Embed Size (px)
Citation preview

1
DEVELOPMENTAL PROGRAMMING OF THE PREIMPLANTATION BOVINE EMBRYO BY COLONY STIMULATING FACTOR 2
By
KYLE BRADLEY DOBBS
A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY
UNIVERSITY OF FLORIDA
2014

2
© 2014 Kyle Bradley Dobbs

3
To Lauren, Mom, Dad and Kory

4
ACKNOWLEDGMENTS
I would like to thank my advisor, Dr. Peter J. Hansen, for mentoring me on what it
takes to truly be a scientist. He always challenged me, which brought out my best, but
did so while also being respectful and understanding. His advice, knowledge, expertise
and willingness to take a chance on bringing me down to Florida are some things I will
never forget and for which I am truly grateful. I would also like to thank my committee
members, Dr. Stephanie Wohlgemuth, Dr. James Resnick and Dr. Kenneth Drury for all
the support and advice that was given to me during my PhD program.
I also thank the members of the Hansen lab, both past and present, for all their
help during my program. Among my lab mates deserving thanks are Anna Denicol, Miki
Sakatani, Manabu Ozawa, James Moss, Luciano Bonilla, Maria Padua, Sofia Ortega,
Sarah Cochran, Firdous Khan, Mateus Sudano, Juliana Delgado, Antonio Ruiz, Paula
Tribulo, Veronica Negron-Perez, Luiz Siqueira, Jasmine Francis, Serdal Dikman,
Christine Meyer and Marlon Rodriguez. Without you guys, I would not have been
productive and for that, I am grateful.
I am very grateful to the lab of Dr. Marc-André Sirard and Dr. Claude Robert in
Québec City, QC for their help and assistance teaching me how to do microarray
analysis and helping me survive Québec with my broken French. Merci beaucoup!
Thank you to Justin Fear and Dr. Alice Morse in the Lauren McInytre lab for their
assistance with analyzing microarray data, specifically. I am also thankful to Dr. Alan
Ealy and his lab for assistance on IFNT assays and to Stacey Jones and the IFAS
communications office for designing figures for the dissertation.
Thanks also to Central Packing Co. in Center Hill, FL for providing the lab with
ovaries and fetuses for my experiments and William Rembert for collecting these

5
samples. Special thanks go to Eric Diepersloot for his assistance and guidance at the
University of Florida Dairy Research Unit (Hague, FL). I came into this lab with little
experience handling cattle and Eric showed me the ropes.
I am also grateful for the Animal Molecular and Cellular Biology Graduate
Program for the fellowship that provided me with funding to complete my PhD. Also,
thanks to the Department of Animal Sciences for housing me and providing me with the
resources for my research. I am also grateful to the graduate students of the
department for their camaraderie and support including Dale Kelley, Rafael Bisinotto,
Karun Kaniyamattam, Natalia Patiño, Eduardo Ribeiro and Tao Sha.
My thanks go out to all of my Gainesville friends outside of the lab that my wife
and I were fortunate to meet, especially Rev. Dan and Jen Prugh as well as Dr. Peter
and Natalie Carter. They made us feel like family and were always there for us.
I also want to thank my Mom and Dad. No one has been there for me throughout
my whole life more than those two. They pushed me in all that I did and made me who I
am today. I know they never thought the son of a CPA and cartographer would
graduate with a PhD in science. I also thank the rest of my family, my brother Kory and
my grandmothers for their love and support. Life was made a lot easier during my
program with the love that was available 24/7 from our Jack-chi (dog), Lexi.
Finally, I thank my wife Lauren, for her love and support. It goes without saying
that I could not have done this without her and I am forever indebted to her. She was
and is always there for me and even moved 1,000+ miles from everyone and everything
that she knew to support me.

6
TABLE OF CONTENTS page
ACKNOWLEDGMENTS .................................................................................................. 4
LIST OF TABLES ............................................................................................................ 9
LIST OF FIGURES ........................................................................................................ 10
ABSTRACT ................................................................................................................... 15
CHAPTER
1 LITERATURE REVIEW .......................................................................................... 17
Introduction ............................................................................................................. 17 Development of the Preimplantation Embryo in the Cow ........................................ 19
Time Course of Development ........................................................................... 20 Embryonic Genome Activation ......................................................................... 23 DNA Methylation .............................................................................................. 24
Differentiation of the First Two Cell Lineages in the Blastocyst ........................ 26 Evidence for the Importance of the Maternal Environment for Regulation of
Development ....................................................................................................... 27 In Vitro Produced Embryo ................................................................................ 28
Alterations in Maternal Function ............................................................................. 33
Supplemental Progesterone ............................................................................. 33
Somatotropin .................................................................................................... 34
Lactation ........................................................................................................... 35 Developmental Programming ................................................................................. 35
CSF2 as an Embryokine ......................................................................................... 39 Role of Colony Stimulating Factor 2 as an Embryokine ................................... 40 Signal Transduction .......................................................................................... 40
Production of CSF2 by the Reproductive Tract ................................................ 41 Actions on the Preimplantation Embryo ........................................................... 42 Characteristics of the CSF2R in the Preimplantation Embryo .......................... 45
Goals of the Current Investigation .......................................................................... 46
2 REGULATION OF PLURIPOTENCY OF INNER CELL MASS AND GROWTH AND DIFFERENTIATION OF TROPHECTODERM OF THE BOVINE EMBRYO BY COLONY STIMULATING FACTOR 2 ............................................................... 48
Introduction ............................................................................................................. 48 Materials and Methods............................................................................................ 50
Embryo Production ........................................................................................... 50 Effect of CSF2 on Survival of Isolated Inner Cell Mass .................................... 51
Bovine fetal fibroblast feeder cells ............................................................. 51

7
Isolation of ICM by lysis of trophectoderm using antibody and complement via immunosurgery ............................................................. 52
Culture of isolated ICM .............................................................................. 53
Experiments ..................................................................................................... 54 Analysis of ICM Colonies Surviving Passage ................................................... 54 Expression of Pluripotency Genes in ICM ........................................................ 55 Actions of CSF2 on Competence of Trophectoderm to Form Outgrowths ....... 55
Establishment of outgrowths ...................................................................... 55
Immunolabeling of CDX2 ........................................................................... 56 Assessment of establishment and growth of trophectoderm outgrowths ... 57 Gene expression ........................................................................................ 57 Antiviral assay ............................................................................................ 57
Experiments ..................................................................................................... 58
Expression of CSF2 Receptor Subunit Genes ................................................. 58 Quantitative PCR .............................................................................................. 59
Statistical Analysis ............................................................................................ 60
Results .................................................................................................................... 61 Blastocyst Development ................................................................................... 61 Effect of CSF2 on Survival of Isolated ICM ...................................................... 62
Expression of Genes Involved in Pluripotency and Differentiation ................... 63 TE Outgrowth ................................................................................................... 64
Expression of CSF2 Receptor Subunit Genes ................................................. 65 Discussion .............................................................................................................. 65
3 SEXUAL DIMORPHISM IN DEVELOPMENTAL PROGRAMMING OF THE BOVINE PREIMPLANTATION EMBRYO CAUSED BY COLONY STIMULATING FACTOR 2 ..................................................................................... 77
Introduction ............................................................................................................. 77 Materials and Methods............................................................................................ 80
Embryo Production ........................................................................................... 80 Embryo Transfer ............................................................................................... 80 Embryo Recovery ............................................................................................. 81
IFNT Assay ....................................................................................................... 82 Extraction of gDNA and Total RNA .................................................................. 83 Sexing by PCR ................................................................................................. 83 Overview of Analysis of Transcriptome ............................................................ 84
RNA amplification ...................................................................................... 84
RNA labeling .............................................................................................. 85
RNA microarray hybridization and scanning .............................................. 85 Analysis of transcriptome microarray data ................................................. 86 Validation of trancriptome microarray by PCR ........................................... 87
Overview of Analysis of Methylome .................................................................. 88 Fragmentation and amplification of gDNA ................................................. 88 DNA labeling .............................................................................................. 90 DNA microarray hybridization and scanning .............................................. 90 Analysis of methylome array ...................................................................... 91

8
Results .................................................................................................................... 91
Embryo Length and IFNT Accumulation in the Uterus ..................................... 91 Differentially Expressed Genes ........................................................................ 92
Methylome ........................................................................................................ 94 Discussion .............................................................................................................. 95
4 DYNAMICS OF DNA METHYLATION DURING EARLY DEVELOPMENT OF THE PREIMPLANTATION BOVINE EMBRYO ..................................................... 111
Introduction ........................................................................................................... 111
Experimental Procedures ...................................................................................... 113 Embryo Production ......................................................................................... 113 Immunofluorescent Labeling for 5-Methylcytosine ......................................... 114 Immunofluorescent Labeling for 5-Methylcytosine and CDX2 ........................ 115
Image Analysis Using Imagej ......................................................................... 115 Separation of TE and ICM by Magnetic-Activated Cell Sorting ...................... 116
Reverse Transcription and Quantitative PCR ................................................. 117 High Resolution Melting Analysis ................................................................... 118
Design of Experiments .......................................................................................... 120 Statistical Analysis ................................................................................................ 123 Results .................................................................................................................. 123
Changes in DNA Methylation during Development to the Blastocyst Stage ... 123 Effect of Sex on DNA Methylation .................................................................. 123
DNA Methylation in Blastocysts as Modulated by Sex, CSF2 and Cell Differentiation .............................................................................................. 124
Developmental Changes in DNMT3B Gene Expression ................................ 124
Developmental Changes in Methylation of DNMT3B ..................................... 125 Discussion ............................................................................................................ 125
5 GENERAL DISCUSSION ..................................................................................... 138
REFERENCES ............................................................................................................ 147
BIOGRAPHICAL SKETCH .......................................................................................... 178

9
LIST OF TABLES
Table page 2-1 Effects of CSF2 from Day 5 to 7 or 6 to 8 post-insemination on
characteristics of TE outgrowths. ........................................................................ 71
3-1 List of primers used for qPCR. .......................................................................... 101
3-2 Validation of microarray results by determination of the correlation between fluorescent intensity on the array with CT value in qPCR. ................................. 102

10
LIST OF FIGURES
Figure page 2-1 The effect of CSF2 on the percent of oocytes that became a blastocyst
depends on the overall competence of embryos to develop to the blastocyst stage. .................................................................................................................. 72
2-2 Treatment with CSF2 increases ability of isolated ICM to survive following passage.. ............................................................................................................ 73
2-3 Representative appearance of colonies of cells derived from ICM at Day 7 passage 1. .......................................................................................................... 74
2-4 Lack of effect of CSF2 on expression of genes related to pluripotency and differentiation in ICM isolated from Day 8 blastocysts.. ...................................... 75
2-5 Developmental changes in steady-state mRNA for CSF2RA and CSF2RB.. ..... 76
3-1 Sex by CSF2 interaction affecting length of the embryo and accumulation of IFNT in uterine flushings at day 15 of pregnancy.. ........................................... 103
3-2 Venn diagram illustrating differentially-expressed transcripts at day 15 of pregnancy in male and female embryos treated with CSF2 or control medium from day 5-7 of development.. .......................................................................... 104
3-3 Diagram illustrating genes encoding for proteins in the electron transport chain that were differentially expressed between control female and male embryos at day 15 of pregnancy.. .................................................................... 105
3-4 Biological processes predicted to vary with sex in control embryos... .............. 106
3-5 Predicted changes in cellular function in male embryos at day 15 of pregnancy caused by CSF2 treatment between day 5-7 of development.. ...... 107
3-6 Venn diagram illustrating differentially-methylated probes at day 15 of pregnancy in male and female embryos treated with CSF2 or control medium from day 5-7 of development. ........................................................................... 108
3-7 Non-random clustering of differentially methylated probes – example from bovine chromosome 1.. .................................................................................... 109
3-8 Model illustrating a mechanism for achievement of sex-specific developmental programming in response to a change in maternal environment.. .................................................................................................... 110

11
4-1 Representative images of embryos labeled for immunoreactive 5- methylcytosine at the 2 cell, 4 cell, 6–8 cell, 9–25 cell, 32 cell-morula and blastocyst stages of development.. ................................................................... 130
4-2 Developmental changes in labeling of immunoreactive 5-methylcytosine from the 2-cell to blastocyst stages of development.. ............................................... 131
4-3 Effects of sex on immunoreactive 5-methylcytosine in embryos at the 6–8 cell and blastocyst stages of development.. ............................................................ 132
4-4 Representative images of an embryo in which labeling with CDX2 (blue) was used to evaluate immunoreactive 5-methylcytosine (green) in ICM (CDX2-negative) and TE (CDX2-positive).. .................................................................. 133
4-5 Effects of sex and cell type [ICM and TE] on immunoreactive 5-methylcytosine in blastocysts.. ......................................................................... 134
4-6 Developmental changes in steady-state mRNA for DNMT3B.. ........................ 135
4-7 Characteristics of the high resolution melt analysis for methylation of an intronic region of DNMT3B.. ............................................................................. 136
4-8 Developmental changes in DNA methylation for DNMT3B.. ............................. 137
5-1 Schematic illustration of conclusions derived from the dissertation including developmental changes in methylation and sex-specific actions of CSF2 on embryonic development.. ................................................................................. 146

12
LIST OF ABBREVIATIONS
Note: Official gene symbols are used without definition as recommended by the HUGO Gene Nomenclature Committee (http://www.genenames.org)
2i Two inhibitors
ABAM Anti-bacterial, anti-microbial
aRNA Amplified ribonucleic acid
ATP Adenosine triphosphate
BSA Bovine serum albumin
bST Bovine somatotropin
BTA Bos taurus
cDNA Complementary deoxyribonucleic acid
COC Cumulus oocyte complex
ConA Concanavalin A
CpG Cytosine-guanine dinucleotide
CT Cycle threshold
DAPI 4’,6-diamidino-2-phenylindole
DAVID Database for annotation, visualization and integrated discovery
DEG Differentially expressed genes
DMEM Dulbecco modified eagle medium
DMR Differentially methylated region
DMSO Dimethyl sulfoxide
DNA Deoxyribonucleic acid
DNMT DNA methyltransferase
DPBS Dulbecco’s phosphate buffered saline
ED Embryonic disc

13
EEM Extra-embryonic membrane
EGA Embryonic genome activation
ESC Embryonic stem cell
ESM Embryonic stem cell medium
FBS Fetal bovine serum
FITC Fluorescein isothiocyanate
gDNA Genomic deoxyribonucleic acid
GO Gene ontology
ICM Inner cell mass
IgG Immunoglobulin G
IPA Ingenuity pathway analysis
iPATH Interactive pathway explorer
IVF In vitro fertilization
IVP In vitro produced
JAK Janus kinase
KEGG Kyoto encyclopedia of genes and genomes
MACS Magnetic activated cell sorting
MAPK Mitogen-activated protein kinase
MS-HRM Methylation sensitive high resolution melting analysis
PBS Phosphate buffered saline
pi post-insemination
PI Propidium iodide
PI3K Phosphoinositide 3-kinase
PVA Polyvinyl alcohol
PVP Polyvinylpyrrolidone

14
qPCR Quantitative polymerase chain reaction
RNA Ribonucleic acid
ROX Reactive oxygen species
siRNA Short interfering ribonucleic acid
SOF-BE1 Synthetic oviductal fluid-bovine embryo 1
STAT Signal transduction and activation of transcription
SWI/SNF Switch/Sucrose nonfermentable
TCJ Tight cell junction
TE Trophectoderm
TUNEL Terminal deoxynucleotidyl transferase

15
Abstract of Dissertation Presented to the Graduate School of the University of Florida in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy
DEVELOPMENTAL PROGRAMMING OF THE PREIMPLANTATION BOVINE
EMBRYO BY COLONY STIMULATING FACTOR 2
By
Kyle Bradley Dobbs
May 2014
Chair: Peter J. Hansen Major: Animal Molecular and Cellular Biology
The maternal environment is an important determinant of preimplantation
development of the mammalian embryo. Many actions of the mother are mediated by
alterations in secretion of regulatory molecules called embryokines produced by the
endometrium. One of these molecules, colony stimulating factor 2 (CSF2), causes
several actions on the embryo when added from days 5-7 of development including
altering lineage commitment of the inner cell mass (ICM) and trophectoderm (TE) of the
blastocyst, increasing competence of an embryo to become a blastocyst and to
establish pregnancy after transfer. Multiple experiments were carried out to understand
the mechanisms of CSF2 on the development of the preimplantation bovine embryo.
First, experiments were conducted to evaluate regulation by CSF2 of pluripotency within
the ICM and TE differentiation and growth. The proportion of isolated ICM colonies that
survived until the end of the first cellular passage was greater for ICM derived from
blastocysts produced in the presence of CSF2 than ICM from control blastocysts.
There was little effect of CSF2 on characteristics of TE outgrowths from blastocysts. In
addition, actions of CSF2 were found to occur through a signaling pathway that is likely

16
independent of CSF2RB because CSF2RB was undetectable from the zygote to
blastocyst stages. Second, the actions of CSF2 from day 5-7 of development on
characteristics of the embryo at day 15 of gestation to determine whether CSF2 causes
long-term changes in embryonic development. CSF2 decreased length and IFNT
secretion of female embryos at Day15 but increased length and IFNT secretion of male
embryos. Similarly, CSF2 affected the transcriptome and methylome differently for
female embryos than male embryos. Finally, immunofluorescent labeling with anti-5-
methylcytosine was used to evaluate global DNA methylation during preimplantation
development. Results suggest that DNA methylation undergoes dynamic changes
during the preimplantation period in a manner that is dependent upon sex and that the
TE of the blastocyst is more methylated than the ICM. There was no effect of CSF2 on
global DNA methylation at the blastocyst stage. Overall, these results establish a
mechanism by which the maternally secreted embryokine, CSF2, alters the
developmental trajectory of the bovine preimplantation embryo.

17
CHAPTER 1 LITERATURE REVIEW
Introduction
The mammalian embryo only spends a few days developing from the totipotent
zygote stage to the blastocyst stage in which the first two lineage commitment events
take place. In cattle, the blastocyst, consisting of ICM and TE, forms by Day 6-7 in vivo
[1] and the hypoblast forms about 48 hours later [2]. Although development of the
zygote to the blastocyst stage can occur in the absence of maternal signals (i.e., in
culture media), the maternal environment is important for ensuring optimal
development. Indeed, embryos produced in vitro have several characteristics that are
altered compared to embryos that develop in vivo, including gene expression [3], lipid
content [4, 5], ultrastructure [6, 7], and degree of methylation of CpG islands of specific
genes [8]. Moreover, perturbations in the maternal environment during the
preimplantation period can compromise embryonic survival and amend the
developmental program of the embryo to cause long-term changes in physiology and
health that extend into postnatal life.
In the cow, there are several lines of evidence indicating the importance of
perturbations in the maternal environment for development of the preimplantation
embryo. An experiment in which embryos were transferred into the oviduct indicated
that lactation causes reduced ability of embryos to develop to the blastocyst stage [9].
Cows in which embryos were transferred into the reproductive tract at day 7 after estrus
had a different pattern of endometrial gene expression on the day of transfer than cows
in which the embryo did not survive [10]. The maternal hormonal environment can alter
characteristics of the embryo as well, as shown by studies that premature elevation of

18
progesterone after ovulation hastened elongation of the trophoblast [11] and that
treatment of superovulated cows with somatotropin increased the competence of the
resultant embryos to establish pregnancy after embryos were recovered and transferred
to recipients [12].
Experiments indicating that alterations in maternal environment can reconfigure
the developmental program of the embryo come largely from experiments with rodents.
Female mice or rats fed a low protein diet during the period corresponding to
preimplantation development, produced changes in the resultant offspring, which
persisted into adult life [13-15]. In the sheep, as well, change in nutritional environment
during the peri-conception period (this time by reducing the bioavailability of methionine)
caused changes in the adult offspring that included increased rate of obesity, altered
immune response, elevated blood pressure and resistance to insulin [16].
One of the characteristics of developmental programming caused by changes in
the maternal environment during the preimplantation period is that females are
programmed differently than males. For example, feeding female mice diets low in
protein during embryonic development affected females differently than males [13, 14].
Male adults born from mothers that were fed low protein diets had elevated angiotensin-
converting enzyme [17], while female adults had increased blood pressure [14] and
reduced heart to body weight ratio [13].
One of the assumptions of this dissertation is that a major mechanism by which
changes in uterine environment affects embryonic development is change in the release
of soluble regulators of embryonic development. These molecules, hereafter called
embryokines, are produced by the oviduct and endometrium and are capable of altering

19
the course of embryo development. Indeed, several maternally-derived molecules have
been identified that have beneficial effects on the preimplantation embryo. In the cow,
these include CSF2 [18], IGF1 [19], TGFB [20], hyaluronan [21], LIF [20], FGF2 [22] and
ILB1 [23].
The best-studied embryokine is CSF2, which can improve early development and
embryonic competence for establishment of pregnancy in mice, cows and humans [18,
24-26]. CSF2 is secreted by the endometrial and oviductal epithelium [27-29].
Treatment of in vitro produced bovine embryos with CSF2 alters gene expression [30],
decreases apoptosis [31] and increases ICM to TE cell ratio within the blastocyst [18].
At least two environmental factors can affect CSF2 synthesis in the uterus – seminal
plasma [shown in mice; [32, 33]] and obesity (shown in cow) [34].
This literature review will be focused on understanding how the maternal
reproductive milieu regulates development of the early embryo. The focus will be on the
specific role of CSF2 as a maternal embryokine. While data from a variety of species
will be dealt with, the major research model discussed will be the cow. For that reason,
and to put research on maternal effects on embryonic development in context, the
review will begin with an overview of the process of preimplantation development in the
cow.
Development of the Preimplantation Embryo in the Cow
Preimplantation development involves a specific chain of events. Following
fertilization of the oocyte, the embryo proceeds through symmetric cleavages, doubling
its cell number and decreasing cell size with each cleavage. During this time, the
embryo utilizes maternally inherited mRNA for protein production until transcription is
initiated in a process called embryonic genome activation (EGA). Although the exact

20
timing is unclear, around the same time as EGA, the embryo loses all DNA methylation
marks before regaining them during de novo methylation involving actions of embryo-
specific methyltransferase enzymes. Finally, following blastocyst formation, the embryo
hatches and begins to elongate, increasing its size more than 20 fold [35] and then
begins to attach to the epithelium of the endometrium in a gradual process that is
initiated around Day 20 [36].
Time Course of Development
Immediately after ovulation, the oocyte is picked up by the fimbria of the oviduct
and moved into the oviductal lumen [37]. Fertilization takes place near the junction of
the isthmus and ampulla [38]. Following fertilization, the two pronuclei fuse, and
meiosis is completed by extrusion of the second polar body [39, 40]. The first round of
cleavage following fertilization occurs within 28-35 hours [41]. This is followed by
additional rounds of cell division so that the embryo reaches the 4-cell stage by 40-46
hours after fertilization, the 8-cell stage by 58-86 hours after fertilization and the 16-cell
stage by 106 hours post-fertilization [41]. Cleavage throughout development is
asynchronous [41].
By 134 hours post fertilization, the formation of tight cell junctions (TCJ) have
formed [41]. TCJ formation involves the gathering of TCJ constituents to form a protein
cluster on E-cadherin complex, which is located between blastomeres [42]. The TCJ
becomes sealed during the 32-cell to early blastocyst stage following the addition of
tight junction protein alpha [42]. TCJ formation represents the first step in differentiation
of the embryo because the presence of tight junctions causes a different
microenvironment for cells on the outside of the embryo from cells on the inside of the

21
embryo. In mice, position of blastomeres at the time of morula formation is an important
determinant of whether cells give rise to ICM or TE cells [43].
Blastocyst formation occurs about 168 hours post insemination [1]. Accumulation
of fluid in the blastocoelic cavity is the result of actions of Na+/K+ ATPase [44] as well as
facilitated movement of water by aquaporin channel proteins [45]. The blastocyst
represents the departure point for the embryo. Before the blastocyst stage, all cells of
the embryo are totipotent. Beginning at the blastocyst stage, however, cells becoming
increasingly restricted in the lineages they can form. The first differentiation is at the
blastocyst stage where TE cells become committed towards a placental lineage [46].
Cells of the ICM give rise to the pluripotent epiblast and the hypoblast, which
contributes to placental endoderm. The hypoblast can first be identified at day 8 after
fertilization as a group of cells on the outer face of the ICM that label with GATA6 [47].
Competency of an embryo to develop to the blastocyst stage is in part due to the
timing of cleavage. The longer the time between fertilization and the initial cleavage,
the lower the likelihood that an embryo can develop to the blastocyst stage [48]. In
vitro, embryos that reached the 16-cell stage by 72 hours after insemination produced
more expanded and hatched blastocysts compared to embryos that were only at the 1-4
cell stage during the same time frame [48]. At least some of these differences in
developmental potential are likely to be caused by differences in the transcriptome.
Embryos that reached the 2-cell stage by 29.5 hours post-fertilization had increased
expression of genes relating to cell-cycle control and DNA damage response factors
compared to embryos requiring 46 hours to reach the same cell stage [49].

22
The timing of development is not the same for embryos produced in vitro versus
those produced in vivo. In vitro produced embryos proceed through their first cleavage
about 33 hours after fertilization, about 5-9 hours later than their in vivo counterparts [1].
This delay persists throughout development; for example blastocysts form about one
day sooner in vivo than in vitro [1]. The timing of development is also dependent upon
the sex of the embryo. Female embryos reach the 2-4-cell and blastocyst stages slower
than male embryos [50]. In addition, female blastocysts are less likely to be expanded
at day 8 compared to males [51].
At 192-240 hours post-insemination, the bovine embryo proceeds through a
series of events that induces hatching of the embryo from the surrounding zona
pellucida [48, 52]. Hatching involves two processes: lysis of the zona pellucida with the
embryo-derived enzyme plasmin [1] and expansion of the blastocoel cavity to apply
pressure to the zona pellucida and allows the blastocyst to protrude and eventually
vacate from the structure. After hatching around day 8-10 after fertilization, the
blastocyst undergoes a series of morphological changes to change shape from
spherical to ovoidal [2]. By day 14 following fertilization, the embryo begins a dramatic
elongation of the trophoblast. At first the embryo assumes a tubular shape, with sizes
ranging from 0.5 mm to 19.0 mm in length [2]. Subsequently, the embryo assumes a
filamentous shape and reaches lengths of 6.0 mm to 160.0 mm by day 16 [1]. The
initiation of elongation is reliant upon the maternal environment, as in vitro produced
embryos will not elongate [53].
Rapid elongation also requires rapid cell proliferation. Over 500 genes,
predominantly associated with trophoblast cell proliferation, are upregulated in embryos

23
between day 7 and 14 [54]. By day 14-16, the process of gastrulation has begun with
the formation of the three germ layers [2].
One function of the elongating embryo is production of IFNT to signal to the
mother to block luteolysis and allow continued secretion of progesterone from the
corpus luteum [55]. As the embryo increases in length, secretion of IFNT also
increases [56, 57]. It is not clear whether the increase in IFNT with size is associated
with an increase in INFT gene expression per cell or simply the larger size [54, 57].
Starting at gestational day 20, the elongating bovine embryo begins the process
of apposition and adhesion to the endometrial epithelium [54]. The bovine embryo, like
other species with an epitheliochorial placentation, does not invade through the
epithelial basement membrane to cause implantation. Rather, some embryonic cells
adhere to epithelial cells while others (binucleate cells) migrate into the endometrial
epithelium and fuse with maternal cells [58]. The process of placentation in the cow is
slow, requiring around 12 days to be completed. [59].
Embryonic Genome Activation
One of the key events in development is activation of the embryonic genome.
Transcription in the oocyte ceases when it reaches a size of 110 μm [60]. Therefore,
the oocyte and embryo are dependent upon preformed mRNA for direction of protein
synthesis until the genome is reactivated. The embryo manages protein synthesis by
modifying polyadenylation of maternally-donated mRNA and controlling the cell-cycle
clock [61, 62]. The first round of transcription from the embryonic genome starts at the
2-4-cell stage, where a couple hundred genes are activated [63]. Some of the genes
transcribed during this period of minor genome activation are SARS, IL18, CRABP1,
ACO2, TXN2, SLC38A2 and SLC25A3 [63]. Minor activation is not required for

24
development to the blastocyst stage because treatment with alpha-amanitin to block
transcription prior to major EGA had no effect on the percent of embryos that became
blastocysts [64]. Embryonic RNA transcription begins at the 8-cell stage [65]. Full
activation of the embryonic genome occurs between the 8-cell and 16-cell stage [66].
Development is blocked at this stage if the embryo is exposed to alpha-amanitin [64].
The mechanism by which EGA occurs in the cow is unknown. In other species
such as mice, Drosophila and C. elegans, EGA is proposed to be initiated by removal of
maternal transcripts through destabilization of the poly-adenylated tail [67]. Other
mechanisms that are also likely to be involved including removal of transcriptional
repressors and chromatin modifications such as the SWI/SNF complex from embryonic
DNA, dilution of an unknown EGA repressor caused by cellular division, and the
acquisition of the cellular machinery for transcription [67].
DNA Methylation
DNA methylation involves the addition of a methyl-group to the 5th carbon
position of cytosine when located 5’ to guanine in a DNA sequence, also known as a
CpG dinucleotide [68]. DNA methylation is an important mechanism for regulating gene
expression. Methylated DNA elicits a transcriptionally-repressive response by recruiting
binding proteins to the DNA strand and by physically impeding transcriptional machinery
from proceeding 5’ to 3’ across the DNA [69, 70]. DNA methylation is thus an important
mechanism for controlling a cell’s differentiation status, by controlling repression of
specific DNA regions [71]. In addition, paternal imprinting involves DNA methylation on
chromosomes of specific paternal origin [72]. Given the role of DNA methylation in
controlling differentiation, it is necessary for the embryo to remove methylation marks as

25
part of the process to establish totipotency and restore methylation in a cell lineage-
dependent manner.
The initial loss of global DNA methylation following fertilization occurs by both
active and passive mechanisms [73, 74]. At least in the mouse, the maternally-derived
genome is demethylated more slowly after fertilization than paternally-derived genome
[75]. Loss of methylation in maternally-derived genome is passive due to the absence of
a de novo DNA methyltransferase in the early embryo [76]. In contrast, DNA
demethylation of the paternally-derived genome is accomplished by a host of DNA
repair mechanisms such as base excision repair through DNA glycosylases [77],
nucleotide excision repair [78], deamination of the methylated cytosine [79] in addition
to 5-methylcytosine oxidation [80]. A specific DNA demethylation enzyme has not been
identified. Before embryonic development, the methylation marks on imprinted genes
are established and subsequently are maintained during global demethylation [72] and
are only demethylated during primordial germ cell migration [81].
Following demethylation, the embryo reacquires methylation marks as
development proceeds. Indeed, development will not occur unless those methylation
marks are restored by DNA methyltransferases [82]. DNA methyltransferases Dnmt3a
and Dnmt3b are activated in order to re-establish the methylation status of embryonic
DNA [82]. Unique to the embryo is an imprinting maintenance DNA methyltransferases,
DNMT1o. Although imprinting occurs during oogenesis in the absence of DNMT1o,
these imprinted marks will disappear during early embryo development when DNMT1o
is absent [83].

26
While DNA demethylation and de novo methylation are critical events, there is
wide divergence in the developmental and lineage-specific characteristics of the
process. In the mouse, demethylation is complete by the late morula stage, de novo
methylation begins at the blastocyst stage of development, and the ICM is more
methylated than the TE [84]. For sheep, in contrast, global methylation decreases
continuously from the 2-cell stage until the blastocyst stage [85] while the pig embryo
does not undergo a loss of methylation between the zygote and blastocyst stages [86].
For both pig and sheep, the ICM is more methylated than the TE [85, 86]. In the cow, it
is unclear at what stage demethylation is complete or at what stage de novo methylation
begins [87, 88].
Differentiation of the First Two Cell Lineages in the Blastocyst
Three cell populations emerge during the blastocyst stage: the ICM, TE and
hypoblast [called primitive endoderm (PE) in mice]. The ICM is a population of cells that
will become the embryo proper while the TE and hypoblast will develop into the extra-
embryonic membranes forming the placenta and endodermal structures. Presence of
specific transcription factors can be used to identify cells of the ICM (NANOG and
SOX2) [89-91], TE (CDX2) or hypoblast (GATA6) [92] lineage. Embryos that have
inactivation of CDX2 will fail to develop a TE [93]. CDX2 is required for differentiation of
TE since inactivation of the gene prevents TE formation [93]. Unlike the mouse,
however, POU5F1 is not responsible for maintenance of ICM and is expressed in both
ICM and TE fate [94]. Commitment to the TE lineage is a gradual process; cells at Day
7 of development retain the capacity to revert back to a pluripotent cell type [94]. By
Day 11 of development, the expression of CDX2 is more than 10 times higher than

27
POU5F1, which subsequently inhibits the activity of POU5F1 and causes irreversible
differentiation of TE cells [94].
GATA6 is initially expressed throughout the blastocyst and then becomes
increasingly restricted to the ICM which is characterized by a mixture of cells expressing
either GATA6 or the epiblast marker NANOG [47]. GATA6 is not absent from the TE
until Day 8 of development [47]. Heterogeneity of the ICM could be the result of variable
responses to Fgf4 signaling as shown in the mouse. In that species, the second cells
produced as a result of proliferation have greater expression of Fgfr2 than cells
produced in the first wave [95, 96]. Cells from the second wave become hypoblast
while the cells from the first wave stay dedicated to the ICM [95]. Interestingly, if Fgf4
signaling through mitogen-activated protein kinase (MAPK) is blocked in the epiblast
cell population, all cells become Nanog positive and Gata6 negative [97]. In the bovine,
as well, inhibition of MAPK leads to an ICM where all cells are NANOG positive [98].
Moreover, activation of FGF4 signaling using human recombinant FGF4, increased the
number of cells in the ICM positive for GATA6 [47].
Evidence for the Importance of the Maternal Environment for Regulation of Development
That the reproductive tract environment established by the mother can affect the
trajectory of development of the preimplantation embryo can be deduced from two lines
of evidence. The first comes from examination of the characteristics of embryos
produced in vitro. While an embryo can develop to the blastocyst stage in the complete
absence of maternal signals, it is clear that such embryos experience a range of
abnormalities that affect competence to establish pregnancy and characteristics of the
resultant offspring. In some cases, such aberrations have been shown to be the result

28
of the culture conditions during embryo development rather than to disorders caused by
in vitro maturation or fertilization. The second line of evidence comes from experiments
indicating that alterations in maternal functions can alter embryonic development.
In Vitro Produced Embryo
Although preimplantation embryonic development ordinarily takes place in the
microenvironment of the female reproductive tract, healthy offspring can result when the
embryo resides in an artificial culture environment during the period up to and including
formation of the blastocyst. The embryo produced in vitro (IVP) is derived from an
oocyte that underwent maturation in the absence of maternal signals other than those
provided by the surrounding cumulus cells. Examination of the characteristics of the
IVP embryo make clear the importance of the maternal environment for regulation of
development because embryos produced in vitro experience a variety of abnormalities
compared to embryos produced in vivo. As a result, transfer of an IVP bovine embryo is
associated with lower pregnancy rate [99] and a higher incidence of abnormal calves at
birth [100].
In cattle, one of the consequences of in vitro production is an altered
transcriptome [101]. Alterations in mRNA expression begin as early as the 4-cell stage
[102]. By the blastocyst stage of development, there are hundreds of differentially
expressed genes between embryos produced by in vitro and in vivo methods [101, 103-
105]. Specific ontologies that are enriched for differentially expressed genes between
IVP and in vivo blastocysts include cholesterol synthesis and cell differentiation [101] as
well as apoptosis and stress response [105].
There is also aberrant DNA methylation in a fraction of IVP embryos [8, 88, 106].
Loss of methylation of the maternally-inherited allele of KvDMR1 has been shown to be

29
associated with overgrowth of bovine fetuses [106]. Among the genes that experience
decreased methylation following culture are genes involved in DNA methylation such as
the de novo methylatransferases DNMT3a and DNMT3b and histone
methyltransferases G9a and SUV39H1 [8].
Ultrastructural morphology of the embryo can be disrupted by IVP. By the
morula stage of development, embryos produced in vitro accumulate increased amount
of intracellular lipid and have fewer mitochondria as compared to embryos produced in
vivo [4]. Analysis of lipid profile revealed that IVP embryos had increased abundance of
phosphatidylcholines 32:0 and 34:1 [107]. These lipids contain low amounts of double
bonds so as to reduce membrane fluidity [107]. At the blastocyst stage, IVP has been
reported to decrease the number of microvilli that did not fully cover the plasma
membrane [7], increase the debris in the perivitelline space and lead to mitochondria
that are translucent in appearance [108].
Experiments have been conducted to determine whether the disruption of
development caused by IVP is the result of errors in oocyte maturation, fertilization or
embryonic development. It is clear that maturation in vitro can disrupt the matured
oocyte. First, the ultrastructure of the oocyte matured in vitro is different from oocytes
matured in vivo [109]. In particular, release of corticle granules after fertilization, which
are responsible for helping prevent polyspermy, are delayed [109]. Furthermore, in vitro
matured oocytes have cumulus cells that are smaller in size and less expanded [109].
In addition, gene expression in oocytes and cumulus cells from cumulus oocyte
complexes (COC) matured in vitro were found to differ from that of cumulus cells from
COC matured in vivo [110]. Using microarray, a total of 64 genes were found to be

30
differentially expressed in cumulus cells from COCs in vivo and in vitro [110]. Genes
that are downregulated in oocytes matured in vitro include PKP, GLUT1 and DSC2
[111]. Among differentially expressed genes for in vitro COCs were upregulated genes
related to stress response (HSPA5 and HSP90B1) and downregulated genes related to
anti-apoptosis and ATP binding (YWHAZ and ACTG1) [110]. Finally, oocytes that are
matured in vivo followed by in vitro fertilization (IVF) and culture are more likely to
become blastocysts on day 7 and 8 of development than oocytes that are matured in
vitro [112]. In vitro maturation in other species is also associated with alterations in
gene expression (mouse) [113], ultrastructure (pig) [114], reductions in oocyte
competence to form a blastocyst (mouse) [115], and ability of resultant embryos to
survive after transfer.
In vitro produced embryos also have reduced development because culture
conditions for the embryo following fertilization are inadequate. As compared to
embryos produced in vivo, Rizos et al. (2002c) observed a decrease in blastocyst yield
for oocytes that were matured in vivo and then cultured for IVF and culture [112].
Survival of bovine embryos after freezing could be increased if embryos produced by in
vitro maturation and IVF are placed inside the oviduct of female sheep [6, 105] following
collection at the blastocyst stage.
One characteristic of in vitro produced embryos is increased susceptibility to
damage after cryopreservation [6, 116, 117] and, as a result, cryopreserved IVP
embryos have poor survival after embryo transfer [118]. One key aspect of how well a
cryopreserved embryo develops is its lipid content prior to freezing [119]. Drugs such

31
as phenazine ethosulfate, which decrease the lipid content of the embryo, improve post-
thaw survivability of the IVP embryo [119].
Recently, Gad et al. (2012) performed an extensive experiment to determine the
specific stages of embryonic development relative to EGA in which culture environment
disrupts gene expression in bovine blastocysts. The transcriptome of six groups of
blastocysts were compared. Groups were as follows: 1) blastocysts produced in vitro, 2)
blastocysts produced in vivo, 3) blastocysts produced by in vitro maturation and
fertilization, cultured to the 4-cell stage and then transfer to recipients, 4) blastocysts
produced as for treatment 3 except that embryos were transferred to recipients at the
16-cell stage, 5) blastocysts that were produced in vivo and then placed in culture at the
4-cell stage, and 6) blastocysts produced in vivo and then placed in culture at the 16-
cell stage. The greatest deviation in the transcriptome occurred in the two groups that
were in culture during EGA – embryos cultured until the 16-cell stage or produced in
vivo and placed in culture at the 4-cell stage. In these two groups, there was
upregulation of genes involved in the NRF2-mediated oxidative stress pathway. This
result was interpreted to mean that the oviduct and the uterine environment provide
some protection against reactive oxygen species and the absence of that protection
leads to activation of oxidative stress genes Additionally, lipid metabolism genes
(MSMO1, ANXA1, ANXA3, HMGCR, HSD17B11, LDLR and ACAT2), were down-
regulated when embryo culture spanned EGA [102].
It is important to recognize that the culture conditions associated with IVP not
only compromise the ability of the embryo to establish pregnancy but also change the
nature of the developmental program so that there is a greater risk for fetal loss,

32
neonatal death and developmental abnormalities in offspring produced as a result of
IVP [99, 120-123]. Offspring from IVP have a higher rate of congenital malformations
including incidence of hydroallantois and aberrant limb formation [124]. Fetal [125, 126]
and birth weight [124, 127, 128] are increased in IVP offspring, which coincides with
higher rates of dystocia in recipients of IVP embryos [124, 127]. Calves born from IVP
embryos are also more likely to be born dead within the first 24 hours [129] or 20 days
of life [120].
Disorders in development caused by IVP are not unique to cattle. In humans,
IVP embryos have increased incidence of chromosomal abnormalities [130],
implantation failure [131], loss of DNA methylation imprinting [87], abnormal cell
division, cell allocation, cell death and embryonic arrest and death [132]. In the pig, IVP
embryos also have decreased cell numbers [133], over 588 differentially expressed
genes (DEG) [133], and aberrant ultrastructure including nucleoli located outside of the
nuclear membrane and malformed smooth endoplasmic reticulum [134]. Mouse IVP
embryos also have differential gene expression [135] and abnormal methylation at
imprinted genes [136].
Consequences of being derived from an IVP embryo can persist into adulthood.
This question has not been well examined in the cow but there is evidence for this idea
from other species. Mice born from IVP embryos have increased blood pressure at 21
weeks of age [13] while children between the ages of 8-18 years old born from IVP
embryos have increased blood pressure and blood glucose levels [137]. Sheep
produced from IVP embryos are heavier by day 61 after birth and several major organs
are larger in size than for offspring produced in vivo [138].

