Embed Size (px)
Citation preview

vii
Contents
Preface. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . vContributors. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . xi
1 Three Decades of Studies to Understand the Functions of the Ubiquitin Family . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1Alexander Varshavsky
Part I Enzymology and SubStratES of ubIquItIn famIly modIfIErS
2 Structure and Evolution of Ubiquitin and Ubiquitin-Related Domains . . . . . . . . . . 15A. Maxwell Burroughs, Lakshminarayan M. Iyer, and L. Aravind
3 Deciphering Tissue-Specific Ubiquitylation by Mass Spectrometry . . . . . . . . . . . . . 65Ugo Mayor and Junmin Peng
4 Analysis of Cellular SUMO and SUMO–Ubiquitin Hybrid Conjugates . . . . . . . . . . 81Marion Schnellhardt, Kristina Uzunova, Veronika N. Bade, Anke Krause, Stefan R. Weisshaar, Gerrit J.K. Praefcke, and R. Jürgen Dohmen
5 Recombinant Reconstitution of Sumoylation Reactions In Vitro . . . . . . . . . . . . . . . 93Annette Flotho, Andreas Werner, Tobias Winter, Andrea S. Frank, Heidi Ehret, and Frauke Melchior
6 Reconstitution of SUMO-Dependent Ubiquitylation In Vitro . . . . . . . . . . . . . . . . 111Kirstin Keusekotten and Gerrit J.K. Praefcke
7 Detection and Analysis of FAT10 Modification . . . . . . . . . . . . . . . . . . . . . . . . . . . . 125Annette Aichem and Marcus Groettrup
8 Isolation of NEDDylated Proteins in Human Cells . . . . . . . . . . . . . . . . . . . . . . . . . 133Orsolya Leidecker and Dimitris P. Xirodimas
9 The ISG15 Conjugation System . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 141Larissa A. Durfee and Jon M. Huibregtse
10 Pupylation: Proteasomal Targeting by a Protein Modifier in Bacteria. . . . . . . . . . . . 151Kristin E. Burns and K. Heran Darwin
Part II rEcognItIon and chaIn formatIon of ubf modIfIErS
11 Role of UbL Family Modifiers and Their Binding Proteins in Cell Signaling . . . . . . 163Sjoerd J. L. van Wijk, Magda Bienko, and Ivan Dikic
12 Isolation of Ubiquitylated Proteins Using Tandem Ubiquitin-Binding Entities . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 173Fabienne Aillet, Fernando Lopitz-Otsoa, Roland Hjerpe, Mónica Torres-Ramos, Valérie Lang, and Manuel S. Rodríguez
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126

viii Contents
13 Using Linkage-Specific Monoclonal Antibodies to Analyze Cellular Ubiquitination. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 185Kim Newton, Marissa L. Matsumoto, Ronald E. Ferrando, Katherine E. Wickliffe, Michael Rape, Robert F. Kelley, and Vishva M. Dixit
14 Measuring Rates of Ubiquitin Chain Formation as a Functional Readout of Ligase Activity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 197Virginia P. Ronchi and Arthur L. Haas
15 Synthesis and Analysis of K11-Linked Ubiquitin Chains . . . . . . . . . . . . . . . . . . . . . 219Anja Bremm and David Komander
16 Synthesis and Analysis of Linear Ubiquitin Chains. . . . . . . . . . . . . . . . . . . . . . . . . . 229Kazuhiro Iwai
17 Detection and Quantitation of SUMO Chains by Mass Spectrometry . . . . . . . . . . . 239Ivan Matic and Ronald T. Hay
18 Bioinformatical Detection of Recognition Factors for Ubiquitin and SUMO. . . . . . 249Benjamin Vogt and Kay Hofmann
19 Surface Plasmon Resonance to Measure Interactions of UbFs with Their Binding Partners . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 263Julian Stingele, Uwe W. Roder, and Shahri Raasi
20 Identifying and Studying Ubiquitin Receptors by NMR . . . . . . . . . . . . . . . . . . . . . 279Xiang Chen and Kylie J. Walters
21 Exploring the Role of p97 and Its UBX-Domain Cofactors Through Identification of Their Interacting Proteins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 305Gabriela Alexandru
Part III ProtEaSomE bIogEnESIS, rEgulatIon and functIon
22 Assembly and Function of the Proteasome . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 315Yasushi Saeki and Keiji Tanaka
23 Using Native Gel Electrophoresis and Phosphofluoroimaging to Analyze GFP-Tagged Proteasomes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 339Cordula Enenkel
24 Disulfide Engineering to Map Subunit Interactions in the Proteasome and Other Macromolecular Complexes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 349Mark Hochstrasser and Minoru Funakoshi
25 Using DNA Damage Sensitivity Phenotypes to Characterize Mutations Affecting Proteasome Function. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 363Benoît Le Tallec and Anne Peyroche
26 Analysing Properties of Proteasome Inhibitors Using Kinetic and X-Ray Crystallographic Studies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 373Nerea Gallastegui and Michael Groll
27 Immunoproteasome-Specific Inhibitors and Their Application . . . . . . . . . . . . . . . . 391Michael Basler and Marcus Groettrup
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167

ixContents
28 Binding of Ubiquitin Conjugates to Proteasomes as Visualized with Native Gels . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 403Suzanne Elsasser, Yuan Shi, and Daniel Finley
29 Affinity Purification of Mammalian 26S Proteasomes Using an Ubiquitin-Like Domain . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 423Henrike C. Besche and Alfred L. Goldberg
30 Using siRNA Techniques to Dissect Proteasome Assembly Pathways in Mammalian Cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 433Takeumi Kaneko and Shigeo Murata
31 Reconstitution of PA700, the 19S Regulatory Particle, from Purified Precursor Complexes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 443George N. DeMartino
Part IV ProtEIn qualIty control
32 Cellular Responses to Misfolded Proteins and Protein Aggregates . . . . . . . . . . . . . . 455Scott A. Houck, Sangita Singh, and Douglas M. Cyr
33 Live-Cell Imaging of Ubiquitin–Proteasome System Function. . . . . . . . . . . . . . . . . 463Mark S. Hipp, Kirill Bersuker, and Ron R. Kopito
34 Analysis of Chaperone-Assisted Ubiquitylation . . . . . . . . . . . . . . . . . . . . . . . . . . . . 473Michael Dreiseidler, Niko Dick, and Jörg Höhfeld
35 Use of CPY* and Its Derivatives to Study Protein Quality Control in Various Cell Compartments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 489Alexandra Stolz and Dieter H. Wolf
36 Assays to Measure ER-Associated Degradation in Yeast . . . . . . . . . . . . . . . . . . . . . . 505Joseph R. Tran and Jeffrey L. Brodsky
37 SDS-PAGE Techniques to Study Ubiquitin-Like Conjugation Systems in Yeast Autophagy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 519Hitoshi Nakatogawa and Yoshinori Ohsumi
38 Analysis of Ubiquitin-Dependent Proteolysis in Caenorhabditis elegans . . . . . . . . . . 531Alexandra Segref and Thorsten Hoppe
Part V cEllular and chEmIcal StratEgIES to Study and manIPulatE ubf rElatEd ProcESSES
39 Structural Insights into Functional Modes of Proteins Involved in Ubiquitin Family Pathways . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 547Petra Hänzelmann, Antje Schäfer, Daniel Völler, and Hermann Schindelin
40 Identification and Application of NEDD8 E1 Inhibitors . . . . . . . . . . . . . . . . . . . . . 577Frank J. Bruzzese, Michael A. Milhollen, James M. Gavin, Helen R. Josephine, and James E. Brownell
41 Formation of Ubiquitin Dimers via Azide–Alkyne Click Reaction . . . . . . . . . . . . . . 589Silvia Eger, Martin Scheffner, Andreas Marx, and Marina Rubini
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207

