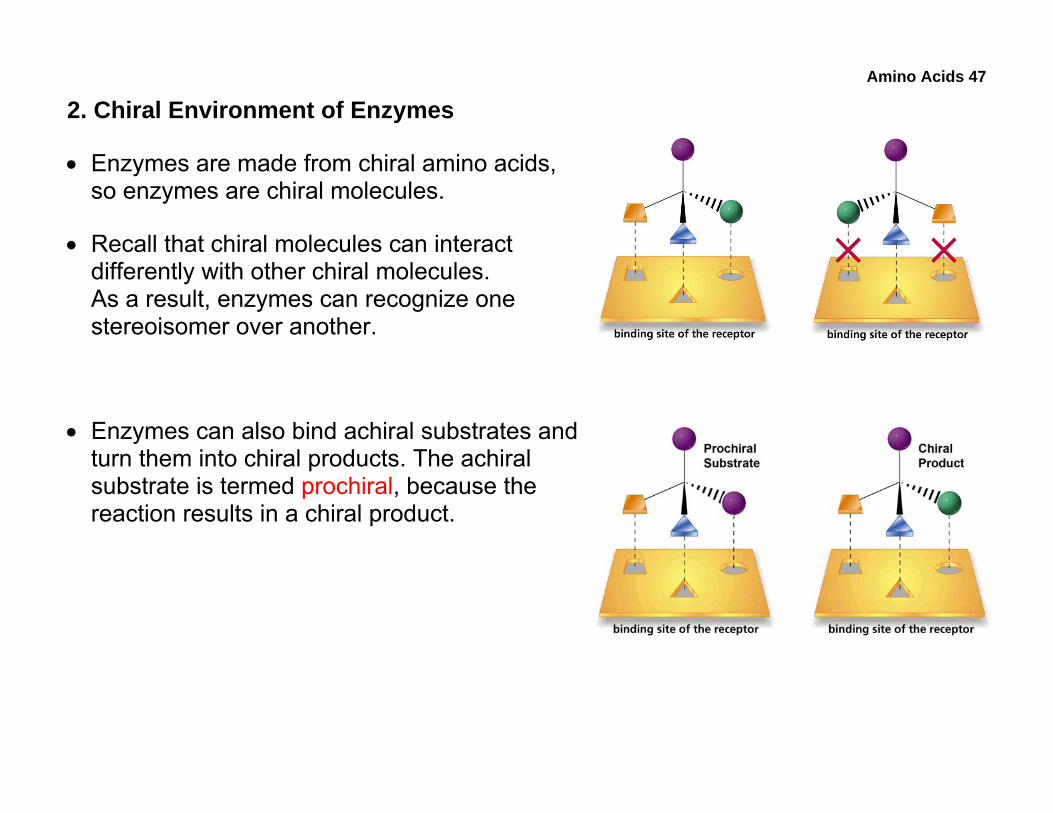
• Enzymes are made from chiral amino acids, so enzymes are chiral molecules.
• Recall that chiral molecules can interact differently with other chiral molecules. As a result, enzymes can recognize one stereoisomer over another.
• Enzymes can also bind achiral substrates and turn them into chiral products. The achiral substrate is termed prochiral, because the reaction results in a chiral product.
Amino Acids 48
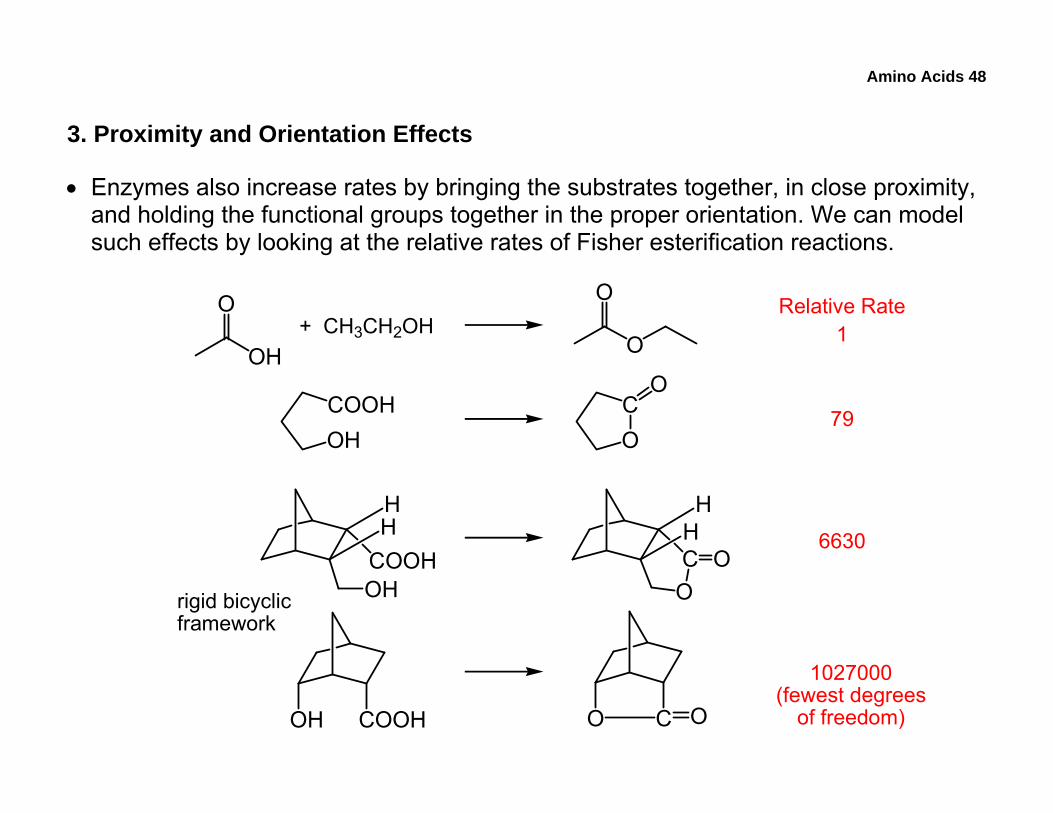
3. Proximity and Orientation Effects
• Enzymes also increase rates by bringing the substrates together, in close proximity, and holding the functional groups together in the proper orientation. We can model such effects by looking at the relative rates of Fisher esterification reactions.
OH
O+ CH3CH2OH
O
O Relative Rate1
OHCOOH
OC
O
COOHOH
HH
CO
HH
O
OH COOH O C O
rigid bicyclicframework
79
6630
1027000(fewest degrees
of freedom)
Amino Acids 49
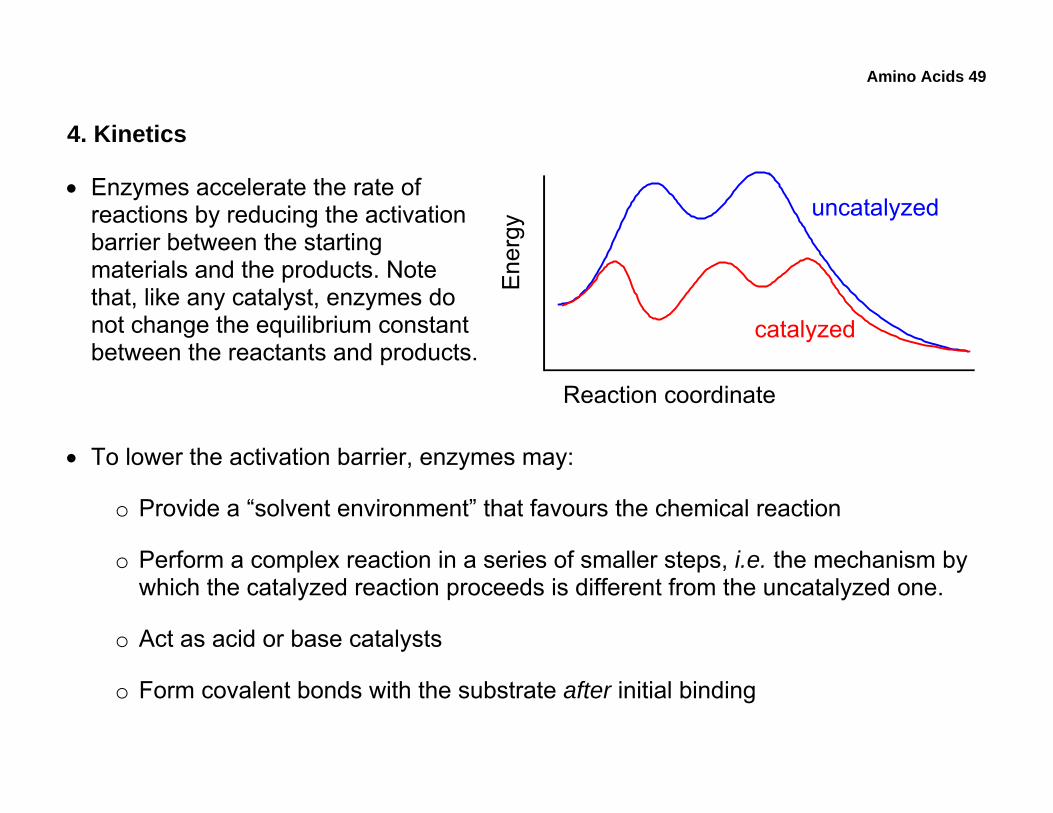
4. Kinetics
• Enzymes accelerate the rate of reactions by reducing the activation barrier between the starting materials and the products. Note that, like any catalyst, enzymes do not change the equilibrium constant between the reactants and products.
• To lower the activation barrier, enzymes may:
o Provide a “solvent environment” that favours the chemical reaction
o Perform a complex reaction in a series of smaller steps, i.e. the mechanism by which the catalyzed reaction proceeds is different from the uncatalyzed one.
o Act as acid or base catalysts
o Form covalent bonds with the substrate after initial binding
Reaction coordinate
Ener
gy
uncatalyzed
catalyzed

Amino Acids 50
5. Example of Enzyme Mechanism: Chymotrypsin (see video online)
• Chymotrypsin belongs to a family of enzymes known as serine proteases, and it is a digestive enzyme that is present in the small intestine.
• The enzyme hydrolyzes peptides with sequence specificity: on the C-terminal side of hydrophobic amino acids (best are Trp, Tyr, and Phe). Note that although Tyr is considered polar, it also has a hydrophobic region, the aromatic ring.
• When the sequence of a peptide is written in the standard N C convention, hydrolysis would occur after the hydrophobic amino acid.
• Hydrolysis creates a new a C-terminal on the N-terminal fragment of the original substrate, and a new N-terminal on the C-terminal fragment of the original substrate.
• A related digestive enzyme, trypsin, hydrolyzes after basic residues (Arg and Lys).