33
Alterations in Maternal Function
An additional line of experimentation to evaluate the importance of the
reproductive tract environment for development of the embryo is to evaluate how
changes in oviductal or uterine function during the preimplantation period affects
embryonic development. Experiments described here indicate that alterations in the
endocrine regulation of the reproductive tract by exogenous administration of hormones
like progesterone and somatotropin can increase embryonic growth and the likelihood
that the embryo will establish pregnancy. Other influences on the reproductive tract, in
particular lactation, can alter the reproductive tract in a way that is detrimental to the
developing embryo.
Supplemental Progesterone
The presence of progesterone during preimplantation embryo development
modulates fertility and elongation of the bovine embryo. The importance of the
reproductive tract for regulating embryonic development can be observed by evaluating
the consequences of hastening the post-ovulation rise in progesterone by providing
supplemental progesterone. Cows receiving supplemental progesterone on Days 1, 2, 3
and 4 of pregnancy have embryos which are developmentally advanced at Day 14
[139], larger in size and have premature IFNT secretion [57, 140].
The mechanism of action is unknown, but is likely to involve changes in
characteristics of the uterine secretome. Forde et al. (2009) conducted a study where
pregnant heifers received either an intravaginal device releasing either high or normal
amounts of progesterone on day 3 after insemination [141]. Tissue was collected from
the endometrium on days 5, 7, 13 or 16 of pregnancy and submitted to microarray
analysis. On day 5 compared to day 7, heifers that received high progesterone had

34
more DEG (36 and 124 respectively) than on day 13 and 16, where heifers that
received high progesterone had only 15 and 25 DEG respectively. The effects of higher
progesterone were greater early in the cycle, when differences in progesterone between
supplemented cows and normal progesterone cows would be greater than at later days
of the cycle. Endometrial tissue collected from high progesterone heifers at day 5 had
increased expression of genes associated with triglyceride synthesis and glucose
transport. One gene that improves glucose secretion, MSTN, increased with high
progesterone treatment. These studies suggest that embryo development is not
increased by progesterone, but that the timeline of development can be shifted by
supplemental progesterone so that embryos reach developmental milestones earlier.
Somatotropin
Another hormone that can improve the competence of the reproductive tract to
support embryonic development is somatotropin. Bovine somatotropin (bST) increases
the systemic and liver production of IGF1, leading to increased milk production [142].
Injections of bST improve pregnancy rates in lactating cows that act as embryo transfer
recipients [143]. An experiment by Ribeiro et al. (2014) showed that injection of bST on
day 0 and 14 following artificial insemination enhanced development of the embryo by
increasing embryonic size at day 34 and 48, decreasing embryonic loss between day 31
and 66, increasing the number of pregnant cows at day 66 and increasing the number
of live calves at birth [144]. The increase in embryonic size was also correlated with an
increase in expression of ISG15 and RTP4 at day 31 and 66, both of which are markers
of IFNT action [145, 146].

35
Lactation
The process of lactation can alter the reproductive tract environment in a way
that compromises embryonic development. One reason lactating cows have decreased
embryonic development during this time is due to corresponding low circulating
progesterone levels [147]. Gene expression in the endometrium differs between
lactating and non-lactating cows at day 17 after estrus in cyclic and pregnant cows
[148]. Several of the upregulated genes in the endometrium of the lactating cow are
involved in immune response and the WNT pathway [148]. That these changes in
endometrial function compromise embryonic development is indicated by results of
embryo transfer experiments. Fewer embryos develop to the blastocyst stage when two
to four-cell embryos were transferred into the oviduct of lactating cows then when
transferred into the oviducts of non-lactating cows [9]. Moreover, pregnancy rates were
lower in lactating recipients when embryo transfer was done at Day 7 after estrus [149-
151].
Developmental Programming
The idea that maternal environment can change the characteristics of
development in a way that affects adult physiology and health is today often called the
Barker hypothesis. This term refers to the scientist who first noted this phenomenon
while studying epidemiological data from adults who were fetuses during the World War
II Dutch famine crisis of 1944-1945 [152, 153]. Caloric intake in Netherlands during that
winter was very low (around 400-800 kcal/d) because of the combination of a harsh
winter and blockade of food delivery by Nazi Germany [152]. A study of middle-aged
adults in the 1990s that were in utero during the famine revealed a variety of changes in
physiology as compared to cohorts born before and after the famine. These changes

36
included increased circulating concentrations of glucose [154], high body mass index
[155], reduced plasma concentrations of factor VII and higher plasma concentrations of
fibrinogen [156].
Since these studies, other epidemiological studies in humans suggest the
importance of nutrition as a programmer of development. For example, adults from
mothers that consumed less protein during pregnancy had higher blood pressure than
those who had a balanced diet [157]. Additionally, mothers that ingested less folic acid
during embryonic development had offspring with an increase rate of congenital
malformations due to defects in the formation of the neural tube [158].
Experimental studies in both laboratory and farm animals have confirmed the role
of prenatal nutrition as a determinant of physiology of the mature animal. Rats fed a low
protein diet during pregnancy had fewer offspring and those offspring had hypertension
at 4 weeks of age [159]. In addition, offspring from rat mothers that were malnourished
during prenatal development had insulinpenia due to a reduction in beta cells [160] and
increased rates of hyperphagia [161]. Caloric restriction from day 28-78 of gestation
resulted in offspring who as six-year adults had decreased insulin sensitivity, increased
body weight and higher feed consumption than sheep from control mothers [162].
Besides nutrition, dehydration, psychological stress and heat stress that are
imposed on the mother can also affect developmental programming of the mature
animal. Sheep that were exposed to hypernatremia from days 110-150 in utero had
offspring with higher plasma osmolality, sodium levels and arterial blood pressure as
neonates [163]. Rhesus monkey mothers that were exposed to unpredictable audible
noise from day 90-145 following conception had offspring with higher levels of

37
circulating ACTH and cortisol as juveniles [164]. Pigs that were exposed to heat stress
in utero whiles fetuses regulated body temperature at a higher set point as adults than
pigs from non-stressed mothers [165].
Developmental programing can occur very early during pregnancy including in
the preimplantation period. Rat mothers that were fed a low protein diet (9% casein)
from fertilization until day 4.25 of gestation had offspring that weighed more at week 4-7
following birth than offspring of mothers fed a control diet (18% casein) [15]. In sheep,
feeding dams a diet deficient in cobalt and sulfur from 30 days before conception
through 6 days following conception resulted in offspring with higher body weight and fat
content as adults than control offspring [16]. Similar finding were found in mice [13].
One characteristic of developmental programming is that the effect can vary with
sex. In one experiment in the rat, four types of offspring were examined at 110 days
after birth 1) from mothers fed low protein (10% casein) during pregnancy and lactation,
2) from mothers fed low protein during pregnancy but a normal diet (20% casein) during
lactation, 3) from mothers fed normal casein during pregnancy but low protein during
lactation and 4) from mothers fed normal protein throughout pregnancy and lactation
[166]. Males but not female offspring from mothers fed low protein during pregnancy
and a normal diet during lactation had insulin resistance compared to controls. In
another experiment, baboon mothers fed a 30% maternal nutrient restricted diet had
male fetuses at mid-gestation with increased renal expression of AT1 compared to
males from control mothers while there was no change in expression for female
offspring [167]. In sheep restricted to 50% of their normal nutrient intake during early to

38
mid-gestation, there was a reduction in renal glomerulus numbers at gestational day
135 in male fetuses but did not affect female fetuses [168].
Sexual dimorphism in developmental programming also occurs during the
preimplantation period. Female offspring from rats that were fed a low protein (9%
casein) diet from day 0-4.25 after mating followed by a normal protein diet for the rest of
gestation weighed less at birth than female offspring fed a normal protein (18% casein)
diet, while male offspring were not affected by the level of protein within the diet [15].
On the other hand, while there was no effect in females, male pups from mothers fed a
low protein diet also had increased systolic blood pressure at weeks 4 and 11 of age
and a higher kidney and lower liver weight compared to the male pups fed the control
diet [15]. Female mouse offspring that were from mothers restricted to a low protein diet
during preimplantation development had higher expression of Igf1r in the retroperitoneal
fat tissue at 1 year of age compared to females from mothers fed a normal protein diet;
In males, however, there was no effect of protein restriction on Igf1r expression [14].
Male mouse offspring born to mothers that were fed a low protein diet (9% casein)
during the pre-implantation period had elevated levels of lung angiotensin-converting
enzyme in lung tissue compared to male offspring from control mothers while there was
no effect of maternal diet on this characteristic in female offspring [17].
In another model, designed to disrupt DNA methylation, rat mothers were fed
methyl-deficient diets that did not contain folic acid from 3 weeks prior to and 5 days
after conception. At 6 months of age, male offspring from these mothers had higher
concentrations of oral glucose than control male offspring while there was no difference

39
in the female offspring [169]. Additionally, sheep mothers that were fed a similar diet
produced offspring with hypertension but only when the sex was male [16].
CSF2 as an Embryokine
Much of this literature review has focused on the evidence that the environment
of the reproductive tract can affect competence of the preimplantation embryo for
development and survival to term. What has not been discussed are the processes by
which the reproductive tract affects embryonic development. The embryo receives all of
its nutrition from the mother through reproductive tract secretions and blood plasma
exudate. The nutritional components of uterine fluid are referred to as histotroph. There
are changes in the composition of the historoph during early pregnancy, including
changes in amino acids, ions, enzymes, hormones, growth factors, proteases and
protease inhibitors, vitamins, glucose, mitogens, lymphokines and cytokines [170]. In
addition to nutritional support, the reproductive tract establishes the pH of the embryo’s
environment as well as providing the epithelial surface to which the embryo will attach
during placentation. The reproductive tract also produces a variety of regulatory
molecules that can affect either endometrial function, other aspects of maternal
physiology or development of the embryo. These include enzymes required for
converting cortisone into cortisol in the sheep [171] and cow [172], prostaglandins,
especially prostaglandin F2 and E2 [173], and a variety of protein growth factors and
cytokines. In the cow, these include IGF2, IGFBP2, PTGER2, VEGFR2 and CST6
[174]. Here we propose the use of the term embryokine to describe regulatory
molecules produced by the reproductive tract that regulate embryonic development.
Several embryokines have been identified that can alter one or more aspects of

40
preimplantation development. In the cow, these include hyaluronan [21], IGF1 [19],
FGF2 [22], LIF [20], TGFB [20] and ILB1 [23].
Role of Colony Stimulating Factor 2 as an Embryokine
Many molecules originally identified as being important for immune function have
since been shown to be produced in the reproductive tract where they could either be
involved in regulation of immune function in the uterus or play a role independent of
regulation of immune function. The best-studied reproductive tract embryokine with
respect to regulation of embryonic development is CSF2. CSF2 is a glycosylated,
monomeric 23-kDa cytokine that was originally described as a product of T lymphocytes
and macrophages, which causes differentiation of granulocytes and macrophages from
hematopoietic stem cells [175-177]. CSF2 is also synthesized by fibroblasts [178]. As
will be described in detail below, CSF2 is also produced by epithelial and stromal cells
of the endometrium [27, 179, 180].
Signal Transduction
CSF2 binds to CSF2R and signals through the JAK2-STAT5ab pathway or the
phosphatidylinositol 3-kinase kinase (PI3K) pathway. Initiation of CSF2 binding begins
first with the attachment of the protein to the trans-membrane receptor colony
stimulating factor alpha (CSF2RA). First described as a protein with 56% homology to
IL-3 by Hayashida in 1990, CSF2RA is an 80 kDa, low-affinity receptor that is specific to
CSF2 [181, 182]. CSF2RA is capable of binding to CSF2 with a dissociation constant of
2-8 nM [182]. There are indications that CSF2 can signal through interactions with
CSFRA alone [183, 184] but the typical signal transduction involves recruitment of the
CSF2RB subunit to the CSF2-CSF2RA complex, which increases the affinity of the
receptor to a dissociation constant to 170 pM [181]. The 120-140 kDa CSF2RB subunit

41
is not specific to CSF2Ra but also serves as part of the receptor complex for IL-3 and
IL-5 [181, 182]. In response to CSF2RB binding, CSF2RA expression is downregulated
[185]. CSF2RA is expressed in the mouse, human, bovine and porcine embryos [186-
189].
Upon binding to CSF2, the CSF2R complex activates Janus kinase (JAK) 2
which in turn causes activation of signal transducer and activator of transcription (STAT)
5 A and B and other Src-homology 2 domain containing proteins [181, 190, 191]. JAK2
associates with the CSF2RB subunit where it is activated and subsequently
phosphorylates tyrosine sites located on the CSF2RB subunit [190, 192].
Phosphorylation of Y612, Y695 and Y750 of CSF2RB subunit allow for binding of
STAT1 and STAT5, with STAT5 being the most critical for CSF2 [191].
Production of CSF2 by the Reproductive Tract
CSF2 has been localized immunochemically to the oviduct and endometrium of
the human [29], cow [27], pig [193] and mouse [194]. In the mouse, CSF2 protein has
been localized to glandular and luminal epithelial cells, while the contribution from the
stroma is minimal; intensity of labeling increases following the post-ovulatory increase in
estrogen [28], which stimulates the secretion of CSF2 while progesterone inhibits it
[194]. Mating can increase the release of CSF2 into uterine fluid through the actions of
TGFB contained in the seminal fluid [33]. In addition, expression of CSF2 in the oviduct
is also decreased following mating with males whose seminal vesicle gland was
surgically impaired [32].
In the human, luminal and glandular epithelium of the endometrium and stroma
express CSF2 mRNA and protein [195]. Glandular epithelial cells have the highest level
of protein and mRNA expression [29, 180]. Secretion of CSF2 from luminal and

42
glandular epithelial cells is highest during the mid-secretory phase of the menstrual
cycle before decreasing during the proliferative phase [195]. However, CSF2 is present
in significantly large quantities in the decidua throughout the first trimester of pregnancy
compared to non-pregnant decidua [196].
In the cow, CSF2 can be localized immunochemically to the luminal and
glandular epithelium and stroma [27, 197]. The majority of CSF2 is localized to the
luminal epithelium [27]. Although not significant, immunoreactive CSF2 in the
endometrium during the estrous cycle had low amounts of CSF2 at estrus (when
progesterone concentrations are low) compared to days 7-18 of the estrous cycle [27].
Therefore, it appears that CSF2 secretion in the cow could be under the control of
progesterone [27].
Another molecule that can modify maternal production of CSF2 in the uterus in
ruminant is IFNT. Transcervical infusions of IFNT into the uterine body increased
amounts of CSF2 in the luminal epithelium [197]. The level of obesity of the cow also
determines CSF2 immunoreactivity and CSF2 mRNA expression in the ampulla: it was
suppressed in the oviduct of obese cows compared to lean cows on the second day
following ovulation [34].
Actions on the Preimplantation Embryo
The first observed action of CSF2 on the preimplantation embryo was an
increase in the proportion of cultured embryos that could develop to the blastocyst
stage. Such a phenomenon has been observed in the cow [18, 24], pig [189, 198, 199],
mouse [26] and human [200]. The mechanism for the increased competence of
embryos to develop to the blastocyst stage is not known. However, in both the mouse
blastocyst [187] and cow morula [31], CSF2 alters expression of genes in a way that

43
would block apoptosis. In the mouse, for example, CSF2 decreased expression and
abundance of genes involved in stress response and apoptosis, such as Hspa5,
Hsp90aa1, Hsp90ab1 and Gas5, and proteins HSP1A/1B and BAX. In cattle, CSF2
increased transcript abundance of several anti-apoptotic genes, such as CD73/NT5E,
PRKAR2B and PGR and decreased the abundance of pro-apoptotic genes such as
MADD, RIPK3, NOD2 and CREM [31]. Furthermore, treatment with CSF2 also
decreased the magnitude of apoptosis in the bovine embryo after exposure to heat
stress [31]. In addition, treatment with CSF2 prior to freezing improved post-thaw
survival of mouse embryos while preventing apoptosis [187, 201]. Thus, CSF2 may
increase competence for development to the blastocyst stage through inhibition of
apoptosis.
CSF2 could also act by changing expression of genes involved in development.
Among the genes whose expression was altered in the bovine morula were genes
associated with neurogenesis, muscle formation, mesenchyme formation and multiple
signaling pathways [31]. In the pig, CSF2 altered gene expression in cloned embryos
with increases in expression of LIF, CDX2, POU5F1, BCL2, DNMT1 and PCNA [198,
199]. In cattle, CSF2 can increase the blastocyst yield regardless of whether it is added
after fertilization or if addition is delayed until day 5 of development [18, 24]. Therefore,
actions of CSF2 probably occur in later cleavage stages and after embryonic gene
activation has occurred.
Not only does CSF2 increase blastocyst yield but also embryos cultured with
CSF2 have increased likelihood of establishing pregnancy after transfer to recipients. In
the mouse, the addition of 2 ng/ml recombinant murine CSF2 to cultured embryos of

44
that were subsequently transferred to recipients resulted in increased litter size and
more viable progeny compared pregnancies after transfer of control embryos in culture
[26]. In the cow, pregnancy rate and calving rate was higher for recipients receiving an
in vitro produced embryos cultured with 10 ng/ml CSF2 from day 5-7 of development
(43% and 37%, respectively) than cows that received control embryos (34% and 23%)
[18]. The effect of CSF2 was only seen when added during days 5-7 of development.
Embryos cultured with CSF2 from days 1-7 did not have superior ability to survive
transfer (35% vs. 35% for controls). An interesting observation was that CSF2 also
reduced the loss of pregnancies occurring after initial pregnancy diagnosis at Day 30-35
of gestation. Pregnancy losses were 22% for control embryos, 11% for embryos treated
with CSF2 from Day 5-7 and 0% for embryos cultured from Day 1-7. [18].
Recently, a similar beneficial effect of CSF2 on embryo competence for survival
after transfer was seen in humans. Treatment of embryos with 2 ng/ml CSF2 throughout
culture increased survival compared to controls at week 7 (23.5% vs. 20%) and 12 of
gestation (23% vs. 18.7%) as well as live births (28.9% vs. 24.1%) [25]. The ongoing
implantation rate in women that previously had one or more miscarriages was also
greater if the embryo transferred had been cultured with CSF2 [25].
The change in the embryo responsible for increased competence to establish
pregnancy is not clear. One possibility is that CSF2 increases number of cells in the
blastocyst, particularly the ICM, and that these increased numbers make the embryo
better able to survive transfer. Culture of mouse embryos with CSF2 resulted in
blastocysts that contained 15 more cells than control blastocysts [186]. When 2-cell
stage embryos were split and allowed to develop to the blastocyst stage, diameter and

45
number of cells of the blastocyst were increased by CSF2 [202]. In the pig, as well,
CSF2 improved total cell number in blastocysts [199]. In both the human [187] and cow
[18], number of cells in the ICM of blastocysts was increased by CSF2. There was also
a reduction in the number of cells undergoing apoptosis in human blastocysts treated
with CSF2 [187].
There is also evidence that treatment with CSF2 can alter the developmental
program of the embryo at a later stage of pregnancy. In one experiment, bovine
embryos were treated with CSF2 from days 5-7 of development, transferred into cows
and then recovered at day 15 of development. Embryos that had been treated with
CSF2 had increased expression of IFNT2 and KRT18, increased IFNT2 secretion into
the uterine lumen and tended to have increased length [30].
Not all studies with CSF2 indicate positive effects of the molecule on embryonic
development. In the one study, the addition of 2, 5, 10 or 50 ng/ml of ovine CSF2 to
cultured bovine embryos did not improve the number of blastocysts that developed at
day 7 after fertilization [203]. In a similar experiment, ovine embryos cultured in the
presence of CSF2 did not reach the blastocyst stage at a rate higher than that of control
embryos. In the mouse, addition of either 5 or 10 ng/ml CSF2 decreased the proportion
of embryos that developed to the blastocyst stage and, although not significant, tended
to increase the rate of aneuploidy [204].
Characteristics of the CSF2R in the Preimplantation Embryo
Although CSF2 can affect embryonic function in a variety of species, it may exert
its actions through a cell signaling mechanism that is distinct from the prototypical signal
transduction pathway initiated by the CSF2RA-CSF2RB complex. This is because, as
has been shown for human and mouse, the preimplantation embryo expresses

46
CSF2RA but not CSF2RB [186, 187]. In the pig, embryos at the blastocyst cell stage
express CSF2RB, but transcript abundance is very low compared to that for CSF2RA
[189].
Other pathways can be activated following the binding of CSF2 to receptor, such
as the MAPK and the PI3K pathway [191, 205, 206]. When CSF2 signaling through
PI3K occurs, the regulatory subunit of PI3K (p85), which is bound to CSFRA through its
C-SH2 domain, is phosphorylated and activates PI3K [183]. CSF2 signaling is blocked
in Day 12 porcine TE cells when treated with the PI3K inhibitor LY294002 [206].
One possibility is that CSF2RB protein is present on the plasma membrane of
embryonic cells despite lack of mRNA. This could be the case because bovine oocytes
contain CSF2RB in trace amounts [188, 189] and some protein that was synthesized by
the oocyte could persist after fertilization.
Goals of the Current Investigation
The aim of this dissertation is to understand the mechanisms responsible for
specific actions of a maternally-derived cytokine, CSF2 on development of the
preimplantation bovine embryo. In cattle, CSF2 treatment from day 5-7 of development
can increase the proportion of embryos becoming a blastocyst, alter gene expression,
decrease apoptosis, increase ICM number, program the embryo to have altered gene
expression and increased trophoblast elongation at day 15 of gestation, and increase
competence of the embryo to maintain pregnancy to term [18, 24, 30, 31]. This
dissertation is focused on understanding key aspects of the biology of some of these
effects of CSF2 as follows:
In Chapter 2, experiments will be described that determine whether CSF2
regulates survival and pluripotency of the ICM or the growth potential and differentiation

47
of the TE. The mechanism by which CSF2 improves the ability of a blastocyst to
establish pregnancy is unknown, but evidence exists that CSF2 affects lineage
commitment of the ICM and TE of the blastocyst in addition to altering expression of
pluripotency and differentiation related genes [18, 31]. In Chapter 3, the nature of the
developmental programming of CSF2 to alter trophoblast elongation and gene
expression at day 15 [30] will be elucidated. In particular, it will be tested whether CSF2
from day 5-7 alters the transcriptome and methylome at day 15. In addition, an
unexpected finding will be described that indicates the nature of developmental
programming by CSF2 depends upon sex, i.e., CSF2 causes different and often
opposite effects in female embryos than male embryos. In Chapter 4, the
developmental pattern of DNA methylation in the preimplantation embryo will be
described as well as possible alterations in the global methylome by CSF2 and sex that
could form the basis for sexual dimorphism in response to CSF2 seen in Chapter 3.

48
CHAPTER 2 REGULATION OF PLURIPOTENCY OF INNER CELL MASS AND GROWTH AND
DIFFERENTIATION OF TROPHECTODERM OF THE BOVINE EMBRYO BY COLONY STIMULATING FACTOR 2
Introduction
Communication between the preimplantation mammalian embryo and
reproductive tract of the mother is important for ability of the embryo to develop into a
blastocyst with the ability to develop to term. Even though embryos can become
blastocysts in vitro, they have reduced competence for establishing pregnancy when
transferred to recipient females than embryos that developed in vivo [26, 207, 208].
Growth factors and cytokines are important components of the embryotropic milieu of
the reproductive tract. Addition of specific growth factors present in the reproductive
tract to culture medium can improve the proportion of embryos that develop to the
blastocyst stage. Examples in the cow include activin A, EGF, FGF2, IGF1, IGF2, IL1B,
and LIF [20, 22, 23, 209-211]. Moreover, competence of in-vitro produced embryos to
establish pregnancy after transfer into females can be improved by LIF and TGFB in the
mouse [212-214] and IGF1 in the cow (when recipients were heat-stressed) [129, 215].
Perhaps the best studied maternally-derived cytokine that regulates embryonic
development is CSF2. Secreted by the oviduct and endometrium [27, 29, 34], CSF2 can
increase the proportion of embryos developing to the blastocyst stage in the cow [18,
24], pig [189, 198, 199], mouse [26] and human [200]. Moreover, CSF2 increased the
proportion of in vitro produced embryos that established pregnancy after transfer into
mice [26], cows [18] and humans [25]. That variations in CSF2 signaling might be a
cause of female infertility is suggested by the recent observation that CSF2 synthesis in
the oviduct is decreased in obese cows [34].

49
The mechanism by which CSF2 improves the ability of a blastocyst to establish
pregnancy is not known but there is evidence for the hypothesis that CSF2 affects
lineage commitment in both the ICM and TE of the blastocyst. Treatment of embryos
with CSF2 at Day 5 post-insemination altered expression of several genes involved in
pluripotency and differentiation [31]. In addition, CSF2 increased the number of ICM
cells in the bovine blastocyst at Day 7 post-insemination without affecting number of TE
cells [18]. At Day 15, the trophoblast of conceptuses formed following treatment with
CSF2 from Day 5-7 post-insemination had higher expression of the maternal signaling
gene IFNT2 and tended to be numerically greater in length than control embryos [30].
Some actions of CSF2 on embryo survival could be the result of regulation of apoptosis
pathways because CSF2 affects expression of genes involved in apoptosis in both
cattle [31] and mice [216] and can block the induction of apoptosis caused by heat
shock in cattle [31].
In most cells, CSF2 signaling is accomplished through a cell surface receptor
composed of a low-affinity subunit (CSF2RA) and a high-affinity subunit (CSF2RB)
[217]. CSF2RA is a low-affinity receptor specific for CSF2 while CSF2RB, which is
involved in signaling for several cytokines, confers increased affinity of the receptor
complex to the ligand [181]. CSF2RA is expressed in preimplantation mouse, pig and
human embryos but expression of CSF2RB is undetectable or nearly so [186, 187,
189]. These observations suggest that CSF2 signaling in the embryo involves an
unknown pathway that is independent of CSF2RB.
The major objective of this study was to determine whether CSF2 could regulate
survival and pluripotency of the ICM and growth potential and differentiation of TE. To

50
examine pluripotency, we utilized a model to evaluate ability of isolated ICM to remain
pluripotent while being cultured and passaged on a layer of fetal fibroblasts in the
presence of MAPK1 and GSKB inhibitors [218]. It was also tested whether CSF2
regulated expression of genes involved in pluripotency (NANOG and SOX2 [46, 219]),
differentiation of TE (CDX2 [46]) and formation of hypoblast (GATA6 [46]) in isolated
ICM. TE growth and differentiation was assessed in a model of TE outgrowth on
Matrigel-coated plates [220]. It was also determined whether both CSF2RA and
CSF2RB were expressed by the preimplantation embryo from the zygote to blastocyst
stage.
Materials and Methods
Embryo Production
IVP of bovine embryos was performed as previously described [22] unless
otherwise noted. Briefly, COCs were obtained by cutting the surface of slaughterhouse-
derived ovaries with a scalpel and vigorously rinsing the ovary through a bath of oocyte
collection medium [tissue culture medium-199 with Earle’s salts without phenol red
(Hyclone, Logan, UT, USA), 2% (v/v) bovine steer serum (Pel-Freez, Rogers, AR), 2
U/ml heparin, 100 U/ml penicillin-G, 0.1 mg/ml streptomycin and 1mM glutamine].
Groups of 10 COCs were matured in 50 μl droplets of oocyte maturation medium [tissue
culture medium-199 with Earle’s salts (Invitrogen, Carlsbad, CA, USA), 10% (v/v)
bovine steer serum, 2 g/ml estradiol 17-β, 20 g/ml bovine follicle stimulating hormone
(Bioniche Life Sciences, Belleville, Ontario, Canada), 22 g/ml sodium pyruvate, 50
g/ml gentamicin sulfate and 1 mM glutamine] covered with mineral oil for 20 hours at
38.5oC and in a humidified atmosphere of 5% (v/v) CO2. Up to 300 matured oocytes

51
were fertilized with Percoll-purified sperm (1.0 x 106/ml) for 8 hours at 38.5ºC in 1.7 ml
of synthetic oviductal fluid-fertilization (SOF-FERT) [221]. Cumulus cells were denuded
after fertilization by vortexing in 600 μl HEPES-TALP containing 1,000 U/ml
hyaluronidase. Putative zygotes were then cultured in 50 µl microdrops of synthetic
oviduct fluid - bovine embryo 1 (SOF-BE1) [22] covered with mineral oil at 38.5oC in a
humidified atmosphere of 5% (v/v) O2 and 5% (v/v) CO2 with the balance N2. Cleavage
rate was assessed at Day 3 post-insemination. Blastocysts were harvested at Day 7 or
8 post-insemination.
Effect of CSF2 on Survival of Isolated Inner Cell Mass
Bovine fetal fibroblast feeder cells
Culture on bovine fetal fibroblasts prevent isolated ICM from losing pluripotency
and undergoing differentiation [218]. Bovine fetal fibroblast (BFF) cells were created
from fetal calf skin (~2-3 months old). Pieces of fetal calf skin (~1 cm3) were cut from
the mediolateral region of the body and transferred into 15 ml tubes of Dulbecco’s
phosphate buffered saline (DPBS) with 1% (v/v) antibiotic-antimycotic [10000 U/ml
penicillin G, 10000 μg/ml streptomycin sulfate and 25 μg/ml amphotericin B] (ABAM)
(Invitrogen). Rinsed fetal calf skin was then placed onto 60 mm petri dishes (BD
Falcon, Franklin Lake, NJ, USA), cut into 5 mm x 5 mm square pieces, and placed onto
scored areas of a new 60 mm petri dish containing 10 ml of a medium consisting of a
modified Dulbecco’s Modified Eagle Medium (DMEM) (Invitrogen) containing 10% (v/v)
fetal bovine serum [FBS] (Atlanta Biologicals, Norcross, GA, USA) and 1% (v/v) ABAM.
On Day 4 after culture, skin pieces were removed from the plate using tweezers. Cells
were passaged by removal of medium by aspiration, washing twice with DPBS
containing 1% (v/v) ABAM, and trypsinization by addition of 5 ml 0.05% (v/v) trypsin with

52
0.38 mg/ml ethylenediamine tetraacetic acid (EDTA) (Invitrogen) and incubation at
38.5°C and 5% (v/v) CO2 in air for 3 min. Trypsin was inhibited by addition of 1.5 ml
FBS. Cells were centrifuged for 5 min at 250 x g, supernatant removed and cells
resuspended with 25 ml modified DMEM. Cells were then transferred into a 175 cm2
flask and cultured until confluent. Medium was changed every two days until 90-100%
confluence was reached. Cells were then trypsinized and passaged at a 1:4 dilution.
For culture of ICM, BFF cells at 90% confluence were treated twice, for 3 hours
each, with 10 µg/ml mitomycin C (Invitrogen) at 38.5oC in a humidified atmosphere of
5% (v/v) CO2 in air. After three washes in medium, cells were cultured overnight,
washed with DPBS with 1% (v/v) ABAM, trypsinized as described above, washed,
collected by centrifugation for 5 min at 250 x g, and resuspended to 1.5 x 106 cells/ml in
modified DMEM that also included 10% (v/v) dimethyl sulfoxide (DMSO) (Sigma-Aldrich,
St. Louis, MO, USA). Cells were frozen in liquid nitrogen in 1 ml aliquots until further
use.
Isolation of ICM by lysis of trophectoderm using antibody and complement via immunosurgery
Procedures were modified from those described by Ozawa et al. [218].
Blastocysts were washed three times in DPBS containing 1% (w/v) polyvinylpyrrolidone
(PVP) (Kodak, Rochester, NY, USA) (DPBS-PVP), treated with 0.1% (w/v) proteinase
solution (proteinase from Streptomyces griseus) in DPBS to remove the zona pellucida,
washed three times in DMEM (Invitrogen) with 1% (v/v) ABAM, washed twice in DMEM
with 1% (v/v) ABAM and 10% (v/v) rabbit anti-bovine serum (Sigma-Aldrich) and then
incubated for 1.5 hours at 38.5oC in a humidified atmosphere of 5% (v/v) O2 and 5%
(v/v) CO2 with the balance N2. Blastocysts were then washed three times in DMEM with

53
1% (v/v) ABAM, washed twice in DMEM with 1% (v/v) ABAM and 20% (v/v) guinea pig
complement (Rockland Immunochemicals Inc., Gilbertsville, PA, USA), and then
incubated in the same medium for 1.5 hours at 38.5oC in a humidified atmosphere of
5% (v/v) O2 and 5% (v/v) CO2 with the balance N2 to lyse TE. Those ICM that were free
of visible TE cells were washed three times in embryonic stem cell medium (ESM)
consisting of knockout-DMEM (Invitrogen) with 15% (v/v) knockout serum replacement
(Invitrogen), 1 mM Glutamax (Sigma-Aldrich), 1% (v/v) ABAM, 0.1 mM minimal
essential medium nonessential amino acids, and 0.1 mM β-mercaptoethanol. Isolated
ICM were then individually transferred into a single well of a 12-well plate (BD Falcon)
coated 24 hours earlier with 1% (w/v) gelatin (BioRad, Hercules, CA, USA) and 2 x 104
cells/cm2 mitomycin C treated fibroblasts in 1 ml of ESM+2i medium [ESM containing 3
μM GSK3B inhibitor CHIR99021 (Stemgent, San Diego, CA, USA) and 1.2 μM MEK1
inhibitor PD0325901 (Stemgent)]. These conditions were used because they promote
maintenance of ICM in a pluripotent state [218].
Culture of isolated ICM
Individual clusters of ICM cells (each from a single blastocyst) were cultured at
38.5oC in a humidified atmosphere of 5% (v/v) O2 and 5% (v/v) CO2 with the balance
N2. On Day 7 following ICM isolation, ICM colonies were passaged using a finely drawn
glass needle to separate the ICM from any surrounding TE that remained and to
dislodge the ICM outgrowth from the BFF layer. Individual ICM outgrowths were then
transferred into a well of a new 12-well plate prepared with gelatin and mitomycin C
inactivated BFF cells as previously described and cultured in 1 ml ESM+2i. Medium
was changed and cells were passaged according to the same procedure above. For
some IVP replicates (i.e, a group of embryos produced on a given day), culture was