x Contents
42 Synthesis of Atypical Diubiquitin Chains . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 597Farid El Oualid, Dharjath S. Hameed, Dris El Atmioui, Henk Hilkmann, and Huib Ovaa
43 TIPI: TEV Protease-Mediated Induction of Protein Instability . . . . . . . . . . . . . . . . 611Christof Taxis and Michael Knop
44 PROTAC-Induced Proteolytic Targeting . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 627Kimberly Cornish Carmony and Kyung-Bo Kim
45 Formation of Nondegradable Forked Ubiquitin Conjugates by Ring-Finger Ligases and Its Prevention by S5a . . . . . . . . . . . . . . . . . . . . . . . . . . 639Hyoung Tae Kim and Alfred L. Goldberg
46 S5a/Rpn10, a UIM-Protein, as a Universal Substrate for Ubiquitination . . . . . . . . 653Hyoung Tae Kim and Alfred L. Goldberg
Index . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 661
208
209
210
211
212
213
214
215
216
217
218
219
220

xi
Contributors
annEttE aIchEm • Biotechnology Institute Thurgau, Kreuzlingen, Switzerland; Department of Molecular Biophysics and Biochemistry, Yale University, New Haven, CT, USA
fabIEnnE aIllEt • Proteomics Unit and Proteomics Platform, CIC bioGUNE CIBERehd, ProteoRed Technology Park of Bizkaia, Derio, Spain
gabrIEla alExandru • Protein Ubiquitylation Unit, The Scottish Institute for Cell Signalling (SCILLS), Dundee, Scotland, UK; University of Dundee, Dundee, Scotland, UK
l. araVInd • National Center for Biotechnology Information, National Library of Medicine, National Institutes of Health, Bethesda, MD, USA
drISElatmIouI • Division of Cell Biology, Netherlands Cancer Institute, Amsterdam, The Netherlands
VEronIka badE • Center for Molecular Medicine Cologne (CMMC), Institute for Genetics, University of Cologne, Cologne, Germany
mIchaEl baSlEr • Department of Biology, Division of Immunology, University of Konstanz, Konstanz, GermanyM. Basler Biotechnology Institute Thurgau, Kreuzlingen, Switzerland
kIrIll bErSukEr • Department of Biology, Stanford University, Stanford, CA, USAhEnrIkE c. bESchE • Department of Cell Biology, Harvard Medical School,
Boston, MA, USAmagda bIEnko • Frankfurt Institute for Molecular Life Sciences and Institute
of Biochemistry II, Goethe University School of Medicine, Frankfurt (Main), Germany; Departments of Physics and of Biology, Massachusetts Institute of Technology, Cambridge, MA, USA
anja brEmm • Protein and Nucleic Acid Chemistry Division, MRC Laboratory of Molecular Biology, Cambridge, UK
jEffrEy l. brodSky • Department of Biological Sciences, University of Pittsburgh, Pittsburgh, PA, USA
jamES E. brownEll • Discovery, Millennium Pharmaceuticals Inc., Cambridge, MA, USA
frank j. bruzzESE • Discovery, Millennium Pharmaceuticals Inc., Cambridge, MA, USA
krIStIn E. burnS • Department of Microbiology, New York University School of Medicine, New York, NY, USA
a. maxwEll burroughS • Omics Science Center (OSC), RIKEN Yokohama Institute, Kanagawa, Japan
kImbErly cornISh carmony • Department of Pharmaceutical Science, College of Pharmacy, University of Kentucky, Lexington, KY, USA
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259

xii Contributors
xIang chEn • Department of Biochemistry, Molecular Biology and Biophysics, University of Minnesota, Minneapolis, MN, USA
douglaS m. cyr • Department of Cell and Developmental Biology, University of North Carolina at Chapel Hill, Chapel Hill, NC, USA
k. hEran darwIn • Department of Microbiology, New York University School of Medicine, New York, NY, USA
gEorgE n. dEmartIno • Department of Physiology, University of Texas Southwestern Medical Center, Dallas, TX, USA
nIko dIck • Institut für Zellbiologie, Rheinische Friedrich-Wilhelms-Universität Bonn, Bonn, Germany
IVan dIkIc • Frankfurt Institute for Molecular Life Sciences and Institute of Biochemistry II, Goethe University School of Medicine, Frankfurt (Main), Germany
VIShVa m. dIxIt • Physiological Chemistry Department, Genentech, Inc., South San Francisco, CA, USA
jürgEn dohmEn • Institute for Genetics, University of Cologne, Cologne, GermanymIchaEl drEISEIdlEr • Institut für Zellbiologie, Rheinische Friedrich-Wilhelms-
Universität Bonn, Bonn, GermanylarISSa a. durfEE • Section of Molecular Genetics and Molecular Biology,
Institute for Cellular and Molecular Biology, University of Texas at Austin, Austin, TX, USA
SIlVIa EgEr • Department of Chemistry, Konstanz Research School Chemical Biology, University of Konstanz, Konstanz, Germany
hEIdI EhrEt • Zentrum für Molekulare Biologie der Universität Heidelberg (ZMBH), DKFZ-ZMBH Alliance, Heidelberg, Germany
SuzannE ElSaSSEr • Department of Cell Biology, Harvard Medical School, Boston, MA, USA
cordula EnEnkEl • Department of Biochemistry, University of Toronto, Toronto, ON, Canada
ronald E. fErrando • Pathology Department, Genentech, Inc., South San Francisco, CA, USA
danIEl fInlEy • Department of Cell Biology, Harvard Medical School, Boston, MA, USA
annEttE flotho • Zentrum für Molekulare Biologie der Universität Heidelberg (ZMBH), DKFZ-ZMBH Alliance, Heidelberg, Germany
andrEa S. frank • Zentrum für Molekulare Biologie der Universität Heidelberg (ZMBH), DKFZ-ZMBH Alliance, Heidelberg, Germany
mInoru funakoShI • Department of Molecular Biophysics and Biochemistry, Yale University, New Haven, CT, USA; Graduate School of Natural Science and Technology, Okayama University, Okayama, Japan
nErEa gallaStEguI • Department Chemie, Technische Universität München, Garching, Germany
jamES m. gaVIn • Discovery, Millennium Pharmaceuticals Inc., Cambridge, MA, USAalfrEd l. goldbErg • Department of Cell Biology, Harvard Medical School,
Boston, MA, USA
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304