Amino Acids 51
• Why does the hydrolysis of an amide need to be enzyme-catalyzed?
o Amide bonds are very stable.
o Hydrolysis by base involves the formation of a tetrahedral intermediate, followed by its collapse and the loss of the leaving group.
R NHR'
O
R NHR'
O
OH
R OH
OHO
NHR'
o Biological pH is ~7, so hydroxide is not present in very large amounts. Chymotrypsin provides the base (Ser) necessary for catalysis.
o The tetrahedral, oxyanion intermediate needs to be stabilized, and the enzyme accomplishes this task by hydrogen-bonding it to the enzyme backbone.
o The amine (the new N-terminal of the original C-terminal fragment) is not a good leaving group, so the enzyme protonates it as it leaves.
Amino Acids 52
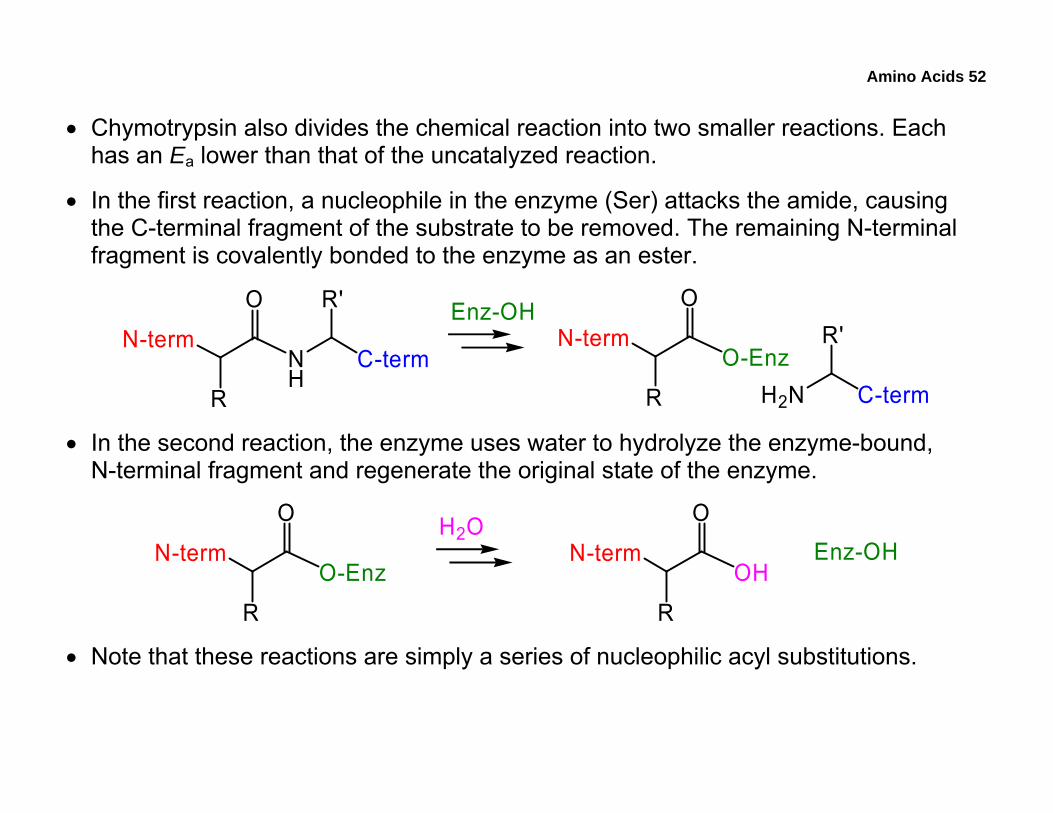
• Chymotrypsin also divides the chemical reaction into two smaller reactions. Each has an Ea lower than that of the uncatalyzed reaction.
• In the first reaction, a nucleophile in the enzyme (Ser) attacks the amide, causing the C-terminal fragment of the substrate to be removed. The remaining N-terminal fragment is covalently bonded to the enzyme as an ester.
N-term
O
R
NH
C-term
R' Enz-OHN-term
O
R
O-EnzH2N C-term
R'
• In the second reaction, the enzyme uses water to hydrolyze the enzyme-bound, N-terminal fragment and regenerate the original state of the enzyme.
N-term
O
R
OHN-term
O
R
O-Enz
H2OEnz-OH
• Note that these reactions are simply a series of nucleophilic acyl substitutions.
Amino Acids 53
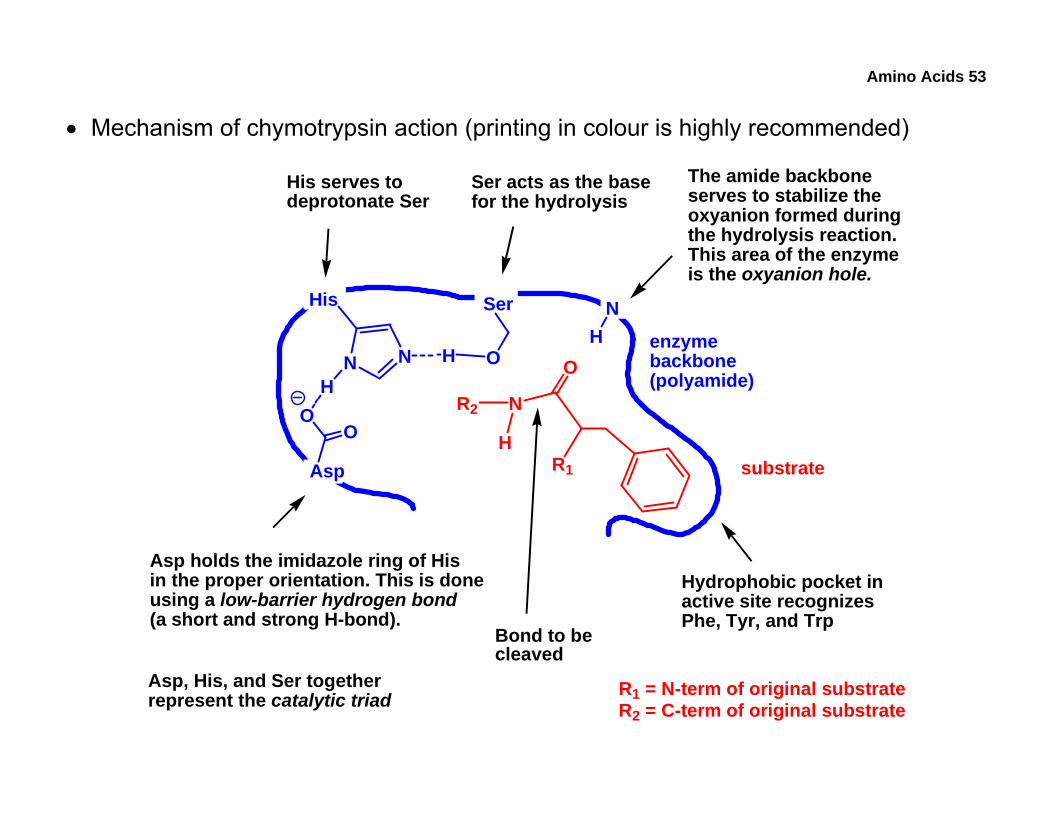
• Mechanism of chymotrypsin action (printing in colour is highly recommended)
enzymebackbone(polyamide)
Asp
OO
His
N N O
N
O
R1
Ser
R2
H
HH
NH
substrate
Hydrophobic pocket inactive site recognizesPhe, Tyr, and Trp
Ser acts as the basefor the hydrolysis
His serves todeprotonate Ser
Asp holds the imidazole ring of Hisin the proper orientation. This is doneusing a low-barrier hydrogen bond(a short and strong H-bond).
The amide backboneserves to stabilize theoxyanion formed duringthe hydrolysis reaction.This area of the enzymeis the oxyanion hole.
Bond to becleaved
R1 = N-term of original substrateR2 = C-term of original substrate
Asp, His, and Ser togetherrepresent the catalytic triad
Amino Acids 54
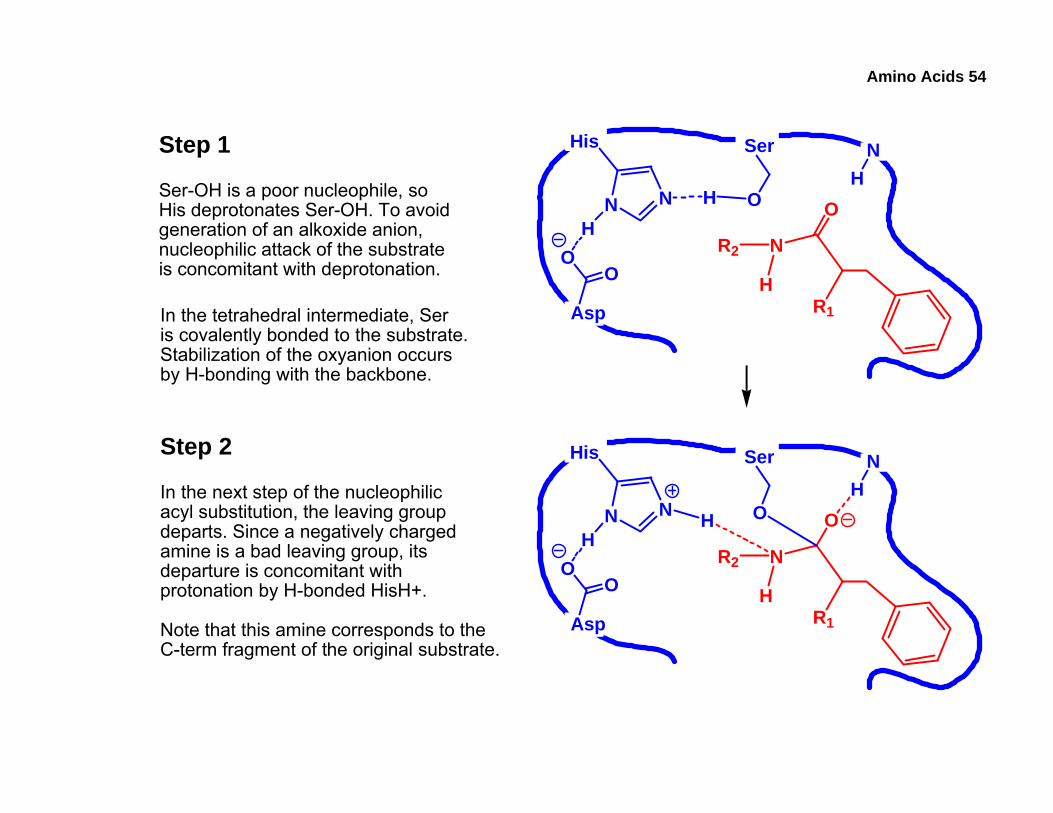
Step 1
Asp
OO
His
N N O
N
O
R1
Ser
R2
H
HH
NHSer-OH is a poor nucleophile, so
His deprotonates Ser-OH. To avoidgeneration of an alkoxide anion,nucleophilic attack of the substrateis concomitant with deprotonation.