54
terminated 7 days after passage 1 while, in others, cells were passaged three times and
cultures stopped at the end of passage 3.
Experiments
In the first experiment, all embryos were moved at Day 6 post-insemination in
groups of 30 to 50 µl microdrops of either 1) 45 µl SOF-BE1and 5 µl recombinant
bovine CSF2 [Novartis, Basle, Switzerland; 100 ng/ml in a 90:10 (v/v) mixture of SOF-
BE1 and DPBS containing 1% (w/v) bovine serum albumin (BSA); final concentration of
CSF2 after dilution in the drops = 10 ng/ml], 2) 45 µl SOF-BE1 and 5 µl vehicle [90:10
SOF-BE1: DPBS containing 1% (w/v) BSA; control treatment) or 3) 50 µl ESM+2i. The
concentration of CSF2 was chosen based on its effectiveness for increasing embryonic
survival after transfer into recipient females [18]. The ESM-2i medium was used as a
positive control for survival of ICM colonies because it had previously been reported to
enhance survival of ICM colonies when used to culture embryos from Day 6-8 post-
insemination [218]. Embryos were harvested at Day 8 post-insemination and ICM
isolated and used for culture as described above. The experiment was replicated on 5
occasions with a total of 12-16 isolated ICM per treatment.
In a second experiment, procedures were similar except all embryos were
harvested on Day 5 post-insemination and cultured with treatments from Day 5 to 7
post-insemination before ICM isolation. The experiment was replicated on 6 occasions
with total experiments of 8-11 isolated ICM per treatment.
Analysis of ICM Colonies Surviving Passage
One colony derived from CSF2-treated Day 7 blastocysts was harvested 7 days
after passage 3, placed in extraction buffer (Arcturus PicoPure RNA isolation kit, Life

55
Technologies, Grand Island, NY, USA) and measured for steady-state amounts of
NANOG, CDX2 and GAPDH by quantitative PCR (qPCR) as described below.
Expression of Pluripotency Genes in ICM
All embryos were moved at Day 6 post-insemination in groups of 30 to 50 µl
microdrops of either 45 µl SOF-BE1 and 5 µl CSF2 (prepared as described above to
yield a final concentration of 10 ng/ml) or 45 µl SOF-BE1 and 5 µl vehicle (control
treatment). Blastocysts were harvested at Day 8 and ICM were isolated by
immunosurgery on Day 8 post-insemination as described above. ICM were immediately
placed in extraction buffer (Life Technologies) and stored at -80°C until ready for RNA
extraction. RNA was extracted and used to measure steady-state amounts of NANOG,
SOX2, CDX2, GATA6 and GAPDH. For each replicate, a total of 3-5 ICM were pooled
for an individual treatment. The experiment was replicated using a total of 7-9 pools per
procedure.
Actions of CSF2 on Competence of Trophectoderm to Form Outgrowths
Establishment of outgrowths
The procedure for analysis of TE outgrowth was modified from that of Yang et al.
[222]. On Day 5 or 6 post-insemination, all embryos in groups of 30 were moved to 50 µl
microdrops of either 45 µl SOF-BE1 and 5 µl CSF2 (prepared as described above to a
final concentration of 10 ng/ml) or 45 µl SOF-BE1 and 5 µl vehicle (control treatment).
After culture for 48 hours at 38.5oC in a humidified atmosphere of 5% (v/v) O2 and 5%
(v/v) CO2 with the balance N2, blastocysts were collected and transferred individually to
wells of 96-well plates (BD Biosciences, Bedford, MA USA) that had been coated with
Matrigel (Matrigel basement membrane matrix growth factor reduced; BD Biosciences).
Embryos were cultured in 100 µl of DMEM (Invitrogen) modified to contain 10% (v/v)

56
FBS (Atlanta Biologicals), 1% (v/v) ABAM, 0.1 mM MEM nonessential amino acids and
0.1 mM β-mercaptoethanol at 38.5oC in a humidified atmosphere of 5% (v/v) O2 and 5%
(v/v) CO2 with the balance N2. After 5-6 d, medium was removed and replenished with
fresh medium. At the end of culture, outgrowths were examined for size, labeling with
anti-CDX2, and gene expression.
Immunolabeling of CDX2
All labeling steps were performed at room temperature unless otherwise stated
and in a volume of 100 µl in the wells in which outgrowths were established. Attached
TE outgrowths were washed in DPBS-PVP and fixed in 4% (w/v) paraformaldehyde in
DPBS-PVP for 15 min. The fixed outgrowths were washed three times in DPBS-PVP,
permeabilized for 20 min in DPBS containing 0.25% Triton X-100 and washed three
times in 0.1% (v/v) Tween 20 with wash buffer [DPBS containing 10 mg/ml fraction V
BSA (Sigma-Aldrich)] followed by three washes with wash buffer. For immunolabeling,
nonspecific binding sites were blocked by incubation with DPBS containing 5 mg/ml
BSA for 1 h. Outgrowths were incubated with affinity-purified mouse monoclonal
antibody against CDX2 (ready to use solution; BioGenex, San Ramon, CA, USA). As a
negative control, anti-CDX2 was replaced with an irrelevant mouse IgG antibody
(Sigma-Aldrich, St. Louis, MO, USA). After 1 h, outgrowths were washed three times in
wash buffer and transferred to 1 μg/ml fluorescein isothiocyanate conjugated anti-
mouse IgG (Abcam, Cambridge, MA, USA). After three washes, nuclei were labeled by
incubation in 5 µg/ml Hoechst 33342 (Sigma-Aldrich) in DPBS-PVP for 15 min.
Outgrowths were washed three times and mounted using ProLong Gold Anti-Fade
mounting medium (Invitrogen) and observed under a Zeiss Axioplan epifluorescence
microscope (Zeiss, Göttingen, Germany). Images were acquired using a 40x objective

57
and green and blue filters. The exposure times were constant for all outgrowths
analyzed in an individual replicate.
Assessment of establishment and growth of trophectoderm outgrowths
The ability of embryos to attach to the Matrigel-coated well was determined for
each IVP replicate as the fraction and percent of embryos that remained attached to the
well after aspiration of medium at Day 13 or 15 post-insemination. Similarly capacity for
formation of TE outgrowths was calculated within IVP replicate as the percent of
blastocysts in which outgrowth could be observed microscopically at either Day 13 or 15
post-insemination. Outgrowth surface area was determined with the aid of a microscope
graticule at 40x magnification.
Gene expression
At Day 15 post-insemination, steady state expression of CDX2 and GATA6 was
determined for a subset of TE outgrowths. Medium was removed, cells washed three
times with DPBS-PVP and the outgrowth removed by adding 100 μl of extraction buffer
(Applied Biosystems, Invitrogen) and pipetting multiple times. For each replicate, pools
of 2-10 TE outgrowths were prepared. The total number of replicates varied from 6-7
depending on the experiment. RNA extraction, DNase treatment and reverse
transcription were completed as described below.
Antiviral assay
The ability of samples of conditioned medium to inhibit vesicular stomatitis virus-
induced death of Madin-Darby bovine kidney cells by 50% was compared with the
activity of recombinant human IFNA standard (3.84x108 IU antiviral activity/mg protein;
(Millipore, Billerica, MA, USA) using a previously-described assay [222]. Assay
sensitivity was 2.6 pg of IFNA standard. The antiviral activity of conditioned medium

58
was adjusted based on this standard to represent the IU activity per mm2 of cell
outgrowth.
Experiments
A total of 4 experiments were conducted. In two experiments, embryos were
treated with 10 ng/ml CSF2 or control from Day 6 to 8 post-insemination. This time
interval was chosen because CSF2 treatment during this time increased capacity of the
ICM to survive culture after isolation from the TE (See Results). In two experiments,
embryos were cultured from Day 5 to 7 post-insemination. This time interval was
chosen because CSF2 treatment during this time increased competence of embryos to
survive after transfer to females [18]. For each treatment period, one experiment was
conducted in which outgrowths were examined at Day 13 post-insemination (i.e., 5-6
days after culture on Matrigel) and one experiment was conducted in which outgrowths
were examined at Day 15 post-insemination (i.e., 7-8 days after culture). We examined
two separate culture periods because of the possibility that effects of CSF2 on TE
characteristics would depend on duration of culture. Each experiment was performed
with 7-15 IVP replicates containing 2-26 embryos per treatment for each replicate. The
replication is described in Table 2-1.
Expression of CSF2 Receptor Subunit Genes
Analysis of gene expression was performed by qPCR using five pools of RNA
extracted from 25-30 embryos/pool at the zygote (0 hpi), 2 cell (32-40 hpi), 3-4 cell, 5-8
cell (48 hpi), 9-16 cell (72 hpi), morula (120 hpi) and blastocyst (168 hpi) stages. Zona
pellucidae were removed and oocytes and embryos processed for RNA extraction,
DNase treatment and reverse transcription as described below. To verify effectiveness

59
of primers, total RNA was also extracted from leukocytes collected from a saphenous
vessel of a Holstein cow.
Quantitative PCR
Total RNA was extracted using the PicoPure RNA isolation kit (Life
Technologies), treated with 2 U of DNase (New England Biolabs, Ipswich, MA, USA) at
37°C for 30 min to remove DNA, and then incubated at 75°C for 15 min to denature
DNase. DNase-treated RNA was then reverse-transcribed using the High Capacity
cDNA Reverse Transcription Kit (Invitrogen). Reverse transcription occurred using
random hexamer primers and involved incubation at 25°C for 10 min, 37°C for 120 min
and 85°C for 5 min. The cDNA was stored at -20°C until further use. Negative controls
for real-time PCR were also performed by incubation without reverse transcriptase.
cDNA was utilized for real-time PCR analysis. For detection of transcript levels,
a CFX96 Real-Time PCR detection System (Bio-Rad, Hercules, CA, USA) was utilized
with SsoFast EvaGreen Supermix with Low ROX (Bio-Rad). Each reaction contained 1
µl forward primer (0.5 μM), 1 µl reverse primer (0.5 μM), 10 µl Evagreen Supermix (Bio-
Rad), 6.8 µl H2O and 1.2 µl cDNA sample (0.6 embryo equivalent). Amplification
conditions were: 95°C for 30 sec, 40 cycles at 95°C for 5 sec, 60°C for 5 sec, and 1
cycle of melt curve analysis at 65-95°C in increments of 0.5°C every 2 sec. Primers for
NANOG, SOX2, CDX2, GATA6, SDHA, and YWHAZ were described and validated
previously [218, 223]. Primers for CSF2RA (F: 5’-
ACGCCGGCCTAAATGTGAAGTTTG-3’, R: 5’- ACGTGCACTGACACTCACTGTCTT-3’,
XM_002686598.2) and CSF2RB (F: 5’- TTCCCAAGAGCTGATGACATGGGT-3’, R: 5’-
AGATGATGCGGCTGGTGTAGTCAT-3’, NM_001192664.1) were designed using
PrimerQuest (Integrated DNA Technologies, Coralville, IA, USA) and validated by

60
evaluation of melt curves, demonstration of standard curves that exhibited a slope of
between -3.0 to -3.5, analysis of amplicon size by agarose gel electrophoresis, and
Sanger sequencing of amplicons to confirm identity. The ΔCT value was determined by
subtracting the CT value of the sample by the CT for GAPDH, or for the experiment on
CSF2RA and CSF2RB expression, by subtracting the CT value of sample by the
geometric mean of the CT for three housekeeping genes, SDHA, GAPDH and YWHAZ
[224].
Statistical Analysis
Most data were analyzed by least-squares analysis of variance using the GLM
procedure of the Statistical Analysis System (SAS) version 9.3 (SAS Institute Inc., Cary,
NC, USA). The exception was for data on survival of ICM after passage, which was
analyzed by the GLIMMIX procedure of SAS using the Binomial feature. Depending on
the experiment, main effects included IVP replicate, stage, treatment and day of
treatment. Interactions were also included. The IVP replicate was considered random.
For real-time PCR results, treatment effects were tested using ΔCT values and graphed
as either fold-change (2-ΔΔCt) relative to control or as the inverse of ΔCT. Data are
shown as least-squares means ± standard error of the means.
For the experiments, a total of 62 IVP replicates were performed in which
embryos were treated with CSF2 from Day 5-7 and 36 IVP replicates in which embryos
were treated from Day 6-8. Statistical analysis was performed to determine whether
response to CSF2 depended on the competence of control embryos in an individual IVP
replicate to develop to the blastocyst stage. Each IVP replicate was classified as to
whether blastocyst development in the control group was within the lowest, middle or

61
highest tercile of percent blastocyst. For Day 5-7, percent of oocytes that became
blastocysts in the control group varied between 10.6-24.9% for the lowest tercile, 25.0-
30.4% for the middle tercile and 30.6-46.8% for the highest tercile. For Day 6-8, ranges
were 12.5-22.7%, 22.9-35.7% and 36.4-60% for the three terciles, respectively. Data
were analyzed as described in the previous paragraph with the inclusion of blastocyst
development class (low, middle and high) and interactions with development class.
Results
Blastocyst Development
A total of 62 embryo culture procedures were performed in which embryos were
cultured in the presence or absence of 10 ng/ml CSF2 from Day 5 to 7. There was no
significant difference between treatments in the proportion of oocytes (i.e., putative
zygotes) that became blastocysts at Day 7 post-insemination (26.9% ± 0.7 for CSF2 vs.
28.3% ± 0.7 for control, P=0.16). However, the effect of CSF2 varied depending on the
percent of oocytes that became blastocysts in the control group. In particular, when
procedures were ranked according to percent blastocyst development in controls and
classified into low, medium and high terciles, there was an interaction between
treatment and development class (P<0.0001). CSF2 increased the proportion of
oocytes that became blastocysts in IVP replicates where blastocyst development in the
control group was in the lowest tercile, had no effect for IVP replicates in the middle
tercile and had negative effects on development when IVP replicates were in the
highest tercile (Figure 2-1A). Moreover, the correlation between the percent of oocytes
that became blastocysts in the control group and the deviation in percent blastocyst
caused by CSF2 was -0.53 (P<0.0001).

62
A total of 36 embryo culture procedures were performed in which embryos were
cultured + 10 ng/ml CSF2 from Day 6 to 8. There was no significant difference between
treatments in the percent of oocytes that became blastocysts at Day 8 post-
insemination (30.8% ± 1.3 for CSF2 vs. 30.1% ± 1.3 for control, P=0.69) and no
interaction between CSF2 treatment and development class. Nonetheless, there was a
correlation of -0.37 between the percent of oocytes that became blastocysts in the
control group and the deviation in percent blastocyst caused by CSF2 (P=0.026) (Figure
2-1B.
Effect of CSF2 on Survival of Isolated ICM
Previously, it was shown that a proportion of ICM isolated from blastocysts could
be maintained in a pluripotent state when cultured on mitocmyin-C treated fetal
fibroblasts in the presence of the GSKB inhibitor CHIR99021 and the MEK1 inhibitor
PD0325901 [218]. Here we tested whether exposure of embryos during the 32-cell to
blastocyst stages of development to CSF2 could improve the rate at which isolated ICM
persisted in a pluripotent state after repeated passage. In the first experiment, embryos
were treated from Day 6 to 8 post-insemination with either control, CSF2 or ESM+2i.
The lattermost treatment was previously reported to increase competence of ICM to
remain pluripotent when used for culture from Day 6 to 8 [218]. The proportion of ICM
colonies that survived until the end of passage 1 was greater for ICM derived from
blastocysts produced in the presence of CSF2 than ICM from control blastocysts
(P=0.002) (Figure 2-2A). In contrast, ICM from blastocysts cultured in ESM+2i had
survival that was not different from controls. Of the ICM that were passaged, a higher
proportion derived from CSF2-treated blastocysts survived to passage 2 (43% vs 7%;
P=0.016) and passage 3 (36% vs 4%; P=0.025) than ICM from control or ESM+2i

63
treated blastocysts. One colony from a CSF2-treated blastocyst that survived to the
end of passage 3 was examined for expression of NANOG and CDX2. The colony
expressed NANOG but not CDX2 (Figure 2-2B).
In the second experiment, treatments were applied from Day 5 to 7 post-
insemination. This timing was chosen because exposure of embryos to CSF2 from Day
5 to 7 improves competence to establish and maintain pregnancy after transfer into
recipients [18]. The microscopic appearance of ICM that survived passage 1 is shown
in Figure 2-3 and the results are summarized in Figure 2-2C. Although not significant,
the percent of Day 7 ICM in the control group that survived passage 1 was higher than
in the previous study using Day 8 ICM (64 vs 33%, P=0.17). While also not significant,
there was a tendency for survival to the end of passage 1 to be numerically greater for
ICM from CSF2-treated blastocysts than control blastocysts.
Data on ICM survival were also analyzed across both experiments. The percent
of established colonies that survived to the end of passage 1 was greater (P=0.031) for
ICM from CSF2 treated embryos than ICM from control embryos and there was no
interaction between treatment and timing of treatment (i.e., CSF2 increased survival
regardless of whether treatment was from Days 5 to 7 or 6 to 8 post-insemination).
Data on ICM survival were scrutinized to determine whether effectiveness of
CSF2 for improving survival of ICM depended on the percent of oocytes that became
blastocysts in the control groups. Effects of CSF2 did not depend on development class
at either Day 7 or 8 post-insemination (results not shown).
Expression of Genes Involved in Pluripotency and Differentiation
To determine whether CSF2 increases survival of ICM because it regulates
genes involved in pluripotency and differentiation, we evaluated the effect of CSF2 from

64
Day 6-8 post-insemination on expression of genes involved in pluripotency (NANOG
and SOX2) and differentiation of TE (CDX2) and hypoblast (GATA6) in isolated ICM.
There was no effect of treatment on steady-state amounts of mRNA for any of the
genes analyzed (Figure 2-4). There was also no interaction between CSF2 and
development class.
TE Outgrowth
When blastocysts are cultured on Matrigel, TE cells spread onto the surrounding
substratum and proliferate [220]. Moreover, exposure to appropriate growth factors
(FGF2) causes accumulation of hypoblast cells in the outgrowths [222]. Since the
trophoblast of Day 15 conceptuses formed following treatment with CSF2 from Day 5-7
post-insemination had higher expression of the maternal signaling gene IFNT2 and
tended to be numerically greater in length than control embryos [30], it was
hypothesized that CSF2 would increase the spread of TE cells, reinforce commitment to
the TE lineage by enhancing expression of CDX2, decrease expression of the hypoblast
marker GATA6, and increase IFNT2 secretion (as measured by antiviral activity in the
medium). Experiments were conducted by culturing embryos with CSF2 or control from
Day 5 to 7 or 6 to 8 post-insemination and culturing the resultant blastocysts individually
until Day 13 or 15 post-insemination. Results are in Table 2-1.
There was no effect of CSF2 on the percent of blastocysts that attached, the
percent of blastocysts or attached blastocysts that experienced TE outgrowth or the
percent of cells in the outgrowth that were positive for CDX2. Similarly, there was no
effect of CSF2 on secretion of antiviral activity into the culture medium.

65
The effect of CSF2 on size of outgrowths depended on the timing of treatment.
CSF2 decreased the size of the outgrowth when treatment was from Day 5 to 7 post-
insemination but had no effect on outgrowth size when treatment was from Day 6 to 8.
Expression of CDX2 was higher for outgrowths of CSF2-treated blastocysts
when treatment was from Day 6 to 8 but not when treatment was from Days 5 to 7
(Table 2-1). There was no effect of CSF2 at either time on expression of GATA6 (Table
2-1).
Effects of CSF2 on characteristics of TE outgrowth were not affected by the
overall development to the blastocyst stage in controls, i.e., there were no CSF2 x
development class interactions at either Day 7 or 8 post-insemination.
Expression of CSF2 Receptor Subunit Genes
Expression of CSF2RA and CSF2RB was examined at the zygote, 2 cell, 3-4
cell, 5-8 cell, 9-16 cell, morula and blastocyst stages of development. CSF2RA was
expressed at all stages. Expression for CSF2RA increased between the zygote to the
9-16 cell stage before declining to the blastocyst stage (P<0.05) (Figure 2-5A). In
contrast, CSF2RB was not detected in any sample of embryos regardless of stage of
development (Figure 2-5B). CSF2RB was expressed in leukocytes, however (Figure 2-
5B), indicating the effectiveness of the PCR procedure.
Discussion
Exposure of bovine embryos from the morula to blastocyst stage of development
with CSF2 increases survival of the embryo after transfer into a recipient female in cows
[18] as well as in mice [26] and humans [25]. These results implicate CSF2 as an
important maternal signal controlling embryonic development. Experiments presented
here suggest two potential mechanisms by which CSF2 could enhance the competence

66
of a blastocyst to establish and maintain pregnancy, namely by regulating pluripotency
and survival of cells derived from the ICM and alteration of TE function. Similar to the
preimplantation mouse, human and pig embryo [186, 187, 189], actions of CSF2 on the
bovine embryo are likely mediated by a signaling mechanism that is distinct from that in
most cells because the CSF2RB subunit was not detected in any sample of embryos
throughout the period culminating in formation of the blastocyst.
Actions of CSF2 to promote survival of isolated ICM in a pluripotent state are
consistent with earlier observations indicating CSF2 regulates ICM formation and
embryo differentiation status. In particular, CSF2 increased the number of cells in the
ICM of the blastocyst [18] and down regulated expression of several genes involved in
differentiation in the Day 6 morula [31]. Actions of CSF2 on the ICM are likely to reflect
an important mechanism for enhancing embryo survival. Inadequate formation or
survival of the ICM appears to be an important cause of pregnancy loss. At Day 14-17
of pregnancy, about 25-33% of conceptuses generated following embryo transfer in
cattle are missing the embryonic disk derived from the ICM [18, 225, 226]. In humans,
blastocysts with larger ICM were more likely to implant after transfer than blastocysts
with smaller ICM [227].
There are several possible mechanisms by which CSF2 could affect the ability of
the ICM to survive in culture in a pluripotent state. The ICM is not a homogenous group
of cells, especially by Day 8 when specific subpopulations can be identified based on
labeling for NANOG and GATA6 [47]. Perhaps CSF2 changes the relative numbers of
cell types in the ICM in a way that promotes survival in a pluripotent state. In the
absence of CSF2, ICM from Day 7 blastocysts tended to be better able to survive in

67
culture than ICM from Day 8. The former is less differentiated [47] and it may be that
actions of CSF2 to prevent differentiation is responsible for its actions on ICM survival.
There was no effect of CSF2 on expression of NANOG or SOX2 in ICM but regulation
of other pluripotency or differentiation factors by CSF2 could be involved. Also, CSF2
might improve survival of ICM by preventing apoptosis because CSF2 can inhibit
apoptosis and induce expression of anti-apoptotic genes in bovine [31] and mouse [216]
embryos. It is also possible that CSF2 induces changes to cells of the ICM that affect
their interaction with the underlying fibroblast layer, for example by affecting cell
adhesion or cell signaling.
The reason for the failure of the ESM+2i treatment to improve survival of ICM,
despite beneficial effects seem earlier [218], is not clear although the low degree of
replication may have obscured small improvements in survival.
One implication of the finding that CSF2 promotes survival of ICM in a pluripotent
state is that the cytokine might prove useful in generation of embryonic stem cells (ESC)
or induced pluripotent stem cells in the cow. There is a need for growth factors that
promote pluripotency in the cow because growth factors that are involved in
maintenance of ESCs in the human and mouse (FGF2 [228] and LIF [229],
respectively) are not effective in the cow [230].
The effects of CSF2 on characteristics of TE outgrowth were small, dependent
upon the timing of treatment and did not involve changes in secretion of antiviral activity
that represents the trophoblast-derived interferon, IFNT2. CSF2 decreased the size of
TE outgrowths when embryos were exposed from Day 5 to 7 but not when embryos
were exposed from Day 6 to 8. In addition, CSF2 increased expression of the

68
transcription factor CDX2 slightly when embryos were treated from Day 6 to 8 but not
when treated from Day 5 to 7. Like the ICM, the TE of the bovine undergoes important
changes between Days 7 and 8. The TE is not yet committed to the TE lineage at Day 7
but rather can give rise to ICM when aggregated with cleavage-stage embryos [94].
Perhaps, the TE is more differentiated at Day 8 than Day 7 and one consequence is the
action of CSF2 on TE function changes. Consistent with a change in differentiation
status of the TE between Day 7 and 8 is the fact that GATA6 is present in both TE and
ICM at Day 7 but becomes localized to the ICM by Day 8 [47]. An increase in CDX2
expression in outgrowths of TE from Day 8 blastocysts caused by CSF2 could reflect
actions of CDX2 to promote TE function. Indeed, embryos treated with CSF2 from Day
5-7 and transferred into cows had higher expression of the trophoblast gene IFNT2 at
Day 15 [30].
There are many reports that CSF2 can increase the proportion of cultured
embryos that develop to the blastocyst stage, including observations in the cow [24],
human [200], pig [189, 198, 199] and mouse [26]. In other experiments, however, CSF2
either did not affect development [203] or was detrimental for development of embryos
to the blastocyst stage [231]. Present results indicate that the effect of CSF2 on
development to the blastocyst state depended on the overall competence of embryos
for development. In particular, CSF2 increased the percent of embryos becoming
blastocysts when the development rate in control blastocysts was low. In contrast,
CSF2 decreased development when blastocyst development rates in controls were
high. This finding is reminiscent of the first report indicating CSF2 improved blastocyst
yield [24]. In that experiment, CSF2 improved the percent of bovine embryos becoming

69
blastocysts when serum was not present in the medium and development was low but
there was no effect of CSF2 when serum was present and development was high. It
appears, therefore, that CSF2 enhances developmental competence of embryos with
characteristics that limit ability for development while reducing the prospects for an
embryo becoming a blastocyst for embryos that have characteristics that allow
development to proceed to the blastocyst stage. Perhaps, the anti-apoptotic effects of
CSF2 [31] increase the ability of a poor embryo to become a blastocyst. If in fact CSF2
promotes pluripotency rather than differentiation [18, 30], it might delay but not prevent
the differentiation of the TE in an embryo with a high developmental capacity. Such an
explanation is consistent with the effects of CSF2 from Days 5-7 on TE outgrowth as
well as the finding that the negative effects of CSF2 on blastocyst development were
lower at Day 8 than Day 7.
As found earlier for the mouse and human [186, 187], the preimplantation bovine
embryo expresses only one of the two subunits of the CSF2R. Typically, CSF2 signaling
occurs through a cell surface receptor consisting of CSF2RA and CSF2RB subunits
[181, 217]. Binding leads to activation of JAK2 followed by phosphorylation of signal
transducer and activation of the STAT5 pathway [190, 232]. CSF2RA binds to CSF2
with low affinity (Kd = 2.7 nM) while recruitment of CSF2RB causes an increase in
affinity to 170 pM [181]. It is possible to achieve signal transduction in the absence of
CSF2RB. In Xenopus oocytes, CSF2 binding to CSF2RA in the absence of CSF2RB
can signal through activation of phosphatidylinositol 3-kinase [183]. Thus, it seems likely
that CSF2 signals through a pathway independent of CSF2RB, even though CSF2 can
affect the embryo at concentrations as low as 2 ng/ml (i.e., 140 pM) in humans [25].

70
Nonetheless, the conclusion that CSF2 signals independent of CSF2RB must be
tempered by two recent observations. Very low levels of mRNA for CSF2RB were
detected in pig embryos at the blastocyst stage [189]. In addition, both CSF2RA and
CSF2RB could be immunolocalized to bovine oocytes [188] and it is possible that the
embryo uses CSF2RB that was inherited from the oocyte. However, immunolabeling
for CSFR2B was much lower than for CSFRA [188] and it seems likely that little
CSF2RB is present even in the oocyte.
In conclusion, CSF2 acts on the bovine preimplantation embryo to alter the ICM
so it is better able to survive and remain pluripotent while cultured on BFF. Accordingly,
it is hypothesized that one mechanism by which CSF2 improves competence of
blastocysts to establish pregnancy involves regulation of ICM survival and differentiation
status. There were also day-of-development specific effects of CSF2 on TE outgrowths,
indicating that CSF2 regulates the function of cells of the TE lineage as well. The nature
of this regulation and its implications for subsequent development remain to be
established. Transcripts for CSF2RB were not detected in embryos in the period
culminating in blastocyst formation, indicating the likelihood that CSF2 signals only
through a CSF2RB-independent mechanism.
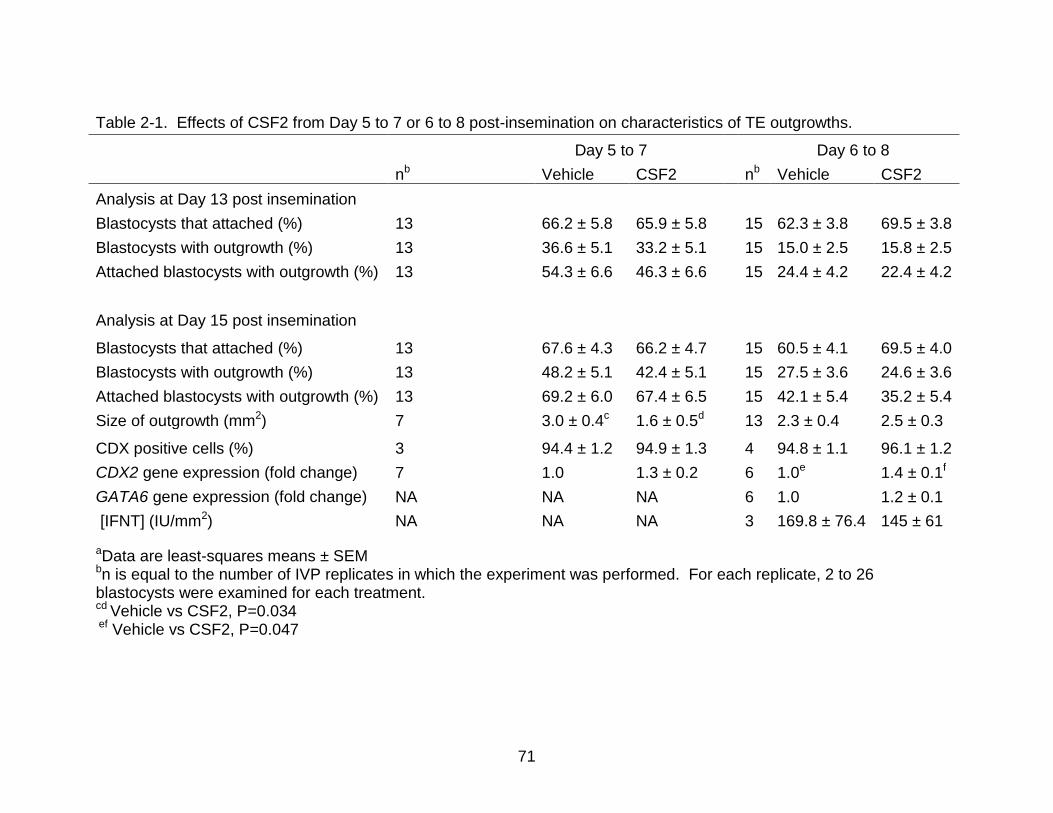
71
Table 2-1. Effects of CSF2 from Day 5 to 7 or 6 to 8 post-insemination on characteristics of TE outgrowths.
aData are least-squares means ± SEM bn is equal to the number of IVP replicates in which the experiment was performed. For each replicate, 2 to 26 blastocysts were examined for each treatment. cd Vehicle vs CSF2, P=0.034 ef Vehicle vs CSF2, P=0.047
Day 5 to 7 Day 6 to 8
nb Vehicle CSF2 nb Vehicle CSF2
Analysis at Day 13 post insemination
Blastocysts that attached (%) 13 66.2 ± 5.8 65.9 ± 5.8 15 62.3 ± 3.8 69.5 ± 3.8
Blastocysts with outgrowth (%) 13 36.6 ± 5.1 33.2 ± 5.1 15 15.0 ± 2.5 15.8 ± 2.5
Attached blastocysts with outgrowth (%) 13 54.3 ± 6.6 46.3 ± 6.6 15 24.4 ± 4.2 22.4 ± 4.2
Analysis at Day 15 post insemination
Blastocysts that attached (%) 13 67.6 ± 4.3 66.2 ± 4.7
15 60.5 ± 4.1 69.5 ± 4.0
Blastocysts with outgrowth (%) 13 48.2 ± 5.1 42.4 ± 5.1 15 27.5 ± 3.6 24.6 ± 3.6
Attached blastocysts with outgrowth (%) 13 69.2 ± 6.0 67.4 ± 6.5 15 42.1 ± 5.4 35.2 ± 5.4
Size of outgrowth (mm2) 7 3.0 ± 0.4c 1.6 ± 0.5d 13 2.3 ± 0.4 2.5 ± 0.3
CDX positive cells (%) 3 94.4 ± 1.2 94.9 ± 1.3
4 94.8 ± 1.1 96.1 ± 1.2
CDX2 gene expression (fold change) 7 1.0 1.3 ± 0.2 6 1.0e 1.4 ± 0.1f
GATA6 gene expression (fold change) NA NA NA 6 1.0 1.2 ± 0.1
[IFNT] (IU/mm2) NA NA NA 3 169.8 ± 76.4 145 ± 61
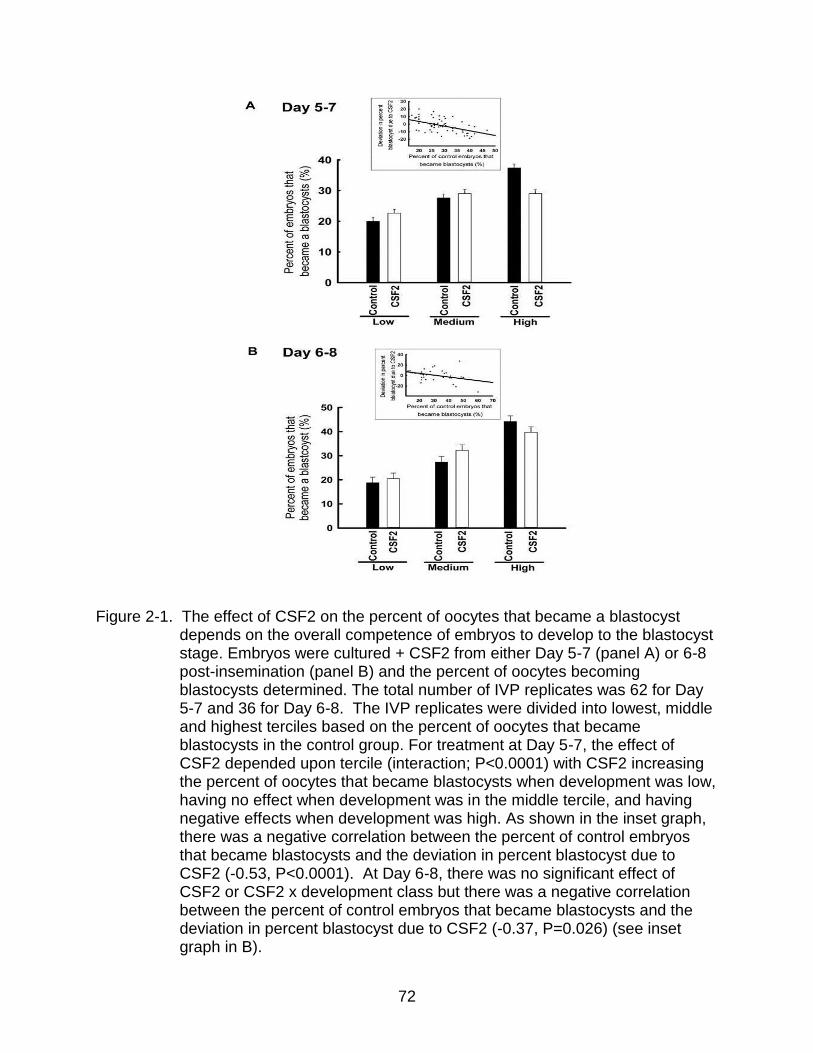
72
Figure 2-1. The effect of CSF2 on the percent of oocytes that became a blastocyst depends on the overall competence of embryos to develop to the blastocyst stage. Embryos were cultured + CSF2 from either Day 5-7 (panel A) or 6-8 post-insemination (panel B) and the percent of oocytes becoming blastocysts determined. The total number of IVP replicates was 62 for Day 5-7 and 36 for Day 6-8. The IVP replicates were divided into lowest, middle and highest terciles based on the percent of oocytes that became blastocysts in the control group. For treatment at Day 5-7, the effect of CSF2 depended upon tercile (interaction; P<0.0001) with CSF2 increasing the percent of oocytes that became blastocysts when development was low, having no effect when development was in the middle tercile, and having negative effects when development was high. As shown in the inset graph, there was a negative correlation between the percent of control embryos that became blastocysts and the deviation in percent blastocyst due to CSF2 (-0.53, P<0.0001). At Day 6-8, there was no significant effect of CSF2 or CSF2 x development class but there was a negative correlation between the percent of control embryos that became blastocysts and the deviation in percent blastocyst due to CSF2 (-0.37, P=0.026) (see inset graph in B).
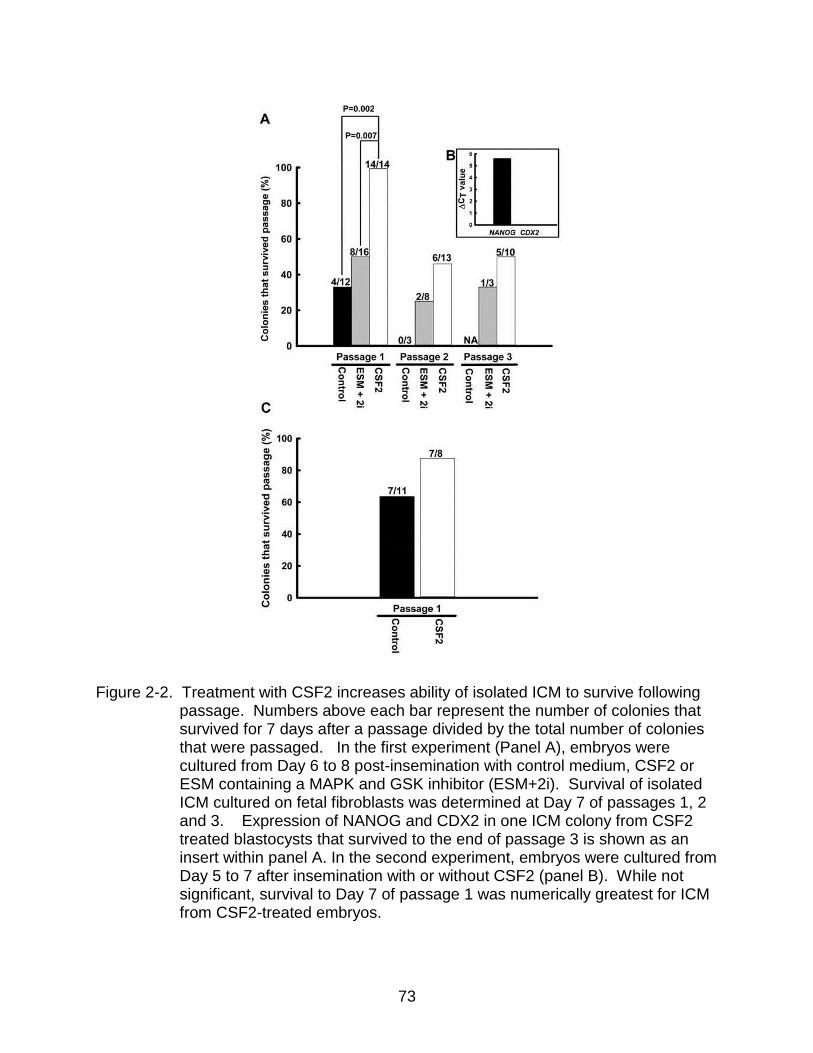
73
Figure 2-2. Treatment with CSF2 increases ability of isolated ICM to survive following
passage. Numbers above each bar represent the number of colonies that survived for 7 days after a passage divided by the total number of colonies that were passaged. In the first experiment (Panel A), embryos were cultured from Day 6 to 8 post-insemination with control medium, CSF2 or ESM containing a MAPK and GSK inhibitor (ESM+2i). Survival of isolated ICM cultured on fetal fibroblasts was determined at Day 7 of passages 1, 2 and 3. Expression of NANOG and CDX2 in one ICM colony from CSF2 treated blastocysts that survived to the end of passage 3 is shown as an insert within panel A. In the second experiment, embryos were cultured from Day 5 to 7 after insemination with or without CSF2 (panel B). While not significant, survival to Day 7 of passage 1 was numerically greatest for ICM from CSF2-treated embryos.