xiiiContributors
marcuS groEttruP • Department of Biology, Division of Immunology, University of Konstanz, Konstanz, Germany; Biotechnology Institute Thurgau, Kreuzlingen, Switzerland
mIchaEl groll • Department Chemie, Technische Universität München, Garching, Germany
arthur l. haaS • Department of Biochemistry and Molecular Biology, Louisiana State University School of Medicine and the Stanley S. Scott Cancer Center, New Orleans, LA, USA
dharjath S. hamEEd • Division of Cell Biology, Netherlands Cancer Institute, Amsterdam, The Netherlands
PEtra hänzElmann • Rudolf Virchow Center for Experimental Biomedicine, University of Würzburg, Würzburg, Germany
ronald t. hay • Wellcome Trust Centre for Gene Regulation and Expression, Sir James Black Centre, College of Life Sciences, University of Dundee, Dundee, Scotland, UK
hEnk hIlkmann • Division of Cell Biology, Netherlands Cancer Institute, Amsterdam, The Netherlands
mark S. hIPP • Department of Biology, Stanford University, Stanford, CA, USA; Department of Cellular Biochemistry, Max Planck Institute of Biochemistry, Martinsried, Germany
roland hjErPE • Proteomics Unit and Proteomics Platform, CIC bioGUNE CIBERehd, ProteoRed, Technology Park of Bizkaia, Derio, Spain
mark hochStraSSEr • Department of Molecular Biophysics and Biochemistry, Yale University, New Haven, CT, USA
kay hofmann • Bioinformatics Group, Miltenyi Biotec GmbH, Bergisch Gladbach, Germany
jörg höhfEld • Institut für Zellbiologie, Rheinische Friedrich-Wilhelms- Universität Bonn, Bonn, Germany
thorStEn hoPPE • Institute for Genetics Cologne Excellence Cluster on Cellular Stress Responses in Aging-Associated Diseases (CECAD), University of Cologne,
Scott a. houck • Department of Cell and Developmental Biology, University of North Carolina at Chapel Hill, Chapel Hill, NC, USA
jon m. huIbrEgtSE • Section of Molecular Genetics and Molecular Biology, Institute for Cellular and Molecular Biology, University of Texas at Austin, Austin, TX, USA
kazuhIro IwaI • Cell Biology and Metabolism Group, Graduate School of Frontier Biosciences, Osaka University, Suita, Japan; Department of Biophysics and Biochemistry, Graduate School of Medicine, Osaka University, Suita, Japan
lakShmInarayan m. IyEr • National Center for Biotechnology Information, National Library of Medicine, National Institutes of Health, Bethesda, MD, USA
hElEn r. joSEPhInE • Discovery, Millennium Pharmaceuticals Inc., Cambridge, MA, USA
takEumI kanEko • Laboratory of Protein Metabolism, Graduate School of Pharmaceutical Sciences, The University of Tokyo, Tokyo, Japan
robErt f. kEllEy • Antibody Engineering Department, Genentech, Inc., South San Francisco, CA, USA
305
306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350

xiv Contributors
kIrStIn kEuSEkottEn • Center for Molecular Medicine Cologne (CMMC), Institute for Genetics, University of Cologne, Cologne, Germany
kyung-bo kIm • Department of Pharmaceutical Science, College of Pharmacy, University of Kentucky, Lexington, KY, USA
mIchaEl knoP • Zentrum für Molekulare Biologie der Universität Heidelberg (ZMBH), ZMBH-DKFZ Alliance, Im Neuenheimer Feld 282, 69120, Heidelberg, Germany
daVId komandEr • Protein and Nucleic Acid Chemistry Division, MRC Laboratory of Molecular Biology, Cambridge, UK
ron r. koPIto • Department of Biology, Stanford University, Stanford, CA, USAankE krauSE • Center for Molecular Medicine Cologne (CMMC),
Institute for Genetics, University of Cologne, Cologne, GermanyValérIE lang • Proteomics Unit and Proteomics Platform, CIC bioGUNE CIBERehd,
ProteoRed, Technology Park of Bizkaia, Derio, SpainorSolya lEIdEckEr • Wellcome Trust Centre for Gene Regulation and Expression,
College of Life Sciences, University of Dundee, Dundee, Scotland, UKfErnando loPItz-otSoa • Proteomics Unit and Proteomics Platform,
CIC bioGUNE CIBERehd, ProteoRed, Technology Park of Bizkaia, Derio, SpainandrEaS marx • Department of Chemistry, Konstanz Research School Chemical
Biology, University of Konstanz, Konstanz, GermanyIVan matIc • Wellcome Trust Centre for Gene Regulation and Expression,
Sir James Black Centre, College of Life Sciences, University of Dundee, Dundee, Scotland, UK
marISSa l. matSumoto • Antibody Engineering Department, Genentech, Inc., South San Francisco, CA, USA
ugo mayor • Proteomics Unit and Proteomics Platform, CIC bioGUNE CIBERehd, ProteoRed, Technology Park of Bizkaia, Derio, Spain
fraukE mElchIor • Zentrum für Molekulare Biologie der Universität Heidelberg (ZMBH), DKFZ-ZMBH Alliance, Heidelberg, Germany
mIchaEl a. mIlhollEn • Discovery, Millennium Pharmaceuticals Inc., Cambridge, MA, USA
ShIgEo murata • Laboratory of Protein Metabolism, Graduate School of Pharmaceutical Sciences, The University of Tokyo, Tokyo, Japan
hItoShI nakatogawa • Frontier Research Center, Tokyo Institute of Technology, Yokohama, Japan
kIm nEwton • Physiological Chemistry Department, Genentech, Inc., South San Francisco, CA, USA
yoShInorI ohSumI • Frontier Research Center, Tokyo Institute of Technology, Yokohama, Japan
farIdEloualId • Division of Cell Biology, Netherlands Cancer Institute, Amsterdam, The Netherlands
huIb oVaa • Division of Cell Biology, Netherlands Cancer Institute, Amsterdam, The Netherlands
junmIn PEng • Department of Structural Biology, St. Jude Proteomics Facility, St. Jude Children’s Research Hospital, Memphis, TN, USA
351
352
353
354
355
356
357
358
359
360
361
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395