In the tetrahedral intermediate, Seris covalently bonded to the substrate.Stabilization of the oxyanion occursby H-bonding with the backbone.
Asp
OO
His
N N O
N
O
R1
Ser
R2
H
HH
NH
Step 2
In the next step of the nucleophilicacyl substitution, the leaving groupdeparts. Since a negatively chargedamine is a bad leaving group, itsdeparture is concomitant withprotonation by H-bonded HisH+.
Note that this amine corresponds to theC-term fragment of the original substrate.
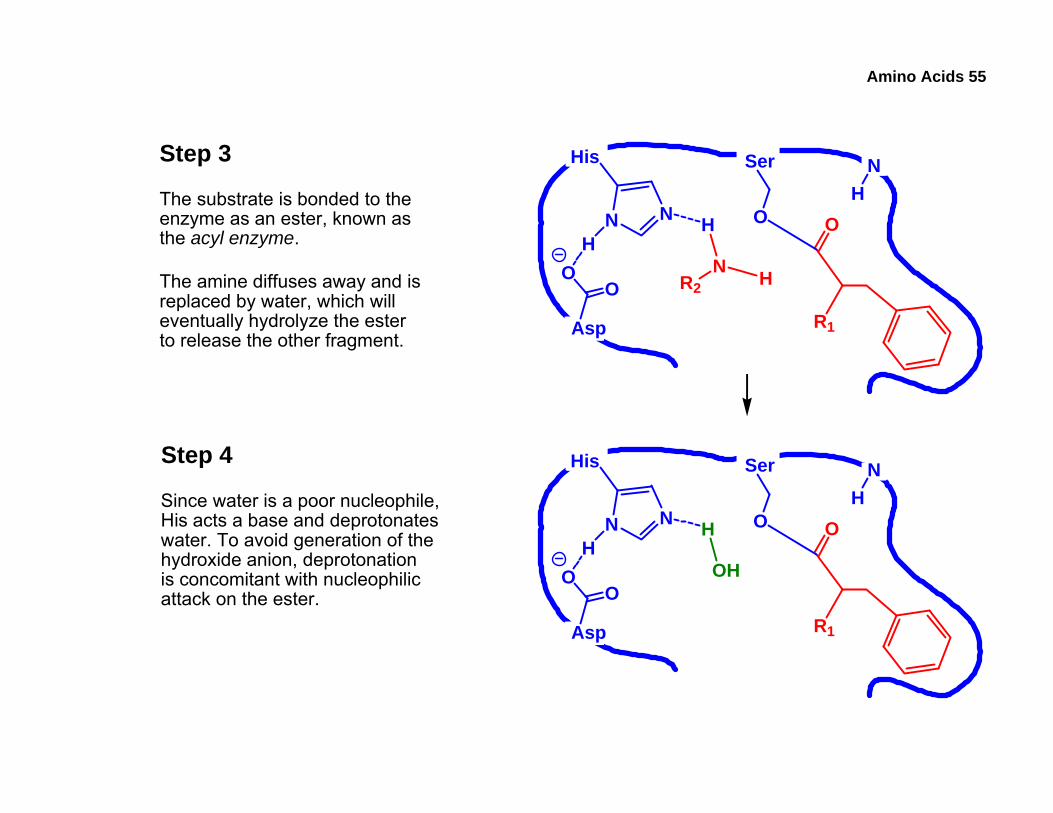
Amino Acids 55
Step 3
The substrate is bonded to theenzyme as an ester, known asthe acyl enzyme.
The amine diffuses away and isreplaced by water, which willeventually hydrolyze the esterto release the other fragment. Asp
OO
His
N N O
N
O
R1
Ser
R2 H
HH
NH
Step 4
Since water is a poor nucleophile,His acts a base and deprotonateswater. To avoid generation of thehydroxide anion, deprotonationis concomitant with nucleophilicattack on the ester.
Asp
OO
His
N N O
OH
O
R1
Ser
HH
NH
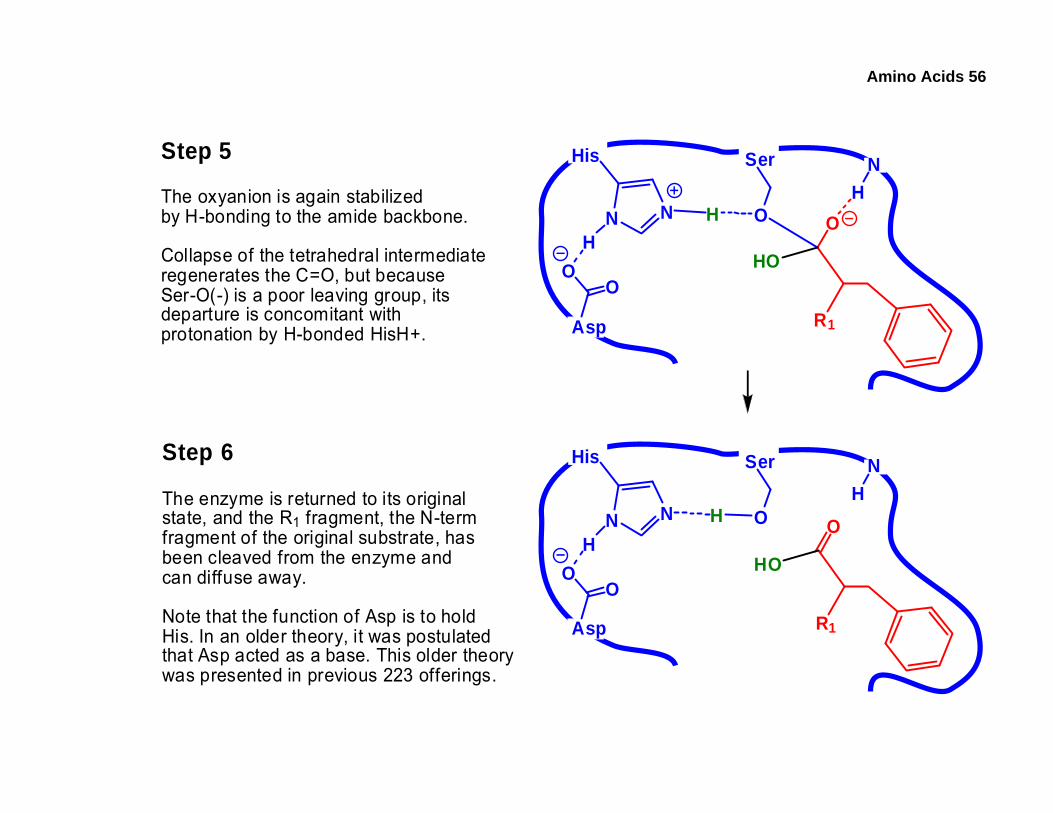
Amino Acids 56
Step 5
The oxyanion is again stabilizedby H-bonding to the amide backbone.
Collapse of the tetrahedral intermediateregenerates the C=O, but becauseSer-O(-) is a poor leaving group, itsdeparture is concomitant withprotonation by H-bonded HisH+.
Step 6
The enzyme is returned to its originalstate, and the R1 fragment, the N-termfragment of the original substrate, hasbeen cleaved from the enzyme andcan diffuse away.
Note that the function of Asp is to holdHis. In an older theory, it was postulatedthat Asp acted as a base. This older theorywas presented in previous 223 offerings.
Asp
OO
His
N N O
HO
O
R1
Ser
HH
NH
Asp
OO
His
N N O
HO
O
R1