74
Figure 2-3. Representative appearance of colonies of cells derived from ICM at Day 7
passage 1. Colonies were derived from blastocysts that were produced in control medium with control, ESM containing a MAPK and GSK inhibitor (ESM+2i) or medium with CSF2. The scale bar represents 50 μm.

75
Figure 2-4. Lack of effect of CSF2 on expression of genes related to pluripotency and
differentiation in ICM isolated from Day 8 blastocysts. Data for mRNA are represented as the inverse of the ΔCT least-squares means ±SEM of results from 7-9 pools of 3-5 isolated ICM per treatment. Black and white bars represent control and CSF2 treated ICMs, respectively. There was no significant effect of treatment on gene expression.

76
Figure 2-5. Developmental changes in steady-state mRNA for CSF2RA and CSF2RB. Panel A: Developmental changes in transcript abundance for CSF2RA expressed as fold-change relative to amount for oocytes. Data are the least-squares means ± SEM of results from pools of 25-30 embryos replicated 5 times. Bars with different superscripts differ (P<0.05). Panel B: Representative electrophoretogram of PCR products for CSF2RA (A), CSF2RB (B), GAPDH, and SDHA. Abbreviations are as follows: blast=blastocyst, leuk.=leukocytes, GAP.=GAPDH. Arrows represent the amplicon location of CSF2RA (A) and CSF2RB (B).

77
CHAPTER 3 SEXUAL DIMORPHISM IN DEVELOPMENTAL PROGRAMMING OF THE BOVINE PREIMPLANTATION EMBRYO CAUSED BY COLONY STIMULATING FACTOR 2
Introduction
Physiological function and health of the adult is first established during prenatal
life: alterations in the maternal environment can lead to changes in the adult that include
susceptibility to hypertension, diabetes, cardiovascular disease, and various cancers
[233-236]. Programming of development occurs as early as the preimplantation period.
Feeding pregnant female mice or rats a low protein diet for a limited period
corresponding to preimplantation development caused changes in the offspring that
persisted into adult life [13-15]. Similarly, postnatal physiology and immune function
were altered in sheep derived from mothers fed a diet deficient in cobalt and sulfur for 8
weeks before and 6 days after mating [16]. Furthermore, embryos that develop in vitro
give rise to neonates that have increased likelihood for a variety of abnormalities as
shown in humans [237, 238], cows [239, 240], sheep [240] and mice [241, 242].
One of the characteristics of developmental programming in the preimplantation
period is that female embryos are programmed differently than male embryos [243]. In
mice, for example, feeding a low protein diet during the first 3.5 days following
conception affected females differently than males. Postnatal growth and body size was
affected by protein diet in female offspring but not male offspring [13, 14]. At adulthood,
females from mothers fed low-protein diets had altered behavioral scores [13],
increased blood pressure [14] and reduced heart to body weight ratio [13] as compared
to animals born from control females. These traits were not affected in male offspring
but males from low-protein mothers had elevated lung angiotensin-converting enzyme
[17]. In rats, feeding a methyl-deficient diet with no folic acid for 3 weeks prior to mating

78
and 5 days after gestation increased the concentration of insulin in males but not female
offspring [169].
Developmental plasticity in response to maternal environment has been
proposed as an evolutionary strategy to allow individuals to adapt to transient changes
in environment while preserving genotype [236]. Sexual dimorphism in developmental
programming may reflect the fact that relative value of female and males for
reproductive fitness varies with environment [244].
The mechanism by which a change in maternal environment alters the
developmental program of a female embryo differently than of a male embryo is not
known. One possibility is that female embryos respond differently to maternally-derived
molecules that regulate embryonic development (hereafter termed embryokines) than
do male embryos. If so, alteration in secretion of maternal embryokines in response to
environmental change could result in a sex-specific deviation in the developmental
program. The possibility of differential responsiveness to maternal embryokines is
strengthened by the observations that gene expression is different between female and
male embryos as early as the blastocyst stage [245, 246].
Colony stimulating factor 2 is a good candidate for an embryokine responsible for
sex-specific programming. This cytokine is produced by the oviductal and endometrial
epithelium of a wide range of mammals including mice [28], cattle [27] and humans [29].
Treatment of preimplantation embryos with CSF2 leads to an increase in number and
pluripotency of cells of the ICM [18, 247, 248], altered gene expression [30] and,
increased resistance to apoptosis [31]. That CSF2 is capable of altering the
developmental program of the embryo is indicated by findings that bovine [18], mouse

79
[26], and human [25] embryos treated with CSF2 have increased likelihood to survive to
term when transferred into females. Endometrial secretion of CSF2 also changes in
response to maternal environment, as indicated by experiments showing reduced
expression of CSF2 in oviduct of obese cows [34] and increased secretion of CSF2
after induction of inflammation [33].
Culture of bovine embryos with CSF2 from day 5-7 of development resulted in
alterations in characteristics of the embryo at a later time point [18, 30]. In particular,
embryos treated with CSF2 tended to be longer and produce more of the anti-luteolytic
signal, IFNT, at day 15 of development than control embryos. For the experiment
reported here, we used this model of CSF2-mediated developmental programming to
explore whether such changes in the trajectory of development depend upon sex. Day
15 is an important point in time for the survival of the bovine embryo. At this time, the
extraembryonic membranes (EEM) of the embryo are in the process of extensive
elongation so that the embryo increases ~50 fold in length between day 13 and day 16
[35]. Coincident with elongation of the EEM is activation of trophoblast transcription of
IFNT, which encodes for a type I interferon that acts on the endometrium to block
secretion of prostaglandin F2 and thereby ensure survival of the corpus luteum and
continued progesterone secretion [55]. The results presented here indicate that the
consequences of exposure to CSF2 from day 5-7 of development are fundamentally
different for the female and male embryo at day 15 with respect to embryo elongation,
IFNT secretion, and characteristics of the transcriptome and methylome. These results
support the contention that sexual dimorphism in response to maternal embryokines is

80
an important mechanism for establishment of sex differences in developmental
programming.
Materials and Methods
Embryo Production
In vitro production of bovine embryos was performed as previously described [22]
unless otherwise noted. Briefly, COC from Holstein females were purchased from
either Sexing Technologies (Navasota, TX, USA) or Ovitra (Midway, TX, USA). The
COC were shipped overnight at 39oC in a portable incubator while cultured in a
proprietary maturation medium. Once arrived, groups of 20-35 matured oocytes were
fertilized with Percoll-purified sperm (1.0 x 106/ml) from a single Holstein bull for 8 hours
at 38.5ºC in 1.7 ml of SOF-FERT [221]. Cumulus cells were denuded after fertilization
by vortexing in 600 μl HEPES-TALP containing 1,000 U/ml hyaluronidase. Putative
zygotes were then cultured in groups of 30 in 45 µl microdrops of SOF-BE1 [22]
covered with mineral oil at 38.5oC in a humidified atmosphere of 5% (v/v) O2 and 5%
(v/v) CO2 with the balance N2. Cleavage rate was assessed at day 3 post-insemination.
On day 5 post-insemination, drops were supplemented with either 5 µl recombinant
bovine CSF2 [Novartis, Basle, Switzerland; 100 ng/ml in DPBS and 1% (w/v) BSA]; final
concentration of CSF2 after dilution in the drops = 10 ng/ml] or 5 µl of the vehicle used
to dilute CSF2 (control treatment). The concentration of CSF2 was chosen based on its
effectiveness for increasing embryonic survival after transfer into recipient females [18].
Embryo Transfer
Grade 1 expanded blastocysts [249] were harvested from culture drops at day 7
of development and loaded into 0.25 ml straws in 250 μl HEPES-TALP [250]
supplemented with 10% (v/v) fetal calf serum and 50 μM dithiothreitol (Sigma-Aldrich).

81
Each straw contained one embryo and was inserted horizontally into a portable
incubator (AI gun warmer, Rusk, TX, USA) at 38.5°C and transported to the Dairy Unit
at the University of Florida (Hague, FL, USA).
For each replicate, ovulation was synchronized using a modified OvSynch 56
protocol [251]. Cows received intramuscular injections of 100 μg gonadotrophin
releasing hormone, (GnRH) (Cystorelin, Merial, Duluth, GA, USA) on the morning of
day -8 (day 0 = expected day of ovulation), 25 mg prostaglandin F2, (Lutalyse,
Zoetis, Madison, NJ, USA) on the morning and afternoon of day -2 and 100 μg GnRH
on the afternoon of Day -1. Cows having a detectable corpus luteum on day 7, as
determined by ultrasonography using a Hitachi Aloka 500 ultrasound with 5 MHz linear
array transducer (Hitachi Aloka, Tokyo, Japan), were randomly assigned to receive a
CSF2-treated or control embryo. The embryo was transferred transcervically to the
uterine horn ipsilateral to the ovary containing the corpus luteum using an embryo
transfer pipette after application of a local epidural block using 2% (w/v) lidocaine.
Embryo Recovery
Embryos were recovered at day 15 of development by flushing the uterine horn
ipsilateral to the corpus luteum with DPBS containing 1% (v/v) polyvinyl alcohol (PVA)
(DPBS-PVA). Flushing medium was introduced into the uterus by means of a 20
French Foley catheter inserted transcervically using a stainless steel blunt stylet placed
inside the catheter. After placement, the stylet was withdrawn and the catheter held in
place by inflation of a cuff at the end of the catheter. The flushing procedure, which
involved introduction of 60 ml into the uterus and recovery of fluid through the catheter,
was repeated four times or until an embryo was recovered.

82
Embryos recovered by flushing were assessed by light microscopy for stage of
development, length, presence of an embryonic disc (ED), and whether the embryo was
recovered intact or in pieces. All recovered embryos were at the filamentous stage of
development. If found, the ED was dissected from the EEM using a sterile scalpel and
tweezers. The EEM was snap frozen in liquid N2 in a minimal amount of DPBS-PVA and
stored at -80°C until extraction of gDNA and total RNA. The fluid from the first uterine
flush was centrifuged for 10 min at 1000 x g and stored at -20°C.
The total number of day 15 embryos collected using the procedures described
above were 16. An additional 14 day 15 embryos that had been treated with CSF2 or
vehicle were used for the analysis of sex and CSF2 effects on embryo length and IFNT
accumulation in the uterus. Details of production of these embryos were reported by
Loureiro et al. (2009). To determine whether sex effects on length and IFNT in embryos
occur earlier in development, we analyzed effect of sex on 10 day 14 control embryos
(i.e., not treated with growth factors during culture). Details of the procedures for
production of these embryos are reported elsewhere [225].
IFNT Assay
Amount of biologically active IFNT in uterine flushings was determined by
measuring the inhibition of vesicular stomatitis virus (VSV) induced lysis of Madin-Darby
bovine kidney (MDBK) cells as described elsewhere [222]. Antiviral activity was
converted to units of IFN (IU/ml) by comparing inhibition to that achieved with a human
IFNA standard (Millipore, Billerica, MA, USA).
Data on embryo length and uterine IFNT were analyzed by least-squares
analysis of variance using the GLM procedure of the Statistical Analysis System (SAS)

83
version 9.3 (SAS Institute Inc., Cary, NC, USA) with effects of sex, treatment and sex x
treatment.
Extraction of gDNA and Total RNA
gDNA and total RNA were extracted using the AllPrep DNA/RNA Mini kit
(Qiagen, Valencia, CA, USA). Embryos were disrupted with Buffer RLT Plus containing
β-mercaptoethanol, vortexed for 30 seconds and homogenized through QIAshredder
columns (Qiagen). Homogenized lysate was then separated into gDNA and total RNA
following manufacturer’s guidelines. Samples were eluted in 100 µl for gDNA and 60 µl
for total RNA. Both gDNA and total RNA were stored at -80ºC until processing. Total
RNA quantity, quality and integrity were determined using an Agilent Bioanalyzer 2100
(Santa Clara, CA, USA). All samples had an RNA integrity >7.5. There was an
absence of DNA contamination of RNA samples as assessed by the Bioanalyzer.
Concentration of gDNA was quantified by Nanodrop spectrophotometer (Thermo Fisher
Scientific, Waltham, MA, USA) with all samples being >76 ng/µl.
Sexing by PCR
Sex for each embryo was determined by PCR with primers specific for the Y
chromosome (forward: 5´-GAT CAC TAT ACA TAC ACC ACT-3´, reverse: 5´-GCT ATG
CTA ACA CAA ATT CTG-3´) and an autosomal satellite DNA sequence (forward: 5´-
TGG AAG CAA AGA ACC CCG CT-3´, reverse: 5´-TCG TCA GAA ACC GCA CAC TG-
3´) [252]. Master-mix was composed of 10X PCR buffer, 2.5 mM dNTP, 50 mM MgCl2,
10 µM of the aforementioned primers and 0.2 µl of Taq DNA polymerase (5 U/µl)
(Invitrogen). A total of 2 µl of DNA sample was combined with the 10.2 µl master mix
and subjected to PCR for 7 min at 95ºC, 40 cycles of 30 sec at 95ºC, 30 sec at 57 ºC
and 30 sec at 72 ºC followed by a final extension at 72ºC for 7 min. Each sample (5 µl)

84
was analyzed on a 1.5% (w/v) agarose gel with 5 µl ethidium bromide (10 mg/ml) using
a 100 bp ladder (New England Biolabs, Ipswich, MA, USA). A male control consisted of
fibroblast or sperm DNA and a female control consisted of DNA extracted from an
ovary.
Overview of Analysis of Transcriptome
Total RNA for each sample was extracted, amplified and labeled before
proceeding to analysis of expression by microarray. The Agilent microarray was
specifically designed by EmbryoGene with probes specific for bovine embryo
development. Microarray data were analyzed and 6 DEG were verified by qPCR. Raw
and normalized data are available on the gene expression omnibus website using the
accession number GSE55364.
RNA amplification
Total RNA from 3 female control embryos, 3 female CSF2-treated embryos, 2
male control embryos and 4 male CSF2-treated embryos were subjected to T7 RNA
amplification using the RiboAmp HSPlus RNA amplification kit as described by the
manufacturer (Life Technologies, Carlsbad, CA, USA). A total of 5 ng RNA was
reversed transcribed using an mRNA-specific first primer and first strand synthesis.
Single-stranded cDNA was made into double-stranded cDNA using second strand
synthesis with an exogenous second primer. aRNA was transcribed in vitro from the
purified cDNA second strand synthesis followed by aRNA purification. Round one
purified aRNA was then re-converted to double-stranded cDNA, purified and subjected
to a second round of in vitro transcription. The aRNA was then purified using PicoPure
RNA isolation kit (Life Technologies), quantified by Nanodrop spectrophotometer
(Thermo Fisher Scientific) and stored at -80ºC until labeling.

85
RNA labeling
The aRNA was labeled using the Universal Linkage System (ULS) (Kreatech
Biotechnology, Apeldoorn Gelderland, Netherlands) as described by the manufacturer
with either ULS-Cy3 or ULS-Cy5. Samples were then incubated at 85ºC for 30 min
followed by incubation on ice for 3 min. aRNA was briefly centrifuged at 16,000 x g and
purified using PicoPure RNA isolation kit (Life Technologies) and eluted in 11 µl elution
buffer. Yield and labeling efficiency was determined using the Nanodrop 1000
spectrophotometer.
RNA microarray hybridization and scanning
Microarray analysis of labeled aRNA was performed using an Agilent 4x44,000
slide microarray (Agilent, Santa Clara, CA, USA) designed by EmbryoGENE to over
represent genes expressed by the bovine preimplantation embryo. The probes for this
array were chosen from one-million high-quality reads from Roche 454 Titanium deep
sequencing datasets from in vitro and in vivo produced bovine embryos from the
germinal vesicle stage, 2-cell, 4-cell, 8-cell, 16-cell, morula, blastocyst and hatched
blastocyst stage [253]. The array contains 42,242 probes which represent 21,139
known reference genes, 9,322 probes for novel transcribed regions, 3,677 alternatively
spliced exons, 3,353 3’-tiling probes and 3,723 controls [253]. For hybridization, , 825
ng of one Cy3 labeled aRNA sample and 825 ng of one Cy5 labeled aRNA sample were
combined with 2.75 µl of 0.01X Agilent spike, 11 µl of 10X blocking agent and 2.2 µl of
25X fragmentation buffer (all from Agilent) and brought to 55 μl with nuclease-free
water. One sample that was CSF2 treated (either Cy3 or Cy5 labeled) was always
combined with a sample that was a control (either Cy5 or Cy3 labeled) and of the same
sex. The mixture was heated at 60°C for 15 min to fragment the RNA, placed on ice,

86
allowed to cool for 1 min and then mixed with an equal volume of 2X GEx hybridization
buffer HI-RPM (Agilent) to stop RNA fragmentation. The solution was centrifuged for 1
min at 16,000 x g and placed on ice.
For each array, 100 μl aRNA solution was added into one of the four chambers.
Hybridization proceeded for 17 hours at 65°C while rotating at 10 rpm. Following
hybridization, array slides were washed as described by the manufacturer, and scanned
with a Tecan PowerScanner microarray scanner (Tecan Group Ltd., Männedorf,
Switzerland).
Analysis of transcriptome microarray data
Microarray images were submitted to BioDiscovery Inc. (Hawthrone, CA, USA) to
generate separate datasets for each color (Cy3 and Cy5). In addition, spots were
aligned to the corresponding GenePix array list, which describes the size, position and
label of each spot on the array, and examined for removal if spots on the array that
were larger than the spot or contained a blemish. The mean intensity for each spot was
corrected by subtracting the mean intensity of background values for each sample to
remove background noise. Mean intensity was then transformed by log2 and analyzed
by using JMP Genomics (SAS, Cary, NC, USA). Data were normalized with loess
normalization for dye-dye correction followed by quantile normalization for between-
array correction. Genes that were differentially expressed were determined by analysis
of variance with a model that included effects of treatment, sex and treatment x sex. P-
values for significant genes were then adjusted for multiple testing by SGoF+
estimation, as described by Carvajal-Rodriguez and Uña-Alvarez [254]. All genes that
had an adjusted P-value of <0.05 and where the difference in expression was ≥ 1.5 or ≤
0.67 were considered differentially expressed.

87
Ingenuity pathway analysis software (Ingenuity Systems, Redwood City, CA,
USA) was used to determine gene ontologies and pathways and to identify pathways
with predicted gene activation or inhibition. Canonical pathways were considered
significant when the –log of the p-value for each pathway was greater than 1.30. Only
pathways with 3 or more differently expressed genes were used for analysis. DAVID
was used to determine gene ontologies and functions in which differentially expressed
genes were overrepresented at P< 0.05 [255]. iPath 2.0 was used to identify KEGG
metabolic pathways that contained overrepresentation of differentially expressed genes
[256].
Validation of trancriptome microarray by PCR
Validation of microarray gene expression was accomplished by quantitative
reverse transcription PCR of 12 samples of embryo RNA. Samples were treated with 1
µl (2 U) of DNase I enzyme (New England Biolabs, Ipswich, MA, USA) in a total volume
of 20 µl volume at 37°C for 30 min to remove DNA, and then incubated at 75°C for 15
min to denature DNase. DNase-treated RNA was then reverse-transcribed using the
High Capacity cDNA Reverse Transcription Kit (Life Technologies). Reverse
transcription occurred using random hexamer primers and involved incubation at 25°C
for 10 min, 37°C for 120 min and 85°C for 5 min. The cDNA was stored at -20°C until
further use. Negative controls for real-time PCR were also performed by incubation
without reverse transcriptase.
cDNA was utilized for real-time PCR analysis. For detection of transcript levels,
a CFX96 Real-Time PCR detection System (Bio-Rad, Hercules, CA, USA) utilizing
SsoFast EvaGreen Supermix with Low ROX (Bio-Rad) was employed. Each reaction
contained 1 µl forward primer (0.5 μM), 1 µl reverse primer (0.5 μM), 10 µl Evagreen

88
Supermix (Bio-Rad), 6.8 µl H2O and 1.2 µl cDNA sample. Amplification conditions were
95°C for 30 sec, 40 cycles at 95°C for 5 sec and 60°C for 5 sec, and 1 cycle of melt
curve analysis at 65-95°C in increments of 0.5°C every 2 sec. The ΔCT value was
determined by subtracting the CT value of the sample by the geometric mean [224] of
the CT for three housekeeping genes, GAPDH, SDHA and YWHAZ, which were utilized
because expression is stable over preimplantation development [31, 257]. Primer
sequences are in Table 3-1. The primers for XIST [258], GAPDH 28, SDHA58 and
YWHAZ 58 were described elsewhere and other primers were designed using
PrimerQuest (IDT DNA, Coralville, IA , USA). The accuracy of results from microarray
analysis was determined by calculating the Pearson’s correlation coefficient between
the ΔCT value as determined by PCR with the normalized intensity as determined by
microarray analysis.
Overview of Analysis of Methylome
To analyze the methylome, gDNA was extracted, digested by MseI and digested
with methylation-sensitive enzymes HpaII, HinpII and AciI. Products were then
amplified, labeled, and then labeled gDNA was subjected to microarray analysis using
an Agilent array designed by EmbryoGene. Raw and normalized data are available on
the gene expression omnibus website using the accession number GSE55873.
Fragmentation and amplification of gDNA
The use of restriction enzymes that are sensitive to 5-methylcytosine was
employed. These restriction enzymes, HpaII, HinpII and AciI, digest when the CpG
dinucleotide is not methylated but leave DNA intact if the CpG is methylated.
Concentrated gDNA from 3 female control embryos, 3 female CSF2-treated embryos, 2
male control embryos and 2 male CSF2-treated embryos were diluted to 10 ng in 15 µl

89
and spiked with a 1.5 µl mix of specific DNA sequences sensitive to the restriction
enzymes to serve as a control for subsequent qPCR. gDNA was digested with 0.5 µl (5
U) MseI in a total volume of 50 µl for 16 hours at 37ºC followed by inactivation for 20
min at 65ºC. MseI-digested gDNA was precipitated using 1 µl of 3 M sodium acetate
and linear acrylamide (Life Technologies). The gDNA pellet was reconstituted in 5 µl
H2O. MseI designed linkers were ligated to the 5’ and 3’ end of the digested gDNA
using 1.0 µl (5 U) T4 DNA ligase and 10 mM ATP at 15ºC for 16 h. Linker-ligated gDNA
was digested by adding 0.5 µl (5 U) HpaII and 0.5 µl (5 U) HinpII at 37 ºC for 12 h. After
12 hours of incubation, an additional restriction enzyme, 0.5 µl (5 U) of AciI, was added
to each sample and incubated at 37 ºC for 4 hours followed by heat inactivation at 80ºC
for 10 min. Digestion efficiency was verified using qPCR on a Roche lightcycler 2.0 with
HpaII, AciI and HinpII specific primers. A difference of a least 5 cycle thresholds was
required to proceed to DNA precipitation. If the results were below 5 threshold cycles,
the gDNA was subjected to one more round of enzyme digestion with the
aforementioned enzymes for 4 hours at 37 ºC.
Digested DNA was precipitated as previously mentioned and amplified by
ligation-mediated PCR. Ligation-mediated PCR was accomplished using MseI primers
that bind to the MseI ligated linkers. A second PCR was performed on the amplified
digested gDNA using only the 3’ MseI primer followed by PCR purification using PCR
purification kit (Qiagen). DNA was eluted in 43.5 µl of elution buffer. MseI linkers were
removed using 0.5 µl MseI (10 U/µl) at 37ºC for 16 hours followed by heat inactivation at
65ºC for 20 min. gDNA was then re-purified using PCR purification kit (Qiagen) and
eluted in 43.5 µl.

90
DNA labeling
Digested gDNA was labeled as described for the procedure to label RNA. gDNA
was purified using QIAquick PCR purification kit (Qiagen) and eluted in 23.5 µl elution
buffer. Yield and labeling efficiency of gDNA was determined using the Nanodrop 1000
spectrophotometer. The degree of labeling of gDNA with Cy3 was between 1.75% and
3.5% while labeling with Cy5 was between 0.75% and 2.5%.
DNA microarray hybridization and scanning
The degree of methylation was determined by measuring hybridization of
restriction-digested, amplified, and labeled DNA to probes for DMR in the bovine
genome using an Agilent 2 x 444,000 slide designed by EmbryoGENE. The microarray
probes were designed based on a methylation-specific deep sequencing analysis of the
bovine genome which generated 1.2 million reads from genomic libraries from day 7
blastocysts and day 12 elongated embryos [259]. The microarray containing 414,566
probes surveying 20,355 genes as well as 34,379 CpG islands. For each slide, 1.0 μg
of Cy3 labeled DNA from one sample (either CSF2 or control treated embryo for the
same sex) and 1.0 μg of Cy5 DNA from a second sample (either control of CSF2
treated embryo for the same sex) were combined with 25 μl of bovine Hybloc DNA (1
mg/ml) (Applied Genetics, Melbourne, FL, USA), 130 μl of 2X Hi-RPM (Agilent) and
100X blocking agent (Agilent); the final volume was brought to 195 μl with nuclease-free
water. This solution was heated for 3 min at 95°C and 30 min at 37°C. Samples were
then centrifuged at 10,000 x g and mixed with 65 μl Agilent-CGH Block solution
(Agilent). A total of 245 μl of each sample was placed into the slide and hybridized at
65°C while rotated at 10 rpm for 40 h. Following incubation, slides were washed
according to manufacturer’s procedure and scanned as described previously.

91
Analysis of methylome array
DNA microarrays were analyzed using the limma package of Bioconductor [260].
Raw data were normalized using within-array loess normalization followed by between-
array quantile normalization. Normalized data were then fit to a linear model before
being subjected to an empirical Bayes correction. Mean intensity of each spot on the
array was transformed by log2 while p-values were determined using a moderated t-
statistic [260]. Regions with an estimated absolute fold-change above 1.5 or below 0.67
and a p-value < 0.05 were considered as differentially methylated. Development of
visualization tools was designed to analyze the quality of both overall genomic and
spiked-in gDNA control digestion. DMR were categorized based on CpG island density
[261], length [262] and distance [263], location of the DMR in the genome and types of
repeated elements [264].
Regions of each chromosome that had a frequency of DMR caused by CSF2
greater than expected were determined by chi-square analysis. For each chromosome,
5 Mbp segments of the genome were examined for observed number of differentially
expressed probes. The expected frequency for chi-square analysis was calculated as
the number of probes in the 5 Mbp segment times the overall percent of probes that
were differentially methylated. The analysis was performed using sliding windows of 5
Mbp that shifted by 500 Kbp at a time until reaching the end of the chromosome.
Analysis was performed separately for female and male embryos.
Results
Embryo Length and IFNT Accumulation in the Uterus
Sex of each embryo was determined by PCR (Figure 3-1A). Embryo length and
amount of IFNT in uterine flushings at day 15 were affected by a treatment x sex

92
interaction (Figure 3-1B, P<0.05). For embryos not treated with CSF2, female embryos
were longer than male embryos and there was greater accumulation of IFNT in the
uterus than for male embryos. In females, CSF2 decreased embryo size and IFNT
accumulation while, in males, CSF2 increased embryo size and IFNT accumulation. As
a result, male embryos treated with CSF2 were larger and secreted more IFNT than
female embryos treated with CSF2.
Another set of embryos collected at day 14 of gestation were examined for sex
differences. In this case, involving embryos not treated with growth factors, there was
no difference in length between female and male embryos (19.4 mm ±11.1 and 23.6
mm ± 11.1 respectively, n=5 for each sex). Thus, it is likely that differences in length
between sexes in control embryos at day 15 represent earlier onset of elongation in
female embryos rather than an inherently-larger size in females.
Differentially Expressed Genes
A Venn diagram illustrating numbers of DEG is presented in Figure 3-2. Among
control embryos, a total of 514 genes differed in expression between female and male
embryos. This represents 1.5% of the probes on the array. Using DAVID (Database for
Annotation, Visualization and Integrated Discovery), DEG genes were overrepresented
in 38 gene ontology terms (P<0.05), with terms related to mitochondrial function, protein
ubiquitination, electron transport and translation being prominent. KEGG pathway
functional annotation terms that were overrepresented in the set of DEG were those for
oxidative phosphorylation (Figure 3-3, P<0.02) and ribosome (P<0.05). Using IPA
software, three canonical pathways having a minimum of three DEG were identified:
EIF2 signaling (P<0.0002), mitochondrial dysfunction (P<0.0003) and fatty acid -

93
oxidation 1 (Figure 3-4A, P<0.007). This finding was reiterated when the list of DEG
was analyzed by iPATH 2.0 [256].
The biofunction analysis feature of IPA was used to predict cellular processes
that would experience a change in activation as a result of changes in gene expression
between female and male embryos. It predicted that synthesis of protein (z-score: -
1.723), translation (z-score: -1.673) and expression of protein (z-score: -1.230) would
be lower for female embryos than male embryos (Figure 3-4B-D).
CSF2 caused differential expression of 54 genes in females and 93 genes in
males. A large proportion of genes regulated by CSF2 in both females and males were
among the set of genes whose expression differed between male and female control
embryos. In particular, 31 of 54 DEG genes in females (57%) and 48 of 93 DEG in
males (52%) were also regulated by sex in control embryos. Only 7 DEG were
regulated by CSF2 in both females and males and regulation was always in opposite
directions. For example, expression of HSP90B1 was increased by CSF2 in females
and decreased in males (Figure 3-2).
Among the genes regulated by CSF2 in females, only 25 of 54 were annotated.
These genes were overrepresented in one DAVID gene ontology, generation of
precursor metabolites and energy (P<0.05), one KEGG pathway, oxidative
phosphorylation (P<0.01). There were no significant IPA canonical pathways or cellular
processes identified by the biofunction analysis feature of IPA. In males, only 60 of 93
were annotated. There were no significant DAVID gene ontology, KEGG pathways or
IPA canonical pathways containing these DEG. However, the biofunction analysis

94
feature of IPA indicated CSF2 increased metabolism of protein and decreased invasion
of cells in male embryos (Figure 3-5).
The accuracy of microarray results was evaluated by determining the correlation
between intensity of hybridization signal in microarray analysis with the ΔCT value from
qPCR for nine genes significantly affected by either CSF2 within sex or between sexes
with control treated samples (Table 3-2). For 7 of 9 genes, an increase in hybdrization
signal was negatively correlated with ΔCT (i.e., positively correlated with amount of
mRNA). The correlation was significant for 6 of 9 genes.
Methylome
A Venn diagram illustrating numbers of DMR is presented in Figure 3-6.
For control embryos, there were 31,706 DMR between females and males. This
represents 7.6% of the probes on the array. Of these DMR, 20,658 (65%) were in open
sea (>4 Kbp from CpG island), 7,824 (24.6%) were in shelf (2-4 Kbp from CpG island),
2,463 (7.7%) in shore (1-2 Kbp from CpG island) and 731 (2.3%) were located in CpG
islands. A total of 30 DMR were not classified.
Treatment with CSF2 from day 5-7 caused hypermethylation at day 15 of 6,227
probes in females and 9,842 probes in males while causing hypomethylation of 3,292
probes in females and 9,322 probes in males (Figure 3-5). In the female, 6,102 (64%)
of these DMR were located in the open sea, 2,452 (25.7%) in the shelf, 787 (8.2%) in
the shore and 181 (1.9%) within a CpG island, while 2 DMR were not classified. In the
male, 11,931 (62%) DMR were located in the open sea, 4,790 (25%) in the shelf, 1,641
(8.5%) in the shore and 792 (4.1%) within a CpG island, while 10 DMR were not
classified.

95
Of the 9519 DMR regulated by CSF2 in females, 2766 (29.1%) were DMR whose
methylation status in controls differed between females and males. Similarly, of the
19164 DMR regulated by CSF2 in males, 6597 (34.4%) were regions that were
differentially methylated between females and males in control embryos. Thus, a
greater percentage of methylation sites regulated by CSF2 are also modified by sex
than is the case for the entire genome (7.6%). A total of 1,186 DMR were affected by
CSF2 in both females and males. Of these, 773 (65.2%) were regulated in the opposite
direction (hypermethylation in one sex and hypomethylation in the other sex).
Chi-square analysis was used to identify whether 1) spatial distribution of DMR on
regions of each chromosome was non-random (as would be expected if DMR were
under regulation) and 2) whether the chromosomal regions that were differentially
methylated by CSF2 in females were distinct from those regions in males. For each
chromosome, there were regions that contained hyper- and/or hypomethylation that was
more frequent than would be expected due to chance if DMR were randomly distributed
across the chromosome. Moreover, the regions of differential methylation caused by
CSF2 were usually different for females than for males, as is illustrated for BTA 1 in
Figure 3-7.
Discussion
It is well known that alterations in the maternal environment during prenatal life
can have long-term effects on the physiology and health of the offspring and that the
nature of these effects differs between females and males [14-16, 169]. From an
evolutionary perspective, sexual dimorphism in developmental programming probably
reflects the differential value of males and females for sexual propagation [244]. While
the phenomenon has been well described, the mechanism by which sexual dimorphism

96
in developmental programming is achieved has hitherto been unknown. Current findings
lead to the hypothesis that sexual dimorphism results because female and male
embryos respond differently to environment-driven alterations in secretion of maternally
derived embryokines. In this model, shown schematically in Figure 3-8, the
reproductive tract uses altered secretion of embryokines as a signal transducer to
transmit information about the maternal environment to the embryo. For certain signals,
as shown here for CSF2, male embryos respond differently than do female embryos so
that the deviation in the developmental trajectory in response to environmental change
varies between sexes.
The evidence for this idea comes from the present experiment that exposure of
bovine preimplantation embryos to CSF2 for a two-day period from day 5-7 of
development, results in changes in embryonic growth, gene expression and DNA
methylation in the EEM of embryos at a later time point in pregnancy, day 15 of
development. It is at this stage of pregnancy that the bovine embryo first signals its
presence to the mother through the secretion of the antiluteolytic molecule, IFNT [55].
Secretion of IFNT is coupled to a process of embryo elongation where the embryo
undergoes a rapid increase in length from about 2-5 mm in length on day 14 to about
150 mm in length on day 16 [265]. Elongation is a prerequisite for the upregulation of
IFNT secretion in the ruminant [56].
Present results suggest that, in the absence of prior exposure to CSF2, the
female embryo is larger and secretes more IFNT into the uterus than the male embryo.
The larger size of female embryos probably reflects earlier onset of elongation rather
than an inherently larger size because there was no difference in embryo length

97
between female and male embryos at day 14, around the time when elongation is being
initiated. Similar lack of difference between female and male embryos in length were
observed for in vitro produced embryos [266] and, in one report, for embryos produced
in vivo [267]. In another report, female in vivo embryos tended to be larger than male
embryos at day 14, although not significantly different [266]. Moreover, IFNT secretion
in vitro was greater for female embryos produced in vivo than male embryos at day 14
[267]. By day 16, male embryos have been reported to be longer than female embryos,
with differences greater for embryos produced in vivo than for those produced in vitro
and subsequently transferred to recipients.
Despite its larger size, analysis of the transcriptome reveals that the female
embryo is less active with respect to oxidative phosphorylation, with NDUFA11,
NDUFS7, UQCRC1, COX6A1 and ATP6V1B2 being downregulated in females and
only NDUFB1 being upregulated. Gene expression in female embryos also indicated
decreased translation and metabolism. The decreased metabolism of female embryos
could reflect the earlier onset of elongation and reduced metabolism as the rate of
growth decreases at the end of the elongation period. However, female embryos have
less mtDNA copies as early as the blastocyst stage [245, 268] and may simply be less
metabolically active than males.
Exposure to CSF2 from day 5-7 reverses differences between female and male
embryos by decreasing length and IFNT release in female embryos while increasing it
in males. Concomitantly, CSF2 programmed gene expression of female embryos
differently than for male embryos. In most cases, genes whose expression at day 15
was altered by prior exposure to CSF2 from day 5-7 were different for female embryos

98
than for male embryos. In the few cases where the same gene was altered in
expression by CSF2 in females and males, regulation was in opposite directions. A
similar phenomenon was observed when examining characteristics of the methylome in
that DMR tended to be different for females and males and when the same, were most
often regulated in opposite directions.
CSF2 is a good candidate for transmitting information about the maternal
environment to the embryo. Produced by the luminal and glandular epithelium of the
endometrium [27], expression of CSF2 is sensitive to perturbation in response to the
maternal environment. In particular, expression of CSF2 in the bovine oviduct was
reduced in obese cows [34] while accumulation of CSF2 or steady-stage amounts of
CSF2 mRNA in the endometrium was increased by deposition of semen in human, pig
and mice [33, 193, 269]. One component of semen responsible for increased
expression of CSF2 in the mouse is Tgfb [33].
Other embryokines exist that could potentially be involved in developmental
programming. Among the embryokines in the cow are IGF1 [270], TGFB [20], LIF [20]
and FGF2 [22]. While each of these molecules can affect embryonic development, it is
not known whether their actions on the embryo also depend upon sex.
It is also not clear how the sex-dependent changes in embryo function seen here
affect the survival of the embryo or the physiology of the resultant neonate. In female
embryos, treatment with CSF2 from day 5-7 increases the proportion of embryos that
establish and maintain pregnancy after transfer to recipient females [18]. Thus, the
reduction in embryo length and IFNT secretion at day 15 is not inimical to embryo
survival. Among the genes upregulated by CSF2 in females were several candidates

99
related to oxidative phosphorylation and it may be that increased capacity of the EEM to
metabolize energy substrates contributes to the embryo’s survival. There is no
comparable information about the effects of CSF2 on subsequent survival of male
embryos. CSF2 altered gene expression in male embryos in a manner suggesting
decreased metabolism of protein and increased invasion of cells in male embryos and
these actions could potentially limit embryo survival.
There is also no information regarding the consequences of CSF2 treatment from
day 5-7 on characteristics of the resultant calves. Data that CSF2 causes sex-specific
changes in the methylome suggest the possibility of long-term changes that could
extend into fetal or postnatal life.
An important unresolved question is the molecular basis for the differential
response of female and male embryos to CSF2. One facet of this response may
involve the nature of genes regulated by CSF2, which include an overrepresentation of
genes and CpGs that are also regulated by sex. Over 50% of the DEG and 35% of the
DMR regulated by CSF2 were among the small pool of genes and DMR that were
regulated by sex. One possibility is that CSF2 regulates transcription factors and other
signaling systems responsible for sex-regulated differences in gene expression.
Alternatively, a large fraction of genes regulated by sex have transcription factors or
regulatory elements also regulated by CSF2.
In conclusion, we present data to indicate a mechanism for sexual dimorphism in
developmental programming. In particular, treatment of preimplantation embryos with
the maternally-derived cytokine, CSF2, from Day 5-7 of development changes
subsequent development so that, at a later stage of pregnancy, Day 15, embryonic

100
development is altered with respect to length, secretion of the pregnancy signal, IFNT,
the transcriptome, and the methylome. Alterations in these characteristics caused by
CSF2 were different between female and male embryos, with effects being often in the
opposite direction. We conclude that alterations in secretion of maternal regulatory
factors caused by change in maternal environment could modify the developmental
program of the female embryo differently than for the male embryo.