xvContributors
annE PEyrochE • CEA, iBiTecS, SBIGeM, Laboratoire du métabolisme de l’ADN et réponses aux génotoxiques, Gif-sur-Yvette, France
gErrIt j. k. PraEfckE • Center for Molecular Medicine Cologne (CMMC), Institute for Genetics, University of Cologne, Cologne, Germany
ShahrI raaSI • Laboratory of Cellular Biochemistry, Department of Biology, University of Konstanz, Konstanz, Germany
mIchaEl raPE • Department of Molecular and Cell Biology, University of California, Berkeley, CA, USA
uwE w. rodEr • GE Healthcare Europe GmbH, Freiburg, GermanymanuEl S. rodríguEz • Proteomics Unit and Proteomics Platform, CIC bioGUNE
CIBERehd, ProteoRed, Technology Park of Bizkaia, Derio, Spain; Biochemistry Department, University of the Basque Country, Leioa, Spain
VIrgInIa P. ronchI • Department of Biochemistry and Molecular Biology, Louisiana State University School of Medicine and the Stanley S. Scott Cancer Center, New Orleans, LA, USA
marIna rubInI • Department of Chemistry, Konstanz Research School Chemical Biology, University of Konstanz, Konstanz, Germany
yaSuShI SaEkI • Laboratory of Protein Metabolism, Tokyo Metropolitan Institute of Medical Science, Tokyo, Japan
antjE SchäfEr • Rudolf Virchow Center for Experimental Biomedicine, University of Würzburg, Würzburg, Germany
martIn SchEffnEr • Department of Biology, Konstanz Research School Chemical Biology, University of Konstanz, Konstanz, Germany
hErmann SchIndElIn • Rudolf Virchow Center for Experimental Biomedicine, University of Würzburg, Würzburg, Germany
marIon SchnEllhardt • Institute for Genetics, University of Cologne, Cologne, Germany
alExandra SEgrEf • Institute for Genetics and Cologne Excellence Cluster on Cellu-lar Stress Responses in Aging-Associated Diseases (CECAD), University of Cologne, Cologne, Germany
yuan ShI • Department of Cell Biology, Harvard Medical School, Boston, MA, USASangIta SIngh • Department of Cell and Developmental Biology, University of North
Carolina at Chapel Hill, Chapel Hill, NC, USAjulIan StIngElE • Department of Molecular Cell Biology, Max-Planck Institute
of Biochemistry, Martinsried, GermanyalExandra Stolz • Institute of Biochemistry, University Stuttgart,
Stuttgart, GermanybEnoît lE tallEc • CEA, iBiTecS, SBIGeM, Laboratoire du métabolisme
de l’ADN et réponses aux génotoxiques, Gif-sur-Yvette, France; Institut Curie, Centre de Recherche, Paris, France
kEIjI tanaka • Laboratory of Protein Metabolism, Tokyo Metropolitan Institute of Medical Science, Tokyo, Japan
chrIStof taxIS • Philipps Universität Marburg, Fakultät Biologie – Genetik, Marburg, Germany
mónIca torrES-ramoS • Proteomics Unit and Proteomics Platform, CIC bioGUNE CIBERehd, ProteoRed, Technology Park of Bizkaia, Derio, Spain
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
424
425
426
427
428
429
430
431
432
433
434
435
436
437
438
439
440
441

xvi Contributors
joSEPh r. tran • Department of Biological Sciences, University of Pittsburgh, Pittsburgh, PA, USA; Graduate Program in Biochemistry and Molecular Genetics, University of Pittsburgh School of Medicine, Pittsburgh, PA, USA
krIStIna uzunoVa • Institute for Genetics, University of Cologne, Cologne, Germany; IMP – Research Institute of Molecular Pathology, Vienna, Austria
alExandEr VarShaVSky • Division of Biology, California Institute of Technology, Pasadena, CA, USA
bEnjamIn Vogt • Institute for Genetics, University of Cologne, Cologne, Germany; Bioinformatics Group, Miltenyi Biotec GmbH, Bergisch Gladbach, Germany
danIEl VöllEr • Rudolf Virchow Center for Experimental Biomedicine, University of Würzburg, Würzburg, Germany
kylIE j. waltErS • Department of Biochemistry, Molecular Biology and Biophysics, University of Minnesota, Minneapolis, MN, USA
StEfan r. wEISShaar • Center for Molecular Medicine Cologne (CMMC), Institute for Genetics, University of Cologne, Cologne, Germany
andrEaS wErnEr • Zentrum für Molekulare Biologie der Universität Heidelberg (ZMBH), DKFZ-ZMBH Alliance, Heidelberg, Germany
kathErInE E. wIcklIffE • Department of Molecular and Cell Biology, University of California, Berkeley, CA, USA
SjoErd j. l. Van wIjk • Frankfurt Institute for Molecular Life Sciences and Institute of Biochemistry II, Goethe University School of Medicine, Frankfurt (Main), Germany
tobIaS wIntEr • Zentrum für Molekulare Biologie der Universität Heidelberg (ZMBH), DKFZ-ZMBH Alliance, Heidelberg, Germany
dIEtEr h. wolf • Institute of Biochemistry, University Stuttgart, Stuttgart, GermanydImItrIS P. xIrodImaS • Wellcome Trust Centre for Gene Regulation and Expression,
College of Life Sciences, University of Dundee, Dundee, Scotland, UK; Centre de Recherche de Biochimie Macromoléculaire – UMR 5237, CNRS, Montpellier, France
442
443
444
445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469

219
R. Jürgen Dohmen and Martin Scheffner (eds.), Ubiquitin Family Modifiers and the Proteasome: Reviews and Protocols, Methods in Molecular Biology, vol. 832, DOI 10.1007/978-1-61779-474-2_15, © Springer Science+Business Media, LLC 2012
Chapter 15
Synthesis and Analysis of K11-Linked Ubiquitin Chains
Anja Bremm and David Komander
Abstract
Much has been learned about protein ubiquitination by studying the structural, biochemical, and bio-physical properties of ubiquitin chains in vitro. However, these analyses were limited to K48-, K63-linked, and linear ubiquitin chains. Only recently, enzymatic and chemical assembly systems for the remaining chain types have been developed. Here, we describe a method to synthesise K11-linked ubiquitin chains in vitro by exploiting the intrinsic K11-specificity of the E2 enzyme UBE2S.
Key words: Ubiquitin, K11 linkage, Atypical ubiquitin chains, UBE2S
Ubiquitin is attached to substrates via an isopeptide linkage between its C terminus and the e-amino group of a lysine residue. Additionally, ubiquitin can be ligated to one of the seven lysine residues or the N-terminal amino group of another ubiquitin molecule, thereby forming polymers of eight different linkage types. It has been demonstrated that all polyubiquitin chains are present in vivo (1). The best-characterised chain types are K48-linked and K63-linked ubiquitin polymers, which have degradative and nondegradative functions, respectively (2, 3). Enzymatic assembly systems have enabled access to large quantities of these polyubiquitin chains. The remaining “atypical” ubiquitin chains have not been studied in great detail and this is partly due to the lack of in vitro assembly systems.
Protein ubiquitination is catalysed by a three-step enzymatic cascade, including an E1 ubiquitin-activating enzyme, an E2 ubiquitin-conjugating enzyme, and E3 ligases (4). Whereas E3s determine substrate specificity, E2s are able to mediate the switch from ubiquitin chain initiation to elongation, regulate the processivity of chain
1. Introduction
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29