Ser
H
NH
H
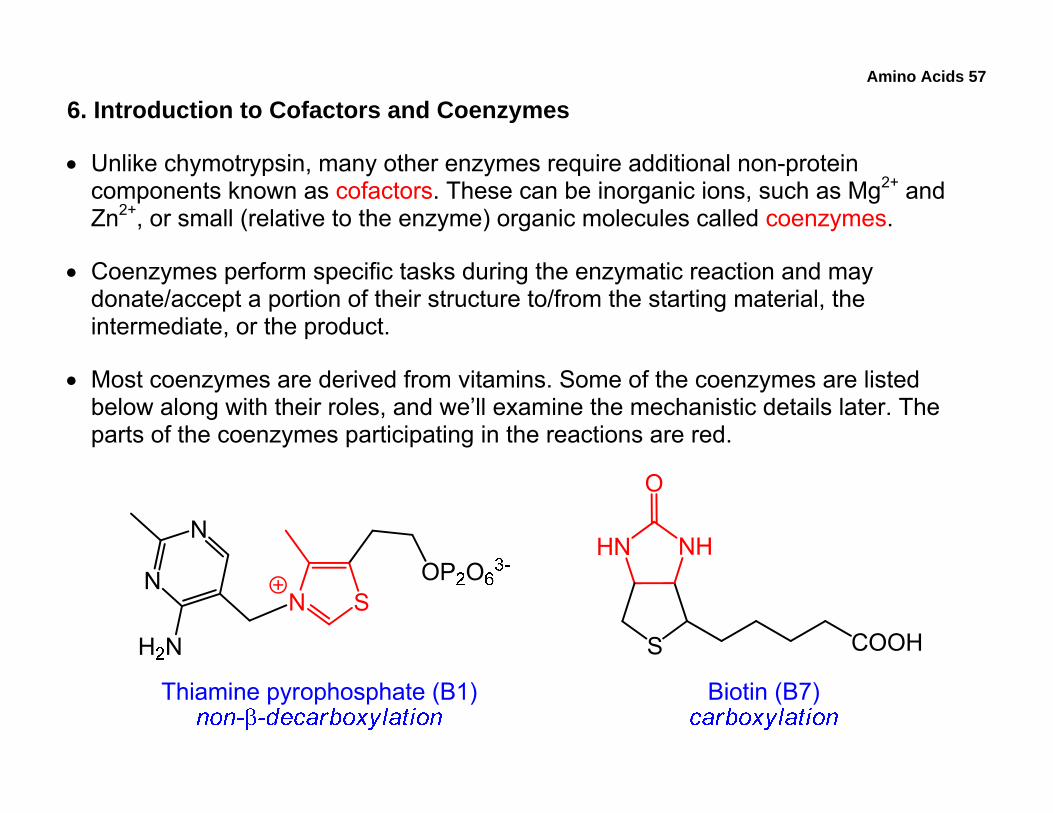
Amino Acids 57
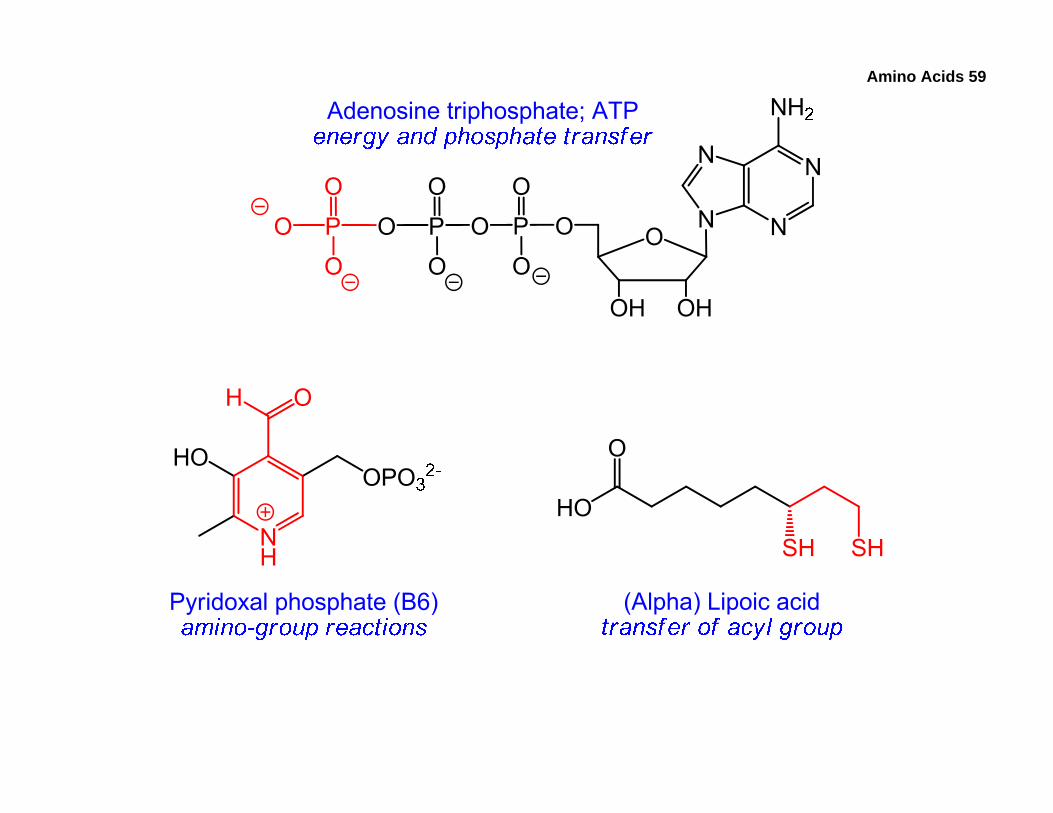
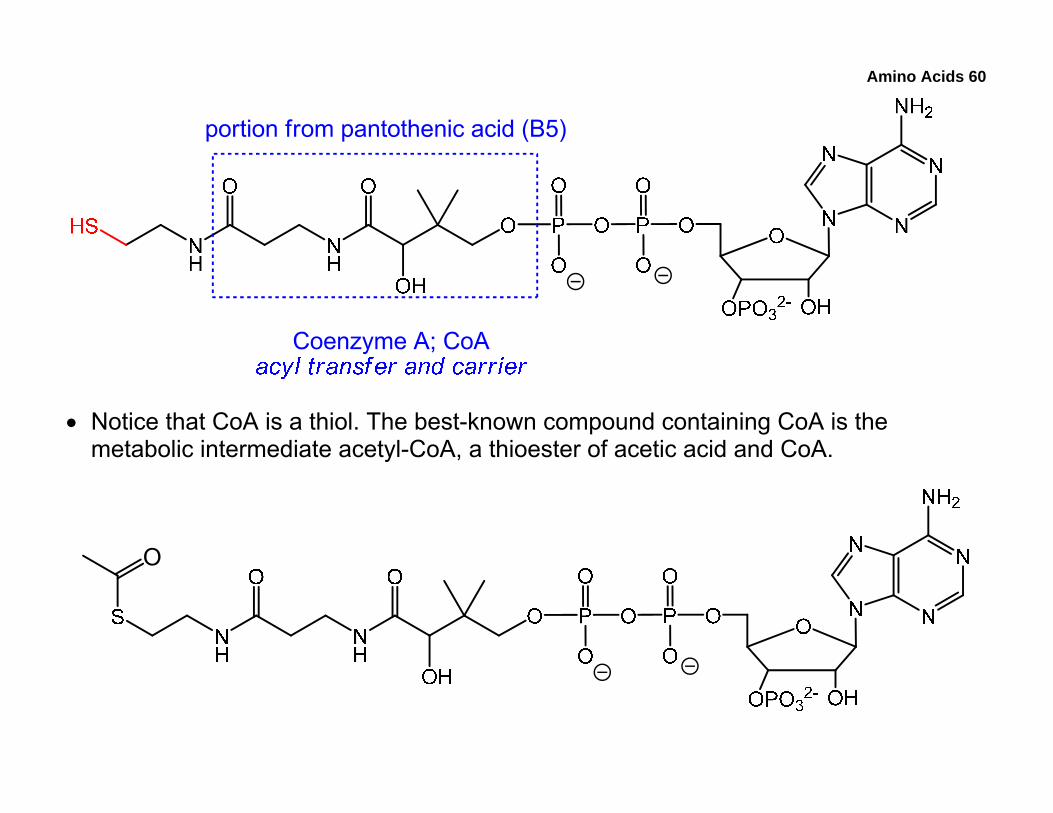
6. Introduction to Cofactors and Coenzymes
• Unlike chymotrypsin, many other enzymes require additional non-protein components known as cofactors. These can be inorganic ions, such as Mg2+ and Zn2+, or small (relative to the enzyme) organic molecules called coenzymes.
• Coenzymes perform specific tasks during the enzymatic reaction and may donate/accept a portion of their structure to/from the starting material, the intermediate, or the product.
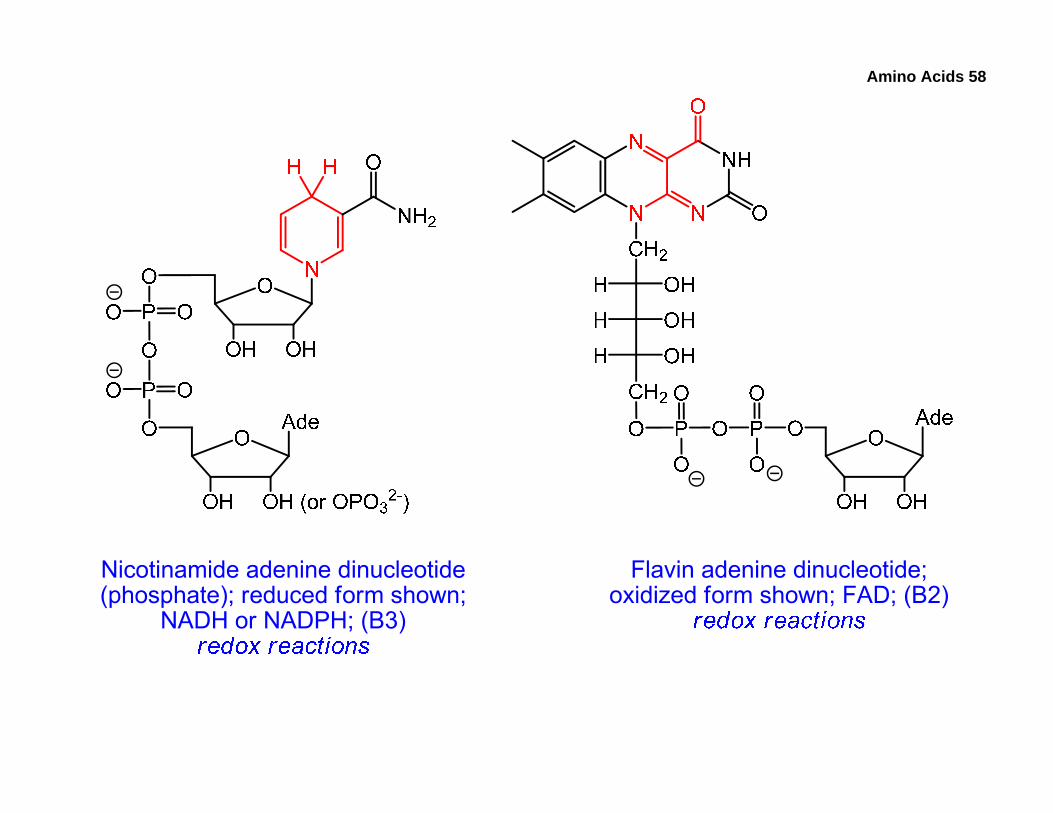
• Most coenzymes are derived from vitamins. Some of the coenzymes are listed below along with their roles, and we’ll examine the mechanistic details later. The parts of the coenzymes participating in the reactions are red.
Amino Acids 58
Amino Acids 59
Amino Acids 60
• Notice that CoA is a thiol. The best-known compound containing CoA is the metabolic intermediate acetyl-CoA, a thioester of acetic acid and CoA.

Amino Acids 61
• Up to now, some of the concepts we’ve covered include:
o Laboratory peptide synthesis
Protecting groups, coupling agents, advantages of solid-phase synthesis
o Enzymatic reactions
Enzyme-substrate interactions, how enzymes can accelerate reactions
o Mechanism of chymotrypsin
o Important cofactors and their biochemical roles
• Using amino-acid structures provided in the lab manual, attempt:
o Practice problems: 23 – 47 (save #34 for later)
o 2008 Midterm: 17 – 21
• For exam-studying purposes, review all the notes and redo the practice problems in the lab manual. Understand and be able to apply the concepts.