101
Table 3-1. List of primers used for qPCR.
Gene Gene ID
Sequence
HSP90B1 NM_174700.2 Forward 5'-CAGGAACAGACGAGGAAGAAC-3' Reverse 5'-CTCTCCACACAGCATCCAAA-3'
PAGE4 NM_001081580.2 Forward 5'-CATGGAACCTGGACAAGAGAAG-3' Reverse 5'-CAAGCTCACCCTCAACCTTT-3'
NDUFB1 NM_175809.2 Forward 5'-TGTACTTGTCCCTATGGGATTTG-3' Reverse 5'-TTGTTCCGGAAGGCAGTTAG-3'
PAG2 NM_176614.1 Forward 5'-CCCTCAAGCCTACATCATCAA-3' Reverse 5'-CCAAGGATCCAGGTGTTTAGAG-3'
EIF3C NM_001034618.2 Forward 5'-TTGCTGAGTGAGGATGAAGAAG-3' Reverse 5'-CATTACGGATGGTCCGGATAAG-3'
PEX10 NM_001076333.1 Forward 5'-CGAACCATGCAGAGGAAAGA-3' Reverse 5'-GGTTATACACTCCCAGCAGAAC-3'
XIST NR_001464.2 Forward 5'-TTGGCTTTTAGATTAATTTGATGAACAGCAT-3' Reverse 5'-CCCTTTAGACTAGGCCCATTTCATA-3'
PCMTD1 NM_001081526.1 Forward 5'-CCCAGTATATCCGCACTGAAAG-3' Reverse 5'-GTCTCGGTAGCCTTCCAAATAG-3'
AMACR XM_002696385.2 Forward 5'-TGGTGGAAGGAACAGCATATC-3'
Reverse 5'-GAAAGGAGCTCCACCATCTAAC-3'
GAPDH NM_001034034.2 Forward Reverse
5’-ACCCAGAAGACTGTGGATGG-3’ 5’- CAACAGACACGTTGGGAGTG-3’
YWHAZ
SDHA
BM446307
NM_174178
Forward Reverse Forward Reverse
5’- GCATCCCACAGACTATTTCC-3’ 5’- GCAAAGACAATGACAGACCA-3’ 5’ GCAGAACCTGATGCTTTGTG-3’ 5’- CGTAGGAGAGCGTGTGCTT-3’

102
Table 3-2. Validation of microarray results by determination of the correlation between fluorescent intensity on the array with CT value in qPCR.
Gene Correlation coefficient P
HSP90B1 -0.86 <0.05 PAGE4 -0.84 <0.05
NDUFB1 -0.74 <0.05 PAG2 -0.74 <0.05 EIF3C -0.47 NS PEX10 0.17 NS XIST -0.89 <0.05
PCMTD1 0.09 NS AMACR -0.73 <0.05

103
Figure 3-1. Sex by CSF2 interaction affecting length of the embryo and accumulation of IFNT in uterine flushings at day 15 of pregnancy. Panel A depicts a representative electrophoretogram of results of the PCR procedure used to sex embryos. Shown are results for three embryos as well as control male and female DNA. The band located at 216 bp represents an amplicon from an automosal satellite DNA sequence while the band at 141 bp represents an amplicon from the Y-chromosome. Interactions between CSF2 treatment from day 5-7 of development and sex on embryo length and concentration of IFNT in uterine flushing are shown in Panel B. Data are the least-squares means ± SEM of results from 7-8 males and 6-8 females per treatment. The treatment x sex interaction was significant (P<0.05) for both endpoints.

104
Figure 3-2. Venn diagram illustrating differentially-expressed transcripts at day 15 of
pregnancy in male and female embryos treated with CSF2 or control medium from day 5-7 of development. Gene symbols represent differentially expressed genes that were annotated. Genes displayed in red increased in expression (female > male or CSF2 > control) while genes displayed in green decreased in expression.

105
Figure 3-3. Diagram illustrating genes encoding for proteins in the electron transport
chain that were differentially expressed between control female and male embryos at day 15 of pregnancy. Genes upregulated in females are in red and genes upregulated in males are in green.

106
Figure 3-4. Biological processes predicted to vary with sex in control embryos. Panel A
shows the canonical pathways containing an overrepresented number of differentially expressed genes. Only pathways with 3 or more genes are shown. Gene symbols represent differentially expressed genes for each pathway; genes in red were upregulated in females and genes in green are upregulated in males. Panels B-D display analyses of predicted differences in cellular function between female and male embryos. Genes in red were upregulated in females while genes in green were upregulated in males (we need to change this). Blue lines represent genes in which the observed change in gene expression are predicted to inhibit the biological function, yellow lines represent genes where the effect of differential expression is inconsistent with the predicted stage of the process and grey lines represent relationships that are not predicted. Biological processes in blue are predicted to be inhibited. Analysis indicates that female embryos were likely to be less active in expression of protein, translation, and translation of protein (panel B), metabolism of protein (panel C) and transport of lipid (panel D) than male embryos.

107
Figure 3-5. Predicted changes in cellular function in male embryos at day 15 of
pregnancy caused by CSF2 treatment between day 5-7 of development. Genes labeled in red were upregulated by CSF2 while genes labeled in green were downregulated by CSF2. Orange lines represent genes where the change in expression is predicted to active the biological function while blue lines represent genes in which the observed change in gene expression is predicted to inhibit the biological function. Yellow lines represent genes where the effect of differential expression is inconsistent with the predicted state of the process and the gray line represents a relationship that is not predicted. Biological processes in blue are predicted to be inhibited. Analysis indicates that embryos treated with CSF2 were likely to experience decreased metabolism of protein and increased invasion of cells.

108
Figure 3-6. Venn diagram illustrating differentially-methylated probes at day 15 of
pregnancy in male and female embryos treated with CSF2 or control medium from day 5-7 of development.
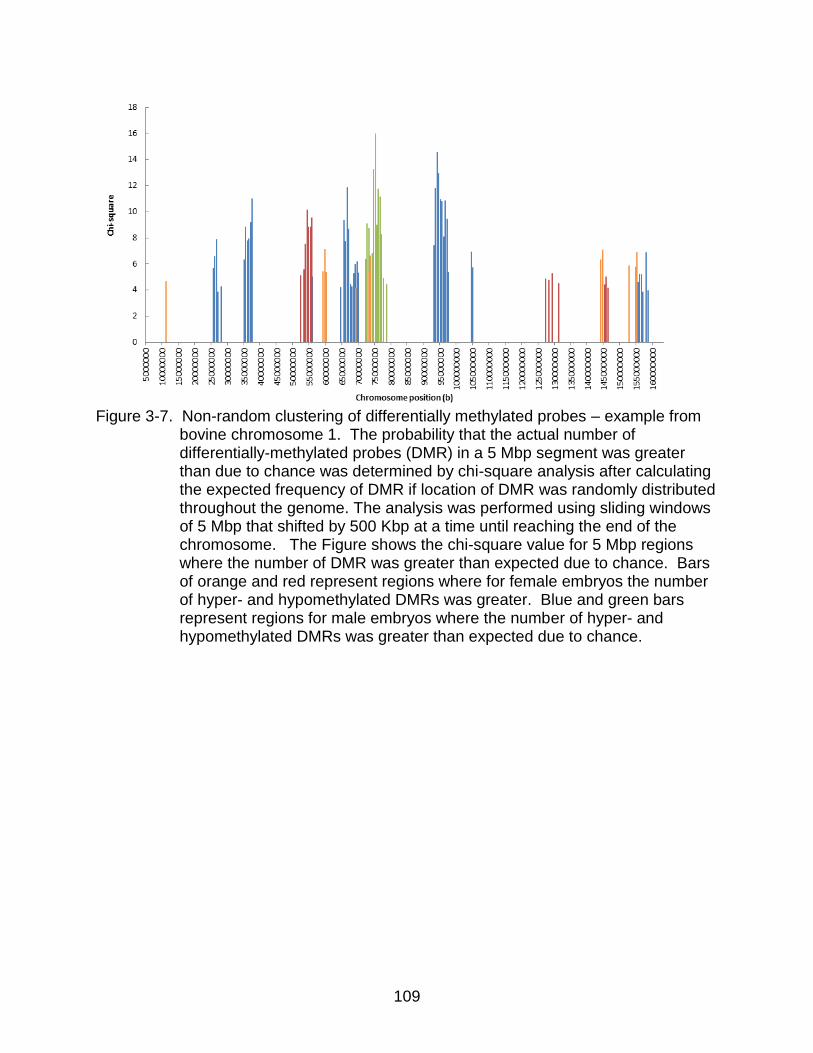
109
Figure 3-7. Non-random clustering of differentially methylated probes – example from
bovine chromosome 1. The probability that the actual number of differentially-methylated probes (DMR) in a 5 Mbp segment was greater than due to chance was determined by chi-square analysis after calculating the expected frequency of DMR if location of DMR was randomly distributed throughout the genome. The analysis was performed using sliding windows of 5 Mbp that shifted by 500 Kbp at a time until reaching the end of the chromosome. The Figure shows the chi-square value for 5 Mbp regions where the number of DMR was greater than expected due to chance. Bars of orange and red represent regions where for female embryos the number of hyper- and hypomethylated DMRs was greater. Blue and green bars represent regions for male embryos where the number of hyper- and hypomethylated DMRs was greater than expected due to chance.
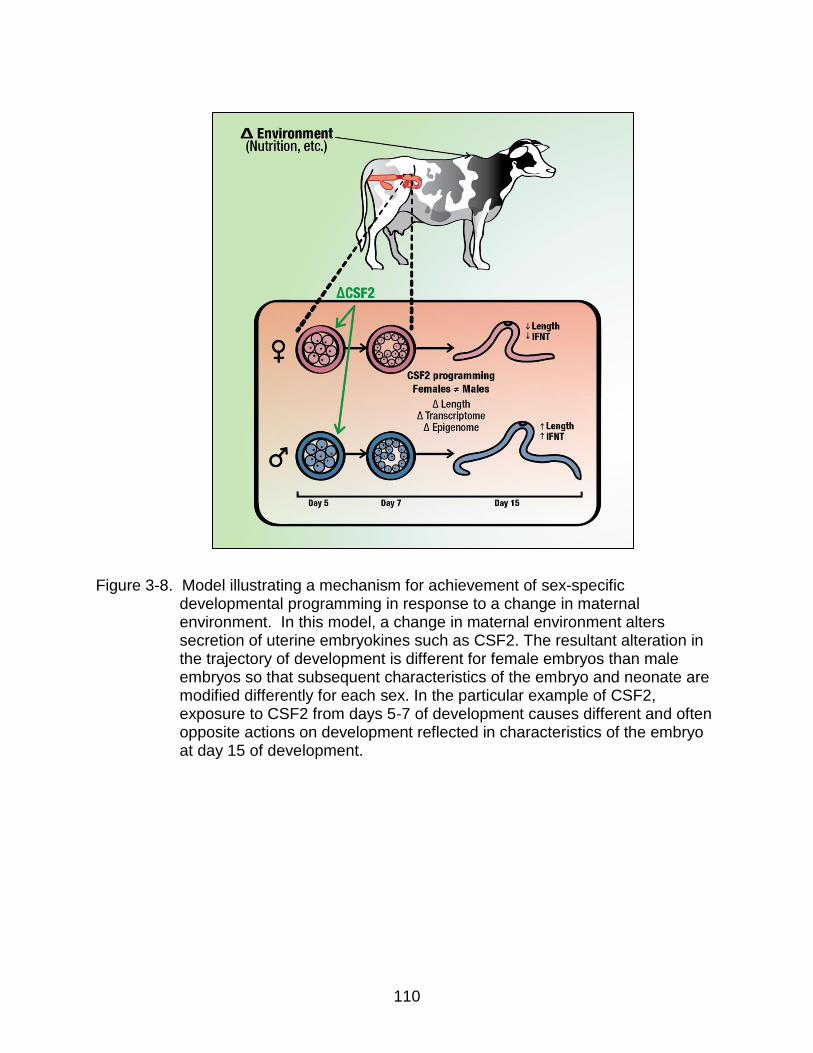
110
Figure 3-8. Model illustrating a mechanism for achievement of sex-specific developmental programming in response to a change in maternal environment. In this model, a change in maternal environment alters secretion of uterine embryokines such as CSF2. The resultant alteration in the trajectory of development is different for female embryos than male embryos so that subsequent characteristics of the embryo and neonate are modified differently for each sex. In the particular example of CSF2, exposure to CSF2 from days 5-7 of development causes different and often opposite actions on development reflected in characteristics of the embryo at day 15 of development.

111
CHAPTER 4 DYNAMICS OF DNA METHYLATION DURING EARLY DEVELOPMENT OF THE
PREIMPLANTATION BOVINE EMBRYO
Introduction
Following fertilization, the embryo remains in a state of transcriptional quiescence
that is maintained until a species-specific stage (8-16 cell stage in the cow, 2-cell stage
in the mouse, 4-cell stage in the pig and 4-8 cell stage in the human) when transcription
is resumed through a process referred to as EGA [271]. In the mouse, activation of
transcription is preceded by a decrease in global methylation of DNA as a result of
active and passive demethylation that begins at the zygote stage and persists through
the morula stage [73, 74]. Demethylation is followed by a wave of DNA methylation
beginning at the blastocyst stage [84] that is mediated by de novo methyltransferases
Dnmt3A and Dnmt3b [82, 84]. Overall levels of methylation in the blastocyst are greater
for ICM than TE [84].
Epigenetic remodeling is important for EGA because mouse embryos in which
the chromatin remodeling gene, Brg1, was knocked out exhibited arrest at the 2-cell
stage and decreased transcription [272]. Moreover, injection of siRNA for the
H3K27me3 demethyltransferase JMJD3 decreased competence of bovine embryos to
develop to the blastocyst stage [273]. Nonetheless, developmental patterns of de novo
methylation in early development are not conserved across mammalian species. The
demethylation occurring during early cleavage-stages in the mouse also occurs in
sheep [85] but not in the pig [86] or rabbit [274]. By the blastocyst stage, the ICM is
more methylated than the TE in the sheep [85] and pig [86] while, in the rabbit, the ICM
is less methylated than TE [274]. De novo methylation is not as clearly understood in
other species. In the sheep, overall DNA methylation declines from the two-cell stage

112
until the blastocyst stage and the ICM is more methylated than the TE [85]. The ICM is
also more methylated than the TE in the pig blastocyst, but unlike the mouse and
sheep, there is no apparent loss of DNA methylation from the two-cell to morula stages
of development [86].
Results are unclear in the bovine embryo, with one report indicating widespread
demethylation occurs through the 8-cell stage before methylation increases at the 16-
cell stage of development [87] while another report indicates that demethylation persists
through the morula stage [88]. The difference in methylation between the ICM and TE
of the bovine blastocyst is also unclear, with one report indicating higher methylation in
the ICM [87] and another indicating higher methylation in TE [88].
Developmental changes in the embryonic methylome are also likely to be
modified by genetics of the embryo and the environment in which it resides. Genetic sex
can have profound effects on development of the embryo as early as the blastocyst
stage when, for example, total transcript abundance in cattle is higher in female
blastocysts than male counterparts [245]. Expression of de novo methyltransferases is
also differentially expressed between the two sexes, with female blastocysts in the cow
having lower expression of DNMT3A and DNMT3B compared to male blastocysts [268].
Signals derived from the mother can also affect embryonic development in a way that
improves competence for survival after transfer into females, as has been demonstrated
in the cow for CSF2 [18], insulin-like growth factor-1 [129] and hyaluronan [21]. The
molecular basis for improvement in competence for long-term development is not
known but could include remodeling of the epigenome. Indirect evidence for epigenetic
programming during early development is the observation that treatment of embryos

113
with CSF2 from Day 5-7 of pregnancy improves late embryonic and fetal survival much
later in pregnancy (after Day 30-35 of gestation) [18].
In this study, we characterized dynamics of DNA methylation during early development
in the cow and determined whether degree of methylation depends upon cell lineage
(ICM vs TE), sex, and exposure to CSF2. In addition, developmental changes in
expression and methylation of DNMT3B were assessed to determine whether
developmental changes in DNA methylation could conceivably be due to differences in
activity of this methylase.
Experimental Procedures
Embryo Production
In vitro production of bovine embryos was performed as previously described [22]
unless otherwise noted. COC were obtained with permission by cutting the surface of
slaughterhouse (Central Beef Packing Co. (Center Hill, FL, USA)-derived ovaries with a
scalpel and vigorously rinsing the ovary through a bath of oocyte collection medium
[tissue culture medium-199 with Earle’s salts without phenol red (Hyclone, Logan UT),
2% (v/v) bovine steer serum (Pel-Freez, Rogers, AR), 2 U/ml heparin, 100 U/ml
penicillin-G, 0.1 mg/ml streptomycin and 1mM glutamine]. Groups of 10 COC were
matured in 50 μl droplets of oocyte maturation medium [tissue culture medium-199 with
Earle’s salts (Invitrogen, Carlsbad, CA), 10% (v/v) bovine steer serum, 2 g/ml estradiol
17-β, 20 g/ml bovine follicle stimulating hormone (Bioniche Life Sciences, Belleville,
Ontario, Canada), 22 g/ml sodium pyruvate, 50 g/ml gentamicin sulfate and 1 mM
glutamine] covered with mineral oil. Maturation proceeded for 20 hours at 38.5oC and in
a humidified atmosphere of 5% (v/v) CO2. Up to 300 matured oocytes were fertilized

114
with Percoll-purified sperm (1 x 106) for 8 hours in 1.7 ml of SOF-FERT [221]. Cumulus
cells were denuded after fertilization for 4 min by vortexing in 600 μl HEPES-TALP [250]
containing 1,000 U/ml hyaluronidase. Embryos were then cultured in 25 µl microdrops
of SOF-BE1 [22] covered with mineral oil at 38.5oC in a humidified atmosphere of 5%
(v/v) O2 and 5% (v/v) CO2 with the balance N2. Cleavage rate was assessed at Day 3
post-insemination (pi) and embryos were cultured until Day 7 pi.
Immunofluorescent Labeling for 5-Methylcytosine
All labeling steps were performed at room temperature unless otherwise stated
and in a volume of 100 µl (either in microdrops covered with oil or in wells of a 96-well
plate). Embryos were harvested from culture drops, washed in DPBS containing 1%
(w/v) polyvinylpyrrolidone (PVP) (Kodak, Rochester, NY, USA) and fixed in 4% (w/v)
paraformaldehyde in DPBS-PVP for 15 min. The fixed embryos were washed three
times in DPBS-PVP, permeabilized for 20 min in 0.25% Triton X-100 (Fisher Scientific,
Waltham, MA, USA) diluted in DPBS, and washed three times in 0.1% (v/v) Tween 20
with wash buffer [DPBS containing 10 mg/ml fraction V BSA (Sigma-Aldrich, St. Louis,
MO, USA)]. Embryos were then incubated for 1 hour in 50 μg/ml RNase A (Qiagen,
Valencia, CA, USA) diluted in DPBS-PVP at 37°C, washed three times in wash buffer
and then incubated at 37°C for 30 min with 3 M HCl/ 0.1% (w/v) PVP. The pH was
neutralized by incubation of embryos for 10 min with 100 mM Tris-HCl, pH 8.5
containing 1% (w/v) PVP followed by three washes with wash buffer.
Nonspecific binding sites were blocked by incubation with DPBS containing 5
mg/ml BSA for 1 h. Embryos were then transferred to a solution of 1 µg/ml anti-5-
methylcytosine (affinity-purified mouse monoclonal antibody; (Calbiochem, Darmstadt,
Germany) diluted in DPBS containing 0.05% (v/v) Tween 20 and 0.01% (w/v) BSA. As

115
a negative control, anti-5-methylcytosine was replaced with an irrelevant mouse IgG1
antibody (Sigma-Aldrich, St. Louis, MO, USA). After 1 h, embryos were washed three
times in wash buffer and transferred to 1 μg/ml fluorescein isothiocyanate (FITC)
conjugated anti-mouse IgG (Abcam, Cambridge, MA, USA). After three washes, nuclei
were labeled by incubation in 50 μg/ml propidium iodide (PI) diluted in DPBS-PVP for 15
min. Embryos were washed three times and mounted with coverslips on slides using
ProLong Gold Anti-Fade mounting medium (Invitrogen, Carlsbad, CA, USA) and
observed under a Zeiss Axioplan epifluorescence microscope (Zeiss, Göttingen,
Germany). Images were acquired using a 40x objective and FITC, blue and rhodamine
filters. The exposure times were constant for all embryos analyzed in an individual
replicate.
Immunofluorescent Labeling for 5-Methylcytosine and CDX2
In one experiment, labeling with antibody to 5-methylcytosine was determined for
ICM and TE using anti-CDX2 to label TE [46]. Embryos were labeled for anti-
methylcytosine as described above, washed six times and transferred to a solution of
antibody against CDX2 (affinity purified mouse monoclonal antibody against CDX-2,
ready to use solution; BioGenex, San Ramon, CA, USA) for 1 h. Embryos were washed
three times and transferred to 10 μg/ml Alexa Fluor 350 labeled goat anti-mouse IgG
diluted in DPBS containing 0.05% (v/v) Tween 20 and 0.01% (w/v) BSA for 1 h.
Embryos were then washed, labeled with PI, mounted on slides and examined as
described above.
Image Analysis Using Imagej
Immunofluorescent intensity was quantified using ImageJ software (version
1.60_41, NIH, Washington DC, USA). For two color images (green for 5-methylcytosine

116
and red for nuclei), individual nuclei were identified by PI labeling and outlined using the
free-hand tool on imageJ. Mean gray intensity of the green and red images were
determined separately. The ratio of intensity for green and red (5-methylcytosine/DNA)
was calculated for each nucleus. Values for all analyzed nuclei from a single embryo
were averaged to obtain the average degree of methylation for that embryo. A similar
process was used for three-color images except that staining for CDX2 was used to
distinguish between TE (blue nuclei) and ICM (no blue labeling) and values were
averaged separately for TE and ICM.
Separation of TE and ICM by Magnetic-Activated Cell Sorting
Magnetic-activated cell sorting was performed as previously described with
modifications [223]. Blastocysts at Day 7 were harvested and incubated in acidic
Tyrode’s solution (Millipore, Billerica, MA, USA) to remove zona pellucidae. Zona
pellucida free blastocysts were washed three times in MACS buffer [DPBS with 0.5%
(w/v) BSA and 2 mM ethylenediaminetetraacetic acid (EDTA), pH 7.2]. Blastocysts
were then incubated for 10 min in concanavalin A conjugated with FITC (Sigma-Aldrich,
St. Louis, MO, USA; ConA-FITC, 1 mg/ml in MACS buffer). Following three washes in
MACS buffer, blastocysts were incubated with 1 μg/ml Hoechst 33342 (Sigma-Aldrich,
St. Louis, MO, USA) in MACS buffer for 3 min. Hoechst 33342 labeled blastocysts were
then washed three times in MACS buffer and incubated in DPBS containing 1 mM
EDTA for 5 min followed by incubation in 0.05% (w/v) trypsin-0.53 mM EDTA solution
(Invitrogen Carlsbad, CA, USA) for 10 min at 38.5°C. Groups of 20-30 blastocysts were
then disaggregated into single blastomeres by vortexing three times for 10 sec each.
Disaggregated blastomeres were transferred into 500 μl DPBS containing 1 mM EDTA
and 10% (v/v) fetal bovine serum (Atlanta Biologicals, Norcross, GA, USA) followed by

117
three washes in MACS buffer by centrifugation at 500 x g for 5 min. Large blastomere
clusters were eliminated by passing the solution through a 0.2 μm cell strainer (BD
Biosciences, San Jose, CA, USA). Single blastomeres that passed through the strainer
were collected, centrifuged at 500 x g for 5 min and resuspended in 110 μl of MACS
buffer. The re-suspended solution was incubated with 10 μl magnetic microbeads
conjugated to mouse anti-FITC (Miltenyi Biotec, Auburn, CA, USA) for 15 min on ice.
Following three washes by centrifugation at 500 x g for 5 min, the magnetic bead
solution was resuspended in 500 μl MACS buffer and passed through MACS separation
columns (Miltenyi biotec, Auburn, CA, USA) attached to a magnetic board (Spherotech,
Lake Forest, IL, USA). The FITC-negative fraction (ICM) was eluted by three 500 μl
MACS buffer washes followed by FITC positive (TE) elution by removing the MACS
separation column from the magnetic board and washing three times with 500 μl MACS
buffer. Purity of eluted ICM and TE using this technique is >91% [223].
Reverse Transcription and Quantitative PCR
Analysis of gene expression was accomplished by quantitative reverse
transcription PCR. Pools of embryos were treated with 0.1% (w/v) proteinase from
Streptomyces griseus to remove the zona pelluicida, washed three times in 50 µl
droplets of DPBS with 1% (w/v) PVP, placed in 100 µl extraction buffer from the
PicoPure RNA isolation kit (Applied Biosystems, Carlsbad, CA, USA) and heated at
42°C for 30 min. Total RNA was extracted using the PicoPure RNA isolation kit
(Applied Biosystems), treated with 2 U of DNase (New England Biolabs, Ipswich, MA,
USA) at 37°C for 30 min to remove DNA, and then incubated at 75°C for 15 min to
denature DNase. DNase-treated RNA was then reverse-transcribed using the High
Capacity cDNA Reverse Transcription Kit (Applied Biosystems). Reverse transcription

118
occurred using random hexamer primers and involved incubation at 25°C for 10 min,
37°C for 120 min and 85°C for 5 min. The cDNA was stored at -20°C until further use.
Negative controls for real-time PCR were also performed by incubation without reverse
transcriptase.
cDNA was utilized for real-time PCR analysis. For detection of transcript levels,
a CFX96 Real-Time PCR detection System (Bio-Rad, Hercules, CA, USA) utilizing
SsoFast EvaGreen Supermix with Low ROX (Bio-Rad). Each reaction contained 1 µl
forward primer (0.5 μM), 1 µl reverse primer (0.5 μM), 10 µl Evagreen Supermix (Bio-
Rad), 6.8 µl H2O and 1.2 µl of cDNA sample (0.6 embryo equivalents). Amplification
conditions were: 95°C for 30 sec, 40 cycles at 95°C for 5 sec, 60°C for 5 sec, and 1
cycle of melt curve analysis at 65-95°C in increments of 0.5°C every 2 sec. The ΔCT
value was determined by subtracting the CT value of the sample by the geometric mean
of the CT for three housekeeping genes, SDHA, GAPDH and YWHAZ [224, 257]. The
primer sequence for DNMT3B (Genbank accession number: AY244711) was 5’-
GGGAAGGAGTTTGGAATAGGAG-3’ and 5’-CGGAGAACTTGCCATCACC-3’ [275].
High Resolution Melting Analysis
The MS-HRM (methylation specific high resolution melting analysis) procedure
was based on Wojdacz and Dobrovic [276]. MS-HRM is a procedure that uses an
intercalcating dye and sensitive thermocycler to detect single nucleotide changes within
an amplicon based on the temperature at which that dsDNA sequence denatures [276,
277]. In the case of methylation analysis, nucleotide changes are induced by bisulfite
conversion of cytosines (but not methylcytosine) to uracil and, following DNA replication,
thymidine.

119
As a preliminary step, 0% and 100% methylated standards were generated for
use in standard curve generation. For the 0% methylation control, bovine embryonic
fibroblast cells derived from skin cells of a fetus at 2-3 months of gestation were
cultured for 1 week in a culture medium composed of 89% (v/v) Dulbecco’s Modified
Eagle’s medium (Invitrogen), 10% (v/v) fetal bovine serum (Atlanta Biologicals,
Norcross, GA, USA), 1% (v/v) of a 100 IU/mL penicillin, 100 μg/ml streptomycin sulfate
and 250 ng/ml amphotericin B solution (Invitrogen) and 12 μM 5-azacytidine (to inhibit
DNA methylation). The concentration of 5-azacytidine used was chosen because it was
the highest concentration that did not cause complete loss of cell growth. Other cells
were cultured without 5-azacytidine. Cells were collected, stored at -80°C until
extraction of DNA and RNA extraction using the AllPrep DNA/RNA Mini kit (Qiagen,
Valencia, CA, USA). gDNA and total RNA were stored at -80°C in extraction buffer and
RNase-free water, respectively, until ready for use. A sample of 100% methylated DNA
was prepared by treating gDNA from fibroblasts cultured in the absence of 12.5 μM 5-
azacytidine with SssI methylase (New England Biolabs, Ipswich, MA, USA) for 1 hour at
37°C. gDNA was then treated with bisulfite using the Imprint DNA modification kit
(Sigma-Aldrich, St. Louis, MO, USA) and frozen at -20°C until further use. Samples of 0
and 100% methylated gDNA were mixed together to create samples of 0, 5, 25, 50, 75
and 100%, methylated gDNA.
High resolution melting analysis was performed using a BioRad C1000 thermal
cycler with CFX96 Real-time system and Precision Melt analysis software (BioRad,
Hercules, CA, USA). PCR conditions were set to the following: 95°C for 2 min, 45
cycles of 95°C for 10 sec, 60°C for 30 sec, 72°C for 30 sec, and heteroduplex formation

120
at 95°C for 30 sec and 60°C for 1 min. Subsequently, high resolution melting was
performed at 0.2°C increments from 65°C to 95°C. Primers were designed using Methyl
Primer Express 1.0 (Applied Biosystems, Carlsbad, CA, USA) with settings to include at
least one nucleotide toward the 5’ end that could bind to both bisulfite-converted and
non-bisulfite converted sequences to decrease bias [278]. Primers were designed to
amplify an intronic 81 bp sequence containing 5 CpG. This region of DNMT3B was
chosen based on the estimation of a CpG island as determined by Methyl Primer
Express (Applied Biosystems, Carlsbad, CA, USA). The primer sequence for HRM
DNMT3B (Genbank accession number NC_007311) were as follows: 5’-
GGAYGGGTTTTAGGTTTGGGTATT-3’ and 5’-CAAACCCCCRCAAAATAATTCT-3’.
Primers were generated by Integrated DNA Technologies (Coralville, Iowa, USA).
A standard curve was run for each PCR procedure using gDNA of 0-100% methylation
and prepared as described above. The difference relative fluorescence units (DRFU)
were recorded at the melting peak for each sample. All samples were analyzed in
duplicate. The relationship between methylation percent and DRFU was exponential.
The standard curve was calculated and used to obtain the degree of methylation for
samples.
Design of Experiments
The first experiment was conducted to determine developmental changes in DNA
methylation. Embryos were fertilized with X-sorted spermatozoa (Genex Cooperative,
Shawano, WI, USA, Accelerated Genetics, Baraboo, WI, USA and Select Sires, Plain
City, OH, USA) from three separate bulls and then placed into culture with SOF-BE1.
Separate drops of embryos were cultured to allow harvest of 2-cell embryos [28-32
hours post-insemination (hpi)], 4-cell embryos (52-56 hpi), 6-8 cell embryos (76-80 hpi),

121
9-25 cell embryos (104 hpi), 32 cell-morula stage embryos (148-152 hpi) and
blastocyst-stage embryos (176 hpi). Embryos were analyzed for DNA methylation by
labeling with anti-5-methylcytosine and PI as described above. The experiment was
replicated on 6 occasions and a total of 11-24 embryos per stage were subjected to
labeling for anti-5-methylcytosine.
The effect of sex on DNA methylation was determined by fertilizing oocytes with
either X-sorted spermatozoa or Y-sorted spermatozoa (Accelerated Genetics, Baraboo,
WI, USA) from the same bull, using three separate bulls for each replicate. Embryos
were then cultured and harvested from separate drops at either the 6-8 cell stage (76-
80 hpi) or blastocyst stage (176 hpi) for analysis of labeling with anti-5-methylcytosine.
The experiment was replicated on 3 occasions and a total of 11-21 embryos per stage
were subjected to labeling for anti-5-methylcytosine.
Effects of CSF2 and cell lineage (ICM and TE) on DNA methylation in the
blastocyst were tested as follows. Oocytes were fertilized with either X-sorted
spermatozoa or Y-sorted spermatozoa (Accelerated Genetics, Baraboo, WI, USA) from
the same bull, using three separate bulls for each replicate. Embryos were then placed
in 45 µl drops of SOF-BE1 and cultured for 7 d. At Day 5 pi, 5 µl of 100 ng/ml
recombinant bovine CSF2 (Novartis) in vehicle [DPBS and 1% (w/v) BSA] or vehicle
alone was added. The concentration of CSF2 was chosen based on its effectiveness
for increasing development to the blastocyst stage and embryonic survival after transfer
into recipient females [24]. Blastocysts were harvested at Day 7 pi and subjected to
labeling with anti-5-methylcytosine, anti-CDX2 and PI. The experiment was replicated

122
on 3 occasions and a total of 14-26 embryos per stage were subjected to labeling for
anti-5-methylcytosine.
Developmental changes in expression of DNMT3B were determined in another
experiment. Embryos were fertilized with conventional semen (Androgenics, Oakdale,
CA, USA and Nebraska Bull Service, McCook, NE, USA) using three different bulls for
each replicate and then placed into culture with SOF-BE1. Separate drops of embryos
were cultured to allow collection of groups of 30 zygotes (0 hpi), 2-cell (28-32 hpi), 3-4
cell (48 hpi), 5-8 cell (56-60 hpi), 9-16 cell (72 hpi), >16 cell (120 hpi) and blastocyst
stage (168 hpi) embryos. RNA was extracted as described above and used for RT-
PCR. The experiment was replicated on 5 occasions.
Methylation of an intronic region of DNMT3B was determined in two experiments.
In the first experiment, embryos were collected at the 6-8 cell stage (76-80 hpi) and at
the blastocyst stage (176 hpi). In the second, blastocysts were harvested at 176 hpi and
ICM and TE isolated using MACS. In both experiments, embryos were fertilized with
conventional semen (Androgenics, Oakdale, CA, USA and Nebraska Bull Service,
McCook, NE, USA) using three different bulls for each replicate and then placed into
culture with SOF-BE1. In the first experiment, embryos were collected in groups of 18-
48, treated to remove the zona pellucida and snap-frozen in liquid N2 before storage at -
80°C until ready for use in MS-HRM analysis. A total of 3 pools of embryos were
collected at each stage of development. In the second experiment, groups of 20-40
blastocysts were subjected to the MACS procedure to isolate ICM and TE and cells
frozen in 20 µl of MACS buffer (DPBS with 0.5% (w/v) BSA and 2 mM EDTA, pH 7.2) at

123
-80°C until ready for MS-HRM. The experiment was replicated with 8 pools of ICM and
TE.
Statistical Analysis
Data were analyzed by least-squares analysis of variance using the GLM
procedure of the Statistical Analysis System version 9.3 (SAS Institute Inc., Cary, NC,
USA). Depending on the experiment, sources of variation included replicate, stage,
sex, cell type (ICM and TE), treatment (CSF2 or vehicle) as well as all interactions.
Replicate was considered a fixed effect. For real-time PCR results, treatment effects
were tested using ΔCT values and graphed as fold-change differences relative to values
for oocytes. All data are shown as least-squares means ± standard error of the means.
Results
Changes in DNA Methylation during Development to the Blastocyst Stage
Representative images of labeling for 5-methylcytosine from the 2-cell stage
through blastocyst development are presented in Figure 4-1 while results from
quantitative analysis are presented in Figure 4-2. There was an effect of stage of
development on immunoreactive 5-methylcytosine (P<0.0001), with amounts
decreasing from a peak at the 2-cell stage to a nadir at the 6-8 cell stage. Thereafter,
immunoreactivity increased to the blastocyst stage.
Effect of Sex on DNA Methylation
Embryos were fertilized separately with X and Y sorted sperm from the same bull
to produce either female or male embryos. DNA methylation was examined at either
the 6-8 cell or blastocyst stages of development by evaluating labeling with anti-5-
methylcytosine. Results are shown in Figure 4-3. DNA methylation was greater for
embryos at the blastocyst stage than at the 6-8 cell stage (stage of development;

124
P<0.0001). There were, however, effects of sex (P=0.0029) and sex x stage
(P=0.0007). The interaction occurred because female embryos had more intense
labeling for 5-methylcytosine than male embryos at the 6-8 cell stage but lower labeling
than male embryos at the blastocyst stage.
DNA Methylation in Blastocysts as Modulated by Sex, CSF2 and Cell Differentiation
Use of CDX2 to distinguish between ICM and TE revealed that fluorescent
activity for immunoreactive 5-methylcytosine was lower in ICM than TE. A
representative blastocyst is shown in Figure 4 and quantitative analysis is summarized
in Figure 5. As seen in the previous experiment, DNA was less methylated in female
embryos than male embryos (P<0.02) and this was apparent in both the ICM and TE
compared to the TE (Figure 4-5). There was no effect of CSF2 or interactions of CSF2
on degree of methylation in either the ICM (0.53 ± 0.09 vs. 0.46 ± 0.07 for CSF2 and
vehicle, respectively) or TE (1.17 ± 0.09 vs. 1.18 ± 0.07 for CSF2 and vehicle,
respectively).
Developmental Changes in DNMT3B Gene Expression
To determine whether developmental changes in DNA methylation could
conceivably be caused by changes in DNMT3B activity, changes in expression of
DNMT3B from the zygote to the blastocyst stage were determined (Figure 4-6). Stage
of development affected (P<0.0001) steady-stage amounts of DNMT3B mRNA.
Expression increased between the zygote stage and 2-cell stage and then declined to a
nadir for embryos at the >16 cell stage of development. Thereafter, steady-state
concentrations of DNMT3B increased at the blastocyst stage.