220 A. Bremm and D. Komander
assembly and define the type of chain linkage (5). E2s specific for K48 or K63 linkages have been identified and are used for the generation of large amounts of these chain types in vitro.
K11-linked ubiquitin chains have been implicated in important biological processes, including cell cycle regulation (6), endoplasmic reticulum-associated degradation (ERAD) (1), and TNFa signalling (7). The RING-E3 anaphase-promoting complex (APC/C) uses the E2 enzymes UBE2C and UBE2S to promote the degradation of mitotic regulators by assembling K11-linked ubiquitin chains (6, 8, 9).
UBE2S also specifically generates free K11-linked diubiquitin in vitro in the absence of an E3 ligase (10). For structural and biophysical studies, the amount of unattached dimers obtained in a chain assembly reaction using wild-type (wt) UBE2S was not sufficient. Therefore, we engineered a UBE2S variant with increased efficiency. We found that wt UBE2S performs autoubiquitination in a cis reaction on its lysine rich C-terminal tail (10). Hence, we replaced the C terminus of UBE2S with a ubiquitin-binding domain (UBD), namely the zinc finger (ZnF) UBP domain of USP5/IsoT (11). This increases the local concentration of free ubiquitin near the active site and enhances formation of free K11-linked chains. The advantage of the ZnF UBP domain is that it binds ubiquitin with nanomolar affinity and that it interacts with the free C termi-nus of ubiquitin, leaving lysine 11, and the TEK box (6) accessible for chain elongation. This engineered UBE2S-UBD fusion protein generates free K11-linked ubiquitin dimers more efficiently than the wt enzyme and also allows the purification of trimers and tetramers in vitro. However, UBE2S-UBD is less specific and incor-porates also K63-linkages. Formation of contaminating K63-linkages can be counteracted by including the K63-specific deubiquitinase (DUB) AMSH in the assembly reactions. This method allows specific generation and purification of K11-linked ubiquitin chains. Taken together, in addition to UBE2R1 (Cdc34) and UBE2N/UBE2V1 (Ubc13/UEV-1), which specifically assemble K48-linked and K63-linked ubiquitin chains (5), respectively, an enzymatic system is now available for the generation of K11-linked ubiquitin chains in vitro.
Prepare all buffers using prechilled ultrapure water, for example Milli-Q water.
1. Plasmids: pGEX6P1-UBE2S to generate pGEX6P1-UBE2S-UBD (see Subheading 3.1), pGEX6P1-AMSH, and pRSETB-IsoT (12).
2. Materials
2.1. Affinity Purification of GST-Fusion Proteins
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71

22115 Synthesis and Analysis of K11-Linked Ubiquitin Chains
2. Competent cells for plasmid DNA preparation (for example, OneShot® Mach1™-T1® cells, Invitrogen) and protein expression (Rosetta™ 2(DE3)pLacI cells, Novagen).
3. LB medium and selective plates containing 100 mg/ml ampicillin.
4. 1 M IPTG stock solution. Dissolve IPTG in H2O, sterile filter, and store aliquots at −20°C.
5. Lysis buffer: 270 mM sucrose, 50 mM Tris–HCl (pH 7.5), 50 mM NaF. Sterile filter and store at 4°C. Before usage add 0.1% v/v b-mercaptoethanol, protease inhibitor cocktail (for example, Complete, Mini, EDTA-free tablets, Roche), 1 mg/ml lysozyme and 0.1 mg/ml DNase I.
6. Glass chromatography columns (Bio-Rad). 7. Glutathione Sepharose™ 4B (GE Healthcare). Store at 4°C. 8. High-salt buffer: 500 mM NaCl, 25 mM Tris–HCl (pH 7.5),
5 mM DTT (see Note 1). 9. Low-salt buffer: 150 mM NaCl, 25 mM Tris–HCl (pH 7.5),
5 mM DTT. 10. PreScission™ protease (GE Healthcare). Store at −20°C (see
Note 2).
1. Ubiquitin storage buffer: 50 mM Tris–HCl (pH 7.5). 2. Ubiquitin from bovine red blood cells (Sigma-Aldrich). Dissolve
ubiquitin in ubiquitin storage buffer and store at −20°C. 3. Recombinant E1 enzyme can be purchased from Ubiquigent,
ENZO Life Sciences or Boston Biochem (see Note 3). 4. 0.1 M ATP stock solution: Dissolve ATP in H2O and adjust
pH to 7.0 with NaOH. Store at −20°C. 5. 10× ligation buffer: 400 mM Tris–HCl (pH 7.5), 100 mM
MgCl2, 6 mM DTT. Store at −20°C. 6. Recombinant UBE2S-UBD and AMSH (see Subheading 3.2). 7. 1 M DTT stock solution. Dissolve DTT in H2O and store
small aliquots at −20°C.
1. Fast protein liquid chromatography (FPLC) system: For exam-ple, the ÄKTApurifier™ system including a Mono S™ 5/50 cation exchange column and a 50-ml superloop (GE Healthcare).
2. Buffer A: 50 mM NH4Ac (pH 4.5). 3. Buffer B: 50 mM NH4Ac (pH 4.5), 1 M NaCl. Adjust pH of
buffer A and B with HCl.
2.2. Large-Scale K11-Linked Ubiquitin Chain Assembly Reaction
2.3. Fast Protein Liquid Chromatography
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110