Amino Acids 62
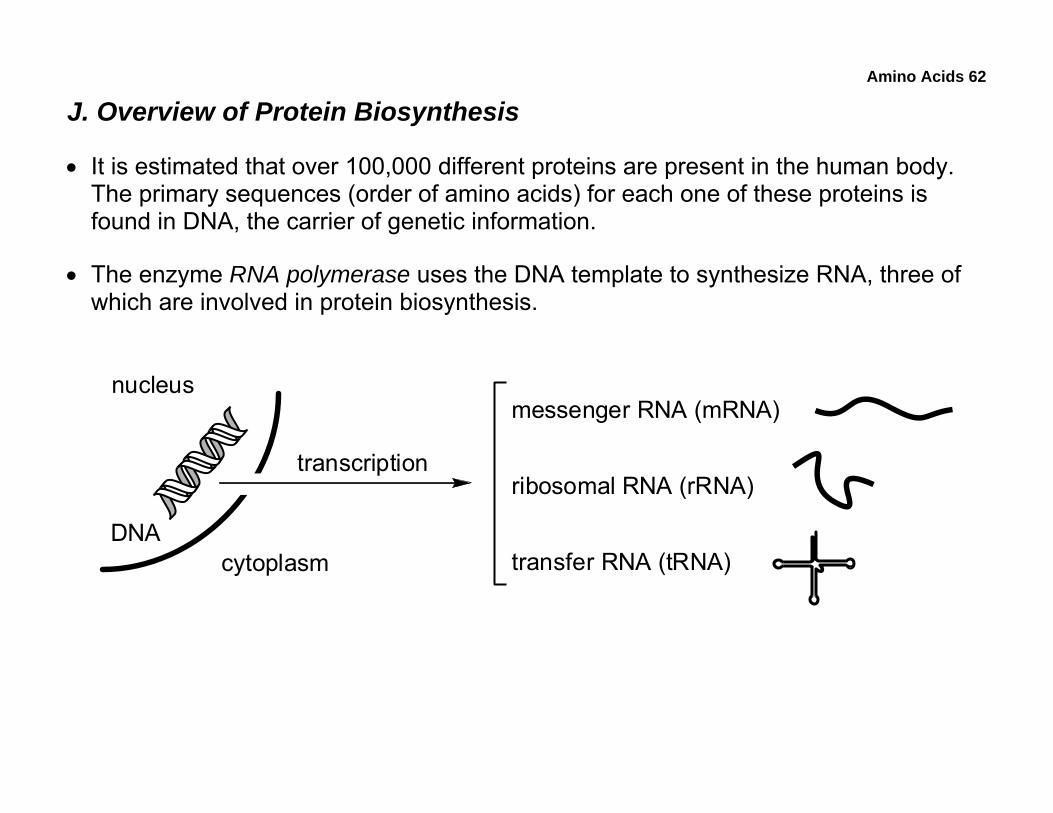
J. Overview of Protein Biosynthesis
• It is estimated that over 100,000 different proteins are present in the human body. The primary sequences (order of amino acids) for each one of these proteins is found in DNA, the carrier of genetic information.
• The enzyme RNA polymerase uses the DNA template to synthesize RNA, three of which are involved in protein biosynthesis.
nucleus
DNA
transcription
messenger RNA (mRNA)
ribosomal RNA (rRNA)
transfer RNA (tRNA)cytoplasm
Amino Acids 63
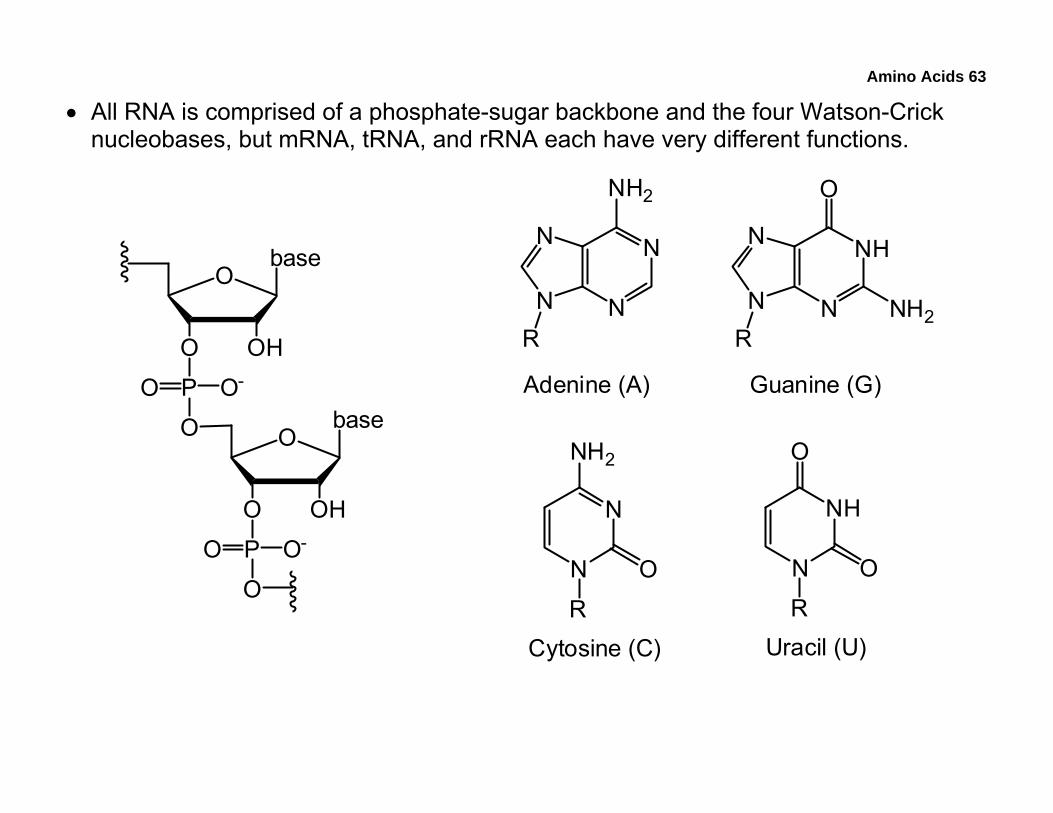
• All RNA is comprised of a phosphate-sugar backbone and the four Watson-Crick nucleobases, but mRNA, tRNA, and rRNA each have very different functions.
N
NN
N
NH2
NH
NN
N
O
NH2
N
N
NH2
O
NH
N
O
O
R R
R R
Adenine (A) Guanine (G)
Cytosine (C) Uracil (U)
O
OHO
PO
O
O-
base
O
OHO
PO
O
O-
base
Amino Acids 64
1. mRNA
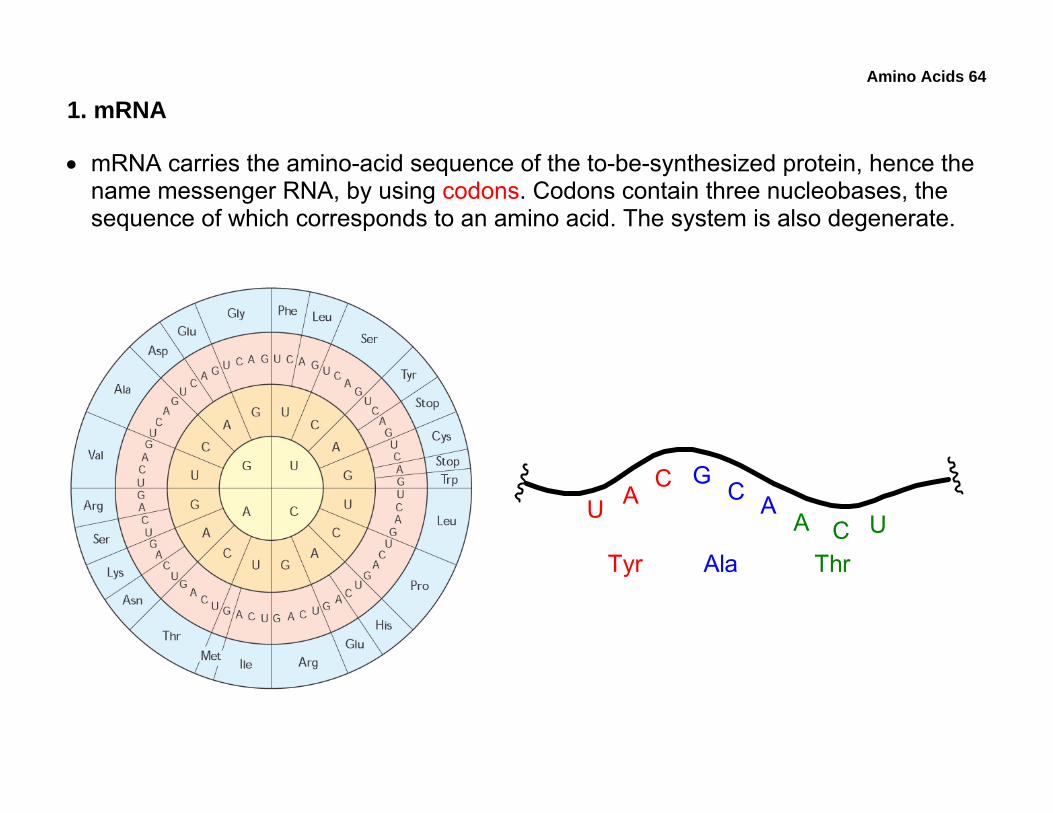
• mRNA carries the amino-acid sequence of the to-be-synthesized protein, hence the name messenger RNA, by using codons. Codons contain three nucleobases, the sequence of which corresponds to an amino acid. The system is also degenerate.
U AC
Tyr
GC A
Ala
A C UThr
Amino Acids 65
2. tRNA
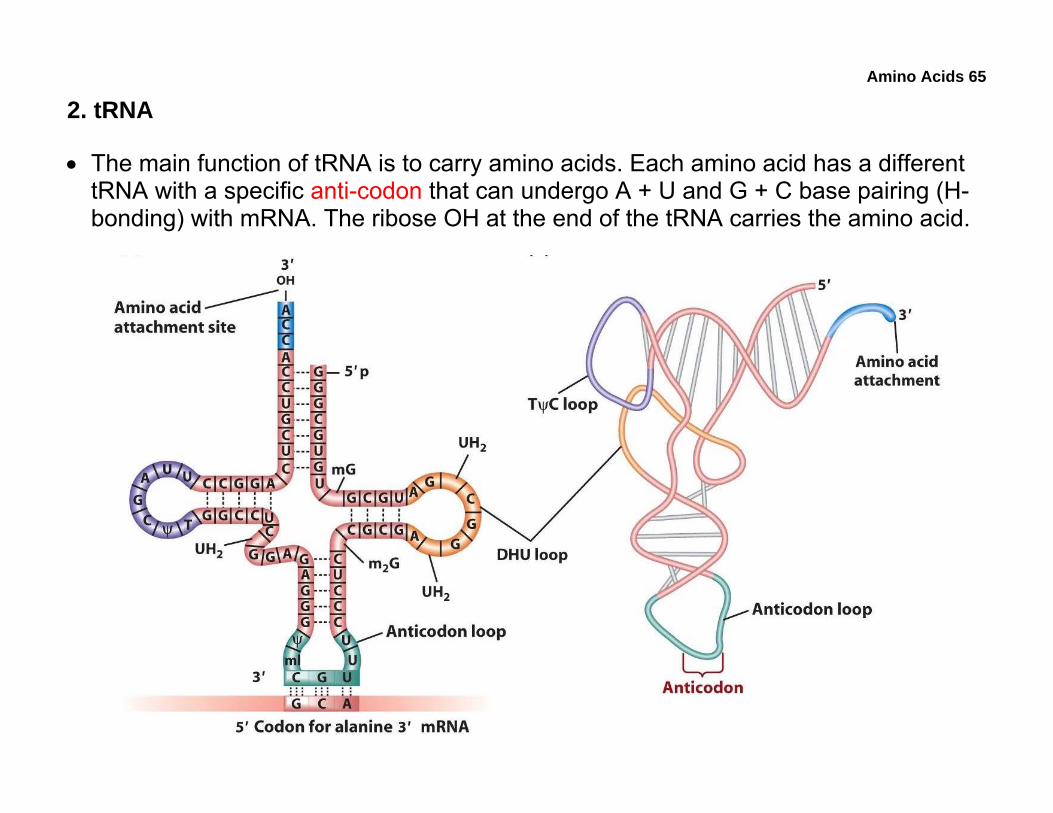
• The main function of tRNA is to carry amino acids. Each amino acid has a different tRNA with a specific anti-codon that can undergo A + U and G + C base pairing (H-bonding) with mRNA. The ribose OH at the end of the tRNA carries the amino acid.