125
Developmental Changes in Methylation of DNMT3B
The possible role of methylation in controlling developmental changes in
expression of DNMT3B was evaluated by determining changes in the degree of
methylation in an 81 bp CpG rich intronic region of DNMT3B (Figure 4-7A). A
representative standard curve is shown in Figure 4-7B and representative results in
Figure 4-8A. Methylation in the analyzed region, which contains 5 CpGs, was higher
(P<0.0001) for 6-8 cell embryos than for embryos at the blastocyst stage (Figure 4-8B).
When ICM and TE were separated by MACS, there was no significant difference
(p=0.25) in methylation of DNMT3B between ICM (70.2 ± 9.8%) or TE (56.3 ± 8.6%).
Discussion
Epigenetic control of gene expression is an important aspect of early embryonic
development. Using the preimplantation bovine embryo as a model, it was observed
that dynamic changes in DNA methylation occur throughout development in a manner
that depends on sex and cell type. Changes in DNA methylation were associated with
similar changes in expression and methylation of DNMT3B, indicating that, as for the
mouse [82], this gene may play an important role in modification of the embryonic
methylome.
There is a steady decrease in overall level of DNA methylation as the mouse
embryo undergoes successive cell division [75]. A similar loss occurs in the developing
sheep embryo [85] and as, shown here, the bovine embryo (Figure 4-2). In contrast,
there appears to be no widespread demethylation during early cleavage divisions in the
pig [86] or rabbit [274]. In the mouse, the loss of methylation during early development
occurs as a result of both passive [279] and active [280] demethylation. Passive
demethylation occurs when DNA replication occurs without the presence of DNMT1 to

126
methylate the newly replicated strand [281] while active DNA demethylation is thought
to occur by base excision repair mechanisms utilizing glycolysis or deamination of 5-
methylcytosine although the exact enzymes responsible are still unknown in mammals
[282].
The bovine embryo is also similar to the mouse embryo in that the period of
demethylation is followed by a period of de novo methylation (Figure 4-2). In the cow,
methylation begins following the 6-8 cell stage and is coincident with activation of the
embryonic genome [283]. Re-establishment of methylation marks on DNA in the mouse
embryo is accomplished by the de novo methyltransferases Dnmt3a and Dnmt3b [82].
Knockout of Dnmt3b caused incomplete methylation of pluripotent genes in the mouse
embryo [82, 284]. Current results implicate DNMT3B as involved in de novo DNA
methylation in the cow based on the observation that steady-state amounts of mRNA for
DNMT3B parallels the pattern of methylation (Figures 4-2 and 4-6). In fact, DNMT3B
itself might be under the control of methylation because methylation of an intronic region
within DNMT3B was lower at the blastocyst stage than at the 6-8 cell stage (Figure 4-
8B). The role of methylation in DNMT3B expression can be further clarified through
experiments to detail the methylation landscape of DNMT3B more fully as well as the
consequences of changes in methylation on DNMT3B expression. It is also likely that
there is regulation of DNMT3B transcription independent of DNA methylation.
Developmental patterns of methylation in the preimplantation embryo diverge
between the mouse and cow at the blastocyst stage of development. In the mouse, the
ICM is more highly methylated than the TE [76] while current results indicate the ICM is
less methylated than TE in the cow (Figure 4-4). Indeed, there is much divergence

127
between species in overall levels of methylation in the ICM and TE, with the sheep and
pig showing a pattern similar to the mouse [85, 86] and with the ICM less methylated
than the TE in the rabbit [274, 285]. One implication of the species variability in DNA
methylation patterns in the blastocyst is that there is likely to be significant differences
between species in the genes that are differentially regulated between ICM and TE.
One example is OCT4 (i.e, POU5F1). In the mouse, Oct4 is strictly expressed in the
ICM while OCT4 in cattle is expressed in both ICM and TE [94]. A genome-wide study
of the bovine ICM and TE revealed genes that exhibited different patterns of differential
expression than is the case in mice or humans [286].
Sex of the bovine embryo is an important factor that affects development as early
as the first cleavage, when the sex ratio is skewed towards males [50]. SRY is
expressed as early as the 8-cell stage of development [287], more male embryos than
female embryos develop past the 8-cell stage of development [288], and male embryos
become blastocysts at a more rapid rate than females [50, 288, 289]. At the blastocyst
stage, gene expression is affected by sex, with the overall level of transcription being
greater for female embryos [245]. Methylation is likely to play a role in establishing sex
patterns of transcription. Earlier studies reported differences in degree of methylation of
specific genes at the blastocyst stage [8, 268, 290]. Here we show that the overall level
of DNA methylation at the blastocyst stage was lower for female embryos than male
blastocysts (Figure 4-3). Such a sex difference would be expected if DNA methylation is
important for the higher transcript abundance for female blastocysts [245]. Earlier in
development, at the 6-8 cell stage, DNA is more methylated for female embryos (Figure
4-3), probably because the rate of demethylation was less for female embryos.

128
DNA methylation can be altered by development in vitro [8]. Thus, it will be
necessary to determine the extent to which changes in DNA methylation caused by
stage of development, cell lineage and sex seen here also occur for embryos
developing in vivo. Moreover, it is possible that regulatory molecules such as CSF2
[18], insulin-like growth factor-1 [129] and hyaluronan [21] that improve competence of
the in vitro produced embryo to survive after transfer into recipients could exert their
action, at least in part, by regulating DNA methylation. This hypothesis was tested in the
present experiment for the maternally derived cytokine CSF2 [27]. There was no effect
of CSF2 treatment on the overall level of DNA methylation at the blastocyst stage
(Figure 4-3C) so actions of CSF2 on the embryo that enhance its survival probably do
not depend upon broad changes in DNA methylation. Nonetheless, it is possible that
CSF2 causes changes in methylation of a more narrow set of genes. There are long-
term consequences of embryonic treatment with CSF2 (decreased pregnancy loss after
Day 35 of gestation [18]) that could represent epigenetic programming.
In conclusion, preimplantation development of the bovine embryo is
characterized by dynamic changes to DNA methylation that are dependent upon sex
and cell lineage. In particular, global methylation declines to a nadir at the 6-8 cell
stage and increases thereafter. Methylation is lower for female embryos than male
embryos at the blastocyst stage and lower for the ICM than TE. The developmental
pattern of DNA methylation in the cow is partially representative of events in the mouse,
with the major difference being in the relative degree of methylation in ICM and TE. Like
the mouse, changes in expression of DNMT3B may be responsible for developmental

129
changes in DNA methylation because levels of methylation are related to expression of
DNMT3B.
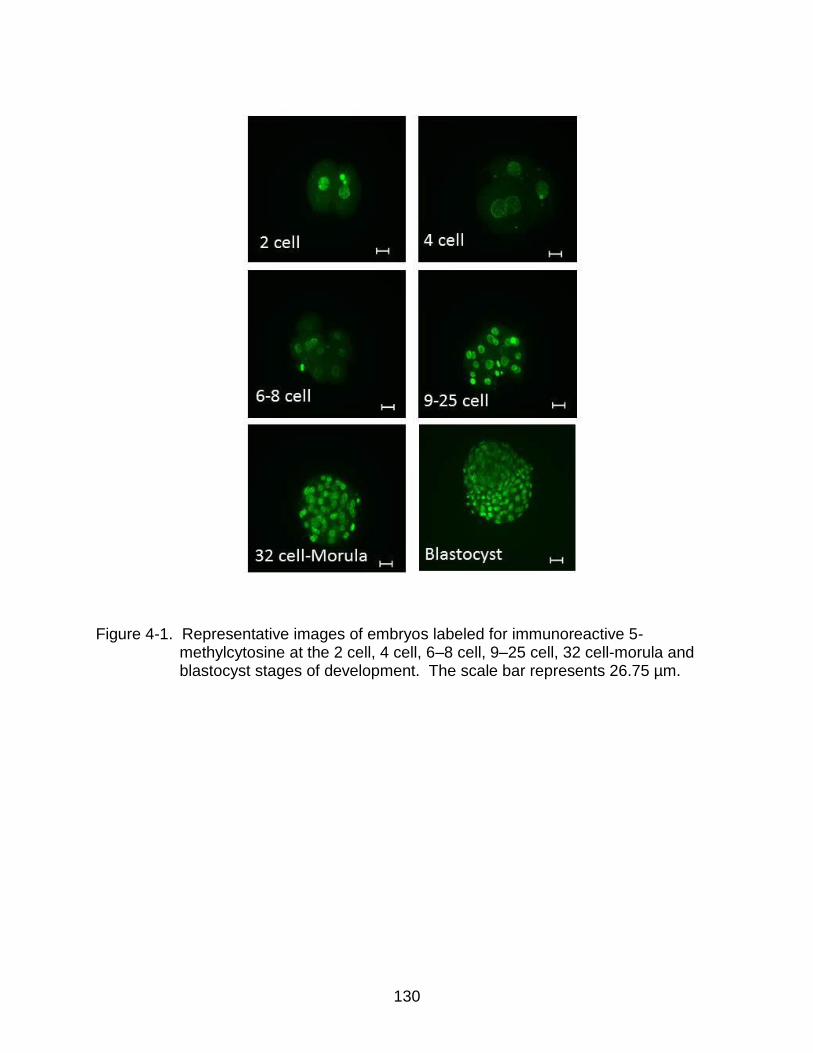
130
Figure 4-1. Representative images of embryos labeled for immunoreactive 5- methylcytosine at the 2 cell, 4 cell, 6–8 cell, 9–25 cell, 32 cell-morula and blastocyst stages of development. The scale bar represents 26.75 µm.
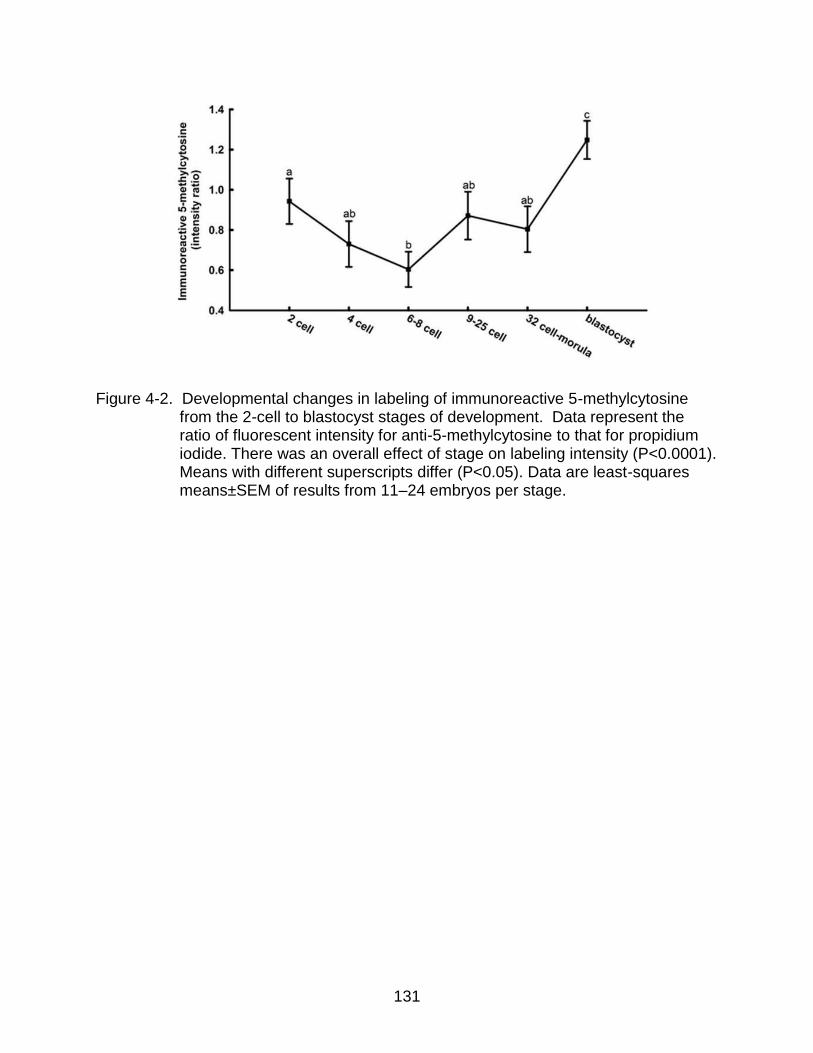
131
Figure 4-2. Developmental changes in labeling of immunoreactive 5-methylcytosine
from the 2-cell to blastocyst stages of development. Data represent the ratio of fluorescent intensity for anti-5-methylcytosine to that for propidium iodide. There was an overall effect of stage on labeling intensity (P<0.0001). Means with different superscripts differ (P<0.05). Data are least-squares means±SEM of results from 11–24 embryos per stage.
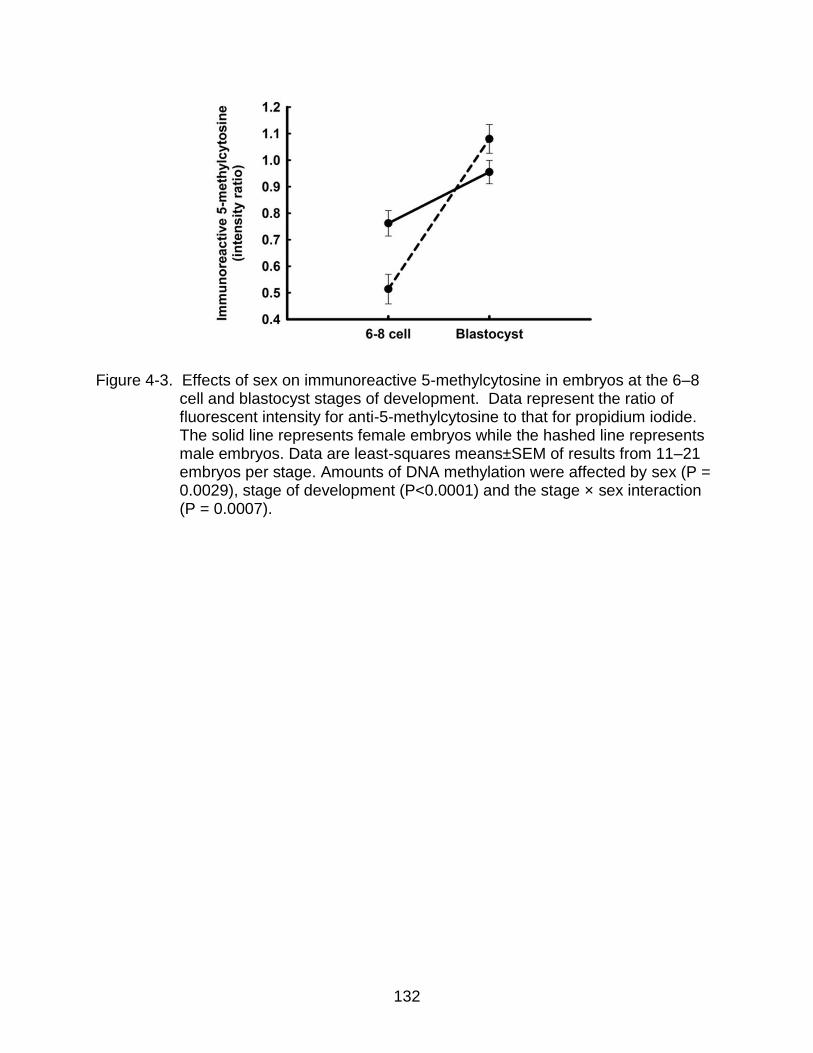
132
Figure 4-3. Effects of sex on immunoreactive 5-methylcytosine in embryos at the 6–8
cell and blastocyst stages of development. Data represent the ratio of fluorescent intensity for anti-5-methylcytosine to that for propidium iodide. The solid line represents female embryos while the hashed line represents male embryos. Data are least-squares means±SEM of results from 11–21 embryos per stage. Amounts of DNA methylation were affected by sex (P = 0.0029), stage of development (P<0.0001) and the stage × sex interaction (P = 0.0007).
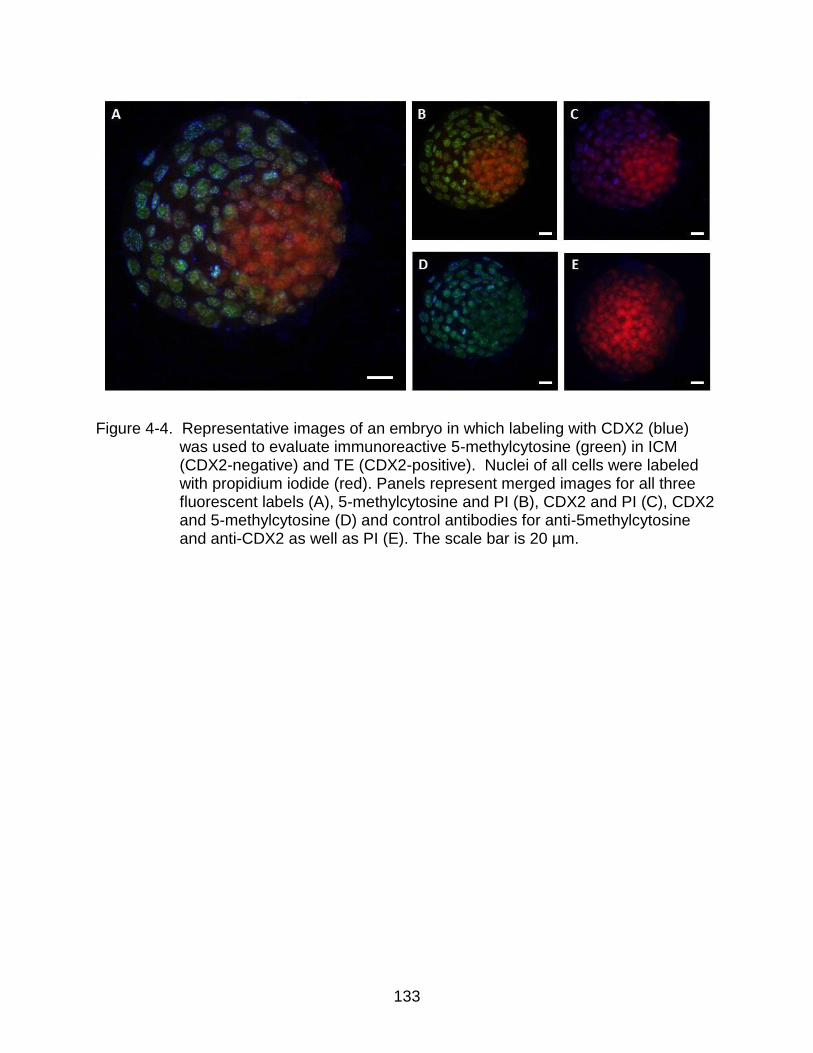
133
Figure 4-4. Representative images of an embryo in which labeling with CDX2 (blue)
was used to evaluate immunoreactive 5-methylcytosine (green) in ICM (CDX2-negative) and TE (CDX2-positive). Nuclei of all cells were labeled with propidium iodide (red). Panels represent merged images for all three fluorescent labels (A), 5-methylcytosine and PI (B), CDX2 and PI (C), CDX2 and 5-methylcytosine (D) and control antibodies for anti-5methylcytosine and anti-CDX2 as well as PI (E). The scale bar is 20 µm.
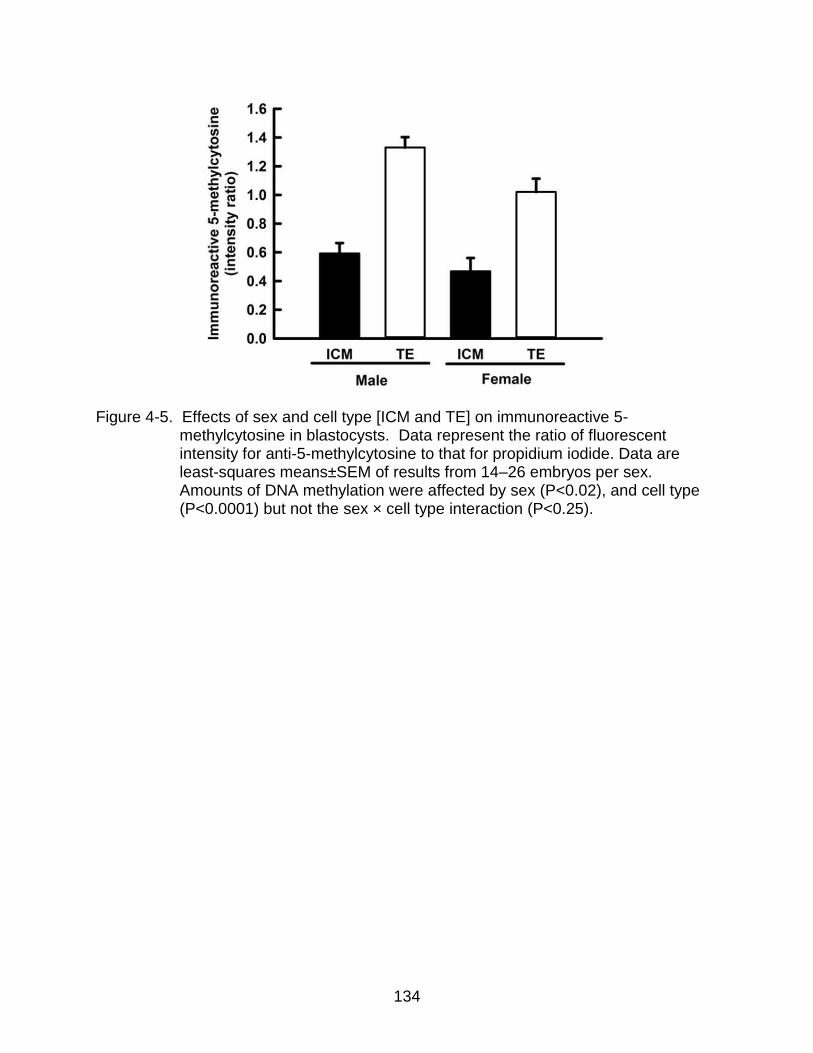
134
Figure 4-5. Effects of sex and cell type [ICM and TE] on immunoreactive 5-
methylcytosine in blastocysts. Data represent the ratio of fluorescent intensity for anti-5-methylcytosine to that for propidium iodide. Data are least-squares means±SEM of results from 14–26 embryos per sex. Amounts of DNA methylation were affected by sex (P<0.02), and cell type (P<0.0001) but not the sex × cell type interaction (P<0.25).
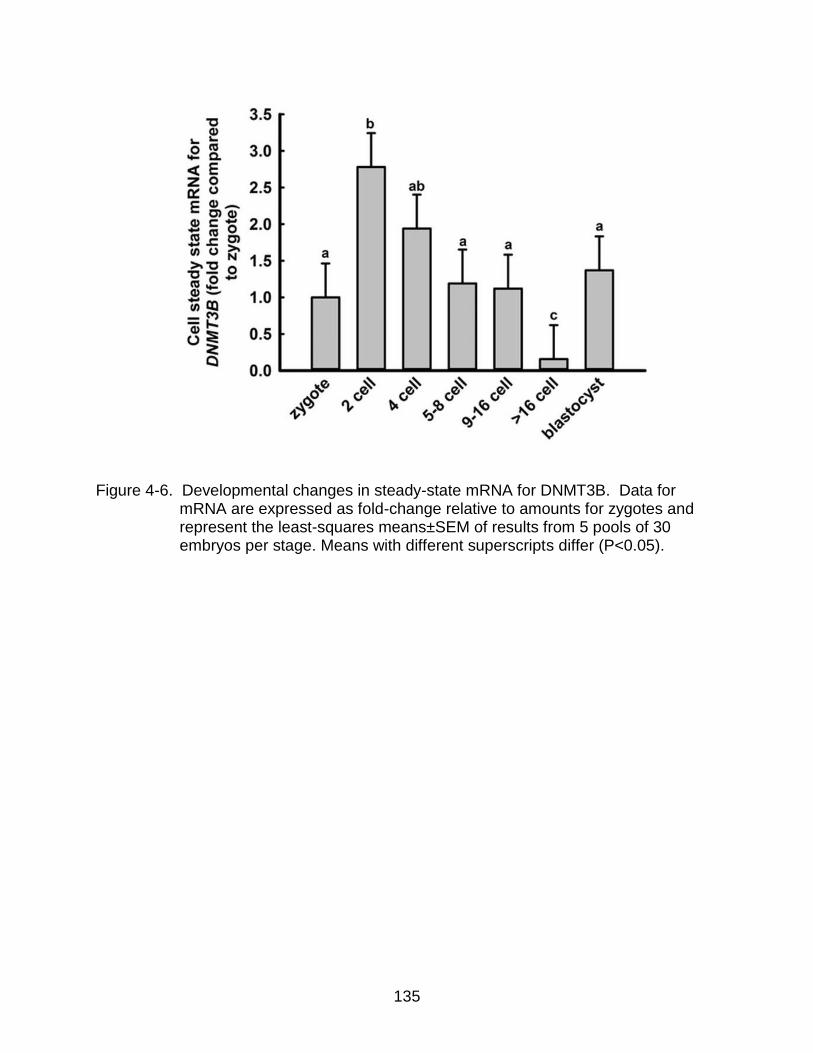
135
Figure 4-6. Developmental changes in steady-state mRNA for DNMT3B. Data for
mRNA are expressed as fold-change relative to amounts for zygotes and represent the least-squares means±SEM of results from 5 pools of 30 embryos per stage. Means with different superscripts differ (P<0.05).
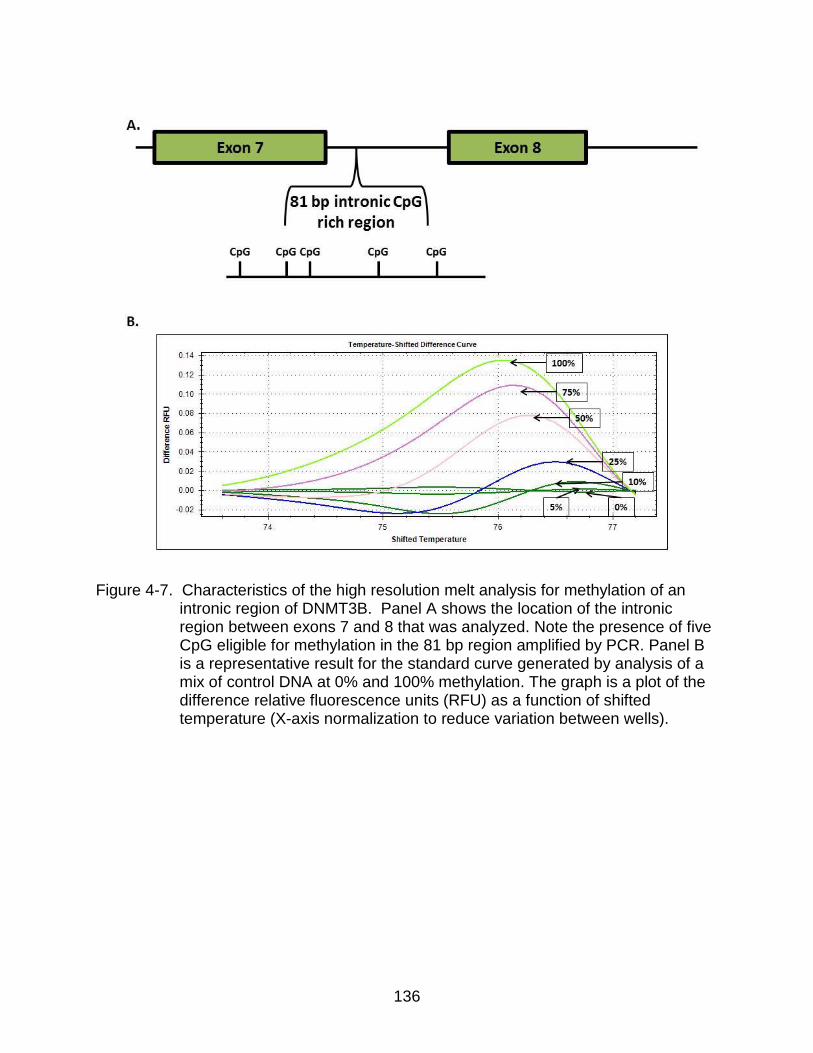
136
Figure 4-7. Characteristics of the high resolution melt analysis for methylation of an
intronic region of DNMT3B. Panel A shows the location of the intronic region between exons 7 and 8 that was analyzed. Note the presence of five CpG eligible for methylation in the 81 bp region amplified by PCR. Panel B is a representative result for the standard curve generated by analysis of a mix of control DNA at 0% and 100% methylation. The graph is a plot of the difference relative fluorescence units (RFU) as a function of shifted temperature (X-axis normalization to reduce variation between wells).

137
Figure 4-8. Developmental changes in DNA methylation for DNMT3B. Panel A is a
representative result of analysis of samples from 6–8 cell embryos (lime green) and blastocysts (red). Panel B shows the degree of methylation between the two stages. Data represent least-squares means±SEM of results from 3 pools of 23–48 embryos per stage. Methylation percent was higher at the 6–8 cell stage than at the blastocyst stage (P<0.0001).

138
CHAPTER 5 GENERAL DISCUSSION
The maternal environment is a critical component for the successful development
of the embryo. Alterations in this microenvironment during preimplantation development
can alter the developmental fate of the fetus, with potential consequences that extend
into post-natal life [13, 15, 99, 291]. It was the assumption of this dissertation that
effects of the maternal environment on the developmental trajectory of the
preimplantation embryo are mediated, at least in part, by altered secretion of regulatory
molecules from the endometrium that affect development. Indeed, a variety of
molecules produced by the endometrium have been identified that can affect
development to the blastocyst stage and competence of the embryo to establish
pregnancy. In the cow, these embryokines, include IGF1 [292], IL1 [23], hyaluronan
[21], TGFB [20], LIF [20] and FGF2 [22]. The most well studied embryokine is CSF2,
which has been shown to have a variety of effects on the preimplantation embryo of
cattle [24], mice [26], pigs [189], and humans [25] including improved ability to develop
to the blastocyst stage [24, 26, 189, 200], increased ICM numbers [18], resistance to
apoptosis [31, 187], altered gene expression [30] and improved competence to
establish pregnancy when embryos are transferred to females [18, 25, 26]. The
purpose of the research described in this dissertation, was to identify mechanisms by
which CSF2 increases the competence of the bovine blastocyst to establish pregnancy.
The conclusions derived from the dissertation are summarized in Figure 5-1. It
was demonstrated in Chapter 4 that early preimplantation development is characterized
by a loss in DNA methylation to a nadir at the 8-16 cell stage. At this time, the female
embryo maintains a higher level of methylation than the male. Subsequently,

139
methylation is increased but in a pattern where, at the blastocyst stage, the TE is more
methylated than the ICM. At this stage, DNA is less methylated for the female than the
male. Sex effects on methylation may be important for actions of CSF2, which occur in
a sex-specific manner. At the blastocyst stage, CSF2 affects characteristics of the ICM
so that its ability to survive passage and remain pluripotent when isolated from TE is
increased (Chapter 2). The mechanisms are unclear but, by the blastocyst stage, the
program of the embryo has been altered by CSF2 differently in female embryos than in
male embryos (Chapter 3). Differential programming is apparent when examining
development of embryos after transfer to recipients. There is no size difference between
female and male embryos at Day 14 of development but, by Day 15, control female
embryos are larger and secrete more IFNT than male embryos. This phenomenon
probably represents earlier elongation for females. Moreover, CSF2 alters the
differences between sexes by decreasing size and IFNT secretion for females but
increasing these characteristics for males. In addition, CSF2 programs the
transcriptome and methylome at day 15 in a sex-specific manner. This observation
leads to the possibility that sex-specific developmental programming [14-17, 169]
occurs because alterations in the maternal milieu during the preimplantation period in
response to changes in environment cause sex-specific effects on the embryo
While several molecules have been shown to increase the competence of an
embryo to establish pregnancy after transfer, including CSF2 [18], IGF1 [292] and
hyaluronan [21], we know nothing of the mechanism by which embryo competence is
increased. Several genes have been identified whose expression at the blastocyst
stage differs between embryos that subsequently establish pregnancy versus those that

140
do not [293], and it may be that certain embryokines alter gene expression in a manner
than improves embryo competence. However, none of the genes that were differentially
expressed between embryos establishing pregnancy or not were found to be regulated
by CSF2 at the morula stage [31]. Results from Chapter 2 suggest that one possible
way CSF2 improves embryo competence for survival is regulation of pluripotency and
capacity for survival of the ICM. Such an action, if exerted by CSF2, could potentially
have a large effect on embryonic survival because 20-25% of in vitro produced embryos
lose the ED derived from the ICM by Day 14-15 of gestation [18, 225, 226]. Perhaps
CSF2 increases pregnancy rate in embryo transfer recipients by reducing loss of the
ED. The evidence for CSF2 regulation of the ICM is based on experiments showing that
CSF2 increases ICM cell number [18], and that isolated ICM cultured on mitomycin C
treated fibroblasts were more likely to survive passage while maintaining pluripotency if
embryos were exposed to CSF2 from Day 5-7 or 6-8 of development (Chapter 2).
The mechanism by which CSF2 affects the ICM is not known. One possibility is
that CSF2 reduced induction of apoptosis. CSF2 can blunt induction of apoptosis by
heat shock in the bovine embryo [31]. Another possibility is that CSF2 regulates
expression of genes associated with pluripotency. Expression for pluripotency related
genes NANOG and SOX2 in isolated ICM were not affected by CSF2, however
(Chapter 2). However, morula-stage embryos collected after 24 hours of CSF2
treatment did contain differentially expressed genes related to pluripotency [31].
Another possible mechanism by which CSF2 could increase embryo competence
to establish pregnancy is through alteration of the methylome of the embryo. As shown
in Chapter 3, some effects of CSF2 treatment from Day 5-7 of development are not

141
realized until several days later (Day 15 in this case) and such effects are consistent
with epigenetic modification. Earlier, Loureiro et al (2011) found a similar long-term
effect of CSF2 since embryos that established pregnancy were less likely to be lost
before term if treated with CSF2 from Day 5-7. Even though there was no effect of
CSF2 on the overall level of methylation in the embryo at the blastocyst stage of
development (Chapter 4), the technique used to assess methylation (labeling with anti-
5-methylcytosine) is crude and it is possible that specific regions of DNA undergo
differential methylation in response to CSF2. Use of the methylome array described in
Chapter 3 could prove effective at examining whether specific CpG undergo differential
methylation at the blastocyst stage in response to CSF2.
One implication of the findings that CSF2 increases survival of isolated ICM is
that CSF2 might be a useful additive to culture media for production of ESCs in the cow.
ESCs from the cow are difficult to keep in the pluripotent state. Regulatory molecules
utilized to maintain pluripotency of mouse [229] or human [228] ESC cultures are
ineffective for that purpose in cow [230]. It is possible that the addition of CSF2 to
culture media could improve the survival of bovine stem cells in an undifferentiated
state.
As has been found earlier for the human [187], the bovine preimplantation
embryo expresses CSF2RA while not expressing CSF2RB. CSF2 binds to CSF2RA,
which combines with the CSF2RB subunit to signal through JAK2/STAT5ab [181].
CSF2RA is capable of signaling in the absence of the CSF2RB subunit, but does so a
much lower affinity [182]. Given that CSF2 can alter the properties of the bovine embryo
despite the absence of CSF2RB mRNA, it is likely that CSF2 signals through a separate

142
pathway other than the JAK-STAT signaling pathway regulated by the CSF2RA-
CSF2RB dimer. The lack of the CSF2RB subunit implies that the embryo does not have
high affinity signaling. However, the dosage of 10 ng/ml of CSF2 used in these
experiments, is below the Kd of the low affinity CSF2RA. In the absence of the CSF2RB
subunit, human myeloid cells are still able to signal through the PI3K pathway [183].
Perhaps a similar pathway is operative in blastomeres of the preimplantation embryo.
As has been alluded to many times in this dissertation, one important rationale
for studying regulation of embryonic development is that the embryo produced in vitro is
abnormal in many respects [3-5, 7, 8] and is less capable of establishing pregnancy
when transferred to recipients [99]. Identification of maternal-derived molecules that
enhance embryonic competence could be useful to improve effectiveness of embryo
transfer using in vitro produced embryos. Few laboratories that study embryonic
development in the cow have the opportunity to transfer embryos to recipients and,
instead use in vitro measures of development, such as the percent of oocytes becoming
blastocysts, as an indicator of the adequacy of culture systems. Results in this
dissertation point out the inadequacy of this approach. There was no consistent effect of
CSF2 on the percent of embryos that progressed to the blastocyst stage of
development. Nonetheless, CSF2 can improve the pregnancy rate after transfer to
recipients in cattle [18], mice [26] and humans [200]. This change in pregnancy rate
after transfer without affecting the percent of embryos that develop to the blastocyst
stage is also true for other molecules such as IGF1 [129] and hyaluronan [21]. In
consequence, optimization of culture systems for production of embryos in vitro will

143
require transfer of embryos to recipient females or development of other in vitro indices
of blastocyst quality more predictive of embryo survival than percent blastocyst.
Perhaps the most important finding of this dissertation was the unexpected
observation that exposure of the morula and blastocyst stage embryo to CSF2 alters the
developmental program in a sex-specific manner (Chapter 3). One implication is that
sexual differentiation occurs as early as Day 5-7. This is to be expected because gene
expression differs significantly between male and female embryos at the blastocyst
stage [245] and other results from this dissertation show differences in global DNA
methylation between female and male embryos as early as the 8-cell stage (Chapter 4).
One possibility is that CSF2 uses signal transduction pathways that differ in
important respects between males and females. While the signaling pathway for CSF2
in the embryo has not been identified, and is likely different than the classical JAK-STAT
pathway (Chapter 2), it is possible that there is more than one pathway and that the
relative activity of one or more varies between male and female embryos. Experiments
to elucidate signaling pathways for CSF2 in the embryo should be conducted to
determine whether there are sex differences. Another possible reason for sex
differences in response to CSF2 would be differences in transcriptional repression of
specific regions of DNA by epigenetic modification. At the blastocyst stage, the male
embryo is methylated at a higher intensity than the female (Chapter 4) while the female
is transcriptionally more active than males at the same stage [245]. Interestingly, over
50% of the genes regulated by CSF2 are also regulated by sex. Thus, CSF2 acts
preferentially on genes that are controlled directly or indirectly by transcription factors
involved in sexual differentiation. There may also be sex-specific methylation of genes

144
encoding transcription factors or enhancers. At least in part, genes regulated by sex
may have transcription factors or regulatory elements that are also regulated by CSF2.
The observation on sex differences in response to CSF2 is important because it
provides a mechanism for the phenomenon by which developmental programming
affects female embryos differently than male embryos. Perhaps changes in nutrition or
stress that the mother is subjected to in her environment lead to alterations in the
embryokine secretion in the uterus, thus preparing the embryo for post-natal life in a sex
specific manner. In addition, this observation implies that much of early development
much be examined in a sex-specific context.
For the phenomenon of sex-specific response to CSF2 to be generally relevant
to preimplantation developmental programming, it should be that the phenomenon
occurs for more than one embryokine and in species other than the cow. Other
embryokines, such as IGF1 [292], IL1B [23], hyaluronan [21], TGFB [20], LIF [20] and
FGF2 [22], have been shown to improve the development of the embryo in the cow, but
it has not been examined whether response of the embryo to these molecules varies
with sex. We do not know whether sex bias responses to embryokines occurs in other
species but at least the embryokine examined, CSF2, causes changes in development
in a wide variety of species, including mice [26], pigs [189] and humans [200].
CSF2 is a good candidate for mediating certain environmental effects on
developmental programming because its expression in the uterus depends on diet and
inflammatory status. In particular, obese cows had lower CSF2 protein and mRNA
present in the oviduct than non-obese cows [34]. Uterine inflammation caused by the
presence of semen, increases CSF2 protein secretion due to TGFB within the seminal

145
plasma [32, 33]. Understanding how CSF2 is regulated by diet may prove useful for
minimizing the effects of undernutrition on fertility [294] as well as on the characteristics
of the resulting offspring.
One implication of the finding that CSF2 affected female embryos differently than
male embryos is that research on regulation of preimplantation development should
now consider the possibility of differences between female and male embryos.
Heretofore, there has been little consideration of embryo sex when trying to understand
preimplantation development, which should change.
Findings from this dissertation provide the groundwork for understanding 1) the
signal transduction system for CSF2 in the preimplantation embryo, 2) mechanisms by
which CSF2 affects ICM survival and pluripotency, 3) and how developmental
programming occurs in a sex-specific manner during preimplantation development.
Among the promising areas of future research would be to address the mechanism by
which CSF2 programs the embryo in a sex specific manner (whether due to sex effects
on signal transduction, transcription factor usage or epigenetic modification) as well as
the mechanisms for actions of CSF2 on survival and pluripotency of the ICM.