222 A. Bremm and D. Komander
Human UBE2S (NM_014501.2) was cloned into the BamHI/NotI restriction sites present in pGEX6P1. The UBE2S sequence has a naturally occurring unique NcoI restriction site just before the lysine-rich tail (residue 196).
1. Digest the pGEX6P1-UBE2S expression plasmid with NcoI and NotI (restriction enzymes for example from New England Biolabs Inc.). Incubate at 37°C for 2 h and purify cleaved plasmid DNA by gel extraction. This releases the lysine-rich C-terminal tail of UBE2S.
2. Amplify the USP5/IsoT ZnF UBP domain (residues 173–289) from cDNA or from pRSETB-IsoT by PCR. Use primers that generate a 5¢ NcoI site and a 3¢ NotI site in the PCR product so that it can be ligated into the NcoI/NotI restriction sites present in pGEX6P1-UBE2S (see Note 4). Purify the PCR product by gel extraction. Elute DNA in 30 ml H2O.
3. Digest the purified PCR product with NcoI and NotI. Incubate at 37°C for 2 h and purify the cleaved DNA by gel extraction.
4. Ligate the cleaved PCR product into linearized pGEX6P1-UBE2S by using for example a Rapid DNA Ligation kit (Roche).
5. Use the ligation reaction mixture for the transformation of competent Mach1 cells. Spread cells on selective ampicillin plates and grow overnight at 37°C.
6. Pick single bacterial colonies and grow cells in 2 ml LB/ampi-cillin medium.
7. Purify plasmid DNA (for example, QIAprep Spin Miniprep Kit, QIAGEN) and verify clones by sequencing.
See Note 5 for additional information.
1. Transform Rosetta cells (see Note 6) with pGEX6P1-UBE2S-UBD or pGEX6P1-AMSH.
2. For each construct, pick a single colony from a selective ampicillin plate to inoculate 50 ml LB/ampicillin medium. Grow cultures at 37°C, 220 rpm, and overnight.
3. Inoculate 1 l LB medium in a 2-l flask with 10 ml overnight culture and grow cultures at 37°C, 220 rpm until OD600 of 0.6 is reached. Take a 5 ml aliquot of the noninduced culture (sample NI) and grow separately.
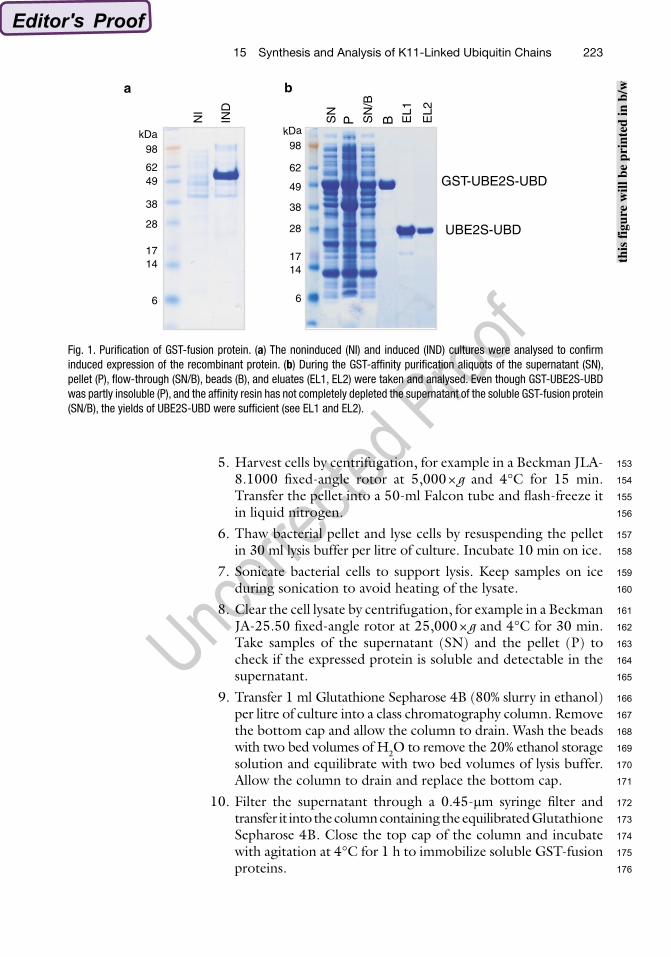
4. Induce expression of recombinant proteins with 250 mM IPTG and grow cultures at 20°C, 220 rpm, overnight. Take a 5 ml aliquot (IND) and analyse it together with 5 ml of sample NI by SDS-PAGE and Coomassie staining to check induced expression of the recombinant protein (see Fig. 1a).
3. Methods
3.1. Cloning of UBE2S-UBD Fusion Construct
3.2. Expression and Affinity Purification of GST-Fusion Proteins
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
22315 Synthesis and Analysis of K11-Linked Ubiquitin Chains
5. Harvest cells by centrifugation, for example in a Beckman JLA-8.1000 fixed-angle rotor at 5,000 × g and 4°C for 15 min. Transfer the pellet into a 50-ml Falcon tube and flash-freeze it in liquid nitrogen.
6. Thaw bacterial pellet and lyse cells by resuspending the pellet in 30 ml lysis buffer per litre of culture. Incubate 10 min on ice.
7. Sonicate bacterial cells to support lysis. Keep samples on ice during sonication to avoid heating of the lysate.
8. Clear the cell lysate by centrifugation, for example in a Beckman JA-25.50 fixed-angle rotor at 25,000 × g and 4°C for 30 min. Take samples of the supernatant (SN) and the pellet (P) to check if the expressed protein is soluble and detectable in the supernatant.
9. Transfer 1 ml Glutathione Sepharose 4B (80% slurry in ethanol) per litre of culture into a class chromatography column. Remove the bottom cap and allow the column to drain. Wash the beads with two bed volumes of H2O to remove the 20% ethanol storage solution and equilibrate with two bed volumes of lysis buffer. Allow the column to drain and replace the bottom cap.
10. Filter the supernatant through a 0.45-mm syringe filter and transfer it into the column containing the equilibrated Glutathione Sepharose 4B. Close the top cap of the column and incubate with agitation at 4°C for 1 h to immobilize soluble GST-fusion proteins.
1714
6
28
38
kDa98
6249 GST-UBE2S-UBD
UBE2S-UBD
NI
IND
SN
P SN
/B
B EL1
EL2
a b
1714
6
28
38
kDa98
62
49
Fig. 1. Purification of GST-fusion protein. (a) The noninduced (NI) and induced (IND) cultures were analysed to confirm induced expression of the recombinant protein. (b) During the GST-affinity purification aliquots of the supernatant (SN), pellet (P), flow-through (SN/B), beads (B), and eluates (EL1, EL2) were taken and analysed. Even though GST-UBE2S-UBD was partly insoluble (P), and the affinity resin has not completely depleted the supernatant of the soluble GST-fusion protein (SN/B), the yields of UBE2S-UBD were sufficient (see EL1 and EL2).
this
figu
re w
ill b
e pr
inte
d in
b/w
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176