Amino Acids 66
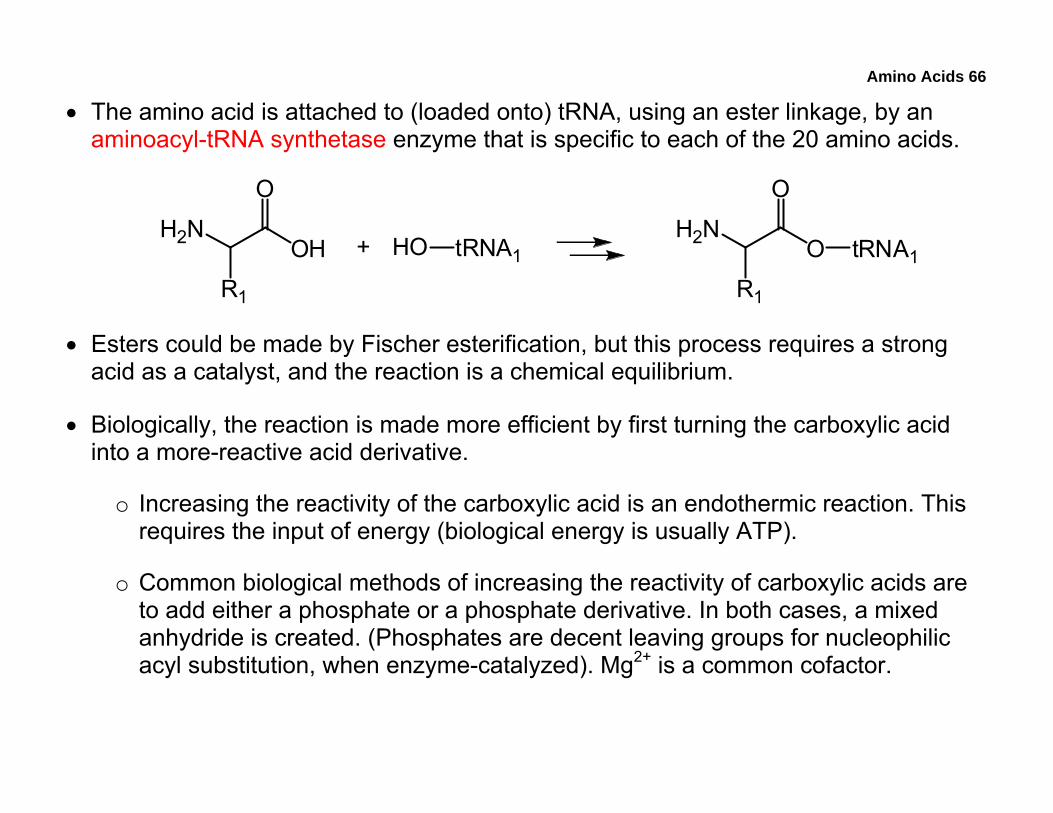
• The amino acid is attached to (loaded onto) tRNA, using an ester linkage, by an aminoacyl-tRNA synthetase enzyme that is specific to each of the 20 amino acids.
H2NOH
O
R1
+H2N
O
O
R1
tRNA1HO tRNA1
• Esters could be made by Fischer esterification, but this process requires a strong acid as a catalyst, and the reaction is a chemical equilibrium.
• Biologically, the reaction is made more efficient by first turning the carboxylic acid into a more-reactive acid derivative.
o Increasing the reactivity of the carboxylic acid is an endothermic reaction. This requires the input of energy (biological energy is usually ATP).
o Common biological methods of increasing the reactivity of carboxylic acids are to add either a phosphate or a phosphate derivative. In both cases, a mixed anhydride is created. (Phosphates are decent leaving groups for nucleophilic acyl substitution, when enzyme-catalyzed). Mg2+ is a common cofactor.
Amino Acids 67
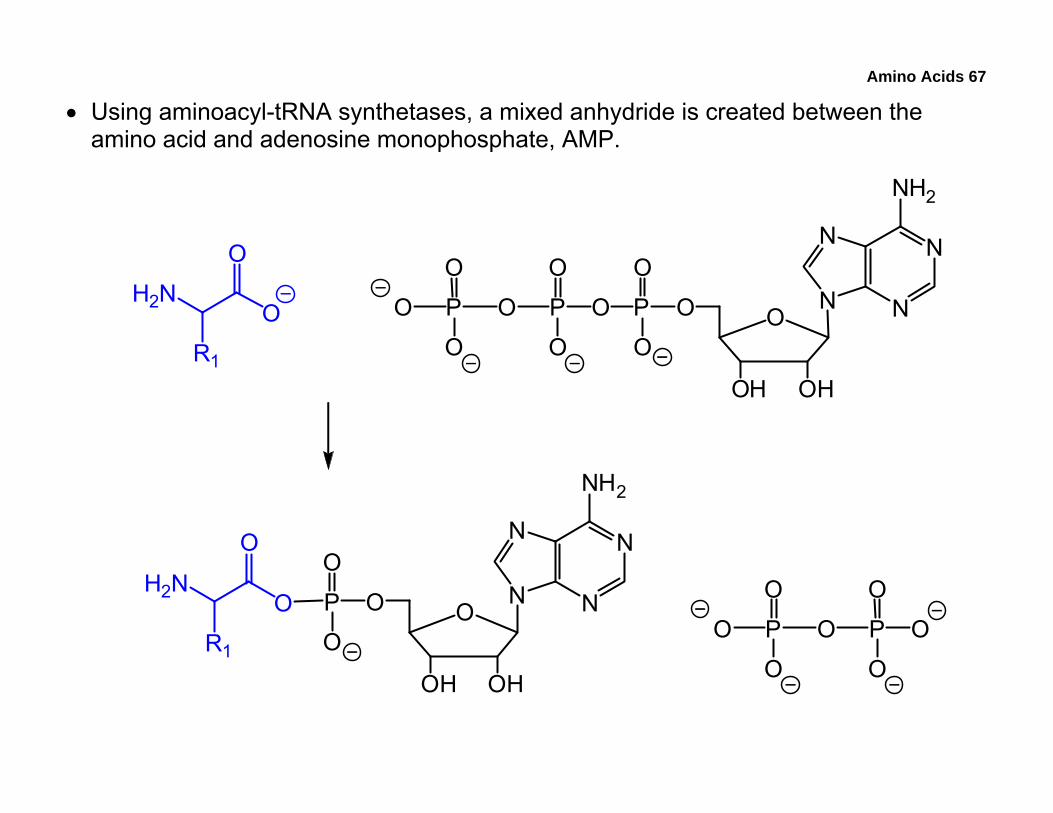
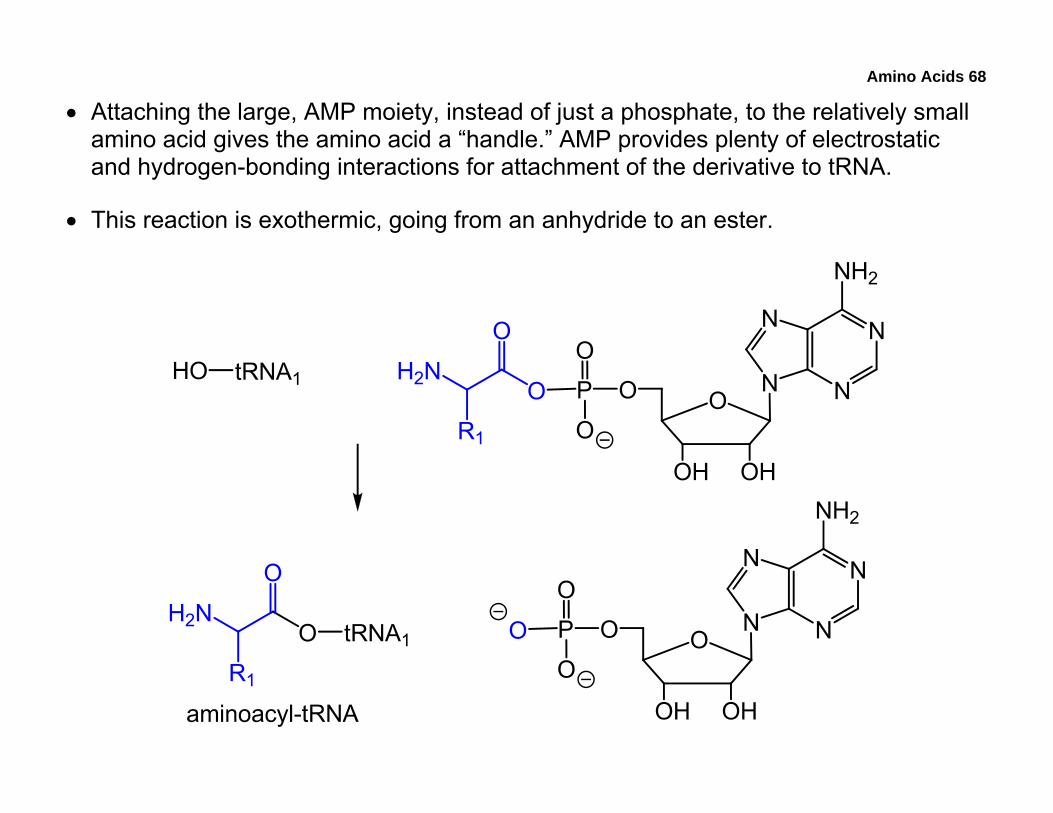
• Using aminoacyl-tRNA synthetases, a mixed anhydride is created between the amino acid and adenosine monophosphate, AMP.
O
OHOH
OPO
O
O
P
O
O
OP
O
O
O
N
NN
N
NH2
H2NO
O
R1
O
OHOH
OP
O
ON
NN
N
NH2
H2NO
O
R1OP
O
O
OP
O
O
O
Amino Acids 68
• Attaching the large, AMP moiety, instead of just a phosphate, to the relatively small amino acid gives the amino acid a “handle.” AMP provides plenty of electrostatic and hydrogen-bonding interactions for attachment of the derivative to tRNA.