146
Figure 5-1. Schematic illustration of conclusions derived from the dissertation including developmental changes in methylation and sex-specific actions of CSF2 on embryonic development. Early preimplantation development is characterized by a loss in DNA methylation to a nadir at the 8-16 cell stage. At this time, the female embryo maintains a higher level of methylation than the male. Subsequently, methylation is increased but in a pattern where, at the blastocyst stage, the TE is more methylated than the ICM. At this stage, DNA is less methylated for the female than the male. Sex effects on methylation may be important for actions of CSF2 which occur in a sex-specific manner. At the blastocyst stage, CSF2 affects characteristics of the ICM so that its ability to survive passage and remain pluripotent when isolated from TE is increased. The mechanisms are unclear but, by the blastocyst stage, the program of the embryo has been altered by CSF2 differently in female embryos than in male embryos. Differential programming is apparent when examining development of embryos after transfer to recipients. There is no size difference between female and male embryos at Day 14 of development but, by Day 15, control female embryos are larger and secrete more IFNT than male embryos. This phenomenon probably represents earlier elongation for females. Moreover, CSF2 alters the differences between sexes by decreasing size and IFNT secretion for females but increasing these characteristics for males. In addition, CSF2 programs the transcriptome and methylome at day 15 in a sex-specific manner.

147
REFERENCES
1. Betteridge KJ, Fléchon JE. The anatomy and physiology of the pre-attachment
bovine embryos. Theriogenology 1988; 29:155-187.
2. Maddox-Hyttel P, Alexopoulos NI, Vajta G, Lewis I, Rogers P, Cann L, Callesen H, Tveden-Nyborg P, Trounson A. Immunohistochemical and ultrastructural characterization of the initial post-hatching development of bovine embryos. Reproduction 2003; 125:607-623.
3. Corcoran D, Fair T, Park S, Rizos D, Patel OV, Smith GW, Coussens PM, Ireland JJ, Boland MP, Evans AC, Lonergan P. Suppressed expression of genes involved in transcription and translation in in vitro compared with in vivo cultured bovine embryos. Reproduction 2006; 131:651-660.
4. Crosier AE, Farin PW, Dykstra MJ, Alexander JE, Farin CE. Ultrastructural morphometry of bovine compact morulae produced in vivo or in vitro. Biol Reprod 2000; 62:1459-1465.
5. Seidel GE. Modifying oocytes and embryos to improve their cryopreservation. Theriogenology 2006; 65:228-235.
6. Enright BP, Lonergan P, Dinnyes A, Fair T, Ward FA, Yang X, Boland MP. Culture of in vitro produced bovine zygotes in vitro vs in vivo: implications for early embryo development and quality. Theriogenology 2000; 54:659-673.
7. Rizos D, Fair T, Papadopoulos S, Boland MP, Lonergan P. Developmental, qualitative, and ultrastructural differences between ovine and bovine embryos produced in vivo or in vitro. Mol Reprod Dev 2002; 62:320-327.
8. Niemann H, Carnwath JW, Herrmann D, Wieczorek G, Lemme E, Lucas-Hahn A, Olek S. DNA methylation patterns reflect epigenetic reprogramming in bovine embryos. Cell Reprogram 2010; 12:33-42.
9. Maillo V, Rizos D, Besenfelder U, Havlicek V, Kelly AK, Garrett M, Lonergan P. Influence of lactation on metabolic characteristics and embryo development in postpartum Holstein dairy cows. J Dairy Sci 2012; 95:3865-3876.

148
10. Ponsuksili S, Murani E, Schwerin M, Schellander K, Tesfaye D, Wimmers K. Gene expression and DNA-methylation of bovine pretransfer endometrium depending on its receptivity after in vitro-produced embryo transfer. PLoS One 2012; 7:e42402.
11. Lonergan P, Woods A, Fair T, Carter F, Rizos D, Ward F, Quinn K, Evans A. Effect of embryo source and recipient progesterone environment on embryo development in cattle. Reprod Fertil Dev 2007; 19:861-868.
12. Thatcher WW, Moreira F, Santos JE, Mattos RC, Lopes FL, Pancarci SM, Risco CA. Effects of hormonal treatments on reproductive performance and embryo production. Theriogenology 2001; 55:75-89.
13. Watkins AJ, Ursell E, Panton R, Papenbrock T, Hollis L, Cunningham C, Wilkins A, Perry VH, Sheth B, Kwong WY, Eckert JJ, Wild AE, et al. Adaptive responses by mouse early embryos to maternal diet protect fetal growth but predispose to adult onset disease. Biol Reprod 2008; 78:299-306.
14. Watkins AJ, Lucas ES, Wilkins A, Cagampang FR, Fleming TP. Maternal periconceptional and gestational low protein diet affects mouse offspring growth, cardiovascular and adipose phenotype at 1 year of age. PLoS One 2011; 6:e28745.
15. Kwong WY, Wild AE, Roberts P, Willis AC, Fleming TP. Maternal undernutrition during the preimplantation period of rat development causes blastocyst abnormalities and programming of postnatal hypertension. Development 2000; 127:4195-4202.
16. Sinclair KD, Allegrucci C, Singh R, Gardner DS, Sebastian S, Bispham J, Thurston A, Huntley JF, Rees WD, Maloney CA, Lea RG, Craigon J, et al. DNA methylation, insulin resistance, and blood pressure in offspring determined by maternal periconceptional B vitamin and methionine status. Proc Natl Acad Sci U S A 2007; 104:19351-19356.
17. Watkins AJ, Lucas ES, Torrens C, Cleal JK, Green L, Osmond C, Eckert JJ, Gray WP, Hanson MA, Fleming TP. Maternal low-protein diet during mouse pre-implantation development induces vascular dysfunction and altered renin-angiotensin-system homeostasis in the offspring. Br J Nutr 2010; 103:1762-1770.

149
18. Loureiro B, Bonilla L, Block J, Fear JM, Bonilla AQ, Hansen PJ. Colony-stimulating factor 2 (CSF-2) improves development and posttransfer survival of bovine embryos produced in vitro. Endocrinology 2009; 150:5046-5054.
19. Bonilla AQ, Oliveira LJ, Ozawa M, Newsom EM, Lucy MC, Hansen PJ. Developmental changes in thermoprotective actions of insulin-like growth factor-1 on the preimplantation bovine embryo. Mol Cell Endocrinol 2011; 332:170-179.
20. Neira JA, Tainturier D, Peña MA, Martal J. Effect of the association of IGF-I, IGF-II, bFGF, TGF-β1, GM-CSF, and LIF on the development of bovine embryos produced in vitro. Theriogenology 2010; 73:595-604.
21. Block J, Bonilla L, Hansen PJ. Effect of addition of hyaluronan to embryo culture medium on survival of bovine embryos in vitro following vitrification and establishment of pregnancy after transfer to recipients. Theriogenology 2009; 71:1063-1071.
22. Fields SD, Hansen PJ, Ealy AD. Fibroblast growth factor requirements for in vitro development of bovine embryos. Theriogenology 2011; 75:1466-1475.
23. Paula-Lopes FF, de Moraes AA, Edwards JL, Justice JE, Hansen PJ. Regulation of preimplantation development of bovine embryos by interleukin-1β. Biol Reprod 1998; 59:1406-1412.
24. de Moraes AA, Hansen PJ. Granulocyte-macrophage colony-stimulating factor promotes development of in vitro produced bovine embryos. Biol Reprod 1997; 57:1060-1065.
25. Ziebe S, Loft A, Povlsen BB, Erb K, Agerholm I, Aasted M, Gabrielsen A, Hnida C, Zobel DP, Munding B, Bendz SH, Robertson SA. A randomized clinical trial to evaluate the effect of granulocyte-macrophage colony-stimulating factor (GM-CSF) in embryo culture medium for in vitro fertilization. Fertil Steril 2013; 99:1600-1609.
26. Sjöblom C, Roberts CT, Wikland M, Robertson SA. Granulocyte-macrophage colony-stimulating factor alleviates adverse consequences of embryo culture on fetal growth trajectory and placental morphogenesis. Endocrinology 2005; 146:2142-2153.

150
27. de Moraes AA, Paula-Lopes FF, Chegini N, Hansen PJ. Localization of granulocyte-macrophage colony-stimulating factor in the bovine reproductive tract. J Reprod Immunol 1999; 42:135-145.
28. Robertson SA, Mayrhofer G, Seamark RF. Uterine epithelial cells synthesize granulocyte-macrophage colony-stimulating factor and interleukin-6 in pregnant and nonpregnant mice. Biol Reprod 1992; 46:1069-1079.
29. Giacomini G, Tabibzadeh SS, Satyaswaroop PG, Bonsi L, Vitale L, Bagnara GP, Strippoli P, Jasonni VM. Epithelial cells are the major source of biologically active granulocyte macrophage colony-stimulating factor in human endometrium. Hum Reprod 1995; 10:3259-3263.
30. Loureiro B, Block J, Favoreto MG, Carambula S, Pennington KA, Ealy AD, Hansen PJ. Consequences of conceptus exposure to colony-stimulating factor 2 on survival, elongation, interferon-τ secretion, and gene expression. Reproduction 2011; 141:617-624.
31. Loureiro B, Oliveira LJ, Favoreto MG, Hansen PJ. Colony-stimulating factor 2 inhibits induction of apoptosis in the bovine preimplantation embryo. Am J Reprod Immunol 2011; 65:578-588.
32. Bromfield JJ, Schjenken JE, Chin PY, Care AS, Jasper MJ, Robertson SA. Maternal tract factors contribute to paternal seminal fluid impact on metabolic phenotype in offspring. Proc Natl Acad Sci U S A 2014.
33. Tremellen KP, Seamark RF, Robertson SA. Seminal transforming growth factor beta1 stimulates granulocyte-macrophage colony-stimulating factor production and inflammatory cell recruitment in the murine uterus. Biol Reprod 1998; 58:1217-1225.
34. Nahar A, Maki S, Kadokawa H. Suppressed expression of granulocyte macrophage colony-stimulating factor in oviduct ampullae of obese cows. Anim Reprod Sci 2013; 139:1-8.
35. Berg DK, van Leeuwen J, Beaumont S, Berg M, Pfeffer PL. Embryo loss in cattle between Days 7 and 16 of pregnancy. Theriogenology 2010; 73:250-260.

151
36. King GJ, Atkinson BA, Robertson HA. Development of the intercaruncular areas during early gestation and establishment of the bovine placenta. J Reprod Fertil 1981; 61:469-474.
37. HAMILTON WJ, LAING JA. Development of the egg of the cow up to the stage of blastocyst formation. J Anat 1946; 80:194-204.
38. Hawk HW. Transport and fate of spermatozoa after insemination of cattle. J Dairy Sci 1987; 70:1487-1503.
39. Hyttel P, Xu KP, Greve T. Scanning electron microscopy of in vitro fertilization in cattle. Anat Embryol (Berl) 1988; 178:41-46.
40. Hyttel P, Greve T, Callesen H. Ultrastructure of in-vivo fertilization in superovulated cattle. J Reprod Fertil 1988; 82:1-13.
41. Holm P, Shukri NN, Vajta G, Booth P, Bendixen C, Callesen H. Developmental kinetics of the first cell cycles of bovine in vitro produced embryos in relation to their in vitro viability and sex. Theriogenology 1998; 50:1285-1299.
42. Eckert JJ, Fleming TP. Tight junction biogenesis during early development. Biochim Biophys Acta 2008; 1778:717-728.
43. Tarkowski AK, Wróblewska J. Development of blastomeres of mouse eggs isolated at the 4- and 8-cell stage. J Embryol Exp Morphol 1967; 18:155-180.
44. Watson AJ, Kidder GM. Immunofluorescence assessment of the timing of appearance and cellular distribution of Na/K-ATPase during mouse embryogenesis. Dev Biol 1988; 126:80-90.
45. Barcroft LC, Offenberg H, Thomsen P, Watson AJ. Aquaporin proteins in murine trophectoderm mediate transepithelial water movements during cavitation. Dev Biol 2003; 256:342-354.
46. Kuijk EW, Du Puy L, Van Tol HT, Oei CH, Haagsman HP, Colenbrander B, Roelen BA. Differences in early lineage segregation between mammals. Dev Dyn 2008; 237:918-927.

152
47. Kuijk EW, van Tol LT, Van de Velde H, Wubbolts R, Welling M, Geijsen N, Roelen BA. The roles of FGF and MAP kinase signaling in the segregation of the epiblast and hypoblast cell lineages in bovine and human embryos. Development 2012; 139:871-882.
48. van Soom A, Ysebaert MT, de Kruif A. Relationship between timing of development, morula morphology, and cell allocation to inner cell mass and trophectoderm in in vitro-produced bovine embryos. Mol Reprod Dev 1997; 47:47-56.
49. Orozco-Lucero E, Dufort I, Robert C, Sirard MA. Rapidly cleaving bovine two-cell embryos have better developmental potential and a distinctive mRNA pattern. Mol Reprod Dev 2014; 81:31-41.
50. Lonergan P, Khatir H, Piumi F, Rieger D, Humblot P, Boland MP. Effect of time interval from insemination to first cleavage on the developmental characteristics, sex ratio and pregnancy rate after transfer of bovine embryos. J Reprod Fertil 1999; 117:159-167.
51. Larson MA, Kimura K, Kubisch HM, Roberts RM. Sexual dimorphism among bovine embryos in their ability to make the transition to expanded blastocyst and
in the expression of the signaling molecule IFN-. Proc Natl Acad Sci U S A 2001; 98:9677-9682.
52. Vejlsted M, Du Y, Vajta G, Maddox-Hyttel P. Post-hatching development of the porcine and bovine embryo-defining criteria for expected development in vivo and in vitro. Theriogenology 2006; 65:153-165.
53. Brandão DO, Maddox-Hyttel P, Løvendahl P, Rumpf R, Stringfellow D, Callesen H. Post hatching development: a novel system for extended in vitro culture of bovine embryos. Biol Reprod 2004; 71:2048-2055.
54. Blomberg L, Hashizume K, Viebahn C. Blastocyst elongation, trophoblastic differentiation, and embryonic pattern formation. Reproduction 2008; 135:181-195.
55. Robinson RS, Hammond AJ, Wathes DC, Hunter MG, Mann GE. Corpus luteum-endometrium-embryo interactions in the dairy cow: underlying mechanisms and clinical relevance. Reprod Domest Anim 2008; 43 Suppl 2:104-112.

153
56. Nephew KP, Cárdenas H, McClure KE, Ott TL, Bazer FW, Pope WF. Effects of administration of human chorionic gonadotropin or progesterone before maternal recognition of pregnancy on blastocyst development and pregnancy in sheep. J Anim Sci 1994; 72:453-458.
57. Robinson RS, Fray MD, Wathes DC, Lamming GE, Mann GE. In vivo expression of interferon tau mRNA by the embryonic trophoblast and uterine concentrations of interferon tau protein during early pregnancy in the cow. Mol Reprod Dev 2006; 73:470-474.
58. Wooding FB, Wathes DC. Binucleate cell migration in the bovine placentome. J Reprod Fertil 1980; 59:425-430.
59. Bazer FW, Spencer TE, Johnson GA, Burghardt RC, Wu G. Comparative aspects of implantation. Reproduction 2009; 138:195-209.
60. Hyttel P, Fair T, Callesen H, Greve T. Oocyte growth, capacitation and final maturation in cattle. Theriogenology 1997; 47:23-32.
61. Memili E, First NL. Zygotic and embryonic gene expression in cow: a review of timing and mechanisms of early gene expression as compared with other species. Zygote 2000; 8:87-96.
62. Weill L, Belloc E, Bava FA, Méndez R. Translational control by changes in poly(A) tail length: recycling mRNAs. Nat Struct Mol Biol 2012; 19:577-585.
63. Kues WA, Sudheer S, Herrmann D, Carnwath JW, Havlicek V, Besenfelder U, Lehrach H, Adjaye J, Niemann H. Genome-wide expression profiling reveals distinct clusters of transcriptional regulation during bovine preimplantation development in vivo. Proc Natl Acad Sci U S A 2008; 105:19768-19773.
64. Memili E, First NL. Developmental changes in RNA polymerase II in bovine oocytes, early embryos, and effect of alpha-amanitin on embryo development. Mol Reprod Dev 1998; 51:381-389.
65. Barnes FL, Eyestone WH. Early cleavage and the maternal zygotic transition in bovine embryos. Theriogenology 1990; 33.

154
66. Kopecný V, Fléchon JE, Camous S, Fulka J. Nucleologenesis and the onset of transcription in the eight-cell bovine embryo: fine-structural autoradiographic study. Mol Reprod Dev 1989; 1:79-90.
67. Tadros W, Lipshitz HD. The maternal-to-zygotic transition: a play in two acts. Development 2009; 136:3033-3042.
68. Bird AP. DNA methylation and the frequency of CpG in animal DNA. Nucleic Acids Res 1980; 8:1499-1504.
69. Boyes J, Bird A. DNA methylation inhibits transcription indirectly via a methyl-CpG binding protein. Cell 1991; 64:1123-1134.
70. Bird A. DNA methylation patterns and epigenetic memory. Genes Dev 2002; 16:6-21.
71. Miranda TB, Jones PA. DNA methylation: the nuts and bolts of repression. J Cell Physiol 2007; 213:384-390.
72. Bartolomei MS, Ferguson-Smith AC. Mammalian genomic imprinting. Cold Spring Harb Perspect Biol 2011; 3.
73. Oswald J, Engemann S, Lane N, Mayer W, Olek A, Fundele R, Dean W, Reik W, Walter J. Active demethylation of the paternal genome in the mouse zygote. Curr Biol 2000; 10:475-478.
74. Mayer W, Niveleau A, Walter J, Fundele R, Haaf T. Demethylation of the zygotic paternal genome. Nature 2000; 403:501-502.
75. Santos F, Dean W. Epigenetic reprogramming during early development in mammals. Reproduction 2004; 127:643-651.
76. Santos F, Hendrich B, Reik W, Dean W. Dynamic reprogramming of DNA methylation in the early mouse embryo. Dev Biol 2002; 241:172-182.
77. Jost JP, Siegmann M, Sun L, Leung R. Mechanisms of DNA demethylation in chicken embryos. Purification and properties of a 5-methylcytosine-DNA glycosylase. J Biol Chem 1995; 270:9734-9739.

155
78. Weiss A, Keshet I, Razin A, Cedar H. DNA demethylation in vitro: involvement of RNA. Cell 1996; 86:709-718.
79. Morgan HD, Dean W, Coker HA, Reik W, Petersen-Mahrt SK. Activation-induced cytidine deaminase deaminates 5-methylcytosine in DNA and is expressed in pluripotent tissues: implications for epigenetic reprogramming. J Biol Chem 2004; 279:52353-52360.
80. Iqbal K, Jin SG, Pfeifer GP, Szabó PE. Reprogramming of the paternal genome upon fertilization involves genome-wide oxidation of 5-methylcytosine. Proc Natl Acad Sci U S A 2011; 108:3642-3647.
81. Seisenberger S, Andrews S, Krueger F, Arand J, Walter J, Santos F, Popp C, Thienpont B, Dean W, Reik W. The dynamics of genome-wide DNA methylation reprogramming in mouse primordial germ cells. Mol Cell 2012; 48:849-862.
82. Okano M, Bell DW, Haber DA, Li E. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell 1999; 99:247-257.
83. Howell CY, Bestor TH, Ding F, Latham KE, Mertineit C, Trasler JM, Chaillet JR. Genomic imprinting disrupted by a maternal effect mutation in the Dnmt1 gene. Cell 2001; 104:829-838.
84. Carlson LL, Page AW, Bestor TH. Properties and localization of DNA methyltransferase in preimplantation mouse embryos: implications for genomic imprinting. Genes Dev 1992; 6:2536-2541.
85. Beaujean N, Taylor J, Gardner J, Wilmut I, Meehan R, Young L. Effect of limited DNA methylation reprogramming in the normal sheep embryo on somatic cell nuclear transfer. Biol Reprod 2004; 71:185-193.
86. Fulka J, Fulka H, Slavik T, Okada K. DNA methylation pattern in pig in vivo produced embryos. Histochem Cell Biol 2006; 126:213-217.
87. Dean W, Santos F, Stojkovic M, Zakhartchenko V, Walter J, Wolf E, Reik W. Conservation of methylation reprogramming in mammalian development: aberrant reprogramming in cloned embryos. Proc Natl Acad Sci U S A 2001; 98:13734-13738.

156
88. Hou J, Liu L, Lei T, Cui X, An X, Chen Y. Genomic DNA methylation patterns in bovine preimplantation embryos derived from in vitro fertilization. Sci China C Life Sci 2007; 50:56-61.
89. Chambers I, Colby D, Robertson M, Nichols J, Lee S, Tweedie S, Smith A. Functional expression cloning of Nanog, a pluripotency sustaining factor in embryonic stem cells. Cell 2003; 113:643-655.
90. Nichols J, Zevnik B, Anastassiadis K, Niwa H, Klewe-Nebenius D, Chambers I, Schöler H, Smith A. Formation of pluripotent stem cells in the mammalian embryo depends on the POU transcription factor Oct4. Cell 1998; 95:379-391.
91. Avilion AA, Nicolis SK, Pevny LH, Perez L, Vivian N, Lovell-Badge R. Multipotent cell lineages in early mouse development depend on SOX2 function. Genes Dev 2003; 17:126-140.
92. Niwa H, Toyooka Y, Shimosato D, Strumpf D, Takahashi K, Yagi R, Rossant J. Interaction between Oct3/4 and Cdx2 determines trophectoderm differentiation. Cell 2005; 123:917-929.
93. Strumpf D, Mao CA, Yamanaka Y, Ralston A, Chawengsaksophak K, Beck F, Rossant J. Cdx2 is required for correct cell fate specification and differentiation of trophectoderm in the mouse blastocyst. Development 2005; 132:2093-2102.
94. Berg DK, Smith CS, Pearton DJ, Wells DN, Broadhurst R, Donnison M, Pfeffer PL. Trophectoderm lineage determination in cattle. Dev Cell 2011; 20:244-255.
95. Morris SA, Graham SJ, Jedrusik A, Zernicka-Goetz M. The differential response to Fgf signalling in cells internalized at different times influences lineage segregation in preimplantation mouse embryos. Open Biol 2013; 3:130104.
96. Krupa M, Mazur E, Szczepańska K, Filimonow K, Maleszewski M, Suwińska A. Allocation of inner cells to epiblast vs primitive endoderm in the mouse embryo is biased but not determined by the round of asymmetric divisions (8→16- and 16→32-cells). Dev Biol 2014; 385:136-148.
97. Yamanaka Y, Lanner F, Rossant J. FGF signal-dependent segregation of primitive endoderm and epiblast in the mouse blastocyst. Development 2010; 137:715-724.

157
98. Harris D, Huang B, Oback B. Inhibition of MAP2K and GSK3 signaling promotes bovine blastocyst development and epiblast-associated expression of pluripotency factors. Biol Reprod 2013; 88:74.
99. Farin PW, Farin CE. Transfer of bovine embryos produced in vivo or in vitro: survival and fetal development. Biol Reprod 1995; 52:676-682.
100. Hansen PJ, Block J. Towards an embryocentric world: the current and potential uses of embryo technologies in dairy production. Reprod Fertil Dev 2004; 16:1-14.
101. Driver AM, Peñagaricano F, Huang W, Ahmad KR, Hackbart KS, Wiltbank MC, Khatib H. RNA-Seq analysis uncovers transcriptomic variations between morphologically similar in vivo- and in vitro-derived bovine blastocysts. BMC Genomics 2012; 13:118.
102. Gad A, Hoelker M, Besenfelder U, Havlicek V, Cinar U, Rings F, Held E, Dufort I, Sirard MA, Schellander K, Tesfaye D. Molecular mechanisms and pathways involved in bovine embryonic genome activation and their regulation by alternative in vivo and in vitro culture conditions. Biol Reprod 2012; 87:100.
103. Lonergan P, Rizos D, Gutiérrez-Adán A, Fair T, Boland MP. Effect of culture environment on embryo quality and gene expression - experience from animal studies. Reprod Biomed Online 2003; 7:657-663.
104. Smith SL, Everts RE, Sung LY, Du F, Page RL, Henderson B, Rodriguez-Zas SL, Nedambale TL, Renard JP, Lewin HA, Yang X, Tian XC. Gene expression profiling of single bovine embryos uncovers significant effects of in vitro maturation, fertilization and culture. Mol Reprod Dev 2009; 76:38-47.
105. Rizos D, Lonergan P, Boland MP, Arroyo-García R, Pintado B, de la Fuente J, Gutiérrez-Adán A. Analysis of differential messenger RNA expression between bovine blastocysts produced in different culture systems: implications for blastocyst quality. Biol Reprod 2002; 66:589-595.
106. Chen Z, Robbins KM, Wells KD, Rivera RM. Large offspring syndrome: a bovine model for the human loss-of-imprinting overgrowth syndrome Beckwith-Wiedemann. Epigenetics 2013; 8:591-601.

158
107. Sudano MJ, Santos VG, Tata A, Ferreira CR, Paschoal DM, Machado R, Buratini J, Eberlin MN, Landim-Alvarenga FD. Phosphatidylcholine and sphingomyelin profiles vary in Bos taurus indicus and Bos taurus taurus in vitro- and in vivo-produced blastocysts. Biol Reprod 2012; 87:130.
108. Gutiérrez-Adán A, Lonergan P, Rizos D, Ward FA, Boland MP, Pintado B, de la Fuente J. Effect of the in vitro culture system on the kinetics of blastocyst development and sex ratio of bovine embryos. Theriogenology 2001; 55:1117-1126.
109. Hyttel P, Xu KP, Smith S, Greve T. Ultrastructure of in-vitro oocyte maturation in cattle. J Reprod Fertil 1986; 78:615-625.
110. Tesfaye D, Ghanem N, Carter F, Fair T, Sirard MA, Hoelker M, Schellander K, Lonergan P. Gene expression profile of cumulus cells derived from cumulus-oocyte complexes matured either in vivo or in vitro. Reprod Fertil Dev 2009; 21:451-461.
111. Knijn HM, Wrenzycki C, Hendriksen PJ, Vos PL, Herrmann D, van der Weijden GC, Niemann H, Dieleman SJ. Effects of oocyte maturation regimen on the relative abundance of gene transcripts in bovine blastocysts derived in vitro or in vivo. Reproduction 2002; 124:365-375.
112. Rizos D, Ward F, Duffy P, Boland MP, Lonergan P. Consequences of bovine oocyte maturation, fertilization or early embryo development in vitro versus in vivo: implications for blastocyst yield and blastocyst quality. Mol Reprod Dev 2002; 61:234-248.
113. Kind KL, Banwell KM, Gebhardt KM, Macpherson A, Gauld A, Russell DL, Thompson JG. Microarray analysis of mRNA from cumulus cells following in vivo or in vitro maturation of mouse cumulus-oocyte complexes. Reprod Fertil Dev 2013; 25:426-438.
114. Motlik J, Fulka J, Fléchon JE. Changes in intercellular coupling between pig oocytes and cumulus cells during maturation in vivo and in vitro. J Reprod Fertil 1986; 76:31-37.
115. Eppig JJ, Schroeder AC. Capacity of mouse oocytes from preantral follicles to undergo embryogenesis and development to live young after growth, maturation, and fertilization in vitro. Biol Reprod 1989; 41:268-276.

159
116. Rizos D, Gutiérrez-Adán A, Pérez-Garnelo S, De La Fuente J, Boland MP, Lonergan P. Bovine embryo culture in the presence or absence of serum: implications for blastocyst development, cryotolerance, and messenger RNA expression. Biol Reprod 2003; 68:236-243.
117. Guyader-Joly C, Ponchon S, Durand M, Heyman Y, Renard JP, Ménézo Y. Effect of lecithin on in vitro and in vivo survival of in vitro produced bovine blastocysts after cryopreservation. Theriogenology 1999; 52:1193-1202.
118. Hasler JF, Henderson WB, Hurtgen PJ, Jin ZQ, McCauley AD, Mower SA, Nelly B, Shuey LS, Stokes JE, Trimmer SA. Production, freezing and transfer if bovine IVF embryos and subsequent calving results. Theriogeneology 1995; 43:141-152.
119. Seidel GE. Modifying oocytes and embryos to improve their cryopreservation. Theriogenology 2006; 65:228-235.
120. Bonilla L, Block J, Denicol AC, Hansen PJ. Consequences of transfer of an in vitro-produced embryo for the dam and resultant calf. J Dairy Sci 2014; 97:229-239.
121. Farin PW, Slenning BD, Britt JH. Estimates of pregnancy outcomes based on selection of bovine embryos produced in vivo or in vitro. Theriogenology 1999; 52:659-670.
122. Siqueira LG, Torres CA, Souza ED, Monteiro PL, Arashiro EK, Camargo LS, Fernandes CA, Viana JH. Pregnancy rates and corpus luteum-related factors affecting pregnancy establishment in bovine recipients synchronized for fixed-time embryo transfer. Theriogenology 2009; 72:949-958.
123. Pontes JH, Nonato-Junior I, Sanches BV, Ereno-Junior JC, Uvo S, Barreiros TR, Oliveira JA, Hasler JF, Seneda MM. Comparison of embryo yield and pregnancy rate between in vivo and in vitro methods in the same Nelore (Bos indicus) donor cows. Theriogenology 2009; 71:690-697.
124. van Wagtendonk-de Leeuw AM, Aerts BJ, den Daas JH. Abnormal offspring following in vitro production of bovine preimplantation embryos: a field study. Theriogenology 1998; 49:883-894.

160
125. Miles JR, Farin CE, Rodriguez KF, Alexander JE, Farin PW. Angiogenesis and morphometry of bovine placentas in late gestation from embryos produced in vivo or in vitro. Biol Reprod 2004; 71:1919-1926.
126. Miles JR, Farin CE, Rodriguez KF, Alexander JE, Farin PW. Effects of embryo culture on angiogenesis and morphometry of bovine placentas during early gestation. Biol Reprod 2005; 73:663-671.
127. Behboodi E, Anderson GB, BonDurant RH, Cargill SL, Kreuscher BR, Medrano JF, Murray JD. Birth of large calves that developed from in vitro-derived bovine embryos. Theriogenology 1995; 44:227-232.
128. Numabe T, Oikawa T, Kikuchi T, Horiuchi T. Birth weight and birth rate of heavy calves conceived by transfer of in vitro or in vivo produced bovine embryos. Anim Reprod Sci 2000; 64:13-20.
129. Block J, Drost M, Monson RL, Rutledge JJ, Rivera RM, Paula-Lopes FF, Ocon OM, Krininger CE, Liu J, Hansen PJ. Use of insulin-like growth factor-I during embryo culture and treatment of recipients with gonadotropin-releasing hormone to increase pregnancy rates following the transfer of in vitro-produced embryos to heat-stressed, lactating cows. J Anim Sci 2003; 81:1590-1602.
130. Ziebe S, Lundin K, Loft A, Bergh C, Nyboe Andersen A, Selleskog U, Nielsen D, Grøndahl C, Kim H, Arce JC, Group CIaS. FISH analysis for chromosomes 13, 16, 18, 21, 22, X and Y in all blastomeres of IVF pre-embryos from 144 randomly selected donated human oocytes and impact on pre-embryo morphology. Hum Reprod 2003; 18:2575-2581.
131. Angell RR, Aitken RJ, van Look PF, Lumsden MA, Templeton AA. Chromosome abnormalities in human embryos after in vitro fertilization. Nature 1983; 303:336-338.
132. Hardy K, Handyside AH, Winston RM. The human blastocyst: cell number, death and allocation during late preimplantation development in vitro. Development 1989; 107:597-604.
133. Bauer BK, Isom SC, Spate LD, Whitworth KM, Spollen WG, Blake SM, Springer GK, Murphy CN, Prather RS. Transcriptional profiling by deep sequencing identifies differences in mRNA transcript abundance in in vivo-derived versus in vitro-cultured porcine blastocyst stage embryos. Biol Reprod 2010; 83:791-798.