224 A. Bremm and D. Komander
11. Remove the top and bottom cap of the column and elute unbound proteins. Take a sample of the flow-through (SN/B).
12. Wash the Glutathione Sepharose 4B with 1 l high-salt buffer and subsequently with 500 ml low-salt buffer to remove non-specifically bound proteins. Allow the column to drain and replace the bottom cap of the column. Take a sample of the beads (B). Samples SN/B and B show if the GST-fusion protein is bound to the Glutathione Sepharose 4B (see Note 7).
13. To cleave the GST tag of the immobilized proteins add 30 mM PreScission protease in 5 ml low-salt buffer to the column, close the top cap, and incubate with agitation at 4°C overnight.
14. Elute cleaved proteins into a fresh tube. This fraction contains most of the cleaved protein (~70%). Wash the beads with 5 ml low-salt buffer to elute the remaining proteins. Repeat this step until all cleaved protein is eluted. This can be monitored by Bradford protein assay. Take an aliquot of each fraction (EL1, EL2, etc.) and analyse them together with samples SN, P, SN/B, and B by SDS-PAGE and Coomassie staining (see Fig. 1b).
15. Pool protein-containing fractions and concentrate them (see Note 8) using for example Vivaspin 15 centrifugal concentrators (Sartorius Stedim Biotech) with a molecular weight cut-off (MWCO) of 10,000 Da (for UBE2S-UBD) or 30,000 Da (for AMSH). Flash-freeze proteins in liquid nitrogen and store them at −80°C (see Note 9).
1. Thaw all enzymes and solutions on ice. 2. Set up a 1 ml reaction: Mix 250 nM E1 enzyme (see Note 3),
4.8 mM UBE2S-UBD, 2.94 mM ubiquitin, 400 nM AMSH, 10 mM ATP, and 10× ligation buffer (400 mM Tris–HCl (pH 7.5), 100 mM MgCl2, 6 mM DTT), and incubate at 37°C for 3 h (see Table 1).
3. After 1 h add another 400 nM AMSH to the reaction to coun-teract the formation of K63-linked ubiquitin chains. Repeat this step after 2 h.
4. Stop the reaction after 3 h by adding 50 mM DTT. Incubate for 10 min on ice. This releases any ubiquitin (monomers or polymers) covalently bound to the active site cysteine of UBE2S-UBD or the E1 enzyme.
5. Subsequently, dilute the reaction with 14 ml buffer A (50 mM NH4Ac [pH 4.5]) and incubate for at least 30 min on ice to precipitate the E1 enzyme and UBE2S-UBD (see Note 10).
6. Filter the diluted assembly reaction through a 0.2-mm syringe filter to remove precipitated enzymes.
3.3. Large-Scale Assembly Reaction
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
22515 Synthesis and Analysis of K11-Linked Ubiquitin Chains
1. Load the filtrate into a 50-ml superloop. 2. Purify K11-linked ubiquitin chains by cation exchange chroma-
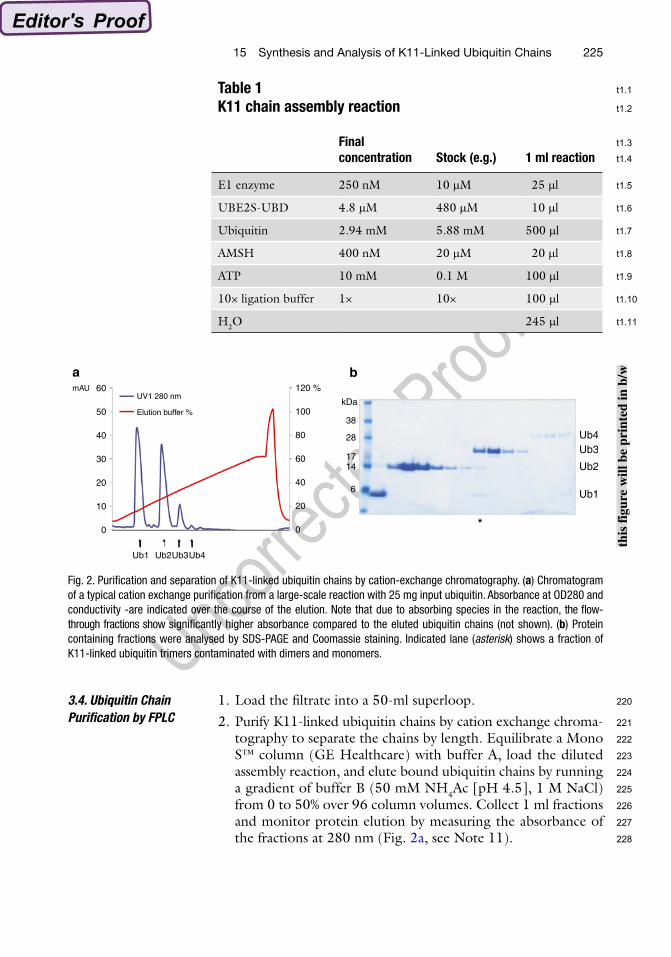
tography to separate the chains by length. Equilibrate a Mono S™ column (GE Healthcare) with buffer A, load the diluted assembly reaction, and elute bound ubiquitin chains by running a gradient of buffer B (50 mM NH4Ac [pH 4.5], 1 M NaCl) from 0 to 50% over 96 column volumes. Collect 1 ml fractions and monitor protein elution by measuring the absorbance of the fractions at 280 nm (Fig. 2a, see Note 11).
3.4. Ubiquitin Chain Purification by FPLC
Table 1K11 chain assembly reaction
Final concentration Stock (e.g.) 1 ml reaction
E1 enzyme 250 nM 10 mM 25 ml
UBE2S-UBD 4.8 mM 480 mM 10 ml
Ubiquitin 2.94 mM 5.88 mM 500 ml
AMSH 400 nM 20 mM 20 ml
ATP 10 mM 0.1 M 100 ml
10× ligation buffer 1× 10× 100 ml
H2O 245 ml
1714
6
28
38
kDa
120 %
Elution buffer %
UV1 280 nm
100
60
50
40
30
20
10
0
80
60
40
20
0
Ub4Ub3
Ub1
Ub2
Ub1
mAU
Ub3Ub2 Ub4
*
ba
Fig. 2. Purification and separation of K11-linked ubiquitin chains by cation-exchange chromatography. (a) Chromatogram of a typical cation exchange purification from a large-scale reaction with 25 mg input ubiquitin. Absorbance at OD280 and conductivity -are indicated over the course of the elution. Note that due to absorbing species in the reaction, the flow-through fractions show significantly higher absorbance compared to the eluted ubiquitin chains (not shown). (b) Protein containing fractions were analysed by SDS-PAGE and Coomassie staining. Indicated lane (asterisk) shows a fraction of K11-linked ubiquitin trimers contaminated with dimers and monomers.
this
figu
re w
ill b
e pr
inte
d in
b/w
220
221
222
223
224
225
226
227
228
t1.1
t1.2
t1.3
t1.4
t1.5
t1.6
t1.7
t1.8
t1.9
t1.10
t1.11