• This reaction is exothermic, going from an anhydride to an ester.
2
2
1
2
1
1
1
2
Amino Acids 69
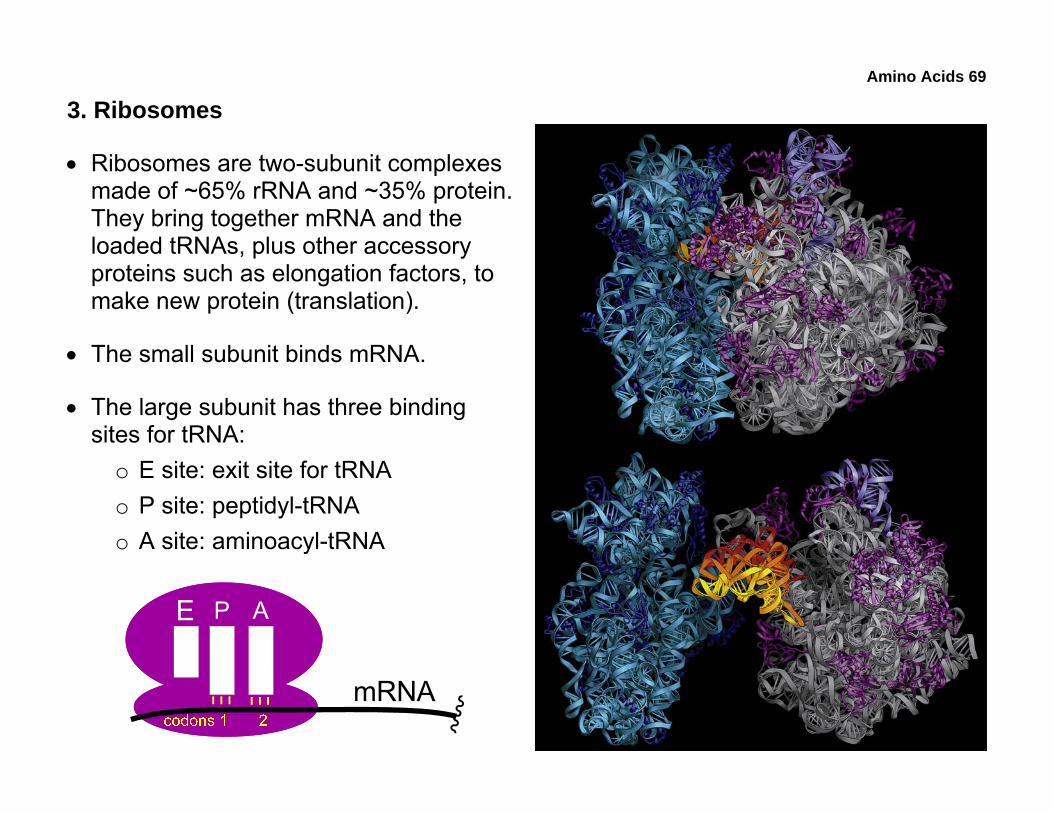
3. Ribosomes
• Ribosomes are two-subunit complexes made of ~65% rRNA and ~35% protein. They bring together mRNA and the loaded tRNAs, plus other accessory proteins such as elongation factors, to make new protein (translation).
• The small subunit binds mRNA.
• The large subunit has three binding sites for tRNA:
o E site: exit site for tRNA o P site: peptidyl-tRNA o A site: aminoacyl-tRNA
Amino Acids 70
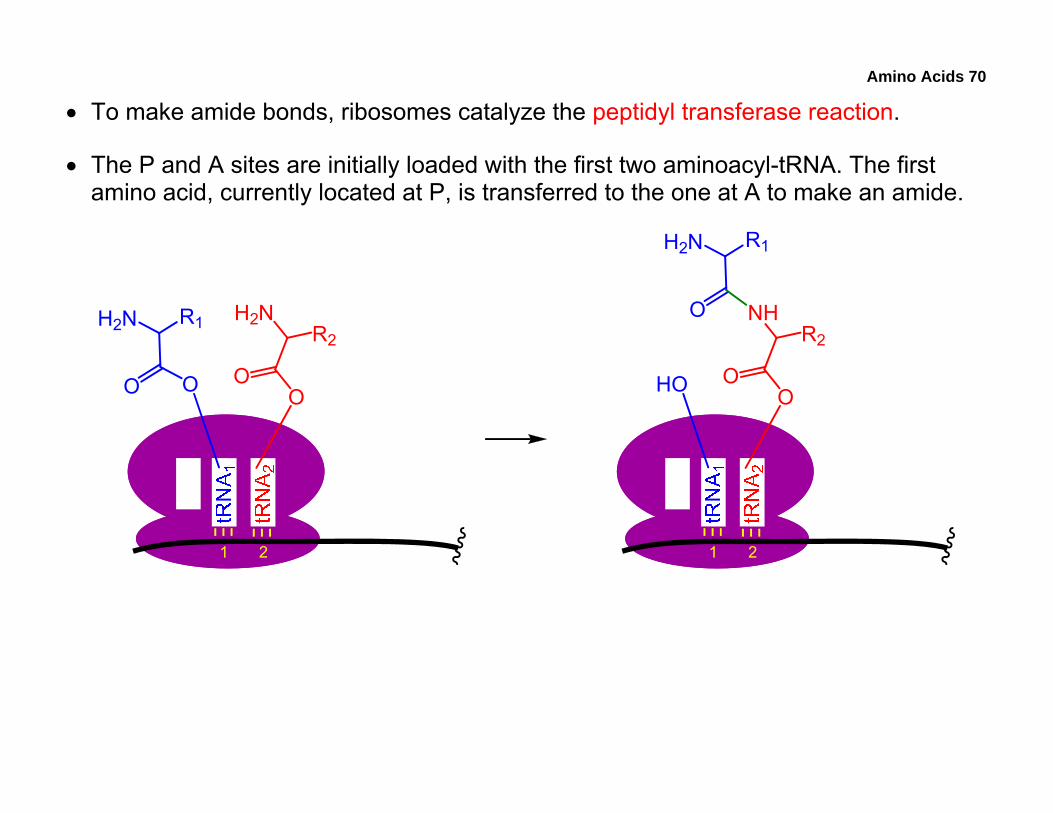
• To make amide bonds, ribosomes catalyze the peptidyl transferase reaction.
• The P and A sites are initially loaded with the first two aminoacyl-tRNA. The first amino acid, currently located at P, is transferred to the one at A to make an amide.
1 2
H2N
OO
R1 H2N
OO
R2
1 2
HO
NH
OO
R2
H2N
O
R1
Amino Acids 71
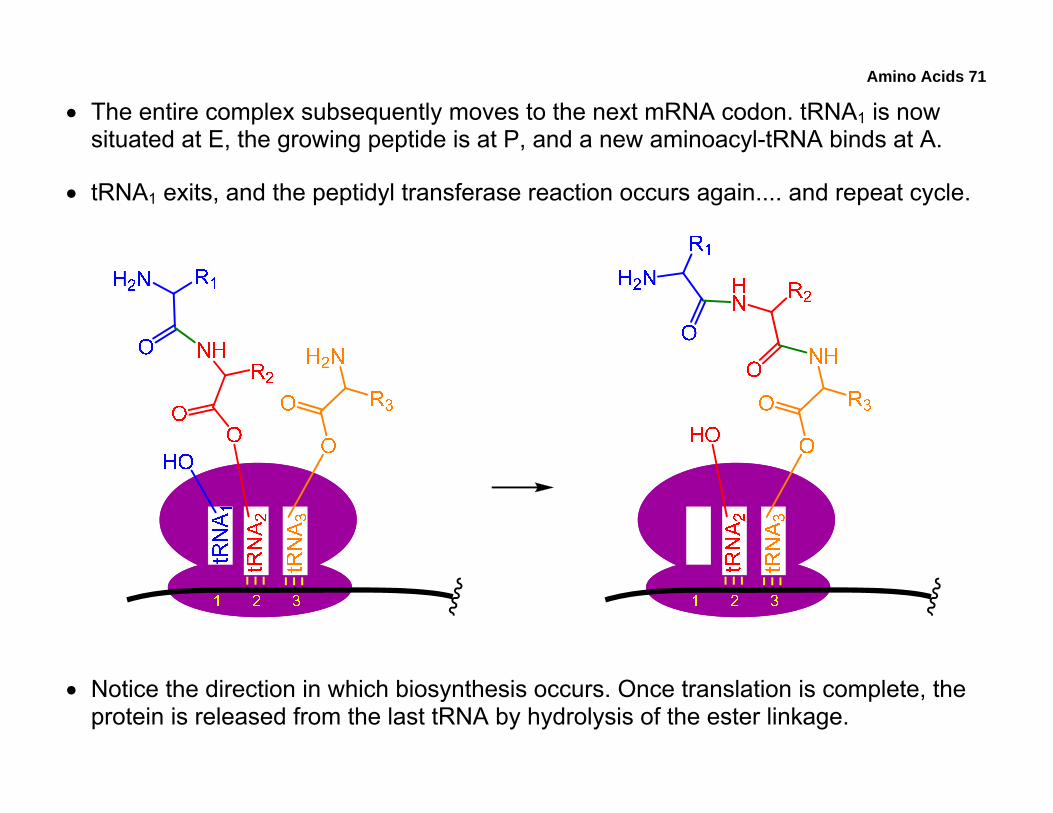
• The entire complex subsequently moves to the next mRNA codon. tRNA1 is now situated at E, the growing peptide is at P, and a new aminoacyl-tRNA binds at A.
• tRNA1 exits, and the peptidyl transferase reaction occurs again.... and repeat cycle.
• Notice the direction in which biosynthesis occurs. Once translation is complete, the protein is released from the last tRNA by hydrolysis of the ester linkage.
Amino Acids 72
• What is the mechanism of the peptidyl transferase reaction?
• Ribosomes were discovered in the 1950’s with the aid of the electron microscope, and by the late 1960’s, scientists had a relatively good knowledge of the general structure of the ribosome and its RNA and protein components.