161
134. Hyttel P, Niemann H. Ultrastructure of porcine embryos following development in vitro versus in vivo. Mol Reprod Dev 1990; 27:136-144.
135. Giritharan G, Talbi S, Donjacour A, Di Sebastiano F, Dobson AT, Rinaudo PF. Effect of in vitro fertilization on gene expression and development of mouse preimplantation embryos. Reproduction 2007; 134:63-72.
136. Doherty AS, Mann MR, Tremblay KD, Bartolomei MS, Schultz RM. Differential effects of culture on imprinted H19 expression in the preimplantation mouse embryo. Biol Reprod 2000; 62:1526-1535.
137. Ceelen M, van Weissenbruch MM, Vermeiden JP, van Leeuwen FE, Delemarre-van de Waal HA. Cardiometabolic differences in children born after in vitro fertilization: follow-up study. J Clin Endocrinol Metab 2008; 93:1682-1688.
138. Sinclair KD, McEvoy TG, Maxfield EK, Maltin CA, Young LE, Wilmut I, Broadbent PJ, Robinson JJ. Aberrant fetal growth and development after in vitro culture of sheep zygotes. J Reprod Fertil 1999; 116:177-186.
139. Garrett JE, Geisert RD, Zavy MT, Morgan GL. Evidence for maternal regulation of early conceptus growth and development in beef cattle. J Reprod Fertil 1988; 84:437-446.
140. Carter F, Forde N, Duffy P, Wade M, Fair T, Crowe MA, Evans AC, Kenny DA, Roche JF, Lonergan P. Effect of increasing progesterone concentration from Day 3 of pregnancy on subsequent embryo survival and development in beef heifers. Reprod Fertil Dev 2008; 20:368-375.
141. Forde N, Carter F, Fair T, Crowe MA, Evans AC, Spencer TE, Bazer FW, McBride R, Boland MP, O'Gaora P, Lonergan P, Roche JF. Progesterone-regulated changes in endometrial gene expression contribute to advanced conceptus development in cattle. Biol Reprod 2009; 81:784-794.
142. Bauman DE. Bovine somatotropin and lactation: from basic science to commercial application. Domest Anim Endocrinol 1999; 17:101-116.
143. Moreira F, Badinga L, Burnley C, Thatcher WW. Bovine somatotropin increases embryonic development in superovulated cows and improves post-transfer pregnancy rates when given to lactating recipient cows. Theriogenology 2002; 57:1371-1387.

162
144. Ribeiro ES, Bruno RG, Farias AM, Hernández-Rivera JA, Gomes GC, Surjus R, Becker LF, Birt A, Ott TL, Branen JR, Sasser RG, Keisler DH, et al. Low Doses of Bovine Somatotropin Enhance Conceptus Development and Fertility in Lactating Dairy Cows. Biol Reprod 2013.
145. Gifford CA, Racicot K, Clark DS, Austin KJ, Hansen TR, Lucy MC, Davies CJ, Ott TL. Regulation of interferon-stimulated genes in peripheral blood leukocytes in pregnant and bred, nonpregnant dairy cows. J Dairy Sci 2007; 90:274-280.
146. Gifford CA, Assiri AM, Satterfield MC, Spencer TE, Ott TL. Receptor transporter protein 4 (RTP4) in endometrium, ovary, and peripheral blood leukocytes of pregnant and cyclic ewes. Biol Reprod 2008; 79:518-524.
147. Lopes da Costa L, Chagas e Silva J, Robalo Silva J. Superovulatory response, embryo quality and fertility after treatment with different gonadotrophins in native cattle. Theriogenology 2001; 56:65-77.
148. Cerri RL, Thompson IM, Kim IH, Ealy AD, Hansen PJ, Staples CR, Li JL, Santos JE, Thatcher WW. Effects of lactation and pregnancy on gene expression of endometrium of Holstein cows at day 17 of the estrous cycle or pregnancy. J Dairy Sci 2012; 95:5657-5675.
149. Hasler J. Factors affecting frozen and fresh embryo transfer pregnancy rates in cattle. Theriogenology 2001; 56:1401-1415.
150. Chagas e Silva J, Lopes da Costa L, Robalo Silva J. Plasma progesterone profiles and factors affecting embryo-fetal mortality following embryo transfer in dairy cattle. Theriogenology 2002; 58:51-59.
151. Wilson RD, Fricke PM, Leibfried-Rutledge ML, Rutledge JJ, Penfield CM, Weigel KA. In vitro production of bovine embryos using sex-sorted sperm. Theriogenology 2006; 65:1007-1015.
152. Roseboom TJ, van der Meulen JH, Ravelli AC, Osmond C, Barker DJ, Bleker OP. Effects of prenatal exposure to the Dutch famine on adult disease in later life: an overview. Mol Cell Endocrinol 2001; 185:93-98.
153. Barker DJP. Mothers, Babies and Disease in Later Life. Edinburgh: Churchill Livingstone; 1998.

163
154. Ravelli AC, van der Meulen JH, Michels RP, Osmond C, Barker DJ, Hales CN, Bleker OP. Glucose tolerance in adults after prenatal exposure to famine. Lancet 1998; 351:173-177.
155. Ravelli AC, van Der Meulen JH, Osmond C, Barker DJ, Bleker OP. Obesity at the age of 50 y in men and women exposed to famine prenatally. Am J Clin Nutr 1999; 70:811-816.
156. Roseboom TJ, van der Meulen JH, Ravelli AC, Osmond C, Barker DJ, Bleker OP. Plasma fibrinogen and factor VII concentrations in adults after prenatal exposure to famine. Br J Haematol 2000; 111:112-117.
157. Roseboom TJ, van der Meulen JH, van Montfrans GA, Ravelli AC, Osmond C, Barker DJ, Bleker OP. Maternal nutrition during gestation and blood pressure in later life. J Hypertens 2001; 19:29-34.
158. Shaw GM, Schaffer D, Velie EM, Morland K, Harris JA. Periconceptional vitamin use, dietary folate, and the occurrence of neural tube defects. Epidemiology 1995; 6:219-226.
159. Langley-Evans SC, Phillips GJ, Jackson AA. In utero exposure to maternal low protein diets induces hypertension in weanling rats, independently of maternal blood pressure changes. Clin Nutr 1994; 13:319-324.
160. Garofano A, Czernichow P, Bréant B. Effect of ageing on beta-cell mass and function in rats malnourished during the perinatal period. Diabetologia 1999; 42:711-718.
161. Vickers MH, Breier BH, Cutfield WS, Hofman PL, Gluckman PD. Fetal origins of hyperphagia, obesity, and hypertension and postnatal amplification by hypercaloric nutrition. Am J Physiol Endocrinol Metab 2000; 279:E83-87.
162. George LA, Zhang L, Tuersunjiang N, Ma Y, Long NM, Uthlaut AB, Smith DT, Nathanielsz PW, Ford SP. Early maternal undernutrition programs increased feed intake, altered glucose metabolism and insulin secretion, and liver function in aged female offspring. Am J Physiol Regul Integr Comp Physiol 2012; 302:R795-804.

164
163. Ross MG, Desai M, Guerra C, Wang S. Prenatal programming of hypernatremia and hypertension in neonatal lambs. Am J Physiol Regul Integr Comp Physiol 2005; 288:R97-103.
164. Clarke AS, Wittwer DJ, Abbott DH, Schneider ML. Long-term effects of prenatal stress on HPA axis activity in juvenile rhesus monkeys. Dev Psychobiol 1994; 27:257-269.
165. Johnson J, Ross J, Selsby J, Boddicker R, Sanz-Fernandez V, Baumgard L. Effects of in-utero heat stress on porcine post-natal thermoregualtion. In. Iowa State University Animal Industry Report 2013: Iowa State University; 2013.
166. Zambrano E, Bautista CJ, Deás M, Martínez-Samayoa PM, González-Zamorano M, Ledesma H, Morales J, Larrea F, Nathanielsz PW. A low maternal protein diet during pregnancy and lactation has sex- and window of exposure-specific effects on offspring growth and food intake, glucose metabolism and serum leptin in the rat. J Physiol 2006; 571:221-230.
167. Gilbert J, Cox L, Babcock S, Shade R, Nathanielsz P, Nijland M. Gender-specific effects of moderate maternal nutrient restriction (NR) in the first half of pregnancy on fetal renal renin-angiotensin system (RAS). Journal of the Society for Gynecologic Investigation 2006; 13:207A.
168. Gilbert JS, Ford SP, Lang AL, Pahl LR, Drumhiller MC, Babcock SA, Nathanielsz PW, Nijland MJ. Nutrient restriction impairs nephrogenesis in a gender-specific manner in the ovine fetus. Pediatr Res 2007; 61:42-47.
169. Maloney CA, Hay SM, Young LE, Sinclair KD, Rees WD. A methyl-deficient diet fed to rat dams during the peri-conception period programs glucose homeostasis in adult male but not female offspring. J Nutr 2011; 141:95-100.
170. Bazer FW, Wu G, Johnson GA, Kim J, Song G. Uterine histotroph and conceptus development: select nutrients and secreted phosphoprotein 1 affect mechanistic target of rapamycin cell signaling in ewes. Biol Reprod 2011; 85:1094-1107.
171. Simmons RM, Satterfield MC, Welsh TH, Bazer FW, Spencer TE. HSD11B1, HSD11B2, PTGS2, and NR3C1 expression in the peri-implantation ovine uterus: effects of pregnancy, progesterone, and interferon tau. Biol Reprod 2010; 82:35-43.

165
172. Lee HY, Acosta TJ, Skarzynski DJ, Okuda K. Prostaglandin F2alpha stimulates 11Beta-hydroxysteroid dehydrogenase 1 enzyme bioactivity and protein expression in bovine endometrial stromal cells. Biol Reprod 2009; 80:657-664.
173. Thatcher WW, Guzeloglu A, Mattos R, Binelli M, Hansen TR, Pru JK. Uterine-conceptus interactions and reproductive failure in cattle. Theriogenology 2001; 56:1435-1450.
174. Forde N, Lonergan P. Transcriptomic analysis of the bovine endometrium: What is required to establish uterine receptivity to implantation in cattle? J Reprod Dev 2012; 58:189-195.
175. Gough NM, Gough J, Metcalf D, Kelso A, Grail D, Nicola NA, Burgess AW, Dunn AR. Molecular cloning of cDNA encoding a murine haematopoietic growth regulator, granulocyte-macrophage colony stimulating factor. Nature 1984; 309:763-767.
176. Ruef C, Coleman DL. Granulocyte-macrophage colony-stimulating factor: pleiotropic cytokine with potential clinical usefulness. Rev Infect Dis 1990; 12:41-62.
177. Trapnell BC, Whitsett JA. Gm-CSF regulates pulmonary surfactant homeostasis and alveolar macrophage-mediated innate host defense. Annu Rev Physiol 2002; 64:775-802.
178. Burgess AW, Metcalf D. The nature and action of granulocyte-macrophage colony stimulating factors. Blood 1980; 56:947-958.
179. Kanzaki H, Crainie M, Lin H, Yui J, Guilbert LJ, Mori T, Wegmann TG. The in situ expression of granulocyte-macrophage colony-stimulating factor (GM-CSF) mRNA at the maternal-fetal interface. Growth Factors 1991; 5:69-74.
180. Chegini N, Tang XM, Dou Q. The expression, activity and regulation of granulocyte macrophage-colony stimulating factor in human endometrial epithelial and stromal cells. Mol Hum Reprod 1999; 5:459-466.
181. Hayashida K, Kitamura T, Gorman DM, Arai K, Yokota T, Miyajima A. Molecular cloning of a second subunit of the receptor for human granulocyte-macrophage colony-stimulating factor (GM-CSF): reconstitution of a high-affinity GM-CSF receptor. Proc Natl Acad Sci U S A 1990; 87:9655-9659.

166
182. Gearing DP, King JA, Gough NM, Nicola NA. Expression cloning of a receptor for human granulocyte-macrophage colony-stimulating factor. EMBO J 1989; 8:3667-3676.
183. Dhar-Mascareno M, Chen J, Zhang RH, Cárcamo JM, Golde DW. Granulocyte-macrophage colony-stimulating factor signals for increased glucose transport via phosphatidylinositol 3-kinase- and hydrogen peroxide-dependent mechanisms. J Biol Chem 2003; 278:11107-11114.
184. Kitamura T, Hayashida K, Sakamaki K, Yokota T, Arai K, Miyajima A. Reconstitution of functional receptors for human granulocyte/macrophage colony-stimulating factor (GM-CSF): evidence that the protein encoded by the AIC2B cDNA is a subunit of the murine GM-CSF receptor. Proc Natl Acad Sci U S A 1991; 88:5082-5086.
185. Barreda DR, Hanington PC, Belosevic M. Regulation of myeloid development and function by colony stimulating factors. Dev Comp Immunol 2004; 28:509-554.
186. Robertson SA, Sjöblom C, Jasper MJ, Norman RJ, Seamark RF. Granulocyte-macrophage colony-stimulating factor promotes glucose transport and blastomere viability in murine preimplantation embryos. Biol Reprod 2001; 64:1206-1215.
187. Sjöblom C, Wikland M, Robertson SA. Granulocyte-macrophage colony-stimulating factor (GM-CSF) acts independently of the beta common subunit of the GM-CSF receptor to prevent inner cell mass apoptosis in human embryos. Biol Reprod 2002; 67:1817-1823.
188. Peralta OA, Bucher D, Fernandez A, Berland M, Strobel P, Ramirez A, Ratto MH, Concha I. Granulocyte-macrophage colony stimulating factor (GM-CSF) enhances cumulus cell expansion in bovine oocytes. Reprod Biol Endocrinol 2013; 11:55.
189. Lee K, Redel BK, Spate L, Teson J, Brown AN, Park KW, Walters E, Samuel M, Murphy CN, Prather RS. Piglets produced from cloned blastocysts cultured in vitro with GM-CSF. Mol Reprod Dev 2013; 80:145-154.

167
190. Quelle FW, Sato N, Witthuhn BA, Inhorn RC, Eder M, Miyajima A, Griffin JD, Ihle JN. JAK2 associates with the beta c chain of the receptor for granulocyte-macrophage colony-stimulating factor, and its activation requires the membrane-proximal region. Mol Cell Biol 1994; 14:4335-4341.
191. Martinez-Moczygemba M, Huston DP. Biology of common beta receptor-signaling cytokines: IL-3, IL-5, and GM-CSF. J Allergy Clin Immunol 2003; 112:653-665; quiz 666.
192. Sakamaki K, Miyajima I, Kitamura T, Miyajima A. Critical cytoplasmic domains of the common beta subunit of the human GM-CSF, IL-3 and IL-5 receptors for growth signal transduction and tyrosine phosphorylation. EMBO J 1992; 11:3541-3549.
193. O'Leary S, Jasper MJ, Warnes GM, Armstrong DT, Robertson SA. Seminal plasma regulates endometrial cytokine expression, leukocyte recruitment and embryo development in the pig. Reproduction 2004; 128:237-247.
194. Robertson SA, Mayrhofer G, Seamark RF. Ovarian steroid hormones regulate granulocyte-macrophage colony-stimulating factor synthesis by uterine epithelial cells in the mouse. Biol Reprod 1996; 54:183-196.
195. Zhao Y, Chegini N. The expression of granulocyte macrophage-colony stimulating factor (GM-CSF) and receptors in human endometrium. Am J Reprod Immunol 1999; 42:303-311.
196. Segerer S, Kammerer U, Kapp M, Dietl J, Rieger L. Upregulation of chemokine and cytokine production during pregnancy. Gynecol Obstet Invest 2009; 67:145-150.
197. Emond V, MacLaren LA, Kimmins S, Arosh JA, Fortier MA, Lambert RD. Expression of cyclooxygenase-2 and granulocyte-macrophage colony-stimulating factor in the endometrial epithelium of the cow is up-regulated during early pregnancy and in response to intrauterine infusions of interferon-tau. Biol Reprod 2004; 70:54-64.
198. Cui XS, Lee JY, Choi SH, Kwon MS, Kim T, Kim NH. Mouse granulocyte-macrophage colony-stimulating factor enhances viability of porcine embryos in defined culture conditions. Anim Reprod Sci 2004; 84:169-177.

168
199. Kwak SS, Cheong SA, Jeon Y, Hyun SH. Porcine granulocyte-macrophage colony-stimulating factor improves the in vitro development of cloned porcine embryos. J Vet Med Sci 2012; 74:1095-1102.
200. Sjöblom C, Wikland M, Robertson SA. Granulocyte-macrophage colony-stimulating factor promotes human blastocyst development in vitro. Hum Reprod 1999; 14:3069-3076.
201. Desai N, Kattal N, AbdelHafez FF, Szeptycki-Lawson J, Goldfarb J. Granulocyte-macrophage colony stimulating factor (GM-CSF) and co-culture can affect post-thaw development and apoptosis in cryopreserved embryos. J Assist Reprod Genet 2007; 24:215-222.
202. Sheikholslami B, Salehnia M, Valojerdi MR, Ramezanzadeh M. Developmental potential of isolated blastomeres from early mouse embryos in the presence and absence of LIF and GM-CSF. J Assist Reprod Genet 2008; 25:7-12.
203. Hickman CF, Ainslie A, Ealy AD, Ashworth CJ, Rooke JA. Effect of ovine granulocyte-macrophage colony-stimulating factor on bovine in vitro embryo development and blastocyst interferon-τ secretion. Reprod Domest Anim 2011; 46:608-615.
204. Elaimi A, Gardner K, Kistnareddy K, Harper J. The effect of GM-CSF on development and aneuploidy in murine blastocysts. Hum Reprod 2012; 27:1590-1595.
205. Dhar-Mascareno M, Pedraza A, Golde DW. PI3-kinase activation by GM-CSF in endothelium is upstream of Jak/Stat pathway: role of alphaGMR. Biochem Biophys Res Commun 2005; 337:551-556.
206. Jeong W, Kim J, Bazer FW, Song G. Proliferation-stimulating effect of colony stimulating factor 2 on porcine trophectoderm cells is mediated by activation of phosphatidylinositol 3-kinase and extracellular signal-regulated kinase 1/2 mitogen-activated protein kinase. PLoS One 2014; 9:e88731.
207. Lazzari G, Wrenzycki C, Herrmann D, Duchi R, Kruip T, Niemann H, Galli C. Cellular and molecular deviations in bovine in vitro-produced embryos are related to the large offspring syndrome. Biol Reprod 2002; 67:767-775.

169
208. Papadopoulos S, Rizos D, Duffy P, Wade M, Quinn K, Boland MP, Lonergan P. Embryo survival and recipient pregnancy rates after transfer of fresh or vitrified, in vivo or in vitro produced ovine blastocysts. Anim Reprod Sci 2002; 74:35-44.
209. Lonergan P, Carolan C, Van Langendonckt A, Donnay I, Khatir H, Mermillod P. Role of epidermal growth factor in bovine oocyte maturation and preimplantation embryo development in vitro. Biol Reprod 1996; 54:1420-1429.
210. Bonilla AQ, Ozawa M, Hansen PJ. Timing and dependence upon mitogen-activated protein kinase signaling for pro-developmental actions of insulin-like growth factor 1 on the preimplantation bovine embryo. Growth Horm IGF Res 2011; 21:107-111.
211. Trigal B, Gómez E, Díez C, Caamaño JN, Martín D, Carrocera S, Muñoz M. In vitro development of bovine embryos cultured with activin A. Theriogenology 2011; 75:584-588.
212. Luo S, Yin HN, Li SW. [Effect of TGF-beta1 on embryo implantation and development in mice in vitro]. Sichuan Da Xue Xue Bao Yi Xue Ban 2010; 41:265-268.
213. Cheung LP, Leung HY, Bongso A. Effect of supplementation of leukemia inhibitory factor and epidermal growth factor on murine embryonic development in vitro, implantation, and outcome of offspring. Fertil Steril 2003; 80 Suppl 2:727-735.
214. Lavranos TC, Rathjen PD, Seamark RF. Trophic effects of myeloid leukaemia inhibitory factor (LIF) on mouse embryos. J Reprod Fertil 1995; 105:331-338.
215. Block J, Hansen PJ. Interaction between season and culture with insulin-like growth factor-1 on survival of in vitro produced embryos following transfer to lactating dairy cows. Theriogenology 2007; 67:1518-1529.
216. Chin PY, Macpherson AM, Thompson JG, Lane M, Robertson SA. Stress response genes are suppressed in mouse preimplantation embryos by granulocyte-macrophage colony-stimulating factor (GM-CSF). Hum Reprod 2009; 24:2997-3009.

170
217. McClure BJ, Hercus TR, Cambareri BA, Woodcock JM, Bagley CJ, Howlett GJ, Lopez AF. Molecular assembly of the ternary granulocyte-macrophage colony-stimulating factor receptor complex. Blood 2003; 101:1308-1315.
218. Ozawa M, Sakatani M, Hankowski KE, Terada N, Dobbs KB, Hansen PJ. Importance of culture conditions during the morula-to-blastocyst period on capacity of inner cell-mass cells of bovine blastocysts for establishment of self-renewing pluripotent cells. Theriogenology 2012.
219. Khan DR, Dubé D, Gall L, Peynot N, Ruffini S, Laffont L, Le Bourhis D, Degrelle S, Jouneau A, Duranthon V. Expression of pluripotency master regulators during two key developmental transitions: EGA and early lineage specification in the bovine embryo. PLoS One 2012; 7:e34110.
220. Yang QE, Fields SD, Zhang K, Ozawa M, Johnson SE, Ealy AD. Fibroblast growth factor 2 promotes primitive endoderm development in bovine blastocyst outgrowths. Biol Reprod 2011; 85:946-953.
221. Sakatani M, Alvarez NV, Takahashi M, Hansen PJ. Consequences of physiological heat shock beginning at the zygote stage on embryonic development and expression of stress response genes in cattle. J Dairy Sci 2012; 95:3080-3091.
222. Yang QE, Johnson SE, Ealy AD. Protein kinase C delta mediates fibroblast growth factor-2-induced interferon-tau expression in bovine trophoblast. Biol Reprod 2011; 84:933-943.
223. Ozawa M, Hansen PJ. A novel method for purification of inner cell mass and trophectoderm cells from blastocysts using magnetic activated cell sorting. Fertil Steril 2011; 95:799-802.
224. Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, Speleman F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol 2002; 3:RESEARCH0034.
225. Block J, Fischer-Brown AE, Rodina TM, Ealy AD, Hansen PJ. The effect of in vitro treatment of bovine embryos with IGF-1 on subsequent development in utero to Day 14 of gestation. Theriogenology 2007; 68:153-161.

171
226. Fischer-Brown AE, Lindsey BR, Ireland FA, Northey DL, Monson RL, Clark SG, Wheeler MB, Kesler DJ, Lane SJ, Weigel KA, Rutledge JJ. Embryonic disc development and subsequent viability of cattle embryos following culture in two media under two oxygen concentrations. Reprod Fertil Dev 2004; 16:787-793.
227. Richter KS, Harris DC, Daneshmand ST, Shapiro BS. Quantitative grading of a human blastocyst: optimal inner cell mass size and shape. Fertil Steril 2001; 76:1157-1167.
228. Vallier L, Alexander M, Pedersen RA. Activin/Nodal and FGF pathways cooperate to maintain pluripotency of human embryonic stem cells. J Cell Sci 2005; 118:4495-4509.
229. Smith AG, Nichols J, Robertson M, Rathjen PD. Differentiation inhibiting activity (DIA/LIF) and mouse development. Dev Biol 1992; 151:339-351.
230. Gong G, Roach ML, Jiang L, Yang X, Tian XC. Culture conditions and enzymatic passaging of bovine ESC-like cells. Cell Reprogram 2010; 12:151-160.
231. Rooke J, Ewen M, McEvoy T, Entrican G, Ashworth C. Effect of inclusion of serum and granulocyte-macrophage colony stimulating factor on secretion of interferon-tau during the in vitro culture of ovine embryos. Reprod Fertil Dev 2005; 17:513-521.
232. Mui AL, Wakao H, Harada N, O'Farrell AM, Miyajima A. Interleukin-3, granulocyte-macrophage colony-stimulating factor, and interleukin-5 transduce signals through two forms of STAT5. J Leukoc Biol 1995; 57:799-803.
233. Gluckman PD, Hanson MA, Cooper C, Thornburg KL. Effect of in utero and early-life conditions on adult health and disease. N Engl J Med 2008; 359:61-73.
234. Gluckman PD, Hanson MA. Living with the past: evolution, development, and patterns of disease. Science 2004; 305:1733-1736.
235. Gillman MW. Developmental origins of health and disease. N Engl J Med 2005; 353:1848-1850.

172
236. Gluckman PD, Hanson MA, Beedle AS. Early life events and their consequences for later disease: a life history and evolutionary perspective. Am J Hum Biol 2007; 19:1-19.
237. Scherrer U, Rimoldi SF, Rexhaj E, Stuber T, Duplain H, Garcin S, de Marchi SF, Nicod P, Germond M, Allemann Y, Sartori C. Systemic and pulmonary vascular dysfunction in children conceived by assisted reproductive technologies. Circulation 2012; 125:1890-1896.
238. Reefhuis J, Honein MA, Schieve LA, Correa A, Hobbs CA, Rasmussen SA, Study NBDP. Assisted reproductive technology and major structural birth defects in the United States. Hum Reprod 2009; 24:360-366.
239. Farin PW, Crosier AE, Farin CE. Influence of in vitro systems on embryo survival and fetal development in cattle. Theriogenology 2001; 55:151-170.
240. Young LE, Sinclair KD, Wilmut I. Large offspring syndrome in cattle and sheep. Rev Reprod 1998; 3:155-163.
241. Sinclair KD, Young LE, Wilmut I, McEvoy TG. In-utero overgrowth in ruminants following embryo culture: lessons from mice and a warning to men. Hum Reprod 2000; 15 Suppl 5:68-86.
242. Fleming TP, Kwong WY, Porter R, Ursell E, Fesenko I, Wilkins A, Miller DJ, Watkins AJ, Eckert JJ. The embryo and its future. Biol Reprod 2004; 71:1046-1054.
243. Aiken CE, Ozanne SE. Sex differences in developmental programming models. Reproduction 2013; 145:R1-13.
244. Trivers RL, Willard DE. Natural selection of parental ability to vary the sex ratio of offspring. Science 1973; 179:90-92.
245. Bermejo-Alvarez P, Rizos D, Rath D, Lonergan P, Gutierrez-Adan A. Sex determines the expression level of one third of the actively expressed genes in bovine blastocysts. Proc Natl Acad Sci U S A 2010; 107:3394-3399.

173
246. Kobayashi S, Isotani A, Mise N, Yamamoto M, Fujihara Y, Kaseda K, Nakanishi T, Ikawa M, Hamada H, Abe K, Okabe M. Comparison of gene expression in male and female mouse blastocysts revealed imprinting of the X-linked gene, Rhox5/Pem, at preimplantation stages. Curr Biol 2006; 16:166-172.
247. Dobbs KB, Khan FA, Sakatani M, Moss JI, Ozawa M, Ealy AD, Hansen PJ. Regulation of Pluripotency of Inner Cell Mass and Growth and Differentiation of Trophectoderm of the Bovine Embryo by Colony Stimulating Factor 2. Biol Reprod 2013.
248. Karagenc L, Lane M, Gardner DK. Granulocyte-macrophage colony-stimulating factor stimulates mouse blastocyst inner cell mass development only when media lack human serum albumin. Reprod Biomed Online 2005; 10:511-518.
249. Robertson I, Nelson R. Certification and identification of the embryo. Manual of the International Embryo Transfer Society: International Embryo Transfer Society; 1998: 103-116.
250. Parrish JJ, Susko-Parrish JL, Leibfried-Rutledge ML, Critser ES, Eyestone WH, First NL. Bovine in vitro fertilization with frozen-thawed semen. Theriogenology 1986; 25:591-600.
251. Bisinotto RS, Ribeiro ES, Martins LT, Marsola RS, Greco LF, Favoreto MG, Risco CA, Thatcher WW, Santos JE. Effect of interval between induction of ovulation and artificial insemination (AI) and supplemental progesterone for resynchronization on fertility of dairy cows subjected to a 5-d timed AI program. J Dairy Sci 2010; 93:5798-5808.
252. Park JH, Lee JH, Choi KM, Joung SY, Kim JY, Chung GM, Jin DI, Im KS. Rapid sexing of preimplantation bovine embryo using consecutive and multiplex polymerase chain reaction (PCR) with biopsied single blastomere. Theriogenology 2001; 55:1843-1853.
253. Robert C, Nieminen J, Dufort I, Gagné D, Grant JR, Cagnone G, Plourde D, Nivet AL, Fournier É, Paquet É, Blazejczyk M, Rigault P, et al. Combining resources to obtain a comprehensive survey of the bovine embryo transcriptome through deep sequencing and microarrays. Mol Reprod Dev 2011; 78:651-664.
254. Carvajal-Rodriguez A, de Uña-Alvarez J. Assessing significance in high-throughput experiments by sequential goodness of fit and q-value estimation. PLoS One 2011; 6:e24700.

174
255. Dennis G, Sherman BT, Hosack DA, Yang J, Gao W, Lane HC, Lempicki RA. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol 2003; 4:P3.
256. Yamada T, Letunic I, Okuda S, Kanehisa M, Bork P. iPath2.0: interactive pathway explorer. Nucleic Acids Res 2011; 39:W412-415.
257. Goossens K, Van Poucke M, Van Soom A, Vandesompele J, Van Zeveren A, Peelman LJ. Selection of reference genes for quantitative real-time PCR in bovine preimplantation embryos. BMC Dev Biol 2005; 5:27.
258. Oliveira CS, Saraiva NZ, Cruz MH, Mazeti B, Oliveira LZ, Lopes FL, Garcia JM. HDAC inhibition decreases XIST expression on female IVP bovine blastocysts. Reproduction 2013; 145:9-17.
259. de Montera B, Fournier E, Shojaei Saadi HA, Gagné D, Laflamme I, Blondin P, Sirard MA, Robert C. Combined methylation mapping of 5mC and 5hmC during early embryonic stages in bovine. BMC Genomics 2013; 14:406.
260. Smyth GK. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat Appl Genet Mol Biol 2004; 3:Article3.
261. Smith ZD, Chan MM, Mikkelsen TS, Gu H, Gnirke A, Regev A, Meissner A. A unique regulatory phase of DNA methylation in the early mammalian embryo. Nature 2012; 484:339-344.
262. Elango N, Yi SV. Functional relevance of CpG island length for regulation of gene expression. Genetics 2011; 187:1077-1083.
263. Krausz C, Sandoval J, Sayols S, Chianese C, Giachini C, Heyn H, Esteller M. Novel insights into DNA methylation features in spermatozoa: stability and peculiarities. PLoS One 2012; 7:e44479.
264. Jurka J, Kapitonov VV, Pavlicek A, Klonowski P, Kohany O, Walichiewicz J. Repbase Update, a database of eukaryotic repetitive elements. Cytogenet Genome Res 2005; 110:462-467.

175
265. Betteridge KJ, Eaglesome MD, Randall GC, Mitchell D. Collection, description and transfer of embryos from cattle 10--16 days after oestrus. J Reprod Fertil 1980; 59:205-216.
266. Machado GM, Ferreira AR, Guardieiro MM, Bastos MR, Carvalho JO, Lucci CM, Diesel TO, Sartori R, Rumpf R, Franco MM, Dode MA. Morphology, sex ratio and gene expression of day 14 in vivo and in vitro bovine embryos. Reprod Fertil Dev 2013; 25:600-608.
267. Kimura K, Spate LD, Green MP, Murphy CN, Seidel GE, Jr., Roberts RM. Sexual dimorphism in interferon-tau production by in vivo-derived bovine embryos. Mol Reprod Dev 2004; 67:193-199.
268. Bermejo-Alvarez P, Rizos D, Rath D, Lonergan P, Gutierrez-Adan A. Epigenetic differences between male and female bovine blastocysts produced in vitro. Physiol Genomics 2008; 32:264-272.
269. Sharkey DJ, Macpherson AM, Tremellen KP, Robertson SA. Seminal plasma differentially regulates inflammatory cytokine gene expression in human cervical and vaginal epithelial cells. Mol Hum Reprod 2007; 13:491-501.
270. Palma GA, Müller M, Brem G. Effect of insulin-like growth factor I (IGF-I) at high concentrations on blastocyst development of bovine embryos produced in vitro. J Reprod Fertil 1997; 110:347-353.
271. Morgan HD, Santos F, Green K, Dean W, Reik W. Epigenetic reprogramming in mammals. Hum Mol Genet 2005; 14 Spec No 1:R47-58.
272. Bultman SJ, Gebuhr TC, Pan H, Svoboda P, Schultz RM, Magnuson T. Maternal BRG1 regulates zygotic genome activation in the mouse. Genes Dev 2006; 20:1744-1754.
273. Canovas S, Cibelli JB, Ross PJ. Jumonji domain-containing protein 3 regulates histone 3 lysine 27 methylation during bovine preimplantation development. Proc Natl Acad Sci U S A 2012; 109:2400-2405.
274. Shi W, Dirim F, Wolf E, Zakhartchenko V, Haaf T. Methylation reprogramming and chromosomal aneuploidy in in vivo fertilized and cloned rabbit preimplantation embryos. Biol Reprod 2004; 71:340-347.

176
275. Giraldo AM, Hylan DA, Ballard CB, Purpera MN, Vaught TD, Lynn JW, Godke RA, Bondioli KR. Effect of epigenetic modifications of donor somatic cells on the subsequent chromatin remodeling of cloned bovine embryos. Biol Reprod 2008; 78:832-840.
276. Wojdacz TK, Dobrovic A. Methylation-sensitive high resolution melting (MS-HRM): a new approach for sensitive and high-throughput assessment of methylation. Nucleic Acids Res 2007; 35:e41.
277. Wittwer CT, Reed GH, Gundry CN, Vandersteen JG, Pryor RJ. High-resolution genotyping by amplicon melting analysis using LCGreen. Clin Chem 2003; 49:853-860.
278. Wojdacz TK, Dobrovic A, Hansen LL. Methylation-sensitive high-resolution melting. Nat Protoc 2008; 3:1903-1908.
279. Rougier N, Bourc'his D, Gomes DM, Niveleau A, Plachot M, Pàldi A, Viegas-Péquignot E. Chromosome methylation patterns during mammalian preimplantation development. Genes Dev 1998; 12:2108-2113.
280. Dean W, Santos F, Reik W. Epigenetic reprogramming in early mammalian development and following somatic nuclear transfer. Semin Cell Dev Biol 2003; 14:93-100.
281. Bestor TH. The DNA methyltransferases of mammals. Hum Mol Genet 2000; 9:2395-2402.
282. Zhu JK. Active DNA demethylation mediated by DNA glycosylases. Annu Rev Genet 2009; 43:143-166.
283. Camous S, Kopecný V, Fléchon JE. Autoradiographic detection of the earliest stage of [3H]-uridine incorporation into the cow embryo. Biol Cell 1986; 58:195-200.
284. Borgel J, Guibert S, Li Y, Chiba H, Schübeler D, Sasaki H, Forné T, Weber M. Targets and dynamics of promoter DNA methylation during early mouse development. Nat Genet 2010; 42:1093-1100.

177
285. Chen T, Zhang YL, Jiang Y, Liu SZ, Schatten H, Chen DY, Sun QY. The DNA methylation events in normal and cloned rabbit embryos. FEBS Lett 2004; 578:69-72.
286. Ozawa M, Sakatani M, Yao J, Shanker S, Yu F, Yamashita R, Wakabayashi S, Nakai K, Dobbs KB, Sudano MJ, Farmerie WG, Hansen PJ. Global gene expression of the inner cell mass and trophectoderm of the bovine blastocyst. BMC Dev Biol 2012; 12:33.
287. Gutiérrez-Adán A, Behboodi E, Murray JD, Anderson GB. Early transcription of the SRY gene by bovine preimplantation embryos. Mol Reprod Dev 1997; 48:246-250.
288. Avery B, Jørgensen CB, Madison V, Greve T. Morphological development and sex of bovine in vitro-fertilized embryos. Mol Reprod Dev 1992; 32:265-270.
289. Gutiérrez-Adán A, Behboodi E, Andersen GB, Medrano JF, Murray JD. Relationship between stage of development and sex of bovine IVM-IVF embryos cultured in vitro versus in the sheep oviduct. Theriogenology 1996; 46:515-525.
290. Gebert C, Wrenzycki C, Herrmann D, Gröger D, Reinhardt R, Hajkova P, Lucas-Hahn A, Carnwath J, Lehrach H, Niemann H. The bovine IGF2 gene is differentially methylated in oocyte and sperm DNA. Genomics 2006; 88:222-229.
291. Hansen PJ, Block J, Loureiro B, Bonilla L, Hendricks KE. Effects of gamete source and culture conditions on the competence of in vitro-produced embryos for post-transfer survival in cattle. Reprod Fertil Dev 2010; 22:59-66.
292. Block J. Use of insulin-like growth factor-1 to improve post-transfer survival of bovine embryos produced in vitro. Theriogenology 2007; 68 Suppl 1:S49-55.
293. El-Sayed A, Hoelker M, Rings F, Salilew D, Jennen D, Tholen E, Sirard MA, Schellander K, Tesfaye D. Large-scale transcriptional analysis of bovine embryo biopsies in relation to pregnancy success after transfer to recipients. Physiol Genomics 2006; 28:84-96.
294. Boland MP, Lonergan P, O'Callaghan D. Effect of nutrition on endocrine parameters, ovarian physiology, and oocyte and embryo development. Theriogenology 2001; 55:1323-1340.

178
BIOGRAPHICAL SKETCH
Kyle Dobbs was born and raised in Webster Groves, MO, a suburb of St. Louis,
to Kathy and Brad Dobbs. He is the older brother of Kory. His father spurred his love of
science and animals at a young age. He attended high school at Lutheran High School
South in Afton, MO where he graduated in 2004. Following graduation, he moved to
Columbia, MO to begin his collegiate career at the University of Missouri where he
graduated in 2008 with his Bachelor of Science degree in biological sciences and a
minor in chemistry. During his undergraduate studies, he worked as a researcher in the
laboratory of Dr. Randall Prather. Following graduation, he began his graduate career
in Dr. Prather’s laboratory, where he studied polyadenylation of porcine transcripts
during early embryogenesis. After graduation in 2010 with his Master of Science
degree, he married his high school sweetheart, Lauren (Kienstra) and immediately
moved the next day to Gainesville, FL to begin work on his doctorate with Dr. Peter J.
Hansen in the Animal Molecular and Cellular Biology Graduate Program. While at
Florida, he was supported by a Graduate Fellowship.
One of the experiments for his dissertation was recognized as the outstanding
student poster presentation at the 40th Annual Conference of the International Embryo
Transfer Society held January 11-14, 2014 in Reno NV.
After completing his Doctor of Philosophy degree, Kyle and Lauren will move to
Boston, MA where he will continue his career as a post-doctoral researcher at
Northeastern University in the laboratory of Dr. Jonathan Tilly.