226 A. Bremm and D. Komander
3. Analyse 5 ml of the peak fractions by SDS-PAGE and Coomassie staining (Fig. 2b, see Notes 12 and 13).
4. Pool fractions by chain length and concentrate them to >5 mg/ml if possible (see Note 11). Use for example a Vivaspin 2 (MWCO: 5,000 Da) centrifugal concentrator (Sartorius Stedim Biotech). Either dialyse purified ubiquitin chains against ubiquitin storage buffer (50 mM Tris–HCl [pH 7.5]) at 4°C overnight, or exchange buffer while concentrating the samples (i.e., repeatedly dilute the concentrated sample with ubiquitin storage buffer and concentrate again, see Notes 14 and 15). Flash-freeze the ubiquitin chains in liquid nitrogen and store them at −20°C.
1. Prepare high- and low-salt buffer freshly each time and add DTT to protect free SH-groups from oxidation during the iso-lation of proteins. The pH was adjusted by taking the theo-retical isoelectrical point of AMSH and UBE2S-UBD into account.
2. PreScission™ protease (GE Healthcare) is used for low-temperature, site-specific separation of the GST tag from proteins expressed using pGEX6P vectors. Since PreScission™ protease contains a noncleavable GST-tag, it is retained on the Glutathione Sepharose 4B resin and only the cleaved protein elutes from the column.
3. We purified recombinant human E1 enzyme using a baculovirus expression system (13). The concentration of E1 enzyme in the K11-chain assembly reaction can be reduced if commercial enzyme is being used.
4. Primer sequence for USP5/IsoT ZnF UBP domain amplification:
UBP-FW 5-CCAAGGTTCCATGGTACGGCAGGTGTCT AAGCATGCC-3;
UBP-RV 5-GCCTAGCGGCCGCTTATGTCTTCTGCAT CT-TCAGCATGTCGATG-3.
5. Different UBDs were fused to UBE2S, but neither the ubiq-uitin-associated (UBA) domain of Cezanne (an OTU-family deubiquitinase) nor the first ubiquitin-interacting motif (UIM) of the 26S proteasome regulatory subunit S5A improved free K11-linked ubiquitin chain assembly by UBE2S.
6. GST-fusion proteins were expressed in Rosetta 2 (DE3) pLacI cells from Novagen. These cells are BL21 derivates and carry a
4. Notes
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269

22715 Synthesis and Analysis of K11-Linked Ubiquitin Chains
plasmid with tRNA genes that decode seven rare codons. The additional tRNAs can significantly improve the yield of recom-binant human proteins.
7. To instantly check if the GST-fusion protein bound to the Glutathione Sepharose 4B, a small aliquot (1 ml) of resin can be mixed with 1 ml Bradford protein assay reagent.
8. Concentration of GST-fusion proteins is important since the 5 mM DTT in the low-salt buffer can interfere with the ubiquitin chain assembly reaction. Purified UBE2S variants and AMSH are stable at high concentrations such as ~10 mg/ml, (295 mM UBE2S-UBD and 200 mM AMSH). From 1 l cultures roughly 5–15 mg protein can be purified.
9. Freshly purified UBE2S-UBD can be tested in small-scale autoubiquitination reactions using the set of ubiquitin K-only mutants (10). It is also advisable to test the large-scale assem-bly reaction at 20-fold smaller scale, which should result in visible bands of di- and triubiquitin in Coomassie stained SDS-PAGE gels.
10. Dilution of the reaction with buffer A drops the pH to 4.5 and causes precipitation of all proteins except mono- and polyubiquitin.
11. The absorbance of ubiquitin at 280 nm is very low since the molecule contains only one tyrosine residue and no tryptophan or cysteine residues. An extinction coefficient of 0.16 should be used to estimate ubiquitin concentration from OD280 measurements, e.g. using a Nanodrop spectrophotometer.
12. We experienced that the separation of K11-linked ubiquitin chains by cation-exchange chromatography was not always complete and that for example ubiquitin monomers and dim-ers eluted together from the Mono S™ column in the same fractions (see Fig. 2b). In this case, all fractions containing dimers were pooled, concentrated, diluted in buffer A to reduce salt concentration, and repurified by cation-exchange chromatography.
13. Analysing the eluted peak fractions by SDS-PAGE and Coomassie staining reveals the quality of the assembled K11-linked ubiquitin chains. Double bands for tri- and tetraubiq-uitin indicate contaminating K63 linkages as differently linked ubiquitin chains show distinct electrophoretic mobility (10).
14. Ubiquitin chains generated by the UBE2S-UBD fusion protein can be verified by tryptic digest and LC-MS/MS to verify the K11 linkage (10).
15. To obtain larger amounts of K11-linked tetramers, it is possible to use the purified dimers as input material in a second chain assembly reaction.
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314

228 A. Bremm and D. Komander
16. K11-linked ubiquitin chains can be synthesised from mutant K63R ubiquitin. This avoids the generation of contaminating K63 linkages by UBE2S-UBD without including AMSH in the assembly reaction.
References
1. Xu P, et al (2009) Quantitative proteomics reveals the function of unconventional ubiq-uitin chains in proteasomal degradation. Cell 137:133–145
2. Chen ZJ, Sun LJ (2009) Nonproteolytic func-tions of ubiquitin in cell signaling. Mol Cell 33: 275–286
3. Hershko A, Ciechanover A (1998) The ubiq-uitin system. Annu Rev Biochem 67:425–479
4. Dye BT Schulman BA (2007) Structural mechanisms underlying posttranslational modification by ubiquitin-like proteins. Annu Rev Biophys Biomol Struct 36: 131–150.
5. Ye Y, Rape M (2009) Building ubiquitin chains: E2 enzymes at work. Nat Rev Mol Cell Biol 10:755–764
6. Jin L, et al (2008) Mechanism of ubiquitin-chain formation by the human anaphase-promoting complex. Cell 133:653–665
7. Dynek JN, et al (2010) c-IAP1 and UbcH5 promote K11-linked polyubiquitination of RIP1 in TNF signalling. EMBO J 29(24): 4198–209
8. Williamson A, et al (2009) Identification of a physiological E2 module for the human ana-phase-promoting complex. Proc Natl Acad Sci U S A 106:18213–18218
9. Garnett MJ, et al (2009) UBE2S elongates ubiq-uitin chains on APC/C substrates to promote mitotic exit. Nat Cell Biol 11: 1363–1369
10. Bremm A, Freund SM, Komander D (2010) Lys11-linked ubiquitin chains adopt compact conformations and are preferentially hydro-lyzed by the deubiquitinase Cezanne. Nat Struct Mol Biol 17:939–947
11. Reyes-Turcu FE, et al (2006) The ubiquitin binding domain ZnF UBP recognizes the C-terminal diglycine motif of unanchored ubiq-uitin. Cell 124: 1197–1208
12. Reyes-Turcu FE, et al (2008) Recognition of polyubiquitin isoforms by the multiple ubiq-uitin binding modules of isopeptidase T. J Biol Chem 283:19581–19592
13. Komander D, et al (2008) The structure of the CYLD USP domain explains its specificity for Lys63-linked polyubiquitin and reveals a B box module. Mol Cell 29:451–464
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
359
360
361
362
363
364
365
366
367





