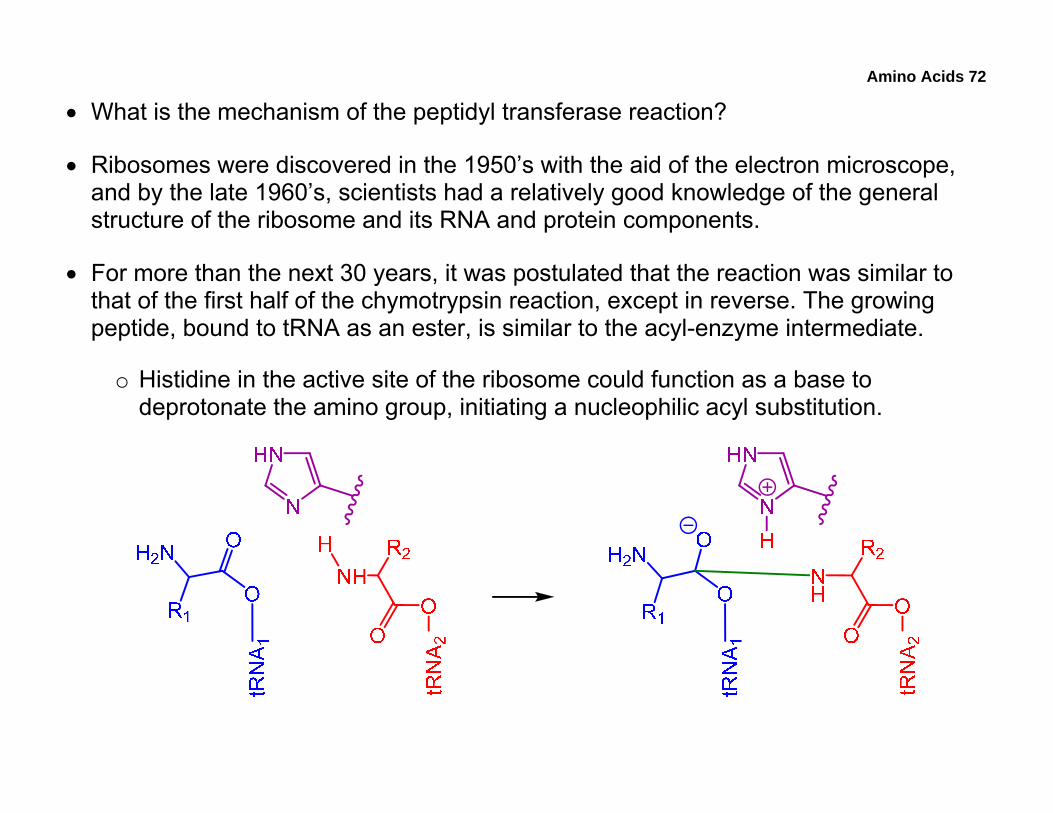
• For more than the next 30 years, it was postulated that the reaction was similar to that of the first half of the chymotrypsin reaction, except in reverse. The growing peptide, bound to tRNA as an ester, is similar to the acyl-enzyme intermediate.
o Histidine in the active site of the ribosome could function as a base to deprotonate the amino group, initiating a nucleophilic acyl substitution.
Amino Acids 73
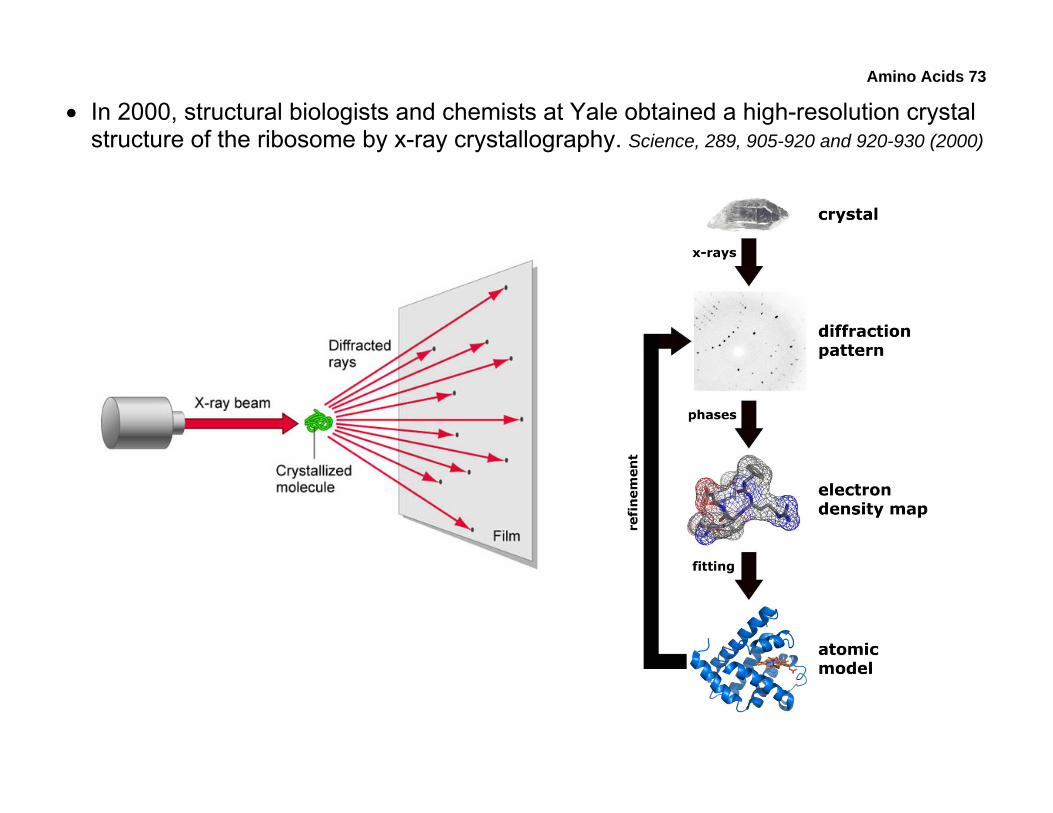
• In 2000, structural biologists and chemists at Yale obtained a high-resolution crystal structure of the ribosome by x-ray crystallography. Science, 289, 905-920 and 920-930 (2000)
Amino Acids 74
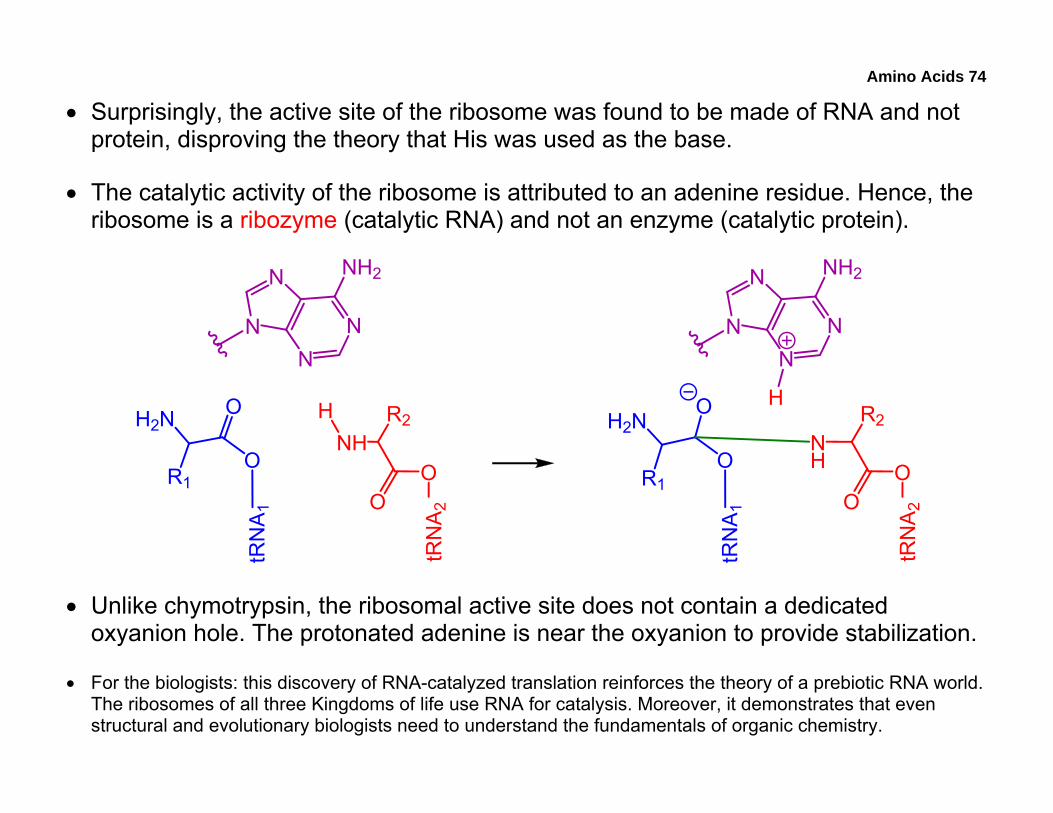
• Surprisingly, the active site of the ribosome was found to be made of RNA and not protein, disproving the theory that His was used as the base.
• The catalytic activity of the ribosome is attributed to an adenine residue. Hence, the ribosome is a ribozyme (catalytic RNA) and not an enzyme (catalytic protein).
H2N
O
O
R1
tRN
A1
NHO
O
R2tR
NA
2
H H2N
O
O
R1
tRN
A1
NH O
O
R2
tRN
A2
NN
N
N NH2
H
NN
N
N NH2
• Unlike chymotrypsin, the ribosomal active site does not contain a dedicated oxyanion hole. The protonated adenine is near the oxyanion to provide stabilization.
• For the biologists: this discovery of RNA-catalyzed translation reinforces the theory of a prebiotic RNA world. The ribosomes of all three Kingdoms of life use RNA for catalysis. Moreover, it demonstrates that even structural and evolutionary biologists need to understand the fundamentals of organic chemistry.

Amino Acids 75
• Up to now, some of the concepts we’ve covered include:
o Function of mRNA
o Basic structure of tRNA
o Mechanism of aminoacyl-tRNA synthetases
o Structure and composition of ribosomes
o Peptidyl transferase reaction
o Direction of peptide biosynthesis (compare to lab synthesis)
o Overview of x-ray crystallography
• Using amino-acid structures provided in the lab manual, attempt:
o 2008 Midterm: 22
• If you are really interested in the biology and/or the chemistry of ribosomes, check out the references provided on page 73.




![Protein Crystallography - instruct.uwo.ca · Protein Crystallization • Principles of protein solubility [PPt] [protein] Undersaturated solubility Supersaturated Precipitation Nucleation](https://img.pdfslide.us/doc/110x75/5e18b58cfac19c6065246f42/protein-crystallography-protein-crystallization-a-principles-of-protein-solubility.jpg)






![[PPT]Lecture 1 Thursday Jan. 4, 2001 - University of Western …instruct.uwo.ca/biology/3404f/Lecture18.ppt · Web viewEVOLUTION OF PLANTS Fall 2008 Lecture 18 Tuesday November 25,](https://img.pdfslide.us/doc/110x75/5ae9e4287f8b9ad73f8c6d42/pptlecture-1-thursday-jan-4-2001-university-of-western-viewevolution-of.jpg)
